

Журнал общей биологии, 2021, T. 82, № 5, стр. 323-336
Эффективность распределительной системы гидроида Dynamena pumila (L., 1758) при различных абиотических воздействиях
В. С. Дементьев 1, 2, Н. Н. Марфенин 1, *
1 Московский государственный университет им. М.В. Ломоносова,
биологический факультет, кафедра зоологии беспозвоночных
119234 Москва, Ленинские горы, 1, Россия
2 Национальный исследовательский университет “Высшая школа экономики”, факультет биологии и биотехнологии
101000 Москва, Мясницкая ул., 20, Россия
* E-mail: marf47@mail.ru
Поступила в редакцию 25.03.2021
После доработки 01.06.2021
Принята к публикации 12.06.2021
Аннотация
Колониальные гидроиды – пример децентрализованных организмов модульного типа. В них нет органов централизованной регуляции. Распределительная система функционирует за счет множества равнозначных пульсаторов, соединенных трубчатым ценосарком. В результате неуправляемого взаимодействия пульсаторов возникают протяженные течения гидроплазмы (ГПТ) в общей полости колонии. Регулярность и протяженность ГПТ могут быть показателями эффективности децентрализованной организации. Колониальных гидроидов можно рассматривать как удобную модель любых нецентрализованных биологических систем, а распределительную систему колониальных организмов – как разновидность коммуникационных структур и процессов, что и определяет актуальность результатов данного исследования. Работа посвящена определению эффективности (дальности и объемам) транспорта взвешенных в гидроплазме пищевых частиц по столону обитающего на литорали колониального гидроида Dynamena pumila в зависимости от четырех факторов среды. Это температура (10, 15, 20, 25, 28°С), соленость (26, 20, 15, 10‰), осушение (длительностью 5, 10, 20, 30 мин), наличие или отсутствие водообмена. Способность к эффективному транспорту определяется расчетной протяженностью локальных токов гидроплазмы, из цепочки которых складываются непрерывные интегральные гидроплазматические течения (ГПТ). Локальные токи, охватывающие один–два модуля столона и теоретически достаточные для генерации совместных протяженных ГПТ, одинаково представлены почти при всех значениях изученных факторов среды, кроме 28°С и 10‰. Это означает, что распределительная система D. pumila эффективна в широком спектре изменений температуры и солености, а также при продолжительном осушении и при содержании колоний в стоячей воде. Наибольшая эффективность интеграции колонии достигается при 15–20°С, 26–20‰ и после осушения на 5–10 мин. По степени интеграции содержание колоний в стоячей воде и на течении практически не различается. Перенос колоний из одной кюветы с водой в другую не сказывается на интенсивности системы ГПТ.
Децентрализованные модульные организмы, примером которых являются колониальные гидроиды, демонстрируют высокую устойчивость системы за счет повышения пластичности как морфологической, так и функциональной. Благодаря множественному дублированию различных органов (модулей), децентрализованные организмы устойчивы к внешним воздействиям, легко переносят повреждения, распад на фрагменты (Марфенин, 2016). Такие организмы продолжают эффективно и слаженно функционировать даже при, казалось бы, неблагоприятных условиях. Толерантность модульных организмов, как правило, иллюстрируют на примере их роста, размножения и выживания в неблагоприятных условиях, т.е. на примере долгосрочных процессов. Однако модульные организмы – не сообщества особей, а особый тип целостных децентрализованных организмов, в которых есть общеорганизменные системы. У гидроидов это общеколониальная распределительная система. Можно ожидать, что в ее функционировании должен проявляться отклик на внешние воздействия. Для проверки этого предположения можно использовать, по крайней мере, два показателя, а именно: пульсации тела и перемещение гидроплазмы в гастроваскулярной полости. Оба показателя позволяют определить эффективность функционирования всей распределительной системы. Эффективная и слаженная работа распределительной системы необходима для перемещения пищи от места ее приема к удаленным частям колонии, в частности, к растущим верхушкам и молодым побегам (Марфенин, Бурыкин, 1979; Косевич, 1991; Dudgeon, Buss, 1996; Dudgeon et al., 1999).
Эффективность работы распределительной системы выражается в скорости и дальности переноса взвешенных в гидроплазме частиц по колонии за один акт однонаправленного непрерывного гидроплазматического течения, т.е. в формировании быстрых и протяженных течений, связывающих отдаленные части колонии между собой. Протяженные гидроплазматические течения (ГПТ), возникающие время от времени в колонии гидроидов, имеют первостепенное значение для интеграции организма (Бурыкин, 2013; Марфенин, Дементьев, 2017, 2019, 2020). Благодаря им частицы пищи достаточно быстро доставляются к зонам роста – прежде всего, к верхушкам роста столонов и прилегающим к ним участкам, в которых формируются новые побеги или гидранты (Косевич, 1991; Марфенин, 1993; Wagner et al., 1998). Первоочередное обеспечение пищей периферических зон роста позволяет колониальным гидроидам не только увеличиваться в размерах, но и смещаться по субстрату за счет роста столонов, уходя из одних зон и осваивая другие. Поэтому протяженные ГПТ играют ключевую роль в жизнедеятельности колониального организма гидроидов. Необходимо знать пределы изменения основных параметров среды обитания, в которых функционирование системы ГПТ остается эффективным. По реакции распределительной системы на изменение условий существования можно определить степень эврибионтности колониального организма, в частности, степень эвритермности, эвригалинности, а также способность выдерживать осушение. Дополнительно к перечисленным задачам мы стремились определить скорость целостной реакции модульного организма на внешние воздействия и способность сохранять эффективный режим функционирования за пределами преферентных значений показателей среды обитания.
Есть основания предполагать, что выбранный нами объект исследования хорошо адаптирован к крайним отклонениям абиотических показателей от оптимума.
Гидроид Dynamena pumila (L., 1758) приспособлен к жизни в нестабильном биотопе литорали и ежедневно подвергается разнофакторному абиотическому воздействию. Сохранение гидроплазматических течений после внешнего воздействия на колонию свидетельствует о том, что распределительная система продолжает функционировать даже в условиях стресса (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б). Но насколько эффективно? В сущности, эффективность мы определяем через способность распределительной системы быстро доставлять пищевые частицы от одного конца колонии к другому. Чем дальше и в бóльшем количестве могут быть перемещены частицы, тем эффективнее функционирует система ГПТ.
Цель работы – сравнение эффективности функционирования распределительной системы в изученном диапазоне изменения абиотических факторов среды. Мы хотим выяснить, насколько варьирует указанная эффективность при резких изменениях параметров среды и, особенно, при крайних значениях этих параметров, за пределами которых рост колониального организма останавливается.
Используемый нами метод исследования позволяет определять почти моментальную реакцию распределительной системы на изменение параметров среды. Об этой реакции можно судить по характеру пульсаций ценосарка, а также скорости и дальности перемещения гидроплазмы в столонах. Достоверные изменения указанных показателей можно определить с помощью микровидеосъемки в течение одного часа.
МАТЕРИАЛ И МЕТОДЫ
Объект исследования – гидроид Dynamena pumila (L., 1758) (Sertulariidae, Leptothecata, Hydrozoa), многократно использовавшийся ранее в различных экспериментальных исследованиях роста, морфогенеза и экологии. Сбор гидроидов, их культивирование, видеорегистрацию функционирования распределительной системы и обработку данных осуществляли по неоднократно описанной ранее методике (Марфенин, Дементьев, 2017, 2018а, б, 2020; Дементьев, Марфенин, 2018, 2019а, б).
Колонии гидроидов были собраны на малой воде вблизи ББС МГУ на Еремеевском пороге Великой Салмы в Кандалакшском заливе Белого моря. В лаборатории гидроидов выращивали на предметных стеклах в аквариумах с циркулирующей морской водой при постоянной температуре 15 ± 1.5°C, примерно соответствующей температуре моря во второй половине лета. Интенсивную циркуляцию воды в аквариумах осуществляли с помощью аэрации микрокомпрессорами через пористые распылители.
Колонии выращивали из фрагментов отдельных побегов с участка столона: стекло обматывали ниткой несколько раз и подводили под натянутую нитку фрагмент побега, ориентируя его плоскостью среза к стеклу. В процессе репаративной регенерации колония на месте среза прилипала к стеклу выделяемым на верхушке роста хитином, формирующим перисарк. В дальнейшем фиксирующую нить удаляли. Избыточное кормление гидроидов осуществляли ежедневно свежевылупившимися науплиями Artemia salina. Замену воды в аквариумах производили раз в три дня.
Для съемки стекло с колониями помещали в непроточную термостатируемую кювету водоизмещением 0.25 л. Видеосъемка проводилась камерой Arecont-AV3100 через микроскоп при увеличении ×100 с записью изображения на компьютер. Продолжительность съемки каждого эпизода составляла от 1.5 до 8 ч. Режим цейтраферной съемки – 4 кадра/с. Обработку видеозаписей, включающую измерения расстояний верхушки роста от границы кадра, просвета ценосарка, перемещения частиц в гидроплазме, осуществляли вручную.
В опытах использовали маленькие прямолинейные колонии с неразветвленными столонами, большими материнскими побегами (обычно 20–30 пар гидрантов) и небольшими дочерними, включающими по 1–2 паре гидрантов каждый (рис. 1). Таким образом, каждая колония состояла из двух побегов: большого материнского (который мог быть разветвленным) и одного дочернего, а также двух модулей столона – участка между двумя последовательными побегами и участка от самого молодого побега до верхушки роста столона. Использование предельно коротких колоний позволяло яснее представить реакцию распределительной системы на изменение значений факторов среды, ограничиваясь регистрацией ГПТ, исходящих из единственного крупного побега.
Рис. 1.
Обобщенная схема колоний D. pumila, использованных для эксперимента с воздействием факторов среды. Пунктирная стрелка указывает место регистрации.
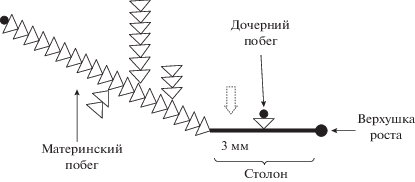
В исследованных колониях были проведены цейтраферные микровидеосъемки первого модуля столона (между первым, считая от верхушки столона, и материнским побегами). Участок между верхушкой столона и первым от нее побегом называется нулевым модулем. В нем мы не проводили регистрацию пульсаций ценосарка и перемещения гидроплазмы. Первый модуль представляет собой сформированный участок колонии (Марфенин, Дементьев, 2017; Marfenin, Dementyev, 2017).
Метод определения основных параметров и эффективности (скорости и дальности перемещения частиц) работы распределительной системы основан на продолжительной регистрации в одном месте всего трех показателей: 1) поперечных пульсаций ценосарка столона; 2) направления и 3) скорости локального ГПТ.
Скорость гидроплазматического течения ($v_{{HF}}^{ - }$, мкм/c) определяли как расстояние, которое проходят распознаваемые частицы в полости столона или побега за 1 с. Этот показатель ($v_{{HF}}^{ - }$) все время изменяется: от 0 мкм/c до максимальных значений и затем снова до 0 мкм/c. Для сравнения можно использовать как усредненные значения $v_{{HF}}^{ - }$ за один цикл пульсации ГПТ, так и максимальные. Последние мы называем амплитудой колебаний скорости ГПТ (АHF). Она равна максимальному значению скорости однонаправленного течения.
Аккумулятивными показателями работы распределительной системы являются: а) протяженность переноса частиц (Z) за один акт однонаправленного локального ГПТ и б) объем перенесенной гидроплазмы (V), содержащей эти частицы.
Расчетная протяженность переноса частиц (Z, мкм) определена на основе регулярных определений скорости перемещения частиц в гидроплазме ($v_{{HF}}^{ - }$). Был выбран интервал регистрации, равный 30 с. Допуская, что в пределах этого интервала скорость неизменна, мы перемножали $v_{{HF}}^{ - }$ на 30 с и получали протяженность перемещенной гидроплазмы за 30 с. Полученные таким способом значения Z складывали и получали расчетную протяженность переноса частиц за однонаправленное ГПТ от старта до его завершения. Следовательно, Z = ${{\sum\nolimits_{i = 1}^n {\left( {v_{{HF}}^{ - }} \right)} }_{i}}$ × ∆t, где i – порядковый номер измерения скорости ГПТ, $v_{{HF}}^{ - }$ – скорость ГПТ (протяженность пробега частицы за 1 с), ∆t – шаг регистрации (в нашем случае 30 с).
Показатель Z представляет сумму непрерывных пробегов частиц в одном направлении с разными скоростями от нулевой до максимальной (показатель АHF) и затем снова до нулевой. Если АHF позволяет количественно оценить интенсивность процесса перемещения частиц по максимальной $v_{{HF}}^{ - }$ за один цикл, то Z дает представление о дальности перемещения фронта ГПТ, так как у ГПТ ясно определяется фронт – граница между пришедшими в продольное движение частицами и продолжающими крутиться на месте.
Объем гидроплазмы (V, мкм3) – объем перенесенной гидроплазмы через поперечное сечение столона за интервал, соответствующий одному ГПТ. Вычисляется, исходя из данных о скорости ГПТ ($v_{{HF}}^{ - }$), продолжительности однонаправленного ГПТ и величины просвета ценосарка столона (D). Принимая условно скорость ГПТ постоянной между двумя последовательными учетами, т.е. за 30 с, получаем сумму значений объемов перемещенной гидроплазмы за каждый полуминутный интервал на протяжении однонаправленного ГПТ по формуле $V = \sum\nolimits_{i = 1}^n {{{(\pi D_{i}^{2})} \mathord{\left/ {\vphantom {{(\pi D_{i}^{2})} 4}} \right. \kern-0em} 4}} $ ($v_{{HF}}^{ - }$)i ∆t, где i – порядковый номер измерения $v_{{HF}}^{ - }$ и D; n – число шагов регистрации; ∆t – шаг регистрации (в нашем случае 30 с).
Протяженность переноса является более информативным показателем эффективности распределительной системы, поскольку без возможности транспортировки по колонии даже значительные объемы гидроплазмы, насыщенные пищей, оказываются бесполезными. Они просто будут скапливаться вблизи получившего пищу гидранта.
Направленные к верхушке столона течения мы называем магистральными (+) ГПТ, от верхушки – компенсаторными (–) ГПТ. Соответственно, мы различаем показатели ГПТ, движущихся дистально (А(+)HF, Z(+)HF, V(+)HF) и проксимально (А(–)HF, Z(–)HF, V(–)HF). Результаты настоящей работы базируются только на характеристиках магистральных (+) ГПТ.
Рассуждая о возможности переноса частиц на дальние расстояния, мы также употребляем термин “протяженность течения”, но подразумеваем в таких случаях протяженность генерализованного ГПТ, состоящего из совокупности локальных ГПТ. Протяженность локальных ГПТ может быть определена численно, исходя из данных о скорости непрерывного перемещения частиц в одном направлении, определенных по результатам видеорегистрации в одном месте. Протяженность генерализованных ГПТ определяют визуально под бинокуляром на малом увеличении, перемещая кювету с колонией (Марфенин, Дементьев, 2019).
В исследовании используются опубликованные ранее данные о влиянии условий среды на D. pumila. Новыми в данной статье являются: во-первых, сравнение реакции распределительной системы колонии на воздействие разных факторов среды и, во-вторых, анализ эффективности функционирования распределительной системы при отклонениях от оптимальных значений показателей состояния среды обитания.
Исследована реакция распределительной системы на четыре абиотических фактора среды в следующих диапазонах:
• температура (10, 15, 20, 25, 28°С);
• опреснение (26, 20, 15, 10‰);
• осушение (длительностью 5, 10, 20, 30 мин);
• отсутствие или наличие водообмена в кювете.
За контрольные значения мы принимаем среднюю соленость воды в месте сбора материала (26‰), температуру культивирования материала (15°С), отсутствие осушения и наличие водообмена.
Всего было исследовано 29 разных колоний D. pumila: шесть – в эксперименте с температурой, шесть – в эксперименте с соленостью, восемь – в эксперименте с осушением, девять – в эксперименте с водообменом. Мы не ставили задачу по изучению адаптации колоний к постепенной смене условий (например, нагреванию или опреснению) и исследовали только кратковременную первичную реакцию на изменение значения фактора среды.
Статистические характеристики представленных в работе результатов (среднее арифметическое значение $\bar {x}$, стандартная ошибка среднего ±SE, выборка n) протяженности и объема гидроплазматических течений в первом модуле столона D. pumila вычислены в программе Excel для всех отснятых при одинаковых условиях (например, при одном и том же значении солености) эпизодов в пределах эксперимента совместно. Эти данные были получены и опубликованы нами ранее (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б). Сводные данные представлены в табл. 1. Несмотря на то, что эксперименты были проведены в разные годы и на различных колониях D. pumila, вполне допустимо сопоставлять данные разных статей, поскольку мы применяли одинаковые условия культивирования, методику регистрации (дистанционную), способы обработки данных и их количественного анализа. Для оценки достоверности полученных величин использовали непараметрический критерий Манна–Уитни.
Таблица 1.
Наибольшая скорость или амплитуда ГПТ (A, мкм/c), протяженность (Z, мкм) и объем ГПТ (V, мкм3) в первом модуле столона D. pumila. Для сравнения указана величина прироста верхушки столона за одну пульсацию (mgp, мкм)
| Фактор среды | Значение фактора | Показатель распределительной системы | mgp | ||||||
|---|---|---|---|---|---|---|---|---|---|
| A(+)HF | Z(+)HF | V(+)HF | |||||||
| $\bar {x}$ ± SE | n | $\bar {x}$ ± SE | n | $\bar {x}$ ± SE | n | $\bar {x}$ ± SE | n | ||
| Температура | 10°С | 25.1 ± 2.4 | 33 | 2192 ± 273 | 40 | 5 505 885 ± 910 439 | 40 | 14.0 ± 0.8 | 51 |
| 15°С | 41.1 ± 4.5 | 45 | 3101 ± 255 | 50 | 7 764 190 ± 1 093 795 | 50 | 15.3 ± 1.1 | 52 | |
| 20°С | 37.3 ± 4.2 | 61 | 2232 ± 248 | 67 | 4 500 050 ± 667 303 | 68 | 15.4 ± 1.0 | 79 | |
| 25°С | 27.4 ± 2.3 | 82 | 1560 ± 134 | 90 | 3 462 456 ± 483 495 | 89 | 7.2 ± 1.8 | 105 | |
| 28°С | 17.0 ± 1.4 | 70 | 1018 ± 95 | 74 | 1 038 013 ± 192 082 | 74 | 1.6 ± 0.8 | 49 | |
| Соленость | 26‰ | 33.8 ± 2.9 | 55 | 2919 ± 300 | 61 | 4 170 077 ± 563 206 | 59 | 12.7 ± 1.1 | 43 |
| 20‰ | 38.9 ± 4.3 | 55 | 2656 ± 252 | 67 | 5 584 866 ± 749 314 | 67 | 15.4 ± 1.2 | 42 | |
| 15‰ | 30.5 ± 2.1 | 57 | 2298 ± 153 | 64 | 4 082 916 ± 385 662 | 63 | 4.9 ± 0.7 | 161 | |
| 10‰ | 23.1 ± 4.2 | 31 | 2257 ± 358 | 34 | 2 935 789 ± 496 458 | 34 | 0.7 ± 0.7 | 9 | |
| Осушение | Без осушения | 23.3 ± 1.6 | 66 | 1994 ± 152 | 79 | 3 888 090 ± 413 221 | 76 | 12.1 ± 1.0 | 56 |
| На 5 мин | 29.5 ± 3.1 | 35 | 2608 ± 284 | 39 | 4 011 455 ± 501 344 | 38 | 11.8 ± 0.9 | 36 | |
| На 10 мин | 29.4 ± 2.7 | 27 | 2612 ± 258 | 41 | 4 139 642 ± 583 980 | 41 | 13.8 ± 0.9 | 69 | |
| На 20 мин | 26.4 ± 2.0 | 53 | 1782 ± 176 | 68 | 2 665 393 ± 355 461 | 68 | 7.4 ± 0.8 | 38 | |
| На 30 мин | 30.5 ± 1.7 | 146 | 2208 ± 121 | 179 | 3 383 962 ± 275 239 | 178 | 3.7 ± 0.7 | 121 | |
| Водообмен | В стоячей воде | 40.5 ± 2.7 | 73 | 2218 ± 152 | 79 | 6 544 997 ± 537 849 | 78 | 14.4 ± 2.7 | 129 |
| В проточной воде | 33.5 ± 2.2 | 51 | 2056 ± 153 | 77 | 5 028 507 ± 396 721 | 76 | 8.0 ± 3.1 | 119 | |
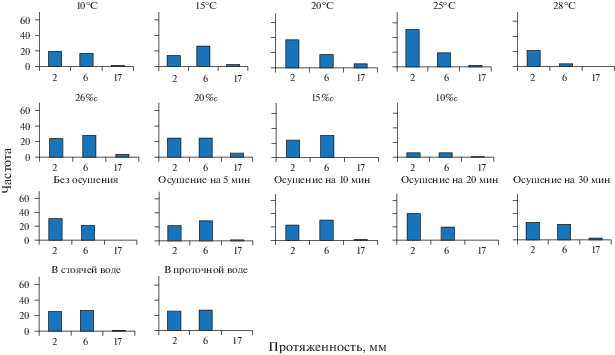
По ходу исследования было выяснено, что средние значения протяженности ГПТ недостаточны для понимания формирования магистральных течений, проходящих по столону через всю колонию. Поэтому среди всех ГПТ были выделены протяженные (более 3 мм) (рис. 2) и “объемные” (более 4 800 000 и 9 600 000 мкм3) (рис. 3) для того, чтобы определить их долю от общего числа учтенных в опыте ГПТ. Дополнительно для сравнения всех опытов использовали подразделение ГПТ по протяженности на три группы: менее 2, 2–6 и 6–17 мм (рис. 4).
Рис. 2.
Доля протяженных (более 3 мм) течений (а) и индекс эффективности интеграции IntHF (б) в первом модуле столона D. pumila при исследованных условиях среды.
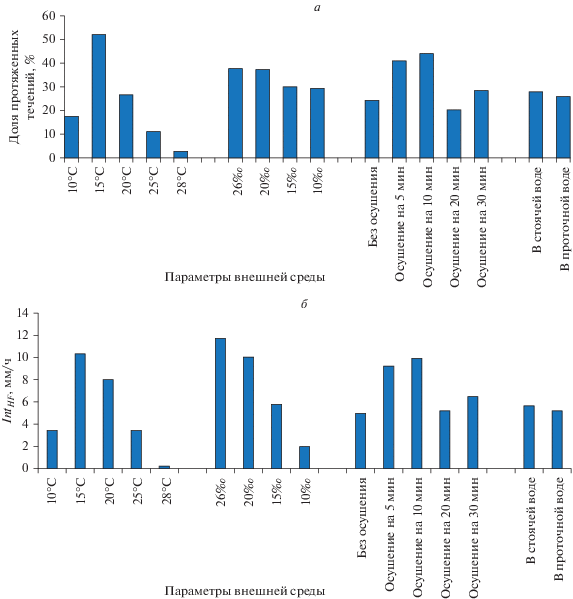
Рис. 3.
Доля течений объемом более 4 800 000 (а) и 9 600 000 (б) мкм3, поступающих в первый модуль столона D. pumila при исследованных условиях среды.
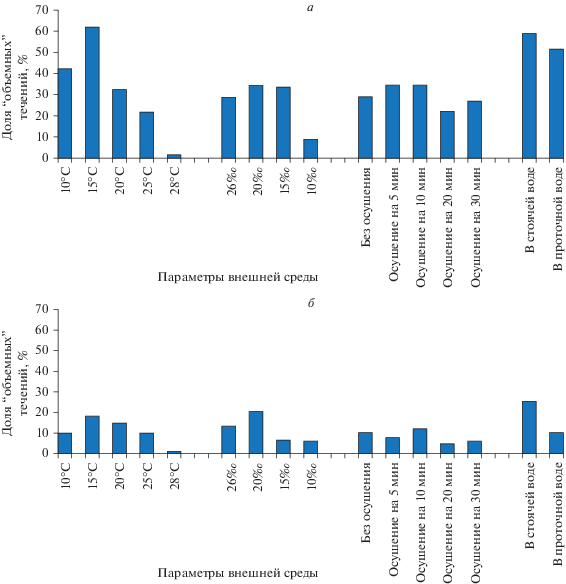
Рис. 4.
Интервальные вариационные ряды значений протяженности ГПТ в столонах D. pumila при исследованных условиях среды. Обозначения: частота – число ГПТ с данной протяженностью за 10 ч; протяженность “2” – менее 2 мм (протяженность менее длины одного модуля), “6” – 2–6 мм (один–два модуля), “17” – 6–17 мм (более двух модулей столона).
Для сравнения реакции распределительной системы на условия содержания колоний дополнительно приведены средние значения показателя прироста столона за один цикл ростовых пульсаций (mgp). Ростовые пульсации верхушки столона определены по результатам видеорегистрации при тех же условиях в тех же колониях, что и ГПТ. Подробности метода описаны ранее (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б). Съемки верхушки роста столона происходили не позже 1.5 ч после съемок ГПТ.
Для определения эффективности распределительной системы мы руководствовались моделью ее функционирования, опубликованной ранее (Марфенин, 1985, 1993; Марфенин, Дементьев, 2017).
Перемещение частиц через всю колонию возможно при последовательном подключении промежуточных побегов, каждый из которых помогает проталкивать столб гидроплазмы, когда тот окажется в непосредственной близости от побега. Чтобы каждый последующий побег “сработал”, порция гидроплазмы должна пройти участок столона, отделяющий данный побег от предыдущего. Если Z – протяженность ГПТ – больше длины одного модуля столона, то возможен дальний перенос частиц. Если же Z составляет менее величины одного модуля столона, то дальний перенос теоретически невозможен: частицы будут перемещаться вперед и назад между двумя побегами в пределах одного модуля столона. Длина модуля столона у D. pumila может варьировать в пределах от 1.5 до 6 мм, но средняя длина сформированного модуля составляет 2.98 ± 0.12 мм, т.е. около 3 мм (Марфенин, Дементьев, 2017). Еще несформированный нулевой модуль столона всегда более короткий. Эта модель приемлема и при объяснении механизма перемещения гидроплазмы в коротких колониях, состоящих всего из двух модулей столона. В тех случаях, когда ГПТ больше длины одного модуля столона, оно может достичь верхушки роста столона без участия промежуточного молодого побега. Но такой вариант, скорее, исключение из общего правила, согласно которому часть гидроплазмы при движении от материнского побега к верхушке столона все же зайдет в молодой побег.
Основываясь на описанной выше модели, можно определить объем ГПТ. Фактически объем ГПТ (VHF) представляет собой произведение протяженности (т.е. длины) течения Z на площадь поперечного сечения ценосарка, через которое это течение проходит. Поскольку величина просвета ценосарка изменяется из-за латеральных пульсаций, это следует учитывать при определении среднего значения площади его поперечного сечения. Интенсивность латеральных пульсаций ценосарка уменьшается по мере удаления от верхушки роста. Латеральные пульсации ценосарка во втором и последующих модулях столона значительно слабее, чем в первом и особенно нулевом модулях (Marfenin, Dementyev, 2017). Ранее нами было установлено на примере более крупных колоний D. pumila, что диаметр просвета ценосарка (D) в первом и последующих модулях столона составляет в среднем около 45 мкм (Марфенин, Дементьев, 2017). Площадь поперечного сечения ценосарка ((πD2)/4) в таком случае равна 1590 мкм2. Распределительная система эффективна, если протяженность локальных ГПТ покрывает расстояние между соседними побегами (3 мм или 3000 мкм) в колонии D. pumila. Перемножив протяженность “эффективного” течения на площадь сечения ценосарка столона, получаем объем “эффективного” ГПТ – 4 800 000 мкм3. Аналогичным способом можно вычислить и объем ГПТ (9 600 000 мкм3), проходящего два модуля столона (6 мм или 6000 мкм).
На основании количественных данных (табл. 1) мы высчитали доли протяженных (более 3 мм) и объемных (более 4 800 000 и 9 600 000 мкм3) от общего числа течений.
Также мы построили интервальные вариационные ряды частот встречаемости ГПТ различной протяженности в столонах D. pumila при исследованных значениях факторов среды. Для этого мы сгруппировали количественные значения протяженности в три варианты. Выбор вариант обусловлен средней длиной модуля столона D. pumila (3 мм): 1) менее 2 мм (протяженность менее длины одного модуля); 2) 2–6 мм (один–два модуля); 3) 6–17 мм (более длины двух модулей)11.
Поскольку длительность эпизодов видеосъемки различалась существенно (от 1.5 до 8 ч), объединение выборок по каждому эпизоду в пределах конкретного значения фактора среды создавало значительный разброс по времени (от 10 до 30 ч). В таком случае использование всех вариант было нецелесообразным: большие по продолжительности эпизоды обнаружили бы значительное превосходство по количеству зарегистрированных течений. Для устранения подобной статистической ошибки частоты встречаемости ГПТ для всех параметров среды были пересчитаны на постоянный отрезок времени – 10 ч.
Для оценки эффективности распределительной системы мы впервые ввели специальный индекс.
Индекс эффективности интеграции (IntHF) – произведение средней протяженности гидроплазматических течений, превышающих заданный лимит (3 мм – длина модуля столона), на число таких ГПТ в час (т.е. на частоту протяженных ГПТ).
Описанный выше метод был использован нами ранее при изучении реакции одного и того же вида (D. pumila) на изменение температуры, солености, водообмена и осушения. Результаты исследований были опубликованы в разное время по мере завершения экспериментов и обработки результатов по каждому из указанных выше параметров среды обитания, но не были сведены вместе. Только сейчас мы можем ответить на более общий вопрос – насколько характерно для D. pumila сохранение эффективности функционирования распределительной системы при резких изменениях различных параметров среды.
РЕЗУЛЬТАТЫ
Расчетная протяженность переноса частиц (показатель Z) в первом модуле столона в колониях варьирует, но в целом условия среды в исследованном диапазоне влияют незначительно на протяженность ГПТ. В тех же пределах изменения параметров среды другие жизненно важные показатели, например прирост столона за цикл ростовых пульсаций (mgp), реагируют активнее, изменяясь в 2–3 раза (табл. 1).
При контрольных значениях параметров среды (26‰, 15°С), близких к естественным в местах обитания вида в летний период, показатель Z в столоне примерно равен 3 мм, т.е. расстоянию между соседними побегами (табл. 1). Наиболее протяженные ГПТ составляют 10.6 мм к верхушке столона и 12.8 мм от верхушки к материнскому побегу (при 26‰). Протяженность перемещенной гидроплазмы в первом модуле столона при различной солености достоверно не различается (p > 0.05) (табл. 1) и оказывается устойчивой к опреснению. Даже при 10‰ усредненное значение Z(+)HF, т.е. ГПТ, направленных к верхушке столона, все еще достаточно велико, а доля протяженных Z(+)HF составляет почти 30% (рис. 2а). Усредненная наибольшая скорость за цикл пульсации, т.е. амплитуда ГПТ (А), в первом модуле столонов проявляет слабую и недостоверную тенденцию (p > 0.05) к снижению по мере опреснения (табл. 1).
В столонах величина Z существенно уменьшается (p < 0.01) при температуре выше 15°С (табл. 1). Хотя усредненная расчетная протяженность перемещения частиц к верхушке столона Z(+)HF при 10 и 20°С составляет менее 3 мм, работа распределительной системы не должна нарушаться, поскольку велика доля отдельных течений с большей протяженностью (рис. 2а), т.е. верхушки достигают не все течения, а только наиболее мощные из них. Наиболее протяженные ГПТ составляют 8.9 мм к верхушке столона (при 15°С) и 16.6 мм от верхушки к материнскому побегу (при 20°С). При дальнейшем нагревании до 25 и 28°С величина Z снижается до 1–1.5 мм, что покрывает лишь половину длины модуля столона. При этом доля отдельных протяженных ГПТ оказывается незначительной – не более 10%. Эти показатели температуры оказываются пороговыми, при их превышении перемещение гидроплазмы становится неэффективным для транспортировки частиц к верхушкам роста столона и ближайшего к нему молодого побега.
В первом модуле столонов амплитуда направленных к верхушке ГПТ (A(+)HF) достигает максимума при 15°С, а при дальнейшем нагревании воды снижается.
От длительности осушения протяженность течений (табл. 1) в изученном диапазоне зависит слабо. Амплитуда ГПТ возрастает с увеличением продолжительности осушения (табл. 1), однако достоверные различия есть лишь между крайними вариантами эксперимента (контроль и тридцатиминутное осушение). Амплитуда направленных к верхушке столона магистральных течений А(+)HF в стоячей воде выше, чем в проточной. В стоячей и проточной воде (табл. 1) протяженность переноса частиц составляет 2–2.5 мм.
Доля протяженных течений (более 3 мм) обычно составляет 10–40% от общего числа ГПТ (рис. 2а), но может быть и меньше (при 28°С), и больше (при 15°С).
Расчет индекса эффективности интеграции (IntHF) колонии посредством работы распределительной системы обнаружил следующие результаты в зависимости от показателей среды (рис. 2б):
• оптимальная температура для интеграции колонии составляет 15°С;
• опреснение приводит к линейному понижению эффективной интеграции;
• после непродолжительного осушения интеграция возрастает;
• в стоячей и проточной воде интеграция однонаправленных течений одинакова.
Изменение объемов V(+)HF в целом демонстрирует ту же динамику (табл. 1, рис. 3), что и изменение протяженностей в соответствующем эксперименте (нагревании, опреснении, осушении). Эти объемы позволяют представить, какой объем гидроплазмы должен поступать в первый модуль столона (где проводили видеорегистрацию) для эффективной транспортировки частиц по колонии. “Объемные” ГПТ составляют значительную долю (иногда до 30–40%) от общего числа течений в изученном диапазоне факторов среды (рис. 3а), хотя ГПТ более 9 600 000 мкм3 (т.е. более 6 мм) встречаются реже (рис. 3б). Исключение составляет температура 28°С, при которой “объемных” протяженных ГПТ очень мало или вообще нет. Таким образом, интенсивные протяженные ГПТ (более одного модуля столона) в колонии существуют, они не единичны, и на них основывается дальний транспорт частиц.
Рассмотрим интервальные вариационные ряды значений протяженности ГПТ в столонах D. pumila при исследованных значениях факторов среды (рис. 4), взаимно дополняющие результаты расчета IntHF .
При повышении температуры от 10 до 25°С частота ГПТ протяженностью более 2 мм изменяется незначительно, хотя частота коротких ГПТ (менее 2 мм) возрастает в 2–3 раза при 20 и 25°С. При температуре 28°С частота коротких ГПТ снижается, а протяженные ГПТ становятся редкими (рис. 4).
При опреснении с 26 до 15‰ существенных изменений частоты коротких и протяженных ГПТ не происходит, т.е. распределительная система хорошо адаптирована к опреснению и продолжает полноценно работать (по крайней мере, в коротких модельных колониях). При 10‰ частота всех ГПТ, как коротких, так и протяженных, снижается почти в 2 раза. Следовательно, эффективность функционирования распределительной системы при 10‰ существенно уменьшается (рис. 4).
Осушение длительностью до 30 мин включительно практически не влияет на частоту ГПТ. Параметр водообмена (стоячая или проточная вода) не сказывается на эффективности распределительной системы как для коротких (менее 2 мм), так и для более протяженных течений (рис. 4).
Средние значения прироста столона за один акт ростовых пульсаций (mgp), зарегистрированные примерно в то же время, что и ГПТ, оказываются более чувствительными к условиям содержания. Существенное уменьшение mgp происходит уже при 25°С, а также при 15‰ и при осушении на 20 мин. При 28°С или при 10‰ рост столона почти прекращается, причем всего лишь за несколько часов после перевода колоний в такие условия содержания (табл. 1).
ОБСУЖДЕНИЕ
Сравнительное изучение реакции распределительной системы колониальных гидроидов на внешние воздействия проводится впервые. Раньше уделяли больше внимания росту и ветвлению колонии, как показателям воздействия на колониальный организм (Бурыкин, 1978а, б, 1979) или популяционным показателям для неколониальных гидроидов (Канаев, 1952). Изменения в росте и ветвлении существующими методами обнаруживаются медленно – спустя сутки и более (Марфенин, Бурыкин, 1979). Они отражают существенную перестройку всего колониального организма, которая становится заметной не сразу и продолжается долго – порой неделю и больше, что в жизни колониальных гидроидов составляет значительный срок.
Быстрее реакция гидроидов на внешние воздействия выражается в пульсации ценосарка и в поведении гидрантов. При наступлении любых неблагоприятных обстоятельств гидранты сжимаются. Так они реагируют на осушение, резкое изменение температуры, токи воды и пр. Пульсации ценосарка также чувствительны к внешним воздействиям – особенно пульсации верхушек роста (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б). Однако и поведение гидрантов, и пульсации ценосарка в значительной мере отражают локальные обстоятельства. По ним нельзя судить об эффекте воздействий на весь организм в целом.
Эффективность функционирования распределительной системы, выраженная в скорости и объемах переносимой гидроплазмой частиц – базовый показатель жизнедеятельности колониального организма, такой же как рост и ветвление, но определяемый намного быстрее (за несколько часов). Описанные в литературе быстрые методы изучения ГПТ – путем введения в пищу радиоактивной метки (Rees et al., 1970) или флуоресцентных красителей (Макаренкова, 1988; Buss et al., 2015) – требуют наличия специального оборудования. Кроме того, эти методы недостаточно содержательны, поскольку с их помощью можно определить только генеральное направление, дистанцию и продолжительность перемещения меток без детализации процесса формирования ГПТ, его зависимости от локальных составляющих ГПТ, без детальной динамики функционирования распределительной системы.
Используя метод видеорегистрации перемещения гидроплазмы в столонах у колониальных гидроидов можно просто и бесконтактно выявлять реакцию распределительной системы на различные воздействия, которым подвергается колониальный организм. Период задержки реакции на воздействие соответствует одному–двум циклам ГПТ, т.е. от получаса до часа. Фактически это моментальная реакция организма, так как все процессы у колониальных гидроидов, кроме поведения гидрантов, происходят медленно – на протяжении десятков минут.
Используя ключевую роль распределительной системы в жизнедеятельности гидроидов и возможность быстрой регистрации ее реакции на внешние воздействия, мы можем подойти к решению ряда задач, как фундаментальных, так и прикладных. Фундаментальная задача – определение степени толерантности модульного организма, способности переживать неблагоприятные периоды, некоторой нечувствительности к стрессовым воздействиям.
Прикладная задача – выяснение продолжительности допустимой смены условий среды, при которой колониальный организм не реагирует перестраиванием своих процессов жизнедеятельности на непродолжительное осушение, изменение температуры или солености, пребывание в неподвижной воде и пр.
В этой связи рассмотрим два вопроса: 1) как система ГПТ реагирует на резкие изменения условий среды; и 2) остается ли эта система эффективной.
Анализ показателей перемещения гидроплазмы позволил выявить экологические пределы нормального существования D. pumila по нескольким параметрам. Под нормальным существованием мы подразумеваем сохранение эффективной транспортировки внутриполостной жидкости по колонии, т.е. способность доставлять частицы от самого большого побега к нулевому модулю, ближайшему к верхушке роста столона.
Способность к протяженному транспорту пищевых частиц в столоне колониального гидроида в общем случае определяется расчетной протяженностью локальных ГПТ, регистрируемых при микровидеосъемке. Из локальных ГПТ, как было показано ранее (Марфенин, Дементьев, 2017), складываются непрерывные однонаправленные интегральные ГПТ. Они распространяются на значительные расстояния – максимально на длину всего столона от его проксимальной части, где находится самый крупный (материнский) побег, из которого была выращена модельная колония, до верхушки роста столона на дистальном его конце (Марфенин, Дементьев, 2019).
Если расчетная протяженность локального ГПТ больше, чем средняя длина модуля столона, то имеются условия последовательного участия побегов вдоль по столону в перемещении гидроплазмы внутри него. Согласно этой гипотезе, локальное ГПТ частично поступает в ближайший побег, вызывая ответную реакцию выталкивания из него излишка гидроплазмы, т.е. генерируя следующее локальное ГПТ и так далее (Марфенин, 1985).
Если же расчетная протяженность локального ГПТ меньше средней длины модуля столона, то отсутствуют необходимые условия для формирования однонаправленного протяженного ГПТ. Такое течение означает всего лишь небольшой сдвиг гидроплазмы в столоне в пределах одного модуля, после чего следует его “откат” назад. Частицы пищи при этом остаются в пределах одного модуля столона. Следовательно, система ГПТ становится неэффективной при протяженности локальных ГПТ менее средней длины модуля столона. Та же система ГПТ при средних значениях протяженности локальных ГПТ больше длины модуля столона признается нами эффективной, т.е. достаточной для образования непрерывных протяженных течений.
Между двумя определенными выше состояниями системы ГПТ – эффективной и неэффективной – могут быть промежуточные состояния. Сравнивая средние значения длины модуля столона и расчетной протяженности локального ГПТ, следует помнить, что при усреднении сводится вместе все разнообразие усредняемых значений. Среди них есть как короткие, так и длинные модули (Марфенин, Дементьев, 2017), а также короткие и длинные локальные ГПТ.
Стандартная ошибка среднего значения дает некоторое представление о реальном разнообразии усредняемых величин, но не полностью. Несколько больше информации можно получить при учете доли протяженных ГПТ (более 3 мм) от общего числа течений в первом модуле столона. Этот показатель оказывается значительным при всех испытанных значениях абиотических факторов (рис. 2).
Больше всего информации мы получили из вариационных рядов, демонстрирующих соотношение между усредняемыми значениями (рис. 4); в нашем случае это соотношение протяженностей локальных ГПТ, подразделенных на три группы:
• первая группа (до 2 мм) – течения, недостаточные для генерации протяженных ГПТ;
• вторая группа (от 2 до 6 мм) – средние по длине локальные ГПТ, охватывающие один–два модуля столона, теоретически достаточные для генерации цепочки локальных ГПТ, составляющих совместно протяженное ГПТ;
• третья группа (от 6 до 17 мм) – течения, которые распространяются на два и более модулей столона.
Результаты сравнения, представленные на рис. 4, убедительно свидетельствуют о том, что главное условие для генерации протяженных ГПТ – течения второй группы – одинаково представлены почти при всех значениях изученных факторов среды, кроме 28°С и 10‰.
Следовательно, даже в тех случаях, когда средняя величина локальных ГПТ меньше средней длины модуля столона, могут быть отдельные локальные ГПТ, достаточные для преодоления расстояния между соседними побегами. В этих случаях формируются интегральные ГПТ, которые переносят частицы пищи на значительное расстояние по столону. Поскольку подобные интегральные ГПТ возникают нерегулярно, то и эффективность системы ГПТ, очевидно, меньше по сравнению с теми случаями, когда средние значения локальных ГПТ выше средних значений длины модуля столона.
При констатации неэффективности системы ГПТ следует все же обращать внимание на величину стандартной ошибки ±SE (табл. 1) и результаты анализа данных с помощью вариационных рядов (рис. 4). Чем больше величина стандартной ошибки, тем вероятнее, что система ГПТ, даже при недостаточном среднем значении расчетной протяженности локального ГПТ, время от времени генерирует течения, способные обеспечить протяженную “эстафетную”22 передачу частиц на значительное расстояние по столону.
Очевидно, что интеграция колонии определяется не только протяженными ГПТ, но и их частотой. Чем выше доля протяженных ГПТ, превышающих среднюю длину одного модуля столона (3 мм), тем эффективнее обеспечение пищей дистально расположенных зон роста. Для совместного учета двух составляющих мы ввели индекс эффективности интеграции IntHF, который представляет собой совокупную длину всех протяженных ГПТ за час (рис. 2б). Сравнивая на рис. 2 попарно ход аналогичных диаграмм, мы видим, что для опытов с температурой, осушением и гидродинамикой они почти совпадают, а в эксперименте с соленостью существенно различаются. Это означает, что по мере опреснения снижается средняя протяженность ГПТ, превышающих 3 мм. Поэтому индекс эффективности интеграции IntHF более информативен, чем показатель частоты протяженных ГПТ, и его можно рекомендовать для анализа физиологической интеграции у колониальных гидроидов.
Следуя этой рекомендации, можно учитывать промежуточные состояния эффективности распределительной системы, а не только выделять два крайних варианта: эффективная и неэффективная системы ГПТ. В таком случае представленные результаты (рис. 2б) позволяют считать, что распределительная система наиболее эффективна при 15 и 20°С, 26‰ и 20‰, осушении на 5 и 10 мин. При содержании колоний в стоячей воде эффективность такая же, как и при наличии водообмена.
Анализируя протяженность ГПТ и объемы переносимой течениями гидроплазмы, следует помнить, что все опыты были поставлены на предельно коротких колониях, состоящих из одного крупного (материнского) побега и одного маленького (дочернего). Длина столона в этих колониях была 5–6 мм. Тем более удивительно, что и в таких крошечных колониях расчетная протяженность исходящего из материнского побега ГПТ в среднем составляет 2–3 мм, а максимально 10.6 мм. Как такое возможно? Объем нулевого модуля столона – участка позади верхушки роста – значительно больше, чем объем любого другого модуля столона. Он эквивалентен трем–пяти модулям столона между побегами (Марфенин, Дементьев, 2017). Также и верхушка побега, вернее, участок столона ниже нее, значительно больше модуля ценосарка побега и модуля столона (наши неопубликованные данные). Совместно этот комплекс (нулевой модуль столона и ближайший молодой побег) способен принять значительные объемы гидроплазмы, вышедшей из материнского побега. Установленные нами максимальные значения расчетного объема одного ГПТ как раз соответствуют максимальным значениям объема нулевого модуля столона.
Аналогичных опытов на длинных колониях не проводилось. В больших колониях перисарк удаленной от верхушки части столона становится темным, что мешает регистрации ГПТ с помощью микровидеосъемки. Однако методом непосредственного визуального отслеживания движения частиц в столоне под бинокуляром было установлено, что эти частицы могут без перерыва перемещаться от одного конца колонии, состоящей из восьми модулей столона (семи сформированных и растущего нулевого), до противоположного ее конца (Марфенин, Дементьев, 2019).
Изучение воздействия факторов среды на колониальный организм гидроида D. pumila показало, что пульсации ценосарка и показатели перемещения гидроплазмы зачастую существенно варьировали (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б), в том числе в случаях, когда значения абиотических факторов были близки к естественным для колонии. Отмеченная вариабельность данных может являться следствием постоянных процессов саморегуляции в модульном организме (Марфенин, 2016). Это важное обстоятельство следует учитывать при анализе реакции распределительной системы на внешние воздействия. Сравнивая систему ГПТ в изменившихся условиях существования, необходимо считаться с тем, что и при неизменных условиях скорости ГПТ, их периодичность и дальность варьируют значительно. Это характерная особенность децентрализованных организмов (Марфенин, 2016).
Проводя данный сравнительный анализ, мы основываемся на первичных результатах осуществленных ранее частных исследований реакции D. pumila на нагревание, опреснение, осушение и изменение водообмена (табл. 1). Характерная вариабельность показателей ГПТ сглаживается наличием трендов, описанных нами ранее (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б).
Оптимальный диапазон, при котором зарегистрированы интенсивные перемещения гидроплазмы и большие объемы перенесенной гидроплазмы, составляет от 10 до 20°С и от 26 до 15‰. При дальнейшем нагревании (25 и 28°С) и опреснении (10‰) перемещения гидроплазмы теряют активность, становясь менее быстрыми, менее протяженными и менее объемными (табл. 1). Осушение на непродолжительный период (до 30 мин включительно) явным образом не сказывается на работе распределительной системы, хотя и приводит к замедлению роста столонов (Дементьев, Марфенин, 2019а). В стоячей воде распределительная система D. pumila функционирует более энергично, а именно: с более высокой частотой возникновения мощных ГПТ, бóльшей их скоростью и бóльшими объемами переносимой гидроплазмы (Марфенин, Дементьев, 2018б).
Только при достижении температуры 25–28°С наблюдается снижение эффективности функционирования распределительной системы, но даже при столь высокой для гидроида температуре протяженные ГПТ не исчезают полностью (хотя становятся редкими). В остальных случаях распределительная система остается эффективной, т.е. способной обеспечить перенос частиц к зонам роста, несмотря на уменьшение прироста и даже остановку роста колонии (Дементьев, Марфенин, 2018, 2019а, б).
На изменение условий среды показатели ГПТ реагируют “мягко”. При пошаговом изменении температуры, солености, постепенном возрастании продолжительности осушения и отсутствии водообмена ожидаемо было бы увидеть линейную реакцию (например, плавное снижение всех показателей). Однако этого не происходит (рис. 4), за исключением опыта с опреснением (рис. 2б). Изменение характера ГПТ при каждом новом воздействии на отдельную колонию в исследуемом диапазоне факторов среды практически незаметно. По анализу ГПТ мы не можем установить, что в данный конкретный момент система почувствовала смену условий среды. Реакция сразу после произведенного изменения параметра не заметна. Показатели течений меняются не быстро и не сильно (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б). Лишь при усреднении множества данных по многим колониям можно выделить некоторые тренды, но и они, впрочем, не вполне отчетливые.
Вблизи экстремальных значений параметров среды гидроплазматические течения продолжаются (рис. 4), хотя другие физиологические показатели нарушаются. Рост снижается вплоть до остановки (mgp в табл. 1); более того, у ростовых и латеральных пульсаций тела изменяются период и амплитуда (Дементьев, Марфенин, 2018, 2019а, б; Марфенин, Дементьев, 2018б).
Сохранение гидроплазматических течений свидетельствует о том, что распределительная система продолжает функционировать даже в условиях стресса.
Являясь характерными организмами, обитающими на границе нижней литорали и сублиторали, исследованные нами колонии гидроида D. pumila подвергаются сильному абиотическому воздействию. До сих пор оставалось совершенно не ясно, вызывает ли абиотический стресс какие-либо нарушения в работе распределительной системы, а если и вызывает, то насколько сильные. Анализ параметров функционирования распределительной системы позволил показать сколь устойчивыми могут быть гидроплазматические течения при внешних воздействиях (температурных, соленостных и т.п.), близких к естественным в местах обитания вида в летний период.
Это служит подтверждением лабильности децентрализованной распределительной системы. В модульной организации, присущей гидроидам, становятся возможными нецентрализованные регуляции, обусловливающие более высокую устойчивость системы за счет развития пластичности физиологических процессов. Гидроиды способны адаптироваться к особенностям среды обитания благодаря своей “природной” вариабельности (Марфенин, 2002, 2016). Относительно просто устроенный организм гидроида демонстрирует пластичность ответа на внешнее воздействие: совершенно разные факторы среды в исследованном диапазоне не могут дестабилизировать систему. Физиологический процесс переноса пищи по колониальному организму существенно не нарушается даже при отклонении условий от оптимальных для гидроида. Хотя D. pumila и является литоральной формой, обитающей в нестабильных условиях приливно-отливной полосы, все же можно выделить наиболее оптимальные для нее показатели абиотических факторов. Использованные в экспериментах значения факторов среды (10–28°С, 10–26‰) соответствуют температурным и соленостным режимам на литорали беломорского побережья в летний период (Наумов, Оленев, 1981; Цетлин и др., 2010). Однако оптимальный диапазон, при котором наблюдаются высокая скорость роста, интенсивные перемещения гидроплазмы и наибольший объем перенесенной гидроплазмы, составляет от 10 до 20°С. Гидроид D. pumila способен существовать без проявления признаков угнетения в диапазоне солености от 26 до 20‰ в течение по крайней мере 8 ч (Дементьев, Марфенин, 2018, 2019а, б). Колонии выдерживают осушение до 30 мин включительно: происходит замедление прироста, но значимые изменения в пульсациях тела и перемещении гидроплазмы отсутствуют даже спустя 7–9 ч наблюдений.
Установленная толерантность распределительной системы D. pumila к абиотическим воздействиям имеет практическое значение. Она позволяет использовать такие процедуры при экспериментальной работе с гидроидами, как: 1) перенос стекол с колониями из одной кюветы в другую; 2) манипуляции с гидроидами в непроточной кювете со стоячей водой; 3) кормление гидроидов в отдельном аквариуме при температуре, отличной от температуры культивирования; 4) вынужденное содержание в воде с разной соленостью (взятой из моря и не стандартизированной) и т.п. Полученные данные позволяют снять многие вопросы о допустимости проведения тех или иных процедур при лабораторных исследованиях гидроидов – важных модельных объектов (по крайней мере, часто используемого вида D. pumila).
Мы рассмотрели эффективность интеграции колонии не только на предмет ее наличия или отсутствия, но и используя критерий частоты интеграционных протяженных ГПТ. Формирование локальных течений, достаточных для эффективного перемещения гидроплазмы по колонии, возможно при всех исследованных условиях среды, хотя и с разной частотой. Исследованные показатели среды не приводят к полному прекращению транспортировки гидроплазмы по колонии (даже при 10‰ в столонах сохраняются ГПТ, а при 28°С встречаются течения протяженностью более 3 мм). Распределительная система остается теоретически эффективной при любых испытанных значениях температуры, солености, длительности осушения и отсутствии водообмена. Установленная стойкость распределительной системы (в первые часы воздействия фактора) – доказательство толерантности гидроида к отклонению показателей среды от оптимальных для организма значений. По-видимому, толерантность является характерной особенностью распределительной системы литорального гидроида D. pumila.
ВЫВОДЫ
Сопоставление результатов исследований реакции гидроида D. pumila на изменения условий среды показало, что распределительная система этого вида слабо реагирует в первые часы резкого отклонения от оптимума по нескольким главным факторам среды, а именно: температуре, солености, осушению и водообмену.
Распределительная система D. pumila остается теоретически эффективной в диапазоне температуры от 10 до 28°С, опреснении с 26 до 10‰, осушении длительностью до 30 мин и отсутствии водообмена. Однако оптимальными условиями функционирования системы ГПТ являются 15–20°С, 26–20‰, а также кратковременное осушение на 5–10 мин. Различия в эффективности функционирования распределительной системы при содержании колоний в стоячей воде и на течении не установлены.
Исследование выполнено в рамках научного проекта государственного задания МГУ имени М.В. Ломоносова № 121032300118-0.
Список литературы
Бурыкин Ю.Б., 1978а. Рост колонии Dynamena pumila (L.) (Hydrozoa, Sertulariidae) при различной солености воды // Деп. ВИНИТИ. № 2426-78. Деп. от 14.07.1978. С. 1–7.
Бурыкин Ю.Б., 1978б. Рост и половое созревание колонии Dynamena pumila (L.) (Hydrozoa, Sertulariidae) при различной температуре воды // Деп. ВИНИТИ. № 2427-78. Деп. от 14.07.1978. С. 1–10.
Бурыкин Ю.Б., 1979. Регулирующая роль некоторых экологических факторов в процессе роста колониальных гидроидов. Дис. … канд. биол. наук. М.: МГУ. 168 с.
Бурыкин Ю.Б., 2013. Эстафетный способ перемещения гидроплазмы в колониях гидроидных полипов // Онтогенез. Т. 44. № 2. С. 115–125.
Дементьев В.С., Марфенин Н.Н., 2018. Влияние опреснения на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 79. № 5. С. 376–392.
Дементьев В.С., Марфенин Н.Н., 2019а. Влияние факторов среды на рост и функционирование распределительной системы гидроида Dynamena pumila (Linnaeus, 1758) // Тр. VII Междунар. науч.-практ. конф. “Морские исследования и образование (MARESEDU-2018)”. Т. IV. Тверь: ООО “ПолиПРЕСС”. С. 282–284.
Дементьев В.С., Марфенин Н.Н., 2019б. Воздействие температуры на рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 80. № 1. С. 22–42.
Канаев И.И., 1952. Гидра: очерки по биологии пресноводных полипов. М.; Л.: Изд-во АН СССР. 372 с.
Косевич И.А., 1991. Сравнение функционирования верхушек роста побегов и столонов в колонии Obelia loveni (Allm.) (Hydrozoa, Campanulariidae) // Вестн. МГУ. Сер. 16. Биол. № 2. С. 44–52.
Макаренкова Е.П., 1988. Определение скорости движения гидроплазмы у колониальных гидроидов рода Obelia с помощью флуоресцентных красителей // ДАН СССР. Т. 302. № 5. С. 1275–1277.
Марфенин Н.Н., 1985. Функционирование распределительной системы пульсаторно-перистальтического типа у колониальных гидроидов // Журн. общ. биологии. Т. 46. № 2. С. 153–164.
Марфенин Н.Н., 1993. Функциональная морфология колониальных гидроидов. СПб.: ЗИН РАН. 151 с.
Марфенин Н.Н., 2002. Нецентрализованная саморегуляция целостности колониальных организмов // Журн. общ. биологии. Т. 63. № 1. С. 26–39.
Марфенин Н.Н., 2016. Децентрализованный организм на примере колониальных гидроидов // Биосфера. Т. 8. № 3. С. 315–337.
Марфенин Н.Н., Бурыкин Ю.Б., 1979. Зависимость роста колонии Dynamena pumila (L.) (Hydrozoa, Sertulariidae) от количества получаемой пищи // Вестн. МГУ. Сер. 16. Биол. № 1. С. 61–68.
Марфенин Н.Н., Дементьев В.С., 2017. Парадокс протяженных течений гидроплазмы в колониальном гидроиде Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 78. № 4. С. 3–20.
Марфенин Н.Н., Дементьев В.С., 2018а. Продольные пульсации столона у колониального гидроида Dynamena pumila (Linnaeus, 1758) // Журн. общ. биологии. Т. 79. № 2. С. 85–96.
Марфенин Н.Н., Дементьев В.С., 2018б. Рост, пульсации ценосарка и перемещение гидроплазмы у колониального гидроида Dynamena pumila (L., 1758) в проточной и непроточной кюветах // Журн. общ. биологии. Т. 79. № 2. С. 97–107.
Марфенин Н.Н., Дементьев В.С., 2019. К вопросу о протяженности гидроплазматических течений у колониального гидроида Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 80. № 5. С. 348–363.
Марфенин Н.Н., Дементьев В.С., 2020. Побеги как генераторы гидроплазматических течений в колониальном гидроиде Dynamena pumila (L., 1758) // Журн. общ. биологии. Т. 81. № 6. С. 421–443.
Наумов А.Д., Оленев А.В., 1981. Зоологические экскурсии на Белом море: пособие для летней учебной практики по зоологии беспозвоночных. Л.: Изд-во ЛГУ. 175 с.
Цетлин А.Б., Жадан А.Э., Марфенин Н.Н., 2010. Флора и фауна Белого моря. Иллюстрированный атлас. М.: Т-во науч. изд. КМК. 471 с.
Buss L.W., Anderson C.P., Perry E.K., Buss E.D., Bolton E.W., 2015. Nutrient distribution and absorption in the colonial hydroid Podocoryna carnea is sequentially diffusive and directional // PLoS One. V. 10. № 9. Art. e0136814.
Dudgeon S.R., Buss L.W., 1996. Growing with the flow: on the maintenance and malleability of colony form in the hydroid Hydractinia // Am. Nat. V. 147. № 5. P. 667–691.
Dudgeon S.R., Wagner A., Vaisnys J.R., Buss J.W., 1999. Dynamics of gastrovascular circulation in the hydrozoan Podocoryne carnea: The one-polyp case // Biol. Bull. V. 196. P. 1–17.
Marfenin N.N., Dementyev V.S., 2017. Functional morphology of hydrozoan stolons: Stolonal growth, contractility, and hydroplasmic movement in Gonothyraea loveni (Allman, 1859) // Mar. Biol. Res. V. 13. № 5. P. 521–537.
Rees J., Davis L.V., Lenhoff H.M., 1970. Paths and rates of food distribution in the colonial hydroid Pennaria // Comp. Biochem. Physiol. V. 34. P. 309–316.
Wagner A., Dudgeon S., Vaisnys J.R., Buss L.W., 1998. Nonlinear oscillations in polyps of the colonial hydroid Podocoryne carnea // Naturwissenschaften. V. 85. P. 117–120.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии