

Палеонтологический журнал, 2021, № 5, стр. 3-9
Псевдоколонии силурийских ругоз Microplasma schmidti Dybowski из Эстонии: морфология и происхождение
Е. С. Казанцева *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: kazantseva@paleo.ru
Поступила в редакцию 18.11.2020
После доработки 10.12.2020
Принята к публикации 05.02.2021
Аннотация
Кратко описан Microplasma schmidti Dybowski, 1874 (Rugosa: Stauriida: Cystiphyllidae). Вид характеризуется примитивным септальным аппаратом, диссепиментами, занимающими центральную зону кораллитов, и частой встречаемостью псевдоколоний. На материале из коллекции Таллинского технического университета из венлока Эстонии (разрез Сепайс, региоярус Джаагараху) описан механизм формирования псевдоколоний путем регенеративного почкования материнского кораллита и рассмотрены основные причины их возникновения.
ВВЕДЕНИЕ
В рамках проекта по изучению коллекции ругоз Таллинского технического ун-та (ТТУ) изучен материал, представленный Microplasma schmidti Dybowski, 1874 из местонахождения Сепайс (Эстония, побережье Балтийского моря).
Род Microplasma Dybowski, 1873 представлен одиночными и колониальными кораллитами с многочисленными диссепиментами, заполняющими срединную зону. Он входит в состав семейства Cystiphyllidae Milne-Edwards et Haime, 1850, представленного одиночными и колониальными ругозами с широким и сильно развитым диссепиментарием. В свою очередь, семейство было выделено в составе обширного отряда Stauriida, описанного А. Верриллом (Verrill, 1865) и впервые присоединенного к ругозам в “Treatise…” (Hill, 1981). Отряд объединяет большое число палеозойских (преимущественно пермских и каменноугольных) семейств с толстой эпитекой и септами двух порядков. В некоторых классификациях отряд упразднен (Scrutton, 1997).
Особенность вида Microplasma schmidti Dybowski, 1874 – частая встречаемость псевдоколониальности и сростков, образованных путем разрастания эпитек нескольких кораллитов. Их происхождение и особенности морфогенеза, а также сопровождающие их другие морфогенетические процессы были детально изучены при исследовании тонких шлифов и пришлифовок. Обсуждению полученных новых данных посвящена настоящая статья. Описание многочисленных псевдоколоний и ложных псевдоколоний у ругоз M. schmidti предваряется кратким описанием типичных экземпляров.
МАТЕРИАЛ И МЕСТОНАХОЖДЕНИЕ
Изучена часть коллекции ругоз Таллинского технического ун-та, представленная многочисленными экземплярами Microplasma schmidti, собранными на местонахождении Сепайс (региоярус Джаагараху, силур, венлок) на северо-западном берегу о. Сааремаа Балтийского моря (рис. 1). Свита представлена последовательной сменой нескольких толщ (по: Nestor et al., 2001). В нижней части свиты с мощностью до 8 м залегает светло-серый, массивный, пятнистый известковый доломит с обильным количеством табулят. В средней части маломощная (около 1 м) порода, пористая, с реликтовым биомикритическим строением. Базальная часть мощностью 3–4 м содержит многочисленные скелеты табулят и строматопороидей и сменяется на продолжительную толщу (до 7 м) доломитового известняка средней или большой толщины, состоящего из нескольких осадочных циклитов. Нижняя часть представлена мелкозернистыми, часто биотурбированными, глинистыми вкраплениями с небольшим количеством пирита. Верхняя часть – известняками от изменчивого до хорошо отсортированного, пиритизированного, мелкозернистого. Скелетный материал состоит в основном из фрагментов кораллов, строматопороидей и двустворчатых моллюсков. Исследуемый материал был отобран из этого интервала. Выше залегают биомикритные известняки с волнистыми прослойками (толщиной 1–5 см) и более мощными слоями (до 50 см) известнякового или глинистого мергеля. В породе содержатся пиритизированные скелетные частицы пельматозойных иглокожих, брахиопод, трилобитов; норы и спорадически пиритизированные литокласты. Общая мощность пачки до 18 м. Она заканчивается продолжительным (21.3 м) чередованием циклов верхних слоев известняков и мергелевых пластов. Характерны норы, пиритизированные по периферии. Кровля свиты представлена биомикритовыми известняками с тонкими прослоями мергеля. Этот интервал является самым известковым в формации и протягивается примерно на 7 м.
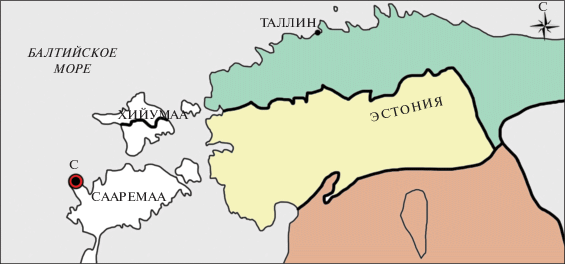
Рис. 1.
Географическое положение изученного материала, показанное на фрагменте карты Эстонии. С – разрез Сепайс. Цветами обозначены отложения: зеленый – ордовикские, желтый – силурийские, коричневый – девонские.
Имеющиеся в коллекции экземпляры отличаются очень хорошей сохранностью скелетов. Материал представлен кораллитами разных стадий развития, преимущественно взрослыми формами, одиночными или псевдоколониальными.
МЕТОДИКА
Хорошая сохранность большинства кораллов позволила успешно применить классическую методику шлифования. Экземпляры фотографировались камерой Nikon D800 с объективом 60 мм сериями с изменением резкости на одном объекте. Полученные серии фотографий были скомпонованы в программе Helicon Focus 6.2.2 и преобразованы в рабочий формат изображений. Шлифы делались сериями из нескольких поперечных и одного продольного. Продольные шлифы проводились строго перпендикулярно осевой части скелета коралла по линии наименьшей кривизны (Сошкина, 1952). Шлифование образцов производилось в несколько этапов. Вначале делались распилы с помощью закрепленного вертикально распиловочного диска. Далее поверхность полировалась на полировочных дисках на аппарате шлифования с помощью двух порошков разной гранулярной размерности. Окончательная полировка поверхности происходила на стекле с применением наиболее тонкого порошка. Готовая пришлифовка подвергалась сушке с помощью грелки Struers и крепилась с помощью растопленного канадского бальзама к предварительно заматированному стеклу. После окончательного затвердевания бальзама не прикрепленная к стеклу часть спила стачивалась на диске до удовлетворительной толщины и окончательно полировалась на стекле.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД STAURIIDA
ПОДОТРЯД CYSTIPHYLLINA
СЕМЕЙСТВО CYSTIPHYLLIDAE MILNE-EDWARDS ET HAIME, 1850
Род Microplasma Dybowski, 1873
Microplasma: Dybowski, 1873, с. 340; McLean, 1976, с. 10.
Pseudomicroplasma: Сошкина, 1949, с. 53.
Cystiplasma: Taylor, 1951, с. 195.
Nardoplasma: Спасский и др., 1974, с. 171.
Типовой вид – Microplasma gotlandicum Dybowski, 1874, по определению Р. Ведекинда (Wedekind, 1927); силур, готланд Швейцарии.
Диагноз. Узкие цилиндрические кораллы, формирующие фацеллоидные колонии или одиночные. Продольный срез заполнен двумя, тремя или четырьмя рядами диссепиментов или днищами (Merriam, 1973).
Состав. Кроме типового, еще 13 видов: M. caespitosum (Schluter, 1882), M. devonica Soshkina, 1937, M. fasciculatum Swartz, 1913, M. flabelliforme Wang, 1948, M. fongi Yoh, 1937, M. fractum Schluter, 1882, M. gotlandica Dybowski, 1874, M. hadron McLean, 1976, M. lovenianum Dybowski, 1874, M. orientalis Ivanovsky, 1974, M. parallelum Etheridge, 1899, M. ronense Mansuy, 1913, M. schlueteri Wedekind, 1922 из силура–девона Урала, Таймыра, США и Балтики.
Сравнение. Сходен с родом Cystiphyllum Lonsdale, 1839 внутренним строением диссепиментов и днищ, но отличен отсутствием на них септальных шипов.
Замечания. В работе Р. МакЛина (McLean, 1976) предлагается объединить большое количество девонских представителей родов Pseudomicroplasma Soshkina, 1949 и Cystiphyllum с родом Microplasma, однако указаны не все необходимые основания для объединения родов.
Microplasma schmidti Dybowski, 1874
Табл. I , фиг. 1–5 (см. вклейку)
Рис. 2.
Взаимное существование кораллитов, где: а – ложная псевдоколония, б – псевдоколония, в – истинная колония. Обозначения: пс1-4 – плоскости симметрии кораллитов согласно порядку их появления; псм – плоскость симетрии материнского кораллита.
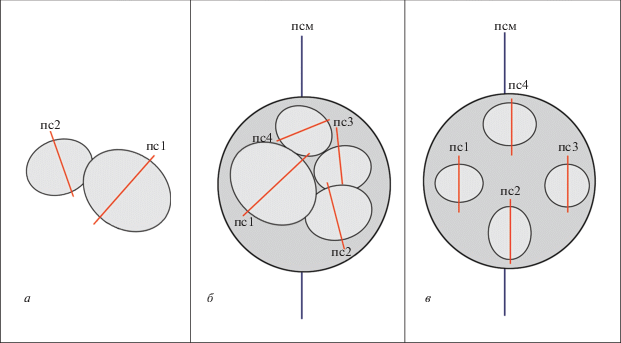
Рис. 3.
Microplasma schmidti Dybowski, 1874, экз. ТТУ, № GIT 397-2019: а, б – поперечные срезы; Эстония, о-в Сааремаа, местонахождение Сепайс; силур, венлок, гомерский и шейнвудский яруса, региоярус Джаагараху.

Рис. 4.
Microplasma schmidti Dybowski, 1874, экз. ТТУ, № GIT 397-1978: а – поперечный срез, б – схема взаимного расположения почек (п1-7) и материнского кораллита (мк); Эстония, о-в Сааремаа, местонахождение Сепайс; силур, венлок, гомерский и шейнвудский яруса, региоярус Джаагараху.
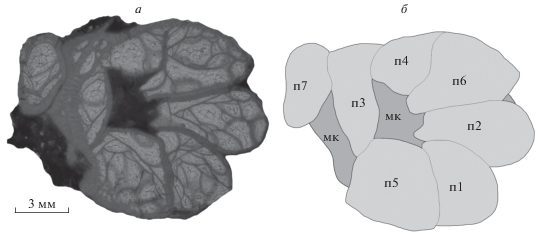
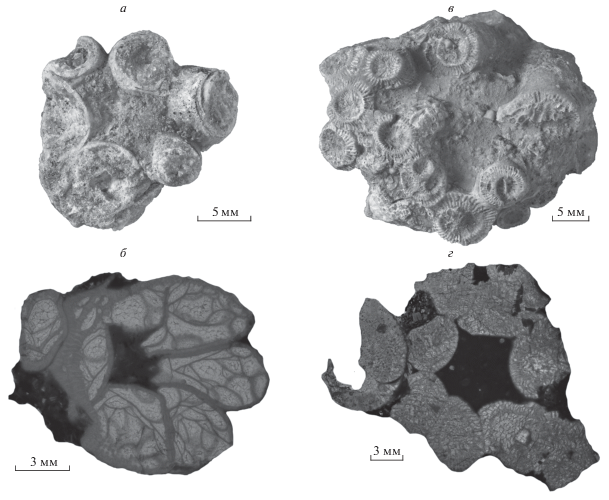
Рис. 5.
Силурийские ругозы: а, б – Microplasma schmidti Dybowski, 1874, экз. ТТУ, № GIT 397-1978: а – общий вид, б – поперечный срез; в, г – Acervularia ananas (Linnaeus, 1758), экз. ТТУ, № GIT 397-1969: в – общий вид, г – поперечный срез; Эстония, о-в Сааремаа, местонахождение Сепайс; венлок, гомерский и шейнвудский яруса, региоярус Джаагараху.
Microplasma schmidti: Dybowski, 1874, с. 97, табл. 5-3 , фиг. 3a-b; Волкова, Латыпов, 1979, с. 20, табл. 10 , фиг. 1a, b; Johannessen, 1874, с. 9.
Microplasma lovenianum: White, 1966, с. 149, табл. 22-3 , фиг. 11 .
Лектотип – экз., изображенный в: Dybowski, 1874, табл. 5-3 , фиг. 3a-b (место хранения неизвестно); о-в Готланд; верхний силур.
Описание (рис. 3; 4; 5, а, б). Одиночные и одиночно-ветвящиеся, многочисленно-почкующиеся кораллы с короткими иглообразными шипиками септ, беспорядочно отходящими от стенки и поверхности диссепиментов. Кораллиты цилиндрические, с неглубокой чашкой. Тонкая ребристая эпитека. Септальный аппарат представлен короткими, тонкими, иглообразными шипиками, реже отходящими от внешней стенки, чаще – от поверхности диссепиментов. Днища крупные и пологие, представлены сильно вогнутыми пластинами, примерно одинаковыми по длине, часто наблюдаются в центральной зоне кораллитов и на поперечных срезах. Диссепименты – вздутые, крупные пузыри, занимающие центральную зону кораллитов и ориентированные радиально вдоль внешней стенки.
Сравнение. Строение скелетных элементов сходно с таковым M. gotlandica Dybowski, 1874, но обилие тонких шипиков септ на диссепиментах свойственно только M. schmidti.
Замечания. Для вида характерно большое количество эпитекальных выростов, регенеративных почек, эпитекальных перешейков, способствующих образованию сложных агрегатов (сростков). Статистически нередки псевдоколонии.
Распространение. Венлок Готланда и Балтики, нижний девон Якутии.
Материал. 31 экз. из разреза Сепайс (о. Сааремаа, Эстония), региярус Джаагараху (силур, венлок, гомерский и шейнвудский яруса).
ИСПОЛЬЗУЕМАЯ ТЕРМИНОЛОГИЯ
Для удобства дальнейшего описания встречаемых у ругоз процессов следующие термины требуют уточнения.
Зачастую встречающиеся у ругоз сростки, образованные в результате разрастания эпитеки, разделены на сростки конкурирующие (где наблюдается подавление одного кораллита другим, или доминирующим в сростке, что часто сопровождается эпиморфозом в зоне повреждения кораллитов, образованной их соприкосновением; и сростки взаимовыгодные, образованные в результате намеренного и взаимно благоприятного срастания ругоз путем образования эпитекальных выростов и перешейков. Зачастую такие сростки визуально сходны с колониями и псевдоколониями, но отличаются в обоих случаях происхождением кораллитов в результате полового размножения. Предполагается отнесение этих сростков к рангу ложных псевдоколоний (рис. 2, а), где кораллиты растут рядом и создают видимость возникновения в результате почкования, но расположены автономно и образованы в результате близкого оседания личинок одного вида кораллов.
Настоящая колония (рис. 2, в), которую порой довольно сложно отличить на ископаемом материале от сростка и множественно-почкующегося индивида, должна рассматриваться как результат генетически запрограммированного процесса бесполого размножения, идущего по определенной схеме (истинная колония). Ложные колонии, в свою очередь, развиваются неупорядоченно, их образование чаще происходило в результате повреждений и последующей регенерации скелета (Кузьмичева, 1988). Такие ложные колонии описаны как псевдоколонии (рис. 2, б) – кластеры индивидуальных кораллитов одного вида, растущих рядом и возникших в результате регенеративного почкования одиночного материнского кораллита (по: Fedorowski, 1971, 1978).
Возникающие в результате вегетативного почкования почки могут быть разделены на несколько основных типов, в соответствии с положением плоскостей их симметрии (проходящих через главную и противоположную септы) относительно таковой у материнского кораллита. Отличаются от вегетативной почки, характерной для колониальных ругоз, способом возникновения и дальнейшим развитием: дубликатная – почка, плоскость симметрии которой параллельна плоскости материнского кораллита; реверсивная – почка, плоскость симметрии которой перпендикулярна таковой у материнского кораллита; сегментная – почка, плоскость симметрии которой параллельна наиболее выраженным септам материнского кораллита в сегменте, где она была сформирована. В некоторых случаях встречаются реверсивные сегментные почки, у которых плоскость симметрии перпендикулярна септам, на которых они образованы (Казанцева, 2019).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Множественное почкование нередко встречается у Microplasma schmidti и выражено в частом формировании регенеративных почек в чашечке. О регенеративном происхождении почек свидетельствует их хаотичное расположение друг относительно друга, взаимная конкуренция и предшествующая строительству почек частичная гибель материнского кораллита, где поврежденная часть обособляется стенкой, как это было, например, описано у Bothrophyllum conicum Trd. из карбона (Rozhnov, 2014; Kazantseva, Rozhnov, 2018). Септальный аппарат у Microplasma schmidti примитивный и выражен в небольших шипиках на поверхности диссепиментов, поэтому изучение плоскостей симметрии недоступно, и определение типа регенеративной почки (дубликатная, реверсивная, сегментная) может быть неточным.
Формирование многократного почкования у M. schmidti зачастую сопровождается срастанием эпитеки автономных кораллитов, где сильно развитые эпитекальные перешейки создают видимость соединительной структуры, но на самом деле являются сплошными отростками без соединительной ткани. Регенеративное почкование путем морфаллаксиса у этого вида начинается с обособления стенки вдоль ряда наиболее полных диссепиментов. Разделенные пополам, они начинают ориентироваться вдоль поверхности эпитеки до формирования упорядоченной структуры (рис. 3). О высокой роли материнского кораллита в росте почек свидетельствует частично сохранившаяся ориентированность диссепиментов материнского кораллита на ранних стадиях роста почек. На более поздних стадиях роста почек развивается ориентированность диссепиментов по окружности к центру, наблюдаются короткие шипы септ, а стенка между кораллитами утолщается, и они обособляются друг от друга. У каждой из почек формируется собственная стенка, и дальнейший рост почек направлен на полное обособление друг от друга. В случаях, когда кораллит формирует всего одну почку, на более взрослых стадиях роста она начинает обособляться и развивается автономно от материнского кораллита, зачастую обуславливая его отклонение в сторону. Такие почки, образованные одновременно в чашке материнского кораллита, при параллельном росте существуют в условиях взаимной конкуренции и сильно деформируются из-за подавления более развитыми и крупными почками. В зоне подавления стенка зачастую расширена, форма и положение диссепиментов соответствует повреждению и отклоняется от нормального. В редких случаях больших размеров достигает только половина из новообразованных почек (рис. 4), чаще всего – одна или две.
Ругозы вида M. schmidti (рис. 5, а, б) встречаются в разрезе преимущественно в одиночной модификации. В то же время, на такую статистику может повлиять частое отнесение псевдоколониальных представителей вида к Acervularia ananas (Linnaeus, 1758) (рис. 5, в, г). Множественные псевдоколонии с частыми регенеративными почками в чашке при отсутствии изготовленных шлифов визуально схожи с колониальными A. ananas, характерным колониальным представителем ругоз венлока, в частности, в разрезе Сепайс. Наибольшей разницей между этими двумя видами можно обозначить истинную колониальность A. ananas, наличие у вида внутренней стенки, а также простую и хаотичную схему строения септального аппарата у Microplasma schmidti.
Несмотря на очевидное отнесение многих представителей M. schmidti к псевдоколониям, в некоторых случаях среди них встречаются сростки двух и более индивидуальных одиночных или псевдоколониальных одиночных кораллитов, образованных половым путем и растущих рядом. На некоторых стадиях их эпитека сливается полностью, периодически – с образованием эпитекальных перешейков; на более поздних стадиях кораллиты расходятся и не соприкасаются друг с другом. В данном случае наблюдается формирование ложных псевдоколоний, определение которых основано на половом происхождении поселившихся рядом кораллитов.
Важным вопросом в понимании процессов формирования псевдоколоний и ложных псевдоколоний является установление причин их возникновения. Любые регенерационные процессы у ругоз возникали в связи с неблагоприятной средой обитания, и зачастую это связано с грунтом или ограничением в пространстве для расселения. Второй наиболее распространенной причиной формирования псевдоколоний может служить резкая смена условий обитания, для которой характерно массовое стремление к формированию новых особей или омоложению взрослых кораллитов. На это указывает большое количество ювенильных особей, встреченных совместно с псевдоколониями одиночных ругоз, или увеличение числа утерянных (недоразвитых) структур у колониальных (Fedorowski, 1978) как результат массового вегетативного почкования с целью спасения колонии и увеличения шансов на выживаемость.
Третьей, вероятно, ключевой причиной возникновения псевдоколониальности становится склонность определенных видов к формированию регенеративных почек. Однозначное отнесение таких таксонов к протоколониальным [промежуточной форме роста между одиночным существованием и колониальным (Fedorowski, Ogar, 2013)] невозможно, поскольку далеко не все представители вида или рода соответствуют тому или иному типу существования. Тем не менее, внутри отряда Stauriida для многих родов, в том числе Microplasma, Bothrophyllum, Gshelia, Paliphyllum и др., регенерационное почкование широко распространено и не зависит от палеогеографического положения области распространения рода.
* * *
Автор благодарит С.В. Рожнова (ПИН РАН) за ценную помощь в написании и редактировании статьи, С.В. Багирова (ПИН РАН) за проведенные сеансы фотографирования в фотолаборатории ПИН РАН. За предоставленный материал автор выражает благодарность У. Тоом и Д. Кальо (Таллинский технический ун-т). Работа выполнена при поддержке гранта РФФИ № 19-34-90003 и проекта Российского научного фонда, № 19-14-00346.
Список литературы
Волкова К.Н., Латыпов Ю.Я. Раннедевонские ругозы и мшанки Селенняхского кряжа. Новосибирск: Наука, 1976. 71 с. (Тр. Ин-та геол. и геофиз. СО АН СССР. Вып. 287).
Казанцева Е.С. Регенерация и колониальность палеозойских кораллов ругоз: реконструкция и терминология // Бюлл. МОИП. Сер. геол. 2019. Т. 94. № 3. С. 37–45.
Кузьмичева Е.А. Основные особенности функциональной морфологии колониальных организмов // Современная палеонтология. Т. 1 / Ред. Меннер В.В., Макридин В.П. М.: Недра, 1988. С. 123–139.
Сошкина Е.Д. Девонские кopaллы Rugosa Урала. М.: Изд-во АН СССР, 1949. 160 с. (Тр. Палеонтол. ин-та АН СССР. Т. 15. Вып. 4).
Сошкина Е.Д. Подкласс Tetracoralla. Четырехлучевые кораллы (Rugosa). Общая часть // Основы палеонтологии. Губки, археоциаты, кишечнополостные, черви / Ред. Соколов Б.С. М.: Изд-во АН СССР, 1962. С. 286–305.
Спасский Н.Я., Кравцов А.Г., Цыганко В.С. Колониальные цистиморфы // Древние Cnidaria. Т. 1. Новосибирск: Наука, 1974. С. 170–172.
Dybowski W.N. Beschreibung eniger nеuеn aus Nordatnerica slammenden devonischen Art der Zoantaгia rugosa // Зап. Имп. Минер. об-ва. 1873. V. 8. P. 153–160.
Dybowski W.N. Monographie der Zoantharia sclerodermata rugosa aus der Silurformation Estlands, Nord-Livlands and der Insel Gotland // Archiv Naturk. Liv-, Ehst- und Kurlands. Ser. 1. 1873–1874. Bd 5. P. 257–414.
Fedorowski J. Aulophyllidae (Tetracoralla) from the Upper Viséan of Sudetes and Holy Cross Mountains // Palaeontol. Pol. 1971. V. 24. 137 p.
Fedorowski J. Some aspects of coloniality in rugose corals // Palaeontology. 1978. V. 21. P. 177–224.
Fedorowski J., Ogar V. Early Bashkirian Rugosa (Anthozoa) from the Donets Basin, Ukraine. Part 4. Cordibia, a new protocolonial genus // Acta Geol. Pol. 2013. V. 63. № 3. P. 297–314.
Hill D. Coelenterata / Cnidaria // Rugosa & Tabulata corals / Ed. Teichert C. Boulder, Lawrence: Geol. Soc. America; Univ. Kansas Press, 1981. 762 p. (Treatise on Invertebrate Paleontology. Pt F. Suppl. 1).
Johannessen W. Silurian dissepimentate cystiphyllid rugose corals from Baltoscandia. Doctor sci. thesis. Bergen Univ., 1998. P. 1–34.
Kazantseva E.S., Rozhnov S.V. From regeneration to coloniality: multiple buds in the solitary coral Bothrophyllum conicum Trautschold, 1879 (Rugosa) in the Carboniferous of the Moscow Basin // Paleontol. J. 2018. V. 52. № 14. P. 1710–1722.
McLean R.F. Middle Devonian cystiphyllid corals from the Hume Formation, Northwestern Canada // Bull. Geol. Surv. Canada. 1976. V. 274. 80 p.
Merriam C.W. Middle Devonian Rugose Corals of the Central Great Basin // Geol. Surv. Wash. Prof. Pap. 1973. V. 799. 53 p.
Nestor H., Einasto R., Nestor V. et al. Description of the type section, cyclity, and correlation of the Riksu formation (Wenlock, Estonia) // Proc. Estonian Acad. Sci. Geol. 2001. V. 50. № 5. P. 149–173.
Rozhnov S.V. Bilateral symmetry in ontogeny and regeneration of solitary Rugosa (Cnidaria; Paleozoic) // Paleontol. J. 2014. V. 48. № 11. P. 1183–1193.
Scrutton C.T. The Paleozoic corals, I: Origins and Relationships // Proc. Yourkshire Geol. Soc. 1997. V. 51. P. 177–208.
Taylor P.W. The Plymouth limestone // Trans. Roy. Geol. Soc. Cornwall. 1951. V. 18. P. 148–159.
Verrill A.E. Classification of polyps (extract condensed from Synopsis of the Polyps and Corals of the North Pacific Exploring Expedition under Commodore C. Ringgold and Captain John Rodgers, U.S.N. // Communications of the Essex Institute. 1865. V. 4. 235 p.
Wedekind R. Die Zoantharia Rugosa von Gotland (bes. Nordgotland) nebst Bemerkungen zur Biostratigraphie des Gotlandium // Sver. Geol. Unders. 1927. V. 19. S. 1–94.
White D.E. The Silurian Rugose coral Microplasma lovenianum Dybowski from Monmouthshire // Palaeontology. 1966. V. 9. Pt 1. P. 148–151.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал