

Журнал общей биологии, 2019, T. 80, № 1, стр. 14-21
Обновляющийся вид. Общее популяционно-генетическое объяснение феномена вида для сингамных и агамных организмов
А. С. Пшеничнов *
Московский государственный университет им. М.В. Ломоносова, биологический факультет,
каф. биологической эволюции
119991 Москва, Ленинские горы, 1, Россия
* E-mail: pxemon@gmail.com
Поступила в редакцию 15.11.2017
После доработки 28.04.2018
Принята к публикации 04.06.2018
Аннотация
Предлагается общий популяционно-генетический механизм, объясняющий существование видов как стабильно воспроизводящихся групп сходных организмов: генетическое обновление в результате действия генетического дрейфа или отбора, ограничивающих генетическое разнообразие в популяции. В отличие от концепций, объясняющих целостность вида через скрещиваемость особей, генетическое обновление за счет дрейфа и отбора объясняет существование видов не только для сингамных (размножающихся половым путем), но и для агамных (размножающихся бесполо) организмов. В терминах генетического обновления определяется популяция, изоляция и вид. Предложенная концепция обновляющегося вида – развитие когезивной концепции вида Алана Темплтона.
Виды биологических организмов представляют собой различимые группы генетически сходных организмов, существующие в течение многих поколений и обособленные от других таких групп. Виды встречаются у разных организмов, независимо от их типа размножения и характера передачи генетического материала: и у сингамных организмов, и у агамных многоклеточных (Fontaneto, Barraclough, 2015), и у прокариот (Rosselló-Móra, Amann, 2015), для многих из которых характерны конъюгация и другие механизмы горизонтального переноса генов (ГПГ). Но до сих пор не существует убедительного универсального объяснения, почему самые разные организмы образуют виды. Цель статьи – предложить такое объяснение.
Существующие концепции вида описывают этот феномен в разнообразных аспектах. По наблюдению Уилкинса (Wilkins, 2011), который рассмотрел 27 разных концепций, большинство из них не ставит целью объяснить универсальную “механику”, благодаря которой виды вообще существуют, причину когезии видов (поддержания их целостности в течение многих поколений). В основном концепции решают иные актуальные биологические задачи. Одни помогают “идентифицировать вид”, провести его границу, определить, относятся ли организмы к одному виду или к разным (например, морфологические и таксономические концепции). Другие описывают эволюционное поведение вида как “черного ящика”, безотносительно механизмов, которые его создают (например, эволюционная концепция).
На “механике вида” фокусируется лишь небольшое число концепций. Все они объясняют когезию вида через элементарные популяционно-генетические процессы. Рассмотрим эти концепции ниже.
Биологическая концепция вида Майра (Mayr, 1942) и концепция распознавания (recognition species, (Paterson, 1985)) объясняют когезию вида скрещиванием. Такое объяснение подходит только для сингамных организмов, но не подходит для организмов с другим способом передачи генетической информации – агамных, с сугубо вертикальной передачей генов или с нерегулярным ГПГ. Однако агамные и сингамные виды как категории не имеют систематических отличий: “мир бесполых организмов в основном так же хорошо (или даже лучше) подразделен на легко различимые биологические таксоны, как и мир организмов, размножающихся половым путем” (Templeton, 1989). К примеру, агамные бделлоидные коловратки имеют лучше различимые виды (Fontaneto, Barraclough, 2015), чем моногононтные, которые иногда размножаются половым путем (Holman, 1987). Есть веские причины ожидать, что механизм, создающий виды, будет общим для сингамных и агамных организмов.
Экологические концепции вида помимо того что дают инструмент для определения конспецификов (т.е. идентифицируют вид) также предлагают механизм когезии: отбор в пользу адаптации к определенной экологической нише, который приводит к поддержанию генетического сходства конспецификов. Экологическое объяснение имеет то преимущество, что не зависит от способа воспроизводства организмов. Однако чаще всего под видообразующей силой в экологических концепциях понимается стабилизирующий отбор в пользу форм, оптимально адаптированных к экологической нише. В этой статье я предложу объяснение сходства организмов в одной экологической нише не столько за счет отбора, сколько за счет воспроизводства популяции в условиях ограниченной емкости экологической ниши.
Когезивная концепция вида Темплтона (Templeton, 1989) стремится объединить биологическую и экологическую концепции, чтобы дать объяснение когезии вида, общее для сингамных и агамных организмов. Концепция не предлагает популяционно-генетического механизма когезии, общего для разных форм воспроизводства, но указывает на аналогию механизмов, действующих у сингамных и агамных организмов.
Я в двух словах опишу когезивную концепцию. Это не является необходимым для понимания популяционной механики, которую я далее описываю в статье. Но когезивная концепция – это основа, от которой я отталкивался при создании концепции обновляющегося вида, и я хочу, чтобы ее общая идея была ясна. Более подробно с когезивной концепцией можно ознакомиться в оригинальной работе Темплтона (Templeton, 1989).
“В когезивной концепции вид – это наиболее всеохватная (most inclusive) популяция индивидов, которые потенциально проявляют фенотипическую когезию через присущие им механизмы когезии”, которые Темплтон далее перечисляет. В результате действия этих механизмов возникает генетическая общность, в поле которой действуют популяционно-генетические силы: “когезивная концепция вида определяет вид как эволюционирующую линию через механизмы, которые очерчивают популяционные границы действия таких элементарных микроэволюционных сил, как генетический поток (в статье Темплтона этот термин используется нестандартно и обозначает половое размножение, см. например, табл. 2 в указанной публикации), естественный отбор и генетический дрейф” (Templeton, 1989, p. 12, 20).
Ряд внутренних когезивных механизмов может “создавать” вид (Templeton, 1989, табл. 2 ). В разных условиях будут действовать разные механизмы когезии, которые, однако, одинаково способны создавать виды:
I. Для сингамных организмов Темплтон выделяет механизмы genetic exchangeability, способности обмениваться генетическим материалом:
а) когезия генетического сходства за счет скрещивания;
б) разрушение когезии генетического сходства за счет половой изоляции.
II. Для агамных организмов выделяются механизмы demographic exchangeability, нахождения организмов в одной фундаментальной нише: “факторы, которые определяют фундаментальную нишу и границы распространения новых генетических вариантов за счет генетического дрейфа и естественного отбора”:
а) когезия за счет генетического дрейфа (идентичность за счет происхождения от общего предка);
б) когезия за счет стабилизирующего отбора;
в) разрушение когезии дрейфа и отбора за счет отбора, разрушающего демографическую взаимозаменяемость.
Темплтон объясняет существование видов действием указанных механизмов когезии и видит видообразование как эволюцию этих механизмов.
Я собираюсь показать, что для объяснения существования видов достаточно единственного механизма когезии, который упоминает Темплтон: когезии за счет генетического дрейфа (пункт IIа в списке выше и в работе Темплтона (Templeton, 1989, табл. 2 )).
КОГЕЗИЯ ОБНОВЛЕНИЯ У АГАМНЫХ ОРГАНИЗМОВ
Последний общий предок постоянно сдвигается в будущее
Этот раздел – выжимка ряда положений, известных в теории нейтральности и теории коалесценции (Futuyama, 2005; Hedrick, 2009). Он не содержит новой научной информации, но консолидирует моменты, существенные для последующих объяснений.
Рассмотрим накопление генетического разнообразия в изолированной агамной популяции. Представим себе популяцию агамных организмов со следующими свойствами:
эффективная численность популяции Ne неизменна от поколения к поколению;
поколения не перекрываются;
организмы размножаются агамно, за счет передачи копии своего генома потомкам (с внесением мутаций);
популяция изолирована, т.е. с течением времени в нее не попадают организмы из других популяций;
популяция происходит от одного общего предка и после этого существует в изоляции достаточно продолжительное время ($ \gg {\kern 1pt} {{N}_{e}}$ поколений).
Представим также, что мы наблюдали популяцию в течение ее исторического развития и точно знаем про каждый существующий сейчас или в прошлом организм, кто его родитель.
На рис. 1,а приведен пример такой популяции с описанными выше свойствами и численностью Ne = 4. Организмы этой популяции и их общие предки отображены на филогенетическом древе. Отметим следующие особенности древа:
Рис. 1.
Динамика изменения T(MRCA), времени последнего общего предка, на филогенетическом древе: а – в поколении t1 последний общий предок всех организмов MRCA1 существовал T(MRCA)1 поколений назад; б – до наступления поколения t2 линии B и D вымерли. В результате время нового последнего общего предка T(MRCA)2 позже, чем аналогичное время T(MRCA)1 для поколения t1.
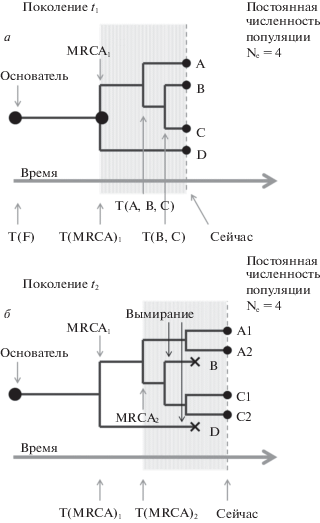
основатель популяции жил T(F) поколений назад;
последний общий предок B и C жил T(B, C) поколений назад. Иными словами, B и C коалесцируют T(B, C) поколений назад;
аналогично, A, B и C коалесцируют T(A, B, C) поколений назад;
все организмы коалесцируют к последнему общему предку (MRCA, the most recent common ancestor) всей популяции, который существовал T(MRCA) поколений назад.
Любой предок последнего общего предка также будет общим предком для всей популяции, например, основатель популяции, который жил T(F) поколений назад. Но они не будут самыми последними.
В каждом поколении одни организмы оставляют больше одного потомка, доживающего до репродукции, а другие не производят ни одного такого потомка. Это может происходить как благодаря отбору в пользу определенных генетических вариантов, так и в силу чисто случайных процессов (генетический дрейф). В результате время от времени целые генетические линии будут исчезать с филогенетического древа. Это иллюстрирует рис. 1,б, на котором та же популяция показана в более позднем поколении t2. За промежуток времени между поколениями t1 и t2 потомки организмов B и D вымерли. Их место заняли потомки A и C. Это не обязательно означает, что A и C лучше адаптированы: элиминация могла быть случайной, т.е. обусловленной исключительно генетическим дрейфом.
В силу описанного процесса время от времени все потомки в пределах одной из ветвей филогенетического древа, происходящей от последнего общего предка, исчезают из популяции. Прежний последний общий предок перестает быть последним. Новым последним общим предком популяции становится другая особь: та, в потомстве которой происходит первое разделение ветвей на филогенетическом древе выживших особей. Время существования нового последнего общего предка всегда позже, чем предыдущего. Таким образом, время существования последнего общего предка периодически сдвигается в более позднее поколение.
Когезия за счет генетического обновления
Согласно теории коалесценции, среднее время существования последнего общего предка T(MRCA) популяции статистически определяется временной динамикой эффективной численности популяции (Ne). Например, если Ne постоянна, то T(MRCA) в среднем равно 2Ne поколений для гаплоидной популяции (см., например, формулы расчета среднего времени коалесценции (Templeton, 2005; Hedrick, 2009)).
Генетическое разнообразие агамной популяции, рассмотренной выше, зависит от того, сколько мутаций накопилось в ветвях, ведущих от последнего общего предка к нынешнему поколению. Но поскольку T(MRCA) периодически перемещается вперед во времени, генетические различия между организмами не увеличиваются бесконечно: приток новых мутаций компенсируется потерей целых ветвей генетического древа. Разнообразие оказывается фиксированным в области значений, которые определяются Ne и скоростью мутаций µ (см. формулы мутационно-дрейфового равновесия, например, Gillespie, 1998; Futuyama, 2005). Поэтому такая популяция будет эволюционировать как группа вечно генетически близких организмов, без накопления генетического разнообразия выше определенного порога.
В рассмотренной модели предполагалось, что вся популяция происходит от одного общего предка. Даже если это не так и предков более одного, через достаточно продолжительное время ($ \gg {\kern 1pt} {{N}_{e}}$) все особи популяции с большой вероятностью будут являться потомками единственного основателя из-за случайной утраты потомков других основателей. Начиная с момента такой утраты популяция начнет эволюционировать как группа генетически близких потомков одного общего предка.
Когезия за счет обновления генов, или когезия обновления – это ограничение генетического разнообразия группы родственных особей в результате их недавнего происхождения от общего предка (в работе Темплтона (Templeton, 1989, табл. 2 ) она указана под номером IIa). При определенных условиях возникновение когезии обновления неизбежно:
1. Генотипы группы воспроизводятся за счет копирования.
2. Группа генотипов изолирована от других таких групп.
Генетическое обновление можно рассмотреть в двух временных аспектах:
в ретроспективе все существующие в настоящий момент генотипы популяции сходятся к единственному последнему общему предку. Время схождения к этому общему предку имеет определенное статистическое распределение, которое определяется динамикой Ne и имеет среднюю величину порядка Ne;
в перспективе любой существующий в определенный момент времени генотип является потенциальным предком всей группы генотипов в каком-либо будущем поколении при условии, что группа не будет разделена непреодолимым барьером. Для разных генотипов вероятность этого может быть различной из-за неодинаковой приспособленности.
Отбор не предотвращает потерю генетического разнообразия
Для действия когезии обновления (т.е. потери генетического разнообразия в результате генетического обновления) достаточно одного лишь генетического дрейфа, даже в отсутствие отбора. Если отбор все-таки есть, то может ли он противодействовать когезии обновления? Отбор в пользу отдельных генотипов – не может. Наоборот, он повышает вероятность быстрой потери менее приспособленных генотипов и может сократить время схождения популяции к общему предку, поскольку вариация приспособленности приводит к уменьшению эффективной численности (Ли, 1978), от которой напрямую зависит T(MRCA).
Вариант отбора, который предотвращает утрату генетического разнообразия – это балансирующий отбор, например когда селективное преимущество получает наименее частый вариант генотипа. Такой отбор может поддерживать сосуществование нескольких генотипов в популяции неопределенно долгое время при условии, что Ne достаточно велика, чтобы случайная утрата одного из генотипов благодаря генетическому дрейфу была маловероятной.
Однако в популяции агамных организмов генотипы, которые поддерживаются балансирующим отбором, начинают эволюционировать как отдельные изолированные субпопуляции. В каждой из них T(MRCA) сдвигается вперед во времени, в то время как последний общий предок этих двух субпопуляций остается в одном и том же историческом моменте. Субпопуляции эволюционируют так, как будто бы они оказались разделены непреодолимым барьером. В итоге между разделившимися популяциями уровень генетических различий начнет возрастать, и генетическое обновление уже не будет поддерживать сходство между ними. Иными словами, в популяции агамных организмов балансирующий отбор не предотвращает когезию обновления, а приводит к разделению популяции, в результате которого формируется несколько субпопуляций.
Когезия обновления в результате действия дрейфа и отбора
В агамной популяции генетическое обновление за счет дрейфа по своему качественному эффекту не отличается от генетического обновления за счет отбора: в обоих случаях лишь часть особей оставляет воспроизводящееся потомство. Случайным образом (дрейф) или под действием отбора, медленнее или быстрее – утрата генотипов в популяции все равно происходит. Качественный итог для генетического разнообразия одинаков: один и только один из существующих в любой момент генотипов станет предком всей популяции для какого-либо будущего поколения.
Изоляция обновления
Одним из условий действия когезии обновления является изоляция (условие 2). В модельной популяции, которую мы рассмотрели выше, изоляция задана как одно из изначальных условий. Возможна ли в естественных условиях изоляция, обеспечивающая действие когезии обновления?
Здесь и далее под изоляцией обновления между агамными группами родственных организмов понимается их способность эволюционировать так, что ни одна из них не может вытеснить другую (т.е. не связана с другой за счет когезии обновления). Степень изоляции обновления может варьировать от полного отсутствия изоляции (аналог панмиксии у сингамных организмов) до полной изоляции обновления, когда особи одной группы не способны оставить потомство в ареале обитания или экологической нише другой. При полной изоляции обновления между двумя близкородственными группами когезия обновления отсутствует, группы эволюционируют независимо, а генетические различия в них накапливаются. Роль факторов, приводящих к изоляции, могут играть:
физические барьеры, которые препятствуют обмену особями между популяциями;
экологические барьеры, когда изолированные популяции состоят из особей, приспособленных к разным экологическим условиям и занимающих разные экологические ниши. Особи одной популяции не способны освоить среду обитания другой и оставить там потомство, которое бы могло эффективно конкурировать с аборигенной популяцией.
Обновляющаяся популяция
Популяция как эволюционирующая единица – это группа организмов, объединенная за счет когезии обновления. Когезия обновления действует, когда популяция занимает определенную экологическую нишу. Границы этой ниши обеспечивают изоляцию данной популяции от других, а “пространство” в пределах этих границ создает поле для действия когезии обновления.
Однако нельзя относить к одной популяции все организмы, занимающие одну и ту же экологическую нишу. Две неродственные группы особей могут временно сосуществовать в одной экологической нише и конкурировать, и это не делает их одной популяцией. Поэтому для определения популяции как эволюционирующей единицы также важно, что ее особи являются недавними потомками общего предка (недавними в том смысле, что их T(MRCA) того же порядка, что и эффективная численность).
Итак, обновляющаяся популяция у агамных организмов – это группа организмов, которые: 1) занимают определенную экологическую нишу и 2) происходят от недавнего общего предка.
КОГЕЗИЯ ОБНОВЛЕНИЯ У СИНГАМНЫХ ОРГАНИЗМОВ
Итак, когезия обновления определена для агамных организмов, генотипы которых воспроизводятся путем копирования и у которых все существующие в настоящий момент особи являются потомками одного общего предка. Однако понятие когезии обновления нельзя напрямую применить к сингамным организмам из-за того, что их геномы достаточно часто рекомбинируют. Невозможно выделить конкретную особь, которая была бы единственным общим предком для всех ныне живущих организмов, потому что геном сохраняет целостность только между актами рекомбинации. Так работает ли когезия обновления в популяциях сингамных организмов?
Когезия обновления в нерекомбинирующем локусе
Хотя понятие когезии обновления неприменимо к рекомбинирующим геномам, оно пригодно для описания эволюционных процессов в пределах их нерекомбинирующих частей. Например, мтДНК воспроизводится копированием, так что на пул мтДНК одного вида действует когезия обновления. Для любого момента времени существует свой последний общий предок всех мтДНК – конкретный ее вариант, который являлся составной частью генома некоей особи в прошлом 11.
Аналогично можно рассматривать генетическое разнообразие любого нерекомбинирующего участка рекомбинирующего генома. Например, у человека это могут быть нерекомбинирующие части ядерных хромосом. У всех гаплотипов (аллелей) такого нерекомбинирующего локуса в любой момент времени есть свой последний общий предковый вариант. Таким образом, аллели нерекомбинирующего локуса образуют хороший обновляющийся пул со свойствами обновляющейся популяции. При этом последние общие предковые варианты разных нерекомбинирующих локусов являются составной частью геномов разных индивидов прошлых поколений, существовавших в разное время.
Представление всего генома как набора нерекомбинирующих участков
Любой участок рекомбинирующего генома можно отнести к определенному обновляющемуся пулу гаплотипов.
Рассмотрим SNP (однонуклеотидный полиморфизм). Он воспроизводится за счет простого копирования родительского варианта. Аллели SNP-локуса произошли от предкового аллеля за счет однонуклеотидной замены. И хотя зачастую невозможно выяснить, какой именно из аллелей был предковым и присутствует ли он все еще в популяции, тем не менее у всего текущего разнообразия аллелей данного SNP-локуса был один предковый вариант. Таким образом пул аллелей SNP-локуса ведет себя как обновляющийся, и его генетическое разнообразие ограничено за счет недавнего происхождения от общего предка. Наблюдаемое генетическое разнообразие будет даже еще ниже, чем у агамных организмов, поскольку у SNP-локуса есть всего четыре возможных состояния (четыре нуклеотида), и вновь возникающие аллели с большой вероятностью могут совпасть с уже имеющимися.
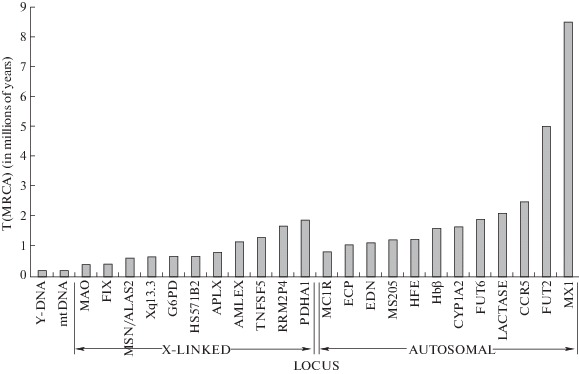
Весь геном можно представить как набор атомарных нерекомбинирующих локусов. Некоторые из них состоят из одного или нескольких нуклеотидов (SNP, микроинсерции, микроделеции, тандемные повторы и т.д.), другие имеют значительную длину (нерекомбинирующие участки Y-хромосомы человека занимают десятки миллионов нуклеотидных пар). Каждый нерекомбинирующий локус формирует обновляющийся пул, генетическое разнообразие которого ограничено когезией обновления и статистически определяется демографическими параметрами популяции. Например, у людей аутосомные локусы представлены в числе 2Ne копий (где Ne – эффективная численность человеческой популяции) и в основном коалесцируют после разделения линий человека и шимпанзе. Время коалесценции для X-сцепленных локусов, локусов нерекомбинирующей части Y-хромосомы и мтДНК в среднем даже более позднее из-за меньшей эффективной численности пулов соответствующих аллелей (рис. 2).
Рис. 2.
Распределение T(MRCA) для 25 нерекомбинирующих локусов человека. Величина T(MRCA) статистически определяется исторической динамикой эффективной численности пула аллелей в локусе. Для аутосом эта численность равна 2Ne, для X-сцепленных локусов – около (3/2)Ne, для мтДНК и Y-хромосомы – около Ne (где Ne – эффективная численность человеческой популяции). Воспроизведен рис. 3 из работы Темплтона (Templeton, 2005).
Это позволяет сделать вывод, что у сингамных организмов, так же как и у агамных, именно когезия обновления является силой, которая ограничивает генетическое разнообразие. Скрещивание же таким фактором не является: этот процесс только определяет границы нерекомбинирующих участков в геноме, на которые действует когезия обновления – точно такая же, которая действует в популяциях агамных организмов.
Эпистаз и рецессивно-доминантные межгенные взаимодействия
Влияют ли межгенные (рецессивно-доминантные и эпистатические) взаимодействия в геноме сингамного организма на действие когезии обновления? Межгенные взаимодействия способны влиять на дисперсию приспособленности и, как следствие, на процесс отбора. Но как показано выше, отбор только способствует ускорению генетического обновления. Таким образом, межгенные взаимодействия в рекомбинантных геномах не препятствуют действию когезии обновления на генетическое разнообразие рекомбинирующих локусов.
Репродуктивная изоляция и изоляция обновления
В панмиктической популяции сингамных организмов атомарные локусы формируют обновляющиеся пулы. Когда сингамная популяция отклоняется от панмиксии и в ней возникает определенный уровень репродуктивной изоляции, в пулах аллелей атомарных локусов формируется соответствующий уровень изоляции обновления. По достижении предельного состояния полной репродуктивной изоляции в атомарных локусах всего генома возникает полная изоляция обновления. Таким образом, репродуктивная изоляция ведет к возникновению изоляции обновления, благодаря которой начинается накопление генетических различий между разделившимися популяциями.
Атомарный локус может выйти из-под действия когезии обновления за счет балансирующего отбора между аллелями так же, как это описано выше для геномов агамных организмов, в результате чего T(MRCA) такого локуса будет увеличено. T(MRCA) аллелей подобных локусов может быть существенно древнее T(MRCA) других локусов. Если этот момент древнее момента наступления репродуктивной изоляции вида от его сестринских видов, то аллельное разнообразие данных локусов может оказаться сходным у этих видов.
ОБНОВЛЯЮЩИЙСЯ ВИД И ВИДООБРАЗОВАНИЕ
Механика возникновения вида за счет генетического обновления
Генетическое обновление как механизм генетической когезии объединяет организмы в популяции. Естественные виды обычно представляют собой системы таких популяций, или подразделенные суперпопуляции. Вид зачастую определяют как наиболее всеохватную популяцию, субпопуляции которой связаны за счет механизмов когезии. Это позволяет дать объяснение вида на основе определения популяции, которое мы дали выше.
Обновляющийся вид – это наиболее всеохватная группа особей, организмы которой объединены действием когезии обновления, т.е. 1) воспроизводятся в определенной экологической нише и 2) генетически сходны за счет недавнего происхождения их геномов (целых или по частям) от геномов предковых особей. Определение обновляющегося вида описывает условия, в которых группа особей образует “видоподобную” эволюционирующую группу – устойчивую во времени и идентифицируемую группу генетически сходных особей. В применении к сингамным организмам концепция обновляющегося вида сводится к биологической концепции, когда в роли среды (“экологической ниши”) для атомарных нерекомбинирующих аллелей выступают локусы в генофонде вида.
Концепция обновляющегося вида включает “экологическое требование” и в этом сходна с экологическими концепциями вида. Например, согласно концепции ecospecies Ван Валена, “вид – это линия (или близкородственная группа линий), которая занимает адаптивную зону, хоть немного отличающуюся от адаптивной зоны любой другой линии в пределах ее ареала, и которая эволюционирует независимо от всех линий за пределами ее ареала” (Van Valen, 1976, p. 233). Однако в рамках экологических концепций вида обычно предполагается, что особи одного вида находятся под одинаковым действием отбора, который заставляет их адаптироваться к одной и той же нише, из-за чего обеспечивается генетическая стабильность вида и его организмы остаются похожими на протяжении многих поколений. В рамках концепции обновляющегося вида важно только то, что границы заселенной ниши определяют границы действия когезии обновления (т.е., в первую очередь – генетического дрейфа).
Насколько сильно могут отличаться экологические ниши разных популяций одного вида, чтобы они еще оставались видом? Если особь, происходящая из одной популяции, может стать предком всех особей в ареале другой популяции (т.е., между популяциями действует когезия обновления), то популяции принадлежат к одному виду.
Видообразование может происходить благодаря возникновению изоляции обновления за счет физических, экологических, а у сингамных – еще и репродуктивных барьеров между группами особей.
Обновляющийся вид и горизонтальный перенос генов (ГПГ)
Популяции агамных организмов с сугубо вертикальным наследованием (такие, как модельная популяция агамных организмов, рассмотренная выше) – редкое, возможно даже исключительное явление, поскольку в природе широко распространен ГПГ. Как же он влияет на когезию обновления и на видообразование?
С генетической точки зрения ГПГ приводит к тому, что отдельные локусы в генофонде вида имеют историю, отличную от истории остальных локусов. Если для простоты рассматривать единичный случай ГПГ, то локусы линии, которая становится акцептором новых генов, будут коалесцировать в одной популяции, а вновь полученные гены – в другой. В реальности ГПГ происходит чаще, каждое такое событие может приводить к появлению в популяции группы аллелей со своей историей. В результате отнесение группы организмов к надвидовым таксонам сталкивается с концептуальными трудностями.
Зато ГПГ не мешает нам идентифицировать вид, потому что его эффект концептуально ничем не отличается от обычной мутации. На идентичность вида можно смотреть с точки зрения того, насколько привнесение новых генов влияет на адаптивную зону. Эффект ГПГ, как и эффект мутаций, варьирует от полного отсутствия фенотипического проявления до значительных изменений адаптивной зоны (например, приводит к появлению устойчивости к антибиотикам, возможности вырабатывать определенную аминокислоту). При изменениях границы занимаемой экологической ниши благодаря генам, приобретенным в процессе ГПГ, часть организмов может приобрести новую видовую идентичность в концепции обновляющегося вида так же, как и в результате мутаций.
Обновляющийся вид среди других концепций вида
Концепция обновляющегося вида – теоретическое обобщение, которое предлагает новое объяснение механики существования видов. Его невозможно практически применять для “идентификации вида” и определения его границ, так же как ради применения биологической концепции Майра невозможно определить возможность организмов скреститься, если по факту они этого не делают. И ценность биологической концепции не в том, что она предлагает эмпирический инструментарий для идентификации конспецификов, а в том, что она предлагает логичное объяснение того, почему сингамные виды существуют. Ценность концепции обновляющегося вида – в этом же, но для более широкой группы организмов.
ЗАКЛЮЧЕНИЕ
В рамках концепции обновляющегося вида существование видов как стабильных популяций генетически сходных особей объясняется действием когезии обновления, независимо от их типа воспроизводства. Внутри агамного вида когезия обновления поддерживает сходство организмов за счет происхождения их геномов от общего предкового генома как такового. В рекомбинирующих геномах сингамных организмов когезия обновления поддерживает аналогичный процесс на уровне отдельных нерекомбинирующих локусов.
Концепция обновляющегося вида – логическое развитие когезивной концепции вида Темплтона. Отличие новой концепции в том, что в ней выделен главный и единственный механизм когезии: когезия обновления.
Можно предположить, что феномен, аналогичный обновляющемуся виду, возникает в любой эволюционирующей системе репликаторов, не обязательно генетических. Примерами таких систем служит масса эволюционирующих культурных явлений: языки и диалекты, музыкальные и художественные стили, типы корпоративной и производственной культуры, традиции воспитания детей, своды летописей и военных уставов, версии машинного кода и так далее. Данные явления демонстрируют ярко выраженную прерывистость в паттерне разнообразия, похожую на прерывистость биологического разнообразия между видами. В механике когезии обновления, на наш взгляд, нет ничего несовместимого с природой этих явлений, хотя выделить в них уровень “атомарных неперемешивающихся элементов”, аналогичных атомарным нерекомбинирующим аллелям, концептуально сложнее. Поиск такой аналогии может стать предметом будущих исследований.
Список литературы
Ли Ч., 1978. Введение в популяционную генетику. М.: Мир. 556 с.
Fontaneto D., Barraclough T.G., 2015. Do Species Exist in Asexuals? Theory and Evidence from Bdelloid Rotifers // Integr. Comp. Biol. V. 55. № 2. P. 253–263.
Futuyama J., 2005. Evolution. Sunderland: Sinauer Associates. 604 p.
Gillespie J.H., 1998. Population Genetics: A Concise Guide. Baltimore; London: Johns Hopkins Univ. Press. 169 p.
Hedrick P., 2009. Population genetics and ecology / Ed. Levin S. Princeton: Princeton Univ. Press. P. 109–116.
Holman E.W., 1987. Recognizability of sexual and asexual species of rotifers // System. Zool. V. 36. P. 381–386.
Mayr E., 1942. Systematics and the origin of species. N.Y.: Columbia Univ. Press. 334 p.
Paterson H.E.H., 1985. The recognition concept of species. Monograph № 4 / Ed. Vrba E.S. Pretoria: Transvaal Museum. P. 21–29.
Rosselló-Móra R., Amann R., 2015. Past and future species definitions for Bacteria and Archaea // Syst. Appl. Microbiol. V. 38. № 4. P. 209–216.
Templeton A., 1989. The meaning of species and speciation: A genetic perspective / Eds Daniel O., Endler J.A. Sunderland: Sinauer Associates. P. 3–27.
Templeton A., 2005. Haplotype trees and modern human origins // Am. J. Phys. Anthropol. № 41. P. 33–59.
Van Valen L., 1976. Ecological Species. Multispecies, and Oaks // Taxon. № 25. P. 233–239.
Wilkins J., 2011. Philosophically speaking, how many species concepts are there? // Zootaxa. V. 2765. P. 58–60.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии