

Палеонтологический журнал, 2021, № 5, стр. 49-57
Новый род Kislovodskoceras (Ammonoidea: Desmoceratidae) из среднего апта окрестностей Кисловодска (Северный Кавказ)
К. С. Полковой *
Саратовский национальный исследовательский государственный университет им. Н.Г. Чернышевского
Саратов, Россия
* E-mail: polkovoykirill@gmail.com
Поступила в редакцию 03.09.2020
После доработки 09.12.2020
Принята к публикации 10.12.2020
Аннотация
Из среднего апта окрестностей Кисловодска описаны аммониты нового рода Kislovodskoceras gen. nov., с типовым видом K. kislovodskense sp. nov. (сем. Desmoceratidae). Приведены данные об онтогенезе раковины и лопастной линии. Находки нового таксона приурочены к средней части зоны Epicheloniceras subnodosocostatum, для которой уточнено положение ее нижней границы в разрезах окрестностей Кисловодска.
ВВЕДЕНИЕ
Аммониты семейства Desmoceratidae Zittel, 1895 широко распространены в аптских отложениях Северного Кавказа, о них нередко упоминается в работах, посвященных стратиграфии региона (Ренгартен, 1926; Атлас…, 1949, 1960; Луппов, 1952; Мордвилко, 1960; Друщиц, Михайлова, 1966; Нижний мел…, 1985; Шарикадзе, 2015). Среди представителей этого семейства наиболее часто отмечались находки Zuercherella zuercheri (Jacob). Однако публикации, содержащие описания и изображения аптских десмоцератид, немногочисленны; они, в большинстве, посвящены лишь наиболее распространенным таксонам (Ренгартен, 1926; Рухадзе, 1938; Атлас…, 1949, 1960; Луппов, 1952; Эристави, 1961; Егоян, 1969). Desmoceratidae Северного Кавказа изучены гораздо хуже, чем представители других групп аммоноидей апта.
В зоне Epicheloniceras subnodosocostatum среднего апта окрестностей Кисловодска на Северном Кавказе обнаружены аммониты, принадлежащие к Desmoceratidae, но по своим морфологическим признакам (поперечное сечение оборотов, конфигурация периодических образований раковины, характер скульптуры и ее развития, лопастная линия) отличающиеся от всех известных аптских представителей рассматриваемого семейства, что послужило основанием для выделения новых рода и вида, описания которых приводятся ниже.
СТРАТИГРАФИЯ
Разрез нижнего мела в целом и апта в частности в окрестностях Кисловодска является классическим для Северного Кавказа, он привлекает внимание геологов с середины XIX в. (Мордвилко, 1960). Аптские отложения здесь достигают 300 м по мощности, литологически выдержаны и обладают богатыми комплексами ископаемой фауны, особенно двустворок и аммонитов. Аммоноидеи нижней зоны Epicheloniceras subnodosocostatum среднего апта характеризуются широким спектром таксономического разнообразия, представленным более чем десятью родами (Полковой, 2016; Полковой, Сельцер, 2018). Зона subnodosocostatum изучена автором в четырех обнажениях в окрестностях Кисловодска (рис. 1); по литологическому составу она подразделяется на три пачки (рис. 2). Нижняя (I) представлена алевролитами (в нижней части глинистыми, в верхней – песчаными) с прослоями известковых конкреций. Средняя пачка (II) представлена песчаниками; в их подошве располагается горизонт крупных караваеобразных известковых стяжений, в толще рассеяны многочисленные небольшие конкреции аналогичного состава. Верхняя пачка (III) представлена переслаиванием известняков-ракушечников и песчаников-ракушечников (мощностью 0.3–1.0 м) с песчаниками (2–11 м), заключающими редкие известковые конкреции. Общая мощность зоны превышает 70 м.
Рис. 1.
Расположение местонахождений (а – общая схема; б – детальная карта): 1 – Толстый бугор, 2 – Васюкова гора, 3 – Мыс Верхнего парка, 4 – “Красная гряда”.
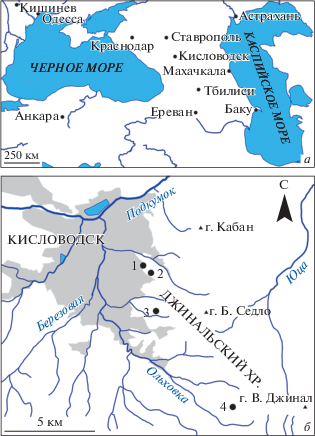
Рис. 2.
Разрез зоны Epicheloniceras subnodosocostatum среднего апта окрестностей Кисловодска (данные автора). Обозначения: 1 – песчаники, 2 – алевролиты, 3 – глины, 4 – песчаник-ракушечник известковый, 5 – известняк-ракушечник песчаный, 6 – известковые конкреции, 7 – интервал распространения Kislovodskoceras kislovodskense gen. et sp. nov. Сокращения: D. f. – Dufrenoyia furcata, P. m. – Parahoplites melchioris.
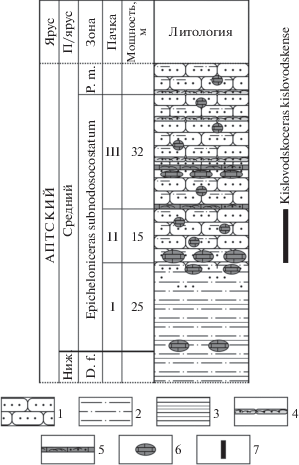
Нижняя граница зоны subnodosocostatum проводилась стратиграфами ранее в подошве пачки II (Мордвилко, 1960; Друщиц, Михайлова, 1966; Baraboshkin et al., 2003), при этом нижележащие алевролиты пачки I считались не охарактеризованными фауной и условно датировались нижним аптом. Вместе с тем, необходимо отметить, что аптский ярус до 1981 г. подразделялся на два подъяруса, но позже было принято его трехчленное деление; объем и границы прежнего верхнего апта по современным представлениям соответствуют среднему подъярусу (Постановления …, 1981, 1997; Михайлова и др., 2013). Встреченные в конкрециях в основании ранее считавшейся “немой” пачки I аммониты Epicheloniceras subnodosocostatum (Sinzow) (рис. 3, а–д), E. orientale (Jacob) (рис. 3, е, ж) и E. cf. tschernyschewi (Sinzow) свидетельствуют о присутствии зоны subnodosocostatum, что, по сравнению со стратификацией предыдущих исследователей, позволяет опустить нижнюю границу зоны (и среднего подъяруса апта) в окрестностях Кисловодска до указанного уровня (рис. 2).
Рис. 3.
Некоторые аммониты рода Epicheloniceras из нижней части зоны Epicheloniceras subnodosocostatum среднего апта: а–д – E. subnodosocostatum (Sinzow): а–в – экз. № 14/13355: а – сбоку, б – с вентральной стороны, в – с устья; г, д – экз. № 15/13355: г – сбоку, д – с вентральной стороны; е, ж – E. orientale (Jacob), экз. № 16/13355: е – сбоку, ж – с устья; Ставропольский край, окр. Кисловодска, “Красная гряда”.

Особенностью распространения Kislovodskoceras kislovodskense gen. et sp. nov. является приуроченность его находок исключительно к пачке II зоны subnodosocostatum в окрестностях Кисловодска (рис. 2). Этот факт может быть использован при детализации местной стратиграфической схемы.
МАТЕРИАЛ
Изученный материал послойно отобран автором в течение полевых сезонов 2009–2020 гг. из четырех местонахождений в окрестностях Кисловодска (рис. 1). Хорошая сохранность аммонитов позволила изучить изменения формы раковины, сечения оборотов, скульптуры, а также лопастной линии в онтогенезе, следуя известной методике (Михайлова, 1982, 1983; Шарикадзе, 2015). Начальная камера, форма сечения оборотов и лопастные линии зарисовывались при помощи бинокуляра МБС-10. Коллекция хранится в Центральном научно-исследовательском геологоразведочном музее им. Ф.Н. Чернышева (ЦНИГР музей), С.-Петербург, под № 13355.
ОБСУЖДЕНИЕ
Представители десмоцератид баррема, апта и альба нередко проявляют значительное морфологическое сходство, что может быть причиной неверных определений не только видов, но и родов, особенно при плохой сохранности материала и без точной стратиграфической привязки. В одних случаях этот факт объяснялся гомеоморфией, например, для Barremites Kilian, 1913 и Beudanticeras Hitzel, 1902 (Casey, 1961, с. 144), в других – происхождением от предполагаемого общего предкового таксона, например, для Zuercherella Casey, 1954 и Pseudohaploceras Hyatt, 1900 (Bogdanova, Hodemaeker, 2004, с. 242). Осложняют данную ситуацию обычно слабая скульптура и ограниченное разнообразие форм раковин у представителей рассматриваемого семейства. Учитывая вышеизложенное, существует необходимость сопоставления нового таксона с некоторыми по разным причинам похожими десмоцератидами.
Среди сопутствующих находкам нового таксона в окрестностях Кисловодска десмоцератид, прежде всего, стоит отметить Zuercherella zuercheri (Jacob) (табл. IX , фиг. 12–14 ; см. вклейку). В окрестностях Кисловодска этот вид встречается и выше по разрезу, вплоть до верхов среднего апта, а в целом на Кавказе он также известен из верхнего апта (Егоян, 1969). Раковины этого вида характеризуются субовальным сечением оборотов с закругленной вентральной стороной и обладают пережимами, начинающимися вблизи умбиликального перегиба и хорошо заметными на ядрах, а также слабо различимыми валиками. На поздней стадии появляются тонкие ребра, хорошо трассируемые на относительно крупных раковинах (табл. IX , фиг. 13 ). У всех изученных экземпляров этого вида наблюдалась симметричная трехраздельная первая умбиликальная лопасть в лопастной линии. Kislovodskoceras представляет иную группу десмоцератид, отличающуюся от Zuercherella zuercheri высокими субтрапецеидальными оборотами со слабоуплощенной и более узкой вентральной стороной, обособленной от боков, более резким перегибом между умбиликальной стенкой и боковой поверхностью, отсутствием валиков и пережимов в приумбиликальной части боковых сторон, а также отчетливых тонких ребер на поздней стадии. В лопастной линии нового таксона трехраздельная первая умбиликальная лопасть асимметрична.
Обращает на себя внимание идентичность формы поперечного сечения оборота на рисунке “Ammonites” impressus d’Orbigny в атласе (d’Orbigny, 1842, табл. 52 , фиг. 1–3) таковой у Kislovodskoceras kislovodskense. Однако на значительно позже представленной фотографии того же экземпляра (лектотипа) конфигурация сечения иная: более широкая и низкая, с заметно скругленным вентральным краем (Gauthier et al., 2006, табл. 32 , фиг. 1). Судя по всему, рисунок этого аммонита со стороны устья в работе А. д’Орбиньи некорректен. Б. Жоли (в: Gauthier et al., 2006) было обосновано отнесение вида “Ammonites” impressus к роду Zuercherella; исследователем отмечено, что лектотип представлен ядром с гладкой поверхностью без следов орнаментации, за исключением двух неглубоких концентрических борозд на боковых сторонах, наличие которых, по его мнению, является важнейшим отличительным признаком этого вида (Gauthier et al., 2006, с. 58–59). Таким образом, от Zuercherella impressa (d’Orbigny) новый таксон отличается менее широкими субтрапецеидальными оборотами с более обособленной и уплощенной вентральной стороной, наличием ясно выраженных валиков и пережимов, отсутствием концентрических борозд на боковых сторонах, а также более узким умбиликусом. Отличия в лопастной линии такие же, как и в приведенном выше сравнении с Z. zuercheri.
Из многочисленных представителей рода Beudanticeras большое сходство с выделенным таксоном проявляет B. revoili (Pervinquière). Прежде всего, Kislovodskoceras kislovodskense отличается от него менее объемлющими оборотами, более широким умбиликусом, более уплощенной, широкой и обособленной от боков вентральной стороной на поздних стадиях. Коллекция автора указанного вида (Pervinquière, 1907) подверглась ревизии Ж.-Л. Латилем (Latil, 2011), который изобразил как оригиналы B. revoili, так и несколько новых топотипов. Все эти экземпляры представлены ядрами со слабыми пережимами на вентральной стороне; боковые стороны практически гладкие. Пережимы на поздних оборотах ядер Kislovodskoceras kislovodskense более глубокие и хорошо выражены не только на вентральной стороне, но и в вентролатеральной области, где наблюдаются также очень слабые ребрышки-складки.
Среди представителей рода Pseudohaploceras (подсемейство Puzosiinae) по субтрапецеидальному поперечному сечению и морфологическим параметрам на средней стадии развития (Д ~ 30–40 мм) довольно похожим на новый таксон является P. gerhardti Bogdanova et Hodemaeker. Наш вид отличается от него более продолжительной стадией с подобным сечением, являющейся для него конечной; более уплощенной и обособленной от боков вентральной стороной; наличием валиков и пережимов только на вентральной стороне и в вентролатеральной области, и отсутствием хорошо различимых ребер между ними. Взрослые раковины P. gerhardti имеют бóльшие размеры, а также иные форму раковины и скульптуру.
ОПИСАНИЕ ТАКСОНОВ
НАДСЕМЕЙСТВО DESMOCERATOIDEA ZITTEL, 1895
СЕМЕЙСТВО DESMOCERATIDAE ZITTEL, 1895
ПОДСЕМЕЙСТВО BEUDANTICERATINAE BREISTROFFER, 1953
Род Kislovodskoceras Polkovoy, gen. nov.
Название рода от г. Кисловодска и keras греч. – рог; с.р.
Типовой вид – K. kislovodskense sp. nov.
Диагноз. Раковины 15–40 мм в диаметре, дисковидные, полуинволютные. Обороты средне объемлющие, высокие, субтрапецеидальные, с узкой и слабо уплощенной вентральной стороной, обособленной от боковых сторон. Умбиликус довольно узкий, ступенчатый, с невысокой и крутой умбиликальной стенкой, отделенной от боковой стороны резким перегибом. Раковины несут дугообразные валики, расположенные только на вентральной стороне и в привентральной части латеральных сторон; на ядрах перед ними или после них размещаются пережимы. Скульптура представлена серповидно изогнутыми линиями роста и низкими, расплывчатыми, трудно различимыми ребрышками-складками на боковых сторонах, которые с возрастом сглаживаются и исчезают. Лопастная линия в значительной степени рассеченная, с асимметричной трехраздельной глубокой первой умбиликальной лопастью и провисающей сутуральной лопастью.
Состав. Типовой вид.
Сравнение. От Zuercherella Casey, 1954 (табл. IX , фиг. 12–14 ; см. вклейку) отличается более высокими субтрапецеидальными оборотами со слабо уплощенной вентральной стороной, обособленной от более плоских боков; регулярностью и большей выраженностью валиков и пережимов, занимающих лишь вентральную сторону и привентральную часть латеральных сторон; отсутствием ясных, хорошо различимых тонких ребер; асимметричной трехраздельной первой умбиликальной лопастью в лопастной линии. Также отличительной чертой Kislovodskoceras является более раннее появление валиков с пережимами и ребрышек-складок, с дальнейшим сглаживанием последних. Представители Zuercherella до Д ~ 25–40 мм лишены характерного, исходя из диагноза автора рода (Casey, 1954, с. 112), орнамента тонких ребер, а также зачастую и валиков с пережимами (они появляются на более поздних оборотах); скульптура поверхности раковины представлена линиями и струйками роста (Jacob, Tobler, 1906; Егоян, 1969; Bogdanova, Hoedemaeker, 2004; Атлас…, 2005).
От Beudanticeras Hitzel, 1902 новый род отличается субтрапецеидальным поперечным сечением со слабо уплощенной вентральной стороной, обособленной от боковых сторон; менее объемлющими оборотами и иной конфигурацией валиков и пережимов, не простирающихся в приумбиликальную часть боковых сторон и характеризующихся регулярностью и хорошей выраженностью. В последнее время проводилась масштабная ревизия Beudanticeras, принадлежность многих ранее установленных видов к этому роду была подвергнута сомнению, в ряде случаев предложено отнесение их к другим известным родовым таксонам (Riccardi, Medina, 2002). В пересмотренном диагнозе указывается наличие узко закругленной вентральной стороны (Riccardi, Medina, 2002, c. 299), что для Kislovodskoceras не типично.
От Uhligella Jacob, 1907 новый род отличается более высокими субтрапецеидальными оборотами со слабо уплощенным вентральным краем, обособленным от менее выпуклых боковых сторон; регулярностью и хорошей выраженностью валиков и пережимов, не наблюдающихся в приумбиликальной части боковых сторон; отсутствием приумбиликальных вздутий и в целом менее рельефной поверхностью раковин.
Kislovodskoceras kislovodskense Polkovoy, sp. nov.
Табл. IX , фиг. 1–11
Рис. 4.
Начальная камера Kislovodskoceras kislovodskense gen. et sp. nov., экз. № 12/13355 (×30): а – сверху, б – со стороны перегородки, в – сбоку; Ставропольский край, окр. Кисловодска, Толстый бугор; средний апт, зона Epicheloniceras subnodosocostatum.
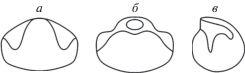
Рис. 5.
Изменение формы поперечного сечения оборотов Kislovodskoceras kislovodskense gen. et sp. nov.: а–з – экз. № 12/13355, и – экз. № 13/13355: а – 0.3 оборота (×13), б – 1.2 оборота (×13), в – 1.7 оборота (×13), г – 2.3 оборота (×9), д – 3.1 оборота (×6), е – 3.5 оборота (×5), ж – 3.9 оборота (×4), з – 4.5 оборота (×3); и – 4.9 оборота (×3); Ставропольский край, окр. Кисловодска, Толстый бугор; средний апт, зона Epicheloniceras subnodosocostatum.
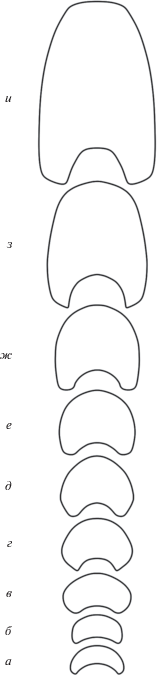
Рис. 6.
Изменение лопастной линии в онтогенезе раковины Kislovodskoceras kislovodskense gen. et sp. nov.: а–л – экз. № 12/13355, м – экз. № 13/13355: а–в – первая, вторая и пятая линии (×64), г – 1.2 оборота (×64), д – 1.7 оборота (×52), е – 2.3 оборота (×29), ж – 2.7 оборота (×26), з – 2.9 оборота (×22), и – 3.4 оборота (×17), к – 3.7 оборота (×13), л – начало 5-го оборота (×11); м – 4.7 оборота (×6); Ставропольский край, окр. Кисловодска, Толстый бугор; средний апт, зона Epicheloniceras subnodosocostatum.
Название вида от г. Кисловодска.
Голотип – ЦНИГР музей, № 1/13355; Ставропольский край, окрестности Кисловодска, Мыс Верхнего парка; средний апт, зона Epicheloniceras subnodosocostatum, в 6 м ниже кровли пачки II.
Форма. Начальная камера валикообразная, с размерами: диаметр 0.36 мм, длина 0.47 мм (рис. 4). Поперечное сечение первых двух оборотов изменяется от полулунного до широкого эллипсоидального (рис. 5, а–в), раковина эволютная. Позже высота оборота постепенно возрастает, при этом сечение сначала становится округлым, с заметно выпуклыми боками (рис. 5, г, д), затем эллипсоидальным, с последующим уплощением боковых сторон (рис. 5, е, ж). К концу четвертого оборота раковина становится полуинволютной (рис. 5, ж). В начале пятого оборота высота начинает преобладать над шириной (рис. 5, з), вентральная сторона сужается и уплощается. Взрослая раковина мелких или средних размеров (~4.8–6 оборотов и ~15–40 мм в Д), дисковидная, полуинволютная. Поперечное сечение оборота высокое, субтрапецеидальное, с плоскими боковыми сторонами, слабо расширяющимися по направлению к умбиликусу, и с узкой, слабо уплощенной вентральной стороной, которая ясно ограничена по краям слегка закругленными вентро-латеральными перегибами (рис. 5, и). Умбиликус довольно узкий, умеренно глубокий, ступенчатый, с невысокой и крутой умбиликальной стенкой, отделенной от боковой стороны резким перегибом. Объемлемость средняя: предыдущий оборот перекрывается на половину его высоты последующим.
Раковины несут регулярные широко расставленные валики, которые на ядрах ограничены сзади и/или спереди пережимами. И те, и другие появляются с последней трети четвертого оборота (Д ~ 5 мм), по мере роста раковины становясь более отчетливыми. Они размещены на вентральном крае и переходят на латеральные стороны, на которых по направлению от вентролатерального перегиба вниз быстро сглаживаются и исчезают, занимая только 1/4–1/3 высоты оборота в привентральной части. Контуры валиков с пережимами повторяют абрисы линий роста. Валики являются самыми высокими образованиями относительно поверхности раковины, особенно на вентральной стороне. У раковин с Д = 20 мм и более насчитывается 5–7 валиков на полуоборот.
Устье наблюдалось фрагментарно у двух экземпляров (№№ 5/13355 и 11/13355). Оно сбоку S‑образное, с широкой округленной и выпуклой вперед частью, максимально выступающей ниже середины (по направлению к умбиликусу) латеральной стороны, и круто загибающееся назад в нижней (приумбиликальной) четверти боков. Вентральная часть устья параболой слабо выдается вперед. Длина жилой камеры у аммонитов с сохранившимся устьем составляет около 3/4 оборота.
Скульптура. Начальные 3.5 оборота гладкие, покрытые лишь тончайшими линиями роста. Последние позже становятся более грубыми, нередко приобретая вид струек, наклонены в сторону устья, узкие S-образные на боковых сторонах и образующие слабый выступ вперед на вентральной стороне. Со стадии 4.8–5 оборота (Д ~ ~ 15–18 мм) и вплоть до устья линии и струйки роста узко-серповидные. В нижней (приумбиликальной) трети латеральной стороны они прямолинейно направлены от умбиликального края с наклоном вперед, затем отгибаются назад под тупым углом, после чего выше, вплоть до вентролатерального перегиба, образуют неширокий синус, выпуклостью направленный от устья. Пересекая вентральную сторону, линии и струйки образуют выступ вперед в виде притупленной параболы.
На ядрах аммонитов обнаруживается рельеф, который состоит из низких, расплывчатых и слабо различимых ребрышек-складок на боковых сторонах, по форме повторяющих струйки роста и разделенных неглубокими депрессиями. Они появляются у умбиликального перегиба в начале пятого оборота (Д ~ 7 мм) и в течение последующих 0.5–0.7 оборота наиболее заметны в приумбиликальной части латеральной стороны, нивелируясь и изредка расщепляясь по направлению к вентральной стороне, редко расставлены; ширина ребер в приумбиликальной части в два–три раза больше ширины депрессий. Позже ребрышки занимают всю боковую поверхность, расширяются или нередко расщепляются в пределах ее середины на две или три ветви по направлению к вентральному краю, и в целом становятся более узкими и частыми. С этой стадии они расставлены довольно беспорядочно, а толщина ребрышек и разделяющих их промежутков непостоянна. Примерно в начале шестого оборота (Д ~ 18 мм) и до устья ребрышки выражены только в привентральной части боковых сторон, постепенно сглаживаясь с ростом раковины вплоть до полного исчезновения. На вентральном крае подобный рельеф не наблюдается. На поверхности раковины, в отличие от ядер, ребрышки и разделяющие их депрессии заметны очень слабо, особенно на поздних стадиях.
Размеры в мм и отношения:
| Экз. № | Д | Ду | В | Ш | Ду/Д | В/Д | Ш/Д | Ш/В |
|---|---|---|---|---|---|---|---|---|
| 12/13355 3.0 оборота |
3.4 | 1.1 | 1.4 | 1.7 | 0.32 | 0.41 | 0.50 | 1.21 |
| 4.0 оборота | 6.9 | 1.9 | 3.1 | 3.1 | 0.28 | 0.45 | 0.45 | 1.00 |
| 4.7 оборота | 13.1 | 2.9 | 6.4 | 5.0 | 0.22 | 0.49 | 0.38 | 0.78 |
| 6/13355 | 15.7 | 3.1 | 7.4 | 5.2 | 0.20 | 0.47 | 0.33 | 0.70 |
| 13/13355 | 17.2 | 3.8 | 8.5 | 5.4 | 0.22 | 0.49 | 0.31 | 0.64 |
| 9/13355 | 17.8 | 3.5 | 8.8 | 6.2 | 0.20 | 0.49 | 0.35 | 0.70 |
| 4/13355 | 18.0 | 3.5 | 9.0 | 6.2 | 0.19 | 0.50 | 0.34 | 0.69 |
| 5/13355 | 19.6 | 3.7 | 9.9 | 6.2 | 0.19 | 0.51 | 0.32 | 0.63 |
| 3/13355 | 20.2 | 4.5 | 9.4 | 6.9 | 0.22 | 0.47 | 0.34 | 0.73 |
| 7/13355 | 21.3 | 4.4 | 10.5 | 6.8 | 0.21 | 0.49 | 0.32 | 0.65 |
| 10/13355 | 23.5 | 4.5 | 12.2 | 7.6 | 0.19 | 0.52 | 0.32 | 0.62 |
| Голотип 1/13355 | 25.2 | 5.4 | 12.4 | 7.8 | 0.21 | 0.49 | 0.31 | 0.63 |
| 8/13355 | 25.6 | 5.4 | 12.6 | 7.7 | 0.21 | 0.49 | 0.30 | 0.61 |
| 2/13355 | 38.0 | 7.6 | 18.9 | 11.5 | 0.20 | 0.50 | 0.30 | 0.61 |
| 11/13355 | 39.3 | 8.8 | 19.4 | 12.4 | 0.22 | 0.49 | 0.32 | 0.64 |
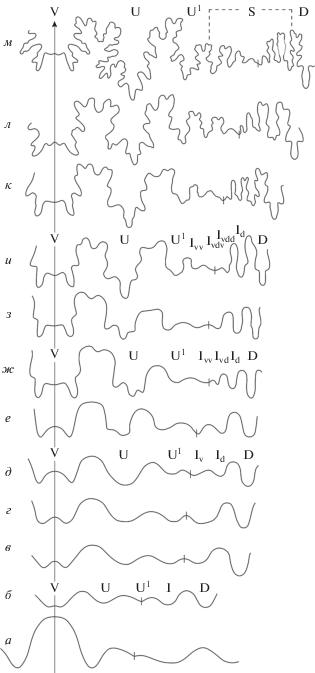
Лопастная линия11 (рис. 6). Просутура трехлопастная, с высоким и умеренно узким вентральным седлом (рис. 6, а). Примасутура пятилопастная, с двураздельной вентральной лопастью. Вторая умбиликальная лопасть наиболее мелкая, рассекается швом посередине (рис. 6, б). На пятой линии она смещается на боковую сторону (рис. 6, в). На стадии 1.2 оборота интернилатеральная лопасть несколько расширяется (рис. 6, г), и примерно в середине второго оборота она почти симметрично делится: I → IvId (рис. 6, д). В начале третьего оборота обособляются ветви интернилатеральной лопасти: внутренняя смещается к дорсальной, внешняя, углубляясь, – к умбиликальному шву; также расширяется седло U1/Iv (рис. 6, е). В интервале 1.7–2.7 оборота асимметрично подразделяется на три ветви первая умбиликальная лопасть, сначала расширяясь и углубляясь (рис. 6, д–ж). На стадии 2.7 оборота внешняя из образованных лопастей также претерпевает деление: Iv → IvvIvd (рис. 6, ж). В дальнейшем в результате аналогичного многократного последовательного расчленения расположенных в пришовной зоне новообразованных ветвей интернилатеральной лопасти формируется провисающая сутуральная лопасть (рис. 6, з–м). В общей сложности на протяжении наблюдавшихся 4.5 оборотов происходит шесть делений интернилатеральной лопасти, вследствие чего возникают семь новых лопастей. На стадии 4.5 оборота лопастная линия в значительной степени рассеченная, с асимметричной трехраздельной глубокой первой умбиликальной лопастью и глубокой дорсальной (рис. 6, м). Формула лопастной линии представляется в виде (полужирным выделены лопасти, претерпевающие деление):
VUU1:ID → VUU1:IvIdD → VUU1Ivv:IvdIdD → →VUU1IvvIvdv:IvddIdD → VUU1IvvIvdvv:IvdvdIvddIdD → → VUU1IvvIvdvvv:IvdvvdIvdvdIvddIdD → → VUU1IvvIvdvvvIvdvvdv:IvdvvddIvdvdIvddIdD.
К настоящему времени лишь у немногих видов Desmoceratidae изучено изменение лопастной линии в онтогенезе, причем среди них нет ни одного представителя Beudanticeratinae. Среди аптских представителей семейства лопастная линия исследована только у Pseudohaploceras douvillei (Fallot) (Шарикадзе, 2015)22, среди альбских – у Puzosia mayoriana (Orbigny) (Михайлова, 1983); оба аммонита относят к подсемейству Puzosiinae. Характер последовательности появления, места заложения новых элементов и их рассеченности в линиях этих таксонов во многом идентичен таковому у Kislovodskoceras kislovodskense. Он также протекает с первым симметричным делением интернилатеральной лопасти и с образованием за счет дальнейшего ее расчленения провисающей сутуральной лопасти. Изменение лопастной линии в онтогенезе нового вида отличается от такового у вышеуказанных формированием асимметричной трехраздельной первой умбиликальной лопасти и несколько более поздним первым делением интернилатеральной лопасти.
Материал. Кроме голотипа, 20 экз. из среднего апта окрестностей Кисловодска (зона Epicheloniceras subnodosocostatum).
Список литературы
Атлас нижнемеловой фауны Северного Кавказа и Крыма. М.: Гостоптехиздат, 1960. 702 с.
Атлас раннемеловой фауны Грузии. Тбилиси, 2005. 788 с. (Тр. Геол. ин-та АН Грузии. Нов. cер. Вып. 120).
Атлас руководящих форм ископаемых фаун СССР. Т. 10. Нижний отдел меловой системы. М.: Госгеолиздат, 1949. 328 с.
Безносов Н.В., Михайлова И.А. Высшие таксоны юрских и меловых Ammonitida // Палеонтол. журн. 1991. № 4. С. 3–18.
Друщиц В.В., Михайлова И.А. Биостратиграфия нижнего мела Северного Кавказа. М.: МГУ, 1966. 189 с.
Егоян В.Л. Аммониты из клансейских слоев Западного Кавказа // Геология и нефтегазоносность Западного Кавказа и Западного Предкавказья. М.: Недра, 1969. С. 126–188 (Тр. Краснодарского фил-ла ВНИИНефть. Вып. 19).
Луппов Н.П. Нижнемеловые отложения Северо-Западного Кавказа и их фауна. Л.–М.: Гостоптехиздат, 1952. 270 с. (Тр. ВНИГРИ. Нов. сер. Вып. 65).
Михайлова И.А. О малоизвестном роде аммонитов из клансейского горизонта (нижний мел) // Палеонтол. журн. 1972. № 3. С. 78–86.
Михайлова И.А. Методика онтогенетического изучения аммоноидей // Бюлл. МОИП. Отд. геол. 1982. Т. 57. № 3. С. 107–113.
Михайлова И.А. Система и филогения меловых аммоноидей. М.: Наука, 1983. 280 с.
Михайлова И.А., Богданова Т.Н., Барабошкин Е.Ю. Аптский ярус (подъярусное и зональное деление, границы ярусов и подъярусов) // Общая стратиграфическая шкала России: состояние и проблемы обустройства. Матер. Всеросс. совещ. (23–25 мая 2013 г., Москва). М.: ГИН РАН, 2013. С. 304–306.
Мордвилко Т.А. Нижнемеловые отложения Северного Кавказа и Предкавказья. Часть 1. М.–Л.: АН СССР, 1960. 239 с.
Нижний мел юга СССР. М.: Наука, 1985. 224 с.
Полковой К.С. Аммоноидеи нижней зоны среднего апта окрестностей г. Кисловодска // Проблемы геологии и освоения недр. Тр. XX междунар. симп. им. акад. М.А. Усова студентов и молодых ученых. Т. I. Томск: ТПУ, 2016. С. 78–80.
Полковой К.С., Сельцер В.Б. О первой находке представителя рода Helicancyloceras Klinger et Kennedy, 1977 (Ammonoidea) в среднем апте Кавказа // Фундаментальная и прикладная палеонтология. Матер. LXIV сессии Палеонтол. об-ва при РАН. СПб.: ВСЕГЕИ, 2018. С. 92–94.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 19. Л.: ВСЕГЕИ, 1981. 89 с.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 29. СПб.: ВСЕГЕИ, 1997. 35 с.
Ренгартен В.П. Фауна меловых отложений Ассинско-Камбилеевского района // Тр. Геол. ком. Нов. сер. 1926. Вып. 147. 132 с.
Рухадзе И.М. Аптские аммониты Северного Кавказа // Бюлл. Геол. ин-та Грузии. 1938. Т. 4. Вып. 2. С. 113–209.
Шарикадзе М.З. Аптские аммониты Кавказа. Тбилиси: Универсал, 2015. 321 с.
Эристави М.С. Аммониты апта и альба Северного Кавказа // Тр. Геол. ин-та АН Груз. ССР. Сер. геол. 1961. Т. 12(17). С. 41–77.
Baraboshkin E.Yu., Alekseev A.S., Kopaevich L.F. Cretaceous paleogeography of the North-Eastern Peri-Tethys // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2003. V. 196. P. 177–208.
Bogdanova T.N., Hoedemaeker P.J. Barremian-Early Albian Deshayesitidae, Oppeliidae, Desmoceratidae and Silesitidae of Colombia // Scripta Geol. 2004. V. 128. P. 183–312.
Casey R. New genera and subgenera of Lower Cretaceous ammonites // J. Wash. Acad. Sci. 1954. V. 44. № 4. P. 106–115.
Casey R. A monograph of the Ammonoidea of the Lower Greensand. Pt III // Monogr. Palaeontogr. Soc. 1961. P. 119–216.
Gauthier H., Busnardo R., Combemorel R. et al. Révision critique de la Paléontologie française d’Alcide d’Orbigny. V. IV. Céphalopodes crétacés. Premiere Partie. Leiden: Backhuys Publ., 2006. 292 p.
Jacob C., Tobler A. Étude stratigraphique et paléontologique du Gault de la Vallée de la Engelberger Aa (Alpes calcaires suisses, environs du Lac des Quatre Cantons) // Mém. Soc. Paléontol. Suisse. 1906. V. 33. 26 p.
Latil J.-L. Early Albian ammonites from Central Tunisia and adjacent areas of Algeria // Rev. Paléobiol. 2011. V. 30. № 1. P. 321–429.
Orbigny A. d'. Paléontologie française. Description zoologique et géologique de tous les animaux mollusqes et rayonnés fossiles de France. Terrains crétacés. T. 1. Céphalopodes. P.: Masson, 1840–1842. 662 p.
Pervinquière L. Études de paléontologie tunisienne. I. Céphalopodes des terrains secondaires // Mém. Carte Géol. Tunisie. 1907. 438 p.
Riccardi A.C., Medina F.A. The Beudanticeratinae and Cleoniceratinae (Ammonitida) from the Lower Albian of Patagonia // Rev. Paléobiol. 2002. V. 21. № 1. P. 291–351.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал