

Палеонтологический журнал, 2020, № 4, стр. 79-88
Новый бентозухид (Amphibia: Temnospondyli) из нижнего триаса Московской синеклизы
Б. И. Морковин *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: prodeo27@yandex.ru
Поступила в редакцию 22.07.2019
После доработки 30.10.2019
Принята к публикации 30.10.2019
Аннотация
Из раннетриасового местонахождения Блудново (Вологодская обл., бассейн р. Юг) описан новый бентозухид Benthosuchus lukyanovi sp. nov. Новый вид обладает более крупными размерами черепа в сравнении с ранее известными находками Benthosuchus. Рассмотрены морфологические отличия от других представителей рода, значительная часть которых связана с выраженностью черт трематозаврового плана строения: сближенные в межноздревом промежутке супраорбитальные каналы (sulci supraorbitales), слабый ангулярный изгиб нижней челюсти, а также удлинение заднего меккелева отверстия и др. Полученные в ходе работы данные расширяют представления о путях морфологической эволюции у бентозухид на ранних этапах становления трематозавридного типа организации.
Бентозухиды – группа трематозавроидных лабиринтодонтов (Amphibia: Temnospondyli), широко распространенная в нижнетриасовых отложениях Восточно-Европейской платформы. На ранних этапах изучения (Быстров, Ефремов, 1940) представители семейства бентозухид были отнесены к неорахитомам, а позднее (Romer, 1947; Очев, 1966) к стереоспондильным капитозавроидам. В дальнейшем их систематические взаимоотношения были пересмотрены (Шишкин, 1980; Гетманов, 1989; Новиков, 2012), группа введена в состав надсемейства Trematosauroidеа, при этом авторами отмечается, что бентозухиды занимают промежуточное положение между капитозавроидами и трематозавроидами. В последние десятилетия систематическое положение и родственные связи семейства Benthosuchidae неоднократно обсуждались, в связи с вопросом о происхождении трематозаврид (Schoch, 2000; Schoch, Milner, 2000; Steyer, 2002; Damiani, Yates, 2003). Большинство упомянутых авторов рассматривают это семейство, как базальную группу трематозавроидов.
Находки представителей рода Benthosuchus приурочены к отложениям рыбинского, слудкинского и, возможно, усть-мыльского горизонтов (Новиков, 2018) оленекского яруса. Род является руководящим для рыбинского горизонта. Наиболее широко здесь представлен типовой вид – B. sushkini Efremov, описанный И.А. Ефремовым из северной части Московской синеклизы (Efremov, 1929; Ефремов, 1937; Быстров, Ефремов, 1940), а также известный из местонахождений южного склона Волго-Уральской антеклизы и Южного Приуралья. Внутри вида, вероятно, существуют две расы (или подвиды), соответствующие северному и южному ареалам их распространения (Шишкин и др., 2006). В.Г. Очев (1958) описал род Parabenthosuchus (P. uralensis) из блюментальской свиты Южного Приуралья, сначала считая его предковым для Benthosuchus (Очев, 1958), но в дальнейшем признал его синонимичность с этим родом (Очев, 1972). Из кзылсайской свиты Башкирского Приуралья Очевым (1972) был описан также B. bashkiriсus, имеющий некоторые прогрессивные отличия от B. sushkini. Локально распространенной, но широко представленной формой является B. korobkovi (Ивахненко, 1972) из прибрежно-озерных отложений рыбинского горизонта Верхнего Поволжья. В 1989 г. по материалам из коллекции А.П. Быстрова, предположительно собранной в отложениях рыбинского горизонта на р. Унже, был выделен вид B. bystrowi (Гетманов, 1989), единственный экземпляр которого (голотип) впоследствии оказался утраченным. Наиболее архаичный представитель Benthosuchus – B. gusevae – описан из низов каменноярской свиты Бузулукской впадины (Новиков, 2012). Указывалось, что этот вид является стратиграфически древнейшим (низы рыбинского горизонта) в составе рода и наиболее близок к капитозавроидному плану строения.
Изложенные представления о видовом составе рода Benthosuchus в настоящий момент могут быть дополнены, благодаря находке его нового представителя в нижнетриасовых отложениях бассейна р. Юг (см. ниже).
МАТЕРИАЛ И МЕТОДИКА
Неполный череп Benthosuchus вместе с остатками плечевого пояса, происходящие из песчанистых глин шилихинской свиты рыбинского горизонта, были собраны автором в 2017 г. на новом местонахождении, расположенном на левом берегу р. Юг у дер. Блудново (Вологодская обл.). Кроме того, в базальной части разреза был найден изолированный гипоцентр позвонка Benthosuchus (Морковин, 2018).
Особенности сохранности. Найденный образец является единственным для бентозухид, сохранившим нижнюю челюсть в прижизненном сочленении с черепом. Отмечается отсутствие следов мацерации, швы между костями трудноразличимы.
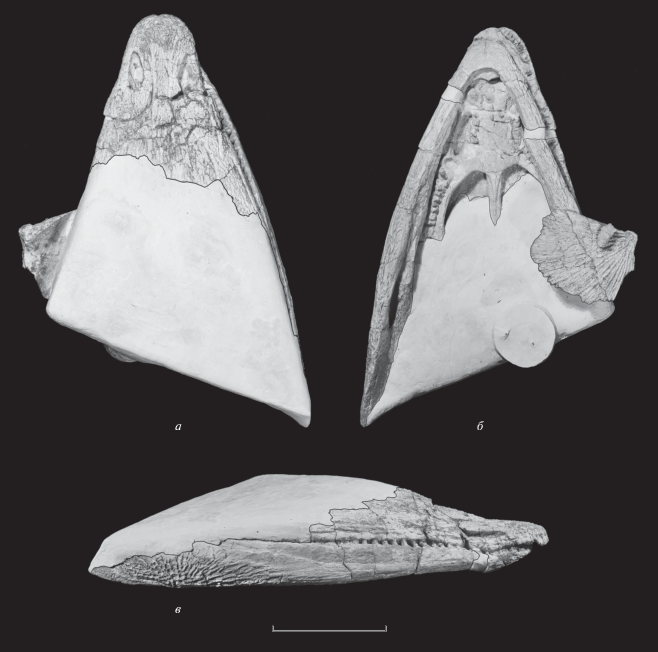
Сохранившаяся часть крыши представлена пренариальной и преорбитальной (значительная часть) зонами; орбитальная, посторбитальная и затылочная части черепа утрачены. Небный комплекс представлен частично, от заднего края fenestra praemaxillaris до переднего края межптеригоидных окон (fenestra interpterygoidea). Нижняя челюсть с сочлененным симфизом. Также, сохранились части дермального плечевого пояса (ключиц и межключицы) и фрагмент кости подъязычного аппарата, незначительно смещенные по сравнению со своим прижизненным положением (рис. 1).
Рис. 1.
Benthosuchus lukyanovi sp. nov., голотип ПИН, № 5618/1, череп с нижней челюстью и элементами посткраниального скелета: а – дорсальная сторона, б – вентральная, в – сбоку. Масштабная шкала 10 см.
Методика измерений. В нижеприведенном сравнении морфометрических особенностей видов Benthosuchus использовались следующие характеристики: осевая длина черепа – L; общая ширина черепа – W; отношение осевой длины к общей ширине черепа – L/W; w/l ch – индекс удлинения хоан; w/l prn – индекс удлинения пренариальной области; w/l nar – индекс удлинения ноздрей; ang nar – угол между длинной осью ноздри и сагиттальной осью черепа (в случае латерального отклонения приводятся положительные значения, медиального – отрицательные); dpf – угол схождения постфенестральных зубных рядов; общая длина нижней челюсти – l mnd; индекс удлинения заднего меккелевого отверстия – w/l fmp; угол схождения нижних челюстей – ang mnd; угол ангулярного изгиба нижней челюсти – ang an; соотношение осевой длины задней пластины к скульптированной зоне нижнечелюстного симфиза в вентральном плане – Lp/Ls. Полученные данные обобщены в табл. 1.
Таблица 1.
Некоторые количественные показатели признаков для видов Benthosuchus и Vyborosurus
| № | Признаки | Benthosuchus sushkini | B. korobkovi | B. gusevae | B. lukyanovi sp. nov. | Vyborosurus mirus |
|---|---|---|---|---|---|---|
| Череп | ||||||
| 1 | L, мм | 229–296 (255) | 180–302 (259) | 165 | ≈400 | * |
| 2 | W, мм | 146–200 (169) | 136–216 (161) | 122 | * | 170 |
| 3 | L/W | 1.47–1.66 (1.52) | 1.39–1.43 (1.41) | 1.35 | * | * |
| 4 | w/l ch | 0.33–0.44 (0.37) | 0.34–0.51 (0.40) | 0.47 | 0.29 | * |
| 5 | w/l prn | 0.55–0.6 | 0.5 | 0.4 | 0.53 | * |
| 6 | w/l nar | 0.26–0.46 (0.38) | 0.33–0.43 (0.41) | 0.46 | 0.38 | * |
| 7 | ang nar., град | от 0 до –10 (–9) | от –6 до –11 (–8) | –15 | 5 | * |
| 8 | Число зубов praemaxillare | 12–17 (14) | 14–17 (15) | 15 | 10 | * |
| 9 | Число зубов постфенестрального зубного ряда | 6–10 (8) | 4–9 (6) | 6 | 5 | * |
| 10 | ∠dpf., град | от 0 до 27 (15) | от 8–79 (50) | 69 | 20 | * |
| Нижняя челюсть | ||||||
| 11 | l mnd, мм | 250–308 (273) | ≈293–360 (322) | 246 | ≈410 | * |
| 12 | w/l fmp | 0.3–0.6 (0.4) | 0.31–0.5 (0.38) | 0.32 | 0.23 | 0.39 |
| 13 | ang man., град | 44–51(48) | 63 | 73 | 51 | * |
| 14 | ang an., град | 153–167 (161) | 153–155 (154) | 157 | 174 | 170 |
| 15 | Число зубов dentale | 30–47 (38) | 34 | 38 | ≈25 | * |
| 16 | Lp/Ls | 0.55–1.5 (0.7) | * | ≈0.57 | 0.45 | * |
Измерения проводились с помощью штангенциркуля в случае линейных промеров, взятых непосредственно на материале, также использовались замеры на плоскостных проекциях цифровых фотографий образцов, сделанные с помощью программы mySize 1.0. Угловые измерения были проведены с использованием электронного транспортира Screen Protractor 4.0. Угол схождения ветвей нижней челюсти, в большинстве случаев, измерялся на фотографиях одной из сохранившихся ветвей, зеркально отображенной и соединенной по симфизному шву в графическом редакторе.
Материал. Для сравнительного анализа были использованы следующие материалы по черепам и нижним челюстям Benthosuchus, доступные для изучения в ПИН РАН: B. sushkini (29 образцов) – экз. ПИН, №№ 2252/2, 3, 5, 9, 11, б/н, 33, 41, 42, 44; 2354/1, 2, 3; 2424/1, 4, 5, 10, 37, 52; 2483/1; 2484/1; 2796/276; 3783/3, 4, 5, 6; 3998/1; 4566/1; 4655/2; B. korobkovi (12 обр.) – экз. ПИН, №№ 3200/1, 63, 66, 169, 179, 207, 258, 275, 276, 323, 324, 517. Вид B. gusevae изучался по голотипу из колл. Самарского областного историко-краеведческого музея (СОИКМ, № КМК КП_23420), а также по экз. ПИН, № 4495/11.
Кроме того, в анализ включены данные по Vyborosaurus mirus (экз. ПИН, №№ 4364/1 и 3360/9). Выборки таксонов, относящиеся к видам рода Benthosuchus, изучались в средних и старших возрастных интервалах, с осевой длиной черепа от 165 до 302 мм (3–5 размерных групп, о методике разделения групп см.: Морковин, 2015, 2017). Нижние челюсти, ввиду своей редкости, были взяты от особей разных индивидуальных возрастов. Помимо перечисленных таксонов, дополнительно рассматривались некоторые другие трематозавроиды: Thoosuchus yakovlevi (6 обр.), экз. ПИН, №№ 3200/35, 93, 213, 456, 473, 477; Angusaurus succedaneus (2 обр.), экз. ПИН, №№ 2428/1, 4362/21; Inflectosaurus amplus (1 обр.), экз. ПИН, № 2242/2.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
НАДОТРЯД TEMNOSPONDYLI
ОТРЯД CAPITOSAUROMORPHA
СЕМЕЙСТВО BENTHOSUCHIDAE EFREMOV, 1937
ПОДСЕМЕЙСТВО BENTHOSUCHINAE EFREMOV, 1937
Род Benthosuchus Efremov, 1937
Rhinesuchus: Яковлев, 1916, c. 158 (part.).
Benthosaurus: Efremov, 1929, c. 757; Hartmann-Weinberg, Kusmin, 1936, c. 56.
Benthosuchus: Ефремов, 1937, с. 131; Быстров, Ефремов, 1940, с. 12; Шишкин, 1964, с. 90; Очев, 1972, c. 155; Каландадзе и др., 1968, c. 77; Ивахненко, 1972, с. 93; Гетманов, 1989, с. 8; Шишкин, 1995, c. 59; Новиков в: Ивахненко и др., 1997, с. 11; Shishkin et al., 2000, c. 49; Schoch, Milner, 2000, c. 103, Новиков, 2012, с. 70; 2018, с. 58.
Rasaurus: Кузьмин, 1938, с. 14 (part.).
Parabenthosuchus: Очев, 1958, с. 486; Шишкин, 1964, с. 91.
Типовой вид — Benthosuchus sushkini Efremov, 1937; нижний триас, рыбинский горизонт Восточной Европы.
Диагноз. См. Новиков, 2018, с. 58.
Видовой состав. Кроме типового вида, из нижнего триаса, ветлужского надгоризонта, рыбинского и слудкинского горизонтов Европейской России B. korobkovi Ivachnenko, 1972; B. bashkiricus Ochev, 1972; B. gusevae Novikov, 2012; B. lukyanovi sp. nov.
Benthosuchus lukyanovi Morkovin, sp. nov.
Вид назван в честь препаратора ПИН РАН И.И. Лукьянова (1940-2013).
Голотип – ПИН, №5618/1, часть черепа в сочленении с нижней челюстью, ассоциированная с фрагментами межключицы, обеих ключиц и, предположительно, элементом подъязычного аппарата; Вологодская обл., верхнее течение бассейна р. Юг, у дер. Блудново; нижний триас, рыбинский горизонт, шилихинская свита.
Описание (рис. 1–4). Череп крупных размеров (L ≈ 400 мм; здесь и далее см. табл. 1), удлиненно-треугольной формы, суженный и вытянутый в носовой части. Передний конец морды имеет округлые очертания, с крупным foramen interpraemaxillare, боковые края черепа, в целом, прямые и плавно сближаются вперед, на уровне ноздрей присутствует слабо выраженный пережим. Пренариальный отдел вытянутый (w/l prn – 0.53). Скульптура дорсальной поверхности в сохранившейся части крыши преимущественно ячеистая, в пренариальной зоне грубая с хорошо развитыми валиками. Кости плотно срастаются; швы между ними слабо различимы. Каналы системы боковой линии хорошо развитые и относительно широкие. Sulci supraorbitales между ноздрями сильно сближены и проходят рядом с сагиттальным швом. Лакримальная флексура sulcus infraorbitalis S-образная. Передняя комиссура выражена слабо. Ноздри вытянутые (w/l nar – 0.38), овальные, сужающиеся вперед. Характерной чертой является латеральное отклонение их длинных осей относительно сагиттальной оси черепа (ang nar – 5°), septomaxillare отсутствует. На вентральной стороне черепа хоаны лежат впереди уровня небных окон и сильно вытянуты (w/l ch – 0.29). Задние сошниковые отростки длинные и, по-видимому, распространяются более, чем на одну треть длины небных окон. Начиная от основания отростков, их медиальный (шовный) край образует продольный киль с хорошо выраженным рельефом, медиальная часть которого имеет уплощенный горизонтальный край. Постфенестральные зубные ряды на сошниках образуют клин с углом 20° и несколько заходят сзади за уровень передних краев хоан. Максимальное число зубов в рядах до 5. Сошниковые клыки крупные, округлого сечения. Palatinum несет крупный клык и ямку, также сохранились до 10 зубов парахоанального ряда (dentes parachoanales). Кость образует с vomer изогнутый шов, ориентированный под углом 48° относительно сагиттальной оси. Зубной ряд praemaxillare несет до 10 относительно крупных зубов на одной из сторон, их основания поперечно расширены. Зубной ряд maxillare скрыт его контактом с нижней челюстью. Нижняя челюсть имеет слабый ангулярный изгиб (ang an – 174°). Угол схождения ее ветвей – 51°, в плане они слабо изогнуты. Заднее меккелево отверстие сильно вытянутое (w/l fmp – 0.23), с заостренным передним краем. От переднего конца отверстия протягивается вперед углубление, над которым postspleniale образует характерный гребень–валик (crista lingualis). Скульптура нижней челюсти преимущественно радиальная, на angulare она заполняет около 2/3 поверхности кости, также наблюдаются небольшие поля ячеистой скульптуры на postspleniale, spleniale и dentale. В системе каналов боковой линии хорошо развит sulcus marginalis, идущий к центру окостенения angulare; слабее выражен sulcus dentalis. Dentale в доступной для обозрения части несет в краевом ряду около 25 крупных, широко расставленных зубов, приблизительно одинаковых по размеру. Лабиальная поверхность dentale гладкая, без выраженных следов наружной челюстной артерии. На вентральной стороне челюсти скульптированный отдел симфиза превышает по длине его гладкую заднюю пластину (Lp/Ls – 0.45), задний край последней заострен. Из элементов посткраниального скелета частично сохранились кости покровного плечевого пояса. Вентральная поверхность ключиц покрыта в основном радиальным орнаментом с небольшими ячеистыми полями у центров окостенений. Латеральный край ключицы (сохранившаяся его часть), несущий processus praescapularis, слабо изогнут в дорсо-вентральном направлении. Передний (суженный) отдел interclavicula несколько расширен. Сохранившийся фрагмент части подъязычного аппарата, предположительно, определен нами как hypobranchiale (рис. 2).
Рис. 2.
Benthosuchus lukyanovi sp. nov., прорисовка голотипа ПИН, № 5618/1: а – сверху, б – снизу, в – сбоку. Обозначения: An – angulare; Ch – choana; Cl – clavicula; с. l. – crista lingualis; D – dentale; f.m.p. – foramen meckelianum posterior; Hb – hypobranchiale; L – lacrimale; Mx – maxillare; N – nasale; Pl –palatinum; Pm – praemaxillare; Psp – postspleniale; Sp – spleniale; c. a. – commissura anterior; s. d. – sulcus dentalis; s. io. – sulcus infraorbitalis; s. m. – sulcus marginalis; s. so. – sulcus supraorbitalis; V – vomer. Масштабная шкала 10 см.
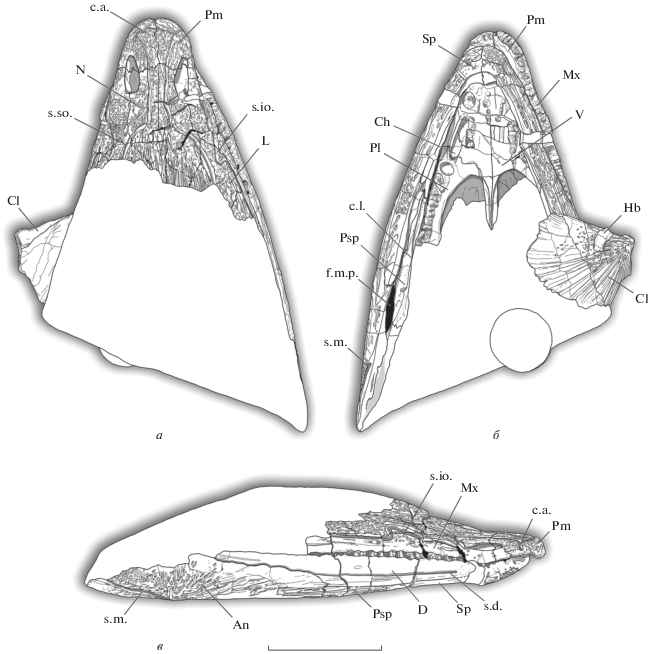
Рис. 3.
Элементы посткраниального скелета Benthosuchus lukyanovi sp. nov., голотип ПИН, № 5618/1; сверху – фотографии, снизу – прорисовки: а – левая ключица (clavicula), б – фрагмент межключицы (interclavicula). Масштабная шкала 5 см.
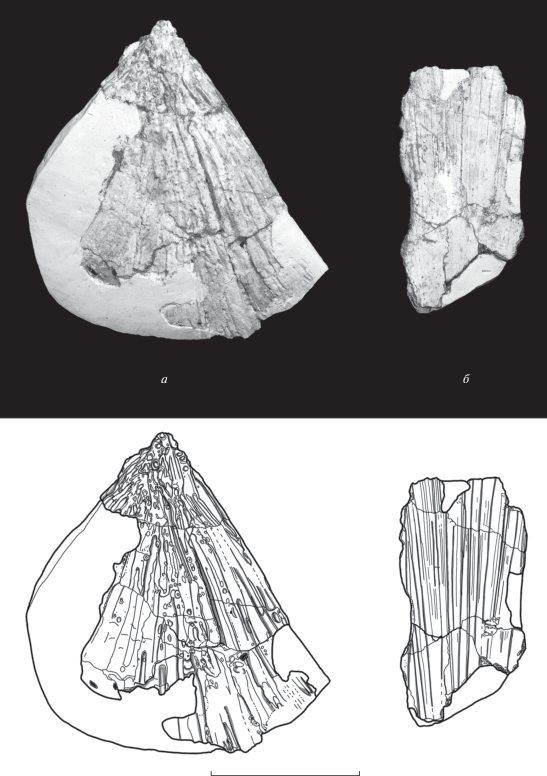
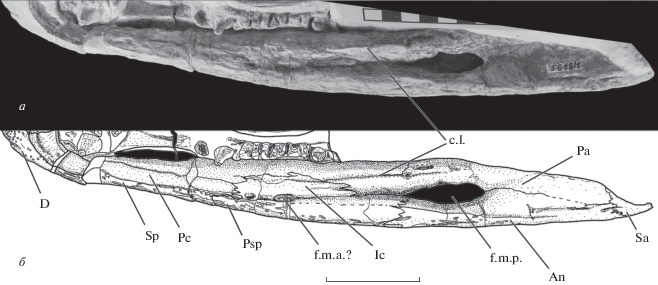
Рис. 4.
Лингвальная поверхность правой ветви нижней челюсти Benthosuchus lukyanovi sp. nov., голотип ПИН, № 5618/1: а – фотография, б – рисунок. Обозначения: An – angulare; D – dentale; Ic – intercoronoideum; Pc – praecoronoideum; Pa –praearticulare; Sa – supraangulare; Sp – spleniale; с.l. – crista lingualis; f.m.a. – foramen meckelianum anterior; f.m.p. – foramen meckelianum posterior. Масштабная шкала 5 см.
Размеры (табл. 1). Реконструированные показатели длины и ширины черепа составляют около 400 и 260 мм, соответственно. Длина хоаны – 42 мм, ширина – 12 мм. Длина ноздри – 34 мм, ширина 13 мм. Длина заднего меккелева отверстия – 42 мм, ширина – 10 мм. Расстояние между ноздрями – 31 мм. Расстояние между хоанами – 40 мм. Реконструированная длина нижней челюсти (на основе сохранившегося отдела правой нижнечелюстной ветви) ≈410 мм.
Сравнение. Новый вид отличается от других представителей рода Benthosuchus, за исключением B. sushkini (Быстров, Ефремов, 1940), более крупными размерами черепа (см. Замечания); L ≈ 400 мм, против максимального значения 302 мм у B. korobkovi, 165 мм у B. gusevae; грубой скульптурой с развитыми валиками в пренариальной зоне; сближенными супраорбитальными желобками в межноздревом промежутке; латерально ориентированными осями ноздрей (5° против среднего значения – 9° у B. sushkini, ‒ 8° у B. korobkovi и – 15° у B. gusevae); слабым ангулярным изгибом нижней челюсти (ang an – 174°, против среднего значения 161° у B. sushkini, 154° у B. korobkovi, 157° у B. gusevae); сильно вытянутым задним меккелевым отверстием (w/l fmp – 0.23, против среднего значения 0.4 у B. sushkini, 0.38 у B. korobkovi, 0.32 у B. gusevae). Еще одной особенностью нового вида является наличие гребня–валика (crista lingualis) на лингвальной поверхности нижней челюсти, подобная структура не отмечена у других представителей рода. Важным отличием от других видов является формирование задними сошниковыми отростками вертикального срединного киля, окаймленного по краям уплощенной горизонтальной зоной. В нижнечелюстном симфизе его скульптированая часть превосходит по протяженности заднюю горизонтальную пластинку в большей мере, чем у других видов рода (Lp/Ls – 0.45, против среднего значения у B. sushkini – 0.7; у B. gusevae – 0.57). От B. korobkovi и B. gusevae новый вид отличается также углом схождения ветвей нижней челюсти, показывая индекс ang Man – 51° против 63° и 73° у этих видов, соответственно. Кроме того, от B. sushkini описанный вид отличается слабо развитой передней комиссурой сенсорной системы; причем существенно, что у типового вида она наиболее хорошо выражена именно у старых особей. К подобным отличиям относится и слабое выражение вогнутости боковых контуров черепа у нового вида в нариальной области. Эта черта, напротив, обычно хорошо заметна у взрослых особей B. sushkini.
По ряду признаков наблюдается сходство описанной формы с типовым видом. Так, удлинение пренариальной зоны (w/l prn), равное 0.53, близко к диапазону у B. sushkini (0.55–0.6); угол схождения постфенестральных зубных рядов (dpf) равен 20° (у B. sushkini от 0° до 27°); удлинение ноздрей (w/l nar) равно 0.38, что соответствует среднему значению выборки типового вида.
Представляет интерес сравнение описанной формы Benthosuchus с более продвинутыми трематозавроидами. Так, сближенные вдоль сагиттальной оси (в межноздревом промежутке) супраорбитальные каналы (sulci supraorbitales) встречены у значительной части выборки тоозухида Thoosuchus yakovlevi. У типичного трематозаврида Trematosaurus brauni такое положение желобков, по-видимому, присутствует в виде нормы (Schoch, Milner, 2000; Schoch, 2019). Слабый ангулярный изгиб нижней челюсти, наблюдаемый у B. lukyanovi (ang an – 174°) (рис. 4), близок по выражению к таковому у тоозухид Thoosuchus yakovlevi и Angusaurus succedaneus (средние значения ang an – 175° и 173° соответственно), а также у позднего бентозухида (яренгиида по: Новиков, 2018) Vyborosurus mirus (ang an – 170°).
Еще одна черта сходства с трематозавроидами касается удлинения заднего меккелева отверстия нижней челюсти. Следует отметить, что исследованный образец (экз. ПИН, № 3360/9), принадлежащий Vyborosurus mirus, имеет относительно крупные размеры заднего меккелевого отверстия, при пропорциях (w/l fmp – 0.39), близких к среднему значению B. sushkini (w/l fmp – 0.4). В свою очередь, сильно вытянутое заднее меккелево отверстие B. lukyanovi (w/l fmp – 0.23) (рис. 4) в общих пропорциях сходно с Angusaurus succedaneus (ср. знач. w/l fmp – 0.22). Лингвальная сторона нижней челюсти типичного трематозаврида Trematosaurus brauni недостаточно изучена из-за особенностей сохранности (Schoch, 2019), но известны данные по близко родственному Inflectosaurus amplus, имеющему пропорции меккелевого отверстия (w/l fmp) – 0.28 (Шишкин, 1960).
Соотношение осевой длины задней пластины к скульптированной зоне нижнечелюстного симфиза (Lp/Ls) у форм с трематозавроидными тенденциями показывает слабое развитие задней пластины. Так, этот показатель у Angusaurus succedaneus равен 0.36, а у Thoosuchus yakovlevi – 0.16. По этому признаку, наиболее вероятным представляется промежуточное положение B. lukyanovi (Lp/Ls–0.45) между формами с развитой задней пластиной (B. gusevae, B. suskini) и указанными выше трематозавроидами с сокращенным вариантом.
Замечания. Поскольку речь идет о крупнейшем из известных черепов Benthosuchus, имеющем длину L ≈ 400 мм, против максимального значения ≈300 мм в основной выборке у B. sushkini и B. korobkovi (см. табл. 1), представляет большой интерес его сравнение с обломком черепной крыши очень крупной особи типового вида (экз. ПИН, № 2243/5-9), описанным Быстровым и Ефремовым (1940), для которого этими авторами предполагалась длина черепа 70–75 см. Исследованный образец был нами переизучен; анализ с использованием выборок по B. sushkini, проведенный на основе сравнения линейных промеров supratemporale относительно общих размеров черепа, показал, что его реконструированные размеры значительно меньше и близки к 40 см. Таким образом, можно предположить, что новый вид и B. sushkini характеризуются размерным сходством.
Материал. Помимо голотипа, найденный в базальной части разреза типового местонахождения гипоцентр позвонка (экз. ПИН, № 5618/2).
ОБСУЖДЕНИЕ
При оценке видовой принадлежности новой находки Benthosuchus можно отметить, что по ряду черт новая форма близка к типовому виду. Принимая во внимание это обстоятельство, мы считаем, что некоторые признаки не могут быть истолкованы как проявления индивидуальной или возрастной изменчивости у B. sushkini. Значительная часть из описанных отличий необычны для ранее известных представителей рода, и это позволяет рассматривать их как систематически значимые. В сумме сюда можно отнести:
1. Слабое выражение вогнутости боковых контуров черепа в нариальной области. Развитие этой черты, типичное для B. sushkini, становится у него хорошо заметным лишь у взрослых особей, т.е. усиливается с возрастом. Учитывая размеры описанного нами черепа, можно было бы ожидать здесь ее наибольшего развития; но она, напротив, выражена слабо, что плохо согласуется с отнесением находки к типовому виду.
2. Одной из важных отличительных особенностей B. lukyanovi, не имеющей аналогий среди доступного нам материала по ранее известным видам рода, является наличие продольного гребня–валика (crista lingualis) на praeаrticulare (рис. 4) над передней частью заднего меккелева отверстия.
3. Такой же особенностью представляется очень слабая выраженность ангулярного изгиба у описанного образца. Достоверно известно, что у взрослых особей рода указанный изгиб обычно хорошо выражен, включая и наиболее крупные образцы нижних челюстей у B. sushkini, а также и у других представителей рода.
4. Необычным отличием является относительная вытянутость заднего меккелева отверстия. У других видов, включая B. sushkini, оно имеет во взрослом состоянии форму овала с очень слабой продольной протяженностью, без ясных признаков сужения спереди.
5. Еще одной важной чертой является строение задних сошниковых отростков, которые в поперечном сечении подразделяются на восстающую медиальную часть и уплощенный (горизонтальный) внешний край. Следует, отметить, что в виде вариации слабое проявление этой особенности, ограниченное только основанием сошниковых отростков, встречено нами у одной особи из “южной расы” B. sushkini (экз. ПИН, № 2424/4), но оно не известно у особей того же вида в северных районах, а также и у других представителей рода.
6. В строении вентральной поверхности нижнечелюстного симфиза отмечается слабое развитие его заднего гладкого отдела по отношению к скульптированному, в сравнении с ранее известными видами Benthosuchus.
7. Отмечается заметное латеральное расхождение длинных осей ноздрей у B. lukyanovi по отношению к сагиттальной линии, не встречающееся у других представителей бентозухид.
Приведенные особенности представляются нам наиболее важными указаниями на принадлежность описанной формы к отдельному виду Benthosuchus. Среди рассмотренных черт нового бентозуха заслуживают специального внимания признаки, которые, независимо от оценки их видовой значимости, отражают общие тенденции в эволюции трематозавроидов. Это, во-первых, расположение передних отделов супраорбитальных желобков близко к осевой линии черепа – черта, очень частая у особей тоозухида Thoosuchus и, по-видимому, постоянная у трематозаврида Trematosaurus (Schoch, Milner, 2000; Steyer, 2002; Schoch, 2019). Еще более определенно это касается удлинения меккелева отверстия и выполаживания ангулярного угла нижней челюсти, поскольку эти признаки типичны для указанных семейств и, кроме того, встречены у Vyborosaurus (Новиков, 1994).
Можно предполагать, что такие изменения, независимо от их статуса в организации у бентозухид, возникали у тех или иных видов, а затем проявлялись у потомков с растущей частотой, становясь в итоге признаками стандартного плана строения у типичных трематозавроидных семейств. Обнаружение ряда таких черт у Benthosuchus lukyanovi расширяет наши представления о путях морфологической эволюции бентозухид и о ранних этапах становления трематозавридного типа организации.
Список литературы
Быстров А.П., Ефремов И.А. Benthosuchus sushkini Efr. – лабиринтодонт из эотриаса р. Шарженги. М.: Изд-во АН СССР, 1940. 102 с. (Тр. Палеонтол. ин-та АН СССР. Т. 10. Вып. 1).
Гетманов С.Н. Триасовые амфибии Восточно-Европейской платформы. М.: Наука, 1989. 102 с. (Тр. Палеонтол. ин-та АН СССР. Т. 236).
Ефремов И.А. О стратиграфическом подразделении континентальной перми и триаса СССР по фауне наземных позвоночных // Докл. АН СССР. Н.С. 1937. Т. 16. № 2. С. 125–132.
Ивахненко М.Ф. Новый бентозухид из нижнего триаса Верхнего Поволжья // Палеонтол. журн. 1972. № 4. С. 93–99.
Ивахненко М.Ф., Голубев В.К., Губин Ю.М. и др. Пермские и триасовые тетраподы Восточной Европы. M.: ГЕОС, 1997. 216 с. (Тр. Палеонтол. ин-та РАН. Т. 268).
Каландадзе Н.Н., Очев В.Г., Татаринов Л.П. и др. Каталог пермских и триасовых тетрапод СССР // Верхнепалеозойские и мезозойские земноводные и пресмыкающиеся СССР. М.: Наука, 1968. С. 72–92.
Кузьмин Ф.М. Примитивные черты в структуре черепа поздних стегоцефалов // Пробл. палеонтологии. 1938. Т. 4. С. 9–45.
Морковин Б.И. О развитии покровного орнамента костей черепа в онтогенезе раннетриасовых бентозухид (Amphibia, Temnospondyli) // Палеонтол. журн. 2015. № 1. С. 52–64.
Морковин Б.И. Особенности эволюционных преобразований на рубеже перми и триаса на основе анализа изменчивости капитозавроморфных лабиринтодонтов (Amphibia: Temnospondyli) Восточно-Европейской платформы // Конф. “Эволюционная и функциональная морфология позвоночных”. М.: Т-во научн. изданий КМК, 2017. С. 227–232.
Морковин Б.И. Новое местонахождение раннетриасовых наземных позвоночных в верховьях р. Юг (Вологодская область) // Бюлл. Моск. об-ва испыт. прир. 2018. Т. 93. Вып. 5–6. С. 50–55.
Новиков И.В. Биостратиграфия континентального триаса Тимано-Североуральского региона по фауне тетрапод. М.: Наука, 1994. 139 с. (Тр. Палеонтол. ин-та РАН. Т. 261).
Новиков И.В. Новые данные по трематозавроидным лабиринтодонтам Восточной Европы. 4. Род Benthosuchus Еfremov, 1937 // Палеонтол. журн. 2012. № 4. С. 68–79.
Новиков И.В. Раннетриасовые амфибии Восточной Европы: эволюция доминантных групп и особенности смены сообществ. М., 2018. 358 с. (Тр. Палеонтол. ин‑та РАН. Т. 296).
Очев В.Г. Новые данные по фауне триасовых позвоночных Оренбургского Приуралья // Докл. АН СССР. 1958. Т. 122. № 3. С. 485–488.
Очев В.Г. Систематика и филогения капитозавроидных лабиринтодонтов. Саратов: Изд-во Сарат. ун-та, 1966. 184 с.
Очев В.Г. Капитозавроидные лабиринтодонты юго-востока европейской части СССР. Саратов: Изд-во Сарат. ун-та, 1972. 269 с.
Шишкин М.А. Новый триасовый трематозаврид Inflectosaurus amplus // Палеонтол. журн. 1960. № 2. С. 130–148.
Шишкин М.А. Подотряд Stereospondyli // Основы палеонтологии. Земноводные, пресмыкающиеся, птицы. М.: Наука, 1964. С. 83–112.
Шишкин М.А. Новое семейство триасовых лабиринтодонтов Luzocephalidae //Палеонтол. журн. 1980. № 1. С. 104–119.
Шишкин М.А. Лабиринтодонты // Биостратиграфия континентального триаса Южного Приуралья. М.: Наука, 1995. С. 56–72.
Шишкин М.А., Сенников А.Г., Новиков И.В., Ильина Н.В. Дифференциация тетраподных сообществ и некоторые особенности биотических событий в раннем триасе Восточной Европы // Палеонтол. журн. 2006. № 1. С. 3–12.
Яковлев Н.Н. Триасовая фауна позвоночных из пестроцветной толщи Вологодской и Костромской губерний // Геол. вестник. 1916. Т. 2. № 4. С. 157–165.
Damiani R., Yates A. The Triassic amphibian Thoosuchus yakovlevi and the relationships of the Trematosauroidea (Temnospondyli: Stereospondyli) // Rec. Austral. Museum. 2003. V. 55. № 33. P. 331–342.
Efremov I.A. Benthosaurus sushkini, ein neuer Labyrinthodont der permotriassischen Ablagerungen der Sharschenga Flusses // Bull. Acad. Sci. USSR. 1929. № 8. S. 757–770.
Hartmann-Weinberg A.P., Kusmin F.M. Untertriadische Stegocephalen der Oka-Zna Antiklinale. Capitosaurus volgensis nov. sp. // Пробл. палеонтол. 1936. Т. 1. С. 35–61.
Romer A.S. Review of the Labyrinthodontia // Bull. Mus. Compar. Zool. 1947. V. 99. № 1. P. 3–368.
Shishkin M.A., Novikov I.V., Gubin Yu.M. Permian and Triassic temnospondyls of Russia // The Age of Dinosaurs in Russia and Mongolia / Ed. Benton M.J., Shishkin M.A., Unwin D.M., Kurochkin E.N. Cambridge: Cambr. Univ. Press, 2000. P. 35–59.
Schoch R.R. The status and osteology of two new cyclotosaurid amphibians from the Upper Moenkopi Formation of Arizona (Amphibia: Temnospondyli; Middle Triassic) // N. Jb. Geol. Paläontol. 2000. Abh. 216. P. 387–411.
Schoch R.R. Osteology of the temnospondyl Trematosaurus brauni Burmeister, 1849 from the Middle Buntsandstein of Bernburg, Germany// Palaeodiversity. Pt 12. Stuttgart, 2019. P. 41–63.
Schoch R.R., Milner A.R. Stereospondyli // Encyclopedia of Paleoherpetology. Pt 3B. München: Pfeil, 2000. P. 1–203.
Steyer J.S. The first articulated trematosaur ‘amphibian’ from the Lower Triassic of Madagascar: implication for the phylogeny of the group // Palaeontology. 2002. V. 45. Pt 4. P. 771–793.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал