

Журнал высшей нервной деятельности им. И.П. Павлова, 2022, T. 72, № 1, стр. 130-146
Субъективное время: природа и нейробиологические механизмы
1 Институт математических проблем биологии РАН
г. Пущино, Россия
* E-mail: v.sergin@mail.ru
Поступила в редакцию 03.03.2021
После доработки 18.08.2021
Принята к публикации 05.10.2021
- EDN: WHDFWA
- DOI: 10.31857/S0044467722010099
Аннотация
В статье рассматриваются природа и нейробиологические механизмы субъективного времени. Показано, что субъективное время является следствием функционирования сознания. Субъективное время порождается сознанием и является одним из его естественных свойств. Строится формальная модель, которая позволяет объяснить психофизиологические механизмы субъективного времени и пока еще загадочные явления, которые оно порождает. Формулируются качественные характеристики субъективного времени, которые хорошо соответствуют обширным экспериментальным данным по восприятию коротких временных интервалов (длительности стимулов и интервалов между ними), а также тем необычным явлениям, которые наблюдаются только эпизодически. Субъективное время – это отдельная сущность, которая непосредственно не зависит от физического времени, хотя может быть связана с ним посредством интенсивности психической активности субъекта.
ВВЕДЕНИЕ
В последние десятилетия феномен восприятия времени становится предметом широкого изучения. В экспериментальной психологии и физиологии сделано уже довольно много для выяснения его свойств. В качестве физиологических механизмов отсчета внутреннего времени обычно рассматриваются специфические пейсмейкеры или периодические процессы, связанные с обработкой информации в мозге. Можно отметить широкое согласие, которое существует в отношении некоторых важных аспектов восприятия времени. Считается, что интенсивность внутреннего времени определяется интенсивностью текущей психической активности субъекта, связанной с интенсивностью восприятия, мышления или эмоциональной нагрузки.
В обзоре (Teki, 2016) анализируется 150 исследований по этой тематике, выполненных после 2000 г. В обзоре указывается, что способность человека оценивать длительность стимулов и интервалов между ними зависит от физических свойств стимулов, новизны, процессов внимания, мотивации, эмоций и других факторов. В отношении постулируемых механизмов отсчета внутреннего времени заметна явная тенденция перехода от рассмотрения специализированных пейсмейкеров к системным нейронным механизмам. Превалирующая тенденция современных исследований состоит в том, что обработка физических свойств стимулов и их длительности рассматривается совместно в единой унифицированной нейронной системе. В недавних исследованиях (Matthews, Meck, 2016; Teki et al., 2012) предложен общий принцип сенсорной обработки, согласно которому различные факторы, улучшающие восприятие стимула, должны увеличивать его воспринимаемую длительность. Факторы, ухудшающие восприятие стимула, должны уменьшать воспринимаемую длительность. Следует отметить также гипотезу (Eagleman, Pariyadath, 2009), согласно которой восприятие длительности стимула связано с количеством энергии, затраченной на отображение стимула.
И. Силькис (2011, 2017) предложила гипотезу совместной обработки физических свойств сенсорных стимулов и их временных характеристик в параллельных сенсорных и лимбических нейронных цепях кора – базальные ганглии – таламус – кора. Время поступления стимула (параметр “когда”) определяется совместно с параметром “что”, характеризующим физические свойства стимула. Время циркуляции возбуждения в этих замкнутых нейронных цепях определяет тактовую частоту обработки сенсорных стимулов. Любой фактор, усиливающий отображение стимула в новой коре, ведет к росту тактовой частоты и переоценке длительности воспринимаемого времени (длительности стимулов или интервалов между ними). Факторы, ослабляющие отображение стимула в коре, ведут к снижению тактовой частоты и недооценке длительности воспринимаемого времени.
Важно заметить, что в современных экспериментальных исследованиях восприятия времени речь идет об измерении или оценке реального физического времени. Тем самым, по умолчанию, время в системе восприятия рассматривается как реальное физическое время. Но именно это вызывает сомнения. Хорошо известно, что любые характеристики сенсорного объекта в восприятии кардинально отличаются от характеристик реального физического объекта. Например, световая вспышка определенного цвета в действительности представляет собой пакет электромагнитных волн определенного спектра частот. Тем не менее, если мы смотрим на световое пятно, то видим не поток электромагнитных волн, источником которого оно в действительности является, а его сенсорную характеристику, которую порождает мозг, – световое пятно. Если цвет пятна меняется, например, на красный или зеленый, мы видим не изменение спектра электромагнитных волн, которое в действительности происходит, а изменение цвета пятна. Световое пятно – это сенсорная категория, обобщенная характеристика потока электромагнитных волн, которая порождается мозгом.
В объективном физическом мире нет ничего синего или красного, горького или сладкого. Цвет, запах, прикосновение, теплое, кислое, соленое – все это биологические интерпретации физических и химических воздействий, обладающие адаптивной ценностью и основанные на эволюционном и прижизненном опыте организма. Биологические интерпретации (сенсорные и моторные категории) выражают специфические характеристики взаимодействия организма с окружающим миром: твердый, мягкий, жидкий, острый, темный, светлый, близкий, далекий и т.п. Эти категории выражают субъективные качества (психические феномены), которые определяют содержательный аспект восприятия.
Простые субъективные характеристики сложных физических явлений обладают высокой адаптивной ценностью, поскольку позволяют организму быстро и адекватно реагировать на события в окружающей среде. Способность биологических организмов порождать субъективные феномены возникла в процессе эволюции и закрепилась естественным отбором в силу высокой эффективности такого способа отображения окружающего мира.
Согласно современным воззрениям, время поступления стимула (параметр “когда”) определяется совместно с параметром “что”, характеризующим физические свойства стимула. Факторы, усиливающие отображение стимула в новой коре, ведут к переоценке длительности воспринимаемого времени, а факторы, ослабляющие отображение стимула, ведут к недооценке длительности воспринимаемого времени. Следовательно, отображение субъективных характеристик стимула и отображение субъективного времени связаны между собой. Принимая во внимание кардинальные различия характеристик “что” в восприятии и в объективном физическом мире, можно утверждать, что время в нашем мозге, субъективное время, может столь же кардинально отличаться от измеряемого физического времени. В некоторых экспериментальных условиях они могут быть связаны и очень близки и тем не менее при других обстоятельствах субъективное время может сколь угодно сильно отличаться от физического времени.
Несмотря на значительные достижения в понимании нейрофизиологических механизмов отсчета временных интервалов, природа субъективного времени остается загадочной. Остается неясным, что может служить непосредственной мерой субъективного времени и какова его шкала. Это делает весьма проблематичным экспериментальное изучение субъективного времени и порождающих его физиологических механизмов.
Каким образом нам удается адекватно оценивать длительность текущего времени, а также правдоподобно оценивать длительность минувших событий или делать предположения о возможной длительности будущих событий? Почему переживаемое нами время то тянется медленно, то несется вскачь? Почему в детстве время тянется бесконечно долго, а в старости мчится невероятно быстро?
Целью данной работы является выяснение природы субъективного времени. В работе не предлагается никаких специальных или экзотических гипотез относительно физиологических или психических механизмов, порождающих субъективное время. Мы полагаем, что субъективное время является следствием функционирования сознания, которое известно нам по биологическим фактам. Для пациентов под наркозом нет времени и нет никаких сознательных явлений. Во время сна без сновидений нет ни времени, ни сознания. Все это позволяет предположить, что субъективное время порождается сознанием и является одним из его естественных свойств.
Если это действительно так, то выяснение природы субъективного времени может быть основано на рассмотрении процессов функционирования сознания и успех такого исследования зависит от обоснованности наших представлений о психобиологических механизмах сознания. В качестве теоретической основы исследования времени примем разработанную в последние годы модель автоотождествления как физиологического механизма осознания, которая хорошо соответствует экспериментальным данным нейрофизиологии и электрофизиологии головного мозга человека (Сергин 1994, 1998, 2009, 2016, 2020; Sergin, 1994, 2000, 2017, 2021). Модель автоотождествления хорошо соответствует обширным экспериментальным данным о временных характеристиках сознательного восприятия и позволяет понять нейронные механизмы широкого круга феноменов сознательной активности мозга. Мы вправе ожидать, что такая модель может неявно содержать в себе и нейронные механизмы формирования субъективного времени.
Таким образом, данную работу можно рассматривать как частную попытку выяснить ключевые свойства субъективного времени, опираясь на модель автоотождествления паттернов нейронной активности в коре головного мозга человека.
АВТООТОЖДЕСТВЛЕНИЕ КАК ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ ОСОЗНАНИЯ
Способность человека осуществлять явное внутреннее представление как внешних объектов, так и конструируемых внутри образов, является наиболее ярким и специфическим свойством сознания. Функционирование сознания всегда связано с явными представлениями образов, символов, звуков, запахов, прикосновений и т.п. и неотделимо от них. Никакая сознательная деятельность мозга невозможна без явного представления данных, которое, вероятно, является ключевым свойством осознания. Если это действительно так, то какой нейробиологический механизм может осуществлять явное представление данных и тем самым порождать психически переживаемый феномен их осознания? Ответу на этот вопрос посвящены недавние работы (Сергин, 2009, 2016; 2020; Sergin, 2000, 2017, 2021). Здесь мы ограничимся только их кратким изложением.
Как известно, в процессе восприятия стимул порождает в одной или нескольких областях коры специфический паттерн возбуждения. Можно предположить, что выходные нейроны этих областей коры формируют паттерн нейронной активности, тождественный паттерну входного возбуждения. Выходной паттерн нейронной активности может передаваться на входы тех же областей коры через автономные массированно-параллельные обратные связи. Тождественные (совпадающие в основных чертах) паттерны возбуждения, порождаемые стимулом и передаваемые по обратным связям, складываются на одних и тех же нейронных структурах (рис. 1), вызывая разрядку все большего числа нейронов и увеличивая интенсивность возбуждения этих структур. Такой циклический процесс с положительной обратной связью взрывообразно увеличивает интенсивность специфического паттерна возбуждения. Специфичность пространственного возбуждения коры акцентирует специфические характеристики стимула, обеспечивая его категоризацию.
Рис. 1.
Локальная схема процесса автоотождествления. Связи с другими областями коры и подкорковыми структурами не показаны (Сергин, 2009).
Fig. 1. Local scheme of the autoidentification process. Links with other areas of the cortex and subcortical structures are not shown (Sergin, 2009).
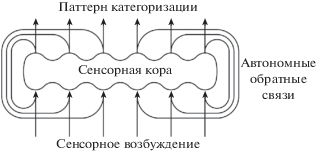
Результат категоризации – цвет, звук, запах – выражает субъективный смысл сенсорного стимула. Паттерн категоризации передается на вход и включается в цикл автоотождествления, обеспечивая интенсивное отображение субъективного смысла стимула. Отображение сенсорных категорий (внутренних данных) паттернами входной нейронной активности коры есть представление этих категорий субъекту в качестве элементов отображения внешнего мира. В результате внешний мир оказывается представленным субъекту не в объективных характеристиках физического мира, а в сенсорных категориях: цвет, вкус, запах, тактильные ощущения и т.п. Это и есть первичные субъективные качества, субъективные ощущения (более глубокое рассмотрение проблемы субъективного качества содержится в статье (Анохин, 2021)).
Следует подчеркнуть, что осознается не входное возбуждение, а сенсорные категории, которые порождаются нейронной структурой коры в ответ на входное возбуждение. Сенсорные категории – это внутренние данные, которые содержатся в памяти, а процесс автоотождествления является способом представления внутренних данных в явной форме. Это значит, что внешнее событие сначала должно быть воспринято, т.е. представлено в сенсорных категориях, и только потом мозг сможет осознать его. Осознание является формой вторичной обработки данных, а процессы неосознаваемого восприятия и осознания оказываются разделенными по времени.
Можно дать априорную оценку вероятной длительности цикла автоотождествления, исходя из экологических условий восприятия. Из постулируемого механизма следует, что любой сигнал должен пройти по крайней мере один цикл автоотождествления, чтобы мозг мог осознать его. Тогда время цикла должно быть меньше характерных временных интервалов изменений в окружающей среде, существенных для выживания организма. В противном случае важные изменения в окружающей обстановке будут происходить, а их осознание не будет успевать за событиями. Адаптивно значимые события, такие как быстрые движения животных, погоня, прыжки, удары лапой, а также время оборонительной реакции характеризуются спектрами, высокочастотные компоненты которых имеют периоды около 0.1 с. Измеренные спектры турбулентности в приземном слое атмосферы, связанные с вихрями и ливнями, колебаниями веток деревьев, кустарников, травы и другими вариациями окружающей среды, имеют резкий спад около 0.1 с. Время цикла автоотождествления – это период квантования непрерывного сигнала. Представление непрерывного сигнала дискретной выборкой требует по крайней мере двух отсчетов за период самой высокочастотной компоненты спектра непрерывного сигнала. Тогда период квантования должен быть меньше или приблизительно равным 50 мс, что является теоретической оценкой времени цикла автоотождествления.
Если сенсорные сигналы подвергаются в аппарате осознания последовательной циклической обработке, то сигналы, попадающие в разные циклы, должны восприниматься как последовательные во времени. Тогда минимальное время, обеспечивающее различение последовательных сигналов, равно длительности цикла. Следовательно, теоретически необходимо существование временного порога различения последовательных событий, величина которого совпадает со временем цикла. Этот временной порог не должен сильно зависеть от модальности сигналов, поскольку порождается универсальным механизмом коры головного мозга.
Порог различения последовательных стимулов был экспериментально установлен еще в 60-е годы прошлого века, он оказался приблизительно одинаковым для слуховой, зрительной и тактильной модальностей, а также для чередующихся стимулов разных модальностей и составляет около 60 мс (Hirsh, Sherrick, 1961; Kristofferson, 1967; Efron, 1973), что близко к теоретической оценке времени цикла автоотождествления. Если порог различения последовательных событий порождается временем синхронной циркуляции возбуждения в нейронных структурах, то он должен изменяться под влиянием тренировки, как и любой физиологический процесс. “Поразительный эффект обучения” (Efron, 1973) обнаружен в самом деле. У тренированных субъектов порог различения последовательных стимулов уменьшается до 15–20 мс для слуховой, зрительной, тактильной и чередующихся модальностей (Hirsh, Sherrick, 1961). Приблизительное равенство порогов различения для разных модальностей, а также их одинаковые изменения под влиянием тренировки, несмотря на кардинальные анатомические и физиологические различия соответствующих перцептивных органов, свидетельствуют об универсальности аппарата осознания в коре головного мозга.
Циклическая обработка информации должна проявляться в особенностях восприятия непрерывности движения (действительного или кажущегося). Примером может служить восприятие кинофильма. Время цикла является минимально различимым временем, поэтому дискретная последовательность кадров кинофильма должна восприниматься как непрерывная, если последовательные кадры попадают в каждый цикл автоотождествления. Если кадры фильма предъявляются через цикл или реже, то такая последовательность может восприниматься как дискретная. Таким образом, из модели автоотождествления следует необходимость существования некоторой критической частоты, выше которой наблюдается непрерывное движение, а ниже – дискретное. Причем численное значение этой критической частоты должно соответствовать частоте циклов. Такая частота действительно существует, она известна как критическая частота слияния мельканий и составляет 10–50 Гц, что соответствует времени цикла 20–100 мс. Эта экспериментальная оценка времени цикла хорошо соответствует нашей теоретической оценке.
Гипотеза автоотождествления влечет за собой точно формулируемые следствия в отношении временных характеристик осознания сигналов. Анализ обширных психофизиологических данных, относящихся к таким феноменам, как движение, временной порог, временная суммация, обратная маскировка, слияние мерцаний и многих других, позволяет оценить время цикла автоотождествления величиной порядка нескольких десятков миллисекунд, которая может варьировать в диапазоне от 10 до 100 мс (Сергин, 1994; 1998; 2016; Sergin, 1994; 2000; 2017). Следовательно, соответствующую полосу частот циклических процессов можно оценить диапазоном приблизительно от 10 до 100 Гц.
Сенсорное осознание – это осознание внешних событий, которые отображаются паттернами нейронной активности коры и становятся внутренними сигналами мозга. В процессе мышления операционная активность мозга также отображается паттернами нейронной активности коры. Поэтому нейрофизиологический механизм осознания результатов операционной активности мозга может быть аналогичен механизму осознания сенсорных сигналов. Механизм автоотождествления оказывается универсальным аппаратом осознания сигналов, как порождаемых сенсорным входом, так и генерируемых самим мозгом (Сергин, 2016; Sergin, 2017).
Частота циклических процессов автоотождествления является физиологической характеристикой, она может быть различной у разных людей и зависеть от состояния субъекта и интенсивности переживаемых событий, аналогично тому, как это имеет место для других периодических процессов, например, частоты пульса и дыхания. На увеличение потока информации $I$ по любому перцептивному каналу механизм автоотождествления должен отвечать увеличением частоты циклов ${{f}_{c}}$, чтобы успевать включать ее в процесс осознания. Следовательно,
Это соотношение имеет две асимптотики. Если поток информации очень велик, частота может достигать значения, увеличение которого физиологически невозможно. При дальнейшем росте входной информации механизм автоотождествления не может справляться с нагрузкой, что означает функциональный отказ сознания как системы обработки данных и физиологически может выражаться стрессом. Если поток входной информации очень мал, частота падает до минимума, соответствующего состоянию релаксации, что может выражаться ощущениями замедленного течения мыслей, ленивой созерцательности, скуки и пустоты. Следовательно, зависимость частоты циклических процессов от потока входной информации можно представить возрастающей функцией с двумя асимптотиками, около 10 и 100 Гц, как это показано на рис. 2.
Рис. 2.
Зависимость частоты циклов автоотождествления ${{f}_{c}}$ от потока входной информации I. Справа показаны альфа- (8–13 Гц), бета- (14–30 Гц) и гамма- (30–100 Гц) диапазоны электрической активности коры головного мозга.
Fig. 2. Dependence of the frequency of auto-identification cycles on the flow of input information I. On the right are shown alpha (8–13 Hz), beta (14–30 Hz) and gamma (30–100 Hz) ranges of electrical activity of the cerebral cortex.
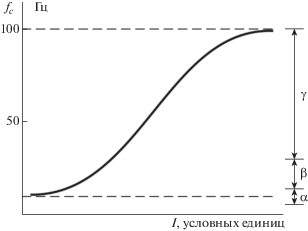
Процессы автоотождествления реализуются в обширных областях коры головного мозга. Если эти циклические процессы действительно существуют, то они могут быть обнаружены прямыми измерениями электрической активности коры. Ф. Крик и К. Кох (Crick, Koch, 1990; Koch, Crick, 1994) выполнили анализ обширных электрофизиологических данных и обнаружили, что зрительное осознание коррелирует с высокочастотной электрической активностью коры головного мозга в диапазоне 30–70 Гц. Этот диапазон точно соответствует центральной полосе частотной характеристики аппарата осознания, представленной на рис. 2.
Данные электро- и магнитоэнцефалографии показывают, что интенсивная сознательная активность мозга характеризуется высокочастотной электрической активностью коры в бета (14–30 Гц) и гамма (30–100 Гц) диапазонах. В состоянии релаксации, отсутствии внешних стимулов или с закрытыми глазами электрическая активность смещается в низкочастотную область с преобладанием альфа-ритма (8–13 Гц) (Кропотов, 2010). Таким образом, высокочастотные колебания электрической активности в диапазоне от 10 до 100 Гц, которые должны порождаться процессами автоотождествления в коре головного мозга, не только существуют в действительности, но и являются доминирующими.
В каких областях коры головного мозга происходит осознание сенсорных данных? Согласно гипотезе автоотождествления, осознание осуществляется посредством автономных обратных связей. Поэтому осознание любой сенсорной характеристики может происходить только в той области коры, которая отображает данное сенсорное качество. Например, осознание цвета должно происходить в основном в зрительной области V4, специализация которой связана с отображением цвета, а осознание движения – преимущественно в специализированной области MT.
Данные первичных зрительных областей коры V1 и V2 также должны осознаваться. Только в этом случае возможно рассматривание деталей зрительного поля с высоким пространственным разрешением. Известно, что больные с пораженной престриарной корой и сохранной областью V1 способны идентифицировать отдельные линии и простые контуры и могут аккуратно их копировать. Это значит, что сенсорные признаки не только отображаются, но и осознаются посредством механизмов зрительной области V1. Но больные неспособны воспринимать совокупность линий как единое целое, например, как дом или собор (Zeki, 1993). Осознание целостных образов возможно только посредством процессов автоотождествления в престриарной и ассоциативной коре, в которых эти образы формируются и которые оказались разрушенными.
Осознание осуществляется в тех же областях коры, в которых происходит неосознаваемое восприятие и формирование моторного ответа. Поэтому автономные обратные связи должны существовать в проекционных и специализированных областях коры всех сенсорных модальностей, а также в моторных и ассоциативных областях, включая височную, теменную и лобную доли. Во всех случаях осознается то содержание, которое соответствует функциям данной области коры.
Вероятным кандидатом на роль анатомической основы аппарата автоотождествления может быть распределенная система кора – базальные ганглии – таламус – кора (Силькис, 2006, 2007, 2011, 2017). В этой системе нейроны каждой области новой коры проецируются в определенную область стриатума, которая затем, через выходные ядра базальных ганглиев, проецируется в область таламуса, которая иннервирует ту же область коры (рис. 3).
Рис. 3.
Упрощенная схема автономных обратных связей в системе кора – базальные ганглии – таламус – кора. Другие связи с подкорковыми структурами мозга не показаны.
Fig. 3. Simplified scheme of autonomous feedbacks in the cortex-basal ganglia-thalamus-cortex system. Other connections with the subcortical structures of the brain are not shown.
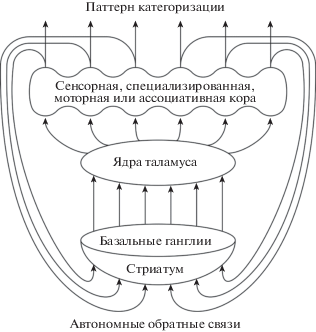
В такой системе интенсивное возбуждение определенной области коры вызывает активацию дофаминергических клеток стриатума, что ведет к последующей реорганизации в цепи, растормаживанию соответствующих клеток таламуса и усилению специфического возбуждения этой области коры. И.Г. Силькис (2006, 2007) показала, что такие дофамин-зависимые положительные обратные связи могут вызывать рост интенсивности и контрастности специфического паттерна возбуждения коры. Время циркуляции возбуждения в замкнутых цепях составляет 20–25 мс и может изменяться в зависимости от физиологических параметров. Взаимодействие холинергической и дофаминергической систем является важнейшим условием функционирования параллельных цепей кора – базальные ганглии – таламус – кора (Силькис, Маркевич, 2017).
Система кора – базальные ганглии – таламус – кора хорошо соответствует априорным условиям аппарата автоотождествления, которые, в свою очередь, соответствуют обширным экспериментальным данным психобиологии и электрофизиологии головного мозга человека. Мы вправе ожидать, что такая система может неявно содержать в себе нейронные механизмы, порождающие специфические свойства субъективного времени.
ПРИРОДА СУБЪЕКТИВНОГО ВРЕМЕНИ
Циклические процессы автоотождествления создают внутреннюю временную шкалу, обеспечивающую различение событий во времени и оценку их продолжительности. Поэтому функционирование механизма автоотождествления должно объяснять способности человека оценивать продолжительность коротких временных интервалов, известные по экспериментальным данным. Согласно модели автоотождествления, сознательное восприятие представляет собой дискретный последовательный процесс. Если это действительно так, то способность человека определять длительность коротких временных интервалов должна быть ограничена ошибкой, равной величине дискретности (времени цикла).
Как известно, поток информации определяется числом различимых состояний сигнала в единицу времени. Если в психофизической задаче оценки временных интервалов экспериментальные условия определены так, что единственной варьируемой характеристикой является их длительность $D$, то поток информации $I\sim {\text{1/}}D$. Частота цикла ${{f}_{c}}$ и его длительность ${{\tau }_{c}}$ связаны соотношением ${{f}_{c}} = {\text{1/}}{{\tau }_{c}}$. Подставляя эти выражения в формулу (1) частотной характеристики аппарата осознания ${{f}_{c}}\sim I$, находим ${\text{1/}}{{\tau }_{c}}\sim {\text{1/}}D$, откуда
То есть время цикла автоотождествления должно возрастать при увеличении длительности оцениваемых интервалов и сокращаться при их уменьшении. Тогда минимальная ошибка оценки интервалов должна соответствовать минимальной длительности цикла и составлять приблизительно 10–12 мс. С ростом длительности оцениваемых интервалов длительность циклов должна возрастать, что может увеличивать минимальную ошибку оценки интервалов до 100 мс. Относительная ошибка оценки длительности интервалов в линейном диапазоне частотной характеристики, представленной на рис. 2, должна оставаться приблизительно постоянной $\delta D{\text{/}}D \approx \operatorname{const} $. Следовательно, при увеличении или уменьшении длительности оцениваемых интервалов в несколько раз минимальная ошибка субъективной оценки должна увеличиваться или уменьшаться во столько же раз.
Эти теоретические предсказания хорошо согласуются с результатами многолетних экспериментальных исследований А. Кристофферсона (Kristofferson, 1967, 1980, 1984), который обнаружил эффект квантования ошибки субъективной оценки длительности временных интервалов. В этих работах установлено, что величина кванта времени является функцией длительности оцениваемых интервалов. Удвоение или деление пополам длительности оцениваемых интервалов заданное число раз удваивает или делит величину кванта в то же число раз. При изменении длительности оцениваемых интервалов от 100 до 800 мс величина кванта изменяется от 12 до 100 мс (Kristofferson, 1980, 1984). То есть экспериментально установленная ошибка субъективной оценки длительности временных интервалов (величина кванта времени) изменяется в точном соответствии со временем цикла автоотождествления. А. Кристофферсон (1984) пришел к заключению, что причиной квантования субъективной оценки длительности интервалов является неизвестный периодический процесс, обеспечивающий внутренний отсчет времени. Из нашей модели становится понятной природа этого периодического процесса, который представляет собой циклический процесс автоотождествления.
Введем понятие интенсивности (скорости) субъективного времени ${{Y}_{t}}$ как отношения субъективного времени к физическому. Согласно современным воззрениям можно предположить, что интенсивность субъективного времени зависит от интенсивности психической активности субъекта ${{Y}_{s}}$. Кроме того, субъективное время связано с частотой циклических процессов автоотождествления ${{f}_{c}}$, поэтому можно написать ${{Y}_{t}} \approx {{Y}_{t}}\left( {{{Y}_{s}},{{f}_{c}}} \right)$.
Интенсивность ${{Y}_{t}}$ не должна содержать размерности времени, поскольку она представляет собой отношение субъективного времени к физическому. Интенсивность психической активности субъекта ${{Y}_{s}}$ можно рассматривать как число событий (образов, символов, знаков и т.п.), отнесенное к единице времени и, следовательно, содержащее размерность времени [1/с]. Из двух определяющих параметров ${{Y}_{s}}$ и ${{f}_{c}}$ можно единственным образом сконструировать функцию, не имеющую размерности времени:
То есть интенсивность субъективного времени определяется интенсивностью психической активности субъекта ${{Y}_{s}}$, отнесенной к частоте циклических процессов автоотождествления.
Субъективное время оказывается привязанным не к физическому времени, а к интенсивности психической активности субъекта. Это значит, что субъективное время является отдельной сущностью, которая непосредственно не зависит от физического времени, хотя может быть связана с ним посредством интенсивности психической активности субъекта.
Рассмотрим частный случай, когда интенсивность психической активности субъекта определяется только интенсивностью потока воспринимаемой информации ${{Y}_{s}}\sim I$, тогда
Поскольку ${{f}_{c}}\sim I$, то для линейного диапазона частотной характеристики, представленной на рис. 2, имеем ${{f}_{c}} \approx kI$, где $k$ – коэффициент пропорциональности. Интенсивность субъективного времени определяется в этом случае так:
(5)
${{Y}_{t}} \approx I{\text{/}}{{f}_{c}} \approx I{\text{/}}kI \approx {\text{1/}}k \approx \operatorname{const} .$Следовательно, в широком диапазоне типичных экологических условий и внутренних параметров субъекта, в которых сохраняются условия линейности частотной характеристики процессов автоотождествления, интенсивность субъективного времени остается постоянной величиной. Это обстоятельство, вероятно, и позволяет нам адекватно оценивать длительность текущего времени, а также правдоподобно оценивать длительность минувших событий или делать предположения о возможной длительности будущих событий.
Способность субъекта правильно оценивать длительность коротких временных интервалов сохраняется только в линейном диапазоне частотной характеристики. Если интенсивность потока информации растет, частота процессов автоотождествления стремится к максимальному значению ${{f}_{c}} \to f_{c}^{{\max }}$. Из частотной характеристики (рис. 2) видно, что при больших значениях частоты величина ${{f}_{c}}$ изменяется мало, и можно написать ${{f}_{c}} \to f_{c}^{{\max }}$ ≈ const. Тогда из выражения (3) следует, что в этой области частот интенсивность субъективного времени ${{Y}_{t}}\sim {{Y}_{s}}{\text{/}}f_{c}^{{\max }}\sim {{Y}_{s}}$ увеличивается с ростом интенсивности психической активности субъекта:
Рост интенсивности субъективного времени означает, что оценка длительности стимулов и интервалов между ними будут преувеличиваться по отношению к реальному физическому времени. Следовательно, внешние события будут казаться происходящими медленнее, чем они происходят в действительности.
Аналогичные рассуждения справедливы и для области низких частот: ${{f}_{c}} \to f_{c}^{{\min }}$ ≈ const. Тогда из выражения (3) также следует ${{Y}_{t}}\sim {{Y}_{s}}{\text{/}}f_{c}^{{\min }}\sim {{Y}_{s}}$ или ${{Y}_{t}}\sim {{Y}_{s}}$. То есть интенсивность субъективного времени уменьшается с уменьшением интенсивности психической активности субъекта. Следовательно, в этой области частот субъективная оценка времени будет преуменьшаться по отношению к реальному физическому времени. Поэтому внешние события будут казаться происходящими быстрее, чем они происходят в действительности.
Таким образом, в линейном диапазоне частотной характеристики интенсивность субъективного времени является постоянной величиной, что обеспечивает способность субъекта правильно оценивать длительность коротких временных интервалов. В области асимптотических продолжений слева и справа от линейного диапазона справедливо выражение (6). Поэтому при возрастании психической активности субъекта оценка длительности коротких интервалов времени будет преувеличиваться по отношению к реальному физическому времени. При снижении интенсивности психической активности субъекта оценка длительности времени будет преуменьшаться по отношению к реальному физическому времени.
Таким образом, из модели автоотождествления (осознания) следуют причинно-следственные отношения, которые разделяют функциональное пространство восприятия временных интервалов на три области: область недооценки длительности временных интервалов, область адекватной оценки и область переоценки длительности интервалов. Эти теоретические следствия модели полностью соответствуют обширным экспериментальным данным по восприятию длительности коротких временных интервалов (Силькис, 2011, 2017; Matthews, Meck, 2016; Teki et al., 2012; Teki, 2016).
ВЫСОКОЧАСТОТНАЯ ОБЛАСТЬ ПРОЦЕССОВ ОСОЗНАНИЯ
Специальный интерес представляет высокочастотная асимптотика процессов автоотождествления. Если интенсивность потока информации растет, частота процессов автоотождествления стремится к максимальному значению ${{f}_{c}} \to f_{c}^{{\max }}$ ≈ const, которое остается приблизительно постоянным. Подставляя это соотношение в (3), получаем
То есть в этом случае интенсивность субъективного времени растет вместе с ростом интенсивности психической активности субъекта ${{Y}_{s}}$. Рост интенсивности субъективного времени может ощущаться как очень быстрое течение субъективного времени по сравнению с реальным физическим временем. В связи с этим рассмотрим важный предельный случай.
Известно, что возникновение ситуации, которая грозит человеку неотвратимой гибелью, вызывает сверхактивацию мозга. Такая сверхактивация мозга вызывается оценкой значимости переживаемых событий, что эквивалентно мгновенному росту величины ${{Y}_{s}} \uparrow $. В то же время частота автоотождествления физически ограничена временем циркуляции возбуждения по замкнутому контуру ${{f}_{c}} \leqslant f_{c}^{{\max }}$, поэтому интенсивность субъективного времени ${{Y}_{t}}$ внезапно возрастает также скачком. Высокая скорость внутреннего субъективного времени может создавать ощущение замедленного течения внешних событий. Окружающий мир вдруг почти останавливается, в то время как мысли протекают чрезвычайно быстро и с исключительной ясностью (благодаря высокой активации мозга). Максимальное количество циклов в единицу времени $f_{c}^{{\max }}$ означает максимальную производительность сознания как системы обработки данных. Становятся доступными для осознания быстро протекающие события и детали окружающей обстановки, обычно недоступные для сознательного восприятия. Становится возможным, за единицу физического времени, больше воспринять, осознать и сделать. На краткий миг открывается возможность сверхбыстрых и адекватных действий в опасной для жизни ситуации.
Аномально высокая активация мозга может происходить в экстремальных ситуациях, в моменты крайней опасности. Известны многочисленные случаи, описанные в литературе, когда за краткие мгновения перед внутренним взором субъекта проносится вся его жизнь. Описаны случаи, когда в условиях неотвратимой угрозы, вызывающей сверхактивацию мозга, внешние события вдруг начинают казаться замедленными, в то время как мысли протекают быстро и с исключительной ясностью. Такая реакция организма обладает высокой адаптивной ценностью, поэтому могла возникнуть и закрепиться в эволюционном процессе. Но уже давно, в более безопасных условиях жизни человеческого общества, потребность в экстремальной реакции уменьшилась и в настоящее время обнаруживается довольно редко и лишь у немногих людей.
Эти рассуждения довольно трудно подтвердить или опровергнуть прямыми экспериментами, поскольку экспериментатор не вправе подвергать жизнь испытуемого неотвратимой угрозе. Тем не менее существуют многочисленные свидетельства участников боевых действий, достоверность которых не вызывает сомнений и которые подтверждают наши рассуждения. Такие свидетельства широко представлены как в биографической литературе участников войны, так и в интернете. Мы не будем на них специально останавливаться, но все же рассмотрим один характерный случай.
Рассказ фронтовика. “Просыпаюсь и слышу немецкую речь… Вот тут все и произошло. Время вдруг страшно замедлилось. Смотрю – из вентиляционного короба показались гранаты. Летят, одна, другая, третья – медленно, словно их на веревочке опускают, только веревочки никакой нету. Падают они, но медленно-медленно! Вращаются, поблескивают, зеленые, с длинными деревянными ручками – на всю жизнь перед глазами. Я как завороженный смотрю и думаю: конец тебе, голубчик. Но спокойно как-то так думаю, без тоски или паники. А они все падают и падают. Ну, сколько гранате падать два с половиной метра? Секунду? А они минут десять падали, я даже устал смотреть. Закрыл глаза и жду, а взрывов все нет и нет. Я даже подумал: надо встать да выйти из землянки. И тут как рванет – один взрыв, второй, на третьем я сознание потерял.” (Ломтев, 2009).
Имеются также редко цитируемые свидетельства ученых, которые в обыденных условиях неожиданно попадали в экстремальные ситуации. Профессор N вспоминает эпизод из школьных лет. Однажды, переходя улицу, он неосторожно перебежал дорогу перед идущим трамваем, поскользнулся на обледенелой поверхности и упал рядом с рельсами. В это время на параллельных путях появился трамвай, идущий в противоположном направлении. N оказался на скользкой поверхности в узком пространстве между движущимися трамваями. Как ни странно, он не испугался и внезапно увидел, что трамваи движутся еле-еле, а колеса проворачиваются чрезвычайно медленно, так что он успевал вытаскивать руку или ногу из-под очередного колеса. Трамваи проехали, он встал и пошел дальше.
Другой эпизод, также относящийся к детству, вспоминает доктор S. “Было мне тогда около 12 лет, и я с приятелем такого же возраста купался в речке. Река текла в красивых и высоких скальных берегах. Сами понимаете, если есть скала, на нее надо забраться. Когда я забрался метров на 10 и до вершины оставалось около метра, то обнаружил, что вверху нет ни трещинки, ни зацепки. Вниз спускаться нельзя, наверняка упаду и разобьюсь. Но никакого испуга не было. Я знал, что умереть никак не могу. Я оглянулся вокруг себя и вдруг увидел, что птицы летят неуклюже и как-то очень медленно. Как коровы, подумалось мне. Крылья еле-еле поднимались и опускались, и можно было рассмотреть каждое перышко. Но было дело и поважнее. Я глянул на скалу и, вероятно, все же обнаружил какие-то зацепки, потому что довольно быстро забрался наверх и бессильно лег на скалу”.
Во всех известных эпизодах доминирования сознания в моменты смертельной угрозы окружающий мир почти останавливается. Ситуация осознается с чрезвычайной ясностью. Нет никакого страха. Мысль работает быстро и очень четко. Такое психофизиологическое состояние прямо противоположно страху, панике и инстинктивному (бессознательному) поведению, которое хорошо известно и доступно экспериментальному исследованию.
Высокочастотная граница процесса автоотождествления $f_{c}^{{\max }}$ в обычных экспериментальных условиях составляет величину порядка 100 Гц. В случае сверхактивации мозга эта величина может на короткое время возрастать скачком. Тем не менее она все равно ограничена временем циркуляции возбуждения по замкнутому контуру. Это значит, что и в экстремальных условиях сверхактивации мозга высокочастотная граница частоты автоотождествления остается жестким ограничивающим параметром.
НИЗКОЧАСТОТНАЯ ВРЕМЕННАЯ ШКАЛА
Восприятие коротких временных интервалов или осознание сенсорных событий формируют высокочастотную шкалу субъективного времени. В отличие от нее, мысли и образы в нашем мозге составляют низкочастотные последовательности. Мы способны осознавать результаты операционной активности мозга и можем произвольно управлять этой активностью, т.е. думать. Примером произвольного управления может служить вербальное повторение: человек произносит слово, слышит его, запоминает, произносит вновь и т.д. Это циклический вербальный процесс сенсорно-моторного повторения. Слова не обязательно произносить вслух, человек может повторять их про себя, что составляет процесс внутреннего сенсорно-моторного повторения. Повторение – управляемый процесс, его можно прерывать и возобновлять вновь, его содержание можно произвольно изменять.
Зрительное повторение представляет собой циклический процесс сенсорно-моторного воспроизведения зрительных образов. Многие зрительные образы, их признаки, свойства и операции над ними выражаются словами и, следовательно, отображаются паттернами речевой моторной системы. Чтобы визуализировать образ, достаточно назвать его про себя или вслух. Называя определенное слово, человек активирует речевую моторную систему, паттерны которой передаются в ассоциативную зрительную кору. Специфическая реакция долговременной зрительной памяти порождает соответствующий образ.
Процедуры преобразования фигур (декомпозиция, сдвиги, повороты и т.п.) также управляются паттернами речевой моторной системы. Эти процессы образного мышления осуществляются посредством механизмов зрительно-вербального повторения. Б. Кохен (Cohen, 1986), анализируя обширный экспериментальный материал, находит, что внутренняя речь играет основную роль в вызове и управлении образами разных модальностей.
Экспериментальные данные показывают, что типичная частота вербального повторения составляет 3–6 Гц, частота зрительного повторения несколько ниже (Klatzky, 1975). Заметим для сравнения, что тактовая частота аппарата осознания на порядок выше и составляет 30–70 Гц, а частота неосознаваемой обработки, вероятно, составляет сотни герц.
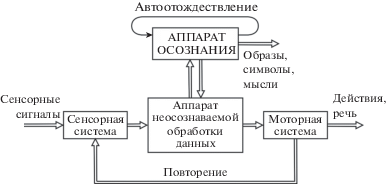
Процедура сенсорно-моторного повторения доступна обозрению и управляема. Физиологически это возможно потому, что повторение является низкочастотным процессом и его содержание доступно обозрению с помощью высокочастотного аппарата автоотождествления (осознания), который надстроен над контуром повторения (рис. 4). Череда мысленных образов, слов или символов управляема посредством программного аппарата моторной системы и доступна обозрению посредством аппарата автоотождествления. Взаимодействующие механизмы сенсорно-моторного повторения и автоотождествления позволяют нам формировать образы, сцены и диалоги, наблюдать и изменять их, порождая подвижный и управляемый мир сознательного опыта (Сергин, 2020, Sergin, 2021).
Рис. 4.
Схема взаимодействия процессов сенсорно-моторного повторения и осознания данных (Сергин, 1994).
Fig. 4. Scheme of interaction of sensory-motor rehearsal processes and data awareness (Sergin, 1994).
В процессе повторения мы можем исключать некоторые элементы или добавлять другие, менять местами элементы выборки, объединять некоторые элементы в группы или разделять их. Повторение нескольких элементов в различных сочетаниях, их сопоставление, манипулирование ими – это и есть процесс мышления. Воспроизводимость и наблюдаемость различных сочетаний позволяют отбирать причинно-следственные отношения. Способность устанавливать причинно-следственные связи является ключевым свойством человеческого разума.
Внутренняя речь и диалоги с самим собой в высшей степени характерны для психики человека. Это та большая рутинная работа, которую постоянно выполняет наш мозг. Внутренний диалог своей назойливой повторяемостью очень напоминает персеверацию – патологическую настойчивость в повторении начатых действий у больных с поражениями префронтальной коры. Избавиться от непроизвольной череды мыслей очень трудно: стоит нам прервать текущую череду мыслей, как она тут же сменяется другой. Внутренний диалог осуществляется посредством сенсорно-моторного повторения, которое является управляемым процессом. Тогда почему мы не можем прекратить череду мыслей и освободить мозг от всяких мыслей вообще просто по своему желанию?
Модель сенсорно-моторного повторения позволяет ответить на этот вопрос. Среди сенсорных функций мозга доминирует детектирование движения, что является наиболее важным для адаптации к изменениям в окружающей среде. Среди множества реакций организма доминирует действие, что обусловлено той же причиной. Внутреннее сенсорно-моторное повторение – это центральный процесс порождения и детектирования действия, поэтому он является доминирующим. Обучение людей с раннего детства основано на многократном повторении звуков, слов, фраз, стихов, песенок, многократном написании черточек, сопрягающих линий, букв, чисел, символов, многократном счете и т.п. Тренируется и межмодальное повторение: слышим-пишем (диктант), видим-говорим (рассказ по картинке) и т.п. Обучение и постоянная тренировка разных способов сенсорно-моторного повторения, обусловленные самим характером нашей цивилизации, превращают естественный доминирующий процесс в стойкий рефлекс.
Внутренний диалог невозможно остановить по желанию, как невозможно прекратить по желанию действие рефлекса. Но можно изменить содержание внутреннего диалога и тем самым изменить набор нейронных структур, осуществляющих сенсорно-моторное повторение (например, при переходе от вербального к зрительному повторению). Изменение содержания диалога осуществляется посредством программного аппарата моторной системы, который доступен произвольному управлению.
Процесс сенсорно-моторного повторения порождает в коре головного мозга взрослых людей доминирующие очаги возбуждения, которые могут препятствовать влияниям префронтальной коры и других отделов мозга или даже подавлять их активность. В результате повторение мыслей и образов оказывается динамически обособленным от других психических процессов. Это и порождает относительную обособленность сознания от остальной психики.
Аппарат рассудочного мышления базируется на данных долговременной памяти и процессах сенсорно-моторного повторения. Повторение создает временную шкалу, которая определяет последовательность событий. Эта низкочастотная шкала принципиально отличается от высокочастотной шкалы процессов осознания. Шкала процессов сенсорно-моторного повторения формируется низкочастотной последовательностью семантических категорий (мыслей и образов), специфические качества которых последовательно осознаются, что позволяет сравнивать их и отличать последующие события от предыдущих. Формируется крупномасштабная шкала семантических категорий, которая позволяет различать прошлое и будущее и оценивать длительность прошедшего времени.
Семантические последовательности могут соответствовать различным масштабам физического времени, от микро- до макроуровней, и относиться к настоящему, прошлому или будущему. В каждом частном случае аппарат мышления, включающий сенсорно-моторное повторение семантических категорий, формирует соответствующую временную шкалу и отображает временную последовательность событий. Семантически значимые временные последовательности многократно повторяются (физически или мысленно) и запоминаются в долговременной памяти. Эти последовательности и составляют основу наших представлений о процессах во времени: биографических, исторических, временах года, перипетий урагана или вспышки молнии. Субъективное время оказывается привязанным не к физическому времени, а к психической активности субъекта.
Интенсивность психической активности субъекта ${{Y}_{s}}$ зависит от интенсивности внутренних событий. Внутренние события – это последовательности мыслей, образов, слов, символов, ощущений и т.п. В этой статье мы рассмотрим самый простой случай, когда интенсивность внутренних событий ${{Y}_{s}}$ определяется только частотой сенсорно-моторного повторения ${{f}_{s}}$. В первом приближении можно написать
С другой стороны, из соотношения (1) следует, что частота циклических процессов автоотождествления ${{f}_{c}}$ зависит от интенсивности потока информации, который в данном случае определяется интенсивностью психической активности субъекта
Тогда
То есть частота автоотождествления пропорциональна частоте сенсорно-моторного повторения, что соответствует смыслу этих процессов: с ростом интенсивности внутренних событий частота автоотождествления должна возрастать, чтобы обеспечивать последовательное осознание этих событий. Данные Кристофферсона (Kristofferson, 1980, 1984), рассмотренные в начале статьи, являются прямым экспериментальным подтверждением этого утверждения.
Возвращаясь к соотношению (3), заметим, что интенсивность субъективного времени растет, если увеличивается психическая активность субъекта ${{Y}_{t}}\sim {{Y}_{s}}{\text{/}}{{f}_{c}}$. Поскольку ${{Y}_{s}}\sim {{f}_{s}}$, получаем
Аппарат автоотождествления должен быть адаптирован к типичной интенсивности внутренних событий. Поэтому в линейном диапазоне частотной характеристики аппарата автоотождествления отношение ${{f}_{s}}{\text{/}}{{f}_{c}} = R$ должно сохраняться приблизительно постоянным. Типичная частота автоотождествления ${{f}_{c}}$ составляет 30–70 Гц. Частота сенсорно-моторного повторения ${{f}_{s}}$ находится в диапазоне 3–6 Гц. Тогда отношение
Эта оценка отношения $R$ соответствует экспериментальным данным, полученным по большому количеству здоровых испытуемых среднего возраста в типичных экологических условиях. Составляющие отношения $R$ являются дофамин-зависимыми функциями, поэтому его величина может быть максимальной в юном возрасте и уменьшаться к старости. Признаками исчерпания ресурсов дофамина и других нейромодуляторов являются известные старческие болезни, такие как болезнь Альцгеймера или Паркинсона.
Неравенство
означает пониженную интенсивность субъективного времени. С увеличением возраста это неравенство становится более сильным, что ведет к дальнейшему уменьшению интенсивности субъективного времени. Поэтому внешние события могут казаться происходящими быстрее, чем это имеет место в действительности. В результате может возникать ощущение, будто физическое время движется слишком быстро и человек просто не успевает за событиями. Только что было утро и вот уже вечер, куда делся целый день? Это может порождать ощущение неудержимо быстрого бега времени: годы летят.Вегетативная нервная система является наиболее древней частью нервной организации млекопитающих. В индивидуальном развитии человека она формируется и достигает максимальной эффективности раньше других частей нервной системы. Ресурсы дофамина и других нейромодуляторов максимальны уже в детском возрасте, что порождает неравенство
Высокая интенсивность субъективного времени может порождать у детей ощущение замедленного течения внешних событий. Время идет, а вокруг ничего не происходит. Нужны игры и сказки, чтобы заполнить время. Дети не ходят шагом, они всегда бегают, что позволяет привести в соответствие высокую интенсивность внутреннего времени с малоподвижным окружающим миром.
Таким образом, ощущения скорости субъективного времени на протяжении жизни человека вполне соответствуют выражению (3). В первой половине статьи показано, что это соотношение хорошо описывает и субъективное восприятие длительности коротких временных интервалов. Все это позволяет утверждать, что соотношение ${{Y}_{t}}\sim {{Y}_{s}}{\text{/}}{{f}_{c}}$ выражает важную фундаментальную закономерность субъективного восприятия времени. В статье даны примеры приложений этого соотношения для анализа процессов субъективного восприятия времени в различных частных случаях.
ЗАКЛЮЧЕНИЕ
Субъективное время, это время, которое мы ощущаем непосредственно, как и другие свойства нашего существования (жарко, холодно, весело, грустно). Это и есть наше внутреннее (психическое) время.
Выполненное исследование показывает, что время в нашем мозге (субъективное время) порождается нейробиологическими механизмами сознания. Такая концепция позволяет объяснить широкое разнообразие феноменов субъективного времени, как доступных систематическому экспериментальному изучению, так и наблюдаемых только эпизодически. Показано, что в широком диапазоне типичных состояний субъекта и окружающей среды интенсивность субъективного времени остается постоянной величиной. Именно поэтому мы способны адекватно оценивать длительность текущего времени, а также правдоподобно оценивать длительность минувших событий или делать предположения о возможной длительности будущих событий.
Если психическая активность субъекта возрастает, интенсивность субъективного времени увеличивается (время движется быстрее), поэтому оценка длительности воспринимаемых интервалов времени преувеличивается по отношению к реальному физическому времени. Внешние события могут казаться протекающими медленнее, чем это имеет место в действительности. Если интенсивность психической активности субъекта снижается, интенсивность субъективного времени уменьшается (время движется медленнее), поэтому оценка длительности воспринимаемых интервалов времени преуменьшается. Внешние события могут казаться протекающими быстрее, чем это имеет место в действительности.
Из нашей концепции следуют причинно-следственные отношения, которые разделяют функциональное пространство восприятия временных интервалов на три области: область недооценки длительности временных интервалов, область адекватной оценки и область переоценки длительности интервалов. Эти теоретические следствия модели полностью соответствуют современным экспериментальным данным по восприятию длительности коротких временных интервалов.
Субъективное время может кардинально отличаться от измеряемого физического времени. В некоторых экспериментальных условиях они могут быть связаны и очень близки, и тем не менее при других обстоятельствах субъективное время может сколь угодно сильно отличаться от физического времени. Субъективное время оказывается привязанным не к физическому времени, а к интенсивности психической активности субъекта. Это значит, что субъективное время является отдельной сущностью, которая непосредственно не зависит от физического времени, хотя может быть связана с ним посредством интенсивности психической активности субъекта.
Возникновение ситуации, которая грозит человеку неотвратимой гибелью, вызывает сверхактивацию мозга. Интенсивность субъективного времени внезапно возрастает скачком. Высокая скорость внутреннего субъективного времени создает впечатление замедленного течения внешних событий. Окружающий мир вдруг почти останавливается, в то время как мысли протекают чрезвычайно быстро и с исключительной ясностью (благодаря высокой активации мозга). Становятся доступными для осознания быстро протекающие события и детали окружающей обстановки, обычно недоступные для сознательного восприятия. Становится возможным, за единицу физического времени, больше воспринять, осознать и сделать. На краткий миг открывается возможность сверхбыстрых и адекватных действий в опасной для жизни ситуации.
Процессы рассудочного и образного мышления реализуются посредством циклических механизмов сенсорно-моторного повторения и автоотождествления. Череда мысленных образов, слов или символов управляема посредством программного аппарата моторной системы и доступна обозрению посредством аппарата автоотождествления. Повторение создает временную шкалу семантических категорий, которая определяет последовательность событий. Низкочастотные последовательности семантических категорий (мыслей и образов) позволяют различать прошлое и будущее и оценивать длительность прошедшего времени.
Семантические последовательности могут соответствовать различным масштабам физического времени, от микро- до макроуровней, и относиться к настоящему, прошлому или будущему. В каждом частном случае аппарат мышления, включающий сенсорно-моторное повторение семантических категорий, формирует соответствующую временную шкалу и отображает временную последовательность событий. Семантически значимые временные последовательности многократно повторяются (физически или мысленно) и запоминаются в долговременной памяти. Эти последовательности и составляют основу наших представлений о процессах во времени: биографических, исторических, временах года, перипетий урагана или вспышки молнии.
В детском возрасте интенсивность субъективного времени велика, что может порождать у детей ощущение замедленного течения внешних событий. Дети не ходят шагом, они всегда бегают, что позволяет привести в соответствие высокую интенсивность внутреннего времени с малоподвижным окружающим миром.
В старости характерна пониженная интенсивность субъективного времени, поэтому внешние события могут казаться происходящими быстрее, чем это имеет место в действительности. В результате возникает ощущение, будто физическое время движется слишком быстро. Это может порождать чувство неудержимо быстрого бега времени: годы летят.
В среднем возрасте физиологические параметры организма наилучшим образом адаптированы к характерным условиям окружающей среды. Нейробиологические механизмы функционируют в линейных диапазонах своих шкал. Человек “шагает в ногу” со временем. Таким образом, существуют три этапа, три возраста в жизни человека, которые выражаются неотъемлемыми специфическими ощущениями течения субъективного времени.
Список литературы
Анохин К.В. Когнитом: в поисках фундаментальной нейронаучной теории сознания. Журн. высш. нервн. деят. им. И.П. Павлова. 2021. 71 (1): 39–71. https://doi.org/10.31857/S00444677210032
Кропотов Ю.Д. Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия. Донецк; издатель Заславский, 2010. 506 с.
Ломтев А. Бег времени. На войне как на войне … Журнал Чудеса и приключения. 2009. 6: 23–25.
Сергин В.Я. Сознание как система внутреннего видения. Журн. высш. нервн. деят. им. И.П. Павлова. 1994. 44 (4–5): 627–639.
Сергин В.Я. Психофизиологические механизмы осознания: гипотеза самоотождествления. Журн. высш. нервн. деят. им. И.П. Павлова. 1998. 48 (3): 558–571.
Сергин В.Я. Нейронные механизмы сознательного восприятия: гипотеза автоотождествления. Лекция на XI Всероссийской научно-технической конференции “Нейроинформатика-2009”. Лекции по нейроинформатике. Ред. Тюменцев Ю.В. М.: МИФИ, 2009. 35–65 с.
Сергин В.Я. Автоотождествление паттернов нейронной активности как физиологический механизм осознания. Журн. высш. нервн. деят. им. И.П. Павлова. 2016. 66 (3): 259–278. https://doi.org/10.7868/S0044467716020076
Сергин В.Я. Автоотождествление и сенсорно-моторное повторение как физиологические механизмы сознания. Журн. высш. нервн. деят. им. И.П. Павлова. 2020. 70 (5): 696–720. https://doi.org/10.31857/S004446772005010X
Силькис И.Г. Вклад синаптической пластичности в базальных ганглиях в обработку зрительной информации (гипотетический механизм). Журн. высш. нервн. деят. им. И.П. Павлова. 2006. 56 (6): 742–756.
Силькис И.Г. Роль дофамин-зависимых перестроек активности в цепях кора-базальные ганглии–таламус-кора в зрительном внимании (гипотетический механизм). Успехи физиол. наук. 2007. 38 (4): 21–38.
Силькис И.Г. Возможный механизм участия цепей кора-базальные ганглии-таламус-кора в восприятии времени. Успехи физиологических наук. 2011. 42 (2): 41–56.
Силькис И.Г. Унифицированный нейронный механизм влияний внимания, эмоций и неврологических заболеваний на восприятие субсекундных временных интервалов (роль базальных ганглиев). Успехи физиологических наук. 2017. 48 (4): 78–94.
Силькис И.Г., Маркевич В.А. Взаимовлияние ацетилхолина, дофамина и ГАМК на функционирование кортико-стрионигральной нейронной сети при болезнях Альцгеймера и Паркинсона (гипотетический механизм). Нейрохимия. 2017. 34 (1): 16–30.
Cohen B.H. The Motor Theory of Voluntary Thinking. Consciousness and self-regulation. Eds R.J. Davidson et al. N. Y.: Plenum Press, 1986. V. 4. 19–37 pp.
Crick F., Koch C. Some reflections on visual awareness. Gold Spring Harbor Symp. on Quantit. Biol. 1990. Vol. LV: 953–962.
Eagleman D.M., Pariyadath V. Is subjective duration a signature of coding efficiency? Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009. 364 (1525): 1841–1851.
Efron R. Conservation of temporal information by perceptual systems. Perception and psychophysics. 1973. 14 (3): 518–530.
Hirsh I.J., Sherrick C.E. Perceived order in different sense modalities. J. of Exp. Psychol. 1961. 62: 423–432.
Klatzky R. Human Memory. Structures and Processes. Freeman, 1975.
Kristofferson A.B. Attention and Psychophysical Time. Acta Psychologica. 1967. 27: 93–100.
Kristofferson A.B. A Quantal Step Function in Duration Discrimination. Perception and Psychophysics. 1980. 27 (4): 300–306.
Kristofferson A.B. Quantal and Determenistic Timing in Human Duration Discrimination. Annals of New York Academy of Sciences. 1984. 423: 3–15.
Koch C., Crick F. Some Further Ideas Regarding the Neuronal Basis of Awareness. Large-Scale Neuronal Theories of the Brain. Eds. Koch, C. and Davis, J. Cambridge, MA: MIT Press. 1994. 93–110 pp.
Matthews W.J., Meck W.H. Temporal cognition: connecting subjective time to perception, attention, and memory. Psychol. Bull. 2016. 142 (8): 865–907.
Sergin V.Ya. Consciousness as a data-processing system. Neural Network World. 1994. 4 (5): 601–608.
Sergin V.Ya. Nature of Sensory Awareness: The Hypothesis of Self-Identification. In Complex Brain Function: Conceptual Advances in Russian Neurosciences. Eds. Miller R., Ivanitsky A.M., Balaban P.M. Harwood Academic Publishers. 2000. 97–112 pp.
Sergin V.Ya. Auto-Identification of Neuronal Activity Patterns as a Physiological Mechanism of Awareness. Neuroscience and Behavioral Physiology. 2017. 47 (7): 737–750. https://doi.org/10.1007/s11055-017-0462-7
Sergin V.Ya. Autoidentification and Sensorimotor Rehearsal as a Physiological Mechanisms of Consciousness. Neuroscience and Behavioral Physiology. 2021. 51 (5): 648–665. https://doi.org/10.1007/s11055-021-1118-x
Teki S., Grube M., Griffiths T.D. A unified model of time perception accounts for duration-based and beat-based timing mechanisms. Front Integr. Neurosci. 2012. V. 5. Article 90.https://doi.org/10.1007/s11055-017-0462-7
Teki S. A citation-based analysis and review of significant papers on timing and time perception. Front. Neurosci. 2016. 95: 58–61.
Zeki S.M. A vision of brain. Blackwell Scientific Publications. 1993. 366 p.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова