

Журнал общей биологии, 2022, T. 83, № 3, стр. 235-240
Пространственные инварианты биологических структур
Ю. Л. Войтеховский 1, 2, *, М. А. Чукаева 2
1 Геологический институт ФИЦ КНЦ РАН
184209 Апатиты, ул. Ферсмана, 14, Россия
2 Санкт-Петербургский горный университет
199106 Санкт-Петербург, 21-я линия, 2, Россия
* E-mail: woyt@geoksc.apatity.ru
Поступила в редакцию 08.04.2022
После доработки 27.04.2022
Принята к публикации 12.05.2022
- EDN: KDJLVA
- DOI: 10.31857/S0044459622330016
Аннотация
Показано, что во многих биологических полигональных и полиэдрических структурах в диапазоне от микрометров (брохосомы, радиолярии, пыльца) до сантиметров (грибы-фуллерены) явно или неявно присутствуют инварианты, обусловленные даже не влиянием физической среды, а геометрией евклидова пространства. По-видимому, их допустимо рассматривать как точки схождения принципов номогенеза и приспособительного онтогенеза биологических индивидов. В качестве первых выступают теоремы Эйлера и Эберхардта. Признаки приспособления в каждом случае следует искать в отклонениях от пространственных инвариантов. Авторы обобщают ранее рассмотренные биологические структуры, обращают внимание на ряд новых и дают интерпретацию найденных закономерностей в свете общебиологической проблематики.
Биологию и минералогию объединяет не только идущее от Линнея выделение видов и индивидов, но и общие принципы развития. Таков, например, принцип Кюри, позволяющий восстановить вещественные потоки и полевые градиенты физической среды по диссимметрии индивида, т.е. по отклонению его симметрии от идеальной, диктуемой внутренним императивом. Нормальная форма минерального индивида – выпуклый полиэдр. Каждая его грань – плоская атомная сетка. Между структурой и формой кристалла есть строгая математическая связь. Среди организмов тоже немало квазиполигональных и квазиполиэдрических структур, образованных сетками, мембранами, оболочками (Геккель, 1902, 2007; Thompson, 1942). Одни различимы непосредственно, другие выявляются несложными преобразованиями. Их анализ обнаруживает инварианты, обусловленные даже не характером физической среды, а более фундаментальной причиной – геометрией евклидова пространства. В данной статье авторы обобщают ранее рассмотренные биологические структуры, обращают внимание на ряд новых и дают интерпретацию найденных закономерностей в свете общебиологической проблематики.
ПОЛИГОНАЛЬНЫЕ СТРУКТУРЫ
Явные
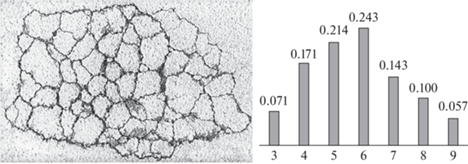
Полигональные разбиения можно видеть на слабо искривленных поверхностях ископаемых (например, Disphyllum caespitosum – верхний девон, Pleurodictyum sp. – нижний карбон, Petalaxis stylaxis – средний карбон, и мн. др.) и современных колониальных кораллов. Нарастая на субстрат, кораллиты теснят друг друга, механически конкурируя за пространство подобно минеральным индивидам на начальной стадии роста, т.е. в зоне геометрического отбора. В обоих случаях выживают индивиды, ориентированные направлением наибольшей скорости роста ортогонально поверхности нарастания. Итог – параллельно-шестоватые агрегаты-колонии с полигональными разбиениями аппроксимирующих поверхностей. Авторами изучены три колонии современного коралла Favites sp. (рис. 1, 15 см по горизонтали) с близкими стандартными статистическими характеристиками: мода (Мо) 6, средняя координация полигона (с) 5.98; среднеквадратическое отклонение (σ) 0.97, асимметрия (As) –0.01, эксцесс (Ex) –0.50. Гистограмма показывает частоту полигонов с координациями от 3 до 9.
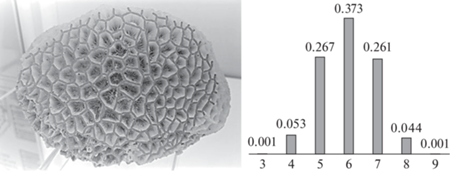
Неявные
Более изогнутая, но все же незамкнутая поверхность характерна для одуванчика обыкновенного Taraxacum officinale. После удаления папиллусов обнаруживается ячеистая структура. Перегородки образуют полигональное разбиение поверхности (рис. 2, диаметр 3 см). Для одного образца получены характеристики: Мо = 6, с = 5.97; σ = 0.66, As = 0.03, Ex = –0.68.

ПОЛИЭДРИЧЕСКИЕ СТРУКТУРЫ
Явные
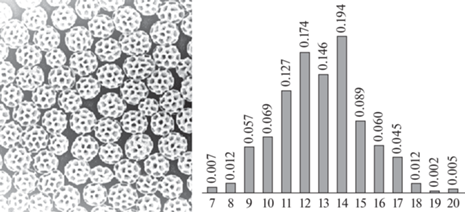
Удивительный пример явной полиэдрической структуры – брохосомы (от др.-греч. βρóχoς – ячея сети, σωμα – тело). Это квазисферические полые образования размером 0.2–0.7 мкм (рис. 3), выделяемые насекомыми семейства цикадок (Cicadellidae). Установлены с помощью электронной микроскопии в начале 1950-х. Состоят из белков и липидов. Механизм формирования и назначение не ясны. Предположительно, насекомые наносят их на тело и кладки яиц, делая их гидрофобными. На первый взгляд, это фуллерены, т.е. выпуклые полиэдры с 5- и 6-угольными гранями, сходящимися по 3. На более чем 400 изображениях подсчитано число видимых граней от 7 до 20. Их общее число (удвоенное число видимых плюс 2–8 боковых, ортогональных к плоскости рисунка) варьирует от 16 до 48. Оценка условна, но заметим, что “нобелевский” фуллерен С60 с 32 гранями попадает в центр диапазона. На некоторых брохосомах есть 4-угольные грани, что заставляет определить их в целом как квазифуллерены (Войтеховский, Степенщиков, 2018). Разнообразие их морфотипов требует изучения.
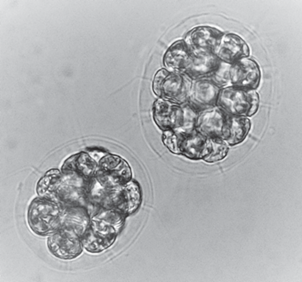
Еще один пример явной полиэдрической структуры – ценобиальная зеленая водоросль Pandorina morum (Müll.) Bory, подробно охарактеризованная ранее (Войтеховский, 2001; Войтеховский и др., 2006). Особенность их морфологии – 16-клеточные ценобии типа фуллеренов (рис. 4) размером около 50 мкм, относящиеся к точечным группам симметрии –43m и 222 в пропорции 3 : 1. Еще более интересными должны быть многоклеточные V. aureus (от 100 до 1000 клеток), V. polychlamys (от 1100 до 2000) и V. globator (от 1500 до 20000). Разнообразие их морфотипов не изучено.
Неявные
Квазисферическую поверхность образует в возбужденном состоянии Diodon holocanthus (сем. Diodontidae, рыбы-ежи, англ. porcupinefishes). Принимая основания игл за точки и используя технику триангуляции поверхности по Делоне и дуального построения по Вороному–Дирихле (Галиулин, 2009), получены полигональные разбиения поверхностей для 8 образцов размером 10–15 см. Их средние характеристики: Мо = 6, с = 5.97; σ = = 0.58, As = 0.01, Ex = –0.17 (рис. 5). Ранее показано, что обитающее рядом с Diodontidae сем. Ostraciontidae (кузовковые, англ. boxfishes) реализует те же полигональные разбиения в покрытиях костными пластинами. Это указывает на родство в составе отряда Tetraodontiformes (Войтеховский, 2009).
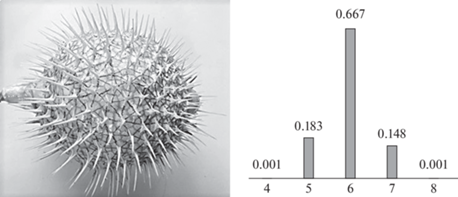
Неизученные
Замечательную полигональную сеть образует съедобный бамбуковый гриб Phallus indusiatus из семейства веселковых (рис. 6а, размер до 25 см). Стандартная характеристика любой сети требует немалого числа клеток. Этот гриб вполне обеспечил бы представительную статистику (порядка 500 полигональных клеток), но в рассматриваемом аспекте совершенно не изучен.
Рис. 6.
Неизученные полиэдрические структуры: а – гриб Phallus indusiatus; б – радиолярия Actinomma antarctica; в, г – морской пузырь Echinosphaerites aurantium; д – гриб Ileodictyon cibarium; е – пыльца растений: 1 – Ipomoea purpurea, 2 – Sidalcea malviflora. Источник: а–в, д, е – интернет, открытый доступ; г – по Бодылевский, 1990, с. 173.

Скелеты радиолярий – широко известный пример ажурных полигональных и полиэдрических структур размером от 50 мкм до 1 мм. Впервые о них заявил Э. Геккель (1902), зафиксировавший полиэдрические мотивы в названиях Circogonia dodecahedra и C. icosahedra. При этом заметим, что додекаэдр – простейший из фуллеренов, а икосаэдр – геометрически дуальная форма. Квазифуллереновые мотивы в строении большого числа других Radiolaria видны на многочисленных микрофотографиях. Например, такова Actinomma antarctica (рис. 6б). Российский математик Д.Д. Мордухай-Болтовской (1936) обосновал их оптимальность как строительной конструкции независимо от американского архитектора Р.Б. Фуллера. Нюансы сложных скелетов (статистика пор по числу соседей и пр.) систематически не изучались. Есть основания полагать, что они имеют отношение к таксономии радиолярий (Voytekhovsky, Stepenshchikov, 2016).
Морской пузырь (Echinosphaerites aurantium) (рис. 6в, г) – ископаемый (средний и верхний ордовик) – организм из класса цистоидей, тип иглокожие. Формировал известковую квазисферическую чашечку диаметром до 4–5 см из многочисленных беспорядочно разбросанных полигональных пластинок (Бодылевский, 1990, с. 49, 173). Ее полиэдрический характер не нарушают ни место прирастания стебля, ни тонкие поры, пронизывающие пластины. Судя по изображениям, полигоны окружены от 4 до 8 соседями с модой 6. Несмотря на множество хорошо сохранившихся образцов, статистика полигонов не изучалась.
Явную полиэдрическую структуру имеют грибы илеодиктион съедобный (Ileodictyon cibarium) (рис. 6д), илеодиктион изящный (I. gracile) и решеточник красный (Clathrus ruber). По сути, их тело размером 5–10 см – это реберная сеть выпуклого полиэдра фуллеренового типа, близкого к С60. На имеющихся изображениях не удалось увидеть граней (ячеек), уверенно отличных от 5- и 6-угольных. Место прирастания гриба не портит структуру. Ее изучение осложнено лишь относительной редкостью в экваториальных и тропических лесах.
Много явных и неявных полиэдрических структур можно видеть в пыльце растений размером от 0.0025 до 0.25 мм. Пример первой – ипомея (Ipomoea purpurea) (рис. 6е, 1), второй – сидальцея (Sidalcea malviflora) (рис. 6е, 2). У последней полиэдрическая структура обнаруживается так же, как у Diodon holocanthus – триангуляцией поверхности по Делоне и дуальным построением по Вороному–Дирихле.
Немного математики
Для любого плоского графа (полигональной сети) выполняется теорема Эйлера:
где f – число граней (клеток), e – ребер, $v$ – вершин. Если из каждой вершины выходят ровно три ребра, тоТогда e = 3f – 6, а средняя координация клетки равна:
Равенство с = 6 в (3) достигается в хорошо известной биологической структуре – пчелиных сотах. Это правильное разбиение поверхности (одно из трех) в кристаллографии имеет символ 63. Каждый трехсвязный плоский граф (т.е. из каждой вершины выходят не менее трех ребер) можно расправить в выпуклый полиэдр. То есть с → 6 – инвариант для простых (в каждой вершине сходятся три грани) плоских графов и выпуклых полиэдров. Выше рассмотрены именно такие биологические структуры. При этом средняя координация в (3) стремится к 6 снизу. Но для трещин в асфальте (рис. 7) имеем полную аналогию: f = 71, e = 207, $v$ = 138, Мо = 6, c = 5.74, σ = 1.59, As = 0.23, Ex = –0.82. Этот пример, не имеющий отношения к биологии, подчеркивает универсальный характер инварианта с = 6.
Для простых сетей и полиэдров найдем еще одно важное соотношение. Умножив (1) на 6 и прибавив удвоенное (2), получим
Далее выразим общее число граней через их разнообразие:
где fn – число n-угольных граней. Через них же выразим число ребер:
Умножив (5) на 6 и вычтя (6), получим:
С учетом (4) преобразуем (7) к виду:
Уравнение Эберхардта (8) охватывает все рассмотренные выше случаи, с одной стороны, допуская вариации статистик n-угольных граней, с другой стороны, налагая на них строгое соотношение. Для фуллеренов из (8) следует жесткое условие f5 = = 12 без видимых ограничений на f6. Оно может быть любым (от 0 у додекаэдра) кроме 1. Последнее доказывается построением реберного графа.
ОБСУЖДЕНИЕ И ВЫВОДЫ
Формы минеральных и биологических индивидов, взятые в деталях, к тому же непрерывно меняющиеся в онтогенезе, бесконечно разнообразны. Менее или более удачные систематики, огрубляя, позволяют обуздать это многообразие и рассмотреть всякую форму как компромисс между внутренним императивом (требованием) и внешней средой. Требование структуры к форме кристалла имеет статус теоремы: точечная группа симметрии формы (их 32) есть фактор-группа пространственной группы симметрии структуры (их 230) по подгруппе трансляций. Аналогичного принципа для биологических структур неизвестно. Опереться на строгие уравнения не удается. Причина – в их несомненно большей сложности. Тем не менее из рассмотренных примеров и математических соотношений следует ряд выводов.
1) В природе действуют менее или более сложные принципы целесообразности, с разной наглядностью проявляющиеся в минеральных и биологических структурах. Полиэдричность – фундаментальное свойство минеральных индивидов, проявляющееся в условиях свободного роста. Не столь тотально, но полигональность и полиэдричность реализуются в биологических (растительных и животных) структурах в диапазоне от микрометров (брохосомы, радиолярии, пыльца) до сантиметров (грибы-фуллерены), охватывающем 5 размерных порядков.
2) Минеральные и биологические индивиды подвержены влияниям среды, имеющим вещественную (потоки вещества) или полевую (потоки тепла и света, сила тяжести) природу. Для тех и других индивидов искажение внешней формы определяется принципом диссимметрии Кюри. При колониальном (агрегатном) росте на субстрате (подложке) биологические и минеральные индивиды борются за пространство, что приводит к полигональным разбиениям огибающих поверхностей.
3) Рассмотренные примеры указывают на еще одно фундаментальное влияние. Оно обусловлено уже тем, что минеральные и биологические индивиды зарождаются и эволюционируют в трехмерном евклидовом пространстве независимо от его физического наполнения. Это влияние всегда маскируется влиянием физической среды, сливается с ним и, как правило, при исследовании объекта не вычленяется.
4) Природа творит минеральные и биологические структуры, испытывая все допустимые возможности. Порождая полиэдрические и полигональные структуры, она попадает в ловушку соотношений, схватываемых теоремами Эйлера и Эберхардта. В смысле неизбежной подчиненности строгим соотношениям их эволюцию можно назвать номотетической (Берг, 1922, 1977). При этом теоремы допускают неограниченное разнообразие структур, разрешающее их широкие приспособительные (возрастные, экологические) вариации. В полигональных и полиэдрических биологических структурах диалектически сочетаются признаки двух стилей эволюции.
5) Целый ряд биологических полиэдрических структур обнаруживает геометрический дуализм путем триангуляции поверхности по Делоне и дуального построения по Вороному–Дирихле. Для рыб семейств Ostraciontidae и Diodontidae (отряд Tetraodontiformes, иглобрюхообразные) он подчеркивает таксономическую близость, установленную по строению челюстного аппарата. Эта абстрактная геометрическая теория лежит в основании современной кристаллографии. Ее применимость в биологии указывает на единство мира.
Список литературы
Берг Л.С., 1922. Номогенез, или эволюция на основе закономерностей. Петроград: Гос. изд-во. 306 с.
Берг Л.С., 1977. Труды по теории эволюции. Л.: Наука. 387 с.
Бодылевский В.И., 1990. Малый атлас руководящих ископаемых. Л.: Недра. 263 с.
Войтеховский Ю.Л., 2001. О морфологическом разнообразии колоний Pandorina morum (Müll.) Bory (Volvocaceae) // Журн. общ. биологии. Т. 62. № 5. С. 425–429.
Войтеховский Ю.Л., 2009. Геометрические мотивы в морфологии рыб Tetraodontiformes // Журн. общ. биологии. Т. 70. № 3. С. 257–261.
Войтеховский Ю.Л., Степенщиков Д.Г., 2018. Брохосомы – биологические фуллерены // Математические исследования в естественных науках. Апатиты: K & M. С. 150–152.
Войтеховский Ю.Л., Тимофеева М.Г., Степенщиков Д.Г., 2006. Принцип Кюри и морфологическое разнообразие колоний Pandorina morum (Müll.) Bory (Volvocaceae) // Журн. общ. биологии. Т. 67. № 3. С. 206–211.
Галиулин Р.В., 2009. Кристаллографическая геометрия. М.: Либроком. 136 с.
Геккель Э., 1902. Красота форм в природе. СПб.: Т-во “Просвещение”. 120 с.
Геккель Э., 2007. Красота форм в природе. СПб.: Изд-во В. Регена. 144 с.
Мордухай-Болтовской Д.Д., 1936. Геометрия радиолярий // Уч. зап. Ростов-н/Д. гос. ун-та. № 8. С. 3–91.
Thompson D’Arsy W., 1942. On Growth and Form. Cambridge: Cambridge Univ. Press. 1116 p.
Voytekhovsky Y.L., Stepenshchikov D.G., 2016. Fullerene transformations as analogues of radiolarian skeleton microevolution // Paleontol. J. V. 50. № 13. P. 1544–1548.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии