

Палеонтологический журнал, 2022, № 2, стр. 9-20
Новое семейство радиолярий позднего палеозоя Provisocyntridae fam. nov.
М. С. Афанасьева *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: afanasieva@paleo.ru
Поступила в редакцию 05.09.2021
После доработки 14.09.2021
Принята к публикации 14.09.2021
- EDN: UIWCRR
- DOI: 10.31857/S0031031X22020027
Аннотация
Проведена ревизия и уточнен диагноз рода Provisocyntra Nazarov et Ormiston, 1987, emend. nov. Проанализированы особенности строения морфотипа рода Provisocyntra, образованного одновременным центробежным и тангентальным способом роста скелета, и подтверждено наличие внутреннего каркаса в виде микросферы. Показано, что род Provisocyntra объединяет 10 видов радиолярий карбона–средней перми. Выполнено описание четырех новых видов P. grandis sp. nov., P. kononovae sp. nov., P. magniporosa sp. nov., P. valminazae sp. nov. из нижнетурнейских отложений Волго-Уральского бассейна. Установлено новое семейство Provisocyntridae fam. nov. и дополнен диагноз и состав отряда Spongiata Afanasieva et Amon, 2003, emend. nov.
ВВЕДЕНИЕ
Достоверные и очень хорошо сохранившиеся радиолярии раннетурнейского возраста впервые встречены в приграничных с девоном отложениях нижнего карбона скважины Мелекесская-1 Волго-Уральского бассейна России (рис. 1).
Радиолярии раннего турне в отложениях скв. Мелекесская-1 объединяют 25 видов из девяти родов: Adamasirad Afanasieva, 2009, Ceratoikiscum Deflandre, 1953, Duodecimentactinia Won, 1997, Entactinia Foreman, 1963, Nestelliana Afanasieva, 2022, Provisocyntra Nazarov et Ormiston, 1987, Radiobisphaera Won, 1997, Spinoalius Afanasieva, 2019 и Spongentactinia Nazarov, 1975.
Конодонты в нижнекаменноугольных, турнейских отложениях скважины Мелекесская-1 представлены только единичными экземплярами родов Siphonodella, Bispathodus, Neopolygnathus, Polygnathus и др. Там же, наряду с радиоляриями, Л.И. Кононова обнаружила зональный конодонт Siphonodella duplicata (Branson et Mehl). Учитывая бедное содержание конодонтов в изученной части разреза, можно лишь условно выделить зоны конодонтов Siphonodilla duplicata и Siphonodilla belkai – Siphonodilla quadruplicata (рис. 2) (Афанасьева, Кононова, 2021; Afanasieva, Kononova, 2021).
Среди радиолярий обращают на себя внимание представители рода Provisocyntra, которые имеют ряд характерных признаков, отличающих их от всех иных губчатых Spumellaria. В предлагаемой работе проанализированы особенности одновременного центробежного и тангентального формирования морфотипа рода Provisocyntra и подтверждено наличие внутреннего каркаса в виде микросферы.
Анализ морфологических особенностей скелетов Provisocyntra позволил: (1) уточнить диагноз рода Provisocyntra Nazarov et Ormiston, 1987, emend. nov., расширить его временнóе и пространственное распространение и увеличить видовой состав до десяти видов; (2) выделить и описать четыре новых вида P. grandis sp. nov., P. kononovae sp. nov., P. magniporosa sp. nov., P. valminazae sp. nov. из нижнетурнейских отложений Волго-Уральского бассейна (рис. 2); (3) установить присутствие вида P. cassicula Nazarov et Ormiston, 1987, известного из нижнекаменноугольных отложений США; (4) установить новое семейство Provisocyntridae fam. nov.; (5) дополнить диагноз и состав отряда Spongiata Afanasieva et Amon, 2003, emend. nov.
МОРФОЛОГИЯ СКЕЛЕТА
Настоящая статья посвящена всестороннему исследованию рода Provisocyntra, очень редкий морфотип которого образован одновременным центробежным и тангентальным способом роста скелета и наличием внутреннего каркаса в виде пористой микросферы.
Морфотип, согласно определению П. Де Вевера с коллегами (De Wever et al., 2001, с. 454), представляет собой таксон, который определяется морфологическими признаками без эволюционных ограничений11. То есть, морфотип – это совокупность морфологических признаков, определяющая общую форму скелета и отраженная в диагнозе таксона.
Формирование скелета
Необычная морфология скелета впервые была детально проанализирована Б.Б. Назаровым и А. Ормистоном (Nazarov, Ormiston, 1987) при описании рода Provisocyntra из каменноугольных отложений: (1) США, Северная Дакота, нижняя часть верхнего турне (зона Siphonodella isosticha), формация Лоджепол, и Оклахома, нижневизейский подъярус; (2) Узбекистан, Тянь-Шань, Пскемский хребет, серпуховский ярус; (3) Северо-Восток России, Магаданская обл., Приколымское поднятие, р. Поповка, московский ярус.
Новые данные и переосмысление взглядов предыдущих исследователей о формировании многочисленных оболочек у раковин радиолярий, образовавшихся в результате центробежного роста скелета (Hollande, Enjumet, 1960; Deflandre, 1964; Kling, 1978), а также в результате более позднего слияния тангентальных элементов по мере роста организма (Enriques, 1931; Stürmer, 1966), позволили Назарову и Ормистону (Nazarov, Ormiston, 1987) выдвинуть предположение о комбинированном механизме формирования скелета Provisocyntra одновременно центробежным и тангентальным способами.
При этом Назаров и Ормистон (Nazarov, Ormiston, 1987) скептически относились к концепции центростремительного развития скелета (Schwartz, 1931; Won, 1983). Автор придерживается того же мнения. Центростремительное формирование скелета прослеживается в ограниченных условиях, например, при зарастании пор скелета: края пор нарастают внутрь, поры постепенно сужаются, и образуется пористая или пластинчатая структура скелета (Afanasieva et al., 2005; Афанасьева, 2006).
Основные иглы скелета Provisocyntra образуются центробежным способом, при этом по мере роста они сначала расширяются, а затем сужаются к терминальной части иглы (рис. 3, а; 4, г).
Рис. 3.
Схема строения морфотипа рода Provisocyntra (по: Nazarov, Ormiston, 1987, с изменениями и дополнениями): а – гипотетическая дифференциация губчатой скелетной ткани на радиальные и тангенциальные элементы, б – фрагмент. Обозначения: c – полигональные ячейки, m – микросфера, s – основные иглы; элементы скелета: r – радиальные, t – тангентальные.
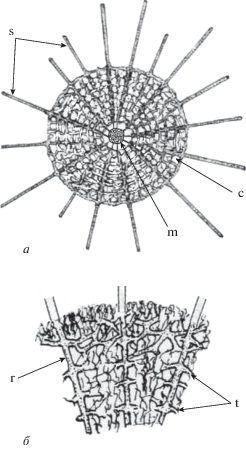
Зарождающиеся внутренние оболочки скелета добавляются тангентальным способом в процессе онтогенетического развития организма. Тангентальные элементы скелета (апофизы) формируются вдоль радиальных центробежных элементов (основные иглы) в направлении, перпендикулярном к их морфологической оси (рис. 3, б; 4, в). По мере развития организма апофизы в разной степени разветвляются, их ветви соединяются и образуют губчатую ткань скелета.
Таким образом, механизм тангентального роста в разной степени разветвляющихся апофизов и зарождение внутренних оболочек у Provisocyntra (рис. 3, б; 4, в) контрастирует с центробежным, последовательным добавлением оболочек у иных радиолярий с несколькими сферами, таких как Copicyntra Nazarov et Ormiston, 1985, Plenoentactinia Won, 1997, Pluristratoentactinia Nazarov, 1981 и других представителей Spumellaria и Sphaerellaria.
Это означает, что, по сути, все зарождающиеся оболочки скелета Provisocyntra могли возникать не последовательно в процессе формирования скелета, а одновременно – после достижения организмом зрелости, т.е. полного законченного развития.
Микросфера
Морфотип рода Provisocyntra представляет собой своеобразную и очень редкую структуру, отличающуюся, помимо комбинированного тангентального и центробежного способа формирования скелета, наличием внутреннего каркаса в форме пористой микросферы (рис. 3, а).
В первоначальном диагнозе рода Provisocyntra Назаров и Ормистон (Nazarov, Ormiston, 1987) написали, что строение внутреннего каркаса не совсем ясно и, вероятно, спикула с очень тонкими лучами эксцентрично расположена внутри небольшой пористой сферы. При этом на рисунках в работе Назарова и Ормистона нарисована спикула внутри предполагаемой микросферы (Nazarov, Ormiston, 1987, рис. 1b и 3e, 3b).
Однако при описании вида Provisocyntra cassicula Назаров и Ормистон (Nazarov, Ormiston, 1987, с. 72, табл. 1 , фиг. 5) указали, что радиальные элементы сходятся на центральном внутреннем каркасе, который в исключительных образцах выглядит как пористая сфера22 диаметром примерно 40 мкм (рис. 3, а). Вместе с тем они (Nazarov, Ormiston, 1987, с. 72) написали, что “сохранность слишком несовершенна, чтобы положительно относиться к центральной сфере”33. Таким образом, отсутствие уверенной идентификации внутреннего каркаса в форме микросферы в работе Назарова и Ормистона (Nazarov, Ormiston, 1987) объясняется неудовлетворительной сохранностью и техническими сложностями изображения скелетов радиолярий.
Рис. 4.
Provisocyntra cassicula Nazarov et Ormiston, 1987 из нижнетурнейских отложений скв. Мелекесская-1: а, б – экз. ПИН, № 5508/6916-04, инт. 1881.7–1883.2 м, обр. 6916: а – штрих = 83 мкм, б – фрагмент, штрих – 28 мкм; в, г – экз. ПИН, № 5508/17603, инт. 1887.5–1888.25 м, обр. 6956: в – штрих = 110 мкм, г – фрагмент, штрих – 50 мкм.
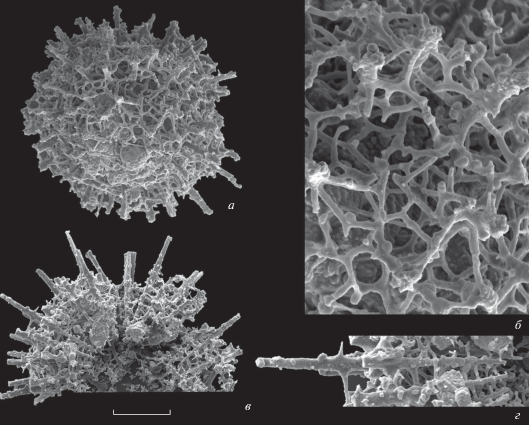
Рис. 5.
Распространение видов из рода Provisocyntra: 1, 2 – Россия, Волго-Уральский бассейн, скв. Мелекесская-1; нижний карбон, нижнетурнейский подъярус: 1 – нижняя экозона радиолярий, зона конодонтов Siphonodella duplicata, 2 – верхняя экозона радиолярий, зоны конодонтов (условно) Siphonodella belkai – Siphonodella quadruplicata (Афанасьева, Кононова, 2021; Afanasieva, Kononova, 2021); 3 – США, Северная Дакота; нижний карбон, верхнетурнейский подъярус, зона конодонтов Siphonodella isosticha (Nazarov, Ormiston, 1987); 4 – Германия, Франкенвальд, Рейнские Сланцевые горы; нижний карбон, верхнетурнейский подъярус, слои Pericyclus-γ, зона конодонтов Scaliognathus anchoralis (Won, 1983); 5 – США, Оклахома; нижний карбон, нижневизейский подъярус, формация Сикамор, зона конодонтов Gnathodus texanus (Nazarov, Ormiston, 1987); 6 – Узбекистан, Тянь-Шань, Пскемский хребет; нижний карбон, серпуховский ярус (Nazarov, Ormiston, 1987); 7 – Северо-Восток России, Приколымское поднятие, Магаданская обл., р. Поповка; верхний карбон, московский ярус (Nazarov, Ormiston, 1987); 8 – США, Западный Техас, Гваделупские горы; средняя пермь (гваделупий), роудский ярус, верхняя часть формации Катофф, зона конодонтов Jinogondolella nankingensis (Nestell, Nestell, 2021).
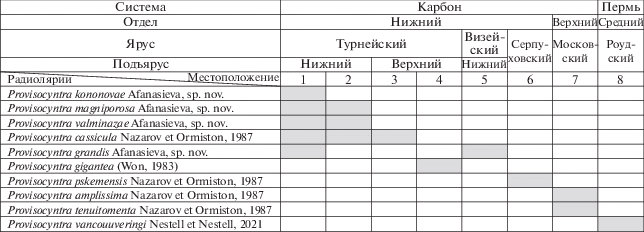
Впервые очень хорошо сохранившуюся крошечную полую микросферу (12 мкм) обнаружили Г.П. и М.К. Нестелл (Nestell, Nestell, 2021, табл. 3 , фиг. 14b ; табл. 4 , фиг. 1b) в скелете Provisocyntra vancouveringi Nestell et Nestell, 2021 из отложений роудского яруса средней перми в Гваделупских горах Западного Техаса, США. Эти авторы также подтвердили образование скелета Provisocyntra одновременным центробежным и тангентальным способами.
В настоящей статье описано четыре новых вида Provisocyntra: P. grandis sp. nov., P. kononovae sp. nov., P. magniporosa sp. nov., P. valminazae sp. nov. из отложений нижнего турне скважины Мелекесская-1 Волго-Уральского бассейна России. На одном из экземпляров P. valminazae sp. nov. отчетливо виден фрагмент достаточно хорошо сохранившейся пористой микросферы диаметром 20 мкм (табл. III , фиг. 1а, черная стрелка; см. вклейку).
ИСТОРИЯ ИЗУЧЕНИЯ РОДА PROVISOCYNTRA
Первоначально род Provisocyntra и пять видов (P. amplissima, P. cassicula, P. pskemensis, P. tenuitomenta, Provisocyntra sp.) были описаны (Nazarov, Ormiston, 1987) из отложений карбона США, Узбекистана и Северо-Востока России. Также в состав рода Provisocyntra Назаров и Ормистон включили вид Centrocubus? giganteus Won, 1983, описанный М.‑З. Вон (Won, 1983, с. 135–136, табл. 7 , фиг. 6–9 ) из топовых верхнетурнейских отложений (слои Pericyclus-γ) Рейнских Сланцевых гор Германии (рис. 5).
Рис. 6.
Основные абсолютные и относительные морфометрические параметры скелетов сферических радиолярий палеозоя (Афанасьева, 2000; Afanasieva et al., 2005): D − диаметр внешней сферы, dp − диаметр полигональный ячеек внешней оболочки, tw − ширина прутьев ячеек внешней оболочки, L − длина основных игл, wL − ширина основания основных игл.
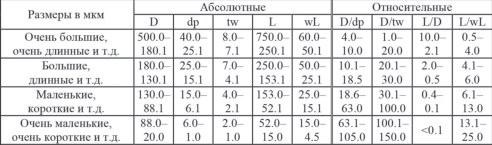
Рис. 7.
Новый вид Provisocyntra grandis sp. nov. из нижнетурнейских отложений скв. Мелекесская-1, Волго-Уральский бассейн, Россия: а–в – голотип ПИН, № 5508/6928-09, инт. 1883.7–1883.8 м, обр. 6928: а – фрагмент, штрих = 50 мкм, б – штрих = 175 мкм, в – фрагмент, штрих = 54 мкм; г–е – экз. ПИН, № 5508/6925-10, инт. 1881.7–1883.2 м, обр. 6925: г – фрагмент, штрих = 50 мкм, д – штрих = 152 мкм, е – фрагмент, штрих = 50 мкм.

По мнению Назарова и Ормистона (Nazarov, Ormiston, 1987), морфотип рода Provisocyntra имеет некоторое сходство с отдельными представителями подсемейства Astroentactiniinae Nazarov et Ormiston, 1985 по наличию внутренней спикулы, строению многочисленных стержневидных игл, апофизов и дифференциации губчатого слоя, и поэтому род был включен в семейство Entactiniidae Riedel, 1967.
Однако П. Нобл с коллегами (Noble et al., 2017) отметили, что поскольку Назаров и Ормистон (Nazarov, Ormiston, 1987) интерпретировали внутренний каркас как спикулу, находящуюся в центре сферы, то последняя могла представлять собой пролокулюм, т. е. начальную камеру. Наличие пролокулюма не позволяет рассматривать род Provisocyntra в составе семейства Entactiniidae Riedel, 1967.
В 1997 г. Вон (Won, 1997) описала новый вид Provisocyntra conferta из верхнедевонской, франской формации Гого, Австралия. Внутренний каркас этого вида представлен шестилучевой спикулой со срединной балкой и апофизами на каждом ее луче, которые образуют субпрямоугольную R-рамку. Основываясь на наличии спикулы, Вон (Won, 1997, с. 383) исправила первоначальный диагноз рода Provisocyntra44. Однако в комментариях к роду она отметила, что “основная структура раковины этого рода такая же, как у Retentactinia gen. nov., кроме более многочисленных игл55”, и отнесла исправленный род к новому подсемейству Retentactiniinae (=Spongentactiniidae Nazarov, 1975, sensu De Wever et al., 2001) в составе семейства Entactiniidae.
Вместе с тем, анализ скелетов Provisocyntra conferta (Won, 1997, рис. 1, табл. 7 , фиг. 17–19 ; табл. 12 , фиг. 16, 17 ) показал, что наличие очень хорошо развитого внутреннего каркаса в виде шестилучевой спикулы со срединной балкой не позволяет отнести данный вид к роду Provisocyntra. Это же сомнение в родовой принадлежности вида P. conferta было высказано Нестелл и Нестеллем (Nestell, Nestell, 2021), предположившими, что данный вид относится либо к роду Retentactinia Won, 1997, либо к другому роду.
По мнению автора данной статьи, вид P. conferta Won, 1997, скорее всего, принадлежат к роду Retentactinia Won, 1997. Кроме этого автор, так же как Нестелл и Нестелл (Nestell, Nestell, 2021), не согласен с исправлением Вон (Won, 1997, с. 383) диагноза рода Provisocyntra.
Автор также присоединяется к точке зрения Нестелл и Нестелла (Nestell, Nestell, 2021) о том, что виды P. densa и P. ormistoni, описанные К. Фенгом и др. (Feng et al., 2007) из верхнечансинской формации Далонг на юге провинции Гуанси, Китай, не принадлежат к роду Provisocyntra. Судя по внутреннему строению скелетов (Feng et al., 2007, табл. 5 , фиг. 2–4, 8 ), эти виды, скорее всего, относятся к роду Copicyntra Nazarov et Ormiston, 1985.
Однако, автор данной статьи не согласен с Нестелл и Нестеллом (Nestell, Nestell, 2021), предлагающими включить род Provisocyntra в состав семейства Copicyntridae. Морфотип рода Provisocyntra характеризуется отсутствием четко тангентально дифференцированной губчатой оболочки на отдельные слои и формированием внутреннего каркаса в форме полой микросферы. В отличие от структуры скелета Provisocyntra, для морфотипа рода Copicyntra присуще наличие нескольких отчетливо выраженных промежуточных оболочек, развивающихся между внутренней пористой оболочкой и внешней губчатой, а также формирование внутреннего каркаса в виде спикулы.
К сожалению, строение внутреннего каркаса в скелете Copicyntra в виде спикулы представлено Назаровым и Ормистоном только на рисунках (Nazarov, Ormiston, 1985, рис. 6; 1987, рис. 1, d; Назаров, 1988, рис. 19 , ж).
Вместе с тем, Нобл и Джин (Noble, Jin, 2010, табл. 6 , фиг. 6, 7) на примере вида Copicyntra erinacea Noble et Jin, 2010 предполагают, что внутренние слои в скелетах Copicyntra формируются по спирали. Поэтому внутренней спикулы в центре скелета быть не может, а начало скелета может быть в форме микросферы. Однако в этом случае представители вида C. erinacea должны быть отнесены к другому роду.
НОВОЕ СЕМЕЙСТВО PROVISOCYNTRIDAE
Современные находки микросферы в скелетах Provisocyntra из отложений нижнетурнейского подъяруса нижнего карбона России (рис. 2) (настоящая работа) и роудского яруса средней перми США (рис. 5) (Nestell, Nestell, 2021) позволяют по-новому оценить систематическое положение рода Provisocyntra.
Достоверный внутренний каркас в виде шестилучевой спикулы у представителей рода Provisocyntra не наблюдался ни Назаровым и Ормистоном (Nazarov, Ormiston, 1987), ни Нестелл и Нестеллом (Nestell, Nestell, 2021). Шестилучевая спикула не была обнаружена и у вновь описываемых в данной работе видов Provisocyntra.
Вместе с тем, у хорошо сохранившегося экземпляра P. cassicula Назаров и Ормистон диагностировали внутренний каркас, который выглядит как пористая сфера (Nazarov, Ormiston, 1987, с. 72, табл. 1 , фиг. 5). При внимательном наблюдении можно рассмотреть внутренний каркас в виде сферы у P. pskemensis (Nazarov, Ormiston, 1987, с. 72, табл. 2 , фиг. 5, 6).
Центральная часть скелета P. vancouveringi Nestell et Nestell, 2021 представлена полой крошечной микросферой диаметром 12 мкм (Nestell, Nestell, 2021, табл. 3 , фиг. 14b ; табл. 4 , фиг. 1b).
На одном из экземпляров нового вида P. valminazae sp. nov. отчетливо виден фрагмент пористой микросферы диаметром 20 мкм (табл. II , фиг. 1а; см. вклейку).
Таким образом, наличие внутреннего каркаса в форме полой перфорированной или решетчатой микросферы у разных возрастных представителей рода Provisocyntra, в том числе у типового вида P. cassicula, позволяет уточнить диагноз рода, расширить его временнóе и пространственное распространение и увеличить видовой состав до десяти видов. Более того, особенности строения редкого для радиолярий палеозоя морфотипа позволяют установить новое семейство Provisocyntridae fam. nov. с типовым родом Provisocyntra.
Новое семейство Provisocyntridae устанавливается в рамках отряда Spongiata Afanasieva et Amon, 2003, что, в свою очередь, предусматривает необходимость дополнить диагноз новым элементом скелета – микросферой, а состав отряда расширить за счет нового семейства Provisocyntridae fam. nov.
ОПИСАНИЕ ТАКСОНОВ
ТИП RADIOLARIA
НАДКЛАСС POLYCYSTINA
КЛАСС SPUMELLARIA
ОТРЯД SPONGIATA AFANASIEVA ET AMON, 2003, EMEND. AFANASIEVA, NOV.
Spongiata: Афанасьева, Амон, 2003, с. 83; Afanasieva et al., 2005, с. 277, Афанасьева, Амон, 2006, с. 115.
Диагноз. Spumellaria с губчатой стенкой внешней сферической оболочки скелета и внутренним каркасом в виде двойной первичной спикулы или микросферы.
Состав. Четыре семейства: Spongentactiniidae Nazarov, 1975; Spongopolyentactiniidae Nazarov, 1975; Conocaryommidae Lipman, 1969, emend. De Wever et al., 2001; Provisocyntridae fam. nov.
Распространение. Средний кембрий–эоцен.
Замечания. Диагноз отряда дополнен новым элементом скелета – микросферой; состав отряда расширен за счет нового семейства Provisocyntridae fam. nov.
СЕМЕЙСТВО PROVISOCYNTRIDAE AFANASIEVA, FAM. NOV.
Типовой род – Provisocyntra Nazarov et Ormiston, 1987, emend. nov.
Диагноз. Spongiata с радиально дифференцированной губчатой тканью, заполняющей всю внутреннюю полость скелета, и начальными элементами зарождающихся внутренних оболочек; тангентальные апофизы расходятся от радиальных основных игл, берущих свое начало на центральном внутреннем каркасе в форме полой перфорированной микросферы; основные иглы многочисленные, стержневидной, конической или трехлопастной формы.
Состав. Типовой род.
Распространение. Карбон–средняя пермь.
Род Provisocyntra Nazarov et Ormiston, 1987, emend. Afanasieva nov.
Provisocyntra: Nazarov, Ormiston, 1987, с. 68; Назаров, 1988, с. 63; Noble et al., 2017, с. 446; Nestell, Nestell, 2021, с. 537; non Won, 1997, с. 383.
Типовой вид – Provisocyntra cassicula Nazarov et Ormiston, 1987; нижний карбон, верхнетурнейский подъярус, зона конодонтов Siphonodella isosticha; США, Северная Дакота.
Диагноз. Сферический скелет с радиально дифференцированной губчатой тканью, заполняющей всю внутреннюю полость скелета и образованной объединением разветвляющихся ветвей апофизов; тангентальные апофизы расходятся перпендикулярно от радиальных основных игл, а объединение разветвляющихся ветвей апофизов является начальным этапом зарождения внутренних оболочек; основные иглы берут свое начало на центральном внутреннем каркасе в форме полой перфорированной микросферы; основные иглы многочисленные, стержневидной, конической или трехлопастной формы с округлым, реже треугольным сечением, слегка сужаются к вершине и основанию (рис. 4, г).
Видовой состав. Десять видов из нижнего карбона–средней перми (рис. 2; 5): P. amplissima Nazarov et Ormiston, 1987, P. cassicula Nazarov et Ormiston, 1987, P. gigantea (Won, 1983), P. grandis sp. nov., P. kononovae sp. nov., P. magniporosa sp. nov., P. pskemensis Nazarov et Ormiston, 1987, P. tenuitomenta Nazarov et Ormiston, 1987, P. valminazae sp. nov., P. vancouveringi Nestell et Nestell, 2021.
Сравнение. От родов Somphoentactinia Nazarov, 1975 и Copicyntra Nazarov et Ormiston, 1985, характеризующихся наличием внутреннего каркаса в виде n-лучевой спикулы, род Provisocyntra отличается развитием внутреннего каркаса в форме полой микросферы. Кроме того, от рода Somphoentactinia, характеризующегося слабой дифференциацией внешней губчатой оболочки, и от рода Copicyntra, демонстрирующего завершенное развитие множества внутренних сфер, род Provisocyntra отличается отчетливой радиальной дифференциацией губчатой ткани, заполняющей всю внутреннюю полость скелета, и начальными элементами тангентально зарождающихся внутренних оболочек.
Замечания. 1. Род Provisocyntra первоначально рассматривался Назаровым и Ормистоном (Nazarov, Ormiston, 1987) в составе подсемейства Astroentactiniinae из семейства Entactiniidae на основании строения многочисленных стержневидных игл, апофизов и дифференциации губчатого слоя.
2. Позже род Provisocyntra вместе с родами Plenoentactinia и Copicyntra были объединены в подсемейство Plenoentactiniinae Afanasieva, 1999, характеризующееся внутренним каркасом в виде 8-12-24- и более лучевой спикулы и формированием скелета из трех и более сфер (Афанасьева, 1999, 2000; Afanasieva et al., 2005; Афанасьева, Амон, 2006).
3. В настоящее время анализ особенностей строения редкого морфотипа радиолярий на основании собственных и литературных данных выявил у разных возрастных представителей рода Provisocyntra наличие внутреннего каркаса в форме полой микросферы, что позволяет уточнить диагноз рода и установить для него новое семейство Provisocyntridae fam. nov.
4. Развитие внутреннего каркаса в форме полой микросферы не позволяет рассматривать род Provisocyntra в качестве предковой формы рода Copicyntra, характеризующегося присутствием внутренней спикулы, как это было предложено Назаровым и Ормистоном (Nazarov, Ormiston, 1987).
Provisocyntra kononovae Afanasieva, sp. nov.
Табл. II , фиг. 1–7
Название вида – в честь палеонтолога и стратиграфа Л.И. Кононовой.
Голотип – ПИН, № 5508/17648; Россия, Волго-Уральский бассейн, скв. Мелекесская-1, инт. 1887.5–1888.25 м, обр. 6956; нижний карбон, нижнетурнейский подъярус, зона конодонтов Siphonodella duplicata (рис. 2).
Описание. Скелет субсферический, большой (D = 132–178 мкм), имеет губчатую структуру и многочисленные основные иглы, которые пронизывают всю внутреннюю полость скелета. Внутренняя полость скелета заполнена соединенными тонкими стержневидными ветвями апофизов (tw = 4 мкм, D/tw = 33–45), формирующих губчатую ткань. Полигональные ячейки губчатой ткани большие (D/dp = 14–18), разного размера (dp = 9–13 мкм). При этом обращает на себя внимание развитие тончайшей сетки на поверхности скелета, образованной соединенными ветвями апофизов толщиной менее 2 мкм, которые формируют крупные полигональные ячейки размером до 20 мкм. Сетка, как правило, частично разрушена (табл. II , фиг. 4б). Апофизы развиваются тангентальным способом перпендикулярно к оси основных радиальных игл (табл. II , фиг. 1б, 2б, 3б, 7б). Основные радиальные иглы на внешней поверхности скелета конической формы, относительно короткие (L/D = 0.1–0.3) и толстые (L/wL = 3–6), с отчетливо выраженным срединным каналом (табл. II , фиг. 2б, 4б). Сечение игл, как правило, округлое, очень редко субтреугольное (табл. II , фиг. 2б). Строение внутреннего каркаса не установлено.
Размеры в мкм и отношения66 (рис. 6):
| Экз. № | D | dp | tw | L | wL | D/dp | D/tw | L/D | L/wL |
|---|---|---|---|---|---|---|---|---|---|
| 5508/17651 | 162 | 9 | 4 | 49 | 8 | 18 | 41 | 0.3 | 6 |
| 5508/17600 | 178 | 13 | 4 | 23 | 7 | 14 | 45 | 0.1 | 3 |
| 5508/17606 | 151 | 10 | 4 | 38 | 8 | 15 | 38 | 0.3 | 5 |
| 5508/17648 (голотип) | 132 | 9 | 4 | 21 | 7 | 15 | 33 | 0.2 | 3 |
Сравнение. От типового вида P. cassicula Nazarov et Ormiston, 1987 из нижнего карбона Северной Дакоты США и из нижнетурнейских отложений Волго-Уральского бассейна, скв. Мелекесская-1 (рис. 4), характеризующегося развитием очень большого скелета (D = 284–455 мкм), длинными стержневидными иглами (L ≤ 225 мкм) и большими полигональными ячейками на внешней поверхности скелета (dp = 20–50 мкм), вид P. kononovae sp. nov. отличается меньшими размерами скелета (D = 132–178 мкм), более тонкими ветвями апофизов (4 мкм против 5–6 мкм у P. cassicula), короткими основными иглами конической формы (L = 21–49 мкм), меньшими размерами полигональных ячеек на внешней поверхности скелета (dp = 9–13 мкм) и развитием тончайшей крупноячеистой (dp ≥ 20 мкм) сетки на поверхности скелета.
Материал: 13 экз. из типового местонахождения (рис. 2).
Provisocyntra valminazae Afanasieva, sp. nov.
Табл. III , фиг. 1–3
Название вида – в честь палеонтолога и стратиграфа Валентины Михайловны Назаровой.
Голотип – ПИН, № 5508/6928-24; Россия, Волго-Уральский бассейн, скв. Мелекесская-1, инт. 1881.7–1883.2 м, обр. 6925; нижний карбон, нижнетурнейский подъярус, зона конодонтов Siphonodella duplicata (рис. 2).
Описание. Скелет субсферический, очень большой (D = 246–340 мкм), имеет губчатую структуру и многочисленные основные иглы, которые пронизывают всю внутреннюю полость скелета. Внутренняя полость скелета заполнена соединенными очень тонкими стержневидными ветвями апофизов (tw = 2–3 мкм, D/tw = 113–142), формирующих губчатую ткань. Полигональные ячейки губчатой ткани большие (D/dp = = 24–36), разного размера (dp = 8–14 мкм). Апофизы развиваются тангентальным способом перпендикулярно к оси основных радиальных игл (табл. III , фиг. 1а, 3а). Основные радиальные иглы на внешней поверхности скелета стержневидной формы, относительно длинные (L/D = 0.4–0.7) и тонкие (L/wL = 8–10), с отчетливо выраженным срединным каналом (табл. III , фиг. 1а, 3а). Сечение игл округлое. Внутренний каркас представлен очень маленькой пористой микросферой размером 20 мкм (табл. III , фиг. 1а).
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | tw | L | wL | D/dp | D/tw | L/D | L/wL |
|---|---|---|---|---|---|---|---|---|---|
| 5508/6928-24 (голотип) |
284 | 8 | 2 | 134 | 16 | 36 | 142 | 0.5 | 8 |
| 5508/6914-27 | 340 | 14 | 3 | 120 | 20 | 24 | 113 | 0.4 | 6 |
| 5508/6928-11 | 246 | 8 | 2 | 175 | 18 | 31 | 123 | 0.7 | 10 |
Сравнение. См. вид P. magniporosa sp. nov.
Материал. Три экз. из нижнетурнейских отложений (зоны конодонтов Siphonodella duplicata и Siphonodella belkai – Siphonodella quadruplicata) скв. Мелекесская-1 (рис. 2).
Provisocyntra magniporosa Afanasieva, sp. nov.
Табл. III , фиг. 4, 5
Название вида от magnus лат. – большой, крупный и porosus лат. – пористый.
Голотип – ПИН, № 5508/6928-10; Россия, Волго-Уральский бассейн, скв. Мелекесская-1, инт. 1883.7–1883.8 м, обр. 6928; нижний карбон, нижнетурнейский подъярус, зона конодонтов Siphonodella duplicata (рис. 2).
Описание. Скелет субсферический, очень большой (D = 210–244 мкм), имеет губчатую структуру и многочисленные основные иглы, которые пронизывают всю внутреннюю полость скелета. Внутренняя полость скелета заполнена соединенными тонкими стержневидными ветвями апофизов (tw = 3 мкм, D/tw = 70–81), формирующих губчатую ткань. Полигональные ячейки губчатой ткани очень большие (D/dp = 8–10), разного размера (dp = 21–30 мкм). Апофизы развиваются тангентальным способом перпендикулярно к оси основных радиальных игл (табл. III , фиг. 4а, 4в, 5а). Основные радиальные иглы на внешней поверхности скелета стержневидной формы, иногда изогнутые (табл. III , фиг. 4б, 5б, 5в), длинные (L/D = 0.5–0.6) и очень тонкие (L/wL = = 15–25), с отчетливо выраженным срединным каналом (табл. III , фиг. 4в, 5а). Сечение игл, как правило, округлое, очень редко субтреугольное (табл. III , фиг. 5а). Строение внутреннего каркаса не установлено.
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | tw | L | wL | D/dp | D/tw | L/D | L/wL |
|---|---|---|---|---|---|---|---|---|---|
| 5508/6928-10 (голотип) | 244 | 30 | 3 | 120 | 7 | 8 | 81 | 0.5 | 17 |
| 5508/6914-13 | 212 | 22 | 3 | 125 | 5 | 10 | 71 | 0.6 | 25 |
| 5508/17620 | 210 | 21 | 3 | 135 | 9 | 10 | 70 | 0.6 | 15 |
Сравнение. От вида P. valminazae sp. nov., характеризующегося большими полигональными ячейками губчатой ткани (dp = 8–14 мкм) и тонкими основными иглами (wL = 16–20 мкм), новый вид P. magniporosa отличается наличием очень больших ячеек губчатой ткани (dp = 21–30 мкм) и очень тонкими основными иглами (wL = 5–9 мкм). При этом морфотипы обоих видов имеют в среднем сопоставимый размер внешней оболочки скелета: P. valminazae, D ≈ 290 мкм и P. magniporosa, D ≈ 222 мкм.
Материал. 4 экз. из нижнетурнейских отложений (зоны конодонтов Siphonodella duplicata и Siphonodella belkai – Siphonodella quadruplicata) скв. Мелекесская-1 (рис. 2).
Provisocyntra grandis Afanasieva, sp. nov.
Provisocyntra sp.: Nazarov, Ormiston, 1987, табл. 2 , фиг. 11 .
Название вида grandis лат. – большой, крупный, огромный.
Голотип – ПИН, № 5508/6928-09; Россия, Волго-Уральский бассейн, скв. Мелекесская-1, инт. 1883.7–1883.8 м, обр. 6928; нижний карбон, нижнетурнейский подъярус, зона конодонтов Siphonodella duplicata (рис. 2).
Описание (рис. 7). Скелет субсферический, очень большой (D = 462–472 мкм), имеет губчатую структуру и многочисленные основные иглы, которые пронизывают всю внутреннюю полость скелета. Внутренняя полость скелета заполнена соединенными очень тонкими стержневидными ветвями апофизов (tw = 3 мкм, D/tw = 154–157), формирующих губчатую ткань. Полигональные ячейки губчатой ткани разного размера (dp = 20–30 мкм), маленькие и большие (D/dp = 16–23). Апофизы развиваются тангентальным способом перпендикулярно к оси основных радиальных игл (рис. 7, а, г). Основные радиальные иглы на внешней поверхности скелета стержневидной формы, короткие (как правило, обломанные) (L/D = 0.2–0.4), тонкие (L/wL = 6–11), с отчетливо выраженным срединным каналом (рис. 7, а, в, г, е). По мере роста основные иглы сначала расширяются, а затем сужаются к терминальной части иглы (рис. 7, а). Сечение игл, как правило, округлое, реже субтреугольное (рис. 7, а, г). Строение внутреннего каркаса не установлено.
Размеры в мкм и отношения (рис. 6):
| Экз. № | D | dp | tw | L | wL | D/dp | D/tw | L/D | L/wL |
|---|---|---|---|---|---|---|---|---|---|
| 5508/6928-09 (голотип) | 472 | 30 | 3 | 182 | 16 | 16 | 157 | 0.4 | 11 |
| 5508/6925-10 | 462 | 20 | 3 | 84 | 13 | 23 | 154 | 0.2 | 6 |
Сравнение. От всех других видов Provisocyntra из нижнетурнейских отложений скв. Мелекесская-1 новый вид P. grandis sp. nov. отличается самым большим скелетом (D = 462–472 мкм) и самыми тонкими ветвями апофизов (D/tw = 154–157), формирующими губчатую ткань.
Замечания. Морфотип нового вида P. grandis sp. nov. похож на очень большой экземпляр Provisocyntra sp. (диаметр скелета 415 мкм) из нижневизейских отложений Оклахомы (Nazarov, Ormiston, 1987). В подписи к изображению Provisocyntra sp. Назаров и Ормистон (Nazarov, Ormiston, 1987, табл. 2 , фиг. 11 ) написали, что это неописанный вид, характеризующийся наличием коротких игл и неправильными маленькими ячейками внешнего губчатого слоя. Особенности морфологии скелета позволяют, вслед за Назаровым и Ормистоном (Nazarov, Ormiston, 1987), уверенно отнести данный экземпляр к роду Provisocyntra, зафиксировать его присутствие в отложениях нижневизейского подъяруса и рассматривать в составе нового вида P. grandis sp. nov. (рис. 5).
Материал. 3 экз.: два экз. из нижнетурнейских отложений (зона конодонтов Siphonodella duplicata) Волго-Уральского бассейна, Россия (рис. 2); один экз. из нижневизейских отложений (зона конодонтов Gnathodus texanus) Оклахомы, США (Nazarov, Ormiston, 1987).
* * *
Автор приносит свою искреннюю благодарность: А.С. Алексееву и Г.П. Нестелл за плодотворное обсуждение вопросов статьи; В.С. Вишневской за ценные советы и конструктивные рекомендации; Л.И. Кононовой за консультации относительно возраста вмещающих пород; Л.И. Кононовой и В.М. Назаровой за предоставленный материал по радиоляриям; А.Ф. Банникову и Я.М. Кузьминой за ценные советы и замечания при подготовке статьи к печати.
Исследование морфологии радиолярий проводилось на сканирующих электронных микроскопах CamScan и TESCAN на базе Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН). Коллекция радиолярий № 5508 (турнейский ярус, нижний карбон) хранится в ПИН РАН. Систематика радиолярий приведена по работам автора (Afanasieva et al., 2005; Афанасьева, Амон, 2006). Измерения элементов скелетов радиолярий и морфометрический анализ абсолютных и относительных значений параметров раковин радиолярий палеозоя проводились по стандартной методике (рис. 6) (Афанасьева, 2000; Afanasieva et al., 2005).
Работа выполнена в рамках бюджетной программы (государственного задания) ПИН РАН.
Список литературы
Афанасьева М.С. Новый вариант систематики радиолярий палеозоя // Геология и минеральные ресурсы европейского северо-востока России: новые результаты и новые перспективы. Матер. XIII Геол. съезда Республики Коми. Сыктывкар: ИГ КНЦ УрО РАН, 1999. Т. II. С. 253–256.
Афанасьева М.С. Атлас радиолярий палеозоя Русской платформы. М.: Научн. мир, 2000. 480 с.
Афанасьева М.С. Скелет радиолярий: формирование и морфология оболочек скелета // Палеонтол. журн. 2006. № 5. С. 13–24.
Афанасьева М.С., Амон Э.О. Новая классификация радиолярий // Палеонтол. журн. 2003. № 6. С. 72–86.
Афанасьева М.С., Амон Э.О. Радиолярии. М.: ПИН РАН, 2006. 320 с.
Афанасьева М.С., Кононова Л.И. О радиоляриях и конодонтах нижнего карбона Волго-Уральского бассейна (Мелекесская скв. 1) // Теоретические и прикладные аспекты палеонтологии. Матер. LXVII сесс. Палеонтол. об-ва при РАН. СПб.: Картфабрика ВСЕГЕИ, 2021. С. 9–10.
Назаров Б.Б. Радиолярии палеозоя. Л.: Недра, 1988. 232 с. (Практическое руководство по микрофауне СССР. Т. 2 / Ред. Жамойда А.И.).
Afanasieva M.S., Amon E.O., Agarkov Yu.V., Boltovskoy D.S. Radiolarians in the geological Record // Paleontol. J. 2005. V. 39. Suppl. 3. P. 135–392.
Afanasieva M.S., Kononova L.I. Early Tournaisian radiolarians and conodonts of the Volga-Ural Basin (Melekesskaya Borehole-1) // Kazan Golovkinsky Stratigr. Meeting, 2021. Sixth All-Russian Conf. “Upper Paleozoic of Russia” October 18–22, 2021, Kazan, Russia: collection of abstracts. P. 11.
Deflandre G. Sur le sens de développement centrifuge ou centripète des éléments de la coque des Radiolaires Sphaerellaires // C. R. Acad. Sci. 1964. V. 259. № 13. P. 2117–2119.
De Wever P., Dumitrica P., Caulet J.P. et al. Radiolarians in the sedimentary record. Amsterdam: Gordon and Breach Sci. Publ., 2001. 533 p.
Enriques P. Formazione e sviluppo dello scheletro siliceo nei radiolari // Boll. Sci. Ital. Biol. Sperimentale. 1931. V. 6. № 4. P. 350–356.
Feng Q., Gu S., He W., Jin Y. Latest Permian Entactinaria (Radiolaria) from southern Guangxi, China // J. Micropalaeontol. 2007. V. 26. № 1. P. 19–37.
Hollande A., Enjumet M. Cytologie, évolution et systématique des Sphaeroïdes (Radiolaires) // Arch. Muséum Nat. Hist. Natur. Sér. 7. 1960. № 7. 134 p.
Kling S.A. Radiolaria // Introduction to Marine Micropaleontology / Eds. Haq B.U., Boersma A. N.-Y.: Elsevier, 1978. P. 202–244.
Nazarov B.B., Ormiston A.R. A new Carboniferous radiolarian genus and its relation to the multishelled entactiniids // Micropaleontol. 1987. V. 33. № 1. P. 66–73.
Nestell G.P., Nestell M.K. Roadian (Earliest Guadalupian, Middle Permian) radiolarians from the Guadalupe Mountains, West Texas, USA. Part II: Spongy radiolarians (Entactinaria? and Spumellaria) // Micropaleontol. 2021. V. 67. № 6. P. 527–555.
Noble P., Aitchison J.C., Danelian T. et al. Taxonomy of Paleozoic radiolarian genera // Catalogue of Paleozoic radiolarian genera / Eds. Danelian T., Caridroit M., Noble P., Aitchison J.C. P., 2017. P. 419–502 (Geodiversitas. V. 39. № 3).
Noble P.J., Jin Y. Radiolarians from the Lamar Limestone, Guadalupe Mountains, West Texas // Micropaleontol. 2010. V. 56. № 1–2. P. 117–147.
Schwarz A. Über den Körperbau der Radiolarien. Ergebnisse paläontologischer Arbeitsmethoden // Abh. Senckenb. Naturforsch. Ges. 1931. Bd 43. № 1. S. 1–17.
Stürmer W. Das Wachstum silurischer Sphaerellarien und ihre späteren chemischen Umwandlung // Paläontol. Z. 1966. Bd 40. № 3/4. S. 257–261.
Won M.-Z. Radiolarien aus dem Unterkarbon des Rheinischen Schiefergebirges (Deutschland) // Palaeontogr. Abt. A. 1983. Bd 182. № 4–6. S. 116–175.
Won M.-Z. The proposed new radiolarian subfamily Retentactiinae (Entactiniidae) from the Late Devonian (Frasnian) Gogo formation, Australia // Micropaleontol. 1997. V. 43. № 4. P. 371–418.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал