

Известия РАН. Серия географическая, 2020, T. 84, № 4, стр. 598-610
Отношения леса и степи в заповеднике “Шайтан-Тау” (Южный Урал)
А. В. Хорошев a, *, Г. М. Леонова a, Д. Е. Шарова a
a Московский государственный университет имени М.В. Ломоносова, географический факультет
Москва, Россия
* E-mail: avkh1970@yandex.ru
Поступила в редакцию 29.05.2019
После доработки 27.03.2020
Принята к публикации 06.04.2020
Аннотация
В условиях климатических изменений лесостепные ландшафты служат эффективной моделью для исследования смещений границ между лесными и степными сообществами и степени равновесности между фитоценозами и почвами при корректирующем вкладе рельефа. В статье исследуется проблема возможности нескольких равновероятностных состояний ландшафта на примере низкогорной лесостепи в государственном заповеднике “Шайтан-Тау” (Южный Урал, Оренбургская область). Проверялaсь гипотеза о существовании несоответствий между мобильными свойствами лесных и степных фитоценозов и более инертными свойствами почв. Построены дискриминантные модели, которые описывают зависимость распространения лесных и степных сообществ от положения в разных типах ландшафтных структур: геоциркуляционной (с использованием морфометрических показателей рельефа, характеризующих аккумуляцию и рассеяние стока), геостационарной (с использованием показателей расчлененности, характеризующих морфологические и генетические свойства рельефа и пород) и биоциркуляционной (с использованием показателей, характеризующих перераспределение солнечной радиации рельефом). Установлено наличие почвенно-фитоценотических признаков, индицирующих наступление или отступление лесных сообществ на участках с высокой вероятностью соответствия экологической нише лесов. В настоящее время степи способны захватывать территории в лесных местоположениях на платообразных участках под действием летнего потепления и учащения пожаров; при этом могут сохраняться свойства лесных почв. На северных склонах и в вогнутых местоположениях леса местами наступают на степи. Кустарники наименее своеобразны по занимаемой геоморфологической нише, примерно с равной вероятностью они встречаются в типично степных и типично лесных местообитаниях.
ВВЕДЕНИЕ
В старой проблеме взаимоотношений леса и степи на границе их ареалов есть два аспекта, стабильно вызывающих повышенный интерес. Во-первых, эта граница часто рассматривается как чуткий индикатор климатических изменений и степени инертности ландшафтов. Во-вторых, в ландшафтоведении решение вопроса об исходе взаимоотношений леса и степи связывается с проблемой возможности нескольких устойчивых состояний, предсказуемости критических порогов и принципиальной невозможности прогноза траектории развития после прохождения точки бифуркации [1, 4, 19]. Поведение лесостепного ландшафта вблизи критического состояния может быть задано случайными погодными колебаниями или внешними воздействиями типа пожаров и вспышек размножения насекомых (часто связанными с последовательностью погод), которые влияют на исход конкуренции двух типов растительности.
Известно, что при прочих равных условиях повышенная конкурентоспособность леса или степи может контролироваться рельефом. Простой пример для горных регионов – солярно-экспозиционные и циркуляционно-экспозиционные различия, которые задают самое общее правило “раздела территории” между конкурентами. В терминологии полиструктурности ландшафта [5] эту закономерность можно рассматривать как вклад лишь одного типа ландшафтных структур – биоциркуляционных. Однако абсолютизация их доминирования вряд ли оправдана, и можно привести немало примеров, когда реальное взаиморасположение лесных и степных урочищ гораздо сложнее. Следовательно, необходимо оценивать также вклады геоциркуляционных и геостационарных структур. Очевидно, что чувствительность типов растительности к климатическим изменениям может существенно отличаться в зависимости от вкладов трех перечисленных типов ландшафтных структур.
После введения в науку понятий “почва–память”, “почва–момент” [20], “характерное время” [2] усилилось внимание к возможности использовать несоответствия между современным фитоценозом и строением почвенного профиля для индикации смещений ландшафтных границ. При изучении отклика ландшафтов на климатические изменения именно разница характерных времен фитоценозов и почв, разделение современных и унаследованных, реликтовых почвенных признаков позволяют увидеть в пространстве образ процесса, происходящего во времени.
Ландшафты низкогорных лесостепей предоставляют богатые возможности для исследования обеих проблем – факторов смещения границы “лес–степь” при климатических изменениях и возможности нескольких устойчивых состояний ландшафта (иногда называемой “пластичностью”). Очевидно, что в лесостепных ландшафтах ключевое значение будут иметь тенденции изменения количества осадков и летних температур, определяющие влагозапас в почве в вегетационный период.
В южной части Восточного макросклона Урала и в Зауралье рост среднегодовых температур за период с 1976 по 2005 гг. составил 1.25–1.5°С [11, 14]. В целом для Южного Урала и Зауралья увеличение среднегодовых температур составило 2–3°С на 100 лет при росте годового количества осадков [23]. Отмечается слабая тенденция к росту осадков за 1976–2005 гг., причем преимущественно за холодный период [14]. На аридных территориях юга Европейской России ожидается увеличение годового количества осадков за счет зимних и весенних осадков, тогда как во второй половине лета и начале осени ожидается их снижение [8]. По данным ГГО для юга Уральского федерального округа прогнозируется рост осадков в 2011–2030 гг. зимой на 6 ± 4%, летом на 0 ± 5%; в период 2041–2060 гг. зимой на 13 ± 8%, летом на 0 ± 6% [9].
Зимние осадки на Урале определяются высокой циклональной активностью в зимние месяцы [10], которая, по некоторым данным, в Европейской России возрастает [17, 24]. Однако площадь залегания снежного покрова и его продолжительность повсеместно сокращаются, особенно в переходные сезоны за счет роста температур зимнего и весеннего сезонов [21]. По материалам ближайшей к заповеднику “Шайтан-Тау” метеостанции Акъяр (51°52′ с.ш., 58°13′ в.д.), расположенной на территории Башкирии в 56 км к северо-востоку, за последние пять лет отмечается рост доли зимних осадков. Доля летних осадков, напротив, сокращается.
Для лесостепных ландшафтов Урала наибольшее внимание привлекает тенденция роста зимних осадков, что должно благоприятствовать экспансии лесов, по крайней мере, в теневых позициях рельефа. С другой стороны, тенденция к повышению температур и зимнего увлажнения может быть причиной активизации степных пожаров из-за роста продуктивности степных фитоценозов: во влажные и прохладные годы активная вегетация растительности способствует накоплению значительных объемов сухой фитомассы, что впоследствии будет провоцировать распространение пожаров [16].
Таким образом, современные климатические тенденции, с одной стороны, могут способствовать улучшению условий для лесов, с другой – росту частоты летних и осенних пожаров, позволяющих степи выигрывать конкуренцию в пригодных для нее местоположениях рельефа. Кроме того, частые для заповедника “Шайтан-Тау” весенне-летние засухи нередко сопровождаются массовым размножением непарного шелкопряда; совместное действие засухи и объедания листьев непарным шелкопрядом приводит к сильному снижению прироста дуба черешчатого [6, 13]. Положение заповедника вблизи юго-восточной границы ареала дуба черешчатого и в целом европейских широколиственных лесов [3] является фактором повышенной чувствительности к внешним воздействиям.
Из этого следует, что взаиморасположение и пропорции лесных и степных участков в заповеднике “Шайтан-Тау” могут находиться в неоднозначной зависимости от климатических изменений. Выдвигается гипотеза о существовании несоответствий между мобильными свойствами фитоценозов и более инертными свойствами почв, что рассматривается как гипотетический индикатор смещений границ. Предполагается, что реакция растительного покрова на внешние воздействия (в том числе, климатические) может отчасти контролироваться и корректироваться условиями рельефа через перераспределение тепла, влаги, твердого и растворенного вещества.
Цель исследования – определение диапазона ландшафтных условий, в которых с наибольшей вероятностью могут происходить смещения границ между широколиственными лесами и степями в низкогорно-лесостепном ландшафте государственного заповедника “Шайтан-Тау”. Если в большинстве работ этот вопрос рассматривается применительно к фитоценозам, то мы ставим задачу шире и концентрируемся на оценке равновесности почвенно-фитоценотических отношений, варьирующих в зависимости от рельефа.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование проведено в низкогорно-лесостепном ландшафте по правобережью меридионального участка долины р. Сакмара в пределах Кувандыкского района Оренбургской области вблизи границы с Башкортостаном (рис. 1).
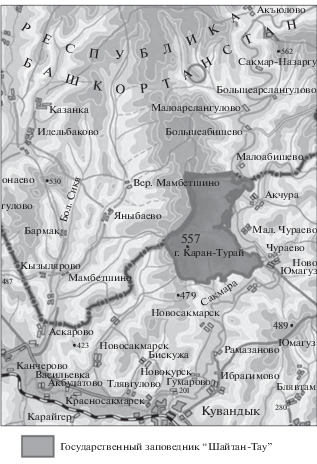
Литогенную основу густорасчлененного эрозионно-денудационного лесостепного ландшафта составляют вулканогенно-осадочные толщи, среди которых преобладают силурийские кремнистые сланцы при участии девонских туффитов и туфов базальтов. На остаточной поверхности пенеплена Зилаирского плато [22], представленной на водораздельном плато с высотами 520–550 м, сохранились верхнемеловые глауконитово-кварцевые пески и галечники. Под широколиственными лесами доминируют серые лесные почвы, под степными и кустарниковыми, как правило, ‒ черноземы выщелоченные, темногумусовые почвы или литоземы.
В автономных урочищах плато на хребте Дзяутобе представлены разнотравно-злаковые степи (Poa transbaicalica, Stipa pennata, Hieracium echioides, Salvia stepposa, Pulsatilla patens). На узких сильнокаменистых гребнях в петрофитных степях доминируют Poa transbaicalica, Stipa pennata, Tanacetum uralense, Scorzonera austriaca, Centaurea marschalliana. В слабовогнутых водосборных понижениях среди вершинных поверхностей обильно произрастает осиновая поросль, многочисленны заросли кустарников (Spiraea hypericifolia).
На склонах северной экспозиции лесные урочища представлены дубравами, дубово-липовыми и липово-березовыми лесами (Quercus robur, Tilia cordata, Ulmus glabra). Под ними сформировались серые, темно-серые почвы, хорошо оструктуренные, слабо- и среднекаменистые. Для почв этих урочищ характерна хорошая выраженность профиля и большая мощность (более 66 см). Небольшие площади на выпуклых участках заняты кустарниками (Spiraea hypericifoia, Cerasus fruticosa, Chamaecytisus ruthenicus) и мезофильными разнотравно-кустарниковыми лугами (Poa transbaicalica, Polygonum alpinum, Filipendula vulgaris, Seseli libanotis). На склонах “теплых” экспозиций дубравы занимают склоны лощин, секущих склон, тогда как степи на неполноразвитых темногумусовых почвах и литоземах встречаются на гребнях, выпуклых и вогнутых участках склонов и в пригребневых позициях.
Алгоритм исследования представлен на рис. 2. Материалы были получены в июне 2016–2018 гг. Составлено 196 ландшафтных описаний, включавших следующие данные: форма рельефа, уклон, экспозиция; почвообразующие породы; обилие видов фитоценоза по шкале Друде; высоты и проективное покрытие ярусов трав и кустарников; максимальные и средние диаметр и высота ярусов древостоя; мощности, цвет по шкале Манселла (Hue, Value, Chroma) и гранулометрический состав до глубины 50 см с измерениями через каждые 5 см, структура генетических горизонтов почв. Описания проведены во всех видах урочищ, репрезентативных для типов растительности, форм рельефа и разновидностей геологического строения. Для 131 фации по результатам опробования генетических горизонтов получены также данные химических анализов почв на содержание органического углерода (по Тюрину), гигроскопической влажности, рН водного, валового содержания микроэлементов (спектральный анализ), содержания обменных форм макроэлементов, общего азота, подвижного фосфора и калия (методами Кирсанова и Гедройца с использованием атомно-адсорбционного спектрометра “Квант-2АТ”).
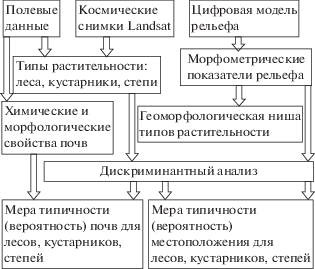
Цифровая модель рельефа (ЦМР) построена в программе SAGA GIS методом простого кригинга. Рассчитаны морфометрические показатели, характеризующие: а) латеральный перенос вещества в локальных окрестностях элемента ЦМР (крутизна, вертикальная и горизонтальная кривизны, индекс конвергенции); б) перераспределениe вещества на уровне местности (топографический позиционный индекс TPI и индекс расчлененности TRI в окрестностях 100, 300, …, 1100 м, водосборная площадь, топографический индекс влажности, эрозионный потенциал рельефа, превышение над водотоком); в) потенциальную теплообеспеченность в инсоляционном поле (доза солнечной радиации, продолжительность освещенности, экспозиция склонов).
Для формализованного выделения ареалов типов растительности использована классификация методом к-средних по космическим снимкам Landsat (29.08.2018, 14.05.2014, 18.06.2015). Результаты классификации снимков и обработки ЦМР сведены в базу данных для 227327 пикселов с разрешением 25 м.
Статистический анализ данных проводился средствами программы Statistica 7.0. Построена серия дискриминантных моделей (метод прямого пошагового выбора при F = 1), которые описывают зависимость распространения типов растительности (группирующая переменная) от положения в разных типах ландшафтных структур (независимые переменные). Для каждого пиксела рассчитана вероятность того, что при имеющейся комбинации морфометрических характеристик рельефа могут существовать степь, кустарники или лес. Методом дисперсионного анализа установлены специфические комбинации свойств рельефа, химических и цветовых свойств почв под лесными, степными и кустарниковыми сообществами.
При оценке значимости геостационарных структур в качестве независимых переменных дискриминантного анализа для распознавания трех типов растительности использованы показатели расчлененности рельефа, тесно связанные с генезисом рельефа и геологическим строением. Вклад геоциркуляционных структур оценивается включением в модель показателей перераспределения вещества. Для учета вклада биоциркуляционных структур в модель включаются показатели инсоляции и экспозиции (выраженные через косинус и синус азимута в радианах).
Проверены гипотезы, что для каждого типа растительности существует своеобразное, присущее только ему сочетание: а) морфометрических признаков рельефа (ниже – “геоморфологическая ниша”), б) химических свойств гумусового горизонта почв по концентрациям макро- и микроэлементов, а также рН, в) морфологических свойств почв по цветовым признакам.
Если в некотором наборе местоположений тот или иной тип растительности преобладает, то есть вероятность встречаемости высокая, то такие местоположения считались наиболее благоприятными, а приуроченность к ним – закономерной. Отсутствие типа растительности в благоприятных местоположениях рассматривалось как следствие действия внешних факторов. Проверялась гипотеза, что при росте благоприятности рельефа для данного типа растительности параллельно происходит рост вероятности того, что химические и морфологические свойства почв соответствуют типичным для него.
Разработанный вероятностный подход к определению геоморфологической ниши типов растительности имел целью сократить необходимость проверки большой серии гипотез о значимости частных геолого-геоморфологических факторов. Значение вероятности принадлежности к геоморфологической нише лесов, кустарников или степей интегрирует большую серию морфометрических показателей и учитывает вовлеченность каждой ландшафтной единицы одновременно в геостационарные, геоциркуляционные и биоциркуляционные структуры. В то же время есть возможность оценить частные вклады трех перечисленных типов структур в дифференциацию растительного покрова, сравнив качество моделей дискриминации, построенных для разных групп морфометрических показателей.
РЕЗУЛЬТАТЫ
Модель геотопологической детерминированности типов растительности
На территории заповедника степь не только закономерно занимает южные склоны, но и большую часть плато выше лесов. При этом часть плато также занято лесами, что заставляет предполагать равноправность двух типов растительности в данном местоположении, что отмечалось для региона еще И.М. Крашенинниковым [12]. Нигде не отмечены явные признаки обильного появления поросли главной лесообразующей породы – дуба – среди степи или среди кустарников, что согласуется с данными [15] для Приволжской возвышенности. При этом довольно часто кайму леса по северным и западным склонам и в водосборных понижениях образует поросль осины, которая, возможно, подготавливает условия для возобновления дубрав в типично лесных местоположениях, задерживая снег и сокращая физическое испарение.
Типичная геоморфологическая ниша степей соответствует либо слаборасчлененным выпуклым водораздельным поверхностям, либо склонам южных и восточных экспозиций со слабовыраженными условиями аккумуляции стока. Леса наиболее часто встречаются на крутых теневых склонах либо в вогнутых или пониженных мезоформах рельефа с максимально выраженными условиями аккумуляции стока при большой водосборной площади и минимальным превышением над тальвегами. Ниша кустарников соответствует промежуточным значениям TPI, высоким значениям TRI, максимально выраженным условиям рассеяния вещества с выпуклых каменистых гребней.
11 типов контуров, показанных на геологической карте заповедника [18], достаточно хорошо распознаются комбинацией показателей TPI и TRI в окрестности от 100 до 1100 м, вертикальной дистанции до тальвега, глубины примыкающих долин и локального базиса эрозии (Wilks’ Lambda 0.14; корректность классификации 57%). При этом практически идеально (на 98%) распознаются участки, сложенные меловыми отложениями на слаборасчлененной останцовой поверхности выравнивания. Поэтому в дальнейшем группа показателей расчлененности рельефа использовалась как индикатор ареалов геостационарных структур. Для характеристики геостационарных и геоциркуляционных структур, как легко заметить, используются не столько собственные свойства конкретной единицы (пиксела), сколько свойства геосистем более высокого ранга, создающих рамочные условия функционирования. Вклад биоциркуляционных структур характеризовался собственными свойствами территориальных единиц. В четвертом варианте в расчет включались показатели всех трех типов структур одновременно, т.е. наиболее полная характеристика рельефа.
Таблица 1 показывает, что наибольший вклад в дифференциацию трех типов растительности вносят биоциркуляционные структуры. К геостационарным и геоциркуляционным структурам наиболее чувствительны леса: особенно заметно их вытеснение степями на девонских основных и ультраосновных эффузивах, а также на меловых глауконитово-кварцевых песчаниках и галечниках. Биоциркуляционные структуры более жестко контролируют распространение степей, чем лесов. Кустарники наименее своеобразны по геоморфологической нише во всех четырех вариантах расчета.
Таблица 1.
Корректность распознавания типов растительности морфометрическими характеристиками рельефа, отражающими принадлежность к геостационарным, геоциркуляционным и биоциркуляционным структурам. Дискриминантная модель. Метод прямого пошагового выбора при F = 1
| Характеристики геостационарных структур (TPI, TRI, Channel base , Vertical distance, Valley depth) | ||||
|---|---|---|---|---|
| Wilks’ Lambda: 0.9054394 F (26,454624) = 890.39 p < 0.000 | ||||
| Наблюдаемые сообщества | Доля корректной классификации | Предсказанные сообщества | ||
| Степи | Кустарники | Леса | ||
| Степи | 44.56351 | 24 702 | 11 211 | 19 518 |
| Кустарники | 22.04818 | 21 802 | 11 936 | 20 398 |
| Леса | 61.29925 | 25 802 | 19 772 | 72 186 |
| ВСЕГО | 47.87113 | 72 306 | 42 919 | 112 102 |
| Характеристики геоциркуляционных структур | ||||
| Wilks’ Lambda: 0.94577 approx. F (18,454632) = 714.09 p < 0.000 | ||||
| Наблюдаемые сообщества | Доля корректной классификации | Предсказанные сообщества | ||
| Степи | Кустарники | Леса | ||
| Степи | 38.90422 | 21 565 | 19 036 | 14 830 |
| Кустарники | 35.70822 | 16 238 | 19 331 | 18 567 |
| Леса | 49.45737 | 27 072 | 32 447 | 58 241 |
| ВСЕГО | 43.60987 | 64 875 | 70 814 | 91 638 |
| Характеристики биоциркуляционных структур | ||||
| Wilks’ Lambda: 0.82621 approx. F (18,454632)=2529.8 p < 0.000 | ||||
| Наблюдаемые сообщества | Доля корректной классификации | Степи | Кустарники | Леса |
| Степи | 64.00931 | 35 481 | 8817 | 11 133 |
| Кустарники | 29.84336 | 18 989 | 16 156 | 18 991 |
| Леса | 56.29670 | 31 991 | 19 474 | 66 295 |
| ВСЕГО | 51.87769 | 86 461 | 44 447 | 96 419 |
| Три типа структур вместе | ||||
| Wilks’ Lambda: 0.73853 approx. F (64,454586) = 1162.3 p < 0.000 | ||||
| Наблюдаемые сообщества | Доля корректной классификации | Предсказанные сообщества | ||
| Степи | Кустарники | Леса | ||
| Степи | 65.79170 | 36 469 | 9845 | 9117 |
| Кустарники | 34.62206 | 18 708 | 18 743 | 16 685 |
| Леса | 63.11990 | 25 842 | 17 588 | 743 30 |
| ВСЕГО | 56.98487 | 81 019 | 46 176 | 100 132 |
Наивысшее качество распознавания трех типов растительности достигается одновременным учетом всех трех типов структур. Дискриминантный анализ для гипотезы о наличии трех устойчивых состояний растительности показал, что 46% площади степей приходится на местоположения в рельефе, которые не являются для них однозначно более благоприятными, чем для лесов и кустарников и, значит, допускают пластичность растительного покрова. Доля лесов в условиях рельефа, допускающих пластичность, составила 36%, а кустарников – 54%. Таким образом, кустарники наименее своеобразны по занимаемой геоморфологической нише и примерно с равной вероятностью встречаются в типично степных и типично лесных местообитаниях.
Для Южного Урала отмечалось, что кустарниковые степи с 1990-х гг. расширяют свой ареал и встречаемость, чему способствовало снижение пастбищных нагрузок [25]. При допущении, что кустарники не имеют своей специфической геоморфологической ниши, расчет вероятностей показал, что основная часть кустарниковых сообществ сформировалась в местоположениях, более типичных для лесов (69% пикселов с вероятностью более 0.5), чем для степей (31% пикселов с вероятностью более 0.5).
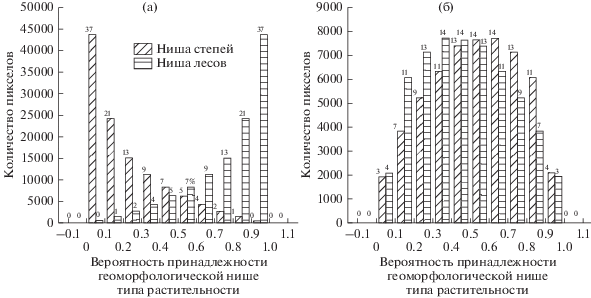
При допущении только двух устойчивых состояний – лесного и степного – оказалось, что 87% пикселов с лесами имеет вероятность принадлежности к лесной геоморфологической нише более 0.5 (рис. 3а). Доля лесов в местоположениях, которые практически исключают формирование степей (вероятность лесов более 0.9) – 37%. Леса могут занимать не только северные, но и южные солнечные склоны, но только при условии достаточной площади водосбора и аккумуляции влаги, т.е. существенно ниже водораздела. Степи, в отличие от лесов, часто встречаются в местоположениях, которые могут быть с примерно равной вероятностью свойственными и степям, и лесам: 26% площади степных урочищ имеют вероятность соответствия степной геоморфологической нише 0.4–0.6 и 55% – вероятность более 0.5 (рис. 3б). Доля степных пикселов в местоположениях, которые практически исключают формирование лесов (вероятность степей более 0.9), – всего лишь 4%.
Геохимические признаки степных и лесных почв
При выявлении геохимической специфики почв под лесными, кустарниковыми и степными урочищами необходимо было сопоставить вклады факторов биогенной аккумуляции и геолого-геоморфологической дифференциации.
В табл. 2 представлены характерные различия химических свойств почв под степями, кустарниками и лесами без учета рельефа. Затем проверялась гипотеза, что рельеф с уклонами менее 8° с устойчивыми радиальными потоками более благоприятен для формирования специфических химических свойств почв под каждым из трех типов растительности, чем рельеф с доминированием латеральных потоков (уклоны более 8°). Выбор крутизны 8° в качестве границы был обусловлен требованием одинакового количества наблюдений в обеих группах.
Таблица 2.
Сравнительная характеристика химических свойств горизонта AU почв под степями (преимущественно черноземы глинисто-иллювиальные), лесами (преимущественно серые лесные) и кустарниками по результатам дисперсионного анализа (p < 0.05)
| Химические свойства | Тип растительности | ||
|---|---|---|---|
| Степи | Кустарники | Леса | |
| Достоверные максимумы | Ga, As | Nобщ., Mn | pH, Pподв., Ba, Ag, La, Zr |
| Достоверные минимумы | Cорг., Kобм., Sr, Ba, Ag | Cr | V, Co, Cu, Ge, Sc |
| Коэффициент радиальной дифференциации (AU/BC) >1 | Cорг., Mn, Zn, P, Y, Yb | – | Kобм., Рподв., Caобм., Mgобм., Кподв., Sr, Ba, Mn |
| Коэффициент радиальной дифференциации (AU/BC) <1 | рН, Mgобм., Naобм. | – | Cr, Mo |
Оказалось, что ландшафтный масштаб (с полным охватом массива данных 196 точек любой крутизны) позволяет наиболее достоверно выявить различия по рН (максимум под лесами), обменному K и валовому Mn (максимум под кустарниками). При этом отдельно на крутых и пологих склонах различия незначимы. Иначе говоря, геоморфологический фактор для этих элементов более значим, чем биологический.
Наоборот, для органического С и общего N различия почв под типами растительности достоверны как в ландшафтном масштабе, так и отдельно по массивам пологих и крутосклонных поверхностей. Следовательно, при любых условиях рельефа растительность способна сформировать специфические для себя химические свойства. Особенно велик вклад растительного покрова на пологих склонах – для общего N и подвижного K (с максимумом под кустарниками), валового Ga (с максимумом под степями); на крутых склонах – для подвижного P, валовых Li, Ba, Ag (с максимумом под лесами), Co и Sc (с максимумом под степями).
Несущественны различия покрова при любом массиве данных для Ca, Mg, Na, Ti, Ni, Zn, Pb, Bi, Be, Mo, W, Y, Yb, Nb, B.
Наиболее устойчиво повторяющиеся ассоциации элементов, показанные в табл. 2, характерны для степей и лесов на плато и на гребнях. Геологические различия оказались менее существенными.
Модель равновесных отношений почв и фитоценозов
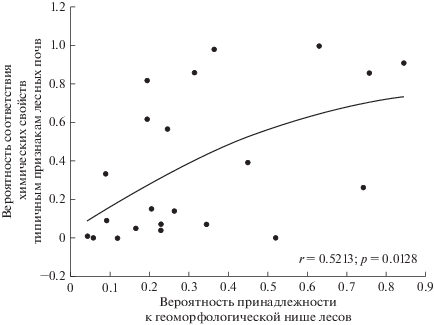
При анализе взаимосоответствий типов растительности и свойств почв сделаны следующие допущения. Если тип растительности занимает обычные для себя местоположения в рельефе и под ним формируются почвы специфического химического состава, то такие почвенно-фитоценотические отношения рассматриваются как равновесные. Если тип растительности занимает обычные для себя местоположения, но почвы несут признаки, характерные для другого типа растительности, то это может свидетельствовать об относительно недавнем восстановлении на месте, временно захваченном другим типом растительности. Если тип растительности занимает чуждые для себя местоположения и имеет почвы, типичные для другого типа растительности, то это свидетельствует о его недавней экспансии. Для краткости будем называть “степными” почвы, в которых содержания химических элементов в горизонте AU имеют более высокую вероятность соответствия типичным значениям для степей, а “лесными” – почвы с более высокой вероятностью соответствия типичным содержаниям элементов для лесов, отмеченным в табл. 2. Цифры, приводимые ниже, соответствуют вероятностям соответствия комбинации химических свойств наиболее часто встречающимся для данного типа растительности (а не встречаемости почв относительно площади исследования).
Рисунок 4а показывает, что ситуация “степь в лесных местоположениях на лесных почвах” (при вероятности “лесных” химических свойств более 0.4) встречается гораздо чаще, чем обратная ситуация (рис. 4б) “лес в степных местоположениях на степных почвах”. Обычно даже если лес по каким-то причинам вторгается в степные местоположения, то вероятность сходства со степными химическими свойствами не превышает 0.4.
Рис. 4.
Встречаемость сочетаний вероятностей принадлежности типа растительности к чуждой геоморфологической нише и вероятности соответствия химических свойств почв типичным для другого типа растительности: (а) степь в лесных местоположениях на лесных почвах, (б) лес в степных местоположениях на степных почвах, (в) кустарники в степных местоположениях на степных почвах.
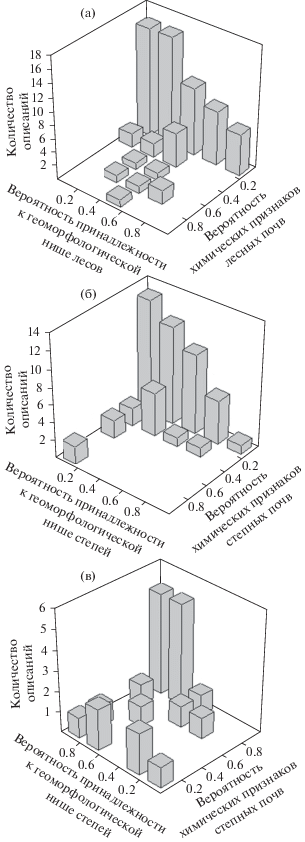
На плато под степями неоднократно встречены почвы, близкие по химическому составу к лесным. В то же время именно на плато наиболее часто зафиксирована и обратная комбинация, когда под дубравами встречаются степные почвы с вероятностями от 0.4 до 1. Другая характерная ситуация несоответствия почв и фитоценозов приурочена к лощинам, где также под лесами могут встречаться степные почвы с вероятностью 0.2–0.4. В почвах склонов вероятность сходства со степными химическими свойствами под лесными фитоценозами менее 0.1. На узких гребнях вероятность степных химических свойств под лесами также всегда меньше 0.1. Однако под кустарниками вероятность степных химических свойств на гребнях в большинстве случаев была в диапазоне 0.8–1. Под кустарниками случаи минимального сходства со степными почвами и максимального – с лесными наблюдались в основном на склонах северных, восточных и южных экспозиций.
Большинство описанных в поле кустарниковых сообществ были приручены к типичным степным местоположениям и при этом имели почвы, сходные с типично степными (рис. 4в). Для безлесных урочищ установлено, что в степных местоположениях, по сравнению с лесными, повышено обилие Spiraea hypericifolia, Cerasus fruticosa, Chamaecytisus ruthenicus, Rosa glabrifolia. Напротив, Caragana frutex и Amygdalus nana могут не уменьшать обилия с переходом в лесную геоморфологическую нишу.
В целом под кустарниками химические свойства чаще ближе к степным почвам (64% исследованных урочищ), чем к лесным (36%). Этот факт свидетельствует в пользу гипотезы об изначально степной природе кустарниковых сообществ: кустарники могут захватывать урочища, которые ранее развивались по степному типу, т.е. с преобладанием травяной растительности. Цветовые признаки могут быть с равной вероятностью близки к степным и лесным почвам. Если кустарники занимают местоположения, хотя бы с небольшой вероятностью соответствующие “лесным”, то могут появляться и химические свойства почв, с вероятностью более 0.4 близкие к лесным (рис. 5). В то же время вероятность того, что морфологические (цветовые) свойства почв соответствуют типичным для лесов не выше, чем для степей. Если же кустарники занимают степные местоположения (с вероятностью 0.6 и более), то очевидная закономерность отсутствует, и возможны разные сочетания степных и лесных химических и морфологических свойств.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные результаты показали, что решающий, но все же не исключительный, вклад в дифференциацию растительного покрова вносит обеспеченность солнечной энергией, зависящая от крутизны и экспозиции поверхности (положение в биоциркуляционной структуре). Это в целом подтверждает закономерность, известную на примере других регионов, однако без учета вклада геостационарных и геоциркуляционных структур (особенно для лесов) понимание размещения типов растительности в лесостепном ландшафте будет неполным. Кустарники не подчиняются со сколько-нибудь значимой достоверностью ни одной из этих трех типов структур, что свидетельствует в пользу их производного характера.
Наши данные свидетельствуют о многообразии и противоречивости результатов отношений между широколиственно-лесными и степными сообществами в пространстве. Основная часть степей приурочена к местоположениям, в которых с равной вероятностью могли бы существовать леса. Степи вообще чаще занимают типичные местоположения лесов, чем наоборот. На субгоризонтальных поверхностях и южных склонах возможны и реально встречаются оба устойчивых состояния. На приводораздельном степном плато лесные свойства в почвах особенно распространены в суженных секторах, где ширина степных участков среди дубрав составляет не более 200 м. В период весеннего снеготаяния такие небольшие степные участки в окружении леса медленнее освобождаются от снежного покрова по сравнению с открытыми склонами и получают дополнительное увлажнение за счет талых вод из окружающего леса, а в летний период испытывают продолжительное затенение [7]. Этот механизм, видимо, создает наиболее благоприятные условия для триггерных эффектов, когда в зависимости от внешних воздействий два типа растительности относительно легко сменяют друг друга.
Основная часть лесов занимает местоположения, в которых у них нет конкурентов, прежде всего – на северных склонах. Группа местоположений, которые являются исключительно степными, занимает небольшие площади и приурочена исключительно к наиболее широкой части плато и глубокорасчлененным местностям, сложенным вулканическими основными породами. Учащение пожаров с ростом летних температур способствует экспансии степей в характерные лесные местоположения при сохранении химических свойств лесных почв. При этом степной фитоценоз препятствует развитию древесной поросли (за исключением водосборных понижений, которые легко захватываются порослью осины), а накопление степного войлока благоприятствует возникновению новых пожаров. Тем самым положительная обратная связь поддерживает устойчивость степных сообществ, захвативших лесные местоположения. В островных дубовых рощах на южных и восточных склонах и на плато возобновление дуба происходит успешно, но прекращается на расстоянии нескольких метров от границы рощи.
Признаки экспансии лесов на степи наиболее часто наблюдались на плато и в лощинах (даже южных склонов), что, видимо, связано с наибольшими возможностями аккумуляции дополнительной влаги, поступающей с растущими зимними осадками. Осиновую поросль можно рассматривать как индикатор экспансии лесной растительности на степь и сохранения в почвах химических свойств, свойственных степям. Прямолинейность границы степных и порослевых сообществ на плато в типично лесных местоположениях свидетельствует о том, что леса были искусственно сведены под распашку, но восстановившаяся после забрасывания степь препятствует восстановлению леса.
Кустарники достаточно “пластичны” в выборе местоположений. Однако основная часть кустарниковых сообществ занимает те же позиции, что и современные степи, а почвы под ними мало отличаются от степных. Не исключается экспансия кустарников из-за роста влагообеспеченности за счет зимних осадков благодаря задержанию снега при сдувании его ветром с выпуклых позиций. В каменистых выпуклых позициях рельефа почвообразование под кустарниками, видимо, принципиально не отличается от почвообразования под степями. Однако если есть хотя бы небольшая вероятность произрастания лесов в данных условиях рельефа, то под кустарниками могут встречаться химические признаки лесных почв. Этому факту также можно дать два объяснения. Во-первых, наличие признаков лесных почв может свидетельствовать лишь о временном отступлении лесов. Широкая распространенность кустарниковых зарослей среди обгоревших мертвых стволов дубов свидетельствует о решающем вкладе пожаров, которые становятся более вероятными по мере роста летнего потепления. Во-вторых, допустим вариант, что повышенное накопление снега в кустарниковых зарослях активизирует проявление признаков лесного почвообразования.
ВЫВОДЫ
1. Основной вклад в распределение лесных, степных и кустарниковых сообществ вносит перераспределение солнечной радиации рельефом (биоциркуляционные ландшафтные структуры) при подчиненном вкладе геоциркуляционных и геостационарных структур.
2. Степные сообщества занимают геоморфологические позиции, в той же степени пригодные и для лесов, что свидетельствует о наличии двух устойчивых состояний лесостепного ландшафта. Внеконкурентная ниша степей занимает лишь небольшие пространства во внутренней части плато. Лесные сообщества теневых склонов и глубоковрезанных лощин занимают позиции, не пригодные для степей.
3. Рост летних температур и частоты пожаров способствует экспансии степей в лесные местоположения при сохранении инертных химических свойств лесных почв. Экспансия лесов в степные местоположения распространена реже, преимущественно по северным склонам; почвы при этом испытывают более быстрые преобразования по лесному типу.
4. Кустарниковые сообщества не имеют специфической геоморфологической ниши, почвы под ними чаще имеют степной характер, чем лесной, что свидетельствует о производном характере этих сообществ.
Список литературы
Арманд А.Д., Ведюшкин М.А. Триггерные геосистемы. М., 1989. 51 с.
Арманд А.Д., Таргульян В.О. Принцип дополнительности и характерное время в географии // Системный подход в географии. М., 1972. С. 18–23.
Горчаковский П.Л. Растения европейских широколиственных лесов на восточном пределе их ареала. Тр. Ин-та экологии растений и животных УрО АН СССР. Свердловск, 1968. 207 с.
Гродзинский М.Д. Устойчивость геосистем: теоретический подход к анализу и методы количественной оценки // Изв. АН СССР. Сер. геогр. 1987. № 6. С. 5–15.
Гродзинський М.Д. Ландшафтна екологія. К.: Знання, 2015. 550 с.
Дубравная лесостепь на хребте Шайтан-Тау и вопросы ее охраны / под ред. Е.В. Кучерова. Уфа, 1994. 186 с.
Золотарева Н.В., Золотарев М.П. Феномен облесения степных участков на Среднем Урале и его вероятные причины // Экология. 2016. № 6. С. 414–425.
Золотокрылин А.Н., Титкова Т.Б., Черенкова Е.А. Увлажнение засушливых земель Европейской территории России: настоящее и будущее // Аридные экосистемы. 2014. Т. 20. № 2 (4). С. 49–54.
Изменение климата в Уральском регионе и предполагаемые изменения его основных составляющих. Последствия изменения климата. Официальный сайт Курганского центра по гидрометеорологии и мониторингу окружающей среды. http://www.kurganpogoda.ru/index.php?option=com_content&view=article&id=13:climat-change&catid=9&Itemid=105) (дата обращения 25.03.2019).
Кильметова Ф.Ш. Циркуляционные условия выпадения осадков и особенности их распределения на Южном Урале / Методы изучения, расчетов и прогнозов водных и климатических ресурсов. Пермь, 1978. С. 86–95.
Кораблева Е.Г. Локальное и региональное потепление на Южном Урале // Вестн. ЧелГУ. 2007. № 6. С. 56–65.
Крашенинников И.М. Физико-географические районы Южного Урала. М.–Л.: Акад. наук СССР. Ч. 1: Предгорья Восточного склона и прилегающие части пенепленов. 1939. 109 с.
Кулакова С.А., Роготнева А.М. Инструментальная оценка состояния дубовых насаждений на территории природного заповедника “Шайтан-Тау” // Проблемы степеведения. 2019. № 15. С. 160–164.
Ленская О.Ю., Ботова М.Г. Особенности текущих климатических изменений в регионе Южного Урала // Вестн. ЧелГУ. 2011. № 5 (220). Экология. Природопользование. Вып. 5. С. 44–49.
Малышева Г.С. Взаимоотношения леса и степи на Приволжской возвышенности (в порядке дискуссии) // Phytodiversity of Eastern Europe. 2018. Т. XII. № 2. С. 170–188.
Павлейчик В.М. К вопросу об активизации степных пожаров (на примере Заволжско-Уральского региона) // Вестн. ВГУ. Сер. География. Геоэкология. 2016. № 3. С. 15–25.
Пищальникова Е.В. Циркуляционные условия формирования обильных снегопадов в Пермском крае // Геогр. вестник. 2016. № 1 (36). С. 70–77.
Проект организации государственного горно-лесостепного заповедника “Шайтан-Тау” (пояснительная записка) / под ред. А.А. Чибилева. Оренбург: Отд. ИЭРИЖ УрО АН СССР, 1991. 217 с.
Пузаченко Ю.Г. Методологические основы географического прогноза и охраны среды. М.: Изд-во УРАО, 1998. 212 с.
Соколов И.А., Таргульян В.О. Взаимодействие почвы и среды: почва–память и почва–момент // Изучение и освоение природной среды. М.: Наука, 1976. С. 150–164.
Титкова Т.Б., Виноградова В.В. Сроки залегания снежного покрова на территории России в начале ХХI века по спутниковым данным // Лёд и Снег. 2017. Т. 57. № 1. С. 25–33.
Чибилев А.А. Заповедник “Шайтан-Тау” – эталон дубравной лесостепи на Южном Урале. Оренбург: Печатный дом “Димур”, 2015. 144 с.
Шкляев В.А., Шкляева Л.С. Оценка изменений температуры воздуха и осадков Среднего и Южного Урала в ХХ веке // Вестн. ЧелГУ. 2011. № 5 (220). Экология. Природопользование. Вып. 5. С. 61–69.
Шмакин А.Б. Климатические характеристики снежного покрова Северной Евразии и их изменения в последние десятилетия // Лёд и Cнег. 2010. № 1. С. 43–57.
Ямалов С.М., Лебедева М.В., Муллагулов Р.Т., Аминев А.Ф. Экологические факторы дифференциации сообществ степных кустарников Зауралья республики Башкортостан // Вестн. Удм. ун-та. Биология. Науки о Земле. 2015. Т. 25. Вып. 4. С. 81–87.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия географическая