

Журнал общей биологии, 2021, T. 82, № 4, стр. 283-296
Проблемы исследования процессов аккумуляции и гипераккумуляции растениями химических элементов
Т. И. Сиромля 1, Ю. В. Загурская 2, *
1 Институт почвоведения и агрохимии СО РАН
630090 Новосибирск, пр. Академика Лаврентьева, 8/2, Россия
2 Федеральный исследовательский центр угля и углехимии СО РАН
650065 Кемерово, пр. Ленинградский, 10, Россия
* E-mail: syjil@mail.ru
Поступила в редакцию 01.02.2021
После доработки 18.02.2021
Принята к публикации 25.02.2021
Аннотация
Рассмотрены терминология, классификации и критерии выделения различных групп растений в связи с различными стратегиями накопления химических элементов (ХЭ). В отличие от неадаптированных растений (“normal plant”), устойчивость к высоким концентрациям ХЭ может определяться различными физиологическими механизмами, которые приводят к формированию основных групп: деконцентраторы (исключатели), биоиндикаторы и концентраторы (аккумуляторы). Среди последних выделяют гиперконцентраторы (гипераккумуляторы), которым уделено особое внимание в данном обзоре. Рассмотрены основания для отнесения таксонов к данной группе: пороговые значения содержания для различных ХЭ, коэффициенты концентрации и транслокации и др. Затронута проблема поверхностного загрязнения растений в зависимости от их биологических особенностей и его влияния на результаты определения содержания ХЭ. Приведены примеры ошибочной интерпретации данных об аккумуляции ХЭ растениями.
Вопросы накопления химических элементов (ХЭ) растениями весьма актуальны в настоящее время и широко рассматриваются как в России, так и за рубежом. Детальное изучение процессов аккумуляции ХЭ необходимо не только для понимания химического круговорота, но также важно и в практическом отношении. Это качество и безопасность пищевых и лекарственных растений (ЛР), поиск биоиндикаторов рудных залежей, оценка уровня антропогенного загрязнения территорий, расширение спектра растений для фиторемедиации и т.п. (Ковальский, 1974; Ковалевский, 1991; Bargagli, 1998; Pollard et al., 2002; Kabata-Pendias, 2010; Гравель и др., 2012; Ловкова и др., 2014; Уфимцева, 2015, и др.). Столь широкий спектр вопросов, связанных с концентрацией ХЭ в растениях, послужил причиной возникновения различных терминологических и классификационных противоречий, требующих выявления и разрешения.
ТЕРМИНОЛОГИЯ И ПРОБЛЕМЫ КЛАССИФИКАЦИИ
В англоязычных работах общеупотребительным является термин “accumulation”, в русскоязычных статьях слова “накопление”, “аккумуляция”, “концентрация” и “концентрирование” используют в качестве синонимов, зачастую единовременно в одной публикации. В наименовании групп растений с различными особенностями аккумуляции ХЭ за рубежом широко используются термины “excluders”, “accumulators”, “indicators” (Baker, 1981). В российских работах терминология намного разнообразнее (Уфимцева, 2015): групповые и селективные (Виноградов, 1957), адаптированные и неадаптированные (Ковальский, 1974), барьерные и безбарьерные (Ковалевский, 1991), концентраторы и исключатели (Серегин и др., 2015) и т.д. Зачастую авторы делают акцент на разных классификационных признаках, что не позволяет провести их адекватное сопоставление.
Весьма логичная концептуальная схема изменения концентрации ХЭ в листьях растений в зависимости от содержания их подвижных форм соединений в почве была предложена в работе Энт с соавт. (Ent et al., 2013) (рис. 1).
Рис. 1.
Взаимосвязь концентрации металлов/металлоидов в листьях/побегах растений с доступной концентрацией металлов/металлоидов в почве (по: Ent et al., 2013). Пунктирной линией указан порог гипераккумуляции для металлов/металлоидов.

Известно, что состояние геохимической среды динамично – формы и концентрации ХЭ в почве постоянно меняются как во времени, так и в пространстве, поэтому один и тот же вид растений может оказаться в самых разнообразных геохимических условиях, где будет вынужден или приспосабливаться к этим условиям, или погибнуть (Ковалевский, 1991; Добровольский, 2003). Выделенные на рис. 1 “нормальные” растения (“normal plant”) могут переносить только низкие концентрации подвижных форм соединений ХЭ в почве. В работе В.В. Ковальского (1974) такие растения называются неадаптированными – при изменении концентраций доступных форм соединений ХЭ в почве у них возникают различные физиологические нарушения, приводящие к эндемическим заболеваниям, угнетается развитие, нарушаются генеративные функции, и популяция вымирает. Незначительное количество особей (1–3% от популяции) может выживать, и в результате постепенного длительного процесса неадаптированные физиологические формы могут перейти в адаптированные.
Среди химических элементов наибольшее внимание исследователей в настоящее время привлекают тяжелые металлы (ТМ). В литературных источниках даются различные определения терминам “тяжелые металлы”, “токсичные элементы”, “микроэлементы” и т.д. Так, например, по разным классификациям к тяжелым металлам относят элементы, относительная атомная масса которых более 40–50, плотность более 5–8 г/см3; исключают из данной группы благородные металлы, учитывают металлоиды и т.п. С экологической точки зрения при включении в эту категорию учитывают не только химические и физические свойства элементов, но и их биологическую активность, токсичность и другие свойства.
Изучение влияния ТМ на экосистемы в целом и на отдельные таксоны тесно связано не только с природными явлениями и процессами, но и с широким спектром антропогенных источников, обуславливающих загрязнение окружающей среды.
Установлено, что в основе токсического действия ТМ на живой организм лежат три основных механизма: ферментотоксическое действие, мембранотропное действие и окислительный стресс (Скугорева и др., 2016). Аналогично этому существует три способа защиты растений от данного неблагоприятного фактора: предотвращение или ограничение проникновения, хелатирование и локализация, а также работа антиоксидантной системы.
Многообразие подходов к выявлению способов адаптации растений в условиях повышенных концентраций ХЭ (в том числе ТМ) обеспечивается сочетанием этих процессов на различных этапах продвижения и различными классификационными критериями. Например, одна из систем основана на различных уровнях ответа: тканевый (обусловлен различной скоростью транспорта ХЭ из корней в побеги), внутриклеточный (связывание ХЭ в клеточной оболочке и вакуоли), межклеточный (активирование механизмов их выведения из клеток) и молекулярный (синтез ферментов, устойчивых к токсическому воздействию ХЭ) и др. (Феник и др., 1995; Серегин, 2001; Серегин, Иванов, 2001). Более интересен подход, сочетающий не только уровни продвижения ХЭ, но и способы их детоксикации (Manara, 2012).
Подробно механизмы адаптации растений к ТМ рассмотрены в работах А.Ф. Титова с соавторами (2007, 2014); выделяют следующие стратегии адаптации: “избегание” (“avoidance”, т.е. ограничение поступления ТМ в ткани) и “устойчивость” (“tolerance”, т.е. механизмы их детоксикации в растительном организме) (Regvar, Vogel-Mikuš, 2008) (рис. 2). Вероятно, реализация этих путей и их сочетание в итоге привело к возникновению адаптированных видов растений, представленных на рис. 1 тремя остальными группами. Они оказались хорошо приспособленными к высоким концентрациям подвижных форм ХЭ, однако для этого они используют разные стратегии, поэтому способность к накоплению элементов выражена у них в различной степени.
Рис. 2.
Основные механизмы адаптации растений к высоким концентрациям тяжелых металлов. Полужирным шрифтом – терминология по: Regvar, Vogel-Mikuš, 2008; обычным – по: Титов и др., 2014; курсив – таксоно- и элементоспецифичные способы. АФК – активные формы кислорода, ПОЛ – перекисное окисление липидов, СР – свободные радикалы.
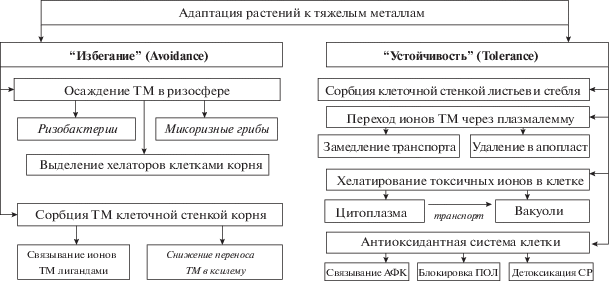
К числу адаптированных относятся прежде всего растения, длительно произрастающие в данной местности, которые в результате естественного отбора приобретают устойчивость к неблагоприятным условиям обитания. В основном это дикорастущая флора и те культурные растения, которые издавна возделываются в данном регионе.
“Исключатели”, или “деконцентраторы” (“excluders”), могут расти в широком диапазоне концентраций подвижных форм соединений ХЭ в почве до тех пор, пока уровень их накопления корректно контролируется физиологическими процессами. Нарушения в данных процессах вызывают нерегулируемое поглощение элементов, приводящее к гибели растений. М.Д. Уфимцева (2015) отмечает, что к этой группе относятся растения с четко функционирующими биогеохимическими барьерами, прежде всего в корневой системе.
“Биоиндикаторы” (“bioindicators”) поглощают ХЭ в более широком диапазоне, и концентрация элементов в листьях определяется их концентрацией в почвах до тех пор, пока не становится фитотоксической, т.е. препятствующей дальнейшей жизнедеятельности растений и приводящей к их гибели.
“Гипераккумуляторы”, или “гиперконцентраторы” (“hyperaccumulators”), способны выдерживать и накапливать гораздо более высокие концентрации ХЭ. Пунктирной линией на рис. 1 отмечены пороговые значения содержания элементов (мкг/г: Cd, Se, Tl – 100; Cu, Co, Cr – 300; Ni, As, Pb – 1000; Zn – 3000; Mn – 10 000) – если эти величины не достигнуты, растения относят к “концентраторам”, или “аккумуляторам” (“accumulators”) (Ent et al., 2013).
В российских монографиях и учебниках, рассматривающих вопросы биогеохимии растений (Виноградов, 1957; Ковальский, 1974; Ковалевский, 1991; Перельман, Касимов, 1999; Добровольский, 2003, и др.), используется другая терминология. Наиболее близким по смыслу к вышеуказанным определениям является выделение среди концентрирующих ХЭ растений привычных (типичных) и непривычных (нетипичных) концентраторов. И те, и другие могут быть как групповыми (одновременное накопление нескольких ХЭ), так и селективными (накопление отдельного ХЭ).
Перечисленные варианты градации адаптивных способностей растений к повышенным концентрациям химических элементов в окружающей среде чаще всего успешно сочетаются. С учетом наиболее распространенных биогеохимических критериев была составлена схема классификации высших растений на основании стратегии накопления ХЭ (рис. 3).
Рис. 3.
Схема классификации высших растений на основании стратегии накопления химических элементов. Полужирным шрифтом – группировка и терминология Энт и соавт. (Ent et al., 2013); обычным – терминология русскоязычных авторов (Виноградов, 1957; Ковальский, 1974; Ковалевский, 1991; Перельман, Касимов, 1999; Добровольский, 2003; Серегин и др., 2015; Уфимцева, 2015).
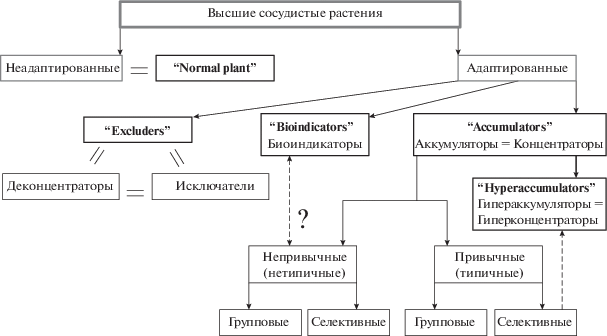
К непривычным концентраторам относится большинство растений местной флоры с широкой амплитудой выносливости к ХЭ. Растения могут произрастать как при повышенных концентрациях элементов в почве, так и при нормальном их содержании. В нормальных условиях из необогащенной почвы растения не извлекают больших количеств элемента. При расселении в зоны, более обогащенные ХЭ, непривычные растения-концентраторы накапливают значительные количества ХЭ, приближаясь по степени накопления к привычным концентраторам. Таким образом, термины “биоиндикатор” и “непривычный (нетипичный) концентратор” весьма близки между собой.
Привычные (типичные) концентраторы, как правило, всегда извлекают значительные количества ХЭ из среды, даже при условии его среднего, нормального содержания в почве. Это связано с тем, что у растений уже выработался особый обмен веществ, обусловленный генетически закрепленной потребностью соответствующих видов в конкретных элементах. Среди них выделяется группа “гиперконцентраторов” – растений с крайне выраженной адаптацией к очень высоким концентрациям ХЭ в среде обитания.
РАСТЕНИЯ-КОНЦЕНТРАТОРЫ (АККУМУЛЯТОРЫ)
Термины “концентратор” и особенно “гиперконцентратор” крайне широко и свободно трактуются и используются разными исследователями. Так, например, А.П. Виноградов (1957) выделял два типа организмов-концентраторов: 1) обитающие в областях с повышенным содержанием какого-либо ХЭ и имеющие повышенное его содержание; 2) накапливающие какой-либо ХЭ в любой среде. А.Л. Ковалевский (1991) предложил сравнивать относительное содержание химических элементов в видах растений (ОСВР), растущих в сопоставимых условиях, и относить к концентраторам те виды, для которых ОСВР превышало 2.5. Использовались также различные биогеохимические коэффициенты. А.И. Перельман и Н.С. Касимов (1999) указывали, что ХЭ накапливаются в растении, если коэффициент биологического поглощения Ах (он же КБП, Кб) – отношение концентрации ХЭ в золе растений к его валовому содержанию в почве – больше 1. М.Я. Ловкова и соавторы (2014) отнесли к концентраторам растения, для которых коэффициент биологического накопления КБН (он же коэффициент биогеохимической подвижности Вх) – отношение концентрации ХЭ в сухом веществе растений к содержанию его подвижных форм в почве – выше 1. При невозможности рассчитать данный коэффициент они считали концентраторами растения, содержание ХЭ в которых было выше кларковых значений в несколько раз, а в гиперконцентраторах – на порядок и более.
Аналогичные подходы использовали и в зарубежной литературе. Маркерт (Markert, 1992) предложил сравнивать содержание ХЭ с элементным составом “обобщенного стандартного растения” (Reference Plant, RP), позднее Данн (Dunn, 2007) несколько уточнил рекомендованные значения. В монографии Баргальи (Bargagli, 1998) приводится коэффициент концентрации (concentration factor, CF). В многочисленных статьях используются термины-синонимы – “коэффициент обогащения” (enrichment factor) (Branquinho et al., 2007), “коэффициент биоаккумуляции” (bioaccumulation factor) (Kovacik et al., 2012), “коэффицент переноса” (transfer factor) (Overesch et al., 2007) и т.п. В общем виде коэффициент рассчитывается как отношение концентрации ХЭ в надземной части растений к его концентрации в почве. Однако авторы используют содержание ХЭ как в сухой, так и в сырой биомассе, и не только их общее количество в почве, но и различные подвижные формы, что значительно затрудняет сравнительный анализ литературных данных. Рейманн и соавторы (Reimann et al., 2001) указывают, что данный показатель необходимо применять с осторожностью, поскольку использование валового содержания ХЭ в почве или подвижных форм соединений ХЭ, извлекаемых различными экстрагентами, приведет к совершенно разным результатам. Отмечается также, что из-за высокого варьирования элементного химического состава почв и растений расчет различных коэффициентов, основанных на содержании ХЭ, не может быть точным и позволяет сделать лишь общие выводы (Zeiner et al., 2015).
РАСТЕНИЯ-ГИПЕРКОНЦЕНТРАТОРЫ (ГИПЕРАККУМУЛЯТОРЫ)
В обзоре Энт с соавторами (Ent et al., 2013) указано, что с середины 1970-х годов термин “гиперконцентратор” использовался миллионы раз тысячами людей, с разной степенью точности, уместности и понимания, что не всегда соответствовало воззрениям создателей терминологии и нынешних авторов. В связи с этим необходимо уточнить обстоятельства, при которых термин “гиперконцентратор” является подходящим, и определить условия, которые должны соблюдаться при использовании данного термина.
Термин “hyperaccumulators” был использован при описании накопления никеля в работе Жаффре с соавт. (Jaffré et al., 1976). В дальнейшем он применялся к растениям, которые накапливали более 1000 мкг/г Ni в сухих листьях (Baker, 1981). Эта пороговая величина в 100–1000 больше, чем обычно встречается у растений на почвах не ультрамафического происхождения, и в 10–100 раз выше, чем у большинства других растений на богатых никелем ультрамафических почвах. Кроме того, данная величина разделяет две моды бимодальной частотной кривой – более или менее логнормально распределенные концентрации до 1000 мкг/г и отдаленный кластер при исключительно высоких концентрациях (Brooks et al., 1979). Из-за относительной редкости гипераккумуляторов других ХЭ, кроме никеля, даже высокий уровень накопления элементов зачастую не позволяет уточнить, образуют ли гипераккумуляторы качественно отличную группу (бимодальная структура) или являются хвостом положительно искаженного непрерывного (ло-гнормального) распределения (Broadley et al., 2001).
Позднее было дано более точное определение (Reeves, 1992): “гипераккумулятор никеля – это растение, в котором концентрация Ni не менее 1000 мкг/г была зарегистрирована в сухом веществе любой надземной ткани по крайней мере в одном экземпляре, растущем в его естественной среде обитания”. Таким образом, использование данного термина неуместно при анализе подземных органов или всего растения полностью. Во-первых, это вызвано сложностью отбора и подготовки образцов, не загрязненных почвенными частицами. Во-вторых, иммобилизация металлов в корневой системе является весьма распространенным явлением, в то время как активное накопление ХЭ в надземных тканях растений делает диагностику гипераккумуляторов более точной (Baker et al., 1994). В дальнейшем было уточнено (Ent et al., 2013):
– для установления статуса гипераккумулятора необходимо использовать листья растений;
– “аккумуляция” должна подразумевать только активное накопление элементов внутри ткани листа растения через корни;
– не следует рассматривать пассивное накопление элементов через атмосферное осаждение на листьях растений.
Дополнительно следует акцентировать внимание на окончании вышеприведенного определения: “… растет в естественной среде обитания” – гиперконцентраторы должны накапливать высокие концентрации металлов, оставаясь при этом достаточно здоровыми для сохранения самоподдерживающейся популяции. В работе Бейкера и Уайтинга (Baker, Whiting, 2002) указано, что зачастую о новых видах растений-гипераккумуляторов ошибочно сообщают по результатам экспериментальных исследований в искусственных условиях, при внесении высоких доз солей тяжелых металлов в почву или питательный раствор. Однако в подобных случаях эксперименты практически никогда не доходят до того, чтобы продемонстрировать образование жизнеспособных семян. Более того, “вынужденное” или “индуцированное” поглощение металлов часто приводит в конечном итоге к гибели растений. Хотя это и представляет интерес для некоторых стратегий фиторемедиации, но может не иметь никакого отношения к продолжающемуся жизненному циклу естественных популяций металлофитов.
Среди гипераккумуляторов выделяют “obligate” (“обязательные”, “строгие”) и “facultative” (“необязательные”) виды (Pollard et al., 2002). Облигатные гипераккумуляторы эндемичны для некоторых типов металлоносных почв и всегда накапливают высокие концентрации элемента. Факультативные гипераккумуляторы являются представителями популяций, в которых лишь некоторые экземпляры растений являются гипераккумуляторами. В обзоре Полларда с соавторами (Pollard et al., 2014), посвященном вопросам гипераккумуляции, отмечено, что, несмотря на попытки понять общие закономерности внутривидовых вариаций, до сих пор существуют виды, поведение которых парадоксально. Такие виды имеют широкие географические ареалы и адаптируются к различным экологическим условиям, в том числе и к “нормальным”, и к “серпентиновым” почвам, однако как гипераккумуляторы проявляют себя лишь в небольшом количестве мест на каждой из них. Предполагается, что факультативная гипераккумуляция может быть связана как с генетическими отличиями между отдельными популяциями вида, так и с разницей в содержании и доступности ХЭ, находящихся в почвах.
КРИТЕРИИ ГИПЕРАККУМУЛЯЦИИ
В настоящее время к гипераккумуляторам относят растения, надземная часть которых содержит ХЭ в количестве (мкг/г сухого вещества): более 100 для Cd, Se и Tl; 300 для Co, Cr и Cu; 1000 для As, Ni и Pb; 3000 для Zn и 10000 для Mn – концентрация должна быть на 2–3 порядка выше, чем в надземной части большинства видов на нормальных почвах, и по крайней мере на один порядок больше, чем в обычном диапазоне, обнаруженном в растениях из металлоносных почв (Ent et al., 2013). Тем не менее номинальные пороговые значения не должны считаться “истиной в последней инстанции” – так, например, растения, проявляющие экстремальное физиологические поведение и стабильно накапливающие 900 мкг/г Ni, вполне могут рассматриваться как гипераккумуляторы этого металла. В настоящее время критерии разработаны для 11 элементов, однако некоторые из них уже подвергаются сомнениям. Слишком высоким считают критерий гипераккумуляции для Cu (Faucon et al., 2007). Крэмер (Krämer, 2010) рекомендовал снизить критерии для Cu и Co, а Энт и соавторы (Ent et al., 2013) поддержали это предложение.
Кроме номинальных пороговых значений, существуют и другие критерии, определяющие гипераккумуляцию. В частности, гипераккумуляторы имеют очень высокий коэффициент концентрации (см. выше – отношение содержания ХЭ в надземной части растений к его содержанию в почве), так как их физиологические особенности обеспечивают активное связывание металлов и увеличение их концентрации. Хоббс и Штрайт (Hobbs, Streit, 1986) предложили считать данный коэффициент одним из показателей гипераккумуляции. Однако позднее было показано, что для определения гипераккумуляции на основе собранного в полевых условиях материала использования только коэффициента концентрации недостаточно (Pollard et al., 2002). Данный коэффициент проявляется как взаимодействие генотипа с окружающей средой и контролируется как генетически детерминированной физиологией растений, так и локальными эдафическими условиями. В связи с этим между содержанием элемента в листьях и почвах нет однозначной связи (Pollard et al., 2002). Кроме того, было установлено, что при низкой концентрации элемента в почве растение интенсивно поглощает имеющееся количество доступных для него форм этого элемента, и только в случае высокого содержания ХЭ в почве начинают срабатывать механизмы регуляции (Bargagli, 1998; Kabata-Pendias, 2010). Осложняющим фактором является то, что ни один химический экстрагент универсально и точно не извлекает доступную для растений фракцию содержащегося в почве металла (Meers et al., 2007; Menzies et al., 2007).
Еще одним критерием, предлагаемым для определения гипераккумуляции, является коэффициент транслокации (“translocation factor”) – в растениях-гипераккумуляторах содержание металлов в корнях ниже по сравнению с надземной частью (Macnair, 2003). Необходимо отметить, что при оценке соотношения концентраций элементов в подземной и надземной частях растений в литературе встречаются различные синонимичные термины. Это коэффициент передвижения (КП) (Ковалевский, 1991), коэффициент корневого барьера (ККБ, Ккб) (Практикум по агрохимии, 2001; Афанасьева, Аюшина, 2018), акропетальный коэффициент (АК) (Сибгатуллина и др., 2014), коэффициент перехода (КП) (Жуйкова, Зиннатова, 2014), коэффициент транслокации (translocation factor, TF) (Branquinho et al., 2007), коэффициент переноса (transfer factor, TF) (Lajayer et al., 2017) и др. При этом рассчитывают как отношение ХЭ в подземной части к надземной, так и наоборот, что приводит к дополнительным противоречиям. Высказываются предположения, что пониженное количество многих ХЭ в надземных частях растений может быть связано как с невысокой потребностью в них, так и с существованием механизмов, регулирующих их накопление и распределение между корнями и надземными органами (Seregin, Kozhevnikova, 2008). Однако низкое содержание доступных форм ХЭ в почве может приводить к более интенсивному поглощению элементов растениями, а эффект “корневого барьера” проявляется лишь при их высоких концентрациях (Bargagli, 1998; Kabata-Pendias, 2010). Использование коэффициента транслокации для доказательства гипераккумуляции также не является достаточным по различным причинам: 1) трудность корректного извлечения корневой массы растений, особенно древесных; 2) трудность полной очистки корней от почвенных частиц; 3) пересчет концентраций ХЭ, содержащихся в различных органах растения, на сухое вещество может быть некорректным, так как зависит от доли в них структурного материала; 4) высокое содержание ХЭ в надземной части может зависеть от внешнего источника поступления и т.д. (Ent et al., 2013).
Высокие значения коэффициентов концентрации и транслокации являются необходимыми, но недостаточными критериями гипераккумуляции, поскольку характерны и для видов-концентраторов (Wei et al., 2008), поэтому номинальные пороговые критерии, несмотря на ряд недостатков и узкий спектр элементов, считаются более обязательным условием для выявления гипераккумуляторов.
В вопросах накопления и детоксикации ХЭ серьезное значение также имеют ткани, в которых локализуется элемент (Isaure et al., 2006; Seregin, Kozhevnikova, 2008; Титов и др., 2014) – для растений-гиперконцентраторов характерно поступление элемента в мезофилл, тогда как в иных случаях скопления наблюдаются только в эпидермальных структурах (Kupper et al., 2000; Choi et al., 2001; Ma et al., 2005).
ПОВЕРХНОСТНОЕ ЗАГРЯЗНЕНИЕ РАСТИТЕЛЬНОГО МАТЕРИАЛА
Определение источника поступления ХЭ в растения является серьезной методической проблемой. Фолиарный путь поступления ХЭ в растения может играть существенную роль в условиях антропогенного загрязнения атмосферы, в том числе мелкодисперсными почвенными частицами (Amato et al., 2009; Копылова, 2013), обогащенными ХЭ (Uzu et al., 2010). Содержание некоторых элементов в этих частицах может быть на порядок выше, чем в почве в целом (Siromlya et al., 2015). В подобных случаях высокие концентрации целого ряда ХЭ в надземной части растений являются результатом их пассивного накопления через атмосферное осаждение. При этом фактически ХЭ не накапливаются в растениях, а содержатся в пылевых частицах на их поверхности (Загурская, Сиромля, 2018).
Пылеудерживающая способность растений зависит не только от типа, размера и количества частиц, но и множества биологических особенностей таксона. Важную роль играют как макроструктурные особенности (высота растений, размер, форма и расположение листьев и т.п.), так и микроструктурные особенности (шероховатость поверхности, различные типы опушения, складчатость, специфические органы выделения, наличие воскового слоя, размер и количество устьиц и т.п.) (Латышевская, Стрекалова, 2006; Sæbø et al., 2012; Leonard et al., 2016, и др.). Так, например, Л.В. Куринская с соавторами (2012) именно архитектоникой листа и площадью ассимиляционного аппарата объясняют наибольшее по сравнению с другими исследуемыми видами содержание Pb в морщинистых листьях Lolium perenne L. В работе Т.А. Корельской и Л.Ф. Поповой (2012) указано, что содержание Zn, Cu и Pb в пылевых частицах, осевших и сорбированных на поверхности растений, увеличивает содержание металлов в листьях и траве более чем на 30%.
Энт и соавторы (Ent et al., 2013) подвергают сомнению результаты ряда опубликованных ранее работ о выявлении новых видов растений-гиперконцентраторов именно из-за возможного загрязнения растительной поверхности частицами атмосферных аэрозолей, содержащими значительные количества ТМ. При этом указывается на необходимость их подтверждения с использованием образцов, обработанных для удаления поверхностного загрязнения. Так, например, большинство видов растений-гипераккумуляторов меди, которые были описаны в Демократической Республике Конго (Brooks et al., 1982), не проявили себя таковыми в более позднем исследовании, в котором использовалась интенсивная промывка растительного листового материала (Faucon et al., 2007). Ученые обнаружили, что 12 видов, которые были ранее изучены и зарегистрированы как гиперконцентраторы меди, действительно имели высокие концентрации данного элемента, но редко превышали номинальные пороговые значения. Они пришли к выводу, что большое изменение концентраций элемента в растениях в пределах одного участка, значительная линейная корреляция между его содержанием в почвах и растениях (характерная для биоиндикаторов) и относительно низкая концентрация меди во многих образцах нехарактерны для гиперконцентраторов, а также предположили, что большинство проанализированных ранее проб было загрязнено пылью. Так, например, добавление 0.2 мг малахита в виде пыли к 100 мг листьев растений, содержащих 10 мкг/г меди, достаточно для повышения кажущейся концентрации Cu более чем на 1.150 мкг/г (Reeves, Baker, 2000). На этом же основании Энт и соавторы (Ent et al., 2013) также подвергают сомнению примеры гипераккумуляции хрома, описанные в ранее опубликованных работах (Zhang et al., 2007; Redondo-Gomez et al., 2011), объясняя подобные явления пассивным накоплением элемента путем осаждения из воздуха на листьях растений.
В условиях юго-востока Западной Сибири влияние поверхностного загрязнения на элементный химический состав растений проявилось при исследовании Leonurus quinquelobatus Gilib. (Загурская, Сиромля, 2018), а также Artemisia sieversiana Willd. и Urtica cannabina L. (Сиромля, 2019). Было установлено, что существенная часть образцов не соответствовала требованиям Государственной Фармакопеи РФ (2018) по общей зольности и остатку золы, нерастворимой в 10% HCl. Последний показатель позволяет оценить долю примесей преимущественно минерального происхождения, т.е. фактически отражает запыленность растений. При сравнении концентрации ХЭ в растительных образцах, соответствующих (“чистых”) и несоответствующих (“запыленных”) вышеуказанным требованиям, была выявлена статистически значимая разница. Также было обнаружено, что количество ХЭ в “чистых” пробах в целом соответствовало верхней части диапазона их содержания в растениях остальных исследованных видов. Подобное явление объясняется специфическими анатомо-морфологическими особенностями вышеуказанных видов – эти растения густо опушены длинными мягкими волосками (Конспект флоры…, 2005). В работе Минкиной с соавторами (Minkina et al., 2017) также показано, что в условиях техногенного загрязнения наибольшие количества ХЭ накапливаются в надземной части опушенных видов растений – Ambrosia artemisiifolia L., Artemisia austriaca Jack. В.Д. Белоногова (2009) отмечает, что среди многочисленных исследованных видов лекарственного растительного сырья повышенное содержание золы общей, нерастворимой в 10% HCl и тяжелых металлов характерно для листьев крапивы (Urtica dioica L.) и мать-и-мачехи (Tussilago farfara L.). Тот факт, что опушенные виды растений концентрируют ХЭ в большей степени, чем менее опушенные, отмечен и в других публикациях (Латышевская, Стрекалова, 2006; Leonard et al., 2016).
Интересно также обратить внимание, что у A. sieversiana и U. cannabina содержание многих исследованных ХЭ в подземной части оказалось ниже, чем в надземной части. То есть значение коэффициента корневого барьера менее 1 в данном случае указывало не на концентрирование ХЭ в надземной части растений, а на их повышенную запыленность и внекорневое поступление значительной доли этих элементов, что также отмечалось и в литературе (Ковалевский, 1991; Ent et al., 2013; Копылова, 2013).
Кроме высокого содержания золы и остатка, нерастворимого в 10% HCl, запыленность растений можно оценивать по содержанию Si (Загурская, Сиромля, 2018) и хром/никелевому соотношению (Сысо, 1998). Рейманн и соавторы (Reimann et al., 2001) указывают, что в качестве первого признака возможного влияния минерагенной пыли на наблюдаемую концентрацию элементов в растениях могут быть приняты высокие значения Al и Fe. Учитывая проблемы обнаружения ряда элементов у некоторых видов (например, Cr, Li, Sc и Th), лучшими показателями запыленности растительного материала являются Al, Fe, Si, V, Y и Zr. Баргальи (Bargagli, 1998) для оценки загрязнения растений почвенными частицами рекомендовал определить в растительных пробах концентрации элементов-индикаторов почвы (Al, Ti, Pu, Sc, Zr, Fe, Si) вместе с интересующими элементами. Затем необходимо рассчитать коэффициент обогащения (EF): концентрация исследуемого элемента в растении отнесена к концентрации Al или Ti в этом же растении, а это отношение, в свою очередь, разделено на соответствующее отношение этих элементов в почве или земной коре. Как правило, чем ближе EF к единице, тем более вероятно почвенное происхождение рассматриваемого элемента, а значения намного выше единицы являются показателем атмосферного загрязнения.
ПРОТИВОРЕЧИЯ ПРИ ВЫЯВЛЕНИИ ВИДОВ-КОНЦЕНТРАТОРОВ
Анализ литературных данных показывает, что использование различными исследователями разных критериев аккумуляции приводит к противоречивым результатам.
Так, в работе Масаровиковой с соавторами (Masarovicova et al., 2010) сообщается о непоследовательной классификации Matricaria chamomilla L. (син. M. recutita L.), которую разные авторы рассматривают и как гипераккумулятор, и как факультативный металлофил, и как вид-исключатель, что объясняется использованием различных параметров и их разнообразием. Несколько позднее Ковачик (Kovacik, 2013) обратил внимание на то, что бессмысленно делать выводы об аккумуляционном потенциале растений при выращивании их в условиях гидропоники. Он также отметил, что нельзя ориентироваться только на величины коэффициентов концентрации и транслокации, не принимая во внимание уменьшение биомассы растений и т.п., а также согласился с рекомендациями (Ent et al., 2013) о необходимости доказательств аккумуляции Cd растениями M. chamomilla, произрастающими на естественно загрязненной почве.
В статье М.Я. Ловковой и Г.Н. Бузук (2013) приведены результаты массового обследования (около 200 видов) лекарственных растений флоры России, из которых к видам-концентраторам отнесено более 80%. При этом основанием для подобных заключений является сравнение содержания ХЭ в ЛР со средними кларковыми величинами. В более ранней публикации Ловковой и Бузук (Lovkova, Buzuk, 2011) указано, что значения кларка меди (5 мг/кг) взяты из монографии A. Кабата-Пендиас и Х. Пендиас (1989), но в данной монографии нет значений кларков, а приводятся диапазоны содержания Cu в зрелых тканях листьев: 2–5 мг/кг – дефицит, 5–30 мг/кг – нормальная концентрация. Ориентируясь на эти значения, можно отметить, что практически все исследованные виды ЛР укладываются в диапазон нормальных концентраций – содержание меди в них составляет 15.5–27.0 мг/кг, однако авторы относят их к концентраторам. К сверхконцентраторам отнесены кубышка желтая – Nuphar lutea L. (33.3 мг/кг), лобелия вздутая – Lobelia inflata L. (41.1 мг/кг), анис обыкновенный – Pimpinella anisum L. (43.3 мг/кг) и сушеница топяная – Gnaphalium uliginosum L. (105.1 мг/кг) (причем у кубышки исследовались корневища, а у аниса – плоды). Однако содержание в них меди намного ниже порогового значения для гиперконцентраторов – 300 мг/кг, предлагаемого в работе Энт с соавт. (Ent et al., 2013).
Данные о концентрировании ЛР других ХЭ (Ловкова, Бузук, 2013) также вступают в значительные противоречия с масштабными исследованиями (Ent et al., 2013) и вызывают много вопросов. Так, например, в качестве кларка хрома приводится значение 0.15 мг/кг, при этом в монографии А. Кабата-Пендиас и Х. Пендиас (1989) указан нормальный диапазон его содержания – 0.1–0.5 мг/кг, а кларковые значения – 1.8 мг/кг (Добровольский, 2003) и 1.5 мг/кг (Markert, 1992; Dunn, 2007). Вероятно, именно использование такого низкого кларкового значения – 0.15 мг/кг – является причиной того, что более половины исследованных видов ЛР отнесены к концентраторам. При этом М.Я. Ловкова и Г.Н. Бузук (2013) указывают, что среди них фактически отсутствуют умеренные накопители, а имеются лишь концентраторы и сверхконцентраторы данного элемента (к последним отнесены растения, накапливающие 6–17 мг/кг хрома). Энт и соавторы (Ent et al., 2013) приводят в качестве порогового значения величину 300 мг/кг и, как отмечено ранее, вообще подвергают сомнению гипераккумуляцию хрома, считая это влиянием запыленности. Полученные нами результаты (Загурская, Сиромля, 2018; Сиромля, 2019) также показали, что для отдельных видов растений, характеризующихся высоким уровнем запыленности в связи с анатомо-морфологическим особенностям, выявляются статистически значимые корреляционные зависимости между валовым содержанием хрома и количеством золы, нерастворимой в 10% HCl. Это вполне соответствует данным А.И. Перельмана и Н.С. Касимова (1999), которые относят хром к элементам с низким уровнем накопления.
HYPЕRICUM PERFORATUM – ГИПЕРКОНЦЕНТРАТОР Cd: ДА ИЛИ НЕТ?
В противоречие с некоторыми литературными данными вступили результаты, полученные нами при исследовании Hypеricum perforatum L. (зверобоя обыкновенного), произрастающего на юго-востоке Западной Сибири. Многие исследователи относят H. perforatum к гиперконцентраторам Cd (Schneider et al., 2002; Jurca et al., 2011; Ловкова и др., 2014; Badea, 2015). Согласно определению (Ent et al., 2013), в таком случае растения должны накапливать более 100 мг/кг Cd. Однако проведенные исследования (Сиромля, 2019; Загурская, Сиромля, 2020) выявили, что повышенные концентрации Cd – до 1.7 мг/кг – встречаются лишь в отдельных образцах H. perforatum (как культивируемых, так и дикорастущих). В основном в регионе преобладают пробы с относительно низким содержанием этого элемента – 0.4–0.5 мг/кг. Подобные высокие внутривидовые различия в накоплении Cd отмечались и у других видов, например у Brassica napus L. (Grispen et al., 2006).
Многочисленные литературные данные показали, что в надземной части H. perforatum диапазон концентраций (мг/кг) данного элемента невелик: например, в России – 0.1–1.7, в странах Азии – менее 0.1, Америки – 0.05–0.26, Европы – 0–4.2 (большинство источников ограничено диапазоном 0.1–1) (Загурская, Сиромля, 2020).
При более подробном изучении публикаций, в которых H. perforatum называют гипераккумулятором Cd, возникают сомнения в обоснованности данного утверждения. Так, в монографии М.Я. Ловковой с соавторами (2014) указано, что H. perforatum – один из восьми видов ЛР, концентрирующих Cd, при этом содержание Cd не приводится, указан лишь коэффициент биологического накопления – 7.2. В статье Шнайдера с соавторами (Schneider et al., 2002) аналогичный коэффициент изменяется от 13 до 888. В работе Бадеа (Badea, 2015) H. perforatum относят к гипераккумуляторам на основании того, что содержание в нем Cd составило 0.13 мг/кг и оказалось выше, чем в других видах исследованных растений (0.04–0.12 мг/кг), ссылаясь при этом на более ранние публикации (Gasser et al., 2009; Đurović et al., 2013). Однако в статье Дурович с соавторами (Đurović et al., 2013) отмечается лишь высокая концентрация Cd (0.73–1.12 мг/кг), которая в 3–4 раза превышает предложенные ВОЗ значения – 0.3 мг/кг сухого вещества. А в работе Гассера с соавторами (Gasser et al., 2009) приводятся данные о том, что количество Cd в 90% исследованных проб не превышает 0.95 мг/кг, и предлагается ввести исключения для его предельно допустимого содержания в траве H. perforatum – 1.0 мг/кг по сравнению с 0.5 мг/кг в Европейской фармакопее 2008 г. Таким образом, ни одна из рассмотренных публикаций не позволяет отнести H. perforatum к гипераккумуляторам Cd. К такому же выводу приходят и авторы другой работы (Pavlova, Karadjova, 2013), указывая, что содержание Cd в образцах H. perforatum не выше, чем в произрастающих при тех же условиях растениях других видов.
Таким образом, H. perforatum не может быть отнесен к гипераккумулярам Cd – содержание данного элемента в надземной части растений не превышает 100 мг/кг. Причины относительно высоких содержаний данного ХЭ в отдельных случаях могут заключаться в том, что пробы отбирались в окрестностях промышленных предприятий (Glavač et al., 2017) или на горном склоне (Đurović et al., 2013), вследствие чего повышенное количество некоторых ХЭ может быть связано с рудопроявлениями (Гравель и др., 2012). В прочих работах (Radanovic et al., 2002; Jurca et al., 2011; Owen et al., 2016; Загурская, Сиромля, 2020) повышенные (т.е. превышающие величины предельно допустимых концентраций для лекарственного растительного сырья и биологически активных добавок на растительной основе) концентрации Cd отмечаются лишь для единичных проб.
ЗАКЛЮЧЕНИЕ
Для корректной научной коммуникации необходима унификация и стандартизация понятийного аппарата и конкретизация терминов, касающихся вопросов накопления химических элементов растениями. Обобщая многочисленные литературные сведения, можно выделить две основные группы растений: адаптированные только к низким/нормальным концентрациям подвижных форм соединений ХЭ в почвах и адаптированные к более широкому диапазону концентраций, в том числе и значительно повышенным. Использование различными исследователями разных критериев аккумуляции зачастую приводит к противоречивым результатам. Для определения видов-концентраторов используются следующие подходы: 1) сравнение содержания ХЭ в растениях с их содержанием в других видах, с кларками, с “обобщенным стандартным растением” и т.п.; 2) различные биогеохимические коэффициенты. Однако они не противоречат друг другу и становятся гораздо информативнее при совместном применении.
Необходимо подчеркнуть, что для достоверного выявления факта аккумуляции ХЭ требуется использовать листья растений, произрастающих в естественной среде обитания и являющихся достаточно здоровыми для сохранения самоподдерживающейся популяции. При этом нужно учитывать только активное накопление элемента внутри ткани листа растения через корни, пассивное накопление через атмосферное осаждение рассматривать при этом не следует. Запыленность растительного материала можно оценить по общей зольности, количеству нерастворимого в 10% HCl остатка золы, хром/никелевому соотношению, коэффициенту обогащения, содержанию Si, Al, Fe и т.д.
Биогеохимические коэффициенты используются и при выявлении видов-гиперконцентраторов, однако первоначальным условием является превышение номинальных пороговых значений содержания ХЭ. К сожалению, в настоящее время данные значения установлены только для 11 элементов, причем зачастую эти границы подвергаются пересмотру. Во всех остальных случаях о гипераккумуляции имеет смысл говорить лишь тогда, когда концентрация элемента в растении на 2–3 порядка выше, чем в надземной части большинства видов на нормальных почвах, и по крайней мере на один порядок больше, чем в обычном диапазоне, обнаруженном в растениях из металлоносных почв.
Таким образом, использование разностороннего и многопланового подхода к оценке аккумуляционного потенциала растений поможет избавиться от большинства существующих в настоящее время противоречий, связанных с выявлением стратегий адаптации растений к высоким концентрациям ХЭ.
Работа выполнена в рамках проектов ГЗ ИПА СО РАН и ГЗ отдела Кузбасский ботанический сад ФИЦ УУХ СО РАН (№ 0286-2021-0010, НИОКТР № АААА-А21-121011590010-5).
Список литературы
Афанасьева Л.В., Аюшина Т.А., 2018. Накопление и распределение микроэлементов в растениях Arctostaphylos uva-ursi // Химия растит. сырья. № 3. С. 123–128.
Белоногова В.Д., 2009. Ресурсы, экологическая безопасность и фитохимические исследования дикорастущих лекарственных растений Пермского края. Автореф. дис. … д-ра фарм. наук. Пермь: Пермская ГФА. 40 с.
Виноградов А.П., 1957. Геохимия редких и рассеянных химических элементов в почвах. М.: Изд-во АН СССР. 298 с.
Государственная фармакопея Российской Федерации, 2018. XIV изд. T. II. М.: Минздрав РФ. 3263 с.
Гравель И.В., Шойхет Я.Н., Яковлев Г.Н., Самылина И.А., 2012. Фармакогонозия. Экотоксиканты в лекарственном растительном сырье и фитопрепаратах. М.: ГЭОТАР-Медиа. 304 с.
Добровольский В.В., 2003. Основы биогеохимии. М.: Изд. центр “Академия”. 400 с.
Жуйкова Т.В., Зиннатова Э.Р., 2014. Аккумулирующая способность растений в условиях техногенного загрязнения почв тяжeлыми металлами // Поволжский экол. журн. № 2. С. 196–207.
Загурская Ю.В., Сиромля Т.И., 2018. Особенности элементного химического состава растений Leonurus quinquelobatus (на примере Западной Сибири) // Учен. зап. Казан. ун-та. Сер. Естеств. науки. Т. 160. Кн. 3. С. 419–435.
Загурская Ю.В., Сиромля Т.И., 2020. Химический элементный состав Hypericum perforatum – нормируемые химические элементы (As, Cd, Hg, Pb) // Химия растит. сырья. № 3. С. 163–170.
Кабата-Пендиас А., Пендиас X., 1989. Микроэлементы в почвах и растениях. Пер. с англ. М.: Мир. 439 с.
Ковалевский А.Л., 1991. Биогеохимия растений. Новосибирск: Наука. 294 с.
Ковальский В.В., 1974. Геохимическая экология. М.: Наука. 302 с.
Конспект флоры Сибири: Сосудистые растения, 2005. Новосибирск: Наука. 362 с.
Копылова Л.В., 2013. Фолиарное поступление тяжелых металлов в древесные растения // Вестн. КрасГАУ. № 12. С. 126–133.
Корельская Т.А., Попова Л.Ф., 2012. Тяжелые металлы в почвенно-растительном покрове селитебного ландшафта г. Архангельска // Арктика и Север. № 7. С. 136–152.
Куринская Л.В., Колесников С.И., Куринская Н.В., 2012. Оценка устойчивости газонных трав к загрязнению почв свинцом на примере придорожных ландшафтов. Saarbrucken: Lap Lambert. 163 с.
Латышевская Н.И., Стрекалова А.С., 2006. Экологические проблемы заготовки лекарственного растительного сырья в рекреационной зоне промышленного центра // Вестн. ОГУ. Приложение “Химия и физические методы, биология, здравоохранение, биоэлементология”. № 5 (55). С. 192–194.
Ловкова М.Я., Бузук Г.Н., 2013. Лекарственные растения – концентраторы микроэлементов. Новые аспекты применения этих видов в медицине // Вопросы биол., мед. и фарм. химии. Т. 11. № 4. С. 43–49.
Ловкова М.Я., Рабинович А.М., Пономарева С.М., Бузук Г.Н., Соколова С.Н., 2014. Почему растения лечат. М.: ЛЕНАНД. 288 с.
Перельман А.И., Касимов Н.С., 1999. Геохимия ландшафта. М.: Астрея-2000. 610 с.
Практикум по агрохимии, 2001 / Под ред. Минеева В.Г. М.: Изд-во МГУ. 689 с.
Серегин И.В., 2001. Фитохелатины и их роль в детоксикации кадмия у высших растений // Успехи биол. химии. Т. 41. С. 283–300.
Серегин И.В., Иванов В.Б., 2001. Физиологические аспекты токсического действия кадмия и свинца на высшие растения // Физиол. растений. Т. 48. С. 461–485.
Серегин И.В., Кожевникова А.Д., Жуковская Н.В., Схат Х., 2015. Устойчивость и накопление кадмия исключателем Thlaspi arvense и различными экотипами гипераккумулятора Noccaea caerulescens // Физиол. растений. Т. 62. № 6. С. 854–864.
Сибгатуллина М.Ш., Александрова А.Б., Иванов Д.В., Валиев В.С., 2014. Оценка биогеохимического состояния травянистых растений и почв Волжско-Камского заповедника // Учен. зап. Казан. ун-та. Сер. Естеств. науки. Т. 156. Кн. 2. С. 87–102.
Сиромля Т.И., 2019. Химические элементы в растениях – фолиарный путь поступления // Тр. XI Междунар. биогеохим. шк. “Биогеохимия – научная основа устойчивого развития и сохранения здоровья человека”. Тула: Тульский ГПУ. С. 262–266.
Скугорева С.Г., Ашихмина Т.Я., Фокина А.И., Лялина Е.И., 2016. Химические основы токсического действия тяжелых металлов (обзор) // Теор. и прикл. экология. № 1. С. 4–13.
Сысо А.И., 1998. Использование отношения Cr : Ni в мониторинге загрязнения природной среды // Агрохимия. № 4. С. 76–83.
Титов А.Ф., Казнина Н.М., Таланова В.В., 2014. Тяжелые металлы и растения. Петрозаводск: Изд-во КарНЦ РАН. 194 с.
Титов А.Ф., Таланова В.В., Казнина Н.М., Лайдинен Г.Ф., 2007. Устойчивость растений к тяжелым металлам. Петрозаводск: Изд-во КарНЦ РАН. 172 с.
Уфимцева М.Д., 2015. Закономерности накопления химических элементов высшими растениями и их реакции в аномальных биогеохимических провинциях // Геохимия. № 5. С. 450–465.
Феник С.И., Трофимяк Т.Б., Блюм Я.Б., 1995. Механизмы формирования устойчивости растений к тяжелым металлам // Успехи соврем. биологии. Т. 115. С. 261–275.
Amato F., Pandolfi M., Viana M., Querol X., Alastuey A., Moreno T., 2009. Spatial and chemical patterns of PM10 in road dust deposited in urban environment // Atmos. Environ. V. 43. P. 1650–1659.
Badea D.N., 2015. Determination of potentially toxic heavy metals (Pb, Hg, Cd) in popular medicinal herbs in the coal power plant area // Rev. Chimie. V. 66. № 8. P. 1132–1136.
Baker A.J.M., 1981. Accumulators and excluders: Strategies in the response of plants to heavy metals // J. Plant Nutr. V. 3. P. 643–654.
Baker A.J.M., Whiting S.N., 2002. In search of the Holy Grail – a further step in understanding metal hyperacumulation // New Phytol. V. 155. P. 1–7.
Baker A.J.M., Reeves R.D., Hajar A.S.M., 1994. Heavy metal accumulation and tolerance in British populations of the metallophytes Thlaspi caerulescens J. and C. Presl (Brassicaceae) // New Phytol. V. 127. № 1. P. 61–68.
Bargagli R., 1998. Trace Elements in Terrestrial Plants: An Ecophysiological Approach to Biomonitoring and Biorecovery. Berlin: Springer-Verlag. 324 p.
Branquinho C., Serrano H.C., Pinto M.J., Martins-Louçao M.A., 2007. Revisiting the plant hyperaccumulation criteria to rare plants and earth abundant elements // Environ. Pollut. V. 146. P. 437–443.
Broadley M.R., Willey N.J., Wilkins J.C., Baker A.J.M., Mead A., White P.J., 2001. Phylogenetic variation in heavy metal accumulation in angiosperms // New Phytol. V. 152. № 1. P. 9–27.
Brooks R.R., Grégoire J., Madi L., Malaisse F., 1982. Phytogéochimie des gisementscupro-cobaltifères de l’anticlinal de Kasonta (Shaba-Zaïre) // Géo-Eco-Trop. V. 6. P. 219–228.
Brooks R.R., Morrison R.S., Reeves R.D., Dudley T.R., Akman Y., 1979. Hyperaccumulation of nickel by Alyssum Linnaeus (Cruciferae) // Proc. Roy. Soc. Lond. B. V. 203. № 1153. P. 387–403.
Choi Y., Harada E., Wada M., Tsuboi H., Morita Y. et al., 2001. Detoxification of cadmium in tobacco plants: Formation and active excretion of crystals containing cadmium and calcium through trichomes // Planta. V. 213. № 1. P. 45–50.
Dunn C.E., 2007. New perspectives on biogeochemical exploration. Paper 12. Advances in prospect-scale geochemical methods // Fifth decennial int. conf. on mineral exploration: Proc. of Exploration 07 conf. Toronto: Decennial Mineral Exploration Conferences. P. 249–261.
Đurović D., Bulat Z., Buha A., Matovic V., 2013. Cadmium, mercury and lead in Hypericum perforatum L. collected in Western Serbia // E3S Web of Conf. V. 1. https://doi.org/10.1051/e3sconf/20130115009
Ent A., van der, Baker A.J.M., Reeves R.D., Pollard J.A., Schat H., 2013. Hyperaccumulators of metal and metalloid trace elements: facts and fiction // Plant Soil. V. 362. № 1–2. P. 319–334.
Faucon M.-P., Shutcha M.N., Meerts P., 2007. Revisiting copper and cobalt concentrations in supposed hyperaccumulators from SC Africa: Influence of washing and metal concentrations in soil // Plant Soil. V. 301. P. 29–36.
Gasser U., Klier B., Kühn A.V., Steinhoff B., 2009. Current findings on the heavy metal content in herbal drugs // Pharmeur. Sci. Notes. V. 1. P. 37–49.
Glavač N.K., Djogo S., Ražić S., Kreft S., Veber M., 2017. Accumulation of heavy metals from soil in medicinal plants // Arch. Industr. Hyg. Toxicol. V. 68. № 3. P. 236–244.
Grispen V.M.J., Nelissen H.J.M., Verkleij J.A.C., 2006. Phytoextraction with Brassica napus L.: A tool for sustainable management of heavy metal contaminated soils // Environ. Pollut. V. 144. P. 77–83.
Hobbs R.H., Streit B., 1986. Heavy metal concentrations in plants growing on a copper mine spoil in the Grand Canyon, Arizona // Am. Midl. Nat. V. 115. № 2. P. 277–281.
Isaure M.-P., Fayard B., Sarret G., Pairis S., Bourguignon J., 2006. Localization and chemical forms of cadmium in plant samples by combining analytical electron microscopy and X-ray spectromicroscopy // Spectrochim. Acta B. V. 61. № 12. P. 1242–1252.
Jaffré T., Brooks R.R., Lee J., Reeves R.D., 1976. Sebertia acuminata: A hyperaccumulator of nickel from New Caledonia // Science. V. 193. № 4253. P. 579–580.
Jurca T., Marian E., Vicas L., Gatea D., 2011. Simultaneous determination of metals in Hypericum perforatum L. by ICP-OES // Rev. Chimie. V. 62. № 12. P. 1154–1156.
Kabata-Pendias A., 2010. Trace Elements in Soils and Plants. Boca Raton, FL: CRC Press. 548 p.
Kovacik J., 2013. Hyperaccumulation of cadmium in Matricaria chamomilla: A never-ending story? // Acta Physiol. Plant. V. 35. P. 1721–1725.
Kovacik J., Gruz J., Klejdus B., Štork F., Hedbavny J., 2012. Accumulation of metals and selected nutritional parameters in the field-grown chamomile anthodia // Food Chem. V. 131. P. 55–62.
Krämer U., 2010. Metal hyperaccumulation in plants // Ann. Rev. Plant. Biol. V. 61. P. 517–534.
Kupper H., Lombi E., Zhao F.J., McGrath S.P., 2000. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri // Planta. V. 212. P. 75–84.
Lajayer B.A., Ghorbanpour M., Nikabadi S., 2017. Heavy metals in contaminated environment: Destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants // Ecotox. Envir. Saf. V. 145. P. 377–390.
Leonard R.J., McArthur C., Hochuli D.F., 2016. Particulate matter deposition on roadside plants and the importanceof leaf trait combinations // Urban For. Urban Gree. V. 20. P. 249–253.
Lovkova M.Y., Buzuk G.N., 2011. Medicinal plants: Concentrators and superconcentrators of Copper and its role in Metabolism of these Species // Appl. Biochem. Microbiol. V. 47. № 2. P. 189–195.
Ma J.F., Ueno D., Zhao F.J., McGrath S.P., 2005. Subcellular localisation of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens // Planta. V. 220. P. 731–736.
Macnair M., 2003. The hyperaccumulation of metals by plants // Adv. Bot. Res. V. 40. P. 63–105.
Manara A., 2012. Plant responses to heavy metal toxicity // Plant and Heavy Metals / Ed. Furini A. N.Y.: Springer. P. 27–52.
Markert B., 1992. Establishing of 'reference plant' for inorganic characterization of different plant species by chemical fingerprinting // Water Air Soil Pollut. V. 64. P. 533–538.
Masarovicova E., Kralova K., Kummerova M., 2010. Principles of classification of medicinal plants as hyperaccumulators or excluders // Acta Physiol. Plant. V. 32. P. 823–829.
Meers E., Samson R., Tack F.M.G., Ruttens A., Vandegehuchte M. et al., 2007. Phytoavailability assessment of heavy metals in soils by single extractions and accumulation by Phaseolus vulgaris // Environ. Exp. Bot. V. 60. № 3. P. 385–396.
Menzies N.W., Donn M.J., Kopittke P.M., 2007. Evaluation of extractants for estimation of the phytoavailable trace metals in soils // Environ. Pollut. V. 145. № 1. P. 121–130.
Minkina T.M., Mandzhieva S.S., Chaplygin V.A., Burachevskaya M.V., Bauer T.V. et al., 2017. Effect of aerotechnogenic emissions on the content of Heavy Metals in herbaceous Plants of the Lower Don Region // Euras. Soil Sci. V. 50. № 6. P. 746–755.
Overesch M., Rinklebe J., Broll G., Neue H.-U., 2007. Metals and arsenic in soils and corresponding vegetation at Central Elbe river floodplains (Germany) // Environ. Pollut. V. 145. P. 800–812.
Owen J.D., Kirton S.B., Evans S.J., Stair J.L., 2016. Elemental fingerprinting of Hypericum perforatum (St. John’s Wort) herb and preparations using ICP-OES and chemometrics // J. Pharm. Biomed. Anal. V. 125. P. 15–21.
Pavlova D., Karadjova I., 2013. Toxic element profiles in selected medicinal plants growing on Serpentines in Bulgaria // Biol. Trace Elem. Res. V. 156. № 1–3. P. 288–297.
Pollard A.J., Powell K.D., Harper F.A., Smith J.A.C., 2002. The genetic basis of metal hyperaccumulation in plants // Crit. Rev. Plant Sci. V. 21. P. 539–566.
Pollard A.J., Reeves R.D., Baker A.J.M., 2014. Facultative hyperaccumulation of heavy metals and metalloids // Plant Sci. V. 217–218. P. 8–17.
Radanovic D., Antic-Mladenovic S., Jakovljevic M., 2002. Influence of some soil characteristics on heavy metal content in Hypericum perforatum L. and Achillea millefolium L. // Acta Hortic. V. 576. P. 295–301.
Redondo-Gomez S., Mateos-Naranjo E., Vecino-Bueno I., Feldman S.R., 2011. Accumulation and tolerance characteristics of chromium in a cordgrass Cr-hyperaccumulator, Spartina argentinensis // J. Hazard Mater. V. 185. P. 862–869.
Reeves R.D., 1992. Hyperaccumulation of nickel by serpentine plants // The Vegetation of Ultramafic (Serpentine) Soils / Eds Baker A.J.M., Proctor J., Reeves R.D. Andover, UK: Intercept. P. 253–277.
Reeves R.D., Baker A.J.M., 2000. Metal-accumulating plants // Phytoremediation of Toxic Metals: Using Plants to Clean up the Environment. N.-Y.: Wiley. P. 193–229.
Regvar M., Vogel-Mikuš K., 2008. Recent advances in understanding of plant responses to excess metals: Exposure, accumulation and tolerance // Sulfur Assimilation and Abiotic Stress in Plants / Ed. Khan N.A. Berlin; Heidelberg: Springer-Verlag. P. 227–251.
Reimann C., Koller F., Frengstad B., Kashulina G., Niskavaara H., Englmaier P., 2001. Comparison of the element composition in several plant species and their substrate from a 1500000-km2 area in Northern Europe // Sci. Total Environ. V. 278. P. 87–112.
Sæbø A., Popek R., Nawrot B., Hanslin H.M., Gawronska H., Gawronski S.W., 2012. Plant species differences in particulate matter accumulation on leaf surfaces // Sci. Total Environ. V. 427–428. P. 347–354.
Schneider E., Pank F., Koball G., Foltys de Garcia E., Dehe M., Bluthner W.D., 2002. Einfluss von Genotyp und Umwelt auf die Cadmium aufnahme des Johanniskrautes (Hypericum perforatum L.) // Z. Arznei- und Gewürzpflanzen. V. 7. P. 329–335.
Seregin I.V., Kozhevnikova A.D., 2008. Roles of root and shoot tissues in transport and accumulation of Cadmium, Lead, Nickel and Strontium // Rus. J. Plant Physiol. V. 55. № 1. P. 1–22.
Siromlya T.I., Khudyaev S.A., Syso A.I., 2015. Using SRXFA in ecological soil studies on the territory of Novosibirsk // Bull. Rus. Acad. Sci. Physics. V. 79. № 1. P. 89–93.
Uzu G., Sobanska S., Sarret G., Muñoz M., Dumat C., 2010. Foliar lead uptake by lettuce exposed to atmospheric fallouts // Environ. Sci. Technol. V. 44. № 3. P. 1036–1042.
Wei S., Zhou Q., Mathews S., 2008. A newly found cadmium accumulator – Taraxacum mongolicum // J. Hazard Mater. V. 159. P. 544–547.
Zeiner M., Cindric I.J., Pozgaj M., Pirkl R., Silic T., Stingeder G., 2015. Influence of soil composition on the major, minor and trace metalcontent of Velebit biomedical plants // J. Pharm. Biomed. Anal. V. 106. P. 153–158.
Zhang X.H., Liu J., Huang H.T., Chen J., Zhu Y., Wang D.Q., 2007. Chromium accumulation by the hyperaccumulator plant Leersia hexandra Swartz // Chemosphere. V. 67. № 6. P. 1138–1143.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии