

Журнал общей биологии, 2021, T. 82, № 4, стр. 297-317
Великая отечественная война как фактор флорогенеза: результаты поиска полемохоров в центральной России
Н. М. Решетникова 1, *, А. А. Нотов 2, С. Р. Майоров 3, А. В. Щербаков 3
1 Главный ботанический сад им. Н.В. Цицина РАН
127276 Москва, ул. Ботаническая, 4, Россия
2 Тверской государственный университет
170100 Тверь, ул. Желябова, 33, Россия
3 Московский государственный университет им. М.В. Ломоносова, биологический факультет
119234 Москва, Ленинские горы, 1, стр. 12, Россия
* E-mail: n.m.reshet@yandex.ru
Поступила в редакцию 08.02.2021
После доработки 17.03.2021
Принята к публикации 26.04.2021
Аннотация
На территории Центральной России в местах дислокации немецких войск выявлено более 45 видов растений, занесенных во время военных действий (видов-полемохоров). Специфика их распространения в регионах Центральной России обусловлена взаимным влиянием военно-исторических и природных факторов. Исходный уровень разнообразия полемохоров определялся количеством занесенных семян, которое зависело от продолжительности оккупации, специфики боевых действий. Наибольшее число видов-полемохоров встречается вблизи станций перевалки грузов с железных дорог на шоссейные и грунтовые, а также участков в местах расположения немецких складов. Основным источником заноса семян полемохоров был фураж для лошадей. Кроме того, на некоторых участках для маскировки позиций, по-видимому, использовали травосмеси. Большое значение для сохранения полемохоров имели тип растительных сообществ, уровень трансформации ландшафтов, характер хозяйственного использования территории и специфика сукцессионной динамики растительного покрова. Полемохоры устойчивы на лугах, полянах и опушках в светлых лесах, не затронутых интенсивной хозяйственной деятельностью и мало трансформированных в ходе сукцессионных смен. Во вторичных лесных сообществах, образовавшихся на месте луговых и рудеральных фитоценозов, сохраняются прежде всего виды с более широкой эколого-фитоценотической амплитудой. Подавляющее большинство полемохоров не проявляет тенденции к дальнейшему распространению, но некоторые виды могут расселяться за пределы исходных местонахождений. На территориях, связанных с ключевыми стратегическими объектами военного времени (например, Ржевско-Вяземский плацдарм), ценотическая роль центральноевропейских видов может быть значительной, а сформировавшиеся фитоценозы могут проявлять сходство со среднеевропейскими луговыми сообществами. Выявлены случаи гибридизации полемохоров с аборигенными видами. Полученные результаты свидетельствует о том, что Великая Отечественная война стала существенным фактором современного генезиса флоры Центральной России.
Прошедшие войны были связаны не только с перемещением огромного количества людей, военной техники, различных грузов, снаряжения, но и лошадей, необходимого для них фуража. Все это неизбежно способствовало масштабному переносу на значительные расстояния диаспор чужеродных растений. Несмотря на очевидность влияния этих процессов на экосистемы разных регионов, роль мировых войн в генезисе современной флоры изучена недостаточно. Хотя в литературе неоднократно приводили примеры заноса растений в результате военных действий и пытались оценить воздействие Второй мировой войны на флору Скандинавских стран (Berg, 1962; Niemi, 1969; Ahti, Hämet-Ahti, 1971; Alm, 2015; Wearn, 2016, и др.), в России этой проблеме не уделяли внимания. Лишь сравнительно недавно А.Н. Сенниковым (2012) высказана гипотеза о связи местонахождений центральноевропейских видов на территории Северо-Западной России с военной историей. К этим видам он применил термин “полемохоры”. В Центральной России осуществленное нами исследование полемохоров проведено впервые.
Цель статьи – обобщить результаты специального поиска полемохоров, проведенного спустя 75 лет в местах дислокации немецких войск во время Великой Отечественной войны.
Задачи: 1) выявить особенности распространения полемохоров в регионах Центральной России; 2) выяснить обусловленность их встречаемости военно-историческими и другими факторами.
ИСТОРИЯ ИЗУЧЕНИЯ ПОЛЕМОХОРОВ
Военные действия как фактор миграции растений. По-видимому, впервые на занос чужеродных растений во время войны как на особое явление было обращено внимание после осады Парижа в 1870–1871 гг. во время франко-прусской войны. В Париже и его окрестностях зафиксировано 190 заносных видов, преимущественно средиземноморского происхождения (Gaudefroy, Mouillefarine, 1871). После проведения дополнительных исследований список был расширен до 268 видов (Gaudefroy, Mouillefarine, 1872). Для этой совокупности растений предложен термин “Florula obsidionalis”, т.е. флора осады (Gaudefroy, Mouillefarine, 1871). На позициях прусской армии было отмечено только три вида, a бóльшая часть растений попала с фуражом для французской кавалерии (Gaudefroy, Mouillefarine, 1871, 1872). K 1874 г. почти все занесенные растения исчезли, уцелело только три вида (Bureau, 1874). Похожий по составу занос чужеродных видов из Средиземноморья с французской кавалерией произошел ранее в Пьемонте (Gras, 1861). Отдельные случаи появления чужеродных видов с войсками регистрировали неоднократно. Так, в окрестностях Парижа обнаружена Bunias orientalis L., занесенная “русскими казаками” во время войны с Наполеоном (Gaudefroy, Mouillefarine, 1871; Thellung, 1917). В России на распространение чужеродных растений во время революции и Гражданской войны обратил внимание М.И. Назаров (1927, с. 223), связав это с “перемещением больших человеческих масс, а с ними груза, фуража и пр.” и, кроме того, со “скученностью населения и трудностью жизни”, которые приводят к “падению культурных привычек”.
Распространение чужеродных растений в период военных действий Теллунг предложил обозначать термином “стратиоботаника” (от греч. στρατιά – армия) (Thellung, 1917). Термин “полемоботаника” (от греч. πολεμος – война) был им отвергнут из-за сходства со словом “полемика”. Тем не менее термин “polemochores” для обозначения растений, занесенных во время военных действий, получил широкое распространение (Mannerkorpi, 1944). Скандинавские ботаники уделили большое внимание полемохорам, попавшим на территорию Норвегии, Финляндии, Швеции во время Второй мировой войны (Berg, 1962; Niemi, 1969; Ahti, Hämet-Ahti, 1971; Sætra, 1972; Uotila, Suominen, 1976; Alm et al., 1989, 2000a, b, c, 2001a, b, 2003, 2009; Bertelsen, 1994; Often, Alm, 1997; Piirainen, 1997a, b, 2001, 2006; Piirainen et al., 1998; Piirainen, Alm, 1998, 2001; Alm, Piirainen, 2000, 2001; Often, 2000; Alm, 2015; Wearn, 2016; Piirainen, Chkalov, 2018).
Изучение полемохоров в России. “В СССР полемохоры не привлекли внимания ботаников, несмотря на длительность оккупации обширной части страны и культовый статус воспоминаний о войне” (Сенников, 2012, с. 183). Отдельные случаи произрастания центральноевропейских видов в местах расположения войск все же были отмечены. Например, А.К. Скворцов (2005, с. 65) предположил, что Poa chaixii у д. Кобелево Смоленской области “вероятно, был занесен во время войны (здесь вообще много и других следов войны)”. Смоленский историк В.Г. Меринков (2003) считал, что найденные им в Красном Бору у Смоленска Geranium phaeum и Primula elatior растут на месте немецких складов и могли быть занесены во время войны. Тем не менее специального сбора и обобщения данных о полемохорах в течение 70 лет никто не осуществил.
Внимание к феномену полемохоров на территории России привлек А.Н. Сенников (2012). Первоначальный список таких видов был предложен им на основе изучения работ финских ботаников и критического анализа флоры северо-запада Европейской России и включал восемь видов (Сенников, 2012). Места скопления этих центральноевропейских видов в Ленинградской области были описаны ранее и стали называться “чудо-полянами”. Их флора была хорошо изучена, на основе чего было высказано предположение о реликтовом характере этих местообитаний (Хааре, 1979), поэтому часть видов предложили к охране. Однако А.Н. Сенников заметил, что все находки приурочены к прифронтовому району, расположенному непосредственно за пределами кольца блокады Ленинграда. По его мнению, среднеевропейский естественный ареал растений “чудо-полян” указывает на источник их заноса, а специфика местообитаний (большинство видов было обнаружено на луговых обочинах и откосах железных дорог, около деревень, станций, перекрестков и т.д.) – на характер заноса. Эти растения могли попасть на оккупированную территорию с сеном для германской армии. Фураж (овес и сено) привозился сюда в огромном количестве. По понятным причинам летом 1941 г. местного сена было заготовлено мало. Уже к середине зим 1941–1942 и 1942–1943 гг. местные запасы фуража, не уничтоженные в результате военных действий населением или партизанами, оказались исчерпанными, и требовался его подвоз из Европы. В частности, в проекте плана операции “Цитадель” от 12.04.1943 указано, что в 9-ю немецкую армию, развернутую в лесостепных районах южнее Орла, требуется подвезти “7000 рег. т [1 регистровая тонна = 2.8 м3] зернового и 12 000 рег. т сухого фуража” (Огненная дуга, 2003, с. 107), т.е. более 50 000 м3.
Необходимость ввоза значительного объема фуража была обусловлена активным использованием лошадей войсками Вермахта. Во время Второй мировой войны кавалерия еще существовала как отдельный род войск, значительная часть артиллерии имела конную тягу, а дороги в лесных областях Восточной Европы бóльшую часть года были непроезжими для автотранспорта. Пехота составляла более двух третей немецкой армии. В пехотной дивизии по штату полагалось 5375 лошадей (Buchner, 2001), а каждая лошадь ежедневно потребляла более 10 кг фуража (Fowler, 2001). По плану Барбаросса на восточном фронте в войсках Германии на июнь 1941 г. в кавалерийских частях было 625 000 лошадей, а в 1942 г. в Вермахте насчитывалось уже 1 250 000 лошадей (Jarymowycz, 2008; Stahel, 2009).
СЛОЖНОСТЬ ВЫЯВЛЕНИЯ ПОЛЕМОХОРОВ
Поиск спустя 75 лет с момента заноса. Со времени окончания Великой Отечественной войны прошло уже три четверти века, и бóльшая часть полемохоров по тем или иным причинам могла исчезнуть (или расселиться) из мест заноса. С одной стороны, это существенно осложняет выявление причинно-следственных связей. С другой стороны, повышается актуальность анализа факторов, которые способствуют продолжительному сохранению чужеродных видов и возможных причин такого запоздалого интереса к исследованию полемохоров.
Полемохоры часто попадали в малонарушенные местообитания, в которых они способны очень долго сохраняться, если экотопы и далее не подвергаются существенной антропогенной трансформации. Это обстоятельство может объяснять отсутствие данных об этой группе в работах, посвященных специальному поиску чужеродных видов. Однако экотопы, в которых встречаются полемохоры, не являются компонентами уникальных природных комплексов с редкими аборигенными видами растений. В этой связи специалисты по аборигенной флоре, как правило, также не уделяют специального внимания местообитаниям с полемохорами. Такое невольное “выпадение” экосистем с полемохорами из поля зрения флористов в некоторой степени может объяснить, почему в течение последних 70 лет эта группа видов так и не была изучена. Даже неоднократные находки чужеродных центральноевропейских видов в Северо-Западной России не стали поводом для их детального анализа. Укрепившееся представление о реликтовом статусе этих местообитаний не позволило поставить задачу специального поиска полемохоров в местонахождениях, связанных с военной историей.
Возможность пропуска полемохоров при инвентаризации флоры, которая не ориентирована на их целенаправленный поиск, мы смогли оценить и в ходе собственных исследований. Например, в 2010 г. при выполнении работ по проектированию памятника природы “Городской бор г. Жиздры” Н.М. Решетниковой и А.В. Крыловым основное внимание уделялось наиболее хорошо сохранившимся природным участкам. Полемохоры, приуроченные к другой части бора, не были обнаружены. Однако в 2019–2020 гг. при обследовании окрестностей бывшей железнодорожной станции Жиздра мы нашли 18 чужеродных видов из этой группы.
Проблема установления полемохорного статуса. Наиболее существенным признаком полемохоров можно считать занос из других регионов во время войны. Однако следует учитывать, что естественная граница распространения некоторых видов, в отношении которых высказано предположение о полемохорном статусе находок, подходит достаточно близко к изучаемым регионам. В связи с этим нельзя исключить вероятность аборигенного статуса части популяций преимущественно центральноевропейских видов в Северо-Западной России или в граничащих с ней районах Центральной России (Нотов и др., 2018). В отношении видов с диффузной и/или подвижной восточной границей ареала вопрос о полемохорном или аборигенном статусе решается еще сложнее. К северо-западной части Центральной России близко подходят северо-восточные границы ареалов центральноевропейских видов, а некоторые виды с более широким долготным распространением на севере могут иметь аборигенный статус. Например, в Тверской области аборигенными видами являются Deschampsia flexuosa, C-arex pilulifera, микровиды Hieracium vulgatum s. l. (Нотов, 1994; Сенников, 2006). С юга-запада по территории Брянской области проходят границы ареалов Carex brizoides и Cruciata glabra.
Занос центральноевропейских видов на территорию Центральной России мог осуществляться разными способами и в разное время. Вид может иметь полемохорное происхождение в одних местонахождениях и иные источники заноса – в других. В этой связи вопрос об установлении или исключении полемохорного статуса должен рассматриваться отдельно, применительно к каждому местонахождению и к каждой популяции вида (Нотов и др., 2018, 2020а; Нотов А., Нотов В., 2019, 2020б). Существенную роль в появлении центральноевропейских видов играли усадебные парки. Такие примеры рассмотрены в работах по Тверской области (Нотов и др., 2018). Состав полемохоров Центральной России достаточно сходен с составом основных интродуцированных видов, которые культивировали в дворянских усадьбах. Это, очевидно, обусловлено общим источником посевного материала из Центральной Европы. Некоторые полемохорные растения нередко трактовались ранее как “беглецы” из усадебных парков (Нотов А., Нотов В., 2020б). В этой связи регионы с обилием старых дворянских усадеб представляют большие трудности для изучения полемохоров. Значительное число старинных парков на территории Тверской, Калужской и Московской областей существенно осложняет дифференциацию полемохоров, особенно в случаях, когда старые усадьбы активно использовались немцами. В них нередко организовывали госпитали и штабы, территории парков пересекали стратегически значимые дороги и окопы (Нотов и др., 2018; Нотов А., Нотов В., 2020б).
Очевидно, что вместе с фуражом в оккупированные районы попадали диаспоры не только центральноевропейских растений, но и более широко распространенных видов, в том числе и встречающихся в России. Итогом такого заноса становились полемохорные популяции некоторых видов, которые представлены в региональной флоре (Нотов А., Нотов В., 2019, 2020б; Нотов и др., 2020а). Однако в настоящее время более или менее определенно установить полемохорный статус можно только для видов с центральноевропейскими ареалами и в редких случаях – для отдельных видов с более широким географическим распространением.
МАТЕРИАЛЫ И МЕТОДЫ
Начальный этап выявления полемохоров. При обобщении материалов по находкам центральноевропейских видов в Центральной России нами предложены критерии отнесения к полемохорам (Щербаков и др., 2013). Основными можно считать следующие критерии: 1) вид обнаружен только в пределах границ оккупированных территорий и только в послевоенное время; 2) местонахождение значительно удалено от основного ареала, и на изучаемой территории вид не встречается во многих других подходящих для него местообитаниях; 3) находки приурочены к природным или малонарушенным местообитаниям, в которые чужеродные растения, как правило, не внедряются; 4) в одном местонахождении обнаруживается сразу несколько видов, соответствующих отмеченным выше критериям.
С учетом этих критериев выявлены виды, которые с большей или меньшей вероятностью могли быть занесены в Центральную Россию войсками немецкой армии во время Великой Отечественной войны. На первых этапах исследований к полемохорам отнесено восемь видов (Щербаков и др., 2013). Для многих видов обнаруженные местонахождения, расположенные в отрыве от основного ареала, ранее считались реликтовыми. В некоторых регионах эти виды были включены в Красные книги или предложены к охране: Holcus mollis (предложен к охране в Калужской области – предварительные списки для издания в 2015 г.), Carex brizoides, Phyteuma spicatum (Красная книга…, 2006), Primula elatior и P. vulgaris (Перечень…, 2012). К полемохорам мы также отнесли Luzula luzuloides, Heracleum sphondylium, Pimpinella major.
В 2014 г. Н.М. Решетниковой при специальных поисках Phyteuma spicatum в рамках мониторинговых исследований по Красной книге Калужской области на реконструированной немцами дороге было отмечено 10 центральноевропейских видов (Решетникова, 2015а). На линии немецких укреплений по рекам Пополта и Перекша на четырех участках был найден охраняемый в регионе центральноевропейский вид Festuca filiformis, известный ранее в Центральной России из трех точек (Алексеев, 1985; Решетникова, 2016). Эти находки подтвердили результативность специального поиска полемохоров даже спустя 70 лет с момента окончания войны.
Полевые работы 2018–2020 гг. Нами составлен алгоритм поиска территорий, перспективных для обнаружения полемохорных видов (Щербаков, Решетникова, 2017). На первом этапе определяли возможные места их заноса. Для этого использовали сведения, полученные при работе в гербариях (известные ранее находки отдельных видов), данные из научной, мемуарной и художественной литературы, анализировали военные карты, учитывали материалы и консультации руководителей поисковых отрядов. Особое внимание уделено территориям, где ранее уже находили центрально- и западноевропейские виды, а также районам продолжительной оккупации, окрестностям военных складов, станциям и старым дорогам, которые активно использовались противником во время войны.
Наиболее детальные исследования проведены в Тверской области, где охвачены значительные по площади ключевые территории бывшего Ржевско-Вяземского плацдарма (Зубцовский, Ржевский, Оленинский районы области). Здесь закартированы все местонахождения полемохоров, выявлены основные типы фитоценозов с их участием (Нотов и др., 2019а, б, 2020б, в, г). Регулярное посещение модельных территорий в течение всего вегетационного периода в Тверской области позволило выявить сезонную динамику сообществ и обнаружить эфемероидную фракцию полемохоров (Нотов и др., 2020а, б). Подробные поиски проведены в Смоленской и Калужской областях, в нескольких районах вдоль линий фронта разных лет. Для каждого изученного участка составлен полный флористический список. Параллельно, но менее интенсивно, изучали полемохоры в Орловской и Брянской областях.
Все встреченные виды, которые мы относили к растениям-полемохорам, а также необычные формы, нуждающиеся в дополнительном изучении, собраны в гербарий. Образцы переданы в Гербарий им. А.К. Скворцова Главного ботанического сада РАН (МНА), дублеты – в Гербарий им. Д.П. Сырейщикова Московского государственного университета (MW).
Принципы отбора видов для анализа. В ходе проведенных исследований собран большой и разноплановый материал по видам, которые в изученных областях в разной степени связаны с пунктами концентрации центральноевропейских растений.
В данной статье при обсуждении результатов рассматриваются только виды и формы, характеризующиеся следующими особенностями: 1) имеют преимущественно центральноевропейский ареал; 2) отмечены в местах дислокации немецких войск в большей части местонахождений или хотя бы в одной из изученных областей; 3) некоторые активно расширяющие свое распространение виды, появление которых в Центральной России могло быть связано с различными процессами (интродукция в дворянских усадьбах и на современных объектах, естественное расширение западной границы ареала, полемохорный занос), если они регулярно встречаются на военных объектах (хотя бы в одной из областей).
Не рассмотрены при анализе результатов следующие группы: 1) виды, которые хотя и встречаются в местах военных действий, но часто приурочены к другим местообитаниям; 2) сложно идентифицируемые виды и формы, распространение которых еще мало изучено; 3) представители агамных комплексов и агрегатов микровидов, нуждающиеся в критическом изучении специалистами.
К первой группе мы относим, например, часто культивируемые и натурализирующиеся в России центральноевропейские виды Narcissus poёticus L., Aquilegia vulgaris L., Bellis perennis L. Широко распространились в Средней России Sieglingia decumbens (L.) Bernh., Poa supina Schrad., Cardamine pratensis L. s. str. и др., их расселение нуждается в дополнительном изучении.
Вторую группу представляют выявленные при полевых работах необычные и нехарактерные формы или малоизученные в Восточной Европе виды. Дальнейшее изучение (в том числе, возможно, и молекулярными методами) поможет установить статус этих находок. При анализе распространения мы не рассматриваем дерновинную форму Helictotrichon pubescens (Huds.) Schult. et Schult. fil. (Решетникова, Майоров, 2020), формы с коротким соцветием у Phleum (возможно, Ph. hubbardii Kovats.) и Alopecurus pratensis L., форма с опушенными влагалищами у Calamagrostis arundinacea (L.) Roth, а также Hylotelephium maximum (L.) Holub, Fragaria semperflorenses Duch., Veronica vindobonensis (M.A. Fisch.) M.A. Fisch., Euphorbia esula L. и др.
К третьей группе относятся малоизученные виды узколистных овсяниц (Festuca duvalii (St.-Yves) Stohr, F. nigrescens Lam. и др.), представители родов Alchemilla, Hieracium, Ranunculus, Taraxacum.
РЕЗУЛЬТАТЫ
Общие итоги и новые данные по регионам. Мы считаем, что полемохорное (или полемохорное в отдельных пунктах) происхождение имеют более 45 видов. Общий список видов-полемохоров Центральной России приведен в табл. 1. В ней обобщена информация о числе находок в изученных областях, годе первой регистрации на их территории, типах местообитаний и связанных с ними военных объектах. Отмечено также общее распространение на территории Средней и Северо-Западной России.
Таблица 1.
Особенности распространения полемохоров в регионах Центральной России
| Вид | Встречаемость в областях |
Типы местообитаний |
Типы военных объектов |
Распространение в Средней (СР) и Северо-Западной России (СЗР) |
|---|---|---|---|---|
| 1. Anthoxanthum amarum Brot. (A. odoratum L. subsp. amarum (Brot.) K. Richt.) | 2018 Смоленская: + 1 | Поляна в сосновом лесу | Склады и окопы | Не отмечен |
| 2. Arrhenatherum elatius (L.) J. et C. Presl # |
1926 Тверская: 25 из 37 р-нов 1884 Калужская: 1960-е, 13 из 24 р-нов 2007 Смоленская: 2 + 4 1973 Брянская: 13 из 27 р-нов 1883 Орловская: все р-ны |
Обочины дорог, луга, опушки, поляны, березняк травяной, рудеральные места | Старые дороги, перевалочные пункты у ж. д., оккупированные населенные пункты, землянки и склады, окопы, шоссе | СР: во всех обл. СЗР: нередко # |
| 3. Brachypodium peregrinum Stanislavsky et Tzvelev | 2019 Калужская: + 2 2020 Брянская: + 1 |
Разреженный березняк травяной, смешанный лес, опушка | Укрепленная “высота” (у землянок и складов), окрестности перевалочного пункта у ж. д. и полевого аэродрома | СЗР: Лен. (1 точка, окр. Гатчины) |
| 4. Brommopsis erecta (Huds.) Fourr. | 2019 Калужская: + 1 | Разреженный березняк травяной | Укрепленная “высота” (у землянок и складов) | СР: не отмечен СЗР: во всех обл. (отд. точки) |
| 5. Bromopsis ramosa (Huds.) Holub subsp. ramosa | 2020 Орловская: + 1 | Разреженная дубрава | Склады | СР: не отмечен СЗР: во всех обл. (отд. точки) |
| 6. Festuca filiformis Pourr. | 1904 Тверская: (2), 1* 2018 Смоленская: + 1 1969 Калужская: (1), 1 + 9 |
Сосняк травяной, опушки, сосново-зеленомошный лес, березняк травяной | На землянках и складах, брустверы окопов, обочины старых немецких дорог | СР: Калуж., Твер. СЗР: Лен. (окр. Выборга) |
| 7. Festuca heterophylla Lam. # |
2020 Тверская: + 2 2020 Калужская: + 1 |
Луга, опушки, мелколиственный лес, обочина дороги в березняке | Перевалочные пункты у ж. д., вблизи немецких складов и землянок | СР: не отмечен, Тамб. губ. (MW #) |
| 8. Festuca trachyphylla (Hack.) Krajina | 1904 Тверская* 1958 Смоленская: 5 + 2 1927 Калужская: [1 ] , с 1970-х из 10 р-нов 1973 Брянская: 4, (1), [2 ] + 1 1977 Орловская: 5 + 3 |
Обочины дорог в лесах, опушки, поляны | Обочины дорог, у землянок и складов | СР: большинство обл. СЗР: во всех обл., в Лен. широко |
| 9. Helictotrichon pratense (L.) Pilger | 1964? Тверская* 2014 Калужская: + 1 |
Луговой склон | Вблизи окопов | СР: Твер.? СЗР: Лен. (аборигенный), Пск. (Себеж) |
| 10. Holcus lanatus L. | 1927 Тверская: (1), 1* XIX в. Смоленская: (1?) 2019 Калужская: + 2 1974 Брянская: 5 западных р-нов из 27 * |
Обочины дорог в зеленомошном сосняке и у травяного березняка | Обочины старых дорог | СР: западные и отдельные обл. к востоку СЗР: занос. у дорог, редко |
| 11. Holcus mollis L. | 1985 Тверская: > 15, 11 из 37 р-нов * 2004 Смоленская: 2 + 1 2010 Калужская: 2 + 3 1975 Брянская: 2 |
Обочина дороги в смешанном лесу, березняк травяной, поляна, луг, лес мелколиственный | Обочина старой дороги, укрепленная высота у окопов, обочина дороги у станции перегрузки | СР: западные и отдельные обл. к востоку СЗР: нередко |
| 12. Koeleria pyramidata (Lam.) Beauv. | 1974 Калужская: 2 + 2 2018 Смоленская: + 3 |
Опушка березняка, открытый склон | Вблизи перевалочного пункты у ж. д., вблизи окопов | СР: не отмечен СЗР: Пск., Новг. (отд. точки) |
| 13. Deschampsia flexuosa (L.) Trin. # |
1895 Тверская: > 50, в 24 из 37 р-нов – *аборигенный 1961 Калужская: 7 + 7 1960?, 1984 Смоленская: 5 + 4 2001 Орловская: 3 1977 Брянская: 4 + 1 |
Сосняки зеленомошные, вблизи дорог в сосняках зеленомошных | Старые дороги, вблизи окопов, укрепленные высоты | СР: спорадически в лесной зоне СЗР: часто |
| 14. Poa chaixii Vill. # |
1870-е Тверская:(>10, 8 из 37 р-нов)* 1997 Смоленская: 1 + 1 1977 Калужская (1) |
Сложный сосняк | Окрестности складов и окопов | СР: отд. обл. (чаще в Моск.) СЗР: Лен., Новг., только # |
| 15. Trisetum flavescens (L.)
P. Beauv. # |
1917 Тверская: >10 + около 10, 13 из 37 р-нов 1958 Калужская: широко c 1980-х, 13 из 24 р-нов 2018 Смоленская: + 3 1975 Брянская: 4 |
Луга, рудеральные сообщества, сосняки, обочины дорог, опушки | Крупные перевалочные пункты, длительно оккупированные населенные пункты, обочина дорог, у шоссе, у землянок | СР: отд. обл. (9 регионов) СЗР: нередко |
| 16. Carex brizoides L. # |
2005 Тверская: 1 1999 Смоленская: 2 + 4 1983 Калужская: 1 + 4 1989 Орловская: 2 1975 Брянская: 13 из 27 р-нов * аборигенный |
Обочины дорог в лесах разного типа, реже – разреженные березняки, сложные сосняки | Около старых дорог, землянок и складов, бруствер противотанкового рва |
СР: только на западе СЗР: Лен., Новг. (интродуцент?), Пск. # |
| 17. Carex disticha Huds. | 2001 Тверская: 1 * 2006 Калужская: 3 1975 Смоленская: 2 + 1 |
Луга: пойменный, у ж. д., около шоссе | Станции перегрузки, вблизи шоссе, у ж. д. | СР: во многих обл., на юго-востоке нередко СЗР: нередко |
| 18. Carex flacca Schreb. | 1928 Тверская: 2 + 1 2006 Брянская: 1 * |
Луга, сероольшаники | Крупные перевалочные пункты у ж. д. | СР: Моск., Твер. (занос.?) СЗР: Лен., Пск. (отд. точки) |
| 19. Carex pilulifera L. | 1990 Тверская: >10 + 1, 6 из 37 р-нов – *аборигенный 2018 Смоленская: + 1 2019 Калужская: + 5 1984 Брянская: 1 |
Обочины дорог в лесах, опушки, разреженный березняк | Укрепленные высоты, вблизи окопов, обочины дорог | СР: отд. обл. СЗР: отд. точки, нередко |
| 20. Carex projecta Mack. | 1964 Тверская: + 1 | Луг | В месте расположения линии фронта | Не отмечен |
| 21. Carex tomentosa L. | 2020 Тверская: + 2 1985 Смоленская: 1 |
Луга (у насыпи ж. д.), опушка | Перевалочные пункты у ж. д., близ шоссе Москва–Минск | СР: отд. обл., много указаний ошибочных СЗР: Лен. (3 точки, Гатчина) |
| 22. Luzula campestris (L.) DC. | 2020 Калужская: + 1 | Поляна в березняке | Вблизи перевалочного пункта у ж. д. и полевого немецкого аэродрома | СР: достоверно неизвестен СЗР: Лен. (4 точки – Выборг, Мга), Пск. (отд. точки) |
| 23. Luzula luzuloídes (Lam.) Dandy et Wilmott. # |
1895 Тверская: (> 15, 10 из 37 р-нов) *парки 2007 Смоленская: 1 + 2 1988 Калужская: 1 + 4 2007 Орловская: 1 |
Березняки, опушки леса, сложный сосняк, ельник | На землянках и по брустверу окопов, склады, у старых дорог, поблизости от шоссе, вблизи полевого аэродрома | СР: отд. обл. СЗР: во всех обл., одич. в лесах и # |
| 24. Colchicum autumnale L. | 2020 Тверская: + 2 | Луга и опушки | Перевалочные пункты у ж. д. | СР: в культуре СЗР: отд. пункты у линии фронта |
| 25. Muscari botryoides (L.) Mill. | 2020 Тверская: + 1 | Луга | Перевалочные пункты у ж. д. | СР: в культуре СЗР: Лен. (1 пункт) |
| 26. Ranunculus bulbosus L. | 1984 Смоленская: 1 2020 Калужская: + 1 |
Поляна в березняке | Перевалочные пункты у ж. д. | СР: Москва (1862, #) СЗР: Лен. (2 точки, интродуцент?) |
| 27. Ranunculus nemorivagus Jord. | 2011 Смоленская: 1 | Близи дорог в сложном сосняке | Немецкие склады | СР: не отмечен СЗР: Лен. (занос., натурализуется) |
| 28. Thlaspi montanum L. (Noccaea montana (L.) F.K. Mey.) | 2019 Калужская: + 1 2020 Брянская: + 1 |
Обочины дорог | Старая дорога и окопы, около перевалочного пункта у ж. д. и шоссе | Не отмечен |
| 29. Lathyrus linifolius (Reichard) Bassler var. montanus (Bernh.) Bassler | 2019 Калужская: + 1 | У дороги в сосняке травяном | Старая дорога к укрепленной высоте | СР: не отмечен СЗР: Лен., Пск. (отд. точки у линии фронта) |
| 30. Geranium phaeum L. # |
2003 Смоленская: 1 + 2 2017 Калужская: + 1 |
Сложный сосняк, березняк травяной, ж. д. насыпь, обочина | Склады, окр. станции, обочины дорог | СР: Москва (#) СЗР: Лен. (по Мге) |
| 31. Chaerophyllum aureum L. # |
1987 Тверская: [2
]
, 1 + 1 1924 Московская: (18) 2019 Смоленская: + 1 2020 Брянская: + 1 |
Обочины дорог, рудеральные сообщества, луга, сложный сосняк, березняк, около дороги |
Обочины старых дорог, окр. складов, перевалочные пункты у ж. д., длительно оккупированные населенные пункты | СР: Моск., Твер. СЗР: Лен., Пск. (#) |
| 32. Chaerophyllum hirsutum L. # |
1992 Тверская: 1* пастбище 2004 Смоленская: 1 + 1 1981 Калужская: 1 |
Сложный сосняк, опушка | Около шоссе, складов и окопов | СР: западные обл., # СЗР: Лен. (#) |
| 33. Heracleum sphondylium L. | 2010 Тверская:1 + >20, 4 из 37 р-нов 2012 Калужская: 4 + 7 2002 Смоленская: 2 + 4 2018Брянская: + 1 2020 Орловская: + 3 |
Обочины дорог, рудеральные местообитания, опушки, луга, разреженные березняки, сложные сосняки | Длительно оккупированные населенные пункты, старые дороги и шоссе, перевалочные пункты у ж. д., укрепленные высоты, склады | СР: Смол., Твер. СЗР: во всех обл. (отд. пункты у линии фронта) |
| 34. Meum athamanticum Jacq. | 2020 Тверская: + 1 2019 Калужская: + 1 |
Луга, поляны в березняке | Перевалочные пункты у ж. д., укрепленная высота у землянок и складов | СР: не отмечен СЗР: Лен. (у линии фронта) |
| 35. Pimpinella major (L.) Huds. # |
2018 Тверская: + >20 2018 Смоленская: + 4 2011 Калужская: 1 + 7 2012 Брянская: + 1 2004 Орловская: 3 |
Обочины дорог, луга, опушки, светлые березняки, смешанные леса, осинники, сероольшаники, кустарниковые ивняки |
Старые дороги, перевалочные пункты у ж. д., длительно оккупированные населенные пункты, укрепленные высоты (у складов и землянок) | СР: Ворон., Моск., Орл. (занос., одич.) СЗР: отд. пункты у линии фронта # |
| 36. Lysimachia punctata L. # |
2006 Тверская: 2 + 1 | Луга, рудеральные сообщества | Длительно оккупированные населенные пункты | СР: только в культуре СЗР: Лен. (#) |
| 37. Primula elatior (L.) Hill. | 2019 Тверская: + 4 2002 Смоленская: 2 + 1 2015 Калужская: + 6 2020 Брянская: + 1 2008 Орловская: 1 + 1 |
Опушки, луга, сложный сосняк, смешанные леса, сероольшаники | Перевалочные пункты у ж. д., около старых дорог и шоссе, укрепленные высоты у землянок и окопов, на месте немецких складов | СР: Орл., Смол. СЗР: все обл. (отд. пункты, интродуцент?) |
| 38. Galium sylvaticum L. | 2020 Калужская: + 1 | Обочина дороги в березняке | Старая дорога к землянкам и складам | Не отмечен |
| 39. Cruciata glabra (L.) Ehrend. | 2002 Смоленская: 2 2016 Калужская: + 2 1854 Брянская: 7 из 22 р-нов * аборигенный 1885 Орловская: 1 + 1 |
Разреженные березняки (с дубом), обочины дорог, ельник | Укрепленная высота (на землянках), старые дороги, окр. перевалочных пунктов у ж. д. | СР: Брян., Смол., Курск. СЗР: все обл. отд. пункты, б. ч. у линии фронта |
| 40. Cruciata laevipes Opiz | 1917 Тверская: [1
]
+ 2 1967 Калужская: 1, [2 ]+ 4 1997 Смоленская: 2 + 2 1982 Брянская: [2 ] , 3 1982 Орловская: [2 ] |
Луга и поляны, опушки, обочины дорог, березняки, сероольшаники, черноольшаник, сложный сосняк | Перевалочные пункты у
ж. д., немецкие склады, обочины дорог, оккупированная деревня |
СР: отд. обл., чаще в черноземной полосе СЗР: все обл. отд. пункты, б. ч. у линии фронта |
| 41. Phyteuma nigrum F.W. Schmidt # |
2020 Тверская: + 2 1986 Брянская: 1 2004 Смоленская: 1 |
Луга, опушки | Перевалочные пункты у ж. д., у землянок и складов | СР: Брян., Смол. СЗР: Лен. (#) |
| 42. Phyteuma spicatum L. # |
2019 Тверская: + 1 1949 Смоленская: 1 1981 Калужская: 1 + 1 2008 Орловская: 2 |
Опушки, обочины дорог в лесах, широколиственные леса | Укрепленная высота (в окр. землянок и складов), старая дорога, перевалочные пункты | СЗР: в 5 обл. СЗР: Лен., Пск. # |
| 43. Achillea ptarmica L. | 1889 Тверская: >10 + >40, 7 из 37 р-нов 2007 Калужская: 1 + 3 |
Обочины дорог, луга у населенных пунктов, опушки | Обочины дорог, перевалочные пункты у ж. д., длительно оккупированные населенные пункты | СР: не изучено, много неверных указаний, редко СЗР: часто |
| 44. Hieracium sylvularum Jord. ex Boreau # |
2003 Калужская: 1 (1) + 1 2002 Смоленская: 1 2020 Орловская: + 2 |
Дубрава, ельник | Окр. высоты, вблизи старых дорог | СР: не изучено, только 6 обл., редко СЗР: Лен. (#) |
| 45. Hieracium vulgatum Fries s.l. # |
С учетом H. jaccardii 1880-е Тверская: >100, >30 из 37 р-нов – *аборигенный 1999 Калужская: 1 2018 Смоленская: 1 |
Сухие сосняки, ельники | Окопы и старые дороги | СР: не изучено, только в 7 обл. СЗР: нередко, распространение отд. видов не изучено |
Примечание. Названия видов расположены согласно индексам Dalla Torre et Harms. # – вид отмечен в старых усадебных парках. Во 2-м столбце приведены год первой регистрации в регионе, число пунктов (точек) или районов, в которых вид найден. Знаком “+” и полужирным шрифтом выделены данные, впервые полученные в ходе наших исследований. В круглых скобках дано число точек из старых парков, в квадратных скобках – приуроченных к железным дорогам. * – регионы, в которых возможность полемохорного заноса пока не установлена (в некоторых случаях вид может быть аборигенным), поэтому местообитания специально не обсуждаются. В 2020 г. часть прежних районов Тверской области приобрели статус муниципальных округов, но мы рассматриваем их как районы. В 3-м столбце отмечены только полемохорные местообитания (прочие исключены), которые приведены в порядке убывания частоты встречаемости и обилия вида. Полужирный шрифт – экотопы, в которых вид хотя бы в одном местонахождении встречен с большим обилием (занимает площадь более 100 м2). В 4-м столбце местонахождения, сопряженные с военными объектами и событиями, которые расположены в порядке убывания частоты встречаемости в них вида. Полужирный шрифт – объекты, на которых обилие вида значительное (занимает площадь более 100 м2). В 5-м столбце даны сведения о распространении вида в Средней (СР) (по: Маевский, 2014; MHA; MW) и Северо-Западной России (СЗР) (по: Цвелев, 2000; Ефимов, Конечная, 2018; LE), информация о встречаемости в усадебных парках (#). Названия областей сокращены (по: Цвелев, 2000; Маевский, 2014). Прочие сокращения: губ. – губерния; занос. – заносные; обл. – область; одич. – одичавший; окр. – окрестности; отд. – отдельные; р-н – район.
При специальных поисках в местах расположения войск всего за три года исследований впервые для Средней России (Маевский, 2014) обнаружено 11 видов (или центральноевропейских форм): Anthoxanthum amarum, Brachypodium peregrinum11, Bromopsis erecta, B. ramosa, Festuca heterophylla, Carex projecta, Luzula campestris, Thlaspi montanum, Lathy-rus linifolius var. montanus, Meum athamanticum, Galium sylvaticum) (Решетникова, 2020; Решетникова, Майоров, 2020; Нотов А., Нотов В., 2020б; Майоров, Решетникова, 2021; Крылов и др., 2021; Решетникова и др., 2021а; Решетникова, Киселева, 2021). Шесть из них были известны к югу от черты блокады Ленинграда на месте дислокации немецких войск (табл. 1). Поиски полемохоров, проведенные в Тверской области в начале вегетационного сезона, позволили выявить эфемероиды, которые являются характерным компонентом центральноевропейских сообществ, а в Центральной России были известны как декоративные растения: Colchicum autumnale, Muscari botryoides (Нотов А., Нотов В., 2020б; Нотов и др., 2020г).
Некоторые виды впервые приводятся нами для отдельных хорошо изученных во флористическом отношении регионов (табл. 1). Включая новые для Средней России находки, впервые отмечено: для Калужской области – 15 видов (Решетникова, 2015б, 2016, 2018, 2020), для Тверской – 10 видов (Нотов и др., 2019а; Нотов А., Нотов В., 2020а, б; Решетникова, Майоров, 2020), для Смоленской – 6 видов (Решетникова, 2019), для Брянской – 6 видов (Решетникова и др., 2021а), для Орловской – 3 вида (Решетникова, Киселева, 2021).
Встречаемость полемохоров в областях Центральной России. Широта распространения полемохоров в изученных регионах различна (табл. 1). В Смоленской области в 2018 г. полемохоры были найдены на 15 из 24 обследованных участков. В Калужской области в 2019–2020 гг. они обнаружены на 18 из 25 обследованных участков. В Тверской области в 2018–2020 гг. выявлено более 50 различных по площади (от 0.5 до 50 га) и степени обособленности участков с полемохорами, которые приурочены к территории бывшего Ржевско-Вяземского плацдарма. В 1942–1943 гг. он был важнейшим стратегическим объектом военных операций Ржевской битвы (Герасимова, 2016; Калашников, 2018). В Орловской области полемохоры были найдены в 5 пунктах, а в Брянской области – в 2 пунктах.
Почти во всех областях найдены Carex brizoides, Cruciata laevipes, Heracleum sphondylium, Pimpinella major, Primula elatior (рис. 1). Именно они были отнесены к группе вероятных полемохоров после первого предварительного анализа (Щербаков и др., 2013).
Рис. 1.
Расположение местонахождений трех наиболее характерных для Центральной России видов-полемохоров по отношению к линии фронта 1942 г.
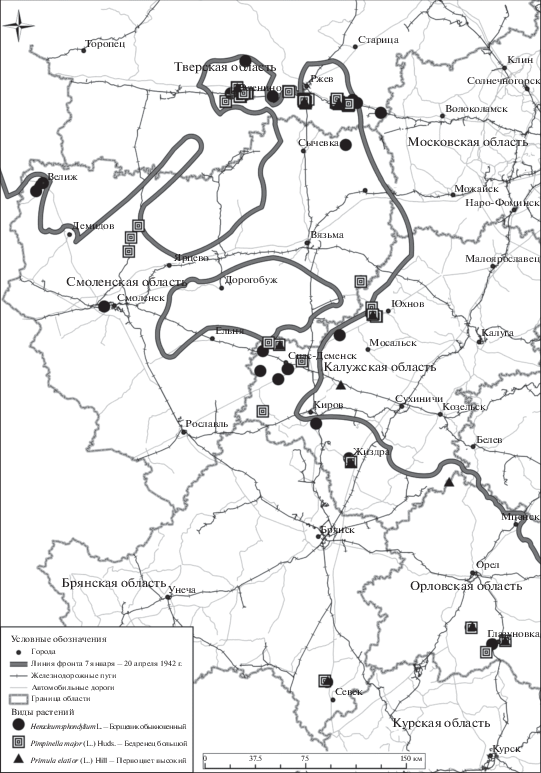
Возможность обнаружить полемохоры в разных частях регионов при специальном поиске в местах дислокации немецких войск при отсутствии их на других территориях подтверждает, что их занос происходил из одного источника, в одно время и сходным путем.
ОБСУЖДЕНИЕ
Особенности распространения полемохоров. В местонахождениях, приуроченных к территории бывших военных объектов, ценотическая роль полемохоров невысока, за исключением Ржевско-Вяземского плацдарма. Участки, на которых активность полемохоров значительна, имеют, как правило, площадь не более 100 м2.
Около дорог полемохорные виды встречались в небольшом числе особей и с низким проективным покрытием. Заросли площадью более 100 м2 сформировали только некоторые преимущественно длиннокорневищные растения – Arrhenatherum elatius (в Калужской и Брянской областях), Holcus mollis (в Калужской области), Carex brizoides (в Калужской и Смоленской областях) и Ranunculus nemorivagus (в Смоленской области).
Вблизи немецких складов в Калужской и Смоленской областях в лесах и на полянах нередко наблюдались единственные старые дерновины Koeleria pyramidata, Carex pilulifera; последняя занимала площадь более 1 м2. Единично (менее десятка особей или на небольшой площади) встречались Meum athamanticum, Bromopsis erecta, Ranunculus bulbosus, Lathyrus linifolius и др. В большом обилии (на площади более 100 м2) на нескольких участках отмечены Arrhenatherum elatius, Festuca filiformis, F. trachyphylla, Holcus mollis, Poa chaixii, Carex bri-zoides, Geranium phaeum, Heracleum sphondylium, Pimpinella major, Primula elatior, Cruciata laevipes, C. glabra22 (табл. 1). В целом большинство полемохоров были приурочены к участкам со старыми или современными нарушениями субстрата (фортификационные сооружения, обочины дорог) (Решетникова и др., 2018, 2019, 2020, 2021б; Панасенко, 2019; Панасенко, Решетникова, 2021). В ненарушенных сообществах поблизости были отмечены только Arrhenatherum elatius, Festuca trachyphylla, Poa chaixii, Trisetum flavescens, Pimpinella major, Primula elatior, Hieracium sp. Аборигенный в Тверской области вид Deschampsia flexuosa в последние годы быстро расселяется в Брянской и Орловской областях, однако в Калужской области он регистрируется преимущественно в местах военных действий.
Наиболее активная натурализация полемохоров отмечена в Тверской области. В пределах бывшего Ржевско-Вяземского плацдарма масштабы заноса диаспор полемохоров были колоссальными. В местах расположения ключевых перевалочных пунктов и в районах, плотно заселенных немцами в период продолжительной оккупации, некоторые полемохоры стали доминантами в луговых и рудеральных сообществах (Нотов и др., 2019а, б, 2020а). Эти сообщества могут встречаться на территориях значительной площади (до 5–10 га). Такие объекты найдены в Зубцовском и Ржевском районах. В числе полемохоров-доминантов – Arrhenatherum elatius, Pimpinella major, Ac-hillea ptarmica. В центральноевропейских сообществах эти виды также могут выступать в качестве доминантов (Mucina, 1997; Mucina et al., 2016; Velev, 2018). Фитоценозы с доминированием Pimpinella major или с его значительным участием отмечены около деревень Папино, Ревякино и Ровное, ж/д станции Платформа 208 км. Этот вид оказался устойчивым даже в крупнотравных травостоях с растениями, обладающими высокой конкурентной способностью (Urtica dioica L., Chamaenerion angustifolium (L.) Scop., Calamagrostis epigeios (L.) Roth, Bromopsis inermis (L.) Holub, Cirsium setosum (Willd.) Bess., Tanacetum vulgare L.) (Нотов и др., 2019а, б). Он сохраняется в осинниках, сероольшаниках, смешанных лесных фитоценозах. На некоторых территориях большие площади занимают сообщества с доминированием Arrhenatherum elatius. Высокого обилия могут достигать Achillea ptarmica, Trisetum flavescens. В окрестностях крупных перевалочных пунктов отмечены сообщества, в которых обильны Cruciata laevipes, Phyteuma nigrum, Primula elatior, Heracleum sphondylium (Нотов и др., 2020б, в, г). На других участках Ржевско-Вяземского плацдарма фитоценотическая роль этих видов незначительна.
Таким образом, в разных областях Центральной России и на различных объектах, связанных с военной историей, частота встречаемости, широта распространения и активность одних и тех же видов может сильно различаться.
Обусловленность распространения военными событиями. Для понимания региональных особенностей распространения полемохоров необходим сопряженный анализ различных факторов, которые оказали влияние на разнообразие, возможность сохранения и динамику их натурализации. В целом обилие и число полемохорных видов снижается с севера на юг. Однако характер взаимосвязи с региональными особенностями физико-географических условий нуждается в специальном изучении.
Изменение линии фронта в 1941–1942 гг. Для понимания специфики распространения полемохоров с центральноевропейским типом ареала особое значение имеют военно-исторические факторы. Расположение войск на территории Центральной России с зимы 1941–1942 гг. до осени 1943 г. последовательно проходило по трем линиям более или менее стабильной обороны. Это предопределило постоянство активной деятельности немецкой армии и, как следствие, локализированный занос диаспор полемохоров.
Наибольшее число полемохоров найдено вблизи линии фронта на Ржевско-Вяземском выступе: Оленино – Ржев (Тверская область) – Гжатск (Смоленская область) – Юхнов – Киров (Калужская область) (рис. 1). Этот объект имел особое стратегическое значение. Здесь линия обороны была оборудована на неподготовленных позициях зимой 1941 – в начале весны 1942 гг., а территория была освобождена только весной 1943 г. Фураж завозился в огромном количестве. Масштабы перевалки грузов на объекты, где шли наиболее напряженные бои – в Ржевском и Зубцовском районах Тверской области, сопоставимы с масштабами на крупных железнодорожных узлах. Однако места перевалки здесь нередко располагались вне населенных пунктов и после войны в хозяйственном отношении не использовались. Некоторые участки в результате военных действий подверглись сильным разрушениям, которые привели к существенной трансформации ландшафтов. Многие населенные пункты были полностью утрачены и после войны не восстанавливались.
Вторая линия немецкой обороны на участке Демидов – Ярцево (Смоленская область) – Спас-Деменск (Калужская область) была построена на инженерно-подготовленных позициях в конце зимы – начале весны 1943 г. Здесь для маскировки позиций использовались привозные материалы. Фураж завозился в меньшем количестве. Этот участок характеризуется наличием полемохорных видов, широко распространенных в газонных смесях.
Орловская, Брянская и бóльшая часть Смоленской области с зимы 1941–1942 гг. и до весны 1943 г. находились в немецком тылу, но здесь активно действовали партизанские отряды и войска Красной Армии, оказавшиеся в окружении. Противопартизанские операции и очистка тылов проводились с широким использованием гужевого транспорта и кавалерийских частей. Поэтому на отдельных участках, где располагались временные склады фуража, а также близ лесных дорог обнаруживаются полемохорные виды.
Меньше всего находок было сделано на немецкой линии обороны Хаген: Киров (Калужская область) – Севск (Брянская область). Оборона немецкими войсками в середине лета 1943 г. была занята на подготовленных позициях, но здесь для маскировки использовались местные материалы. Фураж завозился в небольшом количестве.
Все находки новых и редких для Центральной России полемохоров сделаны преимущественно вдоль линии фронта. Они связаны с местами расположения складов, перевалочных пунктов, на которых осуществлялась перегрузка с железнодорожного транспорта на колесный и гужевой, а также с активно используемыми немцами дорогами.
Пути и способы заноса полемохоров в Центральную Россию. Объем и состав заносимых диаспор полемохоров зависел от уровня стратегической значимости региона, продолжительности оккупации, характера использования территории в этот период, специфики реализованных боевых операций. Эти особенности сопряжены с разной соотносительной ролью путей и способов проникновения полемохоров. Можно выделить как минимум три возможных пути заноса полемохоров, которые охарактеризованы ниже.
1. С фуражом (сеном и др.). Основные поставки фуража шли зимой 1941–1942 гг. и весной 1942 г. Зимой 1942–1943 гг. объемы завоза были несколько меньше. Значительное количество фуража (а значит и диаспор полемохоров) завозилось на склады длительного хранения. Высокое разнообразие полемохоров отмечено нами на месте трех участков со складами. Первый участок расположен в окрестностях Смоленска, в долине Днепра в Красном Бору на примыкающей к городу территории. Это единственный старый лес, который существовал здесь во время войны. По нему или рядом с ним проходят магистральные железные и шоссейные дороги, которые в 1942–1943 гг. были кратчайшими путями, связывавшими Западную и Центральную Европу с центральным участком Восточного фронта. Во время оккупации Смоленска здесь располагались базовые склады группы армий “Центр” и ставка Гитлера Barenhalle (“Медвежья Берлога”), от которой остались развалины фортификационных сооружений. Здесь у дорог и вблизи остатков фортификационных сооружений нами отмечено девять видов полемохоров (Решетникова и др., 2018). Второй участок расположен в сохранившемся ненарушенном сосновом лесу с полянами в долине Угры у д. Кобелево Темкинского района Смоленской области. Здесь располагались склады немецкой дивизии. Кобелево оставалось в ближнем тылу немецких частей в течение почти целого года, до марта 1943 г. Здесь отмечено 15 видов полемохоров (Решетникова и др., 2019). Третий участок находился в долине Угры у д. Харинки Калужской области. Здесь в 1941–1942 гг. немцы держали оборону более года. На этом объекте обнаружено девять видов. Несмотря на то, что все эти пункты расположены в бассейнах разных рек, 80% видового состава их растений-полемохоров совпадает.
Сопоставимы по объему заносимых с фуражом диаспор места, в которых осуществлялась его перевалка с железнодорожного транспорта на гужевой и автомобильный. Вначале мы предполагали, что количество диаспор на них было значительно ниже в связи с коротким периодом нахождения грузов. Однако в Тверской области в районе Ржевско-Вяземского плацдарма у построенной в 1942 г. немцами железной дороги в окрестностях д. Папино (Ржевский район) и на участке около ж/д станции Платформа 208 км (Зубцовский район) было обнаружено не менее чем по 20 видов полемохоров на каждом из этих объектов. Многие из них встречаются обильно (Нотов А., Нотов В., 2020б; Нотов и др., 2020а, б, в, г). В общей сложности на двух модельных территориях вблизи отмеченных перевалочных пунктов было обнаружено 25 видов растений-полемохоров, а также необычных для России форм, нуждающихся в дополнительном изучении. В Калужской области близ бывшей тупиковой железнодорожной станции в Городском бору г. Жиздры было найдено 18 полемохоров (Решетникова и др., 2020, 2021б; Крылов и др., 2021). С этой станции снабжались немецкие войска, воевавшие в районе Людиново–Жиздра, а также располагавшиеся здесь резервные части. Вероятно, большое число видов обусловлено также хорошей сохранностью экотопов со времен войны.
Интересен участок, обнаруженный в Брянской области в урочище “Зеленинский лес” Севского района, где полемохорные растения найдены не у немецких складов, а у землянок партизан или местных жителей. Всего найдено 10 видов (Панасенко, Решетникова, 2021; Решетникова и др., 2021а). Анализ известной нам информации позволяет утверждать, что занос полемохоров на урочище, вероятно, произошел в 1942–1943 гг. с краденым сеном из распложенного поблизости военного склада в с. Пушкино, где находились немецкие конюшни, либо с фуражом, захваченным из двигавшегося к ним обоза. По свидетельству местных жителей, немцы иногда предоставляли крестьянам своих лошадей для вспашки полей и уборки урожая.
Меньшее разнообразие полемохоров выявлено рядом с шоссейными и грунтовыми дорогами, по которым во время войны постоянно осуществлялись перевозки. В Калужской области у Варшавского шоссе, по которому шли основные поставки в сторону Юхнова, найдено только пять видов, и, как правило, на разных участках. В Смоленской области близ прифронтового города Велижа, вдоль Западной Двины на дороге, ведущей в Витебск, отмечено шесть видов. Некоторые из них были найдены экспедицией БИН РАН в 2002–2003 гг. (Бузунова и др., 2004), другие обнаружены нами (Решетникова, 2018). В районах противопартизанских действий в Орловской и Смоленской областях число обнаруженных полемохоров невелико. В большинстве районов мы находили не более 2–5 видов. Максимальное число видов (6) найдено к северу от д. Знаменское, в урочище “Задняя Роща” (Решетникова, Киселева, 2021). Хотя полемохоры у дорог растут обычно в небольшом числе, но протяженность лесных дорог велика и площадь, на которой они могут быть обнаружены, значительна.
В населенных пунктах вблизи долговременной линии фронта в Калужской и Тверской областях регулярно встречается Heracleum sphondylium, который можно считать своеобразным индикатором их продолжительной оккупации. Определенный уровень хозяйственной деятельности в таких деревнях нередко поддерживается до настоящего времени, что способствует сохранению наиболее обычных полемохоров.
2. С травосмесями. Некоторые центральноевропейские растения широко использовали ранее в усадебных парках Центральной России, особенно в дворянских усадьбах Московской и Тверской губерний. Среди них Arrhenatherum elatius, Poa chaixii, Trisetum flavescens, Carex brizoides, Luzula luzuloides, Chaerophyllum hirsutum, Phyteuma spicatum и др. (Ротов, Швецов, 1989а, б; Майоров и др., 2012; Маевский, 2014). На первых этапах исследования значительное сходство с составом полемохоров мы связывали только с общностью источников происхождения. Как правило, посевной материал для усадебных парков заказывали в Центральной и Западной Европе. Оттуда же во время войны, как правило, завозился фураж на Восточный фронт. Однако материалы 2019 г. позволили выдвинуть и другую гипотезу. Большое совпадение состава флоры усадебных парков с флорой военных “высот” на территории Калужской области (по крайней мере 14 видов, отмеченных на высотах) позволяет предположить, что оно не случайно. Согласно канонам деятельности инженерных войск, основная линия обороны должна проходить по господствующим высотам. Противодействуя советской авиаразведке, немецкие войска тщательно маскировали свои позиции, особенно тыловые. Мы предполагаем, что, по крайней мере, в Калужской области при строительстве на ключевых высотах зимой и весной 1943 г. немцы осуществляли специальный подсев газонных травосмесей. Предположение подтверждается обилием и разнообразием центральноевропейских видов именно на земляных сооружениях, построенных в это время, и рядом с ними. Оно значительно превышает таковое в местах обследованных ранее складов, станций снабжения и обочин дорог региона. Укрепления были построены на участках, находящихся в течение года в немецком тылу, где оккупанты имели возможность заготавливать местное сено или отбирать его у населения. Судя по топографическим картам военного времени, прилегающие территории были безлесными, и местного фуража должно было быть вполне достаточно для гужевого транспорта, который использовали при строительстве укреплений и в войсках. В иное время года (и соответственно при создании укреплений на других высотах) земляные сооружения, в соответствии с руководством по инженерной фортификации, можно было маскировать дерном (Franke, 2015; Ястребов, 2019).
После схода снега и до момента развития травяного покрова выброшенная земля хорошо заметна на фоне засохшей прошлогодней травы. Мы предполагаем, что с целью маскировки поверх высеянных трав могли укладывать слой сена или соломы. Эта гипотеза позволяет объяснить появление видов, которые не входили в состав газонных травосмесей, но были широко представлены в завозимом сене. На месте двух укрепленных немцами высот Спас-Деменского района Калужской области – “Высоте 242” у с. Лазинки и на Гнездиловских высотах – найдено по 18 центральноевропейских видов, 10 из них были известны в парковой культуре (Решетникова и др., 2020, 2021б).
Другими объектами, на которых могли высеиваться травосмеси, являются полевые аэродромы, особенно специально построенные в лесу. Лесные травянистые растения мало устойчивы к вытаптыванию, а прочность грунта (устойчивый травяной покров) на летном поле является необходимым условием успешного взлета или посадки самолета. При строительстве и эксплуатации грунтовых аэродромов со времени зарождения авиации и до наших дней во всех странах предусмотрено использование специальных аэродромных травосмесей (Доусон, 1957; Руководство…, 1994; Приказ…, 2008, и др.). Ряд центральноевропейских видов был обнаружен вблизи участка, который обозначен как полевой аэродром в Городском бору г. Жиздра Калужской области (Решетникова и др., 2020, 2021б). Они произрастали на ровных полянах, где не было заметно ни укреплений, ни остатков складов, и действительно напоминали газон.
Газонные растения, использующиеся в травосмесях, как правило, относятся к обычным аборигенным луговым видам злаков или широко распространенным интродуцированным видам (Руководство…, 1994, и др.). Однако только в старых руководствах довоенного времени встречаются рекомендации о высаживании на газонах центральноевропейской Festuca filiformis. Последний вид мы отметили в Калужской области в девяти пунктах (только в местах со следами деятельности немецких войск). На кислых почвах также рекомендовались Deschampsia flexuosa и Sieglingia decumbens. Как примеси на газонах Р.Б. Доусоном (1957) указаны еще Trisetum flavescens, Holcus mollis и H. lanatus. Именно эти виды мы регулярно наблюдали на различных немецких земляных сооружениях и около них, а также на немецких коммуникациях военного времени. Однако некоторые из этих злаков в Средней России в последние десятилетия стали прогрессирующими видами, что затрудняет дифференциацию полемохорных и прочих местообитаний, в которых эти растения оказались в результате расширения естественного ареала (Deschampsia flexuosa) или были занесены иным путем.
3. При перевозке грузов и техники. Уникальной можно считать находку североамериканского вида Carex projecta Mack., который был собран в 1964 г. Г.Ю. Загородней у д. Залужье Западнодвинского района Тверской области и в 2020 г. определен С.Р. Майоровым (Решетникова, Майоров, 2020). За пределами Нового Света этот вид осоки был неизвестен (POWO, 2020). Однако, согласно архивной копии отчетной карты Калининского фронта за середину ноября 1942 г., здесь проходила линия снабжения советских войск. По этой дороге, помимо других поставок, могли поступать и грузы, полученные по ленд-лизу, в том числе из Америки. Иной вектор заноса сложно представить. Если мы правы, то C. projecta – единственный известный нам пример растения-полемохора, занесенного в Россию с другого континента.
Натурализация полемохоров
Предпосылки сохранения в разных экотопах. Устойчивость полемохоров в местах заноса определяется взаимным влиянием различных факторов. Тип наиболее благоприятного для сохранения местообитания в значительной степени обусловлен особенностями биологии и экологии полемохорных видов. Вторым важным фактором можно считать характер динамики экотопа, растительности и ландшафта.
Луга. В фураже и в травосмесях преобладают луговые растения, которым требуется хорошее освещение и богатая почва. Однако луга в Нечерноземье подвержены быстрым изменениям, которые происходят как при смене хозяйственной деятельности, так и в ходе естественных сукцессионных смен. После войны луга распахивались, использовались как сенокосы и пастбища, часто этому сопутствовала мелиорация. При отсутствии хозяйственной деятельности луговые сообщества сменяются лесными. На нашей территории это, как правило, мелколиственные леса. Кроме того, вероятность внедрения в сомкнутые ненарушенные луговые сообщества минимальна, так как в них отсутствуют необходимые для успешного внедрения участки с разреженным или поврежденным травяным покровом. В этой связи на лугах в большинстве областей находки полемохоров немногочисленны. Исключением являются луговые сообщества на ключевых перевалочных пунктах Ржевско-Вяземского плацдарма (Тверская область). На этих объектах колоссальный объем завозимого фуража сочетался с необходимой степенью нарушения растительного покрова. На некоторых участках относительно стабильный уровень умеренного хозяйственного использования луговых сообществ поддерживается до настоящего времени.
Леса и опушки. Полемохоры могут долгое время сохраняться в лесных сообществах в основном на участках, где достаточно света (табл. 1). В Центральной России в местах распространения лесных фитоценозов местообитания, которые в большей степени приближены к луговым экотопам, встречаются на полянах и опушках, в сложных сосняках, травяных березняках или березняках с дубом. При отсутствии подлеска в травяном ярусе здесь преобладают опушечные и луговые растения. Принципиальными, по-видимому, являются два фактора – достаточная освещенность местообитания и разреженный травяной покров. Наибольшее число видов полемохоров зарегистрировано на некоторых участках в березняках с полянами – 18 (Решетникова и др., 2020) и в сложных сосняках – 15 (Решетникова и др., 2018). Большое число видов приурочено к обочинам лесных дорог, где наблюдается более четкое выполнение обоих отмеченных условий. Именно сюда и попадали изначально диаспоры чужеродных видов. Меньше видов отмечено в сосняках зеленомошных. Как правило, это растения не требовательные к плодородию почв, но в достаточной мере светолюбивые. Некоторые теневыносливые виды могут быть устойчивы в ельниках или широколиственных лесах. Вторым важным фактором, определяющим сохранение полемохорных видов в лесах и на полянах, является поддержание в лесах неизменного режима умеренного хозяйственного использования. На месте нарушенных или сведенных со времени войны лесов находки полемохоров невозможны.
Адаптивный потенциал натурализовавшихся полемохоров. Иногда сообщества мелколиственных лесов с полемохорами могут формироваться в результате демутационных сукцессий луговых фитоценозов, на которых они исходно произрастали. На ключевых территориях Ржевско-Вяземского плацдарма с очень мозаичным растительным покровом выявлены модельные участки, на которых по мере возникновения на месте луговых сообществ с полемохорами опушечных и лесных фитоценозов происходило постепенное выпадение центральноевропейских растений, представляющих луговую эколого-фитоценотическую группу (Нотов и др., 2020а, в). В их числе Meum athamanticum, Muscari botryoides, Trisetum flavescens, Achillea ptarmica, Phyteuma nigrum. Некоторые из этих видов на других участках еще встречаются на опушках и в примыкающих к ним луговых фитоценозах (табл. 1). Даже такой массовый и явно доминирующий в луговых и рудеральных сообществах вид, как Arrhenatherum elatius, на опушках встречается редко и необильно, а в лесных фитоценозах отсутствует. Подобные сукцессии могли происходить, по-видимому, и в других регионах.
Устойчивыми во вторичных лесных сообществах, образовавшихся на месте луговых и рудеральных фитоценозов, оказались виды с более широкой эколого-фитоценотической амплитудой (Нотов и др., 2020а, в). Среди них в Тверской области – Carex flacca, а в пределах большинства других изученных областей – Arrhenatherum elatius, Cruciata laevipes, Festuca heterophylla, Pimpinella major, Primula elatior. Почти все они в Центральной Европе распространены не только на лугах, но и отмечены в составе диагностических компонентов сообществ зональных неморальных лесов умеренного пояса или нитрофильных фитоценозов. Однако бóльшая часть этих видов в лесных сообществах изученных территорий встречается сейчас очень локально и с небольшим обилием (табл. 1) (Нотов и др., 2019а, б). Только Pimpinella major в лесных и опушечных фитоценозах распространена более или менее регулярно. В ряде случаев она может играть в данных сообществах существенную фитоценотическую роль.
Наиболее полно высокий адаптивный потенциал реализовали виды, которые могут выступать в центральноевропейских сообществах в качестве доминантов (Mucina, 1997; Mucina et al., 2016; Velev, 2018): Arrhenatherum elatius, Pimpinella major, Achillea ptarmica. По-видимому, все это способствовало тому, что отмеченные полемохоры проявляют в настоящее время тенденцию к расселению, что особенно ярко выражено на территории Ржевско-Вяземского плацдарма (Нотов и др., 2019а, б, 2020а). Участие этих видов во флоре Центральной России в некоторой степени сходно с “поведением” видов, расширяющих ареал.
Сообщества с участием центральноевропейских видов. Благодаря тому, что полемохоры могут продолжительное время удерживаться в местах заноса, в изученных областях Центральной России они стали компонентами разных растительных сообществ (табл. 1). Уровень разнообразия полемохоров и их ценотическая значимость в них различны. На подавляющем большинстве обследованных участков площадь, которую занимают фитоценозы с полемохорами, небольшая, а видовое богатство чужеродных видов низкое. Однако на территориях, приуроченных к ключевым стратегическим объектам военного времени, ценотическая роль полемохоров и фитоценозов с их участием может быть значительной. Такое распространение полемохоров отмечено в окрестностях перевалочных пунктов у железных дорог Ржевско-Вяземского плацдарма в Тверской области и (в меньшей степени) на укрепленных высотах в Калужской области.
В результате масштабного заноса во время войны диаспор из Центральной Европы на ключевых объектах оккупированной территории сформировались фитоценозы, проявляющие сходство со среднеевропейскими сообществами. Они не характерны для Средней России. Даже спустя 70 лет после окончания войны эти своеобразные аналоги луговых фитоценозов Центральной Европы могут сохранять свои функциональные и структурные особенности. На ключевых модельных территориях Ржевско-Вяземского плацдарма нами обнаружены луговые фитоценозы, в которых достаточно полно представлен диагностический компонент среднеевропейских сообществ класса Molinio-Arrhenatheretea Tx. 1937 (Нотов и др., 2020а, б, в, г). В этих фитоценозах встречаются также диагностические виды некоторых соподчиненных синтаксонов этого класса. В их числе союзы Triseto flavescentis-Polygonion bistortae Br.-Bl. et Tüxen ex Marschall 1947 и Arrhenatherion elatioris Luquet 1926. Эти участки представляют уникальную модель для анализа механизмов формирования сообществ, воспроизводящих характерные особенности компонентов растительности других ботанико-географических регионов (Нотов и др., 2020а).
Последствия массового заноса семенного материала из Центральной Европы не ограничиваются только проникновением среднеевропейских видов в Восточную Европу. В регионы Центральной России могли заноситься также диаспоры видов с более широким долготным распространением. Благодаря этому в местах ключевых перевалочных пунктов могли появиться: 1) новые для региона центральноевропейские чужеродные виды; 2) полемохорные популяции уже известных в регионе чужеродных растений с разной широтой долготного распространения; 3) полемохорные популяции встречающихся в регионе аборигенных видов.
Эта проблема нуждается в специальном комплексном исследовании. В местах максимального разнообразия полемохоров отмечены полемохорные популяции редких аборигенных и чужеродных для флоры Тверского региона видов. На ключевых модельных территориях Ржевско-Вяземского плацдарма в числе таких видов Allium angulosum L., Carex hartmanii Cajand., Juncus inflexus L., Salvia pratensis L., Sanguisorba officinalis L. (Нотов А., Нотов В., 2019; Нотов и др., 2020а, б, в). Их популяции нельзя отнести ни к элементам аборигенной фракции флоры, ни к результатам экспансии чужеродных видов.
Расселение полемохоров, гибридизация с аборигенными видами
Расселение полемохоров. Некоторые виды, для которых отмечена возможность полемохорного появления, после Второй мировой войны широко расселились в Центральной России. Однако в их “освоении” этой территории значимую роль могли играть и другие источники заноса. Более или менее определенно связать высокую скорость их распространения с активным внедрением в период военных действий удается только в ключевых с позиции высокого разнообразия полемохоров районах (Нотов и др., 2019б, 2020б, в). В то же время некоторые виды, которые могли появиться на юге Центральной России в результате полемохорного заноса, в Северо-Западной России и Тверской области являются аборигенными. Среди них Deschampsia flexuosa, Carex pilulifera, не исключено, что и Holcus mollis (Цвелев, 2000). Их типичные естественные местообитания – опушки лесов и поляны, реже луга и светлые леса. Отдельные виды в этих регионах встречаются на месте старых парков (табл. 1) (Цвелев, 2000; Ефимов, Конечная, 2018). Некоторые виды в Центральной России распространяются в настоящее время по насыпям железных дорог (Arrhenatherum elatius, Cruciata laevipes, Carex disticha, Festuca trachyphylla). По-видимому, важную роль полемохорные заносы играли в послевоенном расселении Arrhenatherum elatius, Cruciata laevipes. Они по разным причинам прогрессируют в Центральной России. Способность к акклиматизации дала им возможность шире расселиться после войны. Подтверждает гипотезу об их частично полемохорном внедрении и то, что их довоенные сборы приурочены только к паркам или железным дорогам, а в естественных местообитаниях на территории Центральной России они раньше не встречались. В исследованных областях они регистрируются прежде всего в районах дислокации немецких войск (Панасенко, 2019), где выявлены также нетипичные формы, не найденные на других территориях. Например, только в местах расположения немецких частей обнаружены опушенные формы Arrhenatherum elatius. Очевидно, что для большинства видов, широко расселившихся к настоящему времени, во многих местонахождениях сейчас сложно установить возможность появления во время Второй мировой войны.
Гибридизация с аборигенными видами. Дальнейшая судьба центральноевропейских видов и форм может быть неоднозначной. При исследовании полемохоров мы неоднократно наблюдали промежуточные по морфологии между Pimpinella major и P. saxifraga L. экземпляры, рассматриваемые как P. × intermedia Figert. Они обнаружены в трех пунктах в Смоленской, в двух – в Калужской и в двух – в Тверской областях. Гибридное происхождение удалось подтвердить молекулярными данными (по материалам О.И. Кузнецовой). Интересно отметить, что P. × intermedia в Смоленской области был собран на участке, где отсутствовал P. major.
В Калужской и Тверской областях нами неоднократно (в Калужской, по крайней мере, в 4 пунктах, а в Тверской – более чем в 10 пунктах) встречены, по всей вероятности, гибриды Heracleum sphondylium с широко распространенным H. sibiricum L. (Нотов и др., 2019б). Они имели беловатую (но с зеленым оттенком) окраску венчика и не увеличенные краевые лепестки, промежуточную форму листьев и произрастали рядом с родительскими видами.
По-видимому, и для некоторых других полемохорных видов, способных образовывать гибриды с близкими аборигенными видами или формами, можно предполагать дрейф генов из центральноевропейских популяций. Не исключено, что увеличение численности некоторых редких до войны аборигенных видов может быть связано с возросшим в послевоенное время генетическим разнообразием популяций. В качестве объектов для дальнейшего специального изучения и проверки данной гипотезы, на наш взгляд, могут быть интересны Sieglingia decumbens, Cerastium arvense L., Fragaria moschata L. и, возможно, Carex caryophyllea Latourr.
ЗАКЛЮЧЕНИЕ
Возможность предсказать места находок центральноевропейских видов в Центральной России на основе анализа событий военного времени свидетельствует о реальности феномена полемохорного появления этих растений. Хотя с момента окончания Великой Отечественной войны прошло уже три четверти века, на территории Центральной России в местах дислокации немецких войск выявлено более 45 видов.
Специфика современного распространения полемохоров в регионах Центральной России обусловлена различными военно-историческими и природными факторами. Исходный уровень разнообразия полемохоров определялся составом и количеством заносимых диаспор, которые зависели от продолжительности оккупации и специфики боевых действий. Наибольшее число полемохорных видов регистрируется вблизи мест перевалки грузов с железных дорог на шоссейные и грунтовые, а также участков, где располагались немецкие склады. Основным источником заноса семян полемохоров был фураж для лошадей (в первую очередь сено). Однако на некоторых военных объектах, по-видимому, использовались травосмеси для засева фортификационных сооружений и полевых аэродромов.
Большое значение для сохранения полемохоров имели тип растительных сообществ, уровень трансформации ландшафтов, хозяйственное использование территории и специфика сукцессионной динамики растительного покрова. Наибольшее число полемохоров удерживается на лугах, полянах и опушках в светлых лесах, мало изменившихся в ходе хозяйственной деятельности и естественной сукцессии сообществ. Во вторичных лесных сообществах, образовавшихся на месте луговых и рудеральных фитоценозов, устойчивы только некоторые полемохоры с широкой эколого-фитоценотической амплитудой. Стенотопные виды в этих сообществах полностью исчезли или сохранились только на опушках и прилегающих к ним лугах.
Подавляющее большинство полемохоров не проявляет склонности к дальнейшему распространению. В настоящее время тенденция к расселению выявлена лишь у некоторых видов, широко распространенных и способных к доминированию в центральноевропейских сообществах (Arrhenatherum elatius, Pimpinella major). Выявлены случаи гибридизации полемохоров с аборигенными видами.
Полученные результаты подтверждают, что Великая Отечественная война стала важным фактором современного генезиса флоры Центральной России. Они свидетельствуют о недооценке роли военных событий в формировании региональных флор. Некоторые виды, которые ранее считались аборигенными, следует отнести к антропохорным и, соответственно, к чужеродным.
Дальнейшее комплексное изучение полемохорной фракции флоры может внести весомый вклад в выявление закономерностей функционирования экосистем с чужеродными видами и решении общих проблем инвазионной биологии.
Благодарим А.Н. Сенникова (БИН РАН) за проверку определения гербарного материала и за обсуждение результатов работы. За помощь в сборе материала и конструктивное обсуждение мы благодарны В.А. Нотову (ТвГУ), Л.Л. Киселевой (ОГУ), Е.О. Корольковой (МГПУ), Н.Н. Панасенко (БрянГУ), И.А. Фадеевой (СмолГУ). Выражаем признательность А.С. Коваленко, В.Г. Меринкову и В.В. Гариничеву за помощь в работе с военными и картографическими материалами, а также Ю.И. Помазкову за организацию и помощь в исследованиях в окрестностях агробиостанции МПГУ “Лазинки”, А.Д. Нефедовой за работу по идентификации узколистных овсяниц. Благодарим А.В. Крылова и Н.В. Воронкину за собранные и переданные материалы по весенней флоре Гнездиловских высот и Жиздринского городского бора.
Сбор и анализ данных по распространению полемохоров и подготовка рукописи выполнены при финансовой поддержке РФФИ (грант № 18-04-01206-а), а также в рамках Государственного задания ГБС РАН “Биологическое разнообразие природной и культурной флоры: фундаментальные и прикладные вопросы изучения и сохранения” № АААА-А18-118021490111-5 и НИОКР МГУ “Анализ структурного и хорологического разнообразия высших растений в связи с проблемами их филогении, таксономии и устойчивого развития” № АААА-А16-116021660045-2.
Список литературы
Алексеев Е.Б., 1985. Овсяница нитевидная (Festuca filiformis Pourr.) в Европейской части СССР // Вестн. МГУ. Сер. 16. Биология. № 2. С. 29–31.
Бузунова И.О., Конечная Г.Ю., Цвелёв Н.Н., 2004. Дополнение к флоре Смоленской области // Бюлл. МОИП. Отд. биол. Т. 109. № 3. С. 74–75.
Герасимова С.А., 2016. “Я убит подо Ржевом”: трагедия Мончаловского “котла”. М.: Яуза. 377 с.
Доусон Р.Б., 1957. Создание и содержание газона. М.: Изд-во Мин-ва коммунального хоз-ва РСФСР. 217 с.
Ефимов П.Г., Конечная Г.Ю., 2018. Конспект флоры Псковской области (сосудистые растения). М.: Т-во науч. изд. КМК. 471 с.
Калашников А.С., 2018. Первая Ржевско-Вяземская операции на берегах река Осуга. Ч. 1–2. https://ok.ru/osugagrupp/topic/68643282679231
Красная книга Калужской области, 2006. Калуга: Золотая Аллея. 608 с.
Крылов А.В., Воронкина Н.В., Решетникова Н.М., 2021. Дополнения к флоре Калужской области и Средней России по материалам 2020 г. // Бюлл. МОИП. Отд. биол. Т. 126. № 3. С. 24–25.
Маевский П.Ф., 2014. Флора средней полосы европейской части России. 11-е изд. М.: Т-во науч. изд. КМК. 635 с.
Майоров С.Р., Решетникова Н.М., 2021. Дополнения к флоре Средней России из Калужской области (по материалам 2020) // Бюлл. МОИП. Отд. биол. Т. 126. № 3. С. 22–24.
Майоров С.Р., Бочкин В.Д., Насимович Ю.А., Щербаков А.В., 2012. Адвентивная флора Москвы и Московской области. М.: Т-во науч. изд. КМК. 536 с.
Меринков В.Г., 2003. Флористические находки в Смоленской области // Бюлл. МОИП. Отд. биол. Т. 108. № 3. С. 82–83.
Назаров М.И., 1927. Адвентивная флора средней и северной части РСФСР за время войны и революции // Изв. Гл. бот. сада СССР. Т. 26. № 3. С. 223–234.
Нотов А.А., 1994. Дополнения к флоре Тверской области // Флора и растительность Тверской области. Тверь: Изд-во ТвГУ. С. 4–12.
Нотов А.А., Нотов В.А., 2019. О полемохорных и аборигенных популяциях некоторых видов флоры Тверской области // Вестн. ТвГУ. Сер. Биология и экология. № 4 (56). С. 84–102.
Нотов А.А., Нотов В.А., 2020а. Новые данные о флоре Тверской области // Бюлл. МОИП. Отд. биол. Т. 125. № 3. С. 38–41.
Нотов А.А., Нотов В.А., 2020б. Дополнения к флоре Тверской области // Бюлл. МОИП. Отд. биол. Т. 125. № 6. С. 40–45.
Нотов А.А., Мейсурова А.Ф., Зуева Л.В., Андреева Е.А., 2018. Среднеевропейские виды во флоре Тверского региона на рубеже XIX–ХХ веков // Вестн. ТвГУ. Сер. Биология и экология. № 2. С. 204–215.
Нотов А.А., Нотов В.А., Зуева Л.В., Андреева Е.А., 2019а. Полемохоры Тверской области и проблема биологических инвазий // Разнообразие растительного мира. № 3 (3). С. 22–27.
Нотов А.А., Нотов В.А., Зуева Л.В., Андреева Е.А., Мидоренко Д.А., 2019б. О распространении некоторых растений-полемохоров в Тверской области // Вестн. ТвГУ. Сер. Биология и экология. № 3 (55). С. 161–175.
Нотов А.А., Нотов В.А., Зуева Л.В., Иванова С.А., 2020а. Сукцессионная динамика фитоценозов с участием полемохоров // Полевой журн. биолога. Т. 2. № 4. С. 260–271.
Нотов А.А., Нотов В.А., Иванова С.А., Зуева Л.В., Мидоренко Д.А., 2020б. Экология и фитоценология Primula elatior в Тверской области // Вестн. ТвГУ. Сер. Биология и экология. № 2 (58). С. 94–104.
Нотов А.А., Нотов В.А., Иванова С.А., Зуева Л.В., Мидоренко Д.А., 2020в. Cruciata laevipes в экосистемах Тверской области // Вестн. ТвГУ. Сер. Биология и экология. № 3 (59). С. 74–85.
Нотов А.А., Нотов В.А., Петухова Л.В., Иванова С.А., Андреева Е.А., 2020г. О полемохорных популяциях Colchicum autumnale // Вестн. ТвГУ. Сер. Биология и экология. № 4 (60). С. 95–105.
Огненная дуга, 2003. М.: Звонница МГ. 631 с.
Панасенко Н.Н., 2019. Arrhenatherum elatius (L.) J. et C. Presl в Брянской области // Разнообразие растительного мира. № 3 (3). С. 26–38.
Панасенко Н.Н., Решетникова Н.М., 2021. Урочище “Зеленинский лес” – центр разнообразия растений-полемохоров в Брянской области // Бот. журн. Т. 106. (в печати).
Перечень (список) видов грибов, лишайников и растений, занесенных в Красную книгу Смоленской области (по состоянию на 1 марта 2012 г.): Приложение 1 к Приказу и.о. начальника Департамента Смоленской области по охране, контролю и регулированию использования объектов животного мира и среды их обитания № 119 от 29 мая 2012 г. http://oopt.aari.ru/sites/default/files/documents/Departament-Smolenskoy-oblasti-po-ohrane-kontrolyu-i-regulirovaniyu-ispolzovaniya-obektov-zhivotnogo-m/N119_29-05-2012.pdf
Приказ о маскировке аэродромов, войсковых частей и важных военных объектов округов № 0042 от 19.06.1941 по Комиссариату обороны СССР, 2008 // 1941. Борьба за господство в воздухе / Под ред. Хазанова Д.Б. М.: Яуза, ЭКСМО. С. 575–576.
Решетникова Н.М., 2015а. Путь появления некоторых западноевропейских растений в Калужской области – путь следования немецкой армии в 1941–1943 // Рос. журн. биол. инвазий. № 4. С. 95–104.
Решетникова Н.М., 2015б. Дополнения к флоре Калужской области по материалам 2014 г. // Бюлл. МОИП. Отд. биол. Т. 120. № 6. С. 69–74.
Решетникова Н.М., 2016. Новые и редкие для Средней России виды растений, найденные в Калужской области // Бюлл. МОИП. Отд. биол. Т. 121. № 3. С. 66–69.
Решетникова Н.М., 2018. Дополнения к флоре Калужской области по материалам 2015–2016 гг. // Бюлл. МОИП. Отд. биол. Т. 123. № 3. С. 64–70.
Решетникова Н.М., 2019. Новые данные по флоре Смоленской области (2017–2018) // Бюлл. МОИП. Отд. биол. Т. 124. № 3. С. 36–43.
Решетникова Н.М., 2020. Дополнения к флоре Калужской области и Средней России по материалам 2019 г. // Бюлл. МОИП. Отд. биол. Т. 125. № 3. С. 51–57.
Решетникова Н.М., Киселева Л.Л., 2021. Дополнения к флоре Орловской области по материалам 2020 г. // Бюлл. МОИП. Отд. биол. Т. 126. С. 31–33.
Решетникова Н.М., Майоров С.Р., 2020. Дополнения к флоре Средней России // Бюлл. МОИП. Отд. биол. Т. 125. № 3. С. 42–46.
Решетникова Н.М., Щербаков А.В., Фадеева И.Ф., 2018. Материалы к флоре “Красного бора” – уникальной охраняемой территории Смоленской области // Вестн. ТвГУ. Сер. Биология и экология. № 4. С. 160–190.
Решетникова Н.М., Щербаков А.В., Королькова Е.О., 2019. Центральноевропейские виды в окрестностях д. Кобелево (Смоленская область) как следы Великой Отечественной войны // Бот. журн. Т. 104. № 7. С. 1122–1134.
Решетникова Н.М., Щербаков А.В., Королькова Е.О., 2020. Три участка военной истории – растения-полемохоры Калужской области // Вестн. ТвГУ. Сер. Биология и экология. № 4 (60). С. 106–132.
Решетникова Н.М., Панасенко Н.Н., Щербаков А.В., 2021а. Дополнения к флоре Брянской области по материалам 2020 г. // Бюлл. МОИП. Отд. биол. Т. 126. № 3. С. 28–31.
Решетникова Н.М., Щербаков А.В., Королькова Е.О., 2021б. Поиски центральноевропейских растений на месте дислокации немецких войск в Калужской области // Рос. журн. биол. инвазий. Т. 14. (в печати).
Ротов Р.А., Швецов А.Н., 1989a. К флоре усадебных парков Подмосковья // Бюл. Гл. бот. сада АН СССР. Т. 154. С. 27–30.
Ротов Р.А., Швецов, А.Н., 1989б. Очаги натурализации адвентивных видов в усадебных парках Подмосковья // Проблемы изучения адвентивной флоры СССР (Мат-лы совещания 1–3 февраля 1989 г.). М.: Наука. С. 32–33.
Руководство по эксплуатации гражданских аэродромов Российской Федерации (РЭГА РФ-94), утвержденное приказом Директора Департамента воздушного транспорта от 19.09.1994 № ДВ-98. https://docs.cntd.ru/document/1200029032
Сенников А.Н., 2006. Находки новых и редких видов Hieracium (Asteraceae) в Тверской области // Бюлл. МОИП. Отд. биол. Т. 111. № 6. С. 60–63.
Сенников А.Н., 2012. Горькая память земли: Растения-полемохоры в Восточной Фенноскандии и Северо-Западной России // Проблемы изучения адвентивной и синантропной флор России и стран ближнего зарубежья. Ижевск: ИИКИ. С. 182–185.
Скворцов А.К., 2005. Несколько дополнений к флоре Смоленской области // Бюлл. МОИП. Отд. биол. Т. 110. № 2. С. 65–66.
Хааре А.О., 1979. Новое местонахождение реликтовых видов в Ленинградской области // Новости сист. высш. раст. Т. 15. С. 240–247.
Цвелев Н.Н., 2000. Определитель сосудистых растений Северо-Западной России (Ленинградская, Псковская и Новгородская области). СПб.: Изд-во СПХФА. 781 с.
Щербаков А.В., Решетникова Н.М., 2017. Где искать растения-полемохоры в Смоленской области? // Изучение адвентивной и синантропной флор России и стран ближнего зарубежья: итоги, проблемы, перспективы: Мат-лы V междунар. науч. конф. (Ижевск, 6–8 сентября 2017 г.). М.; Ижевск: Ин-т комп. исследований. С. 134–137.
Щербаков А.В., Киселева Л.Л., Панасенко Н.Н., Решетникова Н.М., 2013. Растения – живые следы пребывания группы армий “Центр” на русской земле // Флора и растительность Центрального Черноземья – 2013: Мат-лы межрегион. науч. конф. (г. Курск, 6 апреля 2103 г.). Курск. С. 198–202.
Ястребов В.Н., 2019. Фортификация пехоты вермахта // Немецкие полевые оборонительные рубежи (Западный фронт 1942/43 гг.). М.: Воен. изд-во 1945. http://znamyarodini.ru/stati/stati/fortifikatciia-pekhoty-vermakhta
Ahti T., Hämet-Ahti L., 1971. Hemerophilous flora of the Kuusamo district, northeast Finland, and the adjacent part of Karelia, and its origin // Ann. Bot. Fenn. V. 8. № 1. P. 1–91.
Alm T., 2015. Plant species introduced by foreigners according to folk tradition in Norway and some other European countries: Xenophobic tales or not? // J. Ethnobiol. Ethnomed. V. 11. Art. 72.
Alm T., Piirainen M., 2000. Krigsspredte arter i Sør-Varanger, Finnmark: Knollerteknapp Lathyrus linifolius // Blyttia. V. 58. № 3–4. P. 174–177.
Alm T., Piirainen M., 2001. Krigsspredte arter i Sør-Varan-ger, Finnmark: Storengkall Rhinanthus serotinus // Blyttia. V. 59. № 2. P. 104–107.
Alm T., Edvardsen H., Elven R., 1989. Vaid (Isatis tinctoria) på Nordlandskysten — utbredelse og økologi // Blyttia. V. 47. № 1. P. 3–12.
Alm T., Piirainen M., Often A., 2000a. Krigsspredte arter i Sør-Varanger, Finnmark: Vadderot Phyteuma spicatum og svartvadderot P. nigrum // Blyttia. V. 58. № 1. P. 46–54.
Alm T., Piirainen M., Often A., 2000b. Krigsspredte arter i Sør-Varanger, Finnmark: Stjernemarikepe (Alchemilla acutiloba) — enoversett art? // Polarflokken. V. 24. P. 17–24.
Alm T., Piirainen M., Often A., 2000c. Korsknapp (Glechoma hederacea) i Nord-Norge, med noen kommentarer til funn i nabolandene // Polarflokken. V. 24. P. 49–58.
Alm T., Often A., Piirainen M., 2001a. Krigsspredte arter i Sør-Varanger, Finnmark: Flatsiv Juncus compressus, knappsiv Juncus conglomeratus oglyssiv Juncus effusus // Blyttia. V. 59. № 1. P. 7–14.
Alm T., Often A., Piirainen M., 2001b. Krigsspredte arter i Sør-Varanger, Finnmark: Hvitfrytle Luzula luzuloides – med noen kommentarer til øvrige nordnorske forekomster // Blyttia. V. 59. № 3. P. 147–151.
Alm T., Often A., Piirainen M., 2003. Engtoppklokke Campanula glomerata ssp. glomerata i Sør-Varanger, Finnmark – med noen kommentarer till toppklokkene i Nord-Norge // Blyttia. V. 61. № 1. P. 21–28.
Alm T., Piirainen M., Often A., 2009. Centaurea phrygia subsp. phrygia as a German polemochore in Sør-Varanger, NE Norway, with notes on other taxa of similar origin // Bot. Jahrb. Syst. V. 127. P. 417–432.
Berg R.Y., 1962. Nye utbredelsesdata for norske karplanter // Blyttia. V. 20. № 2. P. 49–82.
Bertelsen O.A., 1994. Russerne forhindret sult og nød // Minner fra en krig. Sør-Varanger 1940–1944 / Ed. Tannvik K. Kirkenes: Andersgrotta. P. 37–38.
Buchner A., 2001. Das Handbuch der Deutschen Infanterie 1939–1945: Gliederung, Uniformen, Bewaffnung, Ausrüstung, Einsätze. Dörfler: Nebel-Verlag. 227 S.
Bureau E., 1874. Séance du 8 Mai 1874 // Bull. Soc. Bot. France. V. 21. P. 152–168.
Fowler J.T., 2001. Axis Cavalry in World War II. Oxford: Osprey Publ. 48 p.
Franke N., 2015. Der Westwall in der Landschaft. Mainz: Ministerium für Umwelt, Landwirtschaft, Ernährung, Weinbau und Forsten Rheinland Pfaltz. 82 S.
Gaudefroy E., Mouillefarine E., 1871. Note sur des plantes méridionales observées aux environs de Paris (Florule obsidionalis) // Bull. Soc. Bot. France. V. 18. P. 246–252.
Gaudefroy E., Mouillefarine E., 1872. La florule obsidionale des environs de Paris en 1872 // Bull. Soc. Bot. France. V. 19. P. 266–277.
Gras A., 1861. Souvenirs d’uneherborisation a Verceil // Bull. Soc. Bot. France. V. 8. P. 684–689.
Jarymowycz R.J., 2008. Cavalry from Hoof to Track. Westport: Greenwood Publishing Group. 276 p.
Mannerkorpi P., 1944. Uhtuan taistelurintamalle saapuneista tulokaskasveista // Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo. V. 20. № 5. P. 39–51.
Mucina L., 1997. Conspectus of classes of the European vegetation // Folia Geobot. Phytotax. V. 32. № 2. P. 117–172.
Mucina L., Bueltmann H., Dierssen K., Theurillat J.P., Raus T. et al., 2016. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities // Appl. Veget. Sci. V. 19. Suppl. 1. P. 3–264.
Niemi B., 1969. Influence of the Soviet tenancy on the flora of the Porkala area // Acta Bot. Fenn. V. 84. P. 1–52.
Often A., 2000. Lodnefølblom (Leontodon hispidus) ogormrot (Bistorta major) krigsspredtetil Sør-Varanger (Finnmark), samtlitt om høyimporttil Kirkenes-områdetunder sistekrig // Polarflokken. V. 24. P. 187–190.
Often A., Alm T., 1997. Russehøymol (Rumex confertus) i Norge og tilgrensende strøk av Russland // Blyttia. V. 55. № 4. P. 189–199.
Piirainen M., 1997a. Briza media in Sør-Varanger, new for Finnmark // Polarflokken. V. 21. P. 273–274.
Piirainen M., 1997b. Centaurea scabiosa och några andra växtfynd vid Grense Jakobselv, Sør-Varanger // Polarflokken. V. 21. P. 279–282.
Piirainen M., 2001. Stor ormrot (Bistorta major) i Neiden, Sør-Varanger — det andra fyndet i Nordnorge // Polarflokken. V. 25. P. 199–201.
Piirainen M., 2006. Baltiskdaggkåpa (baltiskmarikåpe, Alchemilla baltica) i Sør-Varanger // Polarflokken. V. 30. P. 79–82.
Piirainen M., Alm T., 1998. Galium×pomeranicum (= G. album × verum) in Sør-Varanger, Finnmark // Polarflokken. V. 22. P. 93–97.
Piirainen M., Alm T., 2001. Syvhornmarikåpe Alchemilla heptagona Juz. Ogmånemarikåpe A. semilunaris Alechin i Sør-Varanger, Finnmark – to nyearter for Norge // Blyttia. V. 59. № 3. P. 152–161.
Piirainen M., Chkalov A., 2018. Alchemilla parcipila Juz. and A. stellaris Juz. as polemochores in Finland – the first records outside Russia – and the correct identity of A. polemochora S.E. Fröhner // Memo. Soc. Fauna Flora Fenn. V. 94. P. 67–77.
Piirainen M., Alm T., Often A., 1998. Storsyre (Rumex thyrsiflorus Fingerh.) – første funn i Finnmark // Blyttia. V. 56. № 3. P. 166–173.
POWO, 2020. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. http://www.plantsoftheworldonline.org/
Sætra H., 1972. Ny nordgrense for Scirpus silvaticus L. // Blyttia. V. 30. № 2. P. 115–116.
Stahel D., 2009. Operation Barbarossa and Germany’s Defeat in the East. Cambridge: Cambridge Univ. Press. 483 p.
Thellung A., 1917. Stratiobotanik // Vierteljahrsschr. Naturf. Ges. Zürich. V. 62. S. 327–335.
Uotila P., Suominen J., 1976. The Chenopodium species in Finland, their occurrence and means of immigration // Ann. Bot. Fenn. V. 13. № 1. P. 1–25.
Velev N., 2018. Arrhenatheretalia elatioris uncritical checklist of Europe // Phytol. Balcan. V. 24. № 1. P. 99–147.
Wearn J.A., 2016. Seeds of change – polemobotany in the study of war and culture // J. War Cult. Stud. V. 9. № 3. P. 271–284.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии