

Журнал высшей нервной деятельности им. И.П. Павлова, 2021, T. 71, № 5, стр. 637-648
Особенности вызванных потенциалов ЭЭГ при восприятии быстрых и медленных поглаживаний: ответ распознающей и эмоциональной систем механорецепции
А. А. Варламов 1, 2, *, М. А. Иванова 1, И. В. Скороходов 2, А. Н. Семиреченко 1, Г. В. Портнова 3
1 Лаборатория когнитивных и коммуникативных исследований, Государственный институт русского языка
им. А.С. Пушкина
Москва, Россия
2 АНО “Центр реабилитации инвалидов детства “Наш солнечный мир”
Москва, Россия
3 Федеральное государственное бюджетное учреждение науки Институт высшей нервной деятельности
и нейрофизиологии Российской академии наук
Москва, Россия
* E-mail: antonvarlamov@gmail.com
Поступила в редакцию 27.10.2020
После доработки 10.12.2020
Принята к публикации 02.03.2021
Аннотация
За последние 25 лет было убедительно показано, что у человека существует несколько независимых систем кожной механорецепции. Наиболее изучена распознающая система тактильного восприятия, объединяющая кожные механорецепторы, иннервированные быстрыми миелинизированными афферентами типа Aβ, и предоставляющая подробную информацию о месте контакта и характере стимула. Относительно недавно была открыта эмоциональная С-тактильная система, обеспечивающая безусловные положительные эмоции в ответ на нежные медленные прикосновения. Динамика электрического ответа коры на афферентацию от С-тактильной системы почти не изучена, а существующих данных недостаточно для разграничения компонентов вызванного ответа мозга, связанных с деятельностью Aβ- и С-тактильных систем. В данном пилотном исследовании был проведен анализ вызванных потенциалов ЭЭГ в ответ на быстрые (30 см/с) и медленные (5 см/с) поглаживания, осуществляемые роботизированной установкой. В исследовании приняли участие 13 взрослых испытуемых. Первые выделяемые в ответ на поглаживания компоненты ВП наблюдались во временном интервале 400–1800 мс по латеральным фронтальным и центральным отведениям; их амплитуда была выше для быстрых поглаживаний, что указывает на их связь с активностью Aβ-системы. Исключительно для медленных поглаживаний наблюдались комплекс P2100–N2300, вероятно, связанный с реакцией Aβ-системы на прекращение стимуляции, и сверхпоздний положительный потенциал, предположительно связанный с ответом С-тактильной системы. Полученные результаты подтверждают наличие сверхпозднего положительного потенциала, показанного предшествующими исследованиями для медленных поглаживаний, и позволяют дифференцировать этот потенциал от других поздних компонентов ответа.
Тактильное восприятие на сегодняшний день является одним из наиболее активно развивающихся направлений в нейробиологии сенсорных систем. Исследования последних десятилетий помогли установить, что у человека и большинства млекопитающих существует несколько систем и подсистем кожной механорецепции, которые являются относительно независимыми как структурно, так и функционально. Эти системы включают в себя различные типы кожных высокочувствительных механорецепторов, иннервированных различными типами миелинизированных или немиелинизированных афферентов, и характеризуются различными проводящими путями и различным представительством в коре головного мозга (для общего обзора см. (Abraira, Ginty, 2013; McGlone et al., 2014)). Наиболее изученной является распознающая (дискриминативная) система кожной механорецепции, оперативно предоставляющая нам информацию о форме и фактуре поверхности объектов, к которым прикасаемся мы или которые прикасаются к нам, а также о месте соприкосновения нашего тела с такими объектами. Эта система объединяет рецепторы различного типа (тельца Мейснера, диски Меркеля, тельца Паччини, тельца Руффини, комплексы (плексусы) волосяных фолликулов), иннервированные быстрыми миелинизированными волокнами типа Aβ (Abraira, Ginty, 2013), которые передают информацию по трактам лемнисковой системы (заднестолбовому и спинно-цервикальному проводящим путям) в первичную соматосенсорную кору.
Сравнительно недавно была обнаружена другая система кожной механорецепции – С-тактильная система (С-tactile system, CT-system). Она объединяет высокочувствительные рецепторы, иннервированные немиелинизированными волокнами типа C, которые далее передают информацию по спиноталамическому тракту в островковую кору (для обзора на русском языке см. (Варламов и др., 2019), см. также (Löken и др., 2009; McGlone et al., 2014)). С-тактильная система не предоставляет информацию о локализации места прикосновения или особенностях объекта (Olausson et al., 2008), но обеспечивает безусловные приятные эмоциональные ощущения в ответ на медленные прикосновения с умеренной силой воздействия. В отличие от нейронов распознающей Aβ-системы, интенсивность ответа которых возрастает фактически линейно с увеличением скорости движения стимула по коже, зависимость ответа С-тактильных афферентов от скорости стимуляции имеет характер ∩-образной кривой: наибольшая интенсивность ответа наблюдается для скорости 1–10 см/с, а более низкая и более высокая скорость стимуляции вызывают существенно более слабый ответ афферентов, при этом стимулы, приводящие к наиболее интенсивному ответу афферента, описываются как наиболее приятные и на субъективном уровне (Löken et al., 2009). Еще одно принципиальное различие между С-тактильной и Aβ-тактильной системами заключается в плотности распределения рецепторов на разных участках тела, прежде всего, участков с гладкой (ладони, кончики пальцев) и волосистой кожей: Aβ-тактильная система иннервирует и гладкую, и волосистую кожу человека, при этом средняя плотность ее рецепторов на гладкой коже значительно выше, чем на волосистой, что отражается на соматотопических картах (соматосенсорный гомункулус). В свою очередь, по полученным на сегодняшний день данным исследований с использованием микронейрографии, афферентные волокна С-тактильной системы иннервируют только волосистую кожу, но не ладони и внутреннюю сторону пальцев (Vallbo et al., 1999). Предполагается, что основная функция С-тактильной системы – обеспечивать эмоциональное подкрепление нежным прикосновениям и социальным тактильным контактам, способствуя, таким образом, укреплению эмоциональных и социальных связей в малых группах и в целом развитию социального поведения (McGlone et al., 2014; Olausson et al., 2010; Варламов и др., 2019). Различие между дискриминативным и эмоциональным тактильным восприятием возможно также рассмотреть в соответствии с классификацией Г. Хэда, разработанной еще в 20-х годах ХХ века: по этой классификации С-тактильная система обеспечивает протопатическую чувствительность, в отличие от Aβ-тактильной системы, обеспечивающей различающую, эпикритическую чувствительность (Borhani et al., 2017).
Как уже упоминалось выше, информация от афферентов С-тактильной системы поступает по спиноталамическому тракту с первичной проекцией не в соматосенсорную, а в заднюю островковую кору, откуда передается в другие отделы лимбической и паралимбической системы, в первую очередь в переднюю часть поясной извилины, заднюю часть супратемпоральной борозды и медиальную префронтальную кору (Björnsdotter et al., 2009; Gordon et al., 2013; McGlone et al., 2014; Morrison et al., 2010). Большинство исследований, посвященных обработке информации от С-тактильной системы корой головного мозга, использовали нейрометаболические методы, в первую очередь функциональную магнитно-резонансную томографию (фМРТ). Это позволило с высокой точностью определить зоны, принимающие участие в обработке эмоциональных аспектов прикосновений, однако не позволяет полноценно оценить динамику корковой обработки информации, приходящей от СТ-афферентов, в связи с ограниченным временным разрешением фМРТ. При прикосновениях к волосистой коже происходит одновременная стимуляция рецепторов Aβ-тактильной и С-тактильной систем, что в некоторой степени затрудняет интерпретацию получаемых данных. Более того, ряд исследований показали, что стимуляция ладони, иннервированной исключительно Aβ-афферентами, также может приводить к активации островковой и орбитофронтальной коры (Francis et al., 1999; Rolls, 2004) и сопровождаться приятными ощущениями (Krämer et al., 2007). В этой ситуации использование электроэнцефалографии (ЭЭГ), обеспечивающей значительно более высокое временное разрешение, и в особенности методов анализа вызванного электрофизиологического ответа мозга, позволило бы четко дифференцировать ответ коры на афферентацию быстрой Aβ-тактильной и медленной С-тактильной систем. Несмотря на это, исследованиям вызванных изменений ЭЭГ при восприятии тактильных стимулов посвящено всего несколько статей.
В первую очередь следует отметить статью Р. Акерли и соавт. (Ackerley et al., 2013), показавшую, что при восприятии легких медленных поглаживаний предплечья мягкой кистью со скоростью от 2.3 до 4.5 см/с наблюдается сверхпоздний потенциал (ultra-late potential, ULP), предположительно связанный с афферентацией от С-тактильной системы. В этом исследовании использовалась технология роботизированной тактильной стимуляции (РТС), позволяющая предъявлять стимулы с контролируемой скоростью и силой воздействия (Essick et al., 2010; 1999), при этом для каждого стимула перед поглаживанием, в момент начала движения кисти по направлению к руке испытуемого, посылалась синхронизационная метка, позволившая обеспечить приемлемую точность синхронизации для получения средне- и позднелатентных компонентов вызванных потенциалов (ВП) и вызванных изменений ритмов (ВИР) ЭЭГ. Латентность наблюдавшихся компонентов ВП (от 700 мс) в значительной степени зависела от скорости стимуляции, что указывает на то, что эти компоненты могли быть связаны с ответом Aβ-афферентов на динамические изменения силы воздействия и/или на отрыв кисти от руки, а не с первичным ответом СТ-афферентов, скорость проведения стимула по которым не должна зависеть от скорости самого стимула. К сожалению, в этой работе не проводилось сравнение ответа ЭЭГ на медленные поглаживания, оптимальные для стимуляции СТ-афферентов, и на быстрые поглаживания, в меньшей степени активирующие С-тактильную систему и в большей – Aβ-тактильную систему, что в определенной степени ограничивает возможность интерпретации полученных данных. Нам представляется, что без проведения дополнительных исследований невозможно однозначно связать продемонстрированные в этой работе компоненты ЭЭГ с активностью С-тактильной или Aβ-тактильной систем. Кроме того, часть поздних компонентов может быть связана с ответом Aβ-тактильной системы на прекращение стимуляции, так называемым off-ответом (off-response) (Хорунжий, 2014). Этот ответ достаточно хорошо описан для слуховой системы, как на уровне отдельных рецепторов (Ehret, Merzenich, 1988; Бибиков, 2005), так и на уровне когнитивных ВП на слуховые стимулы (Pantev et al., 1996); существование подобного ответа для соматосенсорных систем подтверждается результатами исследования К. Ямаширо и соавт. (Yamashiro et al., 2008), показавшего выраженные компоненты P100 и N140 на прекращение соматосенсорной стимуляции.
В другом исследовании, недавно опубликованном К. Хаггарти и соавт. (Haggarty et al., 2020), где проводилось прямое сравнение ЭЭГ-коррелятов восприятия поглаживаний со скоростью 3 и 30 см/с, было показано, что амплитуда средне- и позднелатентных (до 1000 мс) компонентов ВП значительно выше на быстрые, чем на медленные поглаживания, однако для медленных поглаживаний наблюдался сверхпоздний позитивный потенциал (3000–4000 мс), отсутствовавший для быстрых поглаживаний, при этом амплитуда этого сверхпозднего потенциала отрицательно коррелировала с выраженностью аутистических черт. В этой работе стимуляция осуществлялась вручную, что могло отрицательно отразиться на точности синхронизации и амплитуде среднелатентных компонентов ВП. Поскольку длительность предъявления медленного поглаживания составляла около 3 с, полученные данные также не позволяют однозначно исключить, что наблюдавшийся сверхпоздний потенциал был связан с афферентацией от С-тактильной системы, а не ответом Aβ-тактильной системы на прекращение стимуляции. Нам представляется, что для получения однозначного ответа на вопрос, связан ли этот сверхпоздний потенциал с отрывом кисти от руки испытуемого, следует использовать стимулы, длительность предъявления которых будет заметно больше или заметно меньше, чем 3 с, что позволит разнести по времени момент прекращения тактильной стимуляции и момент, в который предположительный ответ С-тактильных афферентов будет достигать коры головного мозга. Необходимо также отметить, что и в исследовании Р. Акерли, и в исследовании К. Хаггарти испытуемым давалось задание определять редкие отклоняющиеся стимулы, в связи с чем на сенсорные и эмоциональные компоненты ВП могли накладываться когнитивные компоненты, связанные с принятием решения, является ли предъявляемый стимул отклоняющимся.
Наше исследование направлено на то, чтобы сравнить изменения вызванных потенциалов ЭЭГ на медленные и быстрые поглаживания в рамках экспериментальной схемы, позволяющей разграничить по времени момент прекращения тактильной стимуляции и момент поступления афферентации от С-тактильной системы, что позволит дать более однозначную интерпретацию происхождения сверхпоздних потенциалов. Для этого была разработана экспериментальная схема, в которой длительность медленных поглаживаний составляла 1.8 с. Для минимизации когнитивных компонентов ВП, которые могли бы повлиять на интерпретацию результатов, участникам исследования давалась инструкция просто прислушиваться к собственным сенсорным и эмоциональным ощущениям и не давалось задание на различение стимулов, связанное с необходимостью принятия какого бы то ни было решения. В соответствии с экспериментальной гипотезой ожидалось, что медленные поглаживания будут восприниматься как более приятные и менее возбуждающие по сравнению с быстрыми; при этом в ответ на медленные поглаживания будет наблюдаться более низкая амплитуда ранних компонентов ВП, соответствующих ответу Aβ-тактильной системы, но возможно появление более поздних компонентов ВП, связанных с реакцией Aβ-тактильной системы на прекращение тактильной стимуляции с предположительной латентностью 1900–2500 мс, а также поздних компонентов ВП, связанных с обработкой афферентации от С-тактильной системы.
МЕТОДИКА
1. Испытуемые. В исследовании приняли участие 13 здоровых испытуемых – правшей, из них 7 мужчин и 6 женщин, в возрасте от 20 до 49 лет (средний возраст 28.8 года, SD = 8.6 года). Испытуемые были предварительно ознакомлены с процедурой и дали письменное согласие на участие в исследовании. На проведение исследования было получено разрешение этического комитета Гос. ИРЯ им. А.С. Пушкина.
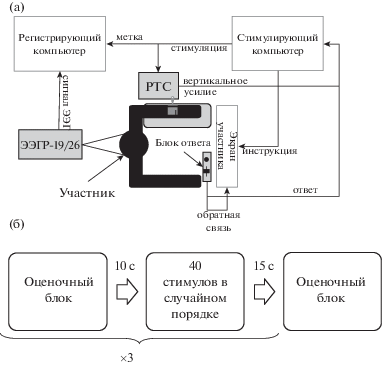
2. Стимулы и экспериментальная процедура. В ходе экспериментальной процедуры испытуемым предъявлялись тактильные стимулы в виде поглаживания кисточкой из искусственного беличьего меха по дорсальной поверхности левого предплечья. Стимуляция осуществлялась при помощи установки роботизированной ротационной тактильной стимуляции (РТС) Dancer Design (см., например, Ackerley et al., 2013), обеспечивающей возможность предъявления стимулов с заданными скоростью и силой воздействия. Стимулы предъявлялись с двумя различными скоростями (5 и 30 см/с) и равной силой (пиковое вертикальное усилие 0.8 Н) в псевдослучайном порядке. Испытуемые располагались на стуле с жесткой спинкой перед 21″ монитором и справа от установки РТС, левая рука располагалась на вакуумной фиксирующей подушке, принимающей форму руки и обеспечивающей комфортное положение и минимизацию движений предплечья (рис. 1 (а)). Испытуемым предъявлялось четыре коротких блока стимулов по четыре стимула (по два стимула со скоростью 5 см/с и со скоростью 30 см/с) с заданием на оценку ощущений; в таких блоках испытуемый должен был оценивать каждое поглаживание по трем аналоговым шкалам: шкала знака эмоции (неприятно – приятно), шкала щекотки (не щекотно – щекотно) и шкала эмоциональной активации (спокоен – возбужден). Ответ на каждый вопрос давался при помощи блока ответа и вспомогательной шкалы, выводимой на экран испытуемого. Блок ответа представлял собой устройство, сочетающее ползунковый потенциометр и кнопку. Положение ползунка регистрировалось в диапазоне от 0 (до упора вниз) до 10 (до упора вверх) с шагом 0.1. Испытуемому демонстрировалось положение метки на вспомогательной шкале, соответствующее положению ползунка. Установив метку в положение, соответствующее степени выраженности ощущения, испытуемый подтверждал свой выбор нажатием на кнопку. Испытуемые не были ограничены во времени при оценке своих ощущений, следующий стимул предъявлялся через 1500 мс после ответа на последний вопрос. Между этими короткими “оценочными” блоками предъявлялось три длинных блока стимулов (40 стимулов в каждом, при восприятии которых испытуемые должны были просто сидеть с закрытыми глазами и прислушиваться к собственным ощущениям. Интервал между блоками составлял 15 с перед началом блока с заданием и 10 с перед началом блока без задания. Схема эксперимента представлена на рис. 1 (б). Всего было предъявлено 136 стимулов, предъявление занимало около 20 мин.
Рис. 1.
Схематическое изображение положения испытуемого во время исследования (а) и общая схема экспериментальной процедуры (б).
Fig. 1. A diagram depicting the layout of the laboratory during the experiment (а) and the experimental scheme (б).
3. Регистрация ЭЭГ и предварительный анализ данных. ЭЭГ регистрировали с использованием 26-канального усилителя “Энцефалан ЭЭГР 19/26” и 19-канальной шапочки с вставными Ag/AgCl электродами (положение и обозначения электродов соответствовали международной схеме 10–20%). Для референтного отведения использовали объединенный мастоидальный электрод, заземляющий электрод располагался в центре лба, сопротивление электродов было не более 10 кОм. При предъявлении каждого стимула посылалась синхронизационная метка, соответствующая типу стимула, при этом роботизированная установка осуществляла регистрацию вертикального усилия на платформе, к которой крепилась стимулирующая насадка, с частотой дискретизации 500 Гц. По этим данным определялся момент реального касания предплечья стимулирующей кистью для каждого предъявления. По нашим оценкам, итоговая точность определения момента касания составляла 10–20 мс, что превышает точность предшествующих исследований (Ackerley et al., 2013; Haggarty et al., 2020). В качестве первого этапа обработки данных осуществлялась полосовая фильтрация (0.3 Гц–35 Гц). Окулографический артефакт удалялся с использованием метода независимых компонент, качество удаления артефакта проверялось экспертом и было устойчиво высоким для всех испытуемых. Для каждого из двух анализируемых экспериментальных условий (быстрых и медленных поглаживаний) выделялись эпохи ЭЭГ, эпохи брались избыточной длины (от –2500 до 5500 мс от начала предъявления стимула), чтобы исключить влияние краевых артефактов, связанных с фильтрацией и сглаживанием. После этого проводилось дополнительное исключение эпох с миографическими, двигательными и иными артефактами (число исключенных эпох по каждому экспериментальному условию составило не более 30% для каждого испытуемого). При анализе ВП коррекция базового уровня проводилась с использованием предстимульного интервала от –200 до 0 мс. Для предварительного анализа данных ЭЭГ использовалось программное обеспечение Brainvision Analyser 2.2 и Matlab.
4. Статистический анализ данных. Временны́е окна компонентов ВП определялись на основании предварительного качественного анализа, после чего для каждого испытуемого были осуществлены определение и экспорт пиковой амплитуды (локальные максимальные или минимальные амплитуды для соответствующего временного интервала) (Luck, 2014) для следующих комплексов компонентов ВП:
а) комплекс, соответствующий первой реакции на касание кожи предплечья кистью: компоненты P600 (временное окно 400–800 мс) и N900 (700–1600 мс);
б) комплекс, наблюдавшийся для медленных поглаживаний и предположительно соответствующий реакции на прекращение стимуляции (отрыв кисти от кожи предплечья): компоненты P2100 (1900–2200 мс) и N2300 (2200–2600 мс).
Дополнительно определялась средняя амплитуда ультрапозднего положительного потенциала для временного окна в 3000–4000 мс.
Для оценки значимости различий для амплитуды каждого из компонентов ВП использовался многофакторный дисперсионный анализ ANOVA с повторными измерениями для отведений, по которым наблюдалась максимальная амплитуда для данного компонента; схемы соответствующих анализов ANOVA приведены в соответствующих разделах. Данные субъективного отчета анализировались отдельно для каждой шкалы с использованием факторов Экспериментальное условие (2) × Блок (4). Сравнения post hoc осуществлялись с использованием критерия Тьюки. Для коррекции значимости полученных результатов при проведении множественных сравнений использовалась поправка Бонферрони (для таких случаев приведенные ниже значения p приводятся уже после коррекции). При проведении статистических анализов использовалось программное обеспечение Statistica 12.0.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
1. Субъективный отчет
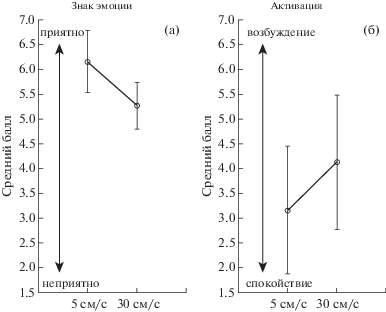
Анализ данных субъективного отчета показал, что для шкалы знака эмоции наблюдался значимый эффект экспериментального условия (F(1, 12) = 11.73, p = 0.005), свидетельствующий о том, что медленные поглаживания оценивались как более приятные по сравнению с быстрыми (см. рис. 2 (а)). Для шкалы эмоциональной активации также наблюдался значимый эффект экспериментального условия (F(1, 12) = 5.91, p = 0.031), показавший, что быстрые поглаживания оценивались как более возбуждающие по сравнению с медленными (см. рис. 2 (б)). Для шкалы щекотки значимых различий получено не было.
Рис. 2.
Средние значения и стандартные ошибки для знака эмоции (а) и степени активации (б) в зависимости от скорости поглаживания.
Fig. 2. Means and SEs for valence (а) and arousal (б) for different velocities of stroking.
2. Анализ данных ВП
Качественный анализ группового усреднения кривых вызванных потенциалов для экспериментальных условий показал, что в ответ на быстрые поглаживания наблюдался один выраженный комплекс компонентов ВП, P600-N900, а в ответ на медленные поглаживания наблюдалось два комплекса компонентов ВП, P600-N900 и P2100-N2300 (см. рис. 3). Максимальная амплитуда этих компонентов наблюдалась по латеральным фронтальным (F3, F4) и передневисочным (F7, F8) отведениям, как для медленных, так и для быстрых поглаживаний. Кроме того, в интервале 3000–4000 мс в ответ на медленные поглаживания наблюдался сверхпоздний положительный компонент, не характерный для быстрых поглаживаний (рис. 4), амплитуда которого была выше для правых фронтальных и центральных отведений (C4, F4). Средние значения и стандартные отклонения амплитуды компонентов для указанных электродов приводятся в табл. 1.
Рис. 3.
Усредненные кривые ВП при быстром и медленном поглаживании для отведения F8 с топографическими картами для компонентов P600, N900, P2100 и N2300.
Fig. 3. ERP grand averages and maps for fast and slow stroking (F8 electrode).
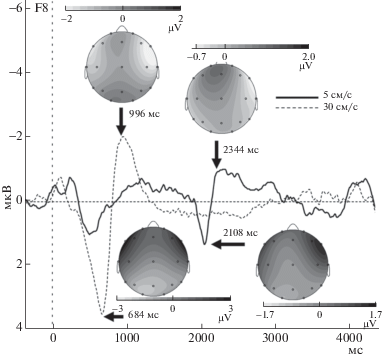
Рис. 4.
Усредненные кривые ВП при быстром и медленном поглаживании для отведения C4.
Fig. 4. ERP grand averages for fast and slow stroking (C4 electrode).
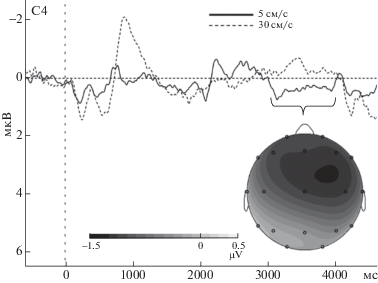
Таблица 1.
Средние значения амплитуды компонентов ВП и стандартные отклонения для релевантных отведений Table 1. Mean amplitudes and SDs for the electrodes of interest
| Быстрые поглаживания | Медленные поглаживания | |||||||
|---|---|---|---|---|---|---|---|---|
| P600 | F3 | F4 | F7 | F8 | F3 | F4 | F7 | F8 |
| Среднее значение | 2.43 | 2.44 | 4.62 | 4.03 | 1.59 | 1.35 | 1.71 | 1.48 |
| Стандартное отклонение | 3.23 | 2.01 | 8.42 | 4.74 | 1.9 | 1.21 | 2.42 | 1.38 |
| N900 | F3 | F4 | F7 | F8 | F3 | F4 | F7 | F8 |
| Среднее значение | –2.52 | –2.57 | –3.01 | –2.35 | –1.13 | –0.83 | –0.89 | –0.83 |
| Стандартное отклонение | 2.12 | 1.35 | 4.86 | 3.02 | 0.87 | 0.73 | 0.93 | 1.11 |
| P2100 | F3 | F4 | F7 | F8 | F3 | F4 | F7 | F8 |
| Среднее значение | 0.71 | 0.89 | 0.33 | 0.52 | 0.93 | 1.43 | 1.79 | 1.48 |
| Стандартное отклонение | 1.08 | 0.62 | 1.06 | 0.83 | 1.45 | 0.95 | 3.5 | 2.17 |
| N2300 | F3 | F4 | F7 | F8 | F3 | F4 | F7 | F8 |
| Среднее значение | –0.73 | –0.4 | –0.62 | –0.22 | –1.15 | –1.4 | –1.41 | –1.95 |
| Стандартное отклонение | 0.82 | 0.59 | 0.76 | 0.68 | 1.22 | 1.38 | 3.16 | 2.08 |
| CПК | F3 | F4 | C3 | C4 | F3 | F4 | C3 | C4 |
| Среднее значение | –0.37 | –0.34 | –0.32 | –0.36 | 0.21 | 0.28 | 0.23 | 0.36 |
| Стандартное отклонение | 0.33 | 0.27 | 0.29 | 0.4 | 0.56 | 0.5 | 0.68 | 0.55 |
2.1. Комплекс P600-N900. Дисперсионный анализ с повторными измерениями, проведенный для фронтальных и передневисочных отведений (F3, F4, F7, F8) для компонента P600 по схеме Электрод (2) × Полушарие (2) × × Экспериментальное условие (2), показал значимый эффект экспериментального условия (F(1, 12) = 7.46, p = 0.018), свидетельствующий, что в этих отведениях амплитуда P600 была выше для быстрых поглаживаний по сравнению с медленными. Такой же анализ для компонента N900 также показал значимый эффект экспериментального условия (F(1, 12) = 6.92, p = 0.022), указывающий на более высокую амплитуду для быстрых поглаживаний. Значимых межполушарных различий получено не было.
2.2. Комплекс P2100-N2300. Дисперсионный анализ с повторными измерениями, проведенный для фронтальных и передневисочных отведений (F3, F4, F7, F8) по схеме Электрод (2) × Полушарие (2) × Экспериментальное условие (2) для компонента P2100, не показал значимых различий между быстрыми и медленными поглаживаниями. Анализ пиковой амплитуды компонента N2300, проведенный по аналогичной схеме, показал наличие значимого взаимодействия Полушарие × Экспериментальное условие (F(1, 12) = = 7.71, p = 0.017); post hoc сравнения показали, что для фронтальных электродов правого полушария амплитуда компонента N2300 была значимо выше для медленных поглаживаний, чем для быстрых (p < 0.01).
2.3. Сверхпоздний позитивный компонент (СПК, 3000–4000 мс). Дисперсионный анализ, проведенный для фронтальных и центральных отведений (F3, F4, C3, C4) по схеме Электрод (2) × Полушарие (2) × Экспериментальное условие (2), показал выраженный значимый эффект экспериментального условия (F(1, 12) = 29.97, p = 0.0001), свидетельствующий о том, что амплитуда СПК была выше для медленных поглаживаний по сравнению с быстрыми. Несмотря на некоторую тенденцию к правополушарной латерализации различий (см. карту на рис. 3), этот эффект не был значимо связан с эффектом полушария.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ данных субъективного отчета показал, что медленные поглаживания воспринимались испытуемыми как более приятные и менее активирующие по сравнению с быстрыми поглаживаниями, что подтверждает нашу экспериментальную гипотезу и соответствует ранее полученным психофизическим данным и современным представлениям о функциях Aβ-тактильной системы и С-тактильной системы. Характер кривых вызванных потенциалов ЭЭГ, полученных в результате группового усреднения, показал, что использованный при анализе данных подход, обеспечивающий синхронизацию данных ЭЭГ и установки РТС, действительно позволяет получить высокую точность синхронизации, достаточную для полноценного анализа относительно ранних компонентов ВП, связанных с активацией Aβ-тактильной системы. Амплитуда первого комплекса компонентов ВП P600–N900 была выше для быстрых поглаживаний, что подтверждает нашу экспериментальную гипотезу и соответствует результатам исследования К. Хаггарти и соавт. (Haggarty et al., 2020). Относительно небольшая латентность данного комплекса и более высокая амплитуда его компонентов, наблюдаемая в ответ на быстрые поглаживания, позволяют однозначно ассоциировать комплекс P600-N900 с реакцией распознающей Aβ-тактильной системы, которая на уровне рецепторов также отличается линейным увеличением амплитуды ответа в зависимости от скорости движения стимула (Löken et al., 2009).
Латентность комплекса компонентов P2100–N2300, наблюдавшегося исключительно для медленных поглаживаний через 200–600 мс после момента отрыва стимулирующей кисти от кожи предплечья испытуемых, позволяет предположить, что он связан с ответом Aβ-тактильной системы на прекращение стимуляции (см. также Yamashiro et al., 2008). На это же указывает и схожесть формы и топографических характеристик этого комплекса компонентов с комплексом P600-N900: подобное сходство формы комплексов наблюдается и для комплексов слуховых потенциалов N100-P200, наблюдающихся в ответ на предъявление слуховых стимулов и на окончание предъявления длящихся звуков (Pantev et al., 1996). Такая специфика реакции Aβ-системы соответствует и субъективному восприятию медленного поглаживания как длящегося стимула, состояния, имеющего начало и конец. Кроме того, “острый” характер пика P2100 показывает, что вариативность латентности этого компонента среди испытуемых была невелика; это тоже является косвенным показателем его связи с Aβ-системой, характеризующейся высокой скоростью проведения импульса, а не с С-тактильной системой, характеризующейся низкой скоростью проведения и, соответственно, более высокой потенциальной вариативностью латентности, связанной с вариативностью как скорости проведения, так и линейной длины проводящих путей у разных испытуемых.
Помимо комплекса P2100-N2300, для медленных поглаживаний была характерна более высокая амплитуда сверхпозднего позитивного компонента, наблюдавшегося в интервале 3000–4000 мс с момента начала предъявления медленных тактильных стимулов. Несмотря на относительно небольшие различия по амплитуде между медленными и быстрыми поглаживаниями (1–1.5 мкв) и небольшой объем выборки нашего пилотного исследования, значимость полученных различий была достаточно высокой, а латентность и топографические характеристики этого компонента соответствовали результатам, ранее полученным К. Хаггарти и соавт. (Haggarty et al., 2020). Поскольку длительность предъявления медленных поглаживаний в данном исследовании и в исследовании К. Хаггарти значительно различалась (≈1800 и ≈3000 мс), а латентность данного компонента была схожей, мы считаем возможным предположить, что его латентность не связана с моментом прекращения стимуляции. Таким образом, полученные данные позволяют считать, что именно этот сверхпоздний позитивный компонент отражает реакцию С-тактильной системы, а его латентность связана с задержкой проведения импульса по немиелинизированным афферентным волокнам. Использование экспериментальных схем, обеспечивающих несколько типов длительных тактильных стимулов с разным временем отрыва кисти, может обеспечить дополнительное подтверждение независимости латентности этого компонента от момента прекращения тактильной стимуляции. В нашем исследовании осуществлялась стимуляция левого предплечья, при этом амплитуда сверхпозднего компонента была несколько больше для правополушарных фронтальных и центральных отведений; в исследовании (Haggarty et al., 2020), наоборот, стимулировалась правая рука, а амплитуда сверхпозднего компонента была выше для левого полушария. Это позволяет предположить, что данный компонент наблюдается контралатерально зоне стимуляции. Дальнейшие исследования с использованием ЭЭГ высокого разрешения и методов локализации источника активности могли бы более полно охарактеризовать особенности локализации и физиологическую природу данного компонента.
Одним из перспективных направлений дальнейших исследований представляется оценка потенциальной взаимосвязи сверхпозднего позитивного компонента, особенностей эмоциональной реакции на воспринимаемые стимулы, устойчивых индивидуальных характеристик и нейрогормонального ответа организма. В исследовании К. Хаггарти было показано, что амплитуда сверхпозднего позитивного компонента отрицательно коррелирует с выраженностью аутистических характеристик, однако представленных данных недостаточно, чтобы установить, связан ли этот эффект непосредственно с аутистическими чертами либо с высоким уровнем тактильной дефензивности, часто характерной для аутизма (Baranek, Foster, 2017), но наблюдающейся не только при расстройствах аутистического спектра, но и при широком круге иных психоэмоциональных нарушений, включающем, например, расстройства аффективного спектра, нарушения пищевого поведения и посттравматические расстройства (Feldman et al., 2004; Field, 2005). Ряд исследований показали, что тактильная стимуляция, социальные тактильные контакты и легкое физическое воздействие, например, массаж, могут снижать уровень стресса и приводить к увеличению уровня окситоцина (для подробного обзора см. (Walker et al., 2017)). В то же время показано, что такое увеличение уровня окситоцина наблюдается не во всех случаях и может модулироваться устойчивыми индивидуальными характеристиками, эмоциональным состоянием и социальным контекстом (Feldman et al., 2010; 2014; Holt-Lunstad et al., 2011; Riem et al., 2017). Более того, установлено, что и выброс окситоцина, и снижение активационных индексов ЭЭГ в ответ на тактильную стимуляцию наблюдаются только у тех испытуемых, для которых характерна выраженная положительная эмоциональная реакция на тактильное воздействие (Portnova et al., 2020). Более глубокое понимание взаимосвязи механизмов эмоционального тактильного восприятия и стресс-протективного нейрогормонального ответа могло бы способствовать разработке новых эффективных методик, использующих терапевтический тактильный контакт и способствующих социальной и психоэмоциональной реабилитации.
ЗАКЛЮЧЕНИЕ
Результаты, полученные при проведении данного пилотного исследования, подтвердили экспериментальную гипотезу, в соответствии с которой ответ на быстрые поглаживания связан с большей амплитудой ранних компонентов вызванных потенциалов ЭЭГ, а ответ на медленные поглаживания – с увеличением амплитуды поздних компонентов ВП. Полученные данные показывают, что некоторые компоненты вызванных потенциалов, наблюдающиеся в ответ на медленные поглаживания, осуществляемые со скоростью, оптимальной для активации медленной, “эмоциональной” С-тактильной системы (а именно – комплекс компонентов P2100-N2300), предположительно связаны не с обработкой афферентации, поступающей от С-тактильной системы, а с ответом быстрой, распознающей Aβ-тактильной системы на прекращение стимуляции. Однако другой компонент, сверхпоздний позитивный потенциал, наблюдавшийся во фронтоцентральных отведениях контралатерально зоне стимуляции в интервале 3000–4000 мс с момента начала предъявления стимула, по-видимому, отражает именно реакцию С-тактильной системы. Дальнейшие исследования могут помочь более полно охарактеризовать специфику этого компонента, а также установить потенциальную взаимосвязь между его амплитудой, устойчивыми индивидуальными характеристиками и нейрогормональной реакцией в ответ на тактильное воздействие.
Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 18-00-01670 (18-00-01616). Авторы благодарят членов военно-исторического клуба “Армэ” за активное участие в исследовании и анонимных рецензентов за ценные замечания и рекомендации.
Список литературы
Бибиков Н.Г. Кодирование длительности сигналов в слуховой системе. Обзор. Сенсорные системы. 2005. 19 (3): 229–239.
Варламов А.А., Портнова Г.В., Макглоун Ф.Ф. С-тактильная система и нейробиологические механизмы “эмоционального” тактильного восприятия: история открытия и современное состояние исследований. Журнал высшей нервной деятельности им. И.П. Павлова. 2019. 69 (3): 280–293.
Хорунжий Г.Д. Динамика временных характеристик активности в рецептивных полях нейронов задних холмов среднего мозга и слуховой коры мыши: дис. канд. биол. наук: 03.03.01. СПб, 2014.
Abraira V.E., Ginty D.D. The sensory neurons of touch. Neuron. 2013.79 (4): 618–639.
Ackerley R., Eriksson E., Wessberg J. Ultra-late EEG potential evoked by preferential activation of unmyelinated tactile afferents in human hairy skin. Neurosci. Lett. 2013. 535: 62–66.
Baranek G.T., Foster L.G. Tactile Defensiveness and Stereotyped Behaviors. 2017. 91–95.
Björnsdotter M., Löken L., Olausson H., Vallbo A., Wessberg J. Somatotopic Organization of Gentle Touch Processing in the Posterior Insular Cortex. J. Neurosci. 2009. 29 (29): 9314–9320.
Borhani K., Làdavas E., Fotopoulou A., Haggard P. “Lacking warmth”: Alexithymia trait is related to warm-specific thermal somatosensory processing. Biological psychology. 2017. 128: 132–140.
Ehret G., Merzenich M.M. Complex sound analysis (frequency resolution, filtering and spectral integration) by single units of the inferior colliculus of the cat. Brain Res. Rev. 1988. 472 (2): 139–163.
Essick G.K., McGlone F., Dancer C., Fabricant D., Ragin Y., Phillips N., Jones T., Guest S. Quantitative assessment of pleasant touch. Neurosci. Biobehav. Rev. 2010. 34 (2): 192–203.
Essick G.K., James A., McGlone F.P. Psychophysical assessment of the affective components of non-painful touch. Neuroreport. 1999. 10 (10): 2083–2087.
Feldman R., Keren M., Gross-Rozval O., Tyano S. Mother-child touch patterns in infant feeding disorders: Relation to maternal, child, and environmental factors. J. Am. Acad. Child Adolesc. Psychiatry. 2004. 43 (9): 1089–1097.
Feldman R., Gordon I., Schneiderman I., Weisman O., Zagoory-Sharon O. Natural variations in maternal and paternal care are associated with systematic changes in oxytocin following parent–infant contact. Psychoneuroendocrinology. 2010. 35 (8): 1133–1141.
Feldman R., Golan O., Hirschler-Guttenberg Y., Ostfeld-Etzion S., Zagoory-Sharon O. Parent–child interaction and oxytocin production in pre-schoolers with autism spectrum disorder. Br. J. Psychiatry. 2014. 205 (2): 107–112.
Field T. Touch deprivation and aggression against self among adolescents. Developmental Psychobiology of Aggression. Под ред. David M. Stoff, Elizabeth J. Susman. Кембридж, Великобритания: Cambridge University Press, 2005. 117–140.
Francis S., Rolls E.T., Bowtell R., McGlone F., O’Doherty J., Browning A., Clare S., Smith E. The representation of pleasant touch in the brain and its relationship with taste and olfactory areas. Neuroreport. 1999. 10 (3): 453–459.
Gordon I., Voos A.C., Bennett R.H., Bolling D.Z., Pelphrey K.A., Kaiser M.D. Brain mechanisms for processing affective touch. Hum. Brain Mapp. 2013. 34 (4): 914–922.
Haggarty C.J., Malinowski P., McGlone F.P., Walker S.C. Autistic traits modulate cortical responses to affective but not discriminative touch. Eur. J. Neurosci. 2020. 51 (8): 1844–1855.
Holt-Lunstad J., Birmingham W., Light K.C. The influence of depressive symptomatology and perceived stress on plasma and salivary oxytocin before, during and after a support enhancement intervention. Psychoneuroendocrinology. 2011. 36 (8): 1249–1256.
Krämer H.H., Lunblad L., Birklein F., Linde M., Karlsson T., Elam M., Olausson H. Activation of the cortical pain network by soft tactile stimulation after injection of sumatriptan. Pain. 2007. 133 (1–3): 72–78.
Löken L.S., Wessberg J., Morrison I., McGlone F., Olausson H. Coding of pleasant touch by unmyelinated afferents in humans. Nat. Neurosci. 2009. 12 (5): 547–548.
Luck S.J. An Introduction to the Event-Related Potential Technique. 2-е изд. Бостон: MIT Press, 2014. 575 p.
McGlone F., Wessberg J., Olausson H. Discriminative and Affective Touch: Sensing and Feeling. Neuron. 2014. 82 (4): 737–755.
Morrison I., Löken L.S., Olausson H. The skin as a social organ. Exp.Brain Res. 2010. 204 (3): 305–314.
Olausson H., Cole J., Rylander K., McGlone F., Lamarre Y., Wallin B.G., Krämer H., Wessberg J., Elam M., Bushnell M.C., Vallbo A. Functional role of unmyelinated tactile afferents in human hairy skin: Sympathetic response and perceptual localization. Exp. Brain Res. 2008. 184 (1): 135–140.
Olausson H., Wessberg J., Morrison I., McGlone F., Vallbo A. The neurophysiology of unmyelinated tactile afferents. Neurosci. Biobehav. Rev. 2010. 34 (2): 185–191.
Pantev C., Eulitz C., Hampson S., Ross B., Roberts L.E. The auditory evoked “off” response: Sources and comparison with the “on” and the “sustained” responses. Ear Hear. 1996. 17 (3): 255–265.
Portnova G.V., Proskurnina E.V., Sokolova S.V., Skorokhodov I.V., Varlamov A.A. Perceived pleasantness of gentle touch in healthy individuals is related to salivary oxytocin response and EEG markers of arousal. Exp. Brain Res. 2020. 238 (10): 2257–2268.
Riem M.M.E., De Carli P., van Ijzendoorn M.H., Linting M., Grewen K.M., Bakermans-Kranenburg M.J. Emotional maltreatment is associated with atypical responding to stimulation of endogenous oxytocin release through mechanically-delivered massage in males. Psychoneuroendocrinology. 2017. 85: 115–122.
Rolls E.T. The functions of the orbitofrontal cortex. Brain Cogn. 2004. 55 (1): 11–29.
Vallbo Å.B., Olausson H., Wessberg J. Unmyelinated afferents constitute a second system coding tactile stimuli of the human hairy skin. J. Neurophysiol. 1999. 81 (6): 2753–2763.
Walker S.C., Trotter P.D., Swaney W.T., Marshall A., McGlone F.P. C-tactile afferents: Cutaneous mediators of oxytocin release during affiliative tactile interactions? Neuropeptides. 2017. 64: 27–38.
Yamashiro K., Inui K., Otsuru N., Kida T., Akatsuka K., Kakigi R. Somatosensory off-response in humans: An ERP study. Exp. Brain Res. 2008. 190 (2): 207–213.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова