

Журнал общей биологии, 2022, T. 83, № 4, стр. 302-320
Эколого-географическая организация и структура летнего населения птиц Северной Евразии
Ю. С. Равкин 1, *, С. М. Цыбулин 1, А. А. Ананин 2, 3, В. Г. Ивлиев 4, Т. К. Железнова 5, Л. Г. Вартапетов 1, В. А. Юдкин 1, 6, В. С. Жуков 1, Е. С. Преображенская 7, М. С. Стишов 7, К. В. Торопов 1, Е. С. Равкин 8, И. Н. Богомолова 1, С. В. Чеснокова 1, М. И. Лялина 1
1 Институт систематики и экологии животных СО РАН
630091 Новосибирск, ул. Фрунзе, 11, Россия
2 ООПТ “Заповедное Подлеморье”
671623 п. Усть-Баргузин, ул. Ленина, 71, Республика Бурятия, Россия
3 Институт общей и экспериментальной биологии СО РАН
670047 Улан-Удэ, ул. Сахьяновой, 6, Республика Бурятия, Россия
4 Институт проблем экологии и недропользования АН РТ
420087 Казань, ул. Даурская, 28, Республика Татарстан, Россия
5 Российский государственный аграрный университет – МСХА им. К.А. Тимирязева
127434 Москва, ул. Тимирязевская, 49, Россия
6 Новосибирский государственный университет
630090 Новосибирск, ул. Пирогова, 1, Россия
7 Институт проблем экологии и эволюции РАН
119071 Москва, Ленинский пр-т, 33, Россия
8 Российский государственный аграрный заочный университет
143907 Московская обл., Балашиха, ш. Энтузиастов, 50, Россия
* E-mail: zm.nsc@yandex.ru
Поступила в редакцию 21.02.2022
После доработки 06.04.2022
Принята к публикации 28.04.2022
- EDN: WRTPKK
- DOI: 10.31857/S0044459622040066
Аннотация
Для выявления пространственно-типологической структуры и организации орнитокомплексов Северной Евразии проанализированы усредненные за первую половину лета (16.05–15.07) результаты учетов птиц, проведенных в период с 1880 по 2019 г. (с перерывами и преимущественно с 1960 г.). В сборе материала участвовали 354 исследователя (в течение 110 лет). Обработка данных проведена с использованием методов многомерной статистики, включая кластерный анализ и линейную качественную аппроксимацию матриц связи. В результате выявлены основные территориальные тренды в сообществах птиц и коррелирующие с ними факторы среды, оценена связь пространственной изменчивости орнитокомплексов с неоднородностью местообитаний. Показана фрактальность пространственной дифференциации населения птиц.
Исследование, послужившее основой для написания статьи, предлагаемой вниманию читателей, выполнено в рамках факторной зоогеографии (Равкин, Ливанов, 2008). Особенность этого направления заключается не только в обязательном выявлении факторов среды, определяющих или объясняющих географическую изменчивость животного населения, но и в переходе при анализе и описании в виртуальное типологическое многомерное факторное пространство из реального хорологического. Методической особенностью работ, выполняемых в рамках факторной зоогеографии, служат факторный (в математическом понимании) и кластерный анализы (последний ранее называли автоматической классификацией). После этого оценивают силу и общность связи изменчивости населения и основных структурообразующих факторов из числа выявленных в процессе анализа и предметной интерпретации его результатов. Эти показатели рассчитывают с помощью линейной качественной аппроксимации – одного из методов регрессионного анализа. Классификацию и оценку связи выполняют на основе матрицы коэффициентов сходства, обычно для количественных признаков.
Подобные исследования, помимо чисто познавательного, теоретического смысла, имеют и важное прикладное значение. Оно сводится к возможности вторичного использования собранных для анализа данных, составления прогнозов в пространстве для заполнения “белых пятен” на необследованных территориях по известным для них факторам среды и во времени для предсказаний грядущих изменений в составе животного населения. Для этого используют прогнозные сведения об изменении площадей местообитаний животных и отличий в основных факторах среды – антропогенных и природно-географических.
Наша статья впервые дает представление об изменчивости орнитокомплексов по всей территории Северной Евразии в границах СССР на 1990 г. Предшествующие обобщения были проведены лишь по отдельным регионам в пределах этой территории или выполнены по орнитофауне, т.е. без использования информации по обилию птиц в местообитаниях ранга ландшафтного урочища. Показана фрактальность населения птиц, в первую очередь самоподобие в его неоднородности как по отдельным регионам, так и в целом по обследованной территории.
От зарубежных аналогов (Diamond, 1975; Connor, Simberloff, 1983, 2014; Gotelli, Graves, 1996; Götzenberger et al., 2012; Gao et al., 2016, и др.) наши исследования отличаются, во-первых, анализом не отдельных параметров животного населения (плотность, разнообразие, видовое богатство), а рассмотрением интегральных показателей сообществ в целом, их облика. Облик населения измеряют коэффициентами сходства для количественных признаков. Во-вторых, за рубежом исследования чаще ведут на островах или ограниченных территориях. Наша работа выполнена на огромной территории, как материковой, так и на прилежащих морских акваториях и островах. В статьях, опубликованных в иностранных журналах, приводятся результаты анализа, проведенного в ограниченных рамках выдвинутых гипотез, которые касаются простых факторов среды и взаимоотношений отдельных групп видов животных. Наши подходы и методы – факторные, компонентные и кластерные, традиционные, но хорошо зарекомендовавшие себя, позволяют целенаправленно выявлять не только основные структурообразующие факторы, но и их неразделимые сочетания – природно-антропогенные режимы. Это не только увеличивает информативность получаемых представлений, что улучшает качество прогнозов, но и сокращает время, затрачиваемое на расчеты и анализ. За рубежом работы не редко проводят по квадратам местности, что позволяет говорить лишь о наименее изменчивых во времени факторах (зональность, континентальность, высотная поясность). В наших исследованиях наименьшей ранговой (масштабной) единицей рассмотрения принято ландшафтное урочище. Это делает возможным проводить более детальное рассмотрение, включая влияние состава растительности и почв, степени антропогенной трансформации среды, кормности угодий, их заболоченности, минерального питания болот, водности и значимые сочетания этих факторов. В свою очередь, это позволяет расширить анализ отдельных факторов до целесообразного уровня рассмотрения с максимально возможной полнотой объяснения.
МАТЕРИАЛЫ И МЕТОДЫ
Для расчетов и анализа взяты материалы из банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН (Равкин, Ефимов, 2009). Значительная часть их опубликована. Всего использованы оценки обилия 649 видов птиц по 8202 орнитокомплексам, часть которых обследована в течение более чем одного года. В сборе материала участвовали 354 орнитолога. Все учеты проведены на территории Советского Союза в границах 1990 г. и на островах архипелага Шпицбергена (Норвегия). Последние обследованы И.В. Покровской и Г.М. Тертицким. Эти материалы включены в анализ в связи с недостатком сведений по островным арктическим тундрам.
Проведение кластерного анализа больших массивов данных сопряжено с рядом серьезных затруднений. В предыдущих работах мы “избавлялись” от обилия материалов, усредняя их по выделам карт растительности после предварительного расчета средних по данным, собранным в одних и тех же местообитаниях в разные годы, поскольку анализ межгодовых отличий в сообществах птиц представляет собой отдельную задачу. Ее можно решать по территориям, обследованным в течение многих лет. Большинство использованных нами данных собрано в один из сезонов.
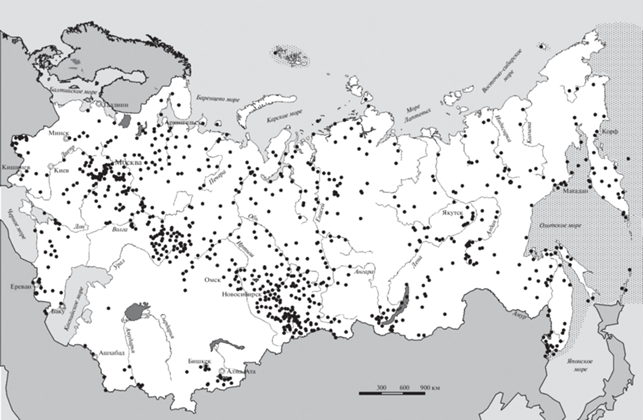
В исследованиях, послуживших основой для написания настоящей статьи, мы попытались ничего не усреднять, за исключением материалов, собранных за один летний сезон с недельной, двухнедельной или подекадной повторяемостью на одних и тех же маршрутах. Для анализа использованы результаты учетов птиц с 16 мая по 15 июля в периоды с 1880 по 1907, с 1935 по 1941 и с 1945 по 2019 гг. При этом большая часть материалов собрана после 1960 г. В список соавторов включены орнитологи, доля материалов которых в общей выборке составила не менее 2% по числу обследованных местообитаний, и специалисты, проводившие обработку и анализ данных. Большая часть остальных участников работ упомянута в наших предыдущих публикациях. В этой статье мы упоминаем только часть обобщающих монографий и статей: Е. Равкин, Ю. Равкин, 2005; Жуков, 2006; Цыбулин, 2009; Торопов, Бочкарева, 2014; Железнова, Леппа, 2016; Равкин и др., 2016, 2020; Железнова, 2017; Железнова, Вартапетов, 2018; Железнова, Костылева, 2018; Климова, Торопов, 2018; Железнова и др., 2021; Ливанов, 2021. В этих публикациях приведен почти полный список участников исследований и библиографическое описание использованных публикаций, откуда взяты сведения по обилию птиц. Места проведения работ показаны на рис. 1. На нем, с одной стороны, четко видна сравнительно высокая “густота” мест проведения учетов на исследуемой территории. С другой стороны, нельзя не отметить некоторую неоднородность их распределения. Так, в окрестностях ряда городов, где больше орнитологов (Москва, Казань, Екатеринбург, Новосибирск, Томск, Владивосток), собран материал, больший по числу мест проведения учетов. В ряде регионов, наоборот, количество таких участков явно недостаточно (часть Средней Азии, а также Средней, Северо-Восточной и Северо-Притихоокеанской Сибири). Однако в целом материала уже достаточно для первого варианта районирования столь обширной и разнородной территории, как Северная Евразия.
Методы подсчета птиц на маршрутах и пересчета результатов относительных учетов на площадь, в том числе в водно-околоводных местообитаниях, описаны ранее (Hayne, 1949; Равкин, 1967; Равкин, Челинцев, 1999; Равкин, Ливанов, 2008; Равкин и др., 2020). При обработке данных для настоящего сообщения применены в основном программы факторной классификации (для выявления трендов в населении птиц и основных структурообразующих факторов) и линейной качественной аппроксимации (для оценки связи изменчивости орнитокомплексов и неоднородности среды) (Куперштох, Трофимов, 1975; Трофимов, 1976; Равкин и др., 1978; Равкин, Ливанов, 2008). Расчеты по программе “Факторная классификация”, при использовании которой объем анализируемой матрицы коэффициентов сходства не ограничен, по всем данным заняли почти шесть суток непрерывного счета. Но сложности возникли не при проведении агрегации, а при интерпретации результатов. Затруднения связаны с объяснением выделенных кластеров из-за многомерности связей их состава с разными факторами среды, одинаково влияющими на степень сходства различных орнитокомплексов, и невозможности “удержать в голове” все или даже основные сочетания факторов.
Попытка анализировать результаты кластерного анализа на подвыборках по 1/8 части не облегчила процесса интерпретации, хотя в целом подтвердила общий характер объяснения пространственной изменчивости орнитокомплексов. Поэтому трудности в интерпретации связаны не столько с объемом анализируемого материала, сколько с постепенностью изменений сообществ, разнообразии и значительности территории, где собраны данные.
Алгоритм линейной качественной аппроксимации матриц связи с помощью выделенных градаций факторов не предусмотрен для работы со столь большими выборками, поэтому расчеты выполнены нами на подвыборках. Для этого вся совокупность вариантов населения перед проведением расчетов бинарным способом (отбором последовательно каждой второй пробы) или с помощью генератора случайных чисел разделена на подсовокупности объемом примерно от половины до 1/16 части выборки. По ним и проведены оценки (подробнее см. Равкин, Ливанов, 2008). Различия их в зависимости от размеров подвыборки колебались в пределах 1, реже 2% дисперсии, поэтому можно считать, что оценки слабо зависят от размеров подвыборки, если она пропорционально отражает неоднородность всей совокупности.
Названия видов птиц даны по А.И. Иванову (1976), кроме отдельных позднее описанных видов, а типов фауны – по Б.К. Штегману (1938) с некоторыми дополнениями.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Итоговая классификация орнитокомплексов первой половины лета
Население птиц местообитаний:
I – Надтипа естественных и агроландшафтов суши11.
1 – Тундрово-стланикового типа (полярных пустынь и тундр, в том числе в сочетании с лугами и болотами, а также участков низкорослой растительности и стлаников, кроме арчовых; лидеры по обилию – лапландский подорожник, доля в населении – 13%, желтая трясогузка, краснозобый конек, камнешарка и пуночка – по 4%; плотность населения, особей/км2/фоновых видов – 189/42; доля в населении преобладающих по обилию типов фауны: арктического – 54%, сибирского – 16%, транспалеарктов – 12%22),
1.1 – островных и равнинных материковых (лапландский подорожник 17, камнешарка 6, краснозобый конек, пуночка и желтая трясогузка по 5; 208/40; арктического типа фауны 69, сибирского 13, транспалеарктов 11),
1.1.1 – островов Шпицбергена (пуночка 32, морской песочник 28, моевка 12, белощекая казарка 8, бургомистр 7; 68/7; арктического типа фауны 100),
1.1.2 – прочих островов (лапландский подорожник и камнешарка по 20, пуночка 16, тулес и чернозобик по 8; 154/15; арктического типа фауны 92),
1.1.3 – материковых равнинных от арктических до северотаежных (лапландский подорожник 17, краснозобый конек 8, желтая трясогузка 7, чечетка и овсянка-крошка по 4; 254/47; арктического типа фауны 60, сибирского 16, транспалеарктов 15),
1.2 – горных материковых (горный конек 15, бурая пеночка 8, варакушка 7, луговой конек и полярная овсянка по 4; 149/34; сибирского типа фауны 25, транспалеарктов 17, тибетского типа 21, китайского 15, европейского 10),
1.2.1 – от Фенноскандии до Урала и Кавказа (луговой конек 39, весничка, варакушка и овсянка-крошка по 7, таловка 5; 91/14; европейского типа фауны 51, сибирского 23, транспалеарктов 13, арктического типа 11),
1.2.2 – северо-притихоокеанских, Северо-Восточной и Средней Сибири (желтая трясогузка 11, чечетка 9, бурая пеночка 8, таловка 6, краснозобый конек 5; 155/38; сибирского типа фауны 34, транспалеарктов 22, арктического и китайского типов по 14),
1.2.3 – прибайкальских, забайкальских и амуро-сахалинских (бурая пеночка 22, щур 10, зеленый конек 7, чечевица и соловей-красношейка по 5; 98/22; китайского типа фауны 50, сибирского 31),
1.2.4 – алтае-саянских (горный конек 28, варакушка 11, бурая пеночка 7, зарничка и полярная овсянка по 6; 187/28; тибетского типа фауны 35, сибирского 19, транспалеарктов 17, китайского 14),
1.2.5 – среднеазиатских (гималайский вьюрок 36, горный конек 17, клушица 10, чечевица 9, черногорлая завирушка 8; 196/15; тибетского типа фауны 53, китайского 18, монгольского 10).
2 – Редколесно-лесного типа (редколесий, лесов, мозаичных по облесенности территорий, в том числе полей среди них, облесенных низинных и переходных болот и рослых рямов; зяблик 8, пухляк, лесной конек, теньковка и большая синица по 4; 511/92; европейского типа фауны 50, сибирского 24),
2.1 – северных (от тундровых до северотаежных и гольцово-подгольцовых; овсянка-крошка 11, юрок 8, чечетка 7, весничка 6, таловка 5; 334/60; сибирского типа фауны 56, европейского 20, транспалеарктов 12),
2.1.1 – всех, кроме колымских и камчатских (овсянка-крошка 12, чечетка и юрок по 8, весничка 6, зарничка 5; 336/57; сибирского типа фауны 55, европейского 21, транспалеарктов 12),
2.1.2 – колымских и камчатских (овсянка-ремез, корольковая пеночка и чечевица по 10, таловка 9, зеленый конек 8; 306/38; сибирского типа фауны 65, китайского 24),
2.2 – срединных (Карпат, Крыма, Кавказа и равнинных от среднетаежных до степных и от западных границ до Забайкалья включительно, кроме передне-, средне- и центральноазиатских; зяблик 11, лесной конек 5, теньковка, большая синица и пухляк по 4; 582/88; европейского типа фауны 60, сибирского 20, транспалеарктов 10),
2.2.1 – хвойных, хвойно-лиственных и мелколиственных лесов и рослых рямов (зяблик 11, пухляк 6, теньковка, лесной конек и большая синица по 5; 560/73; европейского типа фауны 58, сибирского 25),
2.2.2 – широколиственных лесов (зяблик 19, большая синица 7, черноголовая славка, певчий дрозд и садовая славка по 3; 1030/70; европейского типа фауны 85),
2.2.3 – мозаичных по облесенности (лесной конек и зяблик по 5, дубровник, теньковка и чечевица по 4; 514/93; европейского типа фауны 49, транспалеарктов 19, сибирского типа 14, китайского 11),
2.3 – среднеазиатских равнинных (бормотушка 22, бледная пересмешка 20, славка-завирушка и серая синица по 6, скотоцерка 4; 1296/68; средиземноморского типа фауны 65, европейского 23),
2.3.1 – тугаев (бормотушка 24, бледная пересмешка 22, серая синица 7, индийский воробей и славка-завирушка по 4; 1582/66; средиземноморского типа фауны 69, европейского 20),
2.3.2 – зарослей гребенщика (славка-завирушка 26, скотоцерка 17, бормотушка 12, бледная пересмешка 10, тугайный соловей 5; 967/35; средиземноморского типа фауны 60, европейского 35),
2.3.3 – саксаульников (скотоцерка 28, тугайный соловей 24, бормотушка 15, серый жаворонок 11, горлица 4; 259/15; средиземноморского типа фауны 72, монгольского 19),
2.3.4 – садов и парков (черный дрозд 19, большая синица 17, сорока 16, сизый голубь 7, южный соловей 5; 546/30; европейского типа фауны 71, средиземноморского 11),
2.4 – среднеазиатских и переднеазиатских горных (черный дрозд 8, зяблик, зеленая пеночка и серая мухоловка по 5, зарничка 4; 999/84; европейского типа фауны 48, средиземноморского 17, китайского 15),
2.4.1 – пойменных лиственных лесов (скворец, чечевица и зеленая пеночка по 12, князек и зарничка по 7; 884/58; европейского типа фауны 35, китайского 25, средиземноморского 12, транспалеарктов 10),
2.4.2 – хвойных и смешанных лесов (зарничка 14, индийский воробей 11, зеленая пеночка 7, славка-завирушка 6, чечевица 5; 667/53; европейского типа фауны 34, средиземноморского 20, китайского 15, сибирского 14),
2.4.3 – орехово-плодовых лесов (черный дрозд 18, серая мухоловка 11, желтогрудый князек 9, большая горлица 8, рыжешейная синица 6; 1171/51; европейского типа фауны 52, средиземноморского 20, китайского 16),
2.4.4 – арчовых стлаников (красношапочный вьюрок 11, зарничка 10, малая розовая чечевица и чечевица по 9, черногрудая красношейка 8; 608/36; тибетского и китайского типов фауны по 24, средиземноморского и монгольского по 12, сибирского 11, европейского 10),
2.4.5 – арчовых редколесий (горная овсянка 10, рыжешейная синица и серая славка по 9, кеклик 7, плешанка 5; 759/51; европейского типа фауны 34, монгольского 23, китайского 14, средиземноморского 13),
2.4.6 – лугов в сочетании с неарчовыми редколесьями (зарничка и зеленая пеночка по 18, желтоголовый королек 11, московка 9, черногорлая завирушка 8; 696/30; сибирского и европейского типов фауны по 26, китайского 25),
2.4.7 – лесопосадок (индийский воробей 31, черногрудый воробей 11, сорока 7, большая синица 5, большая горлица 3; 524/36; средиземноморского типа фауны 46, европейского 34),
2.4.8 – зарослей облепихи (ополовник 20, южный соловей и широкохвостая камышевка по 12, чечевица и большая горлица по 7; 355/19; европейского типа фауны 37, транспалеарктов 24, китайского типа 18, средиземноморского 12),
2.4.9 – городских парков (большая синица 29, черный дрозд 17, сорока 15, зеленая пеночка 7, городская ласточка 6; 421/21; европейского типа фауны 70, транспалеарктов 10),
2.4.10 – широколиственных лесов (зяблик 18, московка 13, большая синица 12, зеленая пеночка 8, деряба 7; 3372/21; европейского типа фауны 74, средиземноморского 10),
2.5 – прибайкальских, забайкальских, северо-восточносибирских и притихоокеанских (поползень и пухляк по 6, корольковая пеночка и седоголовая овсянка по 5, московка 4; 279/64; сибирского типа фауны 49, китайского 30, европейского 11),
2.5.1 – северо-восточносибирских (овсянка-ремез 17, таловка 9, чечевица 8, горная трясогузка 7, сибирский пепельный улит 6; 382/47; сибирского типа фауны 64, китайского 17, транспалеарктов 14),
2.5.2 – прибайкальских, забайкальских и амуро-сахалинских (поползень и пухляк по 6, корольковая пеночка и седоголовая овсянка по 5, московка 4; 277/64; сибирского типа фауны 46, китайского 34, европейского 10).
3 – Пустынно-луговостепного типа (степей, тундростепей, полупустынь, пустынь, полей, скал, россыпей и кустарников, а также лугов, кроме тундровых; полевой жаворонок 7, желтая трясогузка 6, индийский воробей, сизый голубь и скворец по 3; 447/95; транспалеарктов 31, европейского типа фауны 25, средиземноморского 14),
3.1 – срединных (равнинных, кроме среднеазиатских; полевой жаворонок 10, желтая трясогузка 9, дубровник и скворец по 4, полевой воробей 3; 423/82; транспалеарктов 41, европейского типа фауны 29, китайского 10),
3.1.1 – кустарников и лугов, кроме выпасов, остепненных лугов и солончаков (желтая трясогузка 9, дубровник 7, полевой жаворонок, барсучок и скворец по 3; 519/90; транспалеарктов 35, европейского типа фауны 28, китайского 13, сибирского 10),
3.1.2 – степей, полупустынь, полей, залежей, остепненных лугов, выпасов, осыпей, отвалов и солончаков (полевой жаворонок 18, желтая трясогузка 8, скворец 6, полевой воробей 5, грач 4; 340/56; транспалеарктов 47, европейского типа фауны 30),
3.2 – южных (горных крымско-кавказских, алтае-саянских, центрально- и переднеазиатских, а также среднеазиатских горных и равнинных; индийский воробей и сизый голубь по 8, хохлатый жаворонок 5, полевой воробей и полевой жаворонок по 3; 491/93; средиземноморского типа фауны 31, европейского 20, транспалеарктов 18, монгольского 12).
4 – Болотного типа (болот, кроме рослых рямов и облесенных низинных и переходных; желтая трясогузка 9, лесной конек 5, желтоголовая трясогузка и дубровник по 4, барсучок 3; 380/74; транспалеарктов 34, европейского типа фауны 29, сибирского 11),
4.1 – верховых (лесной конек 15, зяблик 9, белошапочная овсянка 7, весничка и желтая трясогузка по 6; 229/48; европейского типа фауны 50, транспалеарктов 21, сибирского 19),
4.2 – открытых, кроме верховых и марей (желтая трясогузка 11, желтоголовая трясогузка 6, барсучок 5, дубровник и камышевая овсянка по 4; 530/84; транспалеарктов 40, европейского типа фауны 22, арктического 11),
4.3 – марей (бурая пеночка 19, дубровник 8, зеленоголовая трясогузка 7, певчий и пятнистый сверчки 6 и 5; 168/33; китайского типа фауны 42, сибирского 29, транспалеарктов 16).
II – Надтипа антропогенных территорий, кроме аграрных.
5 – Рекреационно-селитебного типа (селитебных, кроме тундровых поселков, промышленных, животноводческих, рудеральных, не среднеазиатских рекреационных, полос отчуждения и виноградников; домовый и полевой воробьи 33 и 15, сизый голубь 12, скворец 4, белая трясогузка 3; 1431/76; транспалеарктов 59, европейского типа фауны 20, средиземноморского 13),
5.1 – северных (в пределах тундровой и лесотундровой зон, а также гольцов и подгольцовья; домовый воробей и белая трясогузка по 19, серебристая чайка 10, желтоголовая трясогузка 7, береговая ласточка 5; 510/34; транспалеарктов 54, арктического типа фауны 18),
5.2 – южных (южнее лесотундровой зоны; домовый и полевой воробьи 33 и 15, сизый голубь 12, скворец 4, белая трясогузка 3; 1413/78; транспалеарктов 54, европейского типа фауны 20, средиземноморского 15).
III – Надтипа материковых водоемов и водотоков.
6 – Озерно-речного типа (озер, водохранилищ, рек, прудов, очистных водоемов, литоралей, полярных и черноморских заливов; береговая ласточка 10, малый зуек, озерная и серебристая чайки по 6, лысуха 5; 272/42; транспалеарктов 64),
6.1 – северных (в пределах тундровой зоны и лесотундры) и гольцово-подгольцовых, кроме среднеазиатских и переднеазиатских (полярная крачка 18, морянка 16, чернозобая гагара 10, круглоносый плавунчик 6, чирок-свистунок 4; 79/16; арктического типа фауны 53, сибирского 23, транспалеарктов 15),
6.2 – срединных (от северной тайги до подтаежных и широколиственных лесов включительно и в пределах лесных поясов гор, кроме средне- и переднеазиатских; береговая ласточка 20, кряква 9, озерная чайка и горная трясогузка по 7, чирок-трескунок 6; 171/28; транспалеарктов 76, сибирского типа фауны 10),
6.3 – южных (лесостепных и степных зон и поясов, кроме средне-, передне- и центральноазиатских; серебристая и озерная чайки 13 и 7, желтая трясогузка, лысуха и береговая ласточка по 6; 420/41; транспалеарктов 53, европейского 11),
6.4 – среднеазиатских и казахстанских в пределах пустынь и полупустынь (малый зуек 30, горная и маскированная трясогузки по 11, перевозчик 9, черный стриж 6; 1436/34; транспалеарктов 66, европейского 10),
6.5 – переднеазиатских и центральноазиатских (лысуха 77, камышевая овсянка 6, серебристая чайка 3, дроздовидная камышевка и горная ласточка по 2; 473/20; транспалеарктов 89).
IV – Надтипа морских акваторий и птичьих базаров.
7 – Морского типа (северных и восточных морских акваторий и птичьих базаров; люрик 47, толстоклювая кайра 40, моевка 10, чистик 0.7, тупик 0.5; 6533/13; арктического типа фауны 100),
7.1 – западных полярных (птичьих базаров Шпицбергена; люрик 48, толстоклювая кайра 41, моевка 10, чистик 0.7, тупик 0.5; 10785/7; арктического типа фауны 100),
7.2 – восточных полярных и тихоокеанских (акваторий и птичьих базаров; моевка 18, тихоокеанская морская чайка 15, топорок 11, глупыш и тонкоклювая кайра по 7; 65/13; арктического типа фауны 61),
7.2.1 – полярных, Берингова моря и севера Охотского (конюга-крошка 24, моевка 19, тонкоклювая кайра 17, большая конюга 14, толстоклювая кайра 8; 60/7; арктического типа фауны 84),
7.2.2 – камчатских, прикомандорских и курильских (глупыш 33, тонкоклювый буревестник 11, толстоклювая кайра и моевка по 10, топорок 8; 89/13; арктического типа фауны 76),
7.2.3 – средней части Охотского моря, сахалинских и шантарских (тихоокеанская морская чайка 21, моевка 19, топорок 14, большая конюга 7, очковый чистик 6; 61/9; арктического типа фауны 53, китайского 11).
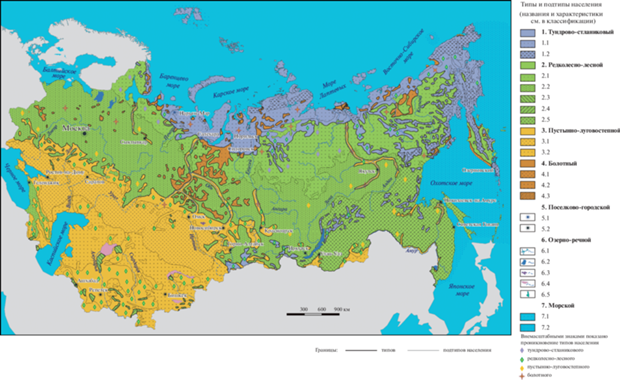
По характеристикам типов и подтипов иерархической классификации составлена обзорная карта, отражающая неоднородность летнего населения птиц Северной Евразии в границах СССР 1990 г. (рис. 2).
Пространственно-типологическая структура орнитокомплексов
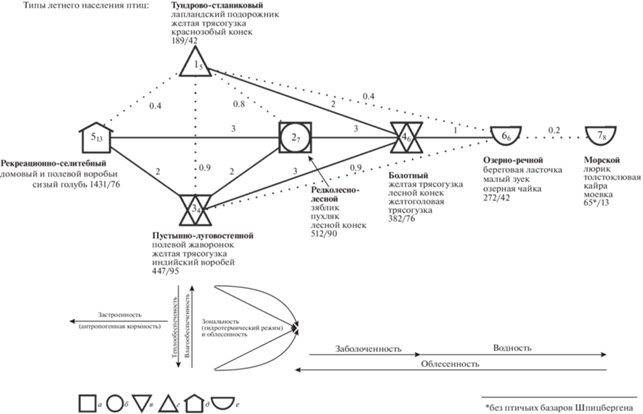
На уровне типа сообществ структура населения птиц представима в виде ряда таксонов от антропогенного (рекреационно-селитебного) типа населения через редколесно-лесной к болотному и далее к водно-околоводным: озерно-речному и морскому (рис. 3). Этот тренд вначале связан с уменьшением застроенности территорий и увеличением облесенности, а потом с возрастанием заболоченности и водности. От него имеется два отклонения вверх (к северу) – в тундрово-стланиковых местообитаниях – и к югу – в пустынных, степных и луговых сообществах. Эти отклонения обусловлены: к северу снижением теплообеспеченности и, соответственно, облесенности, а к югу – уменьшением влагообеспеченности и вследствие этого – облесенности из-за нарастающей аридности. Следует также отметить некую “треугольность” зональных отличий в орнитокомплексах (типы 1–3). В горах она превращается в “полулунность” за счет сходства населения птиц гольцово-подгольцовых и предгорных открытых или закустаренных местообитаний (Ефимов, Равкин, 1980). Эти “искривления” на схеме связаны со сходством экологических условий в открытых и закустаренных местообитаниях как на северных, так и на южных необлесенных территориях для таких видов, как варакушка, бурая пеночка, певчий и пятнистый сверчки и др. По основному (горизонтальному) тренду средняя плотность населения неуклонно уменьшается от застроенных территорий до морских акваторий и птичьих базаров от 1431 до 6444 особей/км2. Почти так же изменяется видовое богатство и число фоновых видов птиц. Исключение составляют сообщества застроенной суши, где видовое богатство ниже, чем в лесных. В вертикальном ряду орнитокомплексов эти показатели убывают к северу, кроме числа фоновых видов, которых в редколесно-лесных сообществах больше, чем к северу и югу.
Рис. 3.
Пространственно-типологическая структура летнего населения птиц Северной Евразии в первой половине лета (на уровне типа). Орнитокомплексы: а – лесов, б – полуоблесенных местообитаний, в – открытых бедных по продуктивности территорий, г – более богатых, д – селитебных ландшафтов, е – рек, материковых водоемов, морей. Для каждого таксона приведены три лидирующих по обилию вида птиц, плотность населения (особей/км2) и число фоновых видов по среднему обилию. Номера типов как в иерархической классификации, индексом у номера типа приведено среднее сходство вошедших в него вариантов. Сходство их по типам показано у связей между типами. Значимое сходство отображено сплошной линией, пунктиром – дополнительная информация; стрелки направлены в сторону усиления влияния фактора среды, коррелирующего с трендом. Нулевым значением обозначено сходство в таксоне, представленном одним вариантом.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Информативность классификационных и структурных представлений о неоднородности орнитокомплексов
Информативность иерархической классификации на уровне надтипа населения птиц равна 10% дисперсии коэффициентов сходства, типа и класса орнитокомплексов примерно вдвое, а подтипа – втрое больше (табл. 1). Вся классификация учитывает 34% дисперсии матрицы сходства (множественный коэффициент корреляции – 0.58, при Р = 0.9).
Таблица 1.
Информативность классификационных представлений о территориальной изменчивости летнего населения птиц Северной Евразии, доля учтенной дисперсии матрицы сходства, %
| Таксон | Иерархическая классификация по 2051 пробам | Структура по 1028 пробам |
|---|---|---|
| Надтип | 10 | 11 |
| Тип | 23 | 26 |
| Подтип | 30 | 34 |
| Класс | 24 | 36 |
| Всего | 34 | 39 |
После бинарного разделения всей совокупности данных примерно на 8 частей среднее значение информативности классификации и на уровне типа, так же как структуры было таким же, как при однократной оценке на вдвое большем объеме – 34% ± 1 сложный процент и – 25% ± 2 сложных процента (Р = 0.9). Тот же порядок отличий свойствен оценкам связи с факторами среды и режимами (табл. 2). Таким образом, информативность оценок, полученных по одной восьмой части выборки и всем подвыборкам в среднем очень сходны.
Таблица 2.
Сила и общность связи территориальной изменчивости летнего населения птиц Северной Евразии и факторов среды (доля учтенной дисперсии матрицы сходства, %, по 515 пробам)
| Фактор, режим | Оценка связи | |
|---|---|---|
| индивидуальная | с нарастающим итогом | |
| Тип растительности | 26 | 26 |
| Облесенность | 21 | 29 |
| Состав лесообразующих пород | 18 | 33 |
| Провинциальность (континентальность, секторность) | 15 | 43 |
| Тепло- и влагообеспеченность (зональность, подзональность, поясность, подпоясность) | 13 | 46 |
| Застроенность | 7 | 47 |
| Водность | 6 | 47 |
| Проточность | 3 | 47 |
| Островной характер территории | 2 | 47 |
| Макрорельеф (горы–равнина) | 1 | 47 |
| Заболоченность | 0.1 | 47 |
| Минеральное питание болот | 0.1 | 47 |
| Всего | 47 | 47 |
Судя по выполненной классификации, несмотря на наличие в выборке многолетних учетов, проведенных в одних и тех же местообитаниях (в окрестностях Казани и на Байкале соответственно до 44 и 28 лет в разных местообитаниях и наличие повторных учетов в разные годы) нигде годовые отличия орнитокомплексов не были больше пространственных. Это лишний раз подтверждает правомерность анализа данных, собранных в разные годы для анализа территориальных отличий на больших площадях. Информативность составленной классификации – 34% дисперсии (коэффициент корреляции 0.58). При использовании карты следует учитывать, что взаимопроникновение различных типов на ней иллюстрирует это явление в принципе и не всегда указывает конкретные места нахождения соответствующих включений.
Информативность структурных представлений на уровне надтипа населения птиц составляет 11% дисперсии, а затем, по мере дробления на таксоны меньшего ранга, оценки увеличиваются в 2, 3 и 3.3 раза. В целом информативность деления больше почти в 3.5 раза по сравнению с надтиповым делением (табл. 1). Множественный коэффициент корреляции – 0.62.
Наше разделение населения птиц на типы очень близко к представлениям Е.С. Птушенко и А.А. Иноземцева (1968) о неоднородности орнитокомплексов Московской области, особенно если исключить типы сообществ, не представленные на этой территории (Тундро-стланниковый и Морской). В оставшейся части типов население птиц, как у Е.С. Птушенко и А.А. Иноземцева, так и у нас, представлено пятью типами (орнитологическими формациями). Четыре из них совпадают почти полностью. Это лесная орнитологическая формация (Редколесно-лесной тип населения в нашем понимании), водная (Озерно-речной тип), открытых пространств (Пустынно-луговостепной тип) и поселений человека (рекреационно-селитебный тип населения). Кустарниковая формация как самостоятельный тип в нашей классификации не представлена. Эти орнитокомплексы входят в Редколесно-лесной или Пустынно-луговостепной типы в зависимости от их зональной принадлежности или поемности. Состав формаций и типов иногда существенно отличаются. Так, в Редколесно-лесной тип входят орнитокомплексы редколесий и мозаичных по облесенности территорий (частично и полностью распаханных участков). Е.С. Птушенко и А.А. Иноземцев относят население пашней к сообществам открытых пространств. Их вариант разделения логичнее, что называется в “чистом” виде. Но в нашем случае объединение связано с особенностями первоначального деления территории. Они выделяли пашни как отдельную консоцию, независимо от площади, занимаемой ими. Мы делили эти местообитания на два урочища – поля в сочетании с перелесками и чистые обширные поля. В соответствии с формализованным алгоритмом из-за непрерывности ряда: леса, редколесья, поля-перелески и поля в их пределах, т.е. распаханные в разной степени местообитания вошли в единый тип населения с лесными орнитокомплексами.
Еще одно отличие касается Болотного типа населения. Е.С. Птушенко и А.А. Иноземцев объединяют их орнитокомплексы с водными сообществами. Дело, видимо, в том, что в Московской области очень мало таких обширных болот как в Западной Сибири, поэтому население этих, как правило, бордюрных биотопов, чаще всего приречных и приозерных, носит переходный (экотонный) характер. Деление населения птиц Московской области на консоции и подтипы по территории СССР в границах 1990 г. не совпадают, поскольку на более обширной территории более значимы широтные (зонально-подзональные) и провинциальные (секторные) отличия, которые не представлены в Московской области.
Пространственная организация орнитокомплексов
Судя по иерархической классификации и структурным графам, территориальная изменчивость летнего населения птиц Северной Евразии коррелирует со следующими факторами среды и природно-антропогенными режимами. В первую очередь это сочетания типов растительного покрова: тундрового, стланикового, редколесного, лесного, пустынного, лугового, степного, болотного и специфика рудерально-рекреационных и водно-околоводных местообитаний – материковых и морских. Фактор – тип растительного покрова, близок к зональности и поясности, но отличается несовпадением границ, объединением части зон и подзон как между собой, так и с горными аналогами по растительности, а также тем, что азональные, интразональные и экстразональные аналоги при зонально-поясном делении входят в соответствующие зоны, подзоны, пояса и подпояса (хорологически), а по населению птиц – в соответствующие таксоны классификации по сходству орнитокомплексов (типологически).
Тепло- и влагообеспеченность определяет распределение типов растительности. Отличия гидротермических условий можно отразить через зональность и подзональность на равнинных участках, поясность и подпоясность в горах, а также провинциальность (континентальность) по удаленности от океанов вглубь материка (на уровне физико-географических стран). Кроме того, в список основных структурообразующих факторов и режимов следует включить антропогенное влияние, в основном через застроенность (антропогенную кормность и промышленную трансформацию местообитаний), а также заболоченность. На уровне подтипа проявляются менее значимые зависимости от зонально-подзональной и физико-географической специфики стран (раздельно), макрорельефа (горы–равнины), трофности и степени облесенности болот.
На уровне класса может быть прослежена связь со спецификой растительности Камчатки, с мозаичностью местообитаний по облесенности, составом зарослей в Средней Азии, заливанием в половодье, степенью аридности, засоленности и распаханности открытых местообитаний, а в населении птиц морских акваторий и птичьих базаров – с особенностями рельефа приморской суши и продуктивностью (кормностью) акваторий.
Обширность исследуемой территории, взаимопроникновение зональных типов растительного покрова в “чуждые” по отношению к преобладающей по зонам растительности, а также плавность и многомерность отличий, в том числе частично связанных с нюансами использованных методик учета птиц, индивидуальными особенностями учетчиков, отличиями в дробности подразделения ими местообитаний и годовых колебаний населения птиц затрудняют интерпретацию результатов кластерного анализа и снижают оценку информативности классификационных представлений после их идеализации.
В целом по территории наиболее велика оценка связи неоднородности населения птиц с типом растительности (26% дисперсии; табл. 2). Несколько меньше связь с различиями в облесенности и с составом лесообразующих пород (21 и 18%), а также с природными особенностями физико-географических стран и тепло- и влагообеспеченностью, которые определяют отличия в изменчивости первых четырех факторов (режимов). Еще почти вдвое меньше сила и общность связи с застроенностью местообитаний и водностью из-за их локальности или линейности и меньшей площади, которую они занимают по сравнению с естественными ландшафтами суши и, соответственно, меньшим числом обследованных местообитаний (7 и 6%). Значимость остальных факторов еще меньше (от 3 до 0.1%). Все факторы и их сочетания учитывают 47% дисперсии матрицы сходства населения птиц (множественный коэффициент корреляции – 0.69). Скоррелированность выявленных факторов существенна. При расчетах нарастающим итогом у шести первых из них она колеблется в пределах от 1 до 10% дисперсии, а начиная с водности, приращение составляет меньше 1%. Последнее в значительной степени связано с невысокой долей в выборке проб, испытывающих влияние этих факторов.
Основные тренды изменчивости летнего населения птиц
Основные тренды территориальных изменений населения птиц, несомненно, связаны с зонально-провинциальной изменчивостью ландшафтов и в самом обобщенном виде могут быть отображены горизонтальными и вертикальными полосами в направлениях север–юг и запад–восток (рис. 4.). Их интегральное взаимовлияние порождает диагональность отличий с северо-запада и северо-востока к центру региона в южной части его. Диагональность (дифферентность) объясняется соответствующим уменьшением отепляющего влияния течений – Северо-Атлантического, Норвежского, Куросио и Северо-Тихоокеанского в северном направлении. Конечно, это очень грубая схема. Тренды в населении птиц, коррелирующие и несомненно определяемые застроенностью, тепло- и влагообеспеченностью (широтным, долготным и высотным гидротермическим режимом) были показаны на рис. 3 и проанализированы ранее. В гораздо большей степени значима связь с типом растительности (тундровым, лесным, пустынным, луговым и степным), что тоже определяется зонально-провинциальными изменениями среды, но не абсолютно совпадает с ними. Так, распространение лесов достигает тундровой зоны и южных границ пустынной зоны, особенно в горах (рис. 5). Тем не менее по населению птиц целесообразно выделить не пять зон, как по растительности, а три, как на схеме, в виде взаимно пересекающихся ромбовидных фигур. При этом открытые местообитания южных зон (лесостепной, степной и пустынной) заняты единым, хотя и не однородным по сходству типом населения птиц. Формирование иного населения птиц на территории чуждых типов отражают представления об интразональности, экстразональности и азональности (Энциклопедический словарь…, 1968). В ландшафтоведении и геоботанике эти понятия используют при описаниях в хорологическом пространстве, а при формализованной классификации растительности и животного населения – в типологическом.
Рис. 4.
Зонально-подзональные, высотно-поясные (подпоясные) и диагональные тренды в неоднородности летнего населения птиц Северной Евразии.
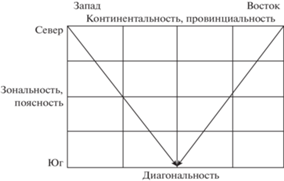
Рис. 5.
Тренды, связанные с типом растительности, застроенностью, водностью, тепло- и влагообеспеченностью, облесенностью и заболоченностью.
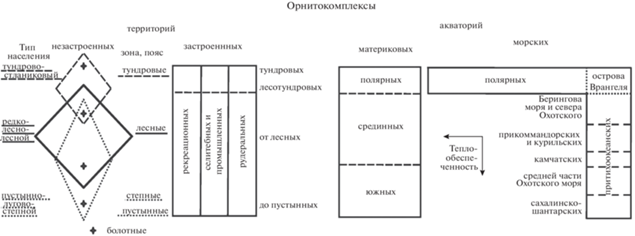
На застроенной территории можно выделить три вертикальных полосы: орнитокомплексов рекреационных, селитебно-промышленных и рудеральных участков, разделенных на две широтных части – северную (тундрово-лесотундровую) и южную (от лесных до пустынных территорий). Население птиц материковых водно-околоводных комплексов разделены на три полосы – полярную, срединную и южную, а морских – на шесть. Эти отличия в целом коррелируют с широтной теплообеспеченностью и, соответственно, с кормностью водоемов.
Аппроксимация неоднородности летнего населения птиц результатами природно-географического районирования
До настоящего времени широко распространено представление о совпадении изменчивости животного населения в целом (и птиц в частности) с зонально-провинциальным изменением тепло- и влагообеспеченности, т.е. с зональностью и континентальностью, иными словами, с отличием в климате с севера на юг и с запада на восток, вплоть до утверждения о полном совпадении границ зон (подзон), провинций, ландшафтов и населения птиц (Кузякин, 1962). Аппроксимация матрицы сходства летнего населения птиц Северной Евразии результатами районирования показала невысокую долю учтенной ими дисперсии. Максимальная оценка свойственна орнито-фаунистическому районированию (Блинова, Равкин, 2008) и по фауне всех наземных позвоночных (Равкин и др., 2015) по 12%, а также по ландшафтному (Кузякин, 1962), геоботаническому, климатическому и физико-географическому районированию (Атлас СССР, 1983) – 11, 10, 9 и 8%. Значительно меньшую долю дисперсии матрицы сходства летнего населения птиц учитывает биотическое районирование (Воронов, Кучерук, 1977) – 5%. По сравнению с классификацией населения птиц, составленной по результатам кластерного анализа, доля учтенной дисперсии в среднем в 10 раз меньше, т.е. степень совпадения изменчивости орнитокомплексов и указанных вариантов районирования очень невелика.
На составленных картах, как по летнему населению птиц Северной Евразии, так и по ее орнитофауне (Блинова, Равкин, 2008), четко видны три зональных полосы: северная (тундровая), срединная (преимущественно лесная) и южная (по фауне среднеазиатская пустынная, а по орнитокомплексам лугово-пустынностепная европейско-азиатская). Однако их границы не совпадают так же, как состав таксонов. Так, по орнитофауне полярные острова и гольцовые участки выделены в отдельные подобласти или провинции, а по населению птиц они входят в единый тундрово-стланниковый тип сообществ. Лесная подобласть по орнитофауне большей частью включает, кроме лесной зоны, лесотундровые, лесостепные и степные территории, а по населению птиц открытые участки лесостепи и степной зоны входят в единый лугово-пустынностепной тип сообществ. Границы территорий провинций и в меньшей степени округов по фауне перекрываются с подтипами и классами орнитокомплексов, хотя тенденции их изменений имеют общую направленность. Естественно, что орнитофаунистическая классификация имеет меньшую дробность, поскольку исходные участки первичного разделения территории взяты в меньшем масштабе, они больше по площади и выполненные по ним усреднения сглаживают (элиминируют) внутренние отличия, например по облесенности и заболоченности. Однако иногда (локально) дробность разделения по фауне выше, чем по населению (например, муссонная провинция, или котловинно-степной округ).
Сопоставление карт, составленных по результатам флоро-фаунистического районирования Северной Евразии (Равкин и др., 2017) и выполненного по летнему населению птиц той же территории, показало, что первое из них больше соответствует зональным представлениям, т.е. совпадение с зональными рубежами выше, чем по населению птиц. В частности, прослежены самостоятельность предтундровых редколесий, а также особенности кавказских и муссонных территорий. По населению птиц эти отличия проявляются на уровне подтипов, степные сообщества более похожи на пустынно-луговые, а по флоре и фауне в целом входят в таежно-степную подобласть. Последнее объединение связано с постепенными изменениями населения птиц внутри этой подсовокупности. При этом уссурийско-сахалинские орнитокомплексы входят в редколесно-лесной тип населения птиц. В принципе, значительное перекрывание таксонов районирования разных групп объектов классификации вполне понятно, так как главным структурообразующим фактором во всех случаях является гидротермический режим, хотя реакция различных животных и растений на него неоднозначна. Кроме того, при комплексном районировании влияют и различия в соотношении разных групп и степень формализации при классифицировании. Сходство и различие выполненных вариантов районирования иллюстрирует многомерность различий биотических групп и “сплетение” тенденций изменений, а также их частичную фрактальность.
Фрактальность орнитокомплексов
Сравнение факторов среды, выявленных на локальных территориях (Е. Равкин, Ю. Равкин, 2005; Цыбулин, 2009; Равкин и др., 2016) и в целом по Северной Евразии, показало значительное сходство факторов и режимов, определяющих территориальную неоднородность орнитокомплексов или коррелирующих с их изменчивостью. В изменениях водно-околоводных и рекреационно-селитебных сообществ тоже прослежено сходство не только отдельных факторов среды, но и их неразделимых сочетаний (природно-антропогенных режимов), а также повторяемость пространственных трендов в населении птиц. Поэтому можно говорить о его географической фрактальности, как значительном сходстве (повторяемости) влияния основных факторов среды независимо от масштаба и ранга рассмотрения. Например, это гидротермический режим, который определяет ландшафтно-зональную неоднородность фитоценозов и, соответственно, населения птиц через кормность, в том числе антропогенную, и растительность в выделенных надтипах населения (застроенной и незастроенной суши, а также водно-околоводных местообитаний). Мандельброт (2002), который ввел понятие “фрактал”, считал подобное масштабной инвариантностью – степенью неправильности, неизменной при всех масштабах рассмотрения. Дело не только в масштабной инвариантности (как в случае в береговой линией на разных картах), но и в ранговой (лист, ветка, дерево или в нашем случае – подзона, зона, регион), причем не только неправильности, но и правильности в повторяемости, по крайней мере, после идеализации, которую проводят для облегчения восприятия (визуализации). Основными признаками фрактальности считают самоподобие частей и целого, вложенность (из-за различий в масштабах) таксонов классификаций друг в друга и почти бесконечную возможность расчленения анализируемых совокупностей (Маврикиди, 2000). В.В. Исаева (2003, с. 40) считает, что “Природные, в частности, биологические структуры – стохастические, хаотические фракталы, или квазифракталы; повторяемость их структуры в разном масштабе неполна и неточна – это “обрубленные” фракталы. Некоторые исследователи, например, С.Д. Хайтун (1996), приходят к заключению, что фракталы не являются реально существующими объектами, а реальные системы могут быть только фракталоподобными”. Географическую квазифрактальность населения птиц образуют сходные тренды на различных территориях: провинциях, регионах, физико-географических странах и при анализе их совокупностей по частям и в целом.
С.М. Цыбулин (2009) отмечал фрактальность населения птиц Алтая, где орнитокомплексы южной провинции представляют собой уменьшенную копию таковых на территории всего Алтая. В остальных провинциях наборы местообитаний и, соответственно, вариантов населения птиц неполны. Так, в Северо-Восточном Алтае, в частности, нет степей, в Юго-Восточном – черневой тайги, в Северо-Предалтайской провинции – темнохвойных лесов. Однако в целом провинции, даже без Южного Алтая, фрактальны по отношению ко всей территории Алтайской горной области. Фрактальность в населении птиц проявляется и в других провинциях Алтая и иных горных системах в общем характере высотно-поясных трендов (полулунности) и параллельности рядов по увлажнению (аридного и гумидного).
Возможность чуть ли не бесконечного дробления анализируемых совокупностей подтверждает иерархия всех классификаций (надтипы, типы, подтипы, классы, подклассы с допустимостью дальнейшего деления), а также возможность сбора и анализа данных с любого уровня ландшафтной или геоботанической классификаций и даже с большей дробностью, чем по их минимальным таксонам (Харвей, 1974). Кроме того, этот признак фрактальности подтверждает возможность анализа животного населения не только на видовом и популяционном уровнях, но и при делении по полу, возрасту, доминированию в популяциях, а также по сезонам года, “историческим” отрезкам времени и т.д., почти до бесконечности.
Фрактальность (квазифрактальность) изменений орнитокомплексов четко видна на графах по притихоокеанской части России (Равкин и др., 2020). Фрактальностью здесь можно считать параллельность широтных и высотных изменений сообществ во всех надтипах населения – на незастроенной и застроенной суше, а также в водно-околоводных материковых и морских сообществах. То же самое прослежено и по всей Северной Евразии. Все типы населения птиц или, по крайней мере, на незастроенной суше, представимы в виде ромбов. Ромбовидность связана с уменьшением площадей, занимаемых тем или иным типом орнитокомплексов, а также плотности населения птиц от середины в распространении типа к северу и югу (в горах вниз и вверх по абсолютным высотам местности) с проникновением в менее свойственные им зоны и пояса. При этом болотный тип, антропогенные и водно-околоводные орнитокомплексы не образуют сплошных полос, а представлены отдельными пятнами или имеют линейный характер.
Орнитокомплексы морских акваторий образуют третий вариант меридионально-широтной неоднородности, соответственно в полярной и тихоокеанской частях. Для более определенной характеристики по ним пока не хватает данных.
Проникновение Тундрово-стланикового типа населения птиц к югу на равнинах отмечено вплоть до северной тайги, а Пустынно-луговостепного – к северу до тундровой зоны за счет пойменных лугов. Проникновение редколесно-лесного типа зарегистрировано вплоть до южной тундры, а к югу – до пустынной зоны в пределах границ рассмотрения. Проникновение Болотного типа, не имеющего сплошного зонального ареала, отмечено до тундровой зоны на севере и до границы рассмотрения к югу.
Таким образом, все типы населения птиц незастроенной суши Северной Евразии могут быть условно (в факторном пространстве) обозначены на схеме ромбами. Они иллюстрируют зонально-подзональную и высотно-поясную континуальность населения птиц (Равкин, 1973). Граница между северными и южными орнитокомплексами застроенных территорий смещена к северу по сравнению с таковыми естественных ландшафтов суши и проходит между лесотундровой и лесной зонами. В населении птиц озер и рек северная граница может быть проведена также между лесотундровой и лесной зонами, но появляется еще одна – южная, между лесной зоной и лесостепью. Широтных границ в населении птиц морских акваторий и базаров больше, но они малообоснованны из-за особенностей учета морских птиц на длительных по протяженности маршрутах, не разделенных на участки.
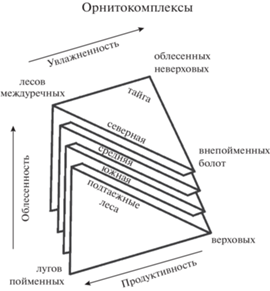
Сходство трендов в изменчивости населения птиц Срединного региона Северной Евразии и алтайских провинций по отдельности и в целом (Равкин, 1973; Цыбулин, 1999, 2009; Равкин и др., 2016) тоже свидетельствует о фрактальности орнитокомплексов. Самоподобие и вложенность таксонов классификации населения птиц четко видны на пространственно-типологических структурах сообществ различных подзон лесной зоны Западной Сибири (Равкин и др., 1978). В двумерном пространстве фрактальность очевидна в “треугольности” основных отличий орнитокомплексов всех четырех подзон. Изменения в них идут между орнитокомплексами лесов междуречий, пойменных лугов и верховых болот. Причины этого – в уменьшении облесенности от междуречий к поймам, а также в увеличении увлажненности и уменьшении продуктивности от тайги и пойменных лугов к верховым болотам. Эти изменения связаны с влиянием фрагментированности из-за сельскохозяйственного освоения территории, а также с заливанием в половодье, заболоченностью и уменьшением трофности болот. В трехмерном факторном пространстве эти изменения иллюстрируют “встроенные” (частично вложенные) друг в друга пирамиды, где на третьей оси отражены изменения населения по отличиям в продуктивности фитоценозов (рис. 6). Эти схемы свидетельствуют и о самоподобии орнитокомплексов лесной зоны и четырех ее подзон, а также о “встроенности” этих структур друг в друга по зоне в целом.
Таким образом, фактически квазифрактальность была четко прослежена нами буквально с первых шагов формализованного анализа населения птиц, хотя сам термин мы не употребляли. Изменчивость орнитокомплексов вполне адекватно может быть описана и без употребления термина “фрактальность”. Отсутствие значимой “новизны” при последовательном анализе сообществ птиц разных провинций, подзон и регионов породило явное разочарование. Фрактальные представления снимают это “недовольство” за счет утверждения, что население птиц исследуемой нами территории в большей или меньшей степени фрактально, поэтому повторы в представлениях закономерны и подтверждают это.
Второй повод для недовольства при исследовании животного населения и, в частности, орнитокомплексов, связан с необходимостью каждый раз начинать все с нуля: с кластерного анализа собранных данных, выявления основных структурообразующих факторов среды и оценок их связи с изменчивостью сообществ на данной территории. При этом увеличение объема анализируемой выборки не приводит к улучшению (увеличению) показателей информативности представлений (учитываемой ими дисперсии матрицы сходства). Даже наоборот увеличение выборки формально ухудшало эти показатели в результате накопления числа случайных, частных и необъяснимых отклонений при интерпретации изменчивости сообществ. Удручало и то, что сведения, полученные на ранних этапах, не используются при оценке информативности более поздних общих представлений, т.е. эти обобщения не включали частностей, нередко четких и “красивых”, которые были отмечены при анализе в более крупном масштабе. Фрактальные представления логически позволяют включать в оценку информативности представлений не только выводы о зональных трендах, но и о подзональных изменениях, не прослеживаемых на обобщенных графах.
Признание квазифрактальности животного населения в целом и орнитокомплексов в частности представляет несомненный теоретический интерес, но хотелось бы оценить информативность этих представлений при описании изменчивости орнитокомплексов рассматриваемых территорий. Это можно сделать, и делалось ранее, через учитываемую (объясненную) дисперсию матрицы коэффициентов сходства сообществ составленными классификациями и расчета значений множественного коэффициента корреляции (регрессии) для каждой из них. Приращение оценок дает включение в расчеты связи территориальной неоднородности животного населения с факторами среды и объяснения результатов кластерного анализа в виде классификаций и структур, т.е. классификаций с учетом значимого сходства между таксонами, приведенными на графах.
Обычно мы сначала отдельно рассчитывали связь с выявленными факторами среды, затем дисперсию, учитываемую классификацией по всем иерархическим уровням, а также и структурой на каком-либо одном уровне классификации. По орнитокомплексам Северной Евразии, рассмотренным в настоящей публикации, объясняемая всеми факторами среды дисперсия равна 48%, а классификациями с учетом деления на надтипы, типы, подтипы и классы населения птиц – 41% (вместе 52%, множественный коэффициент корреляции – 0.72). Естественно, это не полная оценка, рассчитать которую нельзя из-за нереализованной возможности почти бесконечного разделения выборок и их анализа. Однако включение в расчеты структурных представлений, полученных раздельно по физико-географическим странам Северной Евразии, величину вклада фрактальности значимо не увеличило.
Бóльшая оценка вклада фрактальности может быть получена при использовании методов мультифрактального анализа (Гелашвили и др., 2008). Некоторое увеличение общей информативности представлений может быть достигнуто и при дальнейшем делении на подклассы, семейства сообществ и т.д., но это приведет к существенному увеличению числа таксонов классификации и сложности ее объяснения и восприятия.
ЗАКЛЮЧЕНИЕ
Анализ всех материалов по летнему населению птиц Северной Евразии, накопленных в банке данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН, показал или подтвердил ранее выявленные нами по отдельным регионам следующие представления и особенности территориальных изменений орнитокомплексов и связи их с факторами среды.
1. Сравнительно высокую степень корреляции неоднородности среды и летнего населения птиц Северной Евразии (множественный коэффициент 0.72).
2. Большее влияние на неоднородность орнитокомплексов типа растительности по сравнению с зонально-поясными отличиями среды.
3. Диагональное смещение сходства сообществ птиц по отношению к зонально-подзональным границам растительного покрова, как результат интегрального влияния зональности и провинциальности (секторности), а также отепляющего влияния морских течений и связанной с этим меньшей мерзлотностью почвогрунтов.
4. Проявление континуальности (типологической постепенности) изменений населения птиц возрастает в выборках по мере увеличения объема анализируемого материала за счет увеличения ландшафтной разнородности территории.
5. Фрактальность (квазифрактальность) основных трендов изменчивости населения птиц – самоподобие целого и его частей (орнитокомплексов зон, подзон, провинций и физико-географических стран). Это подтверждает повторение на исследованных территориях криволинейности (полулунности) рядов изменений в горах и похожей “треугольности” сходства сообществ “открытых” зон по отношению к лесным на равнине, а не строгую хорологическую вертикальность смены по высотам местности и горизонтальность широтных отличий на равнине. Кроме того, о фрактальности (квазифрактальности) населения птиц свидетельствует идентичность набора и иерархии значимости основных факторов среды, определяющих структуру населения птиц.
6. В предшествующих исследованиях при экспертно-умозрительной оценке сходства орнитокомплексов была несколько завышена значимость ландшафтно-зональной (подзональной и поясной) изменчивости условий среды в формировании населения птиц и в значительной степени орнитогеографических различий по Уралу и Енисею. При формализованном анализе изменчивости населения птиц по коэффициентам сходства существенность этих меридиональных границ не прослежена, но выявлена бóльшая значимость своеобразия орнитокомплексов притихоокеанской территории, за счет проникновения берингийских и китайских видов.
7. Результаты анализа данных за разные годы (до 28 и 44 лет) в одних и тех же местах и биотопах показали, что межгодовые отличия населения птиц не превышают территориальные, что подтверждает правомерность использования данных, собранных в разные годы для анализа пространственной неоднородности орнитокомплексов на больших площадях.
Список литературы
Атлас СССР, 1983. М.: ГУГК при СМ СССР. 260 с.
Блинова Т.К., Равкин Ю.С., 2008. Орнитофаунистическое районирование Северной Евразии // Сиб. экол. журн. Т. 15. № 1. С. 101–121.
Воронов А.Г., Кучерук В.В., 1977. Биотическое разнообразие Палеарктики: проблемы изучения и охраны // Биосферные заповедники. Тр. I сов.-амер. симп. СССР, 15–17 мая 1976 г. Л.: Гидрометеоиздат. С. 7–20.
Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н., Солнцев Л.А., 2008. Основы мультифрактального анализа видовой структуры сообщества // Успехи соврем. биологии. Т. 128. № 1. С. 21–34.
Ефимов В.М., Равкин Ю.С., 1980. Еще раз о пространственной структуре населения птиц Северо-Восточного Алтая // Проблемы зоогеографии и истории фауны. Новосибирск: Наука. С. 59–63.
Железнова Т.К., 2017. Птицы в нефтегазоносных районах Привасюганья. М.: ИПО “У Никитских ворот”. 140 с.
Железнова Т.К., Вартапетов Л.Г., 2018. Птицы среднетаежного Притымья (Западная Сибирь). М.: ИПО “У Никитских ворот”. 196 с.
Железнова Т.К., Костылева Н.А., 2018. Птицы болот лесной зоны Западной Сибири. М.: ИПО “У Никитских ворот”. 160 с.
Железнова Т.К., Леппа В.А., 2016. Птицы Томского Прикетья. М.: Проспект. 224 с.
Железнова Т.К., Миловидов С.П., Блинов Л.В., 2021. Птицы города Томска. Т. 1. М.: ИПО “У Никитских ворот”. 376 с.
Жуков В.С., 2006. Птицы лесостепи Средней Сибири. Новосибирск: Наука. 492 с.
Иванов А.И., 1976. Каталог птиц СССР. Л.: Наука. 276 с.
Исаева В.В., 2003. Синергетика для биологов. Вводный курс. Учеб. пособие. Владивосток: Изд-во ДВФУ. 125 с.
Климова Н.В., Торопов К.В., 2018. Птицы города Кемерово и его окрестностей. Новосибирск: Академическое изд-во “Гео”. 178 с.
Кузякин А.П., 1962. Зоогеография СССР // Учен. зап. МОПИ им. Н.К. Крупской. Т. 109. С. 3–182.
Куперштох В.Л., Трофимов В.А., 1975. Автоматическое выявление макроструктуры системы // Проблемы анализа дискретной информации. Ч. 1. Новосибирск: ИЭиОПП СО АН СССР. С. 67–83.
Ливанов С.Г., 2021. Птицы Урала. Новосибирск: Изд-во СО РАН. 226 с.
Маврикиди Ф.И., 2000. Фракталы: постигая взаимосвязанный мир // Дельфис. № 23. С. 78–85.
Мандельброт Б., 2002. Фрактальная геометрия природы. М.: Ин-т комп. исследований. 656 с. [Mandelbrot B., 1982. The Fractal Geometry of Nature. San Francisco: W.H. Freeman and Co. 460 p.].
Птушенко Е.С., Иноземцев А.А., 1968. Биология и хозяйственное значение птиц Московской области и сопредельных территорий. М.: Изд-во МГУ. 462 с.
Равкин Е.С., Равкин Ю.С., 2005. Птицы равнин Северной Евразии: Численность, распределение и пространственная организация сообществ. Новосибирск: Наука. 304 с.
Равкин Е.С., Челинцев Н.Г., 1999. Методические рекомендации по маршрутному учету населения птиц в заповедниках // Организация научных исследований в заповедниках и национальных парках. Сб. докл. семинара-совещания, г. Пущино-на-Оке. М.: Всемирный форд дикой природы. С. 143–155.
Равкин Ю.С., 1967. К методике учета птиц в лесных ландшафтах // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука. С. 66–75.
Равкин Ю.С., 1973. Птицы Северо-Восточного Алтая. Новосибирск: Наука. 375 с.
Равкин Ю.С., 1978. Птицы лесной зоны Приобья. Новосибирск: Наука. 288 с.
Равкин Ю.С., Ефимов В.М., 2009. Банк данных по численности и распределению животных в пределах бывшего СССР // Формирование баз данных по биоразнообразию – опыт, проблемы, решения. Мат-лы Междунар. науч.-практ. конф. Барнаул: “Артика”. С. 205–214.
Равкин Ю.С., Ливанов С.Г., 2008. Факторная зоогеография. Новосибирск: Наука. 205 с.
Равкин Ю.С., Куперштох В.Л., Трофимов В.А., 1978. Пространственная организация населения птиц // Птицы лесной зоны Приобья / Под ред. Равкина Ю.С. Новосибирск: Наука. С. 253–269.
Равкин Ю.С., Богомолова И.Н., Цыбулин С.М., 2015. Фаунистическое районирование Северной Евразии // Изв. РАН. Сер. Геогр. № 3. С. 29–40.
Равкин Ю.С., Богомолова И.Н., Цыбулин С.М., 2017. Экологическая организация флоро-фаунистической неоднородности Северной Евразии // Принципы экологии. № 1. С. 104–116.
Равкин Ю.С., Бабенко В.Г., Стишов М.С., Пронкевич В.В., Лялина М.И., 2020. Эколого-географическая изменчивость летнего населения птиц притихоокеанской части России // Сиб. экол. журн. № 6. С. 689–703. [Ravkin Yu.S., Babenko V.G., Stishov M.S., Pronkevich V.V., Lyalina M.I., 2020. Ecogeographical vareabelity of the sammer bird population in the pacific part of Russia // Contemp. Probl. Ecol. V. 13. № 6. P. 577–589].
Равкин Ю.С., Богомолова И.Н., Цыбулин С.М., Железнова Т.К., Торопов К.В. и др., 2016. Пространственно-типологическая неоднородность и экологическая организация летнего населения птиц Срединного региона Северной Евразии // Сиб. экол. журн. Т. 9. № 1. С. 103–116. [Ravkin Yu.S., Bogomolova I.N., Tsybulin S.M., Zheleznova T.K., Toropov K.V., et al., 2016. Spatial-typological heterogeneity and environmental organization of the summer population of birds in the Middle Region of Northern Eurasia // Contemp. Probl. Ecol. V. 9. № 1. P. 86–97].
Торопов К.В., Бочкарева Е.Н., 2014. Птицы подтаежных лесов Западной Сибири: 30–40 лет спустя. Новосибирск: Наука-центр. 394 с.
Трофимов В.А., 1976. Качественный факторный анализ матриц связи в пространстве разбиений со структурой // Модели агрегирования социально-экономической информации. Новосибирск: Наука. С. 24–36.
Хайтун С.Д., 1996. Механика и необратимость. М.: Янус. 446 с.
Харвей Д., 1974. Научное объяснение в географии. М.: Прогресс. 503 с.
Цыбулин С.М., 1999. Птицы Северного Алтая. Новосибирск: Наука. 519 с.
Цыбулин С.М., 2009. Птицы Алтая. Новосибирск: Наука. 234 с.
Штегман Б.К., 1938. Основы орнитофаунистического деления Палеарктики / Фауна СССР. Птицы. Т. 1. Вып. 2. М.; Л.: АН СССР. 156 с.
Энциклопедический словарь географических терминов, 1968. М.: Советская энциклопедия. 437 с.
Connor E.F., Simberloff D., 1983. Interspecific competition and species co-occurrence patterns on islands: Null models and the evaluation of evidence // Oikos. V. 41. № 3. P. 455–465.
Connor E.F., Simberloff D., 2014. Neutral models of species' co-occurrence patterns // Ecological Communities: Conceptual Issues and the Evidence / Eds Strong D.R., Simberloff D., Abele L.G., Thistle A.B. Princeton: Princeton Univ. Press. P. 316–331.
Diamond J., 1975. Assembly of species communities // Ecology and Evolution of Communities / Eds Cody M., Diamond J. Cambridge: Belknap. P. 342–444.
Gao M., Liu D., Lin L., Wu D., 2016. The small-scale structure of a soil mite metacommunity // Eur. J. Soil Biol. V. 74. P. 69–75.
Gotelli N.J., Graves G.R., 1996. Null Models in Ecology. Washington: Smithsonian Institution Press. 368 p.
Götzenberger L., Bello F., de, Bråthen K.A., Davison J., Dubuis A., et al., 2012. Ecological assembly rules in plant communities–approaches, patterns and prospects // Biol. Rev. Camb. Philos. Soc. V. 87. № 1. P. 111–127.
Hayne D.W., 1949. An examination of the strip census method for estimating animal populations // J. Wildlife Manag. V. 13. № 2. P. 145–147.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии