

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 4, стр. 446-459
ЛОКАЛИЗАЦИЯ ИСТОЧНИКОВ ГЕНЕРАЦИИ АЛЬФА-РИТМА ЭЭГ ВО ВРЕМЯ НАБЛЮДЕНИЯ, ВЫПОЛНЕНИЯ И ИМИТАЦИИ ИНСТРУМЕНТАЛЬНЫХ ДВИЖЕНИЙ У ИСПЫТУЕМЫХ С РАЗНЫМ УРОВНЕМ ИНТЕЛЛЕКТА
М. А. Начарова 1, *, С. А. Махин 1, В. Б. Павленко 1
1 Крымский федеральный университет им. В.И. Вернадского
Симферополь, Россия
* E-mail: alikina93@gmail.com
Поступила в редакцию 17.12.2019
После доработки 14.02.2020
Принята к публикации 26.02.2020
Аннотация
При помощи метода sLORETA у 62 взрослых испытуемых с разной степенью развития интеллекта локализовали источники генерации ритма в полосе частот 8–12 Гц при наблюдении, самостоятельном выполнении и имитации круговых движений компьютерной мышью экспериментатора. Показана связь степени развития интеллекта с различиями в пространственных паттернах активации коры головного мозга во время выполнения и имитации движений. Выявлена более выраженная и более локализованная активация структур неокортекса у взрослых испытуемых, имеющих высокую степень развития общего интеллекта. Различия в активации корковых регионов в группах с разной степенью развития общего интеллекта в большей степени опосредуют структуры правого полушария, участвующие в процессах зрительно-моторной координации и различения собственных и чужих действий. Бо́льшая вовлеченность прецентральной, поясной и постцентральной извилин левого полушария в процесс имитации движений экспериментатора в группе с высокой степенью развития интеллекта может указывать на бо́льшую активацию “зеркальной” системы мозга.
ВВЕДЕНИЕ
Одной из актуальных проблем когнитивной нейробиологии является поиск ответа на вопрос о том, какие нейрональные механизмы обеспечивают способность человека к имплицитному пониманию эмоциональных состояний и намерений других людей на основе наблюдения лишь отдельных элементов их поведения. Данная проблематика в последние два десятилетия активно изучается в том числе в русле исследований так называемой “зеркальной” системы мозга (ЗСМ) [Лебедева и др., 2017], в основе которой лежит представление о наличии у человека сети нейронов с “зеркальными” характеристиками, под которыми подразумевается их свойство активироваться как при самостоятельном выполнении целенаправленных действий, так и при наблюдении за аналогичными действиями, выполняемыми другими.
Традиционно полагают, что ключевыми зонами ЗСМ как у обезьян, так и у человека являются участки коры, расположенные в премоторной лобной и нижней височной областях головного мозга [Hecht et al., 2015]. При этом возможности непосредственного изучения активности отдельных нейронов у человека имеют существенные ограничения. К настоящему времени опубликованы результаты лишь одного исследования, в котором изучалась активность отдельных нервных клеток в задачах, включающих в себя выполнение и наблюдение за выполнением действий человеком [Mukamel et al., 2010]. При этом электроды располагались лишь в зонах дополнительной моторной области и медиальной височной коры. В ходе эксперимента были обнаружены группы нейронов, которые разряжались сходным образом в ситуациях выполнения и наблюдения действий, тем самым подтвердив гипотезу о наличии у человека нервных клеток, обладающих “зеркальными” свойствами. Более того, в этих же областях были выявлены нейроны, которые возбуждались при выполнении действий, но тормозились при их наблюдении, в связи с чем получили название “антизеркальных” [Keysers et al., 2010]. Данные факты могут говорить о том, что ЗСМ человека имеет более сложное строение и включает в себя дополнительные структуры мозга наряду с описанными ранее в экспериментах с обезьянами. В ряде исследований с использованием различных методов нейровизуализации и транскраниальной магнитной стимуляции получены результаты, также указывающие на высокую степень распределенности ЗСМ [напр., Saygin, 2007; Candidi et al., 2008].
В качестве одной из относительно простых техник оценки активности коры, связанной с функционированием ЗСМ, принято использовать метод регистрации электроэнцефалограммы (ЭЭГ) у людей в процессе выполнения ими специальных экспериментальных задач. Маркером активации ЗСМ считают реакцию десинхронизации сенсомоторного ритма (мю-ритма) ЭЭГ, которая проявляется сходным образом как при выполнении произвольных движений, так и при их мысленном представлении, а также при наблюдении за движениями других [см. обзор Fox et al., 2015; Лебедева и др., 2018]. Однако остаются дискуссионными вопросы локализации эффектов супрессии амплитуды собственно сенсомоторного ритма на фоне изменений амплитуды других видов альфа-подобной активности, имеющих иные источники генерации, особенно в задачах, подразумевающих когнитивные нагрузки и колебания внимания [Hobson et al., 2017]. Следует учитывать, что в отдельных публикациях авторы к мю-ритму относят и альфа-активность в верхне-лобных и теменных регионах, которая имеет сходный с мю-ритмом характер реактивности в аналогичных задачах [Frenkel-Toledo et al., 2013, Кайда и др., 2019].
В настоящее время приобрела широкую популярность гипотеза о наличии связи между активностью ЗСМ и эмпатическими характеристиками личности [Махин и др., 2015; Jeon et al., 2018]. Более того, высказываются гипотезы о том, что система зеркальных нейронов может иметь ключевое значение в обучении и формировании личности человека [см. обзор Лебедева и др., 2017]. Благодаря возможности имплицитного понимания целей и намерений, стоящих за простейшими действиями, повышается эффективность их освоения и ускоряется обучение путем подражания. Мета-анализ результатов исследований, посвященных изучению механизмов подражательного поведения у человека, позволил заключить о существенной вовлеченности структур ЗСМ в данные процессы. При этом указывается на то, что в реализации имитационного поведения существенную роль играет и ряд других структур мозга префронтальной и теменной локализации, которые не являются частью классической ЗСМ [Molenberghs et al., 2012].
Принимая во внимание роль системы “зеркальных” нейронов в процессах социального познания и обучения, можно ожидать, что индивидуальные особенности развития и функционирования ЗСМ в значительной степени предопределяют ход формирования общей способности человека к познанию и решению проблем, которую принято обозначать термином “коэффициент интеллекта” (или фактор g). При этом работы, так или иначе затрагивающие данную проблематику, единичны. Например, опубликованы данные о корреляциях между уровнем невербального интеллекта и степенью десинхронизации мю-ритма при осуществлении целенаправленных движений у взрослых испытуемых (условие наблюдения за действиями других в исследовании не рассматривалось) [Anwar er al., 2015]. Другой группой исследователей были найдены взаимосвязи между уровнем интеллекта и супрессией мю-ритма во время выполнения и наблюдений движений у здоровых детей и у детей с расстройствами аутистического спектра в возрасте от 8 до 13 лет [Raymaekers et al., 2009]. Рассматриваются предположения о том, что когнитивно-моторные способности и интеллект могут иметь один и тот же нервный субстрат [Duncan, 2005]. Было показано, что у людей с более высоким уровнем интеллекта имеет место более выраженная супрессия высокочастотного альфа-ритма в центральных и теменных областях левого полушария во время выполнения задачи на вращение объектов в воображении [Doppelmayr et al., 2005]. А при наблюдении за движениями, осуществляемыми с помощью компьютерной мыши, наиболее выраженная десинхронизация мю-ритма наблюдается у испытуемых с высоким уровнем развития общего интеллекта [Аликина и др., 2018]. Также ранее нами была выявлена положительная связь между показателями общего интеллекта и значениями частоты максимального пика мощности альфа-ритма в лобных отведениях правого полушария (отведения F4 и F8), т.е. в зонах коры с предположительно “зеркальными” свойствами [Начарова и др., 2019].
Результаты опубликованных исследований носят довольно противоречивый характер и не позволяют эффективно дифференцировать вовлеченность “зеркальных” и более высокоуровневых когнитивных процессов в реализацию задач, в которых сопоставляется мозговая активность при выполнении, наблюдении и имитации движений. В связи с этим целью настоящей работы стал анализ источников генерации ЭЭГ-активности в частотном диапазоне 8–12 Гц (именуемой далее альфа-ритмом) у взрослых испытуемых, обладающих разной степенью развития общего интеллекта, в ходе выполнения ими задач, включающих самостоятельное выполнение, имитацию в режиме реального времени и наблюдение движений, осуществляемых с помощью компьютерной мыши. Мы предполагаем, что группы испытуемых, характеризующиеся разной степенью развития общего интеллекта, продемонстрируют паттерны активации неокортекса, различающиеся по степени вовлеченности областей коры, считающихся частью ЗСМ человека, и других структур головного мозга.
МЕТОДИКА
В исследовании приняли участие 62 человека (40 женщин и 22 мужчины) в возрасте от 17 до 35 лет, все правши. От испытуемых было получено информированное согласие на участие в экспериментах. Настоящее исследование соответствовало этическим принципам Хельсинкской декларации и было одобрено этическим комитетом Крымского федерального университета им. В.И. Вернадского.
Оценка степени развития общего интеллекта проводилась с помощью теста “Стандартные прогрессивные матрицы Равена”. Этот тест требует выявления отношений между абстрактными графическими элементами и является одним из традиционных инструментов для измерения фактора g интеллекта [Равен и др., 2012]. Для оценки уровня интеллекта использовалась процентная шкала развития интеллекта, в соответствии с которой высчитывался процент успешно выполненных заданий относительно их общего количества.
Экспериментальная процедура. В ходе эксперимента испытуемый и экспериментатор находились за расположенными рядом столами, экспериментатор справа. На каждом столе были размещены монитор и компьютерная мышь. На мониторе перед испытуемым с помощью веб-камеры демонстрировалось цветное видеоизображение участка поверхности стола экспериментатора с расположенной на нем мышью. Запись ЭЭГ производилась последовательно в следующих экспериментальных ситуациях, каждой из которых предшествовала регистрация в состоянии относительного покоя при зрительной фиксации статичного видеоизображении компьютерной мыши на столе экспериментатора (“фон”):
1. Ситуация “Наблюдение”. Испытуемый наблюдал за видеоизображением круговых движений правой руки экспериментатора с компьютерной мышью, выполняемых с переменной скоростью, и слышал характерные для этого действия звуки.
2. Ситуация “Движение”. Испытуемому давалась инструкция выполнять самостоятельные круговые движений мышью по часовой стрелке с произвольной скоростью. При этом необходимо было удерживать позицию курсора мыши в рамках заданной области монитора размером 25 × 25 см. Во время совершения движений испытуемый видел управляемый им курсор на фоне неподвижного видеоизображения компьютерной мыши и стола экспериментатора.
3. Ситуация “Имитация” в режиме реального времени круговых движений экспериментатора с помощью компьютерной мыши с переменной скоростью.
Перед экспериментальной ситуацией “Наблюдение” испытуемым давалась инструкция подсчитать количество движений против часовой стрелки. При этом для единообразия ситуаций “Наблюдение” и “Имитация” движения экспериментатор выполнял только по часовой стрелке. Наличие инструкции по подсчету движений против часовой стрелки, по нашему мнению, обеспечивало устойчивость внимания испытуемого на всем протяжении выполнения задания в ситуации “Наблюдение”.
Продолжительность каждой из фоновых и экспериментальных записей составляла 30 с.
Регистрация и анализ ЭЭГ. ЭЭГ регистрировалась при помощи 24-канального энцефалографа “Нейрон-Спектр-3” (фирма “Нейрософт”, Иваново) в диапазоне частот от 1 до 30 Гц. ЭЭГ-потенциалы отводились при помощи чашечковых хлор-серебряных электродов монополярно от фронтальных (Fp1, Fp2, F3, F4, Fz, F7, F8), центральных (С3, С4, Cz), височных (T3, T4, T5, T6), теменных (P3, P4, Pz), затылочных (O1, O2) локусов в соответствии с международной системой 10-20. В качестве референтного электрода служили объединенные датчики, закрепленные на мочках ушей. Частота оцифровки ЭЭГ-сигналов составляла 1000 Гц. Участки записей ЭЭГ с амплитудой более 250 мкВ, а также отрезки, содержащие большое количество артефактов, исключались из обработки.
Определение локализации источников генерации ритмов ЭЭГ осуществлялось при помощи метода электромагнитной томографии мозга низкого разрешения sLORETA, который характеризуется минимальной погрешностью в определении их положения [Pascual-Marqui, 2002]. Метод sLORETA реализован в одноименном программном обеспечении (https://www.uzh.ch/keyinst/loreta.htm). С учетом частоты оцифровки и количества каналов записей ЭЭГ безартефактные отрезки длиной 20–30 с преобразовывались к входному формату sLORETA. Частотный анализ ЭЭГ проводился с помощью программного обеспечения sLORETA, предполагающего применение алгоритма быстрого преобразования Фурье с эпохой анализа 5 с. Были выделены частотные диапазоны дельта-ритма (1–4 Гц), тета-ритма (4–8 Гц), альфа-ритма (8–12 Гц), бета-ритма (13–30 Гц) и гамма-ритма (35–45 Гц). Для дальнейшего анализа использовался только диапазон альфа-ритма, для которого в результате обработки был сформирован набор slor-файлов, характеризующих мозговую активность (положение и интенсивность источников тока) во время каждой экспериментальной ситуации. В результате обработки ЭЭГ строилась 3D карта мозга, состоящая из 2394 вокселей, имеющая пространственное разрешение 7 мм. Поля Бродмана (ПБ) описывались в системе трехмерных [x, y, z] координат Монреальского неврологического института с поправкой системы Талайраха.
Статистический анализ данных локализации источников тока проводили в пакете программы sLORETA с помощью метода статистического непараметрического картирования (statistical non-parametrical mapping (SnPM)) (Pascual-Marqui, 2002). Для расчета изменения спектральной плотности источников в парах экспериментальных ситуаций (“Наблюдение” и “Фон”, “Движение” и “Фон”, “Имитация” и “Фон”) был использован регрессионный анализ повторных измерений для связанных выборок (A – A2). В каждой паре ситуаций была рассчитана разница логарифмов спектральной плотности источников тока: из осцилляторных ответов, относящихся к событию А (“Фон”), вычитались осцилляторные ответы, относящиеся к событию А2 (“Наблюдение”, “Движение”, “Имитация”). В результате для каждой группы и пары экспериментальных ситуаций были определены зоны со значимо уменьшенной плотностью источников генерации альфа-ритма. Уменьшение плотности источников генерации альфа-ритма в указанных ситуациях в определенном корковом регионе рассматривалось как его активация в связи с тем, что мощность данного ритма отрицательно коррелирует с уровнем оксигенизации крови соответствующих участков коры [de Munck et al., 2009].
Для расчета различий в распределении плотности источников генерации ритма в группах с разной степенью развития интеллекта был использован анализ повторных измерений для несвязанных выборок (А –А2) – (В – В2), где пары выборок А и А2, В и В2 не связаны и характеризуют мозговую активность в группах с разной степенью развития интеллекта. Статистическая достоверность результатов была оценена с использованием теста рандомизации, который включает в себя поправку на множественные сравнения. В качестве достоверных рассматривались эффекты на уровне значимости p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
По результатам индивидуальной оценки показателей общего интеллекта и в соответствии с руководством к тесту выборка участников была разделена на три группы. Первая группа включала 20 человек, успешно выполнивших 25–74% заданий. Уровень интеллекта данной группы характеризовался как “средний” (ИС). Средний возраст участников данной группы составил 22.0 ± 4.0 года, а показатель интеллекта – 66.7 ± 9.4% (здесь и далее приведены средние значения и стандартное отклонение). Вторая группа состояла из 33 испытуемых, успешно выполнивших 75–90% заданий (интеллект “выше среднего” – ИВС). Средний возраст группы – 20.7 ± 2.7 года, показатель интеллекта – 82.8 ± 4.6. В третью группу входили 9 человек, успешно выполнивших более 90% заданий (интеллект “высокий” – ИВ). Средний возраст группы – 21.8 ± 4.4 года, показатель степени развития интеллекта – 93.7 ± 3.4%. Значимых различий по возрасту в группах обнаружено не было.
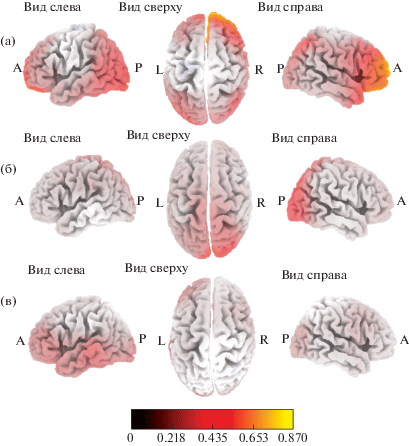
Уменьшение плотности источников генерации альфа-ритма во время наблюдения за выполнением движений. У испытуемых всех групп во время наблюдения за выполнением движений (в сравнении с “фоновой” записью) наблюдались изменения плотности источников генерации альфа-ритма (рис. 1), достигающие уровня статистической значимости. Поля Бродмана, значимо активированные в данной и других исследуемых ситуациях, представлены в табл. 1. В группе ИС уменьшение плотности источников генерации альфа-ритма в ситуации “Наблюдение” происходило в верхней и средней височных извилинах (ПБ 22, 39), островковой доле (ПБ 13) и надкраевой извилине (ПБ 40) правого полушария (рис. 1 (а)). В группе ИВС значимое уменьшение плотности источников происходило билатерально в поясной извилине (ПБ 24, 32) и медиальной лобной извилине (ПБ 9, 10) правого полушария (рис. 1 (б)). В группе ИВ во время наблюдения происходила значимая активация угловой извилины (ПБ 39) и нижней теменной дольки (ПБ 39) правого полушария (рис. 1 (в)). Различий в активации корковых регионов в исследуемых группах выявлено не было.
Рис. 1.
Локализация областей с уменьшенной по сравнению с “фоновой” записью плотностью источников генерации альфа-ритма при наблюдении за выполнением движений у испытуемых со средней (а), выше среднего (б) и высокой (в) степенью развития интеллекта.
Fig. 1. Localization of cortex areas with a decreased alpha rhythm current source density (relative to the baseline condition) when observing others’ movements, in subjects with different IQ levels: average (a), higher than average (б) and high (в).
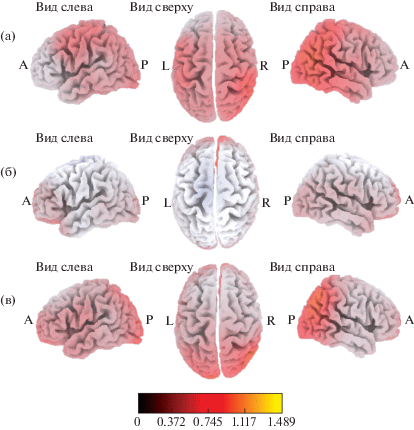
Таблица 1.
Локализация областей с значимым уменьшением плотности источников генерации альфа-ритма по сравнению с “фоновой” записью Table 1. Localization of cortex regions with significantly decreased alpha rhythm current source density values relative to the baseline condition
| Ситуация | Степень развития интеллекта | Максимальная интенсивность активации вокселей | Поля Бродмана | |
|---|---|---|---|---|
| Левое полушарие | Правое полушарие | |||
| “Наблюдение” | средняя | 0.92 | 22, 39, 13, 40 | |
| выше среднего | 0.59 | 24, 32, 10, 9 | ||
| высокая | 0.92 | 39 | ||
| “Движение” | средняя | 0.82 | 40 | |
| выше среднего | 0.87 | 6, 9, 13, 44, 4, 3, 45, 1, 8, 2, 43, 40, 24, 42, 32, 22 | 1, 13, 2, 3, 40, 4, 6, 9, 41, 39, 24, 31, 44, 32, 22, 21, 42, 45, 8, 37, 5, 10, 19, 20, 46, 7, 36, 35, 18, 25 | |
| высокая | 1.39 | 19, 7, 6, 4, 5, 3 | 39, 31, 19, 40, 7, 13, 6, 37, 22, 21, 5, 3, 4, 20, 1 | |
| “Имитация” | средняя | 1.16 | 24, 32, 19, 18 | 40, 31, 24, 32, 10, 39, 7, 9, 5, 25, 22, 13, 11, 2, 47, 19, 29, 3, 23, 27, 18, 30 |
| выше среднего | 1.11 | 6, 9, 13, 4, 44, 3, 45, 1, 2, 7, 8, 43, 40, 31, 5, 23, 19, 24, 25, 39, 17, 45, 22, 18, 42, 30, 33, 34, 11, 13, 46, 37 | 31, 39, 22, 13, 7, 23, 3, 2, 30, 40, 4, 18, 5, 1, 19, 37, 41, 21, 17, 20, 6, 17, 29, 24, 27, 37, 36, 42, 32, 10, 9, 25, 47, 11, 38, 28, 35, 33, 43, 34 | |
| высокая | 1.52 | 19, 7, 18, 6, 5, 3, 4 | 39, 31, 18, 7, 30, 40, 37, 19, 13, 21, 22, 5, 3, 2 | |
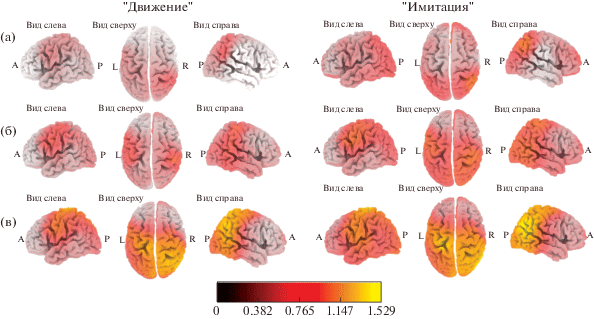
Уменьшение плотности источников генерации альфа-ритма во время выполнения и имитации движений (относительно “фоновой” записи). В ситуациях “Движение” и “Имитация” происходила активация теменно-височных областей правого полушария и лобно-центральных областей левого полушария (рис. 2, табл. 1). Наиболее выраженная активация корковых регионов происходила в группе ИВ, тогда как наибольшее число активированных структур было выявлено в группе ИВС.
Рис. 2.
Локализация областей с уменьшенной по сравнению с “фоновой” записью плотностью источников генерации альфа-ритма в ситуациях “Движение” и “Имитация” у испытуемых с средней (а), выше среднего (б) и высокой (в) степенью развития интеллекта.
Fig. 2. Localization of cortex areas with a decreased alpha rhythm current source density (relative to the baseline condition) when executing (left) and imitating (right) movements, in subjects with different IQ levels: average (a), higher than average (б) and high (в).
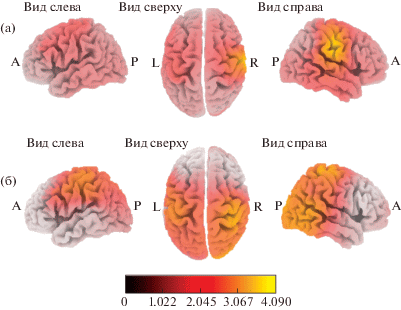
Сравнение активации корковых структур во время выполнения движений в группах с разным уровнем интеллекта. Различия в активации структур в ситуации “Движение” были выявлены в группах ИС и ИВС, а также групп ИС и ИВ. В обоих случаях у испытуемых с более высокой степенью развития интеллекта (ИВС и ИВ) происходила большая активация корковых структур во время выполнения движений по сравнению с активацией структур в группе ИС. В группе ИВС происходила более выраженная активация постцентральной (ПБ 1, 2, 3, 40), прецентральной (4, 6, 43) извилин, нижней теменной дольки (ПБ 40), верхней височной и поперечной височной извилин (ПБ 42) и островковой доли (ПБ 13) правого полушария по сравнению с активацией структур в группе ИС (рис. 3 (а)). В группе ИВ происходила значимо большая по сравнению с группой ИС активация постцентральной (ПБ 2, 40), прецентральной (ПБ 4), клинообразной (ПБ 19), средней затылочной (ПБ 18) и верхней затылочной (ПБ 19) извилин правого полушария (рис. 3 (б)). Статистически значимых различий в корковой активации структур в группах ИВС и ИВ найдено не было.
Рис. 3.
Локализация областей с увеличенной активацией во время выполнения движений (в сравнении с активацией в группе со средней степенью развития интеллекта) в группах со степенью развития интеллекта выше среднего (а) и высоким уровнем интеллекта (б).
Fig. 3. Localization of cortex areas more activated when executing movements in higher than average IQ (a) and high IQ (б) subjects, relative to the average IQ subjects.
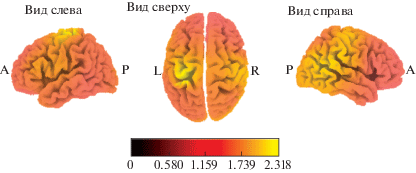
Сравнение активации корковых структур во время имитации движений в группах с разным уровнем интеллекта. Были выявлены различия в активации структур во время имитации движений экспериментатора (относительно “фоновой” записи) в группах ИС и ИВ. У испытуемых с высоким уровнем интеллекта происходила большая активация прецентральной (ПБ 4, 6), поясной (ПБ 24, 31) и постцентральной (ПБ 3) извилины левого полушария и верхней височной извилины (ПБ 22) правого полушария (рис. 4). Статистически значимых различий в корковой активации структур во время имитации движений (относительно “фоновой” записи) в группах ИС и ИВС, а также ИВС и ИВ найдено не было.
Рис. 4.
Локализация областей с увеличенной активацией во время имитации движений в группе с высокой степенью развития интеллекта в сравнении с активацией в группе со средней степенью развития интеллекта.
Fig. 4. Localization of cortex areas more activated when executing movements in high IQ subjects, relative to the average IQ subjects.
Уменьшение плотности источников генерации альфа-ритма во время имитации движений по сравнению с ситуацией самостоятельного выполнения движений. Во время имитации движений экспериментатора в группе ИС была выявлена дополнительная относительно ситуации “Движение” двусторонняя активация поясной извилины (ПБ 10, 24, 32) и медиальной лобной извилины (ПБ 10), а также верхней лобной (9, 10, 11), средней лобной (ПБ 10, 11), нижней лобной (ПБ 11, 47), глазничной (ПБ 11, 47) и прямой (ПБ 11) извилин правого полушария (рис. 5 (а)). В группе ИВС регистрировалась дополнительная активация предклинья (ПБ 31), клинообразной извилины (ПБ 7, 17, 18, 23, 31), поясной извилины (23, 30, 31) и средней затылочной извилины (ПБ 18) правого полушария (рис. 5 (б)). В группе ИВ значимых различий в активации структур во время ситуаций “Движение” и “Подражание” выявлено не было (рис. 5 (в)).
Рис. 5.
Локализация областей с уменьшенной по сравнению с ситуацией “Движение” плотностью источников генерации альфа-ритма при синхронной имитации движений у испытуемых со средней (а), выше среднего (б) и высокой (в) степенью развития интеллекта.
Fig. 5. Localization of cortex areas less activated in average IQ (a), higher than average IQ (б) and high IQ (в) subjects under condition of movement imitation, relative to the condition of movement execution.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Во всех исследуемых ситуациях и экспериментальных группах наиболее выраженная активация регистрировалась в структурах правого полушария, что, как мы полагаем, может объясняться спецификой условий выполнения экспериментальных задач. Полагают, что процессы дифференциации собственных и наблюдаемых действий, а также эффективной зрительно-моторной координации (с учетом их общей продолжительности) в существенной степени обеспечиваются через механизмы контроля, локализованные в правом полушарии [Uddin et al., 2006. Tognoli et al., 2015]. Одной из наиболее вовлеченных структур оказалась область височно-теменного узла (ПБ 7, 21, 22, 38, 39, 40), расположенная на стыке височных и теменных областей и прилегающая к задней части боковой борозды. В ряде исследований было показано, что височно-теменной узел правого полушария играет ключевую роль в анализе и сравнении собственных действий и сигналов внешней среды [Krall et al., 2014] и задействован в процессы идентификации и обработки социально значимых сигналов [Santiesteban et al., 2012; Krall et al., 2014].
Следует подчеркнуть, что в “эталонной” для исследований ЗСМ ситуации зрительного восприятия биологического движения набор значимо активированных структур оказался различен для каждой из экспериментальных групп, а предполагаемая роль этих структур в процессах социального взаимодействия зачастую описывается как носящая разнонаправленный характер. Так, выявленная в группе ИС активация верхней и средней височных и надкраевой извилин (область височно-теменного узла) в ситуации “Наблюдение” может быть связана как с процессами активации ЗСМ [Jeon et al., 2018], так и с обеспечением механизма различения собственных и наблюдаемых действий и контролем имитации и активации ЗСМ [Brass et al., 2009; Duffy et al., 2019]. Одной из попыток объяснить данную мультифункциональность является гипотеза “распределенного” имитационного контроля, согласно которой нейронные сети, обеспечивающие понимание действий других, и система контроля имитации в значительной степени перекрываются в зоне височно-теменного узла [Spengler et al., 2009]. При этом в недавних исследованиях было показано, что активация областей височно-теменного узла правого полушария скорее ингибирует, чем облегчает социальную имитацию [Duffy et al., 2019]. К структурам, обеспечивающим контроль активации “зеркальной” системы мозга, относят и значимо активирующуюся в группе ИС островковую долю правого полушария [Campbell et al., 2018]. Согласно результатам ранее опубликованных исследований, данная область может быть ответственна за выявление несоответствий выполняемых движений – наблюдаемым [Harding et al., 2015; Campbell et al., 2018].
Структуры, значимо активирующиеся в группе ИВС, также связаны с процессами восприятия и сопоставления собственных действий и эмоций с наблюдаемыми и контролем имитации. Имеются данные о том, что медиальные префронтальные области обеспечивают непрерывную оценку собственного поведения и его сопоставление с социальными нормами, причем суждения о себе связаны с активацией вентральной части медиальной префронтальной области, включающей вентральную часть медиальной лобной и поясной извилин (ПБ 10 и 32), а суждения, связанные с другими, связаны с дорсальными областями префронтальной коры (ПБ 8 и 9) [Denny et al., 2012]. Как следствие, у пациентов с повреждениями этих зон наблюдаются нарушения распознавания действий, эмоций и дефицит эмпатии, которые критически необходимы для реализации социальных взаимодействий и самоконтроля [Zald et al., 2010, Варламов и др., 2019]. В ряде исследований [Shamay-Tsoory et al., 2003; Keenan et al., 2000] было продемонстрировано, что процессы формирования эмпатического ответа более тесно связаны с активацией медиальной лобной извилины правого полушария, хотя данная закономерность выявляется не всегда [Denny et al., 2012]. Отдельными исследователями сделан вывод об ингибирующем влиянии медиальной префронтальной коры на фронтальный узел ЗСМ, локализованный в нижней лобной извилине [Cross et al., 2013]. В частности, выдвигается предположение о связи активации медиальных префронтальных областей с генерацией движений, не соответствующих наблюдаемым [Cross et al., 2013].
В группе ИВ во время наблюдения происходила значимая активация угловой извилины (ПБ 39) и нижней теменной дольки (ПБ 39) правого полушария, также являющихся частью височно-теменного узла. Активация нижней теменной дольки правого полушария во многих исследованиях связывается с восприятием собственного тела, а ее поражение приводит к нарушению “схемы тела” и трудностям осознания как собственных ощущений, так и идентификации эмоций и намерений, принадлежащих другим людям [Uddin et al., 2006]. В большинстве исследований нижнюю теменную дольку рассматривают как одну из основных структур ЗСМ [Molenberghs et al., 2012], однако имеются публикации, подчеркивающие ее более частную роль в развитии моторного резонанса, которая заключается в сопоставлении собственных и наблюдаемых действий [Brass et al., 2009]. Отдельные авторы указывают на ее контролирующую функцию [Duffy et al., 2019].
Несмотря на вовлеченность в процессы наблюдения отдельных структур ЗСМ, активация “классических” премоторных зон ЗСМ в данной ситуации не наблюдается ни в одной из экспериментальных групп, а все активированные структуры так или иначе связаны с процессами восприятия, различения собственных и наблюдаемых действий и контроля имитации. На наш взгляд, вовлеченность структур, связанных с различением наблюдаемых действий и контролем имитации, закономерна во время наблюдения за движениями без их одновременной имитации. Учитывая отсутствие различий в активации структур при попарном сравнении экспериментальных групп, можно сделать вывод о сопоставимой вовлеченности системы контроля имитации в группах с разным уровнем интеллекта.
Во время самостоятельного выполнения движений и имитации движений экспериментатора в режиме реального времени пространственные паттерны корковой активации относительно однотипны и выражаются в более высоком уровне вовлеченности теменно-височных структур правого полушария и лобно-центральных структур левого полушария в группах с более высокой степенью развития общего интеллекта.
Примечательно, что структуры, значимо более активированные в группах ИВС и ИВ во время выполнения самостоятельных движений (по сравнению с активацией структур в группе ИС), располагаются исключительно в правом полушарии и, предположительно, связаны с процессами переработки тактильной информации и концентрацией внимания на выполняемых движениях у испытуемых с высокими показателями общего интеллекта. Так, наиболее выраженное уменьшение плотности источников генерации альфа-ритма в этих группах происходит в постцентральной извилине правого полушария, принимающей участие в тактильной обработке текстуры и скорости объекта во время совершения движений [Yang et al., 2017], и интеграции сенсорной и двигательной информации [Kropf et al., 2018]. Значимо бо́льшая активация прецентральной извилины правого полушария в группах ИВС и ИВ во время осуществления самостоятельных движений, предположительно, могла быть следствием бо́льшей вовлеченности испытуемых с более высоким уровнем интеллекта в процесс выполнения экспериментальной задачи.
Интересно отметить, что если во время выполнения самостоятельных движений в группе ИВ происходила бо́льшая по сравнению с группой ИС активация прецентральной и постцентральной извилин в правом полушарии, то во время имитации движений эти же структуры больше активируются в левом, контралатеральном правой руке, которой выполнялись движения. Как известно, прецентральная извилина, включающая в себя первичную и дополнительную моторные области, участвует в процессах планирования и непосредственном выполнении произвольных движений [Banker et al., 2019]. И именно в передней части прецентральной извилины (ПБ 6), предположительно, располагаются нейроны, обладающие “зеркальными” свойствами [Molenberghs et al., 2012]. Постцентральная извилина, значимо больше активирующаяся в группе ИВ, по сравнению с группой ИС, во время имитации движений экспериментатора, по результатам ранее опубликованного мета-анализа [Molenberghs et al., 2012], является областью локализации т.н. “тактильных” зеркальных нейронов и активируется во время восприятия действий, предполагающих тактильный контакт с каким-либо объектом. Тем не менее нельзя исключать возможности интерпретации большей активации постцентральной извилины в группе ИВ как показателя более высокоуровнего восприятия и анализа скорости собственных движений во время совершения движений [Yang et al., 2017], а также интеграции процессов сенсорной обработки и моторного контроля [Kropf et al., 2018]. Значимо больше активирующаяся в группе ИВ передняя часть поясной извилины характеризуется прочными связями с префронтальными, дополнительной моторной и теменными областями и участвует в непрерывном контроле движений [Paus, 2001; Duffy et al., 2019]. Помимо этого, некоторые исследователи указывают на роль дорсальной части передней поясной извилины в процессах обнаружения и оценки социальных процессов [Dedovic et al., 2016]. Показано, что верхняя височная борозда правого полушария участвует в распознавании намерений, лежащих в основе действий путем сравнения собственных и наблюдаемых действий [Jiang et al., 2017]. Совместно с активацией премоторных областей эта структура может отражать активацию ЗСМ [Molenberghs et al., 2012]. Данная структура также является частью височно-теменного узла, а ее активация может быть связана с процессами контроля имитации. Исходя из данных мета-анализа [Molenberghs et al., 2012] все перечисленные области являются частью ЗСМ человека и содержат нейроны, обладающие “зеркальными” свойствами [Mukamel et al., 2010].
Таким образом, можно предположить, что более выраженная активация перечисленных структур у испытуемых с высоким уровнем развития интеллекта во время синхронной имитации движений руки может быть связана как с более активными процессами восприятия и атрибуции наблюдаемых действий (структуры правого полушария), так и с большей активацией ЗСМ.
Более сложными представляются попытки интерпретировать дополнительную активацию корковых структур во время имитации движений экспериментатора по сравнению с их самостоятельным выполнением. Если в группе ИС происходит большая активация фронтальных областей, а в группе ИВС – затылочных, то у испытуемых с высокой степенью развития интеллекта значимых различий в активации структур во время имитации и выполнения движений выявлено не было. Лобные области, активирующиеся в группе ИС, относят как к структурам ЗСМ [Molenberghs et al., 2012], так и к системе контроля активации ЗСМ [Cross et al., 2013; Harding et al., 2015; Campbell et al., 2018]. При этом активирующуюся часть нижней лобной извилины (ПБ 11), в отличие ПБ 44, 45, не связывают с активностью ЗСМ [Jeon et al., 2018]. Несмотря на то что в передней части поясной извилины были найдены нейроны с “зеркальными” свойствами [Mukamel et al., 2010], их количество не превышало порог статистической ошибки. Более того, в более поздних исследованиях подчеркивается участие данной области в процессах нахождения ошибок и контроля имитации [Cross et al., 2013; Campbell et al., 2018]. Таким образом, можно предположить, что дополнительная активация лобных областей во время имитации движений в группе ИС скорее связана с более детальным анализом и контролем движений, чем с активацией ЗСМ.
Дополнительная активация теменно-затылочных структур правого полушария в группе ИВС может быть связана с необходимостью дополнительной обработки визуальной информации и моторной координации во время имитации движений экспериментатора. Так, клинообразная и средняя затылочная извилины вовлечены в базовые процессы обработки зрительной информации, а предклинье участвует в процессах зрительного внимания при совершении движений и переключении внимания между наблюдаемыми объектами движения [Cavanna et al., 2006]. Задняя поясная извилина также может способствовать успешному повторению движений экспериментатора, обеспечивая нисходящий контроль зрительного внимания, движений глаз и выполняемых движений [Leech et al., 2014].
Отсутствие различий в активации структур в группе ИВ во время выполнения и имитации движений экспериментатора, на наш взгляд, может быть связано с отсутствием потребности в дополнительном анализе параметров движений и может свидетельствовать о развитии моторного резонанса в данной группе. Это предположение подтверждается выявленной в данной группе дополнительной активацией премоторных и моторных структур левого полушария во время имитации движений. В то же время отсутствие межгрупповых различий в ситуации “Имитации” может свидетельствовать о необходимости дальнейшего анализа.
ВЫВОДЫ
1. Обнаружена связь между индивидуальным уровнем развития общего интеллекта и различиями в пространственных паттернах активации коры головного мозга во время выполнения и имитации движений. При этом выявленные связи носят сложный характер. Наблюдается более выраженная и более локализованная активация отдельных структур неокортекса у взрослых испытуемых, имеющих высокую степень развития общего интеллекта.
2. Различия между группами участников, характеризовавшихся разным уровнем развития общего интеллекта, в большей степени опосредуют структуры правого полушария, участвующие в процессах зрительно-моторной координации и восприятии действий, выполняемых другими людьми, и различении собственных и чужих действий.
3. Бо́льшая вовлеченность прецентральной, поясной и постцентральной извилин левого полушария в процесс имитации движений экспериментатора в группе с высокой степенью развития интеллекта может указывать на бо́льшую активацию “зеркальной” системы мозга.
Исследование выполнено с использованием оборудования ЦКП “Экспериментальная физиология и биофизика” КФУ им. В.И. Вернадского и при финансовой поддержке РФФИ в рамках научного проекта № 18-015-00074.
Список литературы
Аликина М.А., Махин С.А., Павленко В.Б. Люди с высоким уровнем общего интеллекта демонстрируют более выраженную десинхронизацию мю-ритма при наблюдении за действиями других. Ученые записки Крымского федерального университета им. В.И. Вернадского. Биология. Химия. 2018. 4 (3): 26–34.
Варламов А.А., Портнова Г.В., Макглоун Ф.Ф. С-тактильная система и нейробиологические механизмы “эмоционального” тактильного восприятия: история открытия и современное состояние исследования. Журнал высшей нервной деятельности им. И.П. Павлова. 2019. 69 (3): 280–293.
Кайда А.И., Махин С.А., Эйсмонт Е.В., Павленко В.Б. Возрастная динамика и топография реактивности индивидуального мю-ритма ЭЭГ у детей 4–14 лет. Вестник Томского государственного университета. Биология. 2019. 45: 106–127.
Лебедева Н.Н., Зуфман А.И., Мальцев В.Ю. Система зеркальных нейронов мозга: ключ к обучению, формированию личности и пониманию чужого сознания. Успехи физиологических наук. 2017. 48 (4): 16–28.
Лебедева Н.Н., Каримова Е.Д., Карпычев В.В., Мальцев В.Ю. Зеркальная система мозга при наблюдении, выполнении и представлении моторных задач – нейрофизиологическое отражение восприятия чужого сознания. Журнал высшей нервной деятельности им. И.П. Павлова. 2018. 68 (2): 204–215.
Махин С.А., Макаричева А.А., Луцюк Н.В., Павленко В.Б. Исследование реактивности μ-ритма при наблюдении, слуховом восприятии и имитации движений: взаимосвязь со свойствами личности, определяющими эмпатию. Физиология человека. 2015. 41 (6): 28–35.
Начарова М.А., Махин С.А., Павленко В.Б. Особенности взаимосвязи между индивидуальной пиковой частотой альфа-ритма ЭЭГ и характеристиками общего интеллекта. Ученые записки Крымского федерального университета им. В.И. Вернадского. Биология. Химия. 2019. 5 (2): 132–144.
Равен Дж.К., Корт Дж.Х. Руководство к Прогрессивным Матрицам Равена и Словарным Шкалам. М.: Когито-Центр, 2012. 144 с.
Anwar M.N., Navid S.N., Khan M., Kitajo K. A possible correlation between performance IQ, visuomotor adaptation ability and mu suppression. Brain Research. 2015. 1603 (7): 84–93.
Banker L., Tadi P. Neuroanatomy. Precentral Gyrus. StatPearls Publishing. 2019.
Brass M., Ruby P., Spengler S. Inhibition of imitative behaviour and social cognition. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009. 364 (1528): 2359–2367.
Campbell M.E.J., Mehrkanoon S., Cunnington R. Intentionally not imitating: Insula cortex engaged for top-down control of action mirroring. Neuropsychologia. 2018. 111: 241–251.
Candidi M., Urgesi C., Ionta S., Aglioti S.M. Virtual lesion of ventral premotor cortex impairs visual perception of biomechanically possible but not impossible actions. Social Neuroscience. 2008. 3 (3–4): 388–400.
Cavanna A., Trimble M. The precuneus: a review of its functional anatomy and behavioural correlates. Brain. 2006. 129 (3): 564–583.
Cross K.A., Torrisi S., Reynolds Losin E.A., Iacoboni M. Controlling automatic imitative tendencies: Interactions between mirror neuron and cognitive control systems. NeuroImage. 2013. 83: 493–504.
de Munck J.C., Gonçalves S.I., Mammoliti R., Heethaar R.M., Lopes da Silva F.H. Interactions between different EEG frequency bands and their effect on alpha–fMRI correlations. NeuroImage. 2009. 47 (1): 69–76.
Dedovic K., Slavich G.M., Muscatell K.A., Irwin M.R., Eisenberger N.I. Dorsal Anterior Cingulate Cortex Responses to Repeated Social Evaluative Feedback in Young Women with and without a History of Depression. Frontiers in Behavioral Neuroscience. 2016. 10: 64–76.
Denny B.T., Kober H., Wager T.D., Ochsner K.N. A Meta-Analysis of Functional Neuroimaging Studies of Self and Other Judgments Reveals a Spatial Gradient for Mentalizing in Medial Prefrontal Cortex. J. Cogn. Neurosci. 2012. 24 (8): 1742–1752.
Doppelmayr M., Klimesch W., Hödlmoser K., Sauseng P., Gruber W. Intelligence related upper alpha desynchronization in a semantic memory task. Brain Research Bulletin. 2005. 66 (2): 171–177.
Duffy K.A., Luber B., Adcock R.A., Chartrand T.L. Enhancing activation in the right temporoparietal junction using theta-burst stimulation: Disambiguating between two hypotheses of top-down control of behavioral mimicry. PLOS ONE. 2019. 14 (1): e0211279.
Duncan J. Frontal Lobe Function and General Intelligence: Why it Matters. Cortex. 2005. 41 (2): 215–217.
Fox N.A., Bakermans-Kranenburg M.J., Yoo K.H., Bowman L.C., Cannon E.N., Vanderwert R.E., Ferrari P.F., van IJzendoorn M.H. Assessing human mirror activity with EEG mu rhythm: a meta-analysis. Psychol. Bull. 2016. 142 (3): 291–313.
Frenkel-Toledo S., Bentin S., Perry A., Liebermann D.G., Soroker N. Dynamics of the EEG power in the frequency and spatial domains during observation and execution of manual movements. Brain. Res. 2013. 1509: 43–57.
Harding I.H., Yücel M., Harrison B.J., Pantelis C., Breakspear M. Effective connectivity within the frontoparietal control network differentiates cognitive control and working memory. NeuroImage. 2015. 106: 144–153.
Hecht E.E., Parr L.A. The chimpanzee mirror system and the evolution of frontoparietal circuits for action observation and social learning. In: Ferrari P.F., Rizzolatti G., editors. New frontiers in mirror neurons research. Oxford: Oxford University Press; 2015. pp. 153–181.
Hobson H.M., Bishop D.V.M. The interpretation of mu suppression as an index of mirror neuron activity: past, present and future. R. Soc. open sci. 2017. 4 (3): 160662-83.
Jeon H., Lee S.-H. From Neurons to Social Beings: Short Review of the Mirror Neuron System Research and Its Socio-Psychological and Psychiatric Implications. Clinical Psychopharmacology and Neuroscience. 2018. 16 (1): 18–31.
Jiang J., Borowiak K., Tudge L., Otto C., von Kriegstein K. Neural mechanisms of eye contact when listening to another person talking. Soc Cogn Affect Neurosci. 2017. 12 (2): 319–328.
Keenan J.P., Wheeler M.A., Gallup G.G., Pascual-Leone A. Self-recognition and the right prefrontal cortex. Trends in Cognitive Sciences. 2000. 4 (9): 338–344.
Keysers C., Gazzola V. Social Neuroscience: Mirror Neurons recorded in Humans. Current Biology. 2010. 20 (8): 353–354.
Krall S.C., Rottschy C., Oberwelland E., Bzdok D., Fox P.T., Eickhoff S.B., Konrad K. The role of the right temporoparietal junction in attention and social interaction as revealed by ALE meta-analysis. Brain Structure and Function. 2014. 220 (2): 587–604.
Kropf E., Syan S.K., Minuzzi L., Frey B.N. From anatomy to function: the role of the somatosensory cortex in emotional regulation. Brazilian Journal of Psychiatry. 2018. 41 (3): 261–269.
Lago-Rodriguez A., Cheeran B.J., Koch G., Hortobagy T., Fernández-Del-Olmo M. The role of mirror neurons in observational motor learning: an integrative review. European Journal of Human Movement. 2014. 32: 82–103.
Leech R., Sharp D.J. The role of the posterior cingulate cortex in cognition and disease. Brain. 2014. 137 (1): 12–32.
Molenberghs P., Cunnington R., Mattingley J.B. Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neurosci. Biobehav. Rev. 2012. 36 (1): 341–349.
Mukamel R., Ekstrom A.D., Kaplan J., Iacoboni M., Fried I. Single-neuron responses in humans during execution and observation of actions. Curr. Biol. 2010. 20 (8): 750–756.
Pascual-Marqui R. Standardized low-resolution brain electromagnetic tomography (sLORETA): technical details. Methods Find. Exp. Clin. Pharmacol. 2002. 24 (Suppl D): 5–12.
Paus T. Primate anterior cingulate cortex: where motor control, drive and cognition interface. Nat. Rev. Neurosci. 2001. 2 (6): 417–424.
Raymaekers R., Wiersema J.R., Roeyers H. EEG study of the mirror neuron system in children with high functioning autism. Brain Research. 2009. 1304: 113–121.
Santiesteban I., Banissy M.J., Catmur C., Bird G. Enhancing social ability by stimulating right temporoparietal junction. Current Biology. 2012. 22 (23): 2274–2277.
Saygin A.P. Superior temporal and premotor brain areas necessary for biological motion perception. Brain: A Journal of Neurology. 2007. 130 (9): 2452–2461.
Shamay-Tsoory S.G., Tomer R., Berger B.D., Aharon-Peretz J. Characterization of Empathy Deficits following Prefrontal Brain Damage: The Role of the Right Ventromedial Prefrontal Cortex. Journal of Cognitive Neuroscience. 2003. 15 (3): 324–337.
Spengler S., von Cramon D.Y., Brass M. Control of shared representations relies on key processes involved in mental state attribution. Human Brain Mapping. 2009. 30 (11): 3704–3718.
Tognoli E., Kelso J.A. The coordination dynamics of social neuromarkers. Front. Hum. Neurosci. 2015. 20 (9): 563–578.
Uddin L.Q., Molnar-Szakacs I., Zaidel E., Iacoboni M. rTMS to the right inferior parietal lobule disrupts self–other discrimination. 2006. SCAN. 1 (1): 65–71.
Yang J., Kitada R., Kochiyama T., Yu Y., Makita K., Araki Y., Wu J., Sadato N. Brain networks involved in tactile speed classification of moving dot patterns: the effects of speed and dot periodicity. Sci. Rep. 2017. 7: 40931–40943.
Zald D.H., Andreotti C. Neuropsychological assessment of the orbital and ventromedial prefrontal cortex. Neuropsychologia. 2010. 48 (12): 3377–3391.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова