

Журнал общей биологии, 2023, T. 84, № 2, стр. 127-143
Оценка динамики флоры орхидных (Orchidaceae: Magnoliophyta) России на основании сравнения материалов, накопленных до 1951 г. и в 1961–2010 гг.
Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
* E-mail: efimov@binran.ru
** E-mail: efimov81@mail.ru
Поступила в редакцию 03.10.2022
После доработки 03.11.2022
Принята к публикации 15.11.2022
- EDN: RCUXNF
- DOI: 10.31857/S0044459622060057
Аннотация
Динамика флор является перспективным направлением исследований в ботанике. Работы в этой области имеют практическое значение для диагностики сокращающихся видов, позволяют выявлять конкретные факторы, влияющие на численность растений, и в перспективе разрабатывать меры по сохранению биологического разнообразия in situ. В настоящей статье под динамикой понимается изменение числа местонахождений, т.е. количественные изменения флор. Рассмотрены количественные изменения в составе флоры, выявляемые при сравнении массива данных до 1951 г. и в 1961–2010 гг. Диагностированы виды со статистически значимым сокращением или ростом числа местонахождений, у части видов статистически значимые тенденции не выявляются. Динамика рассматривается как по России в целом, так и отдельно по крупным частям страны, причем случаи, когда у видов выявляется несогласованная динамика числа местонахождений в различных частях страны, редки. Рассматриваются перспективы дальнейших исследований в этой области, в том числе возможности применения разработанной методики к другим группам сосудистых растений России.
Флористический состав растений любой местности претерпевает со временем изменения. Наиболее известными изменениями флоры являются вселение новых, чужеродных элементов и вымирание редких аборигенных видов. Основными причинами вымирания видов называются исчезновение подходящих местообитаний, сбор людьми и климатические изменения (Ceballos et al., 2015). Глобальное потепление, одно из наиболее значимых климатических изменений нашего времени, существенно усилилось в последние десятилетия и наиболее сильно проявляется в северных районах планеты – в арктической и бореальной областях (Masson-Delmotte et al., 2019) – и, таким образом, имеет большое влияние на флору России.
Однако, помимо качественных изменений состава флоры – вселения и вымирания видов, – повсеместно происходят и количественные изменения, выражающиеся в изменении числа местонахождений видов, обозначаемые как прогресс и регресс. Оценка количественных изменений флоры также важна, так как дает наиболее точные представления о характере изменений состава флоры, позволяет изучать причины этих изменений и прогнозировать будущее флоры. Реально используемое практическое приложение сведений о количественных изменениях флоры – оценки уязвимостей видов. Например, сведения по динамике заложены в четырех (A, B, C и E) из пяти критериев IUCN (International Union for Conservation of Nature and Natural Resources – Международный союз охраны природы и природных ресурсов), используемых для категоризации видов по категориям уязвимости (IUCN, 2012).
Изучение количественных изменений флоры сложнее, чем фиксация изменений качественных, так как для этого требуется математический анализ на репрезентативных выборках. Оптимальными для таких исследований являются данные специализированных учетов, как это было осуществлено на материале флоры Великобритании (Braithwaite et al., 2006) и некоторых других стран, либо на материалах национальных баз данных (Tamis et al., 2005; Piessens, Hermy, 2006, и др.).
Изменение числа местонахождений видов сосудистых со временем, именуемое также динамикой числа местонахождений, или для краткости просто динамикой, и является предметом рассмотрения в данной работе. Для России основной тип информации, который имеется для анализа динамики числа местонахождений видов сосудистых растений в исторической перспективе – данные из гербарных коллекций. Ранее нами был разработан сравнительно простой подход к использованию данных такого типа на материале орхидных северо-запада Европейской части России (Efimov, 2010, 2011). Целью настоящего исследования является представить результаты изучения динамики числа местонахождений видов всего семейства орхидных в масштабах всей нашей страны в целом, а также оценить перспективы подобных расчетов для других семейств сосудистых растений.
Орхидные являются хорошим модельным объектом для исследований динамики флоры в связи с их высокой природоохранной значимостью. Также можно считать, что их систематика на сегодняшний день изучена достаточно подробно (имеется новый обзор – Efimov, 2020). К орхидным традиционно наблюдается повышенный интерес как в научной среде, так и со стороны любителей природы, поэтому по данному семейству накоплен довольно значительный объем материала во многих гербарных коллекциях нашей страны.
Насколько можно судить, на настоящий момент исследования динамики флоры России с использованием математических методов не осуществлялись. Существующие оценки динамики, представленные, например, в Красных книгах, являются преимущественно экспертными оценками без статистического анализа, на основании сравнения абсолютных значений числа местонахождений, зарегистрированных в различные периоды времени. Такие оценки вполне адекватны для редких видов, известных из единичных точек, и перспективы такого анализа достаточно велики (Решетникова, 2016), но для более широко распространенных таксонов требуется разработка специальных методов с использованием математической статистики.
Исследуемый в работе интервал времени нацелен на выявление усредненных тенденций в динамике флоры, имеющих место на протяжении XX в. и в сопредельные десятилетия. Не позволяя оценить реакцию флоры на наиболее сильные климатические изменения, связываемые уже с XXI в., наше сравнение охватывает данные за период, когда антропогенное влияние на климат планеты неоднозначно, и период – приблизительно с 1960 г., – когда это влияние считается уже несомненным (Brönnimann et al., 2008). Есть достаточно много данных, показывающих значимость середины XX в. для сопоставлений, связанных с климатом. Так, после 1960 г. наблюдается устойчивый рост средней температуры поверхности Земли без свидетельств цикличности процесса в масштабе десятилетий, наблюдавшихся ранее (Climate change…, 2020). Данные по зимним температурам говорят, что до 1945 г. потепление достаточно локально сказывалось в Северной Америке и в Северной Атлантике, существенно не затрагивая территорию России, в то время как после 1950 г. заметно начало сказываться по всей арктической и бореальной зоне Северного полушария (Jones, Moberg, 2003; Brönnimann et al., 2008).
Отметим, что в России имеется давняя практика популяционных исследований, хорошо разработанных в методическом отношении (Динамика ценопопуляций…, 1985; Заугольнова и др., 1988, и др.). Однако динамику числа местонахождений вида как такового нельзя оценивать на основании динамики числа особей в конкретных, даже нескольких, популяциях. Популяционная динамика во многом зависит от изменения локальных условий произрастания и может не соответствовать общей динамике числа местонахождений вида. Поэтому в данной работе популяционные методы не используются, хотя остаются применимыми для не изучавшихся нами самых редких таксонов, у которых можно напрямую оценить общее число особей и их изменение во всех или хотя бы в большинстве известных популяций.
МАТЕРИАЛЫ И МЕТОДЫ
Техническая реализация исследования основана на анализе базы данных, содержащей сведения о местонахождениях орхидных России, аналогичной той, которая составлялась ранее для работ по орхидным северо-запада Европейской части России (Efimov, 2010, 2011). Основные поля, по которым заносился материал для каждой записи, были следующие: название таксона; ранее присвоенный индивидуальный номер (баркод, номер записи в иных базах данных или иное); регион (область, край и т.п. – если обозначен); местонахождение; дата обнаружения в природе; автор находки; координаты (если исходно были указаны); тип данных (гербарий/литература/наблюдение). Для сведений на основе гербария вносился также номер, проставленный коллектором, и акроним гербарной коллекции. В процессе работы с базой данных региональную принадлежность и географические координаты (с точностью до минут) определяли по возможности для каждой записи. Учитывая общий объем данных, на момент анализа включавший около 62 000 записей, определение географических координат представляло собой наиболее трудоемкий процесс, в особенности для исторических сведений, и было успешным для 92% записей.
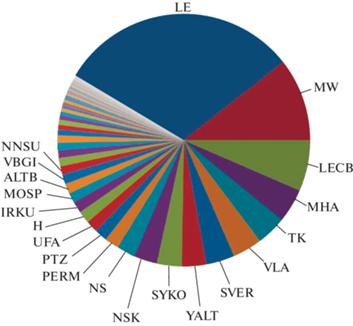
Основное наполнение базы данных осуществляли на основе гербарных коллекций, к моменту анализа были учтены материалы из 59 полностью или почти полностью обработанных фондов (рис. 1), из которых в “Index Herbariorum” (https://sweetgum.nybg.org/science/ih/) зарегистрированы следующие (даны в порядке значимости вклада в рабочую базу данных): LE, MW, LECB, MHA, TK, VLA, SVER, YALT, SYKO, NSK, NS, PERM, PTZ, UFA, H, IRKU, MOSP, ALTB, VBGI, NNSU, KFTA, UDU, IRK, LENUD, KW, KUZ, SIMF, CSR, CSAU, HERZ, KPABG, PSK, PHEO, CNR, UUH, TVBG, KBAI, PKM, PVB, KBHG, DAG, ORIS, KHA, SAKH, SARAT, WIR. Помимо записей на основе гербария, учитывались также данные из литературы, а также вполне достоверные данные непосредственных наблюдений в природе, сделанные как самостоятельно, так и коллегами. Точки с ранее опубликованных карт, а также такие литературные данные, которые могли быть основаны на неверных определениях (например, большая часть информации из литературы по видам р. Dactylorhiza), не учитывались. Определение всех использованных в работе гербарных образцов проверялось – либо при непосредственной работе с гербарной коллекцией, либо по цифровым фотоснимкам.
Рис. 1.
Сравнительный состав материалов из различных гербариев в рабочей базе данных “Орхидные России”.
Расчеты выполнялись в целом по методике, разработанной ранее при исследовании динамики числа местонахождений орхидных северо-запада Европейской части России (Efimov, 2010, 2011). В связи с разной точностью указания местонахождений, использованных в работе (например, на гербарных образцах 100-летней давности с очень грубыми привязками и новых образцах с указанием географических координат места сбора), при сравнениях оперировали не непосредственными числами известных местонахождений видов, а числами сеточных ячеек, в которых вид фиксировался в сравниваемые периоды времени. Размер сеточной ячейки был задан 0.1° в широтном и 0.2° в долготном направлении, что, по-видимому, близко к оптимальному значению размера ячейки, при котором возможна удовлетворительная геопривязка старого материала и не слишком сильное уменьшение объема исходной информации при огрублении изначально более точно привязанных данных. Длина стороны сеточной ячейки сопоставима с приблизительным размером суточного перехода исследователя (5–15 км), во время которого могут фиксироваться множественные точки, являющиеся избыточными при данном анализе. Для Кавказа и Крыма площадь ячеек была взята в 4 раза меньше, для того чтобы получить больше ячеек по сборам с этих территорий, отличающихся малой величиной зоны, где встречаются орхидные. Например, в Крыму, который сам по себе меньше по площади всех остальных рассматриваемых в статье частей страны, орхидные отсутствуют на большей части территории, где доминируют аридные экосистемы, и приурочены почти исключительно к горным районам в южной части полуострова. К тому же, уже при предварительных расчетах выяснилось, что динамика числа местонахождений орхидных в Крыму и на Кавказе, как правило, менее выражена, чем в других частях страны, и для того, чтобы здесь выявить хоть какие-то тенденции, нужно больше исходных данных. В то же время крымский и кавказский материал зачастую удается привязывать точнее, что делает использование более мелких ячеек вполне возможным.
Подсчет чисел сеточных ячеек в сравниваемые периоды времени осуществлялся автоматизированным способом при помощи специально разработанной программы в форме надстройки к MS Excel. При этом учитывались даже такие записи, для которых год находки не был точно известен и был определен в форме интервала – при условии, если интервал полностью попадал в период, по которому осуществлялся подсчет.
Сравнение чисел сеточных ячеек, занимаемых видом в два сравниваемых интервала времени, осуществляли для каждого вида отдельно с использованием критерия хи-квадрат для попарных сравнений, по формуле
Для вычисления ожидаемых чисел сеточных ячеек учитывали различную интенсивность исследований в сравниваемые интервалы времени, что является обязательным условием при проведении подобных исследований (Prendergast et al., 1993; Braithwaite et al., 2006; McInerny et al., 2006; Rich, 2006; Rich, Karran, 2006; Petřík et al., 2010, и др.). Рич и Карран (Rich, Karran, 2006) как раз на примере одного из видов орхидных – Neottia cordata (L.) Rich. – продемонстрировали, насколько сильно могут отличаться от реальности оценки, полученные без учета фактора различной интенсивности исследований в различные периоды времени. В настоящем исследовании интенсивность исследований оценивали исходя из общего числа накопленных сеточных точек суммарно по всем видам орхидных в соответствующие периоды времени, по формулам
Методику расчета можно пояснить на следующем условном примере. Если общее число сеточных ячеек по всем видам орхидных вместе в первый из сравниваемых интервалов времени было, допустим, в 3 раза выше, чем во второй (интенсивность исследований отличается в 3 раза), то соотношение наблюдаемых чисел сеточных ячеек в эти периоды для конкретного вида, равное 3 : 1 (например, 36 ячеек до 1951 г. и 12 ячеек в период 1961–2010 гг.), будет свидетельствовать об отсутствии динамики числа местонахождений у этого вида (значение критерия χ2 будет равно нулю). При наличии расхождения с соотношением 3 : 1 критерий хи-квадрат покажет статистическую значимость различия. Подтвержденным факт изменения числа местонахождений видов считали, если выявлялась не менее чем 90%-ная значимость отличий, но большая часть выявленной динамики подтверждается с 99%-ной и более высокой степенью значимости.
В реальности массивы данных, накопленные в сравниваемые в данном исследовании интервалы времени (т.е. до 1951 г. и в 1961–2010 гг.), имели, как правило, приблизительно одинаковый объем. Так, например, по России в целом суммарно по всем исследованным в работе видам орхидных в эти периоды было зафиксировано 14 159 и 18 120 ячеек соответственно (в период с 1961 по 2010 г. “интенсивность исследований” была всего в 1.28 раза больше, чем до 1951 г.).
Единично встречающиеся виды, зафиксированные в сравниваемые интервалы времени в пределах рассматриваемой территории из 10 ячеек и менее, исключались из расчетов как недостаточно репрезентативные случаи.
В настоящей работе представлена как общая оценка динамики числа местонахождений видов орхидных на территории всей России, так и региональные оценки, осуществленные по отдельности на независимых выборках из общей базы данных для восьми частей страны: Кавказа, Крыма, северо-запада Европейской части, севера Европейской части, центра Европейской части, Урала (включая Тюменскую, Курганскую и Омскую области), Сибири, Дальнего Востока. При региональных оценках динамики интенсивность исследований учитывалась только по соответствующей части страны. Например, для дальневосточного вида орхидных интенсивность исследований имеет значение только в пределах Дальнего Востока.
Указанные выше значения интенсивности исследований в целом по России (14159 и 18120 ячеек) использовались в качестве значений N1 и N2 только для тех видов, которые в России широко распространены, а именно, встречаются во всех указанных выше частях страны не единично (с тем же порогом – 10 сеточных ячеек). Таких видов всего пять: Coeloglossum viride, Corallorhiza trifida, Dactylorhiza incarnata, Goodyera repens и Gymnadenia conopsea. При расчетах динамики числа местонахождений остальных, менее широко распространенных видов интенсивность исследований оценивалась только по тем частям страны, где вид присутствует во флоре, причем также не единично.
Данные за 10 лет между выбранными периодами (с 1951 по 1959 г.) исключены из рассмотрения, чтобы снизить зависимость чисел сеточных ячеек с местонахождениями видов в сравниваемые периоды друг от друга. Период после 2010 г. исключен из анализа в связи с тем, что сведения о местонахождениях приобретают особый характер из-за большого числа данных, опубликованных на краудсорсинговых платформах (iNaturalist и Plantarium), а также целенаправленной фиксации точек автором настоящей работы.
Поскольку критерий хи-квадрат является неполярным, его значение ничего не говорит о направлении изменения числа местонахождений вида (прогресс не отличается от регресса). Направленность изменений оценивалась на основе коэффициента k, принимавшего значение k > 1 при прогрессе и 0 < k < 1 при регрессе. Его вычисляли следующим образом:
РЕЗУЛЬТАТЫ
В связи с выбраковкой редких видов, анализ был осуществлен для 105 видов орхидных флоры России. В последнем чек-листе орхидных России (Efimov, 2020) общее разнообразие орхидных России было оценено в 135 видов; таким образом, использованная методика оказалась применима для 78% видов семейства.
Оценки чисел сеточных ячеек в сравниваемые интервалы времени представлены в табл. 1, а рассчитанные на их основе значения критерия хи-квадрат – в табл. 2.
Таблица 1.
Оценка чисел сеточных ячеек
| Вид | Вся Россия | Кавказ | Крым | Северо-запад Европейской части | Север Европейской части | Центр Европейской части | Урал | Сибирь | Дальний Восток | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | до 1951 г. | 1961–2010 | |
| Anacamptis coriophora (L.) R.M. Bateman, Pridgeon & M.W. Chase | 42 | 28 | 14 | 8 | 17 | 8 | 11 | 12 | ||||||||||
| A. laxiflora (Lam.) R.M. Bateman, Pridgeon & M.W. Chase | 30 | 27 | 4 | 7 | 20 | 10 | 6 | 10 | ||||||||||
| A. morio (L.) R.M. Bateman, Pridgeon & M.W. Chase | 95 | 101 | 33 | 38 | 62 | 63 | ||||||||||||
| A. pyramidalis (L.) Rich. | 93 | 92 | 41 | 35 | 52 | 57 | ||||||||||||
| Calypso bulbosa (L.) Oakes | 172 | 185 | 51 | 37 | 9 | 2 | 29 | 43 | 51 | 60 | 32 | 43 | ||||||
| Cephalanthera damasonium (Mill.) Druce | 65 | 96 | 22 | 31 | 43 | 65 | ||||||||||||
| C. longibracteata Blume | 1 | 16 | 1 | 16 | ||||||||||||||
| C. longifolia (L.) Fritsch | 70 | 91 | 18 | 41 | 28 | 34 | 21 | 7 | 3 | 9 | ||||||||
| C. rubra (L.) Rich. | 143 | 156 | 42 | 42 | 47 | 45 | 30 | 17 | 24 | 52 | ||||||||
| Coeloglossum viride (L.) Hartm. | 570 | 734 | 66 | 42 | 10 | 12 | 63 | 42 | 142 | 243 | 66 | 24 | 71 | 89 | 110 | 161 | 42 | 121 |
| Corallorhiza trifida Châtel. | 433 | 705 | 12 | 17 | 9 | 11 | 103 | 74 | 105 | 191 | 65 | 69 | 41 | 77 | 79 | 170 | 19 | 96 |
| Cremastra appendiculata (D. Don) Makino | 1 | 13 | 1 | 13 | ||||||||||||||
| Cypripedium calceolus L. | 566 | 646 | 8 | 4 | 64 | 56 | 84 | 124 | 107 | 120 | 76 | 121 | 168 | 162 | 59 | 59 | ||
| C. guttatum Sw. | 570 | 572 | 19 | 19 | 33 | 18 | 95 | 87 | 322 | 343 | 101 | 105 | ||||||
| C. macranthos Sw. | 431 | 390 | 61 | 20 | 267 | 257 | 103 | 113 | ||||||||||
| C. shanxiense S.C. Chen | 7 | 11 | 7 | 11 | ||||||||||||||
| C. yatabeanum Makino | 22 | 41 | 22 | 41 | ||||||||||||||
| Dactylorhiza aristata (Fisch. ex Lindl.) Soó | 56 | 118 | 56 | 118 | ||||||||||||||
| D. baltica (Klinge) Nevski | 34 | 262 | 24 | 180 | 10 | 82 | ||||||||||||
| D. euxina (Nevski) Czerep. | 56 | 64 | 56 | 64 | ||||||||||||||
| D. fuchsii (Druce) Soó | 719 | 1189 | 73 | 188 | 97 | 265 | 178 | 193 | 138 | 168 | 233 | 375 | ||||||
| D. iberica (M. Bieb. ex Willd.) Soó | 17 | 10 | 17 | 10 | ||||||||||||||
| D. incarnata (L.) Soó | 785 | 993 | 21 | 16 | 10 | 14 | 119 | 170 | 108 | 163 | 193 | 212 | 137 | 154 | 190 | 258 | 7 | 6 |
| D. maculata (L.) Soó | 425 | 578 | 150 | 215 | 142 | 207 | 107 | 103 | 26 | 53 | ||||||||
| D. psychrophila (Schltr.) Aver. | 52 | 64 | 52 | 64 | ||||||||||||||
| D. romana (Sebast.) Soó | 39 | 56 | 20 | 33 | 19 | 23 | ||||||||||||
| D. saccifera (Brongn.) Soó | 14 | 19 | 14 | 19 | ||||||||||||||
| D. salina (Turcz. ex Lindl.) Soó | 93 | 119 | 9 | 7 | 84 | 112 | ||||||||||||
| D. sibirica Efimov | 37 | 109 | 37 | 109 | ||||||||||||||
| D. traunsteineri (Saut. ex Rchb.) Soó | 95 | 116 | 26 | 23 | 53 | 75 | 12 | 5 | 4 | 13 | ||||||||
| D. urvilleana (Steud.) H. Baumann & Künkele | 55 | 88 | 55 | 88 | ||||||||||||||
| Epipactis atrorubens (Hoffm.) Besser | 176 | 270 | 25 | 23 | 53 | 105 | 25 | 41 | 73 | 101 | ||||||||
| E. condensata Boiss. ex D.P. Young | 6 | 8 | 6 | 8 | ||||||||||||||
| E. helleborine (L.) Crantz | 551 | 680 | 36 | 48 | 51 | 46 | 84 | 95 | 43 | 58 | 219 | 210 | 75 | 135 | 43 | 88 | ||
| E. krymmontana Kreutz, Fateryga & Efimov | 10 | 7 | 10 | 7 | ||||||||||||||
| E. microphylla (Ehrh.) Sw. | 16 | 15 | 16 | 15 | ||||||||||||||
| E. palustris (Mill.) Crantz | 346 | 376 | 20 | 14 | 8 | 4 | 73 | 107 | 32 | 36 | 111 | 118 | 49 | 47 | 53 | 50 | ||
| E. papillosa Franch. & Sav. | 32 | 69 | 32 | 69 | ||||||||||||||
| E. persica (Soó) Hausskn. ex Nannf. | 20 | 25 | 12 | 14 | 8 | 11 | ||||||||||||
| E. thunbergii A. Gray | 10 | 17 | 10 | 17 | ||||||||||||||
| Ephippianthusschmidtii Rchb.f. | 23 | 31 | 23 | 31 | ||||||||||||||
| Epipogium aphyllum Sw. | 153 | 191 | 17 | 12 | 22 | 15 | 36 | 29 | 7 | 15 | 23 | 23 | 32 | 71 | 16 | 26 | ||
| Galearis cyclochila (Franch. & Sav.) Soó | 17 | 21 | 17 | 21 | ||||||||||||||
| Gastrodia elata Blume | 6 | 21 | 6 | 21 | ||||||||||||||
| Goodyera repens (L.) R.Br. | 624 | 1055 | 25 | 32 | 10 | 8 | 84 | 132 | 114 | 216 | 110 | 120 | 71 | 126 | 138 | 282 | 72 | 139 |
| Gymnadenia conopsea (L.) R.Br. | 1195 | 1123 | 104 | 103 | 17 | 16 | 134 | 111 | 149 | 249 | 174 | 78 | 160 | 114 | 326 | 320 | 131 | 132 |
| Habenaria linearifolia Maxim. | 84 | 61 | 84 | 61 | ||||||||||||||
| Hammarbya paludosa (L.) O. Kuntze | 91 | 164 | 32 | 57 | 20 | 44 | 26 | 22 | 6 | 22 | 7 | 7 | 0 | 12 | ||||
| Herminium monorchis (L.) R.Br. | 463 | 274 | 23 | 10 | 52 | 23 | 8 | 0 | 89 | 14 | 44 | 18 | 201 | 176 | 46 | 33 | ||
| Himantoglossum caprinum (M. Bieb.) Spreng. | 25 | 34 | 25 | 34 | ||||||||||||||
| H. comperianum (Steven) P. Delforge | 11 | 17 | 11 | 17 | ||||||||||||||
| Limodorum abortivum (L.) Sw. | 47 | 80 | 14 | 30 | 33 | 50 | ||||||||||||
| Liparis japonica (Miq.) Maxim. | 28 | 60 | 28 | 60 | ||||||||||||||
| L. kumokiri F. Maek. | 5 | 42 | 5 | 42 | ||||||||||||||
| L. loeselii (L.) Rich. | 47 | 59 | 7 | 19 | 22 | 14 | 11 | 16 | 7 | 10 | ||||||||
| L. makinoana Schltr. | 12 | 23 | 12 | 23 | ||||||||||||||
| Malaxis monophyllos (L.) Sw. | 219 | 376 | 9 | 13 | 39 | 41 | 86 | 159 | 17 | 21 | 20 | 39 | 14 | 44 | 34 | 59 | ||
| Neolindleya camtschatica (Cham.) Nevski | 6 | 11 | 6 | 11 | ||||||||||||||
| Neotinea tridentata (Scop.) R.M. Bateman, Pridgeon & M.W. Chase | 468 | 697 | 24 | 37 | 23 | 25 | 136 | 184 | 71 | 182 | 151 | 162 | 33 | 72 | 30 | 35 | ||
| N. ustulata (L.) R.M. Bateman, Pridgeon & M.W. Chase | 29 | 37 | 29 | 37 | ||||||||||||||
| Neottia acuminata Schltr. | 345 | 557 | 93 | 80 | 44 | 59 | 79 | 105 | 30 | 66 | 73 | 180 | 26 | 67 | ||||
| N. camtschatea (L.) Spreng. | 22 | 41 | 22 | 41 | ||||||||||||||
| N. cordata (L.) Rich. | 25 | 27 | 25 | 27 | ||||||||||||||
| N. nidus-avis (L.) Rich. | 12 | 43 | 12 | 31 | 0 | 12 | ||||||||||||
| N. nipponica (Makino) Szlach. | 339 | 420 | 46 | 65 | 31 | 34 | 60 | 73 | 12 | 21 | 155 | 137 | 28 | 71 | 7 | 19 | ||
| N. ovata (L.) Bluff & Fingerh. | 29 | 29 | 29 | 29 | ||||||||||||||
| N. papilligera Schltr. | 75 | 79 | 38 | 35 | 37 | 44 | ||||||||||||
| N. puberula (Maxim.) Szlach. | 96 | 42 | 17 | 19 | 21 | 4 | 24 | 7 | 34 | 12 | ||||||||
| Ophrys apifera Huds. | 8 | 5 | 8 | 5 | ||||||||||||||
| O. insectifera L. | 15 | 12 | 10 | 6 | 5 | 6 | ||||||||||||
| O. mammosa Desf. | 12 | 24 | 5 | 13 | 7 | 11 | ||||||||||||
| O. oestrifera M. Bieb. | 32 | 46 | 8 | 20 | 24 | 26 | ||||||||||||
| Orchis mascula (L.) L. | 69 | 112 | 25 | 55 | 36 | 41 | 8 | 16 | ||||||||||
| O. militaris L. | 368 | 229 | 31 | 40 | 21 | 12 | 29 | 21 | 76 | 24 | 40 | 17 | 171 | 115 | ||||
| O. pallens L. | 13 | 10 | 13 | 10 | ||||||||||||||
| O. provincialis Balb. ex Lam. & DC. | 14 | 13 | 3 | 7 | 11 | 6 | ||||||||||||
| O. punctulata Steven ex Lindl. | 21 | 28 | 7 | 18 | 14 | 10 | ||||||||||||
| O. purpurea Huds. | 73 | 80 | 23 | 20 | 50 | 60 | ||||||||||||
| O. simia Lam. | 49 | 62 | 11 | 13 | 38 | 49 | ||||||||||||
| Oreorchis patens (Lindl.) Lindl. | 25 | 48 | 25 | 48 | ||||||||||||||
| Platanthera bifolia (L.) Rich. | 746 | 930 | 11 | 14 | 127 | 219 | 107 | 174 | 226 | 187 | 136 | 172 | 139 | 164 | ||||
| P. chlorantha (Cust.) Rchb. | 192 | 274 | 69 | 77 | 46 | 47 | 24 | 98 | 53 | 52 | ||||||||
| P. chorisiana (Cham.) Rchb.f. | 16 | 81 | 16 | 81 | ||||||||||||||
| P. convallariifolia Lindl. | 15 | 40 | 15 | 40 | ||||||||||||||
| P. densa Freyn | 57 | 72 | 4 | 10 | 53 | 62 | ||||||||||||
| P. fuscescens (L.) Kraenzl. | 95 | 95 | 27 | 25 | 68 | 70 | ||||||||||||
| P. hologlottis Maxim. | 89 | 83 | 14 | 7 | 75 | 76 | ||||||||||||
| P. komarovii Schltr. | 14 | 24 | 14 | 24 | ||||||||||||||
| P. metabifolia F. Maek. | 31 | 65 | 31 | 65 | ||||||||||||||
| P. oligantha Turcz. | 16 | 58 | 8 | 27 | 8 | 31 | ||||||||||||
| P. ophrydioides F. Schmidt | 5 | 8 | 5 | 8 | ||||||||||||||
| P. sachalinensis F. Schmidt | 2 | 16 | 2 | 16 | ||||||||||||||
| P. tipuloides (L.f.) Lindl. | 60 | 134 | 60 | 134 | ||||||||||||||
| P. ussuriensis (Regel & Maack) Maxim. | 18 | 17 | 18 | 17 | ||||||||||||||
| Pogonia japonica Rchb.f. | 43 | 31 | 43 | 31 | ||||||||||||||
| Ponerorchis chusua (D. Don) Soó | 13 | 3 | 13 | 3 | ||||||||||||||
| P. cucullata (L.) X.H. Jin, Schuit. & W.T. Jin | 221 | 297 | 47 | 29 | 35 | 51 | 98 | 168 | 41 | 49 | ||||||||
| Pseudorchis albida (L.) Á. Löve & D. Löve | 26 | 47 | 26 | 47 | ||||||||||||||
| Serapias orientalis (Greuter) H. Baumann & Künkele | 5 | 9 | 5 | 9 | ||||||||||||||
| Spiranthes australis (R.Br.) Lindl. | 256 | 307 | 9 | 4 | 138 | 152 | 109 | 151 | ||||||||||
| S. spiralis (L.) Chevall. | 3 | 11 | 3 | 11 | ||||||||||||||
| Steveniella satyrioides (Spreng.) Schltr. | 20 | 29 | 6 | 13 | 14 | 16 | ||||||||||||
| Traunsteinera globosa (L.) Rchb. | 16 | 9 | 16 | 9 | ||||||||||||||
| T. sphaerica (M. Bieb.) Schltr. | 50 | 40 | 50 | 40 | ||||||||||||||
Таблица 2.
Динамика числа местонахождений флоры орхидных России (в ячейках проставлены значения критерия хи-квадрат)
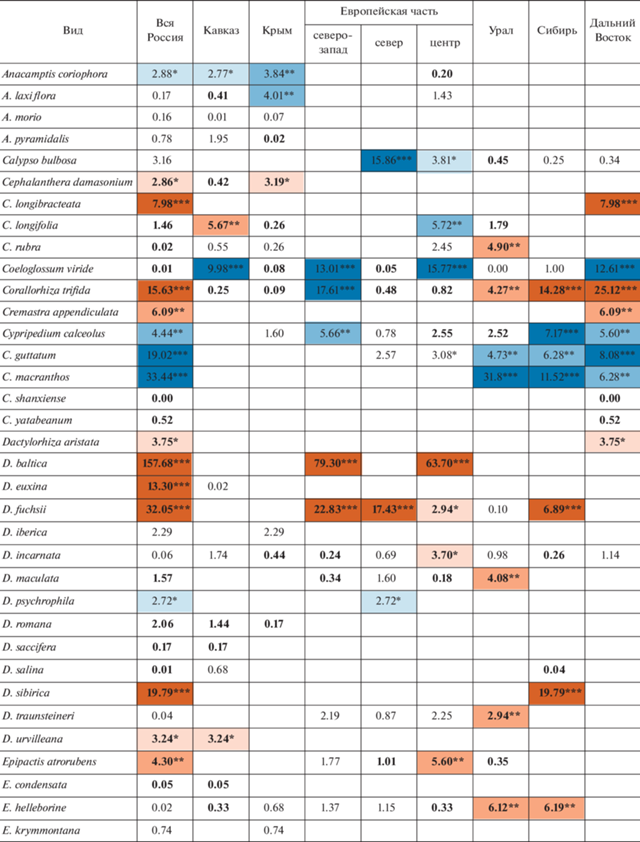
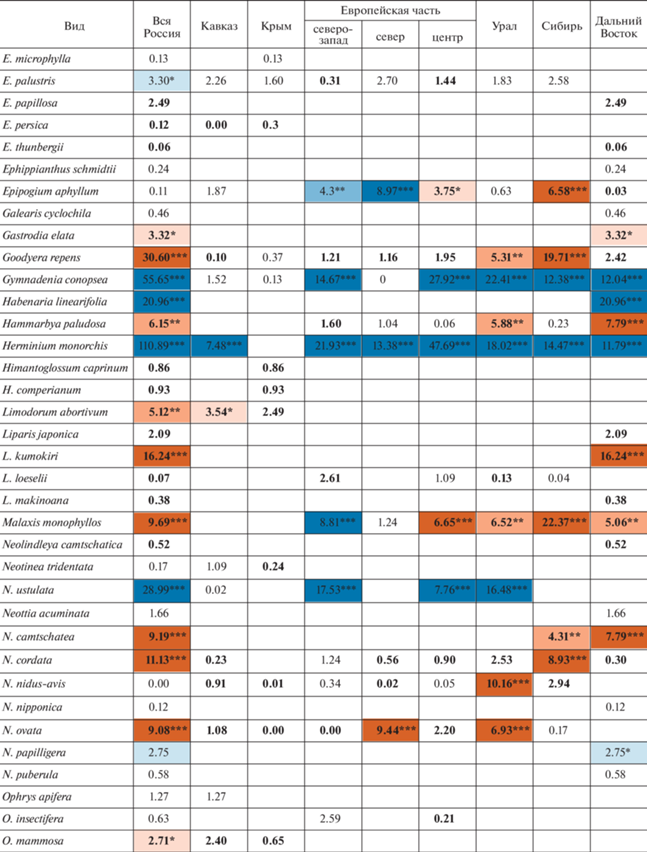
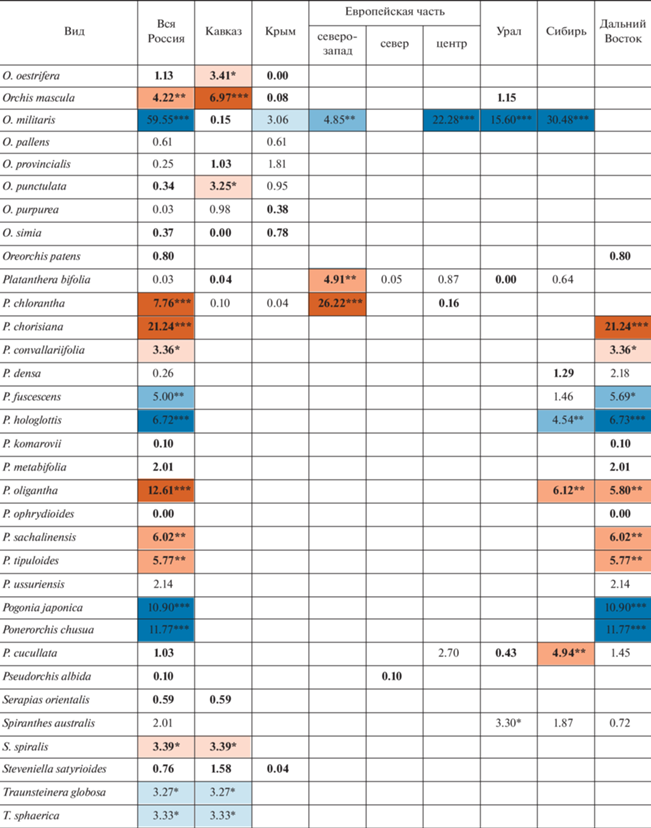
Можно видеть, что в целом по стране статистически значимый регресс в рассмотренный период времени показан у 18 видов. Из них с вероятностью 99% число местонахождений сокращается у 10: Cypripedium guttatum Sw., C. macranthos Sw., Gymnadenia conopsea (L.) R.Br., Habenaria linearifolia Maxim., Herminium monorchis (L.) R.Br., Neotinea ustulata (L.) R.M. Bateman, Pridgeon & M.W. Chase, Orchis militaris L., Platanthera hologlottis Maxim., Pogonia japonica Rchb.f. и Ponerorchis chusua (D. Don) Soó. К этому списку можно добавить Coeloglossum viride (L.) C. Hartm., проявляющий регресс во многих отдельно взятых частях страны. Положительная динамика числа местонахождений отмечена у 27 видов, из которых с вероятностью 99% – у 15: Cephalanthera longibracteata Blume, Corallorhiza trifida Châtel., Dactylorhiza baltica (Klinge) Nevski, D. euxina (Nevski) Czerep., D. fuchsii (Druce) Soó, D. sibirica Efimov, Goodyera repens (L.) R.Br., Liparis kumokiri F. Maek., Malaxis monophyllos (L.) Sw., Neottia camtschatea (L.) Spreng., N. cordata (L.) Rich., N. ovata (L.) Bluff & Fingerh., Platanthera chlorantha (Cust.) Rchb., P. chorisiana (Cham.) Rchb.f., P. oligantha Turcz.
В работе выявляется определенная региональная специфика динамики. Однако видов, у которых в пределах России выявляется динамика числа местонахождений разной направленности, всего четыре (табл. 2): Cephalanthera longifolia (L.) Fritsch характеризуется ростом числа местонахождений на Кавказе и сокращением их числа в центральных областях Европейской части; Corallorhiza trifida проявляет регресс в областях северо-запада Европейской части и прогресс в восточной части страны, начиная с Урала (Урал, Сибирь, Дальний Восток); Epipogium aphyllum Sw. проявляет регресс в областях северо-запада и севера Европейской части и прогресс в областях центра Европейской части и в Сибири; динамика числа местонахождений Malaxis monophyllos сходна с Corallorhiza trifida, но прогресс этого вида дополнительно отмечается еще и в центре Европейской части. В противовес этим случаям имеется 19 видов, у которых выявляется однонаправленная статистически значимая динамика числа местонахождений в двух или более частях страны. Из них наиболее показательно наличие шести широко распространенных видов, у которых статистически значимая динамика числа местонахождений выявляется синхронно в четырех и более частях страны (регресс у Coeloglossum viride, Cypripedium guttatum, Gymnadenia conopsea, Herminium monorchis, Orchis militaris и прогресс у Dactylorhiza fuchsii). Таким образом, динамику числа местонахождений отдельно взятых видов в масштабах страны в основном можно считать синхронной.
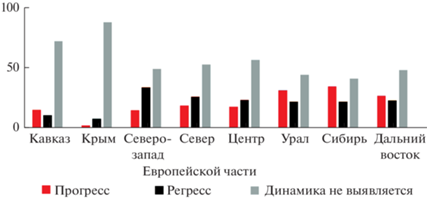
Если сравнивать различные части нашей страны между собой (рис. 2), то наибольший процент сокращающихся видов наблюдается на северо-западе и в центре Европейской части России, а в Азиатской России (Урал, Сибирь, Дальний Восток) он немного ниже. Видов с растущим числом местонахождений, напротив, в Азиатской части страны существенно больше, чем в Европейской. Кавказ, Крым и север Европейской части характеризуются большим процентом видов, у которых динамика числа местонахождений не выявляется, следовательно, можно утверждать, что в этих районах она по меньшей мере более слабая, чем в прочих регионах.
ОБСУЖДЕНИЕ
Полученные оценки динамики числа местонахождений 78% видов семейства орхидных позволяют говорить о том, что использованный метод – эффективный инструмент для оценки количественных изменений флоры сосудистых растений России. Оставшиеся 22% – наиболее редкие виды орхидных, у которых слишком мало сеточных ячеек для корректных сравнений. Но для оценки динамики числа местонахождений таких видов статистические методы и не столь необходимы, здесь может быть достаточно сведений о том, сохранился или нет вид в конкретных точках, или информации об изменении числа особей в имеющихся популяциях (т.е. к ним можно применять популяционные методы, упомянутые ранее).
Таким образом, основной категорией сосудистых растений, для которой использованный метод остается не подходящим, являются наиболее редкие виды. Также спорным является возможность применения методики к микровидам из недостаточно изученных групп, по которым мало достоверно определенного материала (Taraxacum, Hieracium, Ranunculus и др.).
Сопоставление полученных данных со сведениями, полученными другими авторами, затруднительно в связи с тем, что ранее имелись в основном экспертные оценки для конкретных видов, последовательные данные по динамике числа местонахождений растений какой-либо местности очень немногочисленны. Статистический анализ динамики, как указывалось ранее, не осуществлялся вовсе – ни по орхидным, ни по другим видам сосудистых растений флоры России. Однако можно отметить отсутствие явных противоречий полученных данных с имеющимися в литературе предположительными оценками. Так, из 10 видов орхидных, сокращение которых по нашим данным статистически наиболее значимо, 7 внесены в Красную книгу РФ (2008), по многим из них приводятся сведения о регрессе, по крайней мере в Европейской части (Аверьянов, 1999, 2000; Мамаев и др., 2004; Вахрамеева и др., 2014). Особенно часто говорится о сокращении Herminium monorchis и Neotinea ustulata, а также о сокращении Cypripedium guttatum и C. macranthos в Европейской части. Отметим, что H. monorchis и по нашим данным характеризуется статистически наиболее значимым регрессом по России в целом (табл. 2).
Блинова (Blinova, 2008) интерпретирует данные об изменении численности особей в 21 популяции 10 видов орхидных Мурманской области в 1992–2004 гг. в контексте климатических изменений. Как говорилось ранее, популяционные методы позволяют оценить динамику конкретных популяций, которая не обязательно отражает динамику числа местонахождений вида в целом из-за особенностей локальных условий. По данным Блиновой, из исследованных ею видов достоверный положительный тренд был выявлен в популяциях Cypripedium calceolus L. и Platanthera bifolia (L.) Rich., а отрицательный – у Neottia ovata, Coeloglossum viride и Dactylorhiza incarnata (L.) Soó. Наши данные по северу Европейской части сильно отличаются: у перечисленных видов динамика числа местонахождений не выявляется, за исключением N. ovata, показавшего, наоборот, статистически значимый положительный тренд. Такое сильное различие оценок связано с указанным принципиальным отличием использованных методик, а также разницей периодов времени, по которым имеются данные, и возможной спецификой Мурманской области (наши данные – по северу Европейской части в целом).
А.П. Серегин (2014) по результатам сеточного картирования Владимирской области констатирует предполагаемое исчезновение Herminium monorchis, Neottia cordata, Liparis loeselii (L.) Rich. Наши данные по центру Европейской части свидетельствуют о регрессе только первого из них. Здесь уже нет принципиального различия в методах (в обеих работах сопоставляется число сеточных ячеек), но также имеет место региональная специфика, разница в периодах наблюдений, да и просто фактор случайности при находках редких видов растений.
Указанные примеры показывают сложность сопоставления полученных нами данных с результатами других авторов. Сопоставление с нашими предшествующими оценками динамики числа местонахождений орхидных по северо-западу Европейской России (Efimov, 2010, 2011), напротив, малоинтересны из-за того, что в обеих работах использована одна методика и очень схожие исходные данные. Отличия сводятся к тому, что ранее рассматривались несколько иные периоды времени (1700–1949 и 1950–2009 гг.), а также был задействован несколько меньший объем исходных данных (с течением времени база данных непрерывно пополняется). В раннем исследовании не был выявлен статистически значимый прогресс у Platanthera bifolia (сейчас он подтвержден на уровне значимости 95–99%), а у вида Neottia nidus-avis (L.) Rich., напротив, был отмечен регресс на таком же уровне значимости (сейчас он не подтверждается). Также в раннем исследовании отмечается более высокая статистическая значимость регресса у видов Cypripedium calceolus, Dactylorhiza traunsteineri (Saut. ex Rchb.) Soó, Epipactis atrorubens (Hoffm.) Bess. и E. helleborine (L.) Crantz.
В последние годы по редким видам Ponerorchis chusua и Cypripedium macranthos нами были даны прямые оценки изменения числа местонахождений со временем (по C. macranthos – только по Европейской части), показавшие, что число местонахождений обоих видов сократилось во много раз по сравнению с первой половиной XX в. и что оба вида находятся на грани вымирания (Ефимов и др., 2022; Efimov et al., 2022). Эти сведения вполне согласуются с оценками регресса этих видов, что также неудивительно в силу того, что эти оценки сделаны на одних и тех же исходных данных. Однако они показательны как визуализация имеющихся тенденций.
Определенные сопоставления могут быть сделаны с зарубежными территориями. Так, все пять выявленных нами наиболее сокращающихся видов орхидных, представленных в Западной Европе, практически повсеместно считаются сокращающимися или вымирающими, что говорит о сходстве динамики числа местонахождений этих видов по всему их ареалу. Herminium monorchis в Европе в целом характеризуется как один из наиболее уязвимых видов семейства (Pridgeon et al., 2001; Štípková, Kindlmann, 2015; Kull et al., 2016). Coeloglossum viride в горных районах и в высоких широтах не считается уязвимым, но в равнинных областях, напротив, оказывается одним из наиболее сильно сократившихся (Eccarius, 2016), в Германии число его местонахождений упало на 80% (Eccarius, 2016), в Великобритании – на 46% с 1930-х годов (Fay et al., 2015). Neotinea ustulata также почти повсеместно в Европе считается быстро сокращающимся видом (Kretzschmar et al., 2007), возможно, наиболее быстро вымирающим видом орхидных Великобритании (Preston et al., 2002). Gymnadenia conopsea в Западной Европе также сокращается, хотя численность его не опустилась до такого предела, чтобы он расценивался как уязвимый в целом (Meekers et al., 2012). Orchis militaris в целом считается также сокращающимся (Kretzschmar et al., 2007), в Великобритании число его местонахождений уменьшилось во много раз (Farrell, 1985). В сопредельной Эстонии наиболее сильно сократившимися, на основании карт ареалов, можно считать Coeloglossum viride и Herminium monorchis (Kurbel, Hirse, 2017). Сопоставление с картами ареалов дает и ряд других соответствий, но без математического анализа остается субъективным критерием.
Сопоставления с конкретными зарубежными исследованиями динамики числа местонахождений затруднительны как из-за различия в подходах, так и неоднородности условий разных стран, характера антропогенного преобразования местности. Так как одним из важных факторов, влияющих на динамику флоры, является изменение климата, можно ожидать различной реакции на его изменение в различных частях ареала, откуда следует необходимость корректировок при сопоставлении данных по удаленным территориям. Так, исследование изменения числа местонахождений орхидных стран Бенелюкса (Vogt-Schilb et al., 2015), по-видимому, наиболее корректно сравнивать с нашими данными по Крыму и Кавказу. Из изученных в обоих исследованиях видов регресс совпадает у четырех (Anacamptis coriophora (L.) R.M. Bateman, Pridgeon & M.W. Chase, A. laxiflora (Lam.) R.M. Bateman, Pridgeon & M.W. Chase, Herminium monorchis, Traunsteinera globose (L.) Rchb.), а также в этот список может быть добавлен вид Cypripedium calceolus, регресс которого не был нами отмечен в России в изученный интервал времени, но был зафиксирован позже, так как к 2019 г. сохранилось всего одно местонахождение этого вида в Крыму (Фатерыга, 2019). Прогресс орхидных в странах Бенелюкса и на Кавказе или в Крыму не совпадает ни у одного вида, но и противоположная динамика числа местонахождений выявлена всего у одного таксона, у Spiranthes spiralis (L.) Chevall. – регресс в странах Бенелюкса и прогресс на российском Кавказе. Последний вид известен сильным регрессом в северной части ареала – в северной части Великобритании и Франции, Бельгии и Польше практически вымер, при том что в южной части ареала сократился не столь сильно (Jacquemyn, Hutchings, 2010).
Исследование динамики числа местонахождений орхидных Дании (Damgaard et al., 2020), выполненное на огромном массиве популяционных данных за последние десятилетия, можно сравнить с нашими данными по северо-западу Европейской части, наиболее близкому территориально. В Дании примерно у половины видов был выявлен регресс и только у двух – прогресс. Совпадает динамика числа местонахождений (отрицательная) у Corallorhiza trifida, Gymnadenia conopsea, Herminium monorchis и Orchis militaris и не совпадает (прогресс на северо-западе Европейской части и регресс в Дании) у Dactylorhiza fuchsii и Platanthera bifolia. Таким образом, здесь отмечается сходство динамики только на уровне наиболее сильно сокращающихся видов.
По зарубежной Азии данных по динамике меньше, и основываться можно только на списках охраняемых видов, которые, помимо видов с отрицательной динамикой, включают также единично встречающиеся таксоны. Так, по орхидным Монголии в новейшем ранжировании видов по IUCN сведения по динамике не были привлечены вовсе (Baasanmunkh et al., 2021), за исключением сообщения о предположительном вымирании Calypso bulbosa (L.) Oakes. Список охраняемых видов Японии (Global…, 2020) включает 12 видов, динамика числа местонахождений которых нами изучалась, из них на Дальнем Востоке у семи выявлен регресс (Cypripedium calceolus, C. guttatum, C. macranthos, Herminium monorchis, Neottia papilligera Schltr., Platanthera fuscescens (L.) Kraenzl., Pogonia japonica), еще у двух регресс выявлен в других частях страны (Calypso bulbosa, Epipogium aphyllum), но есть и три прогрессирующих таксона (Hammarbya paludosa (L.) O. Kuntze, Platanthera chorisiana, P. convallariifolia (Fisch. ex Lindl.) Lindl.). Таким образом, параллели в динамике числа местонахождений орхидных Дальнего Востока России и Японии также имеют место.
ЗАКЛЮЧЕНИЕ
Сходство динамики исследованных видов в различных частях России является аргументом в пользу корректности полученных оценок, так как для анализа динамики одного и того же вида в различных регионах использовались независимые, никак не связанные друг с другом массивы исходных данных. В этом же убеждают и параллели с литературными данными, имеющиеся главным образом для наиболее сильно регрессирующих таксонов.
Мы считаем, что использованный метод можно рекомендовать для изучения динамики и других видов флоры России. Однако для территорий, где имеются более полные данные о распространении видов, приводящие к высокой повторности регистрации вида в одних и тех же сеточных ячейках, представленный метод может оказаться неприменимым. Дело в том, что если в пределах сеточных ячеек вид фиксируется многократно, то динамика числа местонахождений начнет сказываться на изменении числа занятых сеточных ячеек с сильным запозданием, только тогда, когда вид начнет уже полностью выпадать из флоры отдельных ячеек. В России проблема повторности пока мало актуальна. Слабое покрытие территории исследованиями отражает тот факт, что число необследованных ячеек на порядок может превышать число ячеек с точками видов. Неэффективность метода для детально изученных территорий была ранее показана при применении его к массиву данных по Эстонии – параллели с динамикой числа местонахождений орхидных сопредельного северо-запада Европейской части оказались незначительны (Efimov, 2011).
Существенное препятствие для применения использованной методики для других групп сосудистых растений, отличных от орхидных, – трудоемкость сбора исходных данных. Проблема, во-первых, в том, что требуется привлечение данных из большого числа гербарных коллекций страны. Во-вторых, крайне длительный и кропотливый этап обработки материалов – это их геопривязка. В будущем постепенный перевод гербарных коллекций России в цифровую форму с представлением в геопривязанном формате существенно облегчит такие исследования и может сделать их даже рутинными.
Также можно отметить, что, пока не появится большой объем уже ставших историческими краудсорсинговых данных по России, при исследованиях динамики числа местонахождений любых групп сосудистых растений всегда будет ощущаться существенный недостаток исходных данных. До какой-то степени можно пытаться преодолевать эту проблему, подключая к анализу дополнительные данные из необследованных ранее гербарных коллекций, но в этом можно достаточно быстро достичь предела. Проблему недостатка данных можно преодолевать и путем увеличения географического охвата, однако здесь возникает вопрос о возможности различной динамики числа местонахождений одного и того же вида в различных частях его ареала. Для столь обширной территории, как Россия, эта проблема весьма существенна, что и отражает факт разнородной динамики числа местонахождений в различных частях ареала, выявленный в настоящей работе. Если подразделять территорию страны более дробно в широтном направлении, то не исключено, что географическая разнородность динамики числа местонахождений может проявиться еще более явно. Например, на северной и южной границах ареала можно ожидать даже разнонаправленную динамику, если имеет место смещение ареала как реакция на климатические изменения, например, на глобальное потепление.
Наконец, в силу специфики исходных данных, использованный в работе метод может выявлять артефактную (ложноположительную или ложноотрицательную) динамику числа местонахождений видов, имеющих ярко выраженное неравномерное распространение по территории. Примером может быть ложноположительная динамика числа местонахождений видов, произрастающих в удаленных районах, приходящаяся на периоды экспедиционных исследований, когда флора этих районов исследуется более детально. Или ложноположительная динамика числа местонахождений видов верховых болот в период активизации болотоведческих исследований. Впрочем, подобные артефакты представляются все-таки не очень значимыми, так как они сглаживаются за счет больших площадей изучаемых территорий. Диагностика таких артефактов пока затруднительна из-за отсутствия независимых критериев проверки. В первом приближении маркером возможных артефактов можно считать единичный случай выявления особой динамики числа местонахождений в пределах ареала вида. В настоящем исследовании таких примеров немного.
Недостатком представленного метода можно считать и то, что из-за дефицита данных на коротких отрезках времени приходится оперировать массивами данных, собранными на больших интервалах времени (от 50 лет). За столь долгий период динамика числа местонахождений видов может быть нелинейна, выявляется же только усредненная тенденция. Выявление более тонких особенностей динамики числа местонахождений видов теоретически возможно путем разбивки имеющегося массива данных не на два, а на большее число интервалов, которые можно анализировать множественными попарными сравнениями, либо привлекать регрессионные модели для их анализа. Данные методики не являются предметом обсуждения в настоящей статье и при этом также имеют свои недостатки. Наши предварительные оценки показали, что по многим видам данных для регрессионного анализа недостаточно. Тем не менее выявление более тонких деталей динамики числа местонахождений очень важно, так как позволит шире изучать возможные причины наблюдаемой динамики, например, путем поиска корреляций с изменяющимися климатическими факторами. В дальнейшем анализ динамики числа местонахождений орхидных России будет продолжен.
Список литературы
Аверьянов Л.В., 1999. Род башмачок – Cypripedium (Orchidaceae) на территории России // Turczaninowia. Т. 2. Вып. 2. С. 5–40.
Аверьянов Л.В., 2000. Орхидные (Orchidaceae) Средней России // Turczaninowia. Т. 3. Вып. 1. С. 30–53.
Вахрамеева М.Г., Варлыгина Т.И., Татаренко И.В., 2014. Орхидные России: биология, экология и охрана. М.: Т-во науч. изд. КМК. 437 с.
Динамика ценопопуляций растений, 1985 / Под ред. Серебрякова Т.И. М.: Наука. 208 с.
Ефимов П.Г., Прокопенко С.В., Якубов В.В., 2022. Ponerorchis chusua (Orchidaceae): систематика и распространение в России // Бот. журн. Т. 107. № 5. С. 433–440. https://doi.org/10.31857/S0006813622050015
Заугольнова Л.Б., Жукова А.А., Комарова А.С., Смирнова О.В., 1988. Ценопопуляции растений (очерки популяционной биологии). М.: Наука. 184 с.
Красная книга Российской Федерации (растения и грибы), 2008 / Под ред. Бардунова Л.В., Новикова В.С. М.: Т-во науч. изд. КМК. 855 с.
Мамаев С.А., Князев М.С., Куликов П.В., Филиппов Е.Г., 2004. Орхидные Урала: систематика, биология, охранный статус. Екатеринбург: УрО РАН. 124 с.
Решетникова Н.М., 2016. Динамика флоры средней полосы Европейской части России за последние 100 лет на примере Калужской области. Автореф. дисс. … докт. биол. наук. М.: ГБС РАН. 47 с.
Серегин А.П., 2014. Флора Владимирской области: анализ данных сеточного картирования. М.: Т-во науч. изд. КМК. 441 с.
Фатерыга В.В., 2019. Современное состояние ценопопуляции Cypripedium calceolus (Orchidaceae) в Крыму // Экосистемы. № 20. С. 76–81.
Baasanmunkh S., Oyuntsetseg B., Efimov P., Tsegmed Z., Vandandorj S. et al., 2021. Orchids of Mongolia: Taxonomy, species richness and conservation status // Diversity. V. 13. № 7. https://doi.org/10.3390/d13070302
Blinova I., 2008. Populations of orchids at the northern limit of their distribution (Murmansk Oblast): Effect of climate // Russ. J. Ecol. V. 39. № 1. P. 26–33. https://doi.org/10.1134/S1067413608010050
Braithwaite M.E., Ellis R.W., Preston C.D., 2006. Change in the British Flora 1987–2004. L.: BSBI. 382 p.
Brönnimann S., Ewen T., Luterbacher J., Diaz H.F., Stolarski R.S., Neu U., 2008. A focus on climate during the past 100 years // Climate Variability and Extremes during the Past 100 Years (Adv. Global Change Res. V. 33). Dordrecht: Springer. P. 1–25. https://doi.org/10.1007/978-1-4020-6766-2_1
Ceballos G., Ehrlich P.R., Barnosky A.D., García A., Pringle R.M., Palmer T.M., 2015. Accelerated modern human-induced species losses: Entering the sixth mass extinction // Sci. Adv. V. 1. № 5. https://doi.org/10.1126/sciadv.1400253
Climate change, 2020. Evidence and causes: Update 2020. Washington: National Acad. Press. 24 p. https://doi.org/10.17226/25733
Damgaard C., Moeslund J., Wind P., 2020. Changes in the abundance of Danish Orchids over the past 30 years // Diversity. V. 12. № 6. https://doi.org/244.10.3390/d12060244
Eccarius W., 2016. Die Orchideengattung Dactylorhiza. Eisenach: Selbstverlag des Verfassers. 639 p.
Efimov P.G., 2010. The dynamics of Orchids of NW European Russia // J. Eur. Orch. Bd. 42. H. 3. P. 501–518.
Efimov P.G., 2011. Revealing the decline and expansion of Orchids of NW European Russia // Eur. J. Environ. Sci. V. 1. № 2. P. 7–17.
Efimov P.G., 2020. Orchids of Russia: Annotated checklist and geographic distribution // Nat. Cons. Res. V. 5. Suppl. 1. P. 1–18. https://doi.org/10.24189/ncr.2020.018
Efimov P.G., Panasenko N.N., Gornov A.V., 2022. Remnant populations of Cypripedium macranthos (Orchidaceae) in Eastern Europe: Evidence of almost complete extinction and widespread introgression with Cypripedium calceolus // Nat. Cons. Res. V. 7. № 2. P. 1–20. https://doi.org/10.24189/ncr.2022.014
Farrell L., 1985. Biological flora of the British Isles: Orchis militaris L. // J. Ecol. V. 73. № 3. P. 1041–1053.
Fay M., Sayers B., Taylor I., 2015. Dactylorhiza viridis // Curtis Bot. Mag. V. 32. № 1. P. 33–41. https://doi.org/10.1111/curt.12091
Global Red List of Japanese Threatened Plants, 2020. https://www.kahaku.go.jp/english/research/db/botany/redlist/references.html
IUCN, 2012. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0. Gland: IUCN. 41 p.
Jacquemyn H., Hutchings M., 2010. Biological flora of the British Isles: Spiranthes spiralis (L.) Chevall // J. Ecol. V. 98. № 5. P. 1253–1267. https://doi.org/10.1111/j.1365-2745.2010.01701.x
Jones P.D., Moberg A., 2003. Hemispheric and large-scale surface air temperature variations: an extensive revision and update to 2001 // J. Climate. V. 16. P. 206–223.
Kretzschmar H., Eccarius W., Dietrich H., 2007. The Orchid Genera Anacamptis, Orchis and Neotinea: Phylogeny, Taxonomy, Morphology, Biology, Distribution, Ecology and Hybridisation. Bürgel: EchinoMedia. 544 p.
Kull T., Selgis U., Pecina M.V., Metsare M., Ilves A. et al., 2016. Factors influencing IUCN threat levels to orchids across Europe on the basis of national Red Lists // Ecol. Evol. V. 6. P. 6245–6265.
Kurbel R., Hirse T., 2017. Eesti Orhideede Käsiramaat. Tallinn: MTÜ Käoraamat. 304 p.
Masson-Delmotte V., Zhai P., Pörtner H.-O., Roberts D., Skea J. et al. (eds.), 2019. IPCC, 2018: Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. https://www.ipcc.ch/site/assets/uploads/sites/2/2019/ 06/SR15_Full_Report_Low_Res.pdf
McInerny G.J., Roberts D.L., Davy A.J., Cribb P.J., 2006. Significance of sighting rate in inferring extinction and threat // Cons. Biol. V. 20. № 2. P. 562–567. https://doi.org/10.1111/j.1523-1739.2006.00377.x
Meekers T., Hutchings M.J., Honnay O., Jacquemyn H., 2012. Biological flora of the British Isles: Gymnadenia conopsea s.l. // J. Ecol. V. 100. № 5. P. 1269–1288. https://doi.org/10.1111/j.1365-2745.2012.02006.x
Petřík P., Pergl J., Wild J., 2010. Recording effort biases the species richness cited in plant distribution atlases // Perspect. Plant Ecol. Evol. Syst. V. 12. P. 57–65.
Piessens K., Hermy M., 2006. Does the heathland flora in northwestern Belgium show an extinction debt? // Biol. Conserv. V. 132. № 3. P. 382–394. https://doi.org/10.1016/j.biocon.2006.04.032
Prendergast J.R., Wood S.N., Lawton J.H., Eversham B.C., 1993. Correcting for variation in recording effort in analyses of diversity hotspots // Biodivers. Lett. V. 1. № 2. P. 39–53. https://doi.org/10.2307/2999649
Preston C.D., Pearman D.A., Dines T.D., 2002. New Atlas of the British and Irish Flora. Oxford: Oxford Univ. Press. 922 p.
Pridgeon A.M., Cribb P.J., Chase M.W., Rasmussen F.N., 2001. Genera Orchidacearum. V. 2. Orchidoideae (Part 1). Oxford: Oxford Univ. Press. 464 p.
Rich T., 2006. Floristic changes in vascular plants in the British Isles: Geographical and temporal variation in botanical activity 1836–1988 // Bot. J. Linn. Soc. V. 152. № 3. P. 303–330.
Rich T., Karran A., 2006. Floristic changes in the British Isles: Comparison of techniques for assessing changes in frequency of plants with time // Bot. J. Linn. Soc. V. 152. № 3. P. 279–301. https://doi.org/10.1111/j.1095-8339.2006.00574.x
Štípková S., Kindlmann P., 2015. Extent and reasons for meadows in South Bohemia becoming unsuitable for orchids // Eur. J. Environ. Sci. V. 5. № 2. P. 142–147. https://doi.org/10.14712/23361964.2015.87
Tamis W.L.M., Van’t Zelfde M., Van Der Meijden R., Udo De Haes H.A., 2005. Changes in vascular plant biodiversity in the Netherlands in the 20th century explained by their climatic and other environmental characteristics // Clim. Change. V. 72. № 1–2. P. 37–56. https://doi.org/10.1007/s10584-005-5287-7
Vogt-Schilb H., Munoz F., Richard F., Schatz B., 2015. Recent declines and range changes of orchids in Western Europe (France, Belgium and Luxembourg) // Biol. Conserv. V. 190. P. 133–141. https://doi.org/0.1016/j.biocon.2015.05.002
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии