

Палеонтологический журнал, 2021, № 5, стр. 10-22
Микроструктура трубок кембрийских зоопроблематик семейства Hyolithellidae
Ю. Е. Демиденко *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: juliad@paleo.ru
Поступила в редакцию 25.12.2020
После доработки 15.01.2021
Принята к публикации 15.01.2021
Аннотация
Исследована микроструктура стенки трубок представителей рода Hyolithellus из типовых разрезов томмотского и ботомского ярусов нижнего кембрия Сибирской платформы (рр. Лена, Алдан, Синяя). Показано ее трехслойное строение. Впервые описана микроструктура наружного и внутреннего слоев трубок хиолительминтов.
ВВЕДЕНИЕ
Хиолительминты (отряд Hyolithelmintida Fisher, 1962) – вымершая группа беспозвоночных неясного систематического положения. Таксономическое разнообразие хиолительминтов представлено двумя семействами и менее чем 20 родами, известными из отложений позднего венда – карбона Европы, Азии, Северной Америки, Австралии и Антарктиды. В ископаемом состоянии сохраняется наружный скелет из фосфата кальция – это открытые с обеих сторон конические трубки с округлым (сем. Hyolithellidae) или овальным (сем. Torellellidae) поперечным сечением. Трубки хиолительминтов прямые или изогнутые в одной или нескольких плоскостях, длиной до 15–20 мм, диаметром от 0.2–0.5 до 1–2 мм, с углом расширения 1°–10°. Наружная поверхность трубок с концентрическими линиями роста, поперечными или продольными ребрами, иногда гладкая; внутренняя поверхность трубок гладкая.
Систематическое положение хиолительминтов – предмет дискуссий (табл. 1), что вызвано довольно простым строением скелета и отсутствием однозначных диагностических признаков, по которым группу можно было бы отнести к определенным типам или классам многоклеточных. Чаще всего хиолительминтов помещают в тип кольчатых червей и относят к седентарным полихетам (Fisher, 1962; Есакова, Жегалло, 1996; Demidenko, 2019) на основе внешнего сходства морфологии трубки, однако разный химический состав скелета (у седентарных полихет это карбонат кальция, у хиолительминтов – фосфат кальция) делает этот выбор систематического места для хиолительминтов весьма условным.
Таблица 1.
Варианты систематического положения хиолительминтов
| Таксон | Литература |
|---|---|
| Конулярии | Barrande, 1867; Bouček, 1939 |
| Аннелиды | Matthew, 1885; Walcott, 1911; Cobbold, Pocock, 1934; Poulsen, 1942; Fisher, 1962; Григорьева, Жегалло, 1979; Григорьева, 1980; Есакова, Жегалло, 1996; Skovsted, Peel, 2011; Demidenko, 2019 |
| Хиолиты | Сысоев, 1962 |
| Проблематики | Розанов, Миссаржевский, 1966; Мешкова, 1969; Poulsen, 1970; Bengtson et al., 1990 |
| Погонофоры | Poulsen, 1963; Southward, 1971 |
| Кишечнополостные | Миссаржевский, 1989; Bischoff, 1989; van Iten et al., 1992; Zhu et al., 2000; Vinn, 2006 |
| Нематоды | Новожилова, 2010 |
Ранее нами были изучены представители семейства Torellellidae, рассмотрены морфологические особенности их скелета и сделано предположение об их принадлежности к полихетам (Demidenko, 2019). Было показано наличие трех слоев в структуре стенки трубки. Детальная ультраструктура была выявлена в среднем слое, который состоит из многочисленных ламелл, сложенных разнонаправленными фибрами, создающими шахматный рисунок. Наружный и внутренний слои тореллеллид не удалось изучить с такой степенью детальности. Ниже приводятся результаты исследования микроструктуры представителей другого семейства хиолительминтов – семейства Hyolithellidae.
Изученный материал хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН), колл. №№ 5083, 5279.
МИКРОСТРУКТУРА ТРУБОК HYOLITHELLIDAE
Самое первое исследование микроструктуры трубок представителей семейства было проведено у вида Hyolithellus micans Billings, 1871 из нижнекембрийских глауконитовых песчаников Комли (Англия). В стенке трубки установлено присутствие двух слоев: толстого (15 мкм) внутреннего гранулированного слоя со следами параллельной слоистости и тонкого (3 мкм) наружного слоя, состоящего из апатитовых призм (Hurst, Hewitt, 1977). Позднее Р. Хьюит посчитал наружный слой перекристаллизованным (Hewitt, 1980, с. 663).
При изучении хиолительминтов H. tenuis Missarzhevsky, 1966 из томмотских отложений Сибирской платформы также было обнаружено два слоя (Григорьева, Жегалло, 1979). Внутренний слой представлен пластинами, ориентированными параллельно поверхностям трубки, внешний слой – косо расположенными загибающимися пластинами шевронной формы. Был сделан вывод о том, что хиолительминты по строению стенки трубки ближе к полихетам, нежели к погонофорам, стенка трубки которых толщиной 1.0–1.3 мкм состоит из пластин, параллельных поверхностям трубки. Для сравнения был изучен современный Spirorbis sp. с побережья Бретани (Григорьева, Жегалло, 1979, рис. 1, д), трубка которого состоит из трех слоев: наружный и внутренний толщиной по 2 мкм каждый, а также средний слой, толщиной 100–130 мкм, по строению аналогичный наружному слою хиолительминтов. При этом отмечено, что трубки Spirorbis и Hyolithellus отличаются по химическому составу. Рентгено-структурный анализ хиолительминтов показал, что их трубки состоят из франколита – богатой углеродом разновидности фторапатита (Есакова, Жегалло, 1996).
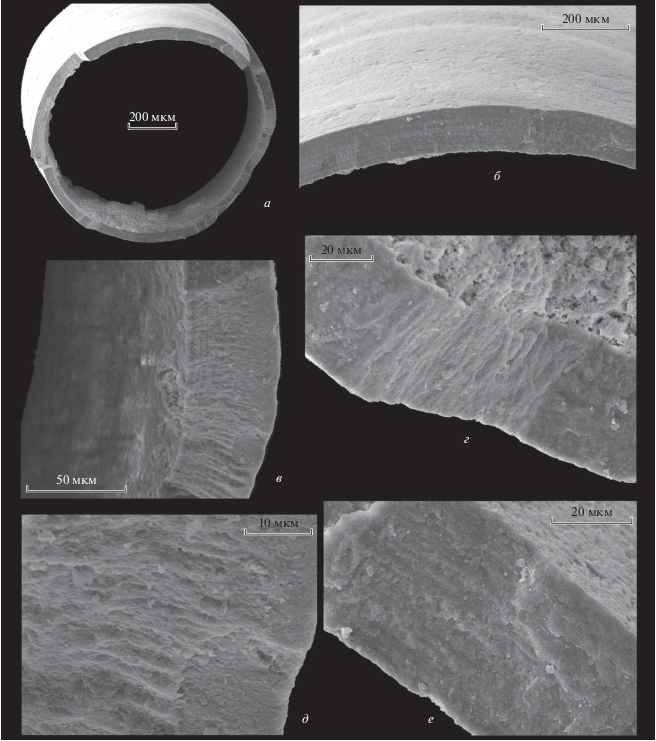
Рис. 1.
Микроструктура стенки трубки Hyolithellus sp., экз. ПИН, № 5279/5307: а – поперечное сечение трубки; б–е – строение среднего слоя: б, е – продольные структуры среднего слоя, в–д – поперечные структуры среднего слоя; Сибирская платформа, р. Лена, разрез Быдьянгая; нижний кембрий, томмотский ярус, зона D. regularis.
С. Бенгтсон усомнился в наличии внешнего слоя у Hyolithellus, ссылаясь на неубедительность приведенного авторами изображения (Bengtson et al., 1990, с. 187). При изучении H. cf. micans из отложений нижнего кембрия Южной Австралии он отметил, что трубки слоистые, имеют фосфатный состав и стенку толщиной 4–30 мкм, состоящую из тонких ламин, параллельных поверхности и сложенных параллельными фибрами диаметром 0.05–0.1 мкм; при этом соседние ламеллы имеют различные направления фиброзности (Bengtson et al., 1990, фиг. 124D ).
К. Сковстед и Дж. Пил при изучении трубок рода Hyolithellus из нижнекембрийских илистых доломитов Северной Гренландии (Aftenstjernesø Fm) впервые отметили прижизненное захоронение этих широко распространенных кембрийских ископаемых (Skovsted, Peel, 2011). Большая часть экземпляров из изученной выборки ориентирована узким концом трубок вниз (при нормальном залегании вмещающего слоя). Разнообразие в направлении линий нарастания объясняется активной регулировкой животным своего роста путем достижения вертикального положения трубки по отношению к поверхности осадка. Увеличение толщины трубки по направлению к зауженному концу сдвигает центр масс вниз и обеспечивает большую устойчивость в осадке. На протяжении всего онтогенеза трубка оставалась открытой с обоих концов. Авторы предположили, что хиолиты – это кольчатые черви, фильтровавшие воду через осадок.
Нами изучена микроструктура трубок Hyolithellus vladimirovae Missarzhevsky, 1966 и H. tenuis Missarzhevsky, 1966, происходящих из разреза руч. Быдьянгая в среднем течение р. Лены, Сибирская платформа. Интервал разреза, из которого отобраны ископаемые, литологически и фаунистически коррелируется со слоем 10 разреза Исить – стратотипом зоны Dokidocyathus regularis томмотского яруса нижнего кембрия (Розанов и др., 1969; Путеводитель …, 1973; Ярусное расчленение …, 1984; Варламов и др., 2008). Также изучены трубки Hyolithellus sp. из подошвы суннагинской пачки разреза Васькина протока, р. Алдан, Сибирская платформа (томмотский ярус, зона Nochoroicyathus sunnaginicus) и III пачки переходной свиты в низовьях р. Синяя, Сибирская платформа (ботомский ярус, зона Bergeroniellus micmacciformis–Erbiella).
Микроструктурное исследование с помощью сканирующего электронного микроскопа показало трехслойное строение стенки трубок у представителей семейства Hyolithellidae. Подобное трехслойное строение стенки трубок у хиолительминтов семейства Torellellidae было установлено ранее для Torellella gracilenta Esakova, 1996 из томмотского яруса Сибирской платформы (Demidenko, 2019).
Наружный слой трубок Hyolithellus состоит из ламин, сформированных плотно расположенными гексагональными призмами высотой от 3 до 5.5–6.5 мкм, толщиной 2–3 мкм (рис. 3, б; 4, ж; 6, ж, з). Подобная микроструктура называется фавозитной, поэтому наружный слой можно также именовать фавозитным. Этот слой в виде тонкой пленки покрывает мощный средний слой, превышающий его по толщине во много раз. Встречаются экземпляры, на наружном слое которых видна трещиноватость (рис. 3, в, г; 4, в), сквозь эти трещины можно четко увидеть гексагональные призмы фавозитной микроструктуры. Часто фавозитный слой присутствует фрагментарно (рис. 3, д, е) или не сохраняется, и мы видим сразу внешнюю поверхность среднего слоя.
Рис. 2.
Микроструктура стенки трубки Hyolithellus sp., экз. ПИН, № 5083/5105: а – обломок трубки, б – увеличенный фрагмент; в – фрагмент трубки изнутри, видны бакулы среднего слоя, г – увеличенный фрагмент; д – увеличенный фрагмент поперечного скола, видна слоистость среднего слоя; е – наружная поверхность среднего слоя; Сибирская платформа, р. Алдан, разрез Васькина протока; нижний кембрий, томмотский ярус, зона N. sunnaginicus.

Рис. 3.
Микроструктура стенки трубки Hyolithellus sp., экз. ПИН, № 5083/5172: а – общий вид трубки, б–г – строение наружного слоя трубки: б – видны гексагональные призмы, слагающие фавозитную микроструктуру наружного слоя; увеличенный фрагмент; в, г – залегающий на среднем слое трещиноватый наружный слой, сложенный гексагональными призмами; д, е – наружный и средний слои: д – наружный и средний слои, е – виден преимущественно средний слой; Сибирская платформа, р. Алдан, разрез Васькина протока; нижний кембрий, томмотский ярус, зона N. sunnaginicus.
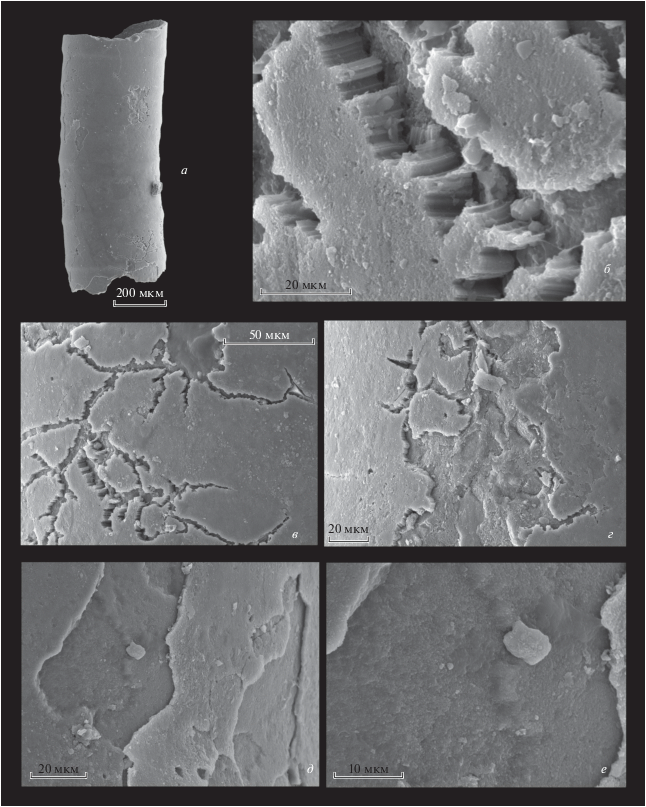
Рис. 4.
Микроструктура стенки трубки Hyolithellus sp.: а, ж – экз. ПИН, № 5083/5205: а – общий вид трубки, ж – наружный фавозитный слой, сложенный гексагональными призмами; видны наружный и средний слои; б–е, з – экз. ПИН, № 5083/5179: б – общий вид трубки, в–е, з – наружный слой, сложенный гексагональными призмами: в – трещиноватость наружного слоя; Сибирская платформа, р. Алдан, разрез Васькина протока; нижний кембрий, томмотский ярус, зона N. sunnaginicus.

Максимальный по толщине средний слой мощностью 30–50 мкм состоит из нескольких (3‒9?) слойков (табл. II , фиг. 3б–3д, табл. III , фиг. 3б, 3в; рис. 1, б–г, е; 2, д; 5, ж), которые, в свою очередь, состоят из многочисленных пластин – ламелл. Каждая ламелла состоит из фибр (рис. 6, и, к), причем у соседних ламелл наблюдается разнонаправленная фиброзность (рис. 6, и). Для среднего слоя представителей рода Hyolithellus подобная ламеллярная микроструктура выявлена впервые. Она представлена чередованием многочисленных ламелл с характерными рисунками из фибр на наружных поверхностях: ламелла с продольной струйчатостью сменяется ламеллой с поперечной струйчатостью (рис. 6, и). Таким образом формируется “шахматный” рисунок среднего слоя. Это отчетливо видно на фрагментах наружной скульптуры двух и более соседних подстилающей и налегающей ламелл (табл. III , фиг. 1б, 1г, 2б–2г; рис. 6, и). На поперечных сколах трубки между ламеллами можно наблюдать остатки органического слоя или пустоты, если этот слой отсутствует. Когда поперечные фибры разрушаются, на их месте можно наблюдать полости или зияния, которые ошибочно можно принять за крупные поровые отверстия. Продольные фибры обычно сохраняются лучше поперечных (рис. 5, ж, з, к). Вывод о том, что соседние ламеллы у хиолительминтов могут иметь различные направления фиброзности, совпадает с данными Бенгтсона, полученными при изучении H. cf. micans, и данными, полученными при изучении рода Torellella (Demidenko, 2019).
Рис. 5.
Микроструктура стенки трубки Hyolithellus sp.: а, ж, з, к – экз. ПИН, № 5083/5192: а – общий вид трубки, ж, з, к – увеличенные фрагменты среднего ламеллярного слоя: вверху видны ламины, состоящие из параллельных ламелл, ниже – соединенные между собой вертикальные столбики ламелл в разрезе, полости между вертикальными столбиками – несохранившиеся горизонтальные ламеллы; б, и – экз. ПИН, № 5083/5202: б – общий вид трубки, и – вертикальные заостренные столбики ламелл среднего ламеллярного слоя; в–е – экз. ПИН, № 5083/5188: в – общий вид трубки, г–е – залегающие вдоль вертикальные столбики ламелл среднего ламеллярного слоя; Сибирская платформа, р. Алдан, разрез Васькина протока; нижний кембрий, томмотский ярус, зона N. sunnaginicus.

Рис. 6.
Микроструктура стенки трубки Hyolithellus sp.: а, б, д, е – экз. ПИН, № 5083/5170: а – общий вид трубки, б – увеличенный фрагмент устьевой части, д, е – фрагменты трубки изнутри, виден внутренний слой; в, ж, з – экз. ПИН, № 5083/5209: в – общий вид трубки, ж, з – увеличенные фрагменты фавозитного наружного слоя, состоящего из гексагональных призм; г, и, к – экз. ПИН, № 5083/5228: г – общий вид трубки, и, к – увеличенные фрагменты среднего ламеллярного слоя; Сибирская платформа, р. Алдан, разрез Васькина протока; нижний кембрий, томмотский ярус, зона N. sunnaginicus.
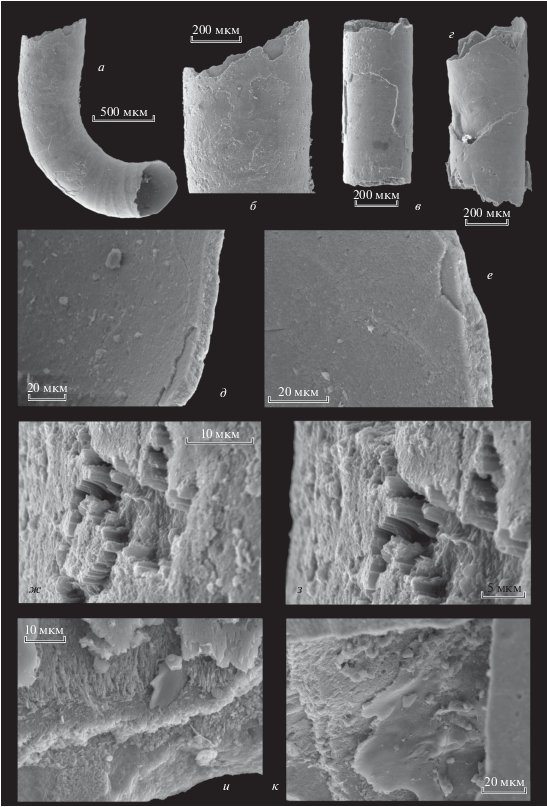
Объяснение к таблице II
Фиг. 1–3. Hyolithellus vladimirovae Missarzhevsky, 1966: 1 – экз. ПИН, № 5279/5353: 1а – общий вид обломанной трубки; 1б – увеличенный фрагмент устьевой части обломанной трубки; 2 – экз. ПИН, № 5279/5340: 2а – общий вид трубки; 2б – поперечный скол трубки: видны наружный и средний слои; Сибирская платформа, разрез Быдьянгая; нижний кембрий, томмотский ярус, зона D. regularis; 3 – экз. ПИН, № 5279/5513: 3а – обломанная часть трубки изнутри; 3б–3д – многослойная структура среднего слоя стенки трубки; Сибирская платформа, разрез по р. Синей; нижний кембрий, ботомский ярус, зона Bergeroniellus micmacciformis–Erbiella, III пачка, 4 слой.

Внутренний слой является аналогом наружного слоя: пористый, обладает фавозитной микроструктурой, сложенной гексагональными призмами, и сферулитовой наружной поверхностью, однако является меньшим по толщине – 2–3 мкм (рис. 6, д, е). Когда сохранность материала неудовлетворительная, мы можем наблюдать так называемую вертикальную столбчатую отдельность наружного и внутреннего слоев, оставшуюся от вертикально расположенных гексагональных призм. Сохраняется внутренний слой редко, вероятно, в силу своей небольшой толщины. Обычно мы видим ламеллярную слоистость среднего слоя без присутствия наружного и внутреннего слоев.
Как показали предыдущие исследования, обычно главными составляющими подобных трубок и других SSF являются кальций и фосфор (Ушатинская, 2018; Demidenko, 2019). Вероятнее всего, наружный и внутренний слои трубки раковины представляли собой при жизни животного тонкие органо-фосфатные слои, которые после его смерти попадали в условия с повышенным содержанием кислорода, повышенной щелочностью и с присутствием бактерий, ускоряющих процессы разложения и замещения. Такие условия способствовали частичному переходу в растворимое состояние ионов фосфора и кальция из мягких тел и из раковин организмов, и дальнейшему осаждению их в виде минерального апатита, замещающего первичные структуры. Уникальная сохранность при захоронении позволяет нам наблюдать все три слоя раковины трубки с детально сохранившейся микроструктурой. Примером этому могут служить сохранившиеся пластины среднего слоя, сформированные соединением вертикальных столбиков ламелл, которые мы можем наблюдать на сколе (рис. 5, ж, з, к). Такая сохранность наблюдается, когда происходит быстрая, почти мгновенная фосфатизация.
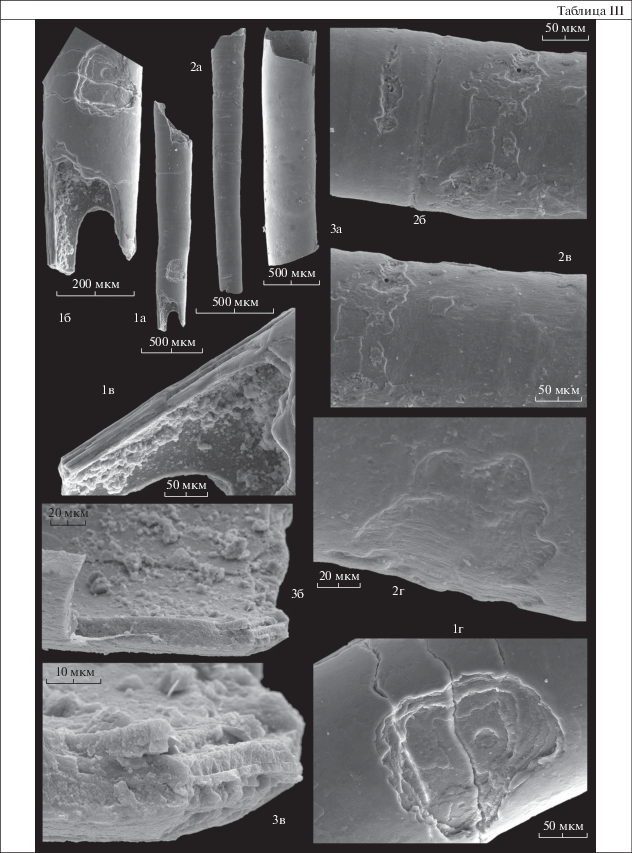
Объяснение к таблице III
Фиг. 1, 2. Hyolithellus tenuis Missarzhevsky, 1966: 1 – экз. ПИН, № 5279/5358: 1а – общий вид трубки; 1б – увеличенный фрагмент начальной части трубки; 1в – продольный скол начальной части трубки; 1г – фрагмент трубки: видна слоистость среднего слоя с разнонаправленной фиброзностью; 2 – экз. ПИН, № 5279/5361: 2а – общий вид трубки; 2б‒2г – увеличенные фрагменты трубки: видна слоистость среднего слоя с разнонаправленной фиброзностью и, возможно, фрагменты ?наружного слоя; Сибирская платформа, разрез Быдьянгая; нижний кембрий, томмотский ярус, зона D. regularis.
Фиг. 3. Hyolithellus vladimirovae Missarzhevsky, 1966, экз. ПИН, № 5279/5523: 3а – общий вид трубки; 3б, 3в – многослойная структура среднего слоя стенки трубки; Сибирская платформа, разрез по р. Синей; нижний кембрий, ботомский ярус, зона Bergeroniellus micmacciformis – Erbiella, III пачка, 5 слой.
Основываясь на изучении микроструктуры стенки раковины представителей двух семейств отряда Hyolithelmintida – Hyolithellidae в данной статье и Torellellidae ранее (Demidenko, 2019), можно предположить, что микроструктура хиолительминтов, скорее всего, является родовым признаком, а не признаком более высоких рангов (класс, тип). Данный вывод получен, исходя из характерной для каждого семейства отличающейся друг от друга микроструктуры наружного и внутреннего слоев, в то время как строение среднего слоя является схожим. Для подтверждения этого заключения требуется более детальное изучение наружного и внутреннего слоев представителей семейства Torellellidae.
Проведенное изучение трубок хиолительминтов семейства Hyolithellidae не противоречит их интерпретации как “домиков” сидячих полихет. Это подтверждается сходством в общей форме трубок с радиальной симметрией и наличием трех слоев стенки различной мощности – фавозитного наружного слоя, ламеллярного среднего слоя и идентичного наружному внутреннего фавозитного слоя. У современных полихет также наблюдается трехслойное строение раковинной трубки со слоями схожей толщины.
* * *
Автор благодарен Н.В. Есаковой за материал из разреза Васькина протока, р. Алдан (сборы 1981 г.), П.Ю. Пархаеву за предоставленные образцы пород с р. Синей (сборы 2009 г.), Р.А. Ракитову за помощь при работе на СЭМ Tescan Vega III кабинета приборной аналитики ПИН РАН. Исследование выполнено при финансовой поддержке РФФИ (проект № 19-04-01027а).
Список литературы
Варламов А.И., Розанов А.Ю., Хоментовский В.В. и др. Кембрий Сибирской платформы. Книга 1: Алдано-Ленский регион. М.: ПИН РАН, 2008. 300 с.
Григорьева Н.В. К вопросу об исследовании микроструктуры хиолительминтов // Палеонтология и стратиграфия: доклады советских геологов к XXVI Междунар. геол. конгр. М.: Наука, 1980. С. 49–55.
Григорьева Н.В., Жегалло Е.А. К исследованию микроструктур некоторых томмотских ископаемых // Палеонтол. журн. 1979. № 2. С. 142–144.
Есакова Н.В., Жегалло Е.А. Биостратиграфия и фауна нижнего кембрия Монголии. М.: Наука, 1996. 216 с. (Тр. Совм. Росс.-Монгол. палеонтол. экспед. Вып. 46).
Мешкова Н.П. К вопросу о палеонтологической характеристике нижнекембрийских отложений Сибирской платформы // Биостратиграфия и палеонтология нижнего кембрия Сибири и Дальнего Востока. М.: Наука, 1969. С. 158–174.
Миссаржевский В.В. Древнейшие скелетные окаменелости и стратиграфия пограничных толщ докембрия и кембрия. М.: Наука, 1989. 237 с. (Тр. Геол. ин-та АН СССР. Вып. 443).
Новожилова Н.В. Новые данные по морфологии и систематическому положению хиолительминтов (кембрийские проблематичные организмы) // Палеонтол. журн. 2010. № 2. С. 6–9.
Путеводитель экскурсии по рекам Алдану и Лене: (Международная экскурсия по проблеме границы кембрия и докембрия). М.; Якутск: ВИЭМС, 1973. 118 с.
Розанов А.Ю., Миссаржевский В.В. Биостратиграфия и фауна нижних горизонтов кембрия. М.: Наука, 1966. 144 с. (Тр. Геол. ин-та АН СССР. Вып. 148).
Розанов А.Ю., Миссаржевский В.В., Волкова Н.А. и др. Томмотский ярус и проблема нижней границы кембрия. М.: Наука, 1969. 380 с. (Тр. Геол. ин-та АН СССР. Вып. 206).
Сысоев В.А. Хиолиты кембрия северного склона Алданского щита. М.: Изд-во АН СССР, 1962. 66 с.
Ушатинская Г.Т. Сохранение минерализованных мягких тканей и их отпечатков в раковинах древних фосфатных брахиопод // Палеонтол. журн. 2018. № 5. С. 23–27.
Ярусное расчленение нижнего кембрия. Стратиграфия / Ред. Розанов А.Ю., Соколов Б.С. М.: Наука, 1984. 184 с.
Barrande J. Système Silurien du centre de la Bohême. 1: Recherches paléontologiques. V. 3: Classe des Mollusques, Ordre des Pteropodes. Prague, 1867. 179 p.
Bengtson S., Conway Morris S., Cooper B. et al. Early Cambrian fossils from South Australia // Mem. Assoc. Austral. Palaeontol. 1990. № 9. P. 1–364.
Bischoff C.G.O. Byroniida new order from early Palaeozoic strata of eastern Australia (Cnidaria, thecate scyphopolyps) // Senkenb. Lethaea. 1989. V. 59. P. 275–327.
Bouček B. Conularida // Handbuch der Paläozoologie. Bd 2A / Ed. Schindewolf O.H. Berlin: Verl. Gebrüder Borntraeger, 1939. S. 113–131.
Cobbold E.S., Pocock R.W. The Cambrian area of Rushton (Shropshire) // Phil. Trans. Roy. Soc. London B. 1934. V. 223. P. 305–411.
Demidenko Yu.E. Morphology, systematic position, and stratigraphic distribution of Hyolithelmintida Fisher, 1962 – Torellella gracilenta Esakova, 1996 // Paleontol. J. 2019. V. 53. № 7. P. 676–688.
Fisher D.W. Small conoidal shells of uncertain affinities // Treatise on Invertebrate Paleontology. Pt W. Miscellanea. Lawrence: Univ. Kansas Press, 1962. P. 98–143.
Hewitt R.A. Microstructural contrasts between some sedimentary francolites // J. Geol. Soc. London. 1980. № 137. P. 661–667.
Hurst J.M., Hewitt R.A. Tubular problematica from the type Caradoc (Ordovician) of England // N. Jb. Geol. Paläontol. Abh. 1977. Bd 153. № 2. S. 147–169.
Matthew G.F. Notice of a new genus of Pteropods from the Saint John Group (Cambrian) // Amer. J. Sci. Arts. Ser. 3. 1885. V. 25. № 178. P. 105–111.
Poulsen C. Some hitherto unknown fossils from the Exsulans limestone of Bornholm // Medd. Dan. Geol. Fören. 1942. V. 10. № 2. P. 212–235.
Poulsen C. The lower Cambrian from Slagelse. 1. Western Sealand // Geol. Surv. Senm. Ser. 2. 1970. V. 93. P. 18–20.
Poulsen V. Notes on Hyolithellus Billings, 1871. Class Pogonophora Johanson, 1937 // Biol. Medd. Dan. Vidensk. Selsk. 1963. V. 23. P. 1–15.
Skovsted C.B., Peel J.S. Hyolithellus in life position from the Lower Cambrian of North Greenland // J. Paleontol. 2011. V. 85. № 1. P. 37–47.
Southward E. Recent researches on the Pogonophora // Oceanogr. Mar. Biol. Ann. Rev. 1971. V. 9. P. 193–220.
van Iten H., Cox R.S., Mapes R.H. New data on the morphology of Sphenothallus Hall: Implications for its affinities // Lethaia. 1992. V. 25. P. 135–144.
Vinn O. Possible cnidarian affinities of Torellella (Hyolithelminthes, Upper Cambrian, Estonia) // Paläontol. Z. 2006. Bd 80. № 4. P. 384–389.
Walcott C.D. Cambrian geology and paleontology. Middle Cambrian annelids // Smithson. Misc. Coll. 1911. V. 57. № 5. P. 109–144.
Zhu M.Y., van Iten H., Cox R.S. et al. Occurrence of Byronia Matthew and Sphenothallus Hall in the Lower Cambrian of China // Paläontol. Z. 2000. Bd 74. P. 227–238.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал