

Палеонтологический журнал, 2020, № 4, стр. 70-78
Новое семейство Salwaichthyidae (Pisces, Perciformes s.l.) из нижнего олигоцена Кавказа и Карпат
А. Ф. Банников *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: aban@paleo.ru
Поступила в редакцию 28.11.2019
После доработки 13.12.2019
Принята к публикации 13.12.2019
Аннотация
Из нижнего олигоцена Кавказа (Абхазия и Ю.-З. Россия, Адыгея) и Польских Карпат по трем экземплярам описан Salwaichthys paratethyensis gen. et sp. nov., представляющий новое монотипичное семейство окунеобразных рыб Salwaichthyidae. Новое семейство характеризуется удлиненным телом с относительно небольшой головой и протяженным хвостовым стеблем, крупным ртом, раздельными спинными плавниками, многочисленными (около 55) позвонками и их ростро-каудальной сжатостью, мелкой чешуей. Salwaichthys разделяет ряд апоморфных признаков с представителями некоторых семейств окунеобразных, в особенности с Propercarinidae. Однако из-за уникальной комбинации его признаков новое семейство не может быть однозначно отнесено к какому-либо из известных подотрядов окунеобразных, и оно помещено incertae sedis среди Perciformes.
Отложения низов майкопской серии Предкавказья и Абхазии (нижний хадум, пшехский региоярус: Popov et al., 2002; Попов и др., 2019; и др.) слабо охарактеризованы малакофауной, но несут весьма обильную фауну морских рыб (Банников, Парин, 1997; Банников, 2010; Попов и др., 2019). К настоящему времени отсюда были определены свыше 70 видовых таксонов костистых, принадлежащих не менее чем 58 родам 18-ти отрядов (Банников, 2010, 2012, 2014, 2018; Bannikov et al., 2017; и др.). Кроме того, пока остаются неописанными некоторые материалы по раннемайкопским рыбам, как из давних сборов, так и собранные относительно недавно, относящиеся к новым для науки таксонам. Один из этих таксонов, описываемый ниже как Salwaichthys paratethyensis gen. et sp. nov., явно заслуживает выделения в особое семейство окунеобразных, причем из-за морфологического своеобразия положение этого семейства внутри отряда Perciformes далеко не очевидно. К новому таксону мы относим также полный скелет хорошей сохранности с противоотпечатком, найденный Г. Сальва (Польша) в нижнем олигоцене Польских Карпат.
Отряд окунеобразных (Perciformes) традиционно считался (Берг, 1940; Greenwood et al., 1966; Nelson, 2006; и др.) самым разнообразным отрядом эволюционно продвинутых костистых рыб, самым крупным из отрядов не только рыб, но и позвоночных в целом. Разные авторы выделяли внутри Perciformes различное количество подотрядов. В последнее время, однако, на смену традиционной классификации организмов зачастую приходит формальная кладистическая систематика, построенная на компьютерной обработке молекулярных данных и часто противоречащая морфологическим свидетельствам. Результат кладистического анализа не всегда отражает реальные родственные связи (Вислобокова, 2019). В кладистической систематике отряд окунеобразных подразделяют на множество отрядов, причудливо сгруппированных в таксоны более высокого ранга (Wiley, Johnson, 2010; Near et al., 2012, 2013; Betancur-R. et al., 2013, 2017; Nelson et al., 2016; и др.). Высокие таксоны кладистической иерархии морфологически гетерогенны, что признают и сами кладисты (Miya et al., 2013); обычно эти таксоны невозможно диагностировать морфологическими признаками. Это побуждает нас использовать здесь традиционную, естественную систематику.
Материал, описанный в настоящей статье, хранится в коллекции Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН) и в частной коллекции Г. Сальва (Grzegorz Salwa, Кросно, Польша); автор благодарен Г. Сальва за возможность ознакомления с его материалом. Фотографии выполнены С.В. Багировым (ПИН).
ОТРЯД PERCIFORMES
PERCIFORMES INCERTAE SEDIS
СЕМЕЙСТВО SALWAICHTHYIDAE BANNIKOV, FAM. NOV.
Типовой род – Salwaichthys gen. nov.
Диагноз. Тело удлиненное, с относительно небольшой головой и протяженным, невысоким хвостовым стеблем. Рот крупный, сочленение нижней челюсти находится позади орбиты. Зубы в челюстях среднего размера, острые, конические. Praeoperculum с ровным постеровентральным краем. Позвонков около 55, из них примерно 28–30 хвостовые. Тела позвонков короткие, сжатые ростро-каудально; остистые отростки тонкие. Средние и задние туловищные позвонки несут сильно развитые парапофизы. Hypuralia слиты в две пластинки. Четыре supraneuralia. Два спинных плавника, в промежутке между ними не менее пяти свободных птеригиофоров. Первый спинной плавник невысокий, в нем около 10 тонких колючих лучей. Начало первого спинного плавника сдвинуто каудально. Второй спинной плавник довольно высокий, несет один или два жестких и 27–28 мягких лучей. Анальный плавник начинается слегка позади мягкого спинного, он немного протяженнее его; в анальном плавнике две–три слабых колючки и около 28 мягких лучей. Грудные плавники небольшие; брюшные средней длины, расположены под грудными. Хвостовой плавник крупный, глубоко вырезанный, с 17 главными лучами. Чешуя мелкая, циклоидная.
Состав. Типовой род.
Сравнение. Комбинацией таких признаков, как удлиненное тело с относительно небольшой головой и протяженным хвостовым стеблем, крупный рот, четыре supraneuralia, раздельные спинные плавники, каудальный сдвиг начала первого спинного плавника, многочисленные позвонки, мелкая циклоидная чешуя, новое семейство отличается от всех других окунеобразных разных подотрядов. Ростро-каудальная сжатость позвонков – несомненная апоморфия нового семейства.
Род Salwaichthys Bannikov, gen. nov.
Род назван в честь Г. Сальва (Польша, Кросно) и от ichthys греч. – рыба; м.р.
Типовой вид – Salwaichthys paratethyensis sp. nov.
Диагноз. Совпадает с диагнозом семейства.
Видовой состав. Типовой вид.
Сравнение. В составе семейства один род.
Salwaichthys paratethyensis Bannikov, sp. nov.
Название вида от Паратетиса, внутриконтинентального бассейна, развитого в олигоцене на территории Европы.
Голотип – ПИН, № 3363/193, двусторонний отпечаток скелета без передней его части; Абхазия, р. Гумиста; нижний олигоцен, пшехский региоярус, эолисковые слои.
Описание (рис. 1–4). Тело удлиненное, с относительно невысоким, протяженным хвостовым стеблем. Длина головы 3.5–4.2 раза укладывается в стандартной длине тела (SL) и у взрослых уступает высоте тела, составляющей 26–33% SL. Высота хвостового стебля в 3.0–3.5 раза меньше высоты тела. Глаза умеренного размера, горизонтальный диаметр орбиты не уступает длине рыла.
Рис. 1.
Salwaichthys paratethyensis sp. nov., голотип ПИН, № 3363/193: а – отпечаток, б – противоотпечаток (длина масштабной линейки – 1 см); Абхазия, р. Гумиста; нижний олигоцен, пшехский региоярус, эолисковые слои.

Рис. 2.
Salwaichthys paratethyensis sp. nov., паратип ПИН, № 1413/831: а – отпечаток, б – противоотпечаток (длина масштабной линейки – 5 мм); Ю.-З. Россия, Адыгея, р. Белая выше ст. Абадзехской; нижний олигоцен, пшехский региоярус, эолисковые слои.

Рис. 3.
Salwaichthys paratethyensis sp. nov., экз. б/н из личной коллекции Г. Сальва: а – отпечаток, б – противоотпечаток (длина масштабной линейки – 2 см); Польские Карпаты, окрестности г. Кросно, Винница; нижний олигоцен, менилитовая свита.

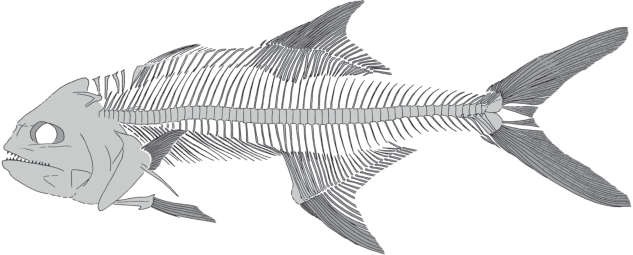
Нейрокраний довольно высокий, почти прямой крепкий парасфеноид проецируется вдоль нижнего края орбиты. Frontalia относительно широкие. Гребень supraoccipitale умеренно высокий, треугольный, закругленный на углу. Сохранность не позволяет различить подглазничные кости. Рот крупный, конечный; нижнечелюстное сочленение у взрослых расположено позади вертикали заднего края орбиты. Praemaxillare узкое, длинное, почти прямое, с невысоким восходящим отростком. Верхнечелюстные зубы конические, сохранились плохо. Maxillare узкое, удлиненное. Длина нижней челюсти составляет около 57–66% длины головы; зубы на dentale относительно крупные, конические, однорядные. Dentale протяженное, умеренно высокое, понижается у симфиза. Ось hyomandibulare слабо наклонена назад. Ectopterygoideum узкое, удлиненное, плавно изогнуто задним концом вниз. Другие птеригоидные кости сохранились плохо. Quadratum субтреугольное, относительно небольшое. Praeoperculum с длинной верхней ветвью и короткой нижней, спереди вогнуто умеренно. Предкрышка с несколькими небольшими шипами вдоль нижнего края у ювенильного экземпляра (паратип; рис. 2) и ровным нижним краем у взрослого экземпляра (рис. 3). Другие кости жаберной крышки сохранились плохо; operculum взрослого экземпляра небольшое и без шипов. В целом жаберная крышка относительно узкая. Кости гиоидного комплекса сохранились плохо, точное число radii branchiostegi неизвестно.
Позвонков 55–57. В туловищной части позвоночника не менее 25 позвонков у паратипа и 27 позвонков у экз. из Польши (начало позвоночника не сохранилось у голотипа). В хвостовой части позвоночника 29–30 позвонков. Длина туловищной части позвоночника примерно в 1.35 раза короче хвостовой его части. Тела позвонков высокие, но резко укороченные ростро-каудально на большей части позвоночника, более изометричные на хвостовом стебле. Большинство остистых отростков тонкие и относительно короткие, почти прямые; слабо изогнутые на хвостовом стебле. Передние гемальные отростки не расширены и отходят почти перпендикулярно к оси позвоночника (как и соответствующие невральные отростки), угол наклона назад остистых отростков слегка увеличивается рострально и каудально. Более половины задних туловищных позвонков (около 16) несут тонкие, почти прямые парапофизы, удлиняющиеся каудально. Ребра тонкие и относительно короткие, довольно сильно наклонены назад, занимая верхнюю половину брюшной полости. Передние ребра прикрепляются к вентральной части тел позвонков, а задние – к парапофизам. Epineuralia имеются в туловищной части позвоночника, но большая часть их скрыты телами позвонков.
Терминальный позвонок образован слиянием тел первого преурального (pu1) и уральных позвонков. Hypuralia 1–2 явно консолидированы в единую пластинку, равно как и hypuralia 3–4; гипаксиальная и эпаксиальная гипуральные пластинки разделены щелью. Гипуральная диастема широкая, но не глубокая. Parhypurale и гемальный отросток pu2 аутогенные; неясно, слит ли с телом третьего преурального позвонка гемальный отросток. Детали строения uroneuralia не прослеживаются. По-видимому, имеются три epuralia. Невральный отросток второго преурального позвонка редуцирован, тогда как невральный отросток pu3 сильный и, по-видимому, длиннее предшествующего. Хвостовой плавник крупный, глубоко вырезанный, состоит из 17 главных лучей (I8-7I). Верхние и нижние дополнительные (“procurrent”) лучи относительно немногочисленные, числом менее 10. Длина лопастей хвостового плавника достигает 30% SL у самого крупного экземпляра (рис. 3). Существенной гипуростегии не отмечается.
Между затылком и спинным плавником расположены четыре клиновидные supraneuralia умеренного размера, два первых из них предшествуют невральному отростку первого позвонка; предорсальная формула (см. Ahlstrom et al., 1976) (по экз. из Польских Карпат) – 0 + 0/0/0//1/. Колючий спинной плавник гораздо короче мягкого и отделен от него большим промежутком, длина которого почти равна длине восьми противолежащих позвонков. Первый спинной плавник отсутствует на голотипе и плохо сохранился на двух других экземплярах, но очевидно, что его шипы тонкие и недлинные, короче самых длинных мягких лучей второго спинного плавника. По-видимому, первый шип сверхштатный. Судя по числу птеригиофоров, в первом спинном плавнике имеется 11 шипов. Первое предорсальное расстояние составляет 33–40% SL, а второе предорсальное расстояние – 55–60% SL. В промежутке между спинными плавниками расположены пять свободных, не несущих лучей птеригиофоров. Высота второго спинного плавника примерно в 1.3 раза короче длины его основания, плавник начинается над последним или предпоследним туловищным позвонком. Во втором спинном плавнике один или два жестких и 27–28 мягких лучей, образующих несколько заостренную лопасть в передней части. Птеригиофоры спинных плавников узкие (первый клиновидно расширен), образуют непрерывную серию, средние из них (в начале второго спинного плавника) длиннее крайних. Вначале птеригиофоры расположены реже, их число примерно равно числу невральных отростков, расположенных под ними; более каудально птеригиофоры расположены чаще, по одному-два птеригиофора второго спинного плавника входят в промежутки между невральными отростками позвонков. Длина птеригиофоров второго спинного плавника уменьшается от среднего птеригиофора к последнему, а их наклон увеличивается.
Анальный плавник начинается слегка позади мягкого спинного, сходен с ним по форме и высоте, но в 1.09–1.17 раза более протяженный. Преанальное расстояние составляет 60–66% SL. Длина основания анального плавника превышает длину хвостового стебля. В анальном плавнике две–три слабых колючки и около 28 мягких членистых лучей. Птеригиофоры анального плавника не заходят в брюшную полость; передние из них длинные, каудально птеригиофоры укорачиваются. По одному-два птеригиофора анального плавника входят в промежутки между гемальными отростками хвостовых позвонков. Первый анальный птеригиофор самый крепкий, пластинчато расширен антеровентрально.
Грудные плавники относительно короткие, заостренные, прикрепляются непосредственно выше середины между линией позвоночника и вентральным краем тела, перед началом спинного плавника. Точное число лучей грудного плавника неизвестно; судя по экз. из Польских Карпат, их около 18. Posttemporale вильчатое. Supracleithrum удлиненное. Cleithrum крупное, прямое в нижней части, в верхней части загнуто вперед. Коракоид довольно протяженный, с прямым постеровентральным краем. Scapula и пекторальные radialia не распознаются. Вентральное postcleithrum довольно крупное, ребровидное, расположен косо; его дистальный конец расположен существенно позади basipterygium.
Брюшные плавники относительно небольшие, но слегка длиннее грудных, прикрепляются под их основанием, состоят из колючки и, очевидно, пяти ветвящихся лучей. Длина брюшных плавников составляет не более 10% SL. Тазовые кости относительно небольшие, клиновидные.
Чешуя образует сплошной покров на теле, мелкая и тонкая, покрыта концентрическими морщинками. Границы индивидуальных чешуй, ctenii и radii не распознаются. Боковая линия прослеживается плохо.
Размеры. Длина голотипа (фрагмент тела) – около 10 см. SL паратипа ПИН, № 1413/831 – 19.5 мм. SL экз. б/н из Польских Карпат – 33 см.
Возрастные изменения. У молоди относительная длина головы больше, чем у взрослых рыб, тело относительно ниже (его высота составляет 26% SL против 33% SL у взрослых), а хвостовой плавник короче (его длина составляет 20% SL против 30% SL у взрослых). Таким образом, длина головы несколько превышает высоту тела у молоди, но явно уступает ей у взрослых. Рот у взрослых рыб относительно крупнее, чем у молоди.
Сравнение. В составе рода один вид.
Замечание. Удлиненное обтекаемое тело и крупный глубоко вырезанный хвостовой плавник нового вида определенно указывают на активно плавающий образ его жизни. Косвенным свидетельством принадлежности Salwaichthys paratethyensis sp. nov. к эпипелагической группировке является редкость и в то же время широкое географическое распространение находок этого вида.
Распространение. Нижний олигоцен Кавказа (Абхазия, Адыгея) и Карпат (Польша).
Материал. Помимо голотипа, паратип ПИН, № 1413/831, двусторонний отпечаток полного скелета ювенильной рыбы с р. Белой (Адыгея), и экз. из личной коллекции Г. Сальва (Кросно, Польша), двусторонний отпечаток полного скелета взрослой рыбы из Польских Карпат.
ОБСУЖДЕНИЕ
Наличие настоящих шипов в спинном и анальном плавниках определенно указывает на то, что Salwaichthys gen. nov. относится к крупной и разнообразной кладе колючеперых костистых, надотряду Acanthopterygii (Greenwood et al., 1966) или “Series Percomorpha” (sensu Nelson, 2006). Новый таксон демонстрирует анатомические свидетельства его принадлежности к отряду Perciformes (в его традиционном понимании) – в частности, общая конфигурация его хвостового скелета, характеризующегося “пятью или менее” hypuralia и аутогенными гемальными отростками второго и третьего преуральных позвонков (Johnson, Patterson, 1993). В то же время морфологическое своеобразие нового семейства: комбинация таких признаков, как удлиненное тело с сильно увеличенным числом позвонков, длинный хвостовой стебель, четыре supraneuralia, раздельные спинные плавники, каудальный сдвиг начала первого спинного плавника, мелкая циклоидная чешуя, ростро-каудальная сжатость позвонков – делает затруднительным выявление его близких родственников и, соответственно, положения этого семейства внутри отряда Perciformes. Salwaichthyidae fam. nov. не разделяет особенностей, определяющих следующие подотряды Perciformes, выделяемые Дж. Нельсоном (Nelson, 2006): Acanthuroidei, Anabantoidei, Blennioidei, Callionymoidei, Caproidei, Channoidei, Elassomatoidei, Gobiesocoidei, Gobioidei, Icosteoidei, Kurtoidei, Labroidei, Notothenioidei, Pholidichthyoidei, Scombrolabracoidei, Trachinoidei и Zoarcoidei. С другой стороны, некоторые представители других подотрядов окунеобразных, Percoidei, Scombroidei и Stromateoidei, разделяют с новым семейством те или иные из отмеченных выше признаков.
Percoidei, “крупнейший и наиболее разнообразный из подотрядов окунеобразных, без сомнения является полифилетичным…” (Johnson, 1993, с. 15) и диагностируется несколькими плезиоморфными для Perciformes признаками; к этой группе относятся те из окунеобразных, которые не могут быть помещены в другие подотряды. В последнее время, ссылаясь на молекулярные данные, многие семейства, традиционно включавшиеся в Percoidei, выносят из этого подотряда (Nelson et al., 2016), либо вовсе ограничивают его состав двумя–тремя семействами (Betancur-R. et al., 2017). По Нельсону (Nelson, 2006), подотряд Percoidei включает в себя 79 современных семейств, и большинство из них характеризуется относительно малым числом позвонков – 23–27. Большее число позвонков (35 и более) могут иметь лишь 12 специализированных семейств перкоидов (Aplodactylidae, Bramidae, Caristiidae, Cepolidae, Congrogadidae, Echeneidae, Latrididae, Percichthyidae, Percidae, Plesiopidae, Pseudochromidae и Sillaginidae) и род Lateolabrax Bleeker (Johnson, 1984, табл. 120 ). Так много, как у нового таксона (около 55 позвонков), бывает лишь у брамид подсемейства Pteraclinae, а у Cepolinae (Cepolidae) и Congrogadidae с их сильно удлиненным телом число позвонков увеличено до 65–80. Позвонки перкоидов обычно либо удлиненные, либо субквадратные в латеральном аспекте (несколько передних позвонков могут быть укорочены). Столь сильной ростро-каудальной сжатости позвонков, как у Salwaichthys (высота тел позвонков вдвое превышает их длину), у перкоидов не отмечено.
У большинства перкоидов имеются от одного до трех супраневралий (предорсальных костей: Smith, Bailey, 1961; Johnson, 1984), иногда исчезающих при сдвиге спинного плавника вперед (Сaristiidae, Coryphaenidae, Priacanthidae, Rachycentridae), либо вовсе отсутствующих. Более трех supraneuralia (как у нового таксона) обнаружено у “…по меньшей мере, некоторых членов 13 групп перкоидов, с соответствующим сдвигом назад начала спинного плавника” (Johnson, 1984, с. 479). У подавляющего большинства перкоидов передний птеригиофор спинного плавника заходит во второй межневральный промежуток, реже наблюдается ростральный (Banjosidae, Caristiidae, Centrogenysidae, Chaetodontidae, Cheilodactylidae, Coryphaenidae, Latrididae, Priacanthidae, Rachycentridae, Scatophagidae и др.) или каудальный (Bathyclupeidae, Leptobramidae, Sillaginidae, Toxotidae, некоторые Apogonidae и Moronidae и др.) сдвиг начала спинного плавника. У нового семейства передний птеригиофор спинного плавника заходит в четвертый межневральный промежуток, то есть наблюдается каудальный сдвиг начала первого спинного плавника. Рот у перкоидов обычно менее крупный, чем у Salwaichthys, но некоторые из них (например, Leptobramidae) также имеют крупный рот. Хвостовой стебель у перкоидов обычно короткий или умеренно длинный, реже бывает столь же длинным, как у нового семейства или даже длиннее: Apogonidae, Eocottidae, Kuhliidae, Mullidae. Спинной плавник большинства перкоидов единый, с выемкой между жесткой и мягкой его частями или без выемки. У некоторых семейств Percoidei, однако, спинной плавник подразделяется на два, причем между колючим и мягким спинными плавниками обычно имеются (как у Salwaichthys) свободные (не несущие лучей) птеригиофоры.
Таким образом, каждая отдельно взятая апоморфия Salwaichthyidae, кроме сильной ростро-каудальной сжатости позвонков, может быть обнаружена у тех или иных перкоидов, что не исключает возможности отнесения нового семейства к подотряду Percoidei.
Некоторое сходство Salwaichthyidae обнаруживает с наиболее генерализованными Scombroidei – семейством Euzaphlegidae, в особенности, с миоценовым подсемейством Euzaphleginae (см. David, 1943; Банников, 1993). Это сходство заключается в наличии двух спинных плавников, удлиненном теле и относительно большом числе позвонков. Кроме того, миоценовые роды Euzaphlegidae имеют почти столь же длинный хвостовой стебель, как у Salwaichthys (David, 1943), небольшую голову и крупный рот. В то же время, остальные апоморфии Salwaichthyidae (в особенности, ростро-каудальная сжатость позвонков и наличие четырех supraneuralia) не обнаружены у каких-либо Scombroidei, что делает сомнительным наличие родства двух этих групп.
Еще один подотряд отряда Perciformes, с которым может быть сближено новое семейство – это Stromateoidei (молекулярные систематики помещают строматеевидных в их “отряд Scombriformes”: Nelson et al., 2016; Betancur-R. et al., 2017). Большинство строматеевидных рыб характеризуются тем, что передняя часть их пищевода, сразу за последней жаберной дугой, имеет боковые мешки, несущие изнутри продольные складки или озубленные сосочки (Haedrich, 1967; и др.). Наличие озубленных глоточных мешков может быть выявлено у ископаемых рыб только в исключительных случаях (Bannikov, 1995), кроме того, такие мешки отсутствуют у современных Amarsipidae. У тех или иных групп строматеевидных встречаются такие признаки Salwaichthys, как небольшая голова, удлиненное тело, раздельные спинные плавники, длинный хвостовой стебель и относительно большое число позвонков. Каудальный сдвиг начала спинного плавника, подобно Salwaichthys, наблюдается у таких строматеевидных, как Amarsipidae и некоторые Centrolophidae (Ahlstrom et al., 1976; Bannikov, 2000). Однако у строматеевидных никогда не бывает четырех supraneuralia (как у Salwaichthys) – их два–три, либо нет вовсе, как у Tetragonuridae); лишь у монотипического рода центролофовых Icichthys 7–11 предорсальных костей (Ahlstrom et al., 1976) (возможно, в их число входят как supraneuralia, так и утратившие шипы передние дорсальные птеригиофоры). Так много позвонков, как у нового таксона (около 55), из Stromateoidei бывает лишь у Tetragonuridae и рода центролофовых Icichthys, но столь сильной ростро-каудальной сжатости позвонков, как у Salwaichthys, у строматеевидных не отмечено; крупный рот также не характерен для Stromateoidei.
Более других строматеевидных новое семейство напоминают современные Amarsipidae и олигоценовые Propercarinidae (см. Bannikov, 1995; Přikryl et al., 2014) удлиненным телом с относительно длинным хвостовым стеблем, увеличенным числом позвонков (до 48 у Amarsipidae, до 36 у Propercarinidae) и раздельными спинными плавниками (у Amarsipidae слегка, у Propercarinidae примерно как у Salwaichthys). Начало спинного плавника у Amarsipidae сдвинуто каудально еще сильнее, чем у Salwaichthys, а начало анального плавника, наоборот, сдвинуто рострально – передние анальные птеригиофоры заходят в брюшную полость (Přikryl et al., 2014, рис. 9 ). Propercarinidae сходны с Salwaichthys и внешним обликом, и сильным развитием парапофизов, но позвонков у них гораздо меньше, а тела позвонков имеют нормальную для колючеперых удлиненную форму, а не резко укорочены, как у Salwaichthys. У последнего основание анального плавника несколько превышает таковое второго спинного плавника, при примерно равном числе лучей в обоих плавниках, тогда как у Propercarinidae, как и у таких строматеевидных, как Amarsipidae, и ископаемых родов, предварительно отнесенных к Centrolophidae (Bannikov, 2000; Бачу, Банников, 2004), в анальном плавнике больше лучей, чем во втором спинном. Таким образом, непосредственных указаний на родство нового семейства с подотрядом Stromateoidei не обнаружено, несмотря на сходство отдельных признаков Salwaichthys с теми или иными строматеевидными, особенно с родом Propercarina.
Формой тела, увеличенным числом позвонков и относительно длинным хвостовым стеблем Salwaichthyidae отдаленно напоминает среднеэоценовое семейство Perciformes incertae sedis Caucasichthyidae (Банников и др., 2011) с Кавказа. Однако существенные различия исключают возможность родства упомянутых семейств: у Caucasichthyidae позвонки не укорочены, как у Salwaichthyidae, и их значительно меньше (не более 40); имеется крупный шип на praeoperculum; чешуя относительно крупная и преимущественно спиноидная; нет supraneuralia; брюшные плавники удлиненные; и т.д.
Таким образом, анализ распределения некоторых апоморфных особенностей Salwaichthyidae показывает крайнюю проблематичность выявления родственных отношений этого семейства. Приведенное выше обсуждение не предоставило убедительных свидетельств принадлежности Salwaichthyidae к какому-либо из известных подотрядов Perciformes (в традиционном понимании отряда). Salwaichthys разделяет некоторые апоморфные признаки с определенными семействами перкоидов, скумбриевидных и строматеевидных. Тем не менее, уникальная комбинация признаков препятствует адекватному установлению места нового семейства внутри какого-либо из существующих подотрядов окунеобразных, и оно рассматривается incertae sedis среди Perciformes (в традиционном понимании этого отряда).
Отнесение к одному видовому таксону кавказского (неполный скелет взрослой и полный скелет ювенильной рыбы) и польского (полный скелет крупной рыбы) материала по Salwaichthys достаточно условно. Только находка сопоставимого с польским по полноте и размеру экземпляра на Кавказе подтвердит или опровергнет их конспецифичность.
Список литературы
Банников А.Ф. Позднеэоценовый вид Palimphyes (Teleostei, Perciformes) из Северного Кавказа // Палеонтол. журн. 1993. № 2. С. 72–78.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. LXI + 244 с.
Банников А.Ф. Первая находка рода Isurichthys (Perciformes, Ariommatidae) в нижнем олигоцене Северного Кавказа // Палеонтол. журн. 2012. № 2. С. 61–67.
Банников А.Ф. Новый род семейства Palaeocentrotidae (Teleostei, Lampridiformes) из олигоцена Северного Кавказа и замечания о других ископаемых Veliferoidei // Палеонтол. журн. 2014. № 6. С. 50–58.
Банников А.Ф. Новый род и вид строматеевидных рыб (Perciformes, Stromateoidei) из нижнего олигоцена Северного Кавказа // Палеонтол. журн. 2018. № 6. С. 41–49.
Банников А.Ф., Карневале Дж., Парин Н.В. Новое семейство Caucasichthyidae (Pisces, Perciformes) из эоцена Северного Кавказа // Палеонтол. журн. 2011. № 1. С. 75–81.
Банников А.Ф., Парин Н.Н. Список морских рыб из кайнозойских (верхний палеоцен–средний миоцен) местонахождений юга европейской части России и сопредельных стран // Вопр. ихтиол. 1997. Т. 37. № 2. С. 149–161.
Бачу Д.-С., Банников А.Ф. Новые строматеевидные рыбы (Perciformes, Stromateoidei) из нижнего олигоцена Румынии // Вопр. ихтиол. 2004. Т. 44. № 2. С. 162–170.
Берг Л.С. Система рыбообразных и рыб, ныне живущих и ископаемых // Ежегодн. Зоол. музея АН СССР. 1940. Т. 5. С. 87–517.
Вислобокова И.А. О новых подходах изучения эволюции макротаксонов и концепции экона // Палеонтол. журн. 2019. № 1. С. 3–12.
Попов С.В., Табачникова И.П., Банников А.Ф. и др. Лектостратотип майкопской серии по р. Белая выше г. Майкопа (Западное Предкавказье) в его олигоценовой части // Стратигр. Геол. корреляция. 2019. Т. 27. № 3. С. 70–92.
Ahlstrom E.H., Butler J.L., Sumida B.Y. Pelagic stromateoid fishes (Pisces, Perciformes) of the eastern Pacific: kinds, distributions and early life histories and observations on five of these from the northwest Atlantic // Bull. Mar. Sci. 1976. V. 26. № 3. P. 285–402.
Bannikov A.F. Morphology and phylogeny of fossil stromateoid fishes (Perciformes) // Geobios. 1995. Mém. spec. № 19. P. 177–181.
Bannikov A.F. A new genus and species of putative centrolophid fish (Perciformes, Stromateoidei) from the Eocene of Bolca, northern Italy // Boll. Mus. Civ. Stor. Natur. Verona. Geol. Paleontol. Preist. 2000. V. 24. P. 37–46.
Bannikov A.F., Carnevale G., Popov Y.A. An extraordinary pipefish (Teleostei, Syngnathidae) with fully developed anal fin from the Oligocene of the North Caucasus (SW Russia) // Boll. Soc. Paleontol. Ital. 2017. V. 56. № 1. P. 79–88. https://doi.org/10.4435/BSPI.2017.08
Betancur-R. R., Broughton R.E., Wiley E.O. et al. The tree of life and a new classification of bony fishes // PLOS Currents Tree of Life. 2013. P. 1–41.
Betancur-R. R., Wiley E.O., Arratia G. et al. Phylogenetic classification of bony fishes // BMC Evol. Biol. 2017. 17:162. P. 1–40. https://doi.org/10.1186/s12862-017-0958-3
David L.R. Miocene fishes of Southern California // Geol. Soc. Amer. Spec. Pap. 1943. № 43. P. 1–143.
Greenwood P.H., Rosen D.E., Weitzman S.H., Myers G.S. Phyletic studies of teleostean fishes, with a provisional classification of living forms // Bull. Amer. Mus. Natur. Hist. 1966. V. 131. Art. 4. P. 339–455.
Haedrich R.L. The stromateoid fishes: systematics and a classification // Bull. Mus. Comp. Zool. Harv. Univ. 1967. V. 135. № 2. P. 31–139.
Johnson G.D. Percoidei: development and relationships // Ontogeny and systematics of fishes / Eds Moser H.G., Richards W.J., Cohen D.M. et al. Lawrence: Allen Press, 1984. P. 464–498 (Amer. Soc. Ichthyol. Herpetol. Spec. Publ. № 1).
Johnson G.D. Percomorph phylogeny: progress and problems // Bull. Mar. Sci. 1993. V. 52. № 1. P. 3–28.
Johnson G.D., Patterson C. Percomorph phylogeny: a survey of acanthomorphs and a new proposal // Bull. Mar. Sci. 1993. V. 52. № 1. P. 554–626.
Miya M., Friedman M., Satoh T.P. et al. Evolutionary origin of the Scombridae (tunas and mackerels): Members of a Paleogene adaptive radiation with 14 other pelagic fish families // PLOS One. 2013. V. 8. № 9. P. e73535:1–19.
Near T.J., Dornburg A., Eytan R.I. et al. Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes // Proc. Nat. Acad. Sci. USA. 2013. V. 110. P. 12738–12743.
Near T.J., Eytan R.I., Dornburg A. et al. Resolution of ray-finned fish phylogeny and timing of diversification // Proc. Nat. Acad. Sci. USA. 2012. V. 109. P. 13698–13703.
Nelson J.S. Fishes of the World. 4 ed. Hoboken, New Jersey: John Wiley and Sons, 2006. 601 p.
Nelson J.S., Grande T.C., Wilson M.V.H. Fishes of the World. 5th ed. Hoboken, NJ: Wiley, 2016. 707 p.
Popov S.V., Akhmetiev M.A., Bugrova E.M. et al. Biogeography of the Northern Peri-Tethys from the Late Eocene to the Early Miocene. Part 2. Early Oligocene // Paleontol. J. 2002. V. 36. Suppl. 3. P. 185–259.
Přikryl T., Bannikov A.F., Grădianu I. et al. Revision of the family Propercarinidae (Perciformes, Stromateoidei) with description of a new species from the Oligocene of the Carpathians // C. R. Palevol. 2014. T. 13. Fasc. 8. P. 691–700.
Smith C.L., Bailey R.M. Evolution of the dorsal-fin supports of percoid fishes // Pap. Mich. Acad. Sci. Arts Lett. 1961. V. 46. P. 345–361.
Wiley E.O., Johnson G.D. A teleost classification based on monophyletic groups // Origin and Phylogenetic Interrelationships of Teleosts / Eds Nelson J.S., Schultze H.-P., Wilson M.V.H. Munich: Verl. Dr. F. Pfeil, 2010. P. 123–182.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал