

Стратиграфия. Геологическая корреляция, 2023, T. 31, № 6, стр. 140-160
Возраст и условия формирования стратотипа верхнепалеогеновой–нижненеогеновой куршской свиты (Калининградская область) по палинологическим данным
О. Б. Кузьмина 1, *, А. И. Яковлева 2, Э. В. Мычко 3, 4
1 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
2 Геологический институт РАН
Москва, Россия
3 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
4 Балтийский федеральный университет им. И. Канта
Калининград, Россия
* E-mail: KuzminaOB@ipgg.sbras.ru
Поступила в редакцию 28.12.2022
После доработки 24.01.2023
Принята к публикации 18.02.2023
- EDN: TLBTZY
- DOI: 10.31857/S0869592X23060078
Аннотация
Представлены результаты палинологического изучения стратотипа куршской свиты (карьер Приморский, Калининградская область). В низах куршской свиты (пачка “шоколадных” глин) выявлен комплекс цист динофлагеллат с Areosphaeridium diktyoplokum, Glaphyrocysta semitecta и Cordosphaeridium funiculatum терминально-эоценового возраста. Свиту характеризуют четыре спорово-пыльцевых комплекса: (1) комплекс Pinuspollenites–Inaperturopollenites–Sciadopityspollenites терминального эоцена в пачке “шоколадных” глин и низах пачки коричневых песков; (2) комплекс Sequoiapollenites–Betulaepollenites betuloides раннеолигоценового возраста в нижней части пачки коричневых песков; (3) комплекс Boehlensipollis hohli–Carpinipites carpinoides раннеолигоценового возраста в средней части пачки коричневых песков; (4) комплекс Alnipollenites–Corylopollis позднеолигоценового–раннемиоценового возраста в верхней части пачки коричневых песков куршской свиты. Общий возраст куршской свиты – терминальный эоцен–ранний миоцен. В низах перекрывающей куршскую свиту замландской свиты выявлен комплекс Pinuspollenites–Tricolporopollenites pseudocingulum–T. euphorii предположительно среднемиоценового возраста. На основе полученных палинологических данных реконструированы условия осадконакопления в самом конце эоцена–олигоцене–раннем миоцене на территории Южной Прибалтики. Регрессия морского бассейна началась в конце приабона, климат оставался все еще достаточно теплым и влажным, близким к субтропическому. По берегам пролива произрастали мезофитные смешанные хвойно-широколиственные леса, низинные участки суши были заняты болотной растительностью. Похолодание на границе эоцена/олигоцена выразилось в появлении в растительных сообществах тсуги, увеличении доли сережкоцветных (ольха, береза, граб). Предположительно, в позднем олигоцене–раннем миоцене в мезофитных лесах резко увеличилась доля мелколиственных древесных, особенно ольхи и лещины, сократилось количество сосновых. Более влажные и теплые климатические условия предполагаются для раннего миоцена: для этого времени характерно увеличение числа ореховых, кипарисовых, цирилловых. В среднем миоцене климат оставался еще достаточно теплым, но более сухим, из растительных сообществ исчезли такие влаголюбивые породы, как ногоплодник, ель, глиптостробус, болотный кипарис.
ВВЕДЕНИЕ
Балтийская синеклиза представляла собой в палеогене крупный бассейн осадконакопления; в ее различных частях формировались морские, прибрежные и континентальные отложения. Южная Прибалтика, включающая Калининградскую область и юго-запад Литвы, на протяжении большей части палеогена являлась северо-восточной окраиной Датско-Польского морского эпиконтинентального бассейна. В позднем палеогене произошли резкие изменения в палеогеографической обстановке Южной Прибалтики: сначала произошло существенное сокращение площади морских акваторий, а затем морской бассейн вообще прекратил свое существование, и в регионе установился континентальный режим осадконакопления (Геологические…, 1996; Харин, Лукашина, 2002).
Несмотря на то, что детальное изучение палеогена Калининградской области, прежде всего Самбийского полуострова, имеет ключевое значение для восстановления палеогеновых палеогеографических обстановок Польско-Литовского морского палеопролива (как части Датско-Польского моря), до настоящего времени существовал ряд сложностей в решении этой задачи. Прежде всего возникающие проблемы были и остаются связанными с определением точного стратиграфического возраста верхнепалеогеновых свит прибрежно-морского, лагунно-континентального и континентального генезиса. Следует отметить, что их последовательность в Калининградской области в целом отличается большой фациальной изменчивостью и слабой палеонтологической насыщенностью, в связи с чем возраст свит в этом регионе по-прежнему достаточно спорен и требует дополнительных исследований. В последнее время, благодаря детальному изучению цист динофлагеллат и континентальных палиноморф, удалось установить приабонский возраст стратотипов прусской и пальвеской свит в промышленном карьере Приморский Калининградского Янтарного комбината, а также датировать низы вышележащей куршской свиты переходным интервалом от эоцена к олигоцену (Iakovleva et al., 2021; Кузьмина, Яковлева, 2023). Тем не менее возраст всего интервала куршской свиты по-прежнему требует уточнения. Актуальность решения этого вопроса связана с тем, что именно в момент накопления куршской свиты произошел переход от морского к континентальному осадконакоплению, что привело к окончательному прекращению морского сообщения между палеобассейнами Пери-Тетиса и Северо-Западной Европы.
История изучения отложений, относимых в настоящее время к куршской свите, насчитывает более 200 лет. Впервые эти толщи, характеризующиеся богатой ископаемой наземной флорой и запасами бурого угля, были упомянуты в работе немецкого натуралиста А. Швейггера (Schweigger, 1819), заметившего, что остатки древних растений встречаются при разработках янтарей на Самбийском полуострове. В дальнейшем находки ископаемых шишек хвойных растений были отмечены в работе геолога К. Томаса (Thomas, 1847), а затем изучены палеоботаниками Г. Гёппертом и Г. Берендтом (Göppert, Berendt, 1845). Позднее Г. Цаддах попытался детально изучить буроугольные отложения, параллельно собрав обширную коллекцию флоры и описав все доступные обнажения Самбии, а также он разделил третичные отложения полуострова на древнюю глауконитовую и более молодую буроугольную формации (Zaddach, 1860, 1868).
В 1966 г. советский геолог В.И. Балтакис (1966) опубликовал подробное литологическое описание, основываясь на преобладающем генетическом типе пород, литологических особенностях и стратиграфическом положении отдельных слоев в разрезе. Он предложил разделять буроугольную формацию в Калининградской области на три основных комплекса (снизу вверх): лагунно-дельтовые отложения (буровато-серые углистые пески), речные образования (косослоистые разнозернистые пески со стволами деревьев, песчано-глинистые алевриты, песчаный бурый уголь) и болотно-озерные отложения (бурые кварцевые пески, переходящие в бурые угли). Что касается возраста буроугольной формации, то нижний комплекс был отнесен В.И. Балтакисом (1966) к олигоцену, средний, по данным изучения остатков флоры (von Heer, 1869) и спорово-пыльцевым данным (Веножинскене, 1960), − к миоцену, а верхний комплекс, с большой долей сомнения, оказался датирован более молодой частью неогена (Балтакис, 1966). Следует отметить, что палеоботаники Л.И. Буданцев и И.Н. Свешникова (1964), детально проанализировав макрофлористические остатки из буроугольной формации в Калининградской области, пришли к выводу о ее олигоценовом возрасте.
Позднее, в процессе изучения буроугольной формации на западе Самбийского полуострова, советский микропалеонтолог В.Ю. Зосимович (1991) заметил, что в нижней части ее разреза (6.5−7 м) четко выделяются две пачки: (1) нижняя, сложенная коричневыми и зеленовато-серыми горизонтально- и тонкослоистыми однородными глинами (так называемые “шоколадные” глины); и (2) верхняя пачка коричневых песков, содержащая кварцевые пески и тонкослоистые глауконитовые глинистые алевриты, с включениями углистых частиц. На основе полученных наблюдений В.Ю. Зосимович (1991) установил в карьере Приморский стратотип куршской свиты предположительно позднеолигоценового (хаттского) возраста.
Задачами настоящей статьи явились детальный палинологический анализ отложений куршской свиты с целью определения стратиграфического возраста ее стратотипа, а также уточнение интервала геологического времени, в течение которого произошел переход от морского к континентальному типу осадконакопления на Самбийском полуострове; фотоиллюстрация наиболее характерных таксонов палиноморф и, наконец, восстановление растительного палеоландшафта этой территории в позднем палеогене–начале неогена.
МАТЕРИАЛ И МЕТОДЫ
Материалом для настоящего исследования послужили 45 палинологических образцов, отобранных в 2019−2021 гг. из отложений стратотипа куршской свиты в карьере Приморский. Карьер Приморский, принадлежащий Калининградскому Янтарному комбинату, расположен на западе Самбийского полуострова, в 40 км к северо-западу от Калининграда и в 1 км восточнее поселка городского типа Янтарный (рис. 1). В основании карьера залегает алкская свита терминальнолютетского–бартонского возраста, которая последовательно перекрывается прусской и пальвеской свитами приабонского возраста. Пальвеская свита, представленная буровато-зелеными и ярко-зелеными глинистыми, слюдистыми глауконит-кварцевыми песками, подстилает рассматриваемую в настоящей работе куршскую свиту. Последняя, в свою очередь, перекрывается породами замландской свиты предположительно неогенового возраста (Зосимович, 1991).
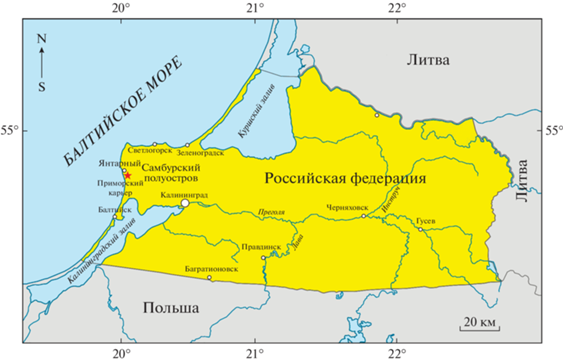
Ниже приводится описание куршской свиты, опробованной в трех соседних точках в северном борту карьера Приморский (рис. 2, 3, 4) на первом, втором и третьем уступах (снизу вверх):


Рис. 4.
Общий вид на чередование тонкозернистых песков замландской свиты. Хорошо заметные черные углистые пески в верхней части обнажения.

–1.7…–1.1 м11 − пачка “шоколадных” глин: глины коричневые, жирные, слюдистые, алевритовые, часто песчанистые; отличаются ленточной слоистостью за счет тонких прослоев серого алеврита.
–1.1...4.7 м – мощная пачка бежевых и светло-серых разнозернистых (в нижней части в основном крупнозернистых) кварцевых песков, местами углистых, с маломощными прослойками и линзочками глин и детрита. В пачке заметно чередование мелко- и среднезернистых слоев.
4.7–6.3 м – глины темно-серые, пластинчатые, тонкие, слюдистые, с ленточной слоистостью, с большим количеством макроостатков растений, обугленных стволов и ветвей деревьев.
6.3–8 м – светло-серые мелкозернистые кварцевые пески, местами алевритистые.
8–12.7 м – пачка так называемых “полосатых” пестрых разнозернистых косо- и перекрестно-косослоистых песков, содержащих большое количество органики в виде мелких обугленных древесных остатков. Внутри пачки в разрезе прослеживается последовательное чередование бурого алеврита, темно-коричневого сильноуглистого песка и светлого мелкозернистого песка с органикой.
12.7–~14 м – слой так называемых “средних глин”, представленный темно-серыми пластинчатыми тонкими слюдистыми глинами с ленточной слоистостью, содержащими немногочисленные макроостатки растений плохой сохранности.
Интервал ~14–16.5 м скрыт в уступе карьера.
Выше по разрезу (16.5–~23 м) куршская свита перекрывается замландской свитой (Зосимович, 1991), контакт которых скрыт в уступе. Замландскую свиту можно разделить на две пачки:
~16.5–21.7 м – крупная пачка ритмично чередующихся слоев светло-бежевых и розовых тонкозернистых слюдистых полуокатанных, хорошо сортированных песков, обогащенных органикой; встречаются линзочки и прослойки углистых песков, а также крупные лигнитизированные стволы и ветви деревьев.
~21.7–~23 м – пески углистые, тонкозернистые, насыщенные сажистыми частицами темного-бурого и черного цвета.
Обработка палинологических образцов была проведена в соответствии со стандартной методикой, принятой в Лаборатории палеонтологии и стратиграфии мезозоя и кайнозоя ИНГГ СО РАН, включающей (1) воздействие на образцы 10%-ной соляной кислотой (HCl) для удаления карбонатов; (2) воздействие горячим раствором пирофосфата натрия (Na4P2O7 · 10H2O) для дисперсии глинистого материала, а затем отмывка каждые 2 ч с целью удаления глинистых частиц; (3) центрифугирование образцов в тяжелой жидкости (K2CdI4) с плотностью 2.25 с целью отделения органической фракции от более тяжелых минеральных частиц; (6) отмывку образца в дистиллированной воде и заливку глицерином. Просеивание мацерата через сита не проводилось.
Палиноморфы изучались в постоянных и временных препаратах с помощью биологических световых микроскопов Микромед 3 (U3) и Микмед-6 при увеличении ×400. Количественный анализ палиноморф включал не менее 200–250 экземпляров на образец. При расчете долевого участия спорово-пыльцевых таксонов за 100% принималась сумма пыльцы голосеменных, покрытосеменных растений и спор мхов и папоротников. Спорово-пыльцевые комплексы (СПК) в куршской свите в разрезе карьера Приморский выделены с учетом наиболее характерных для каждого интервала таксонов. Фотографии палиноморф выполнены с помощью микроскопа ZEISS Axioskop 40 с фотокамерой Canon PowerShot G10. Коллекция препаратов хранится в Институте нефтегазовой геологии и геофизики СО РАН.
РЕЗУЛЬТАТЫ
Во всех изученных палинологических образцах, за исключением обр. 2.20 и 2.22, выявлены количественно представительные ассоциации палиноморф (рис. 5). Следует отметить существенные изменения в соотношениях различных групп морских, водных и континентальных палиноморф. Так, в нижней пачке “шоколадных” глин цисты динофлагеллат составляют до 15% от общего состава палиноморф, в основании пачки коричневых песков они достигают 19%, но выше, в переделах двух следующих метров, их содержание резко сокращается, а затем сводится практически к нулю. Что касается континентальных палиноморф, то в кровле подстилающей пальвеской свиты преобладает пыльца покрытосеменных (Кузьмина, Яковлева, 2023), в то время как для большей части куршской свиты характерно доминирование пыльцы голосеменных, доля которой сокращается лишь в верхней части свиты, уступая пыльце покрытосеменных. В комплексе замландской свиты вновь преобладает пыльца голосеменных. Споры мхов и папоротников составляют лишь небольшую часть выявленных ассоциаций по всему разрезу.
Рис. 5.
Распределение цист динофлагеллат, пыльцы и спор в кровле пальвеской свиты, в куршской и замландской свитах в карьере Приморский. 1 – глины; 2 – пески; 3 – алевриты; 4 – суглинки; 5 – растительные остатки, детрит; 6 – содержание компонента менее 1%; 7 – перерыв в осадконакоплении, несогласие; 8 – отложения скрыты в уступе карьера. Сокращения: Зел. стена – Зеленая стена, pv – пальвеская свита, P. – Platanipollis, C. – Castaneoideaepollis, T. − Tricolpopollenites.
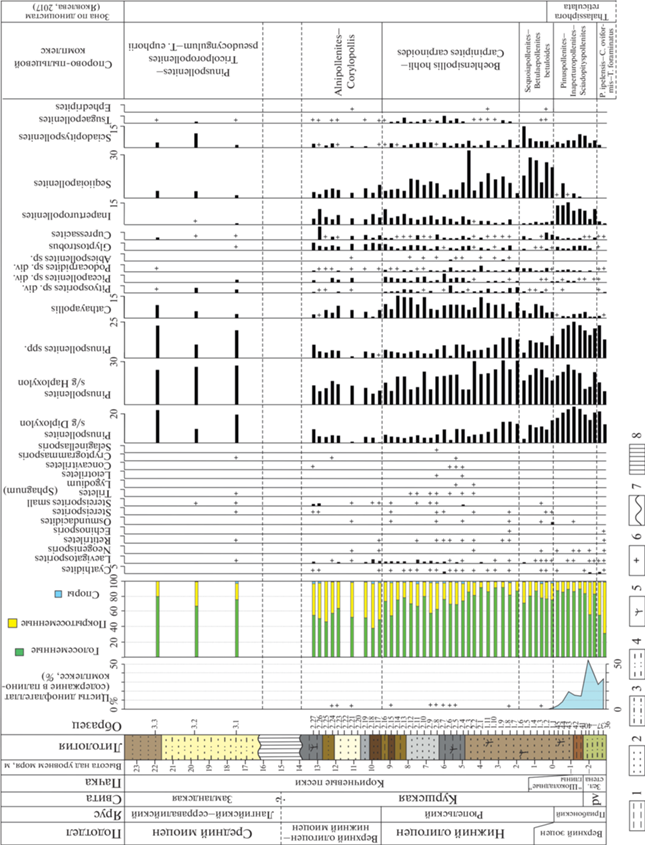
Рис. 5.
Окончание
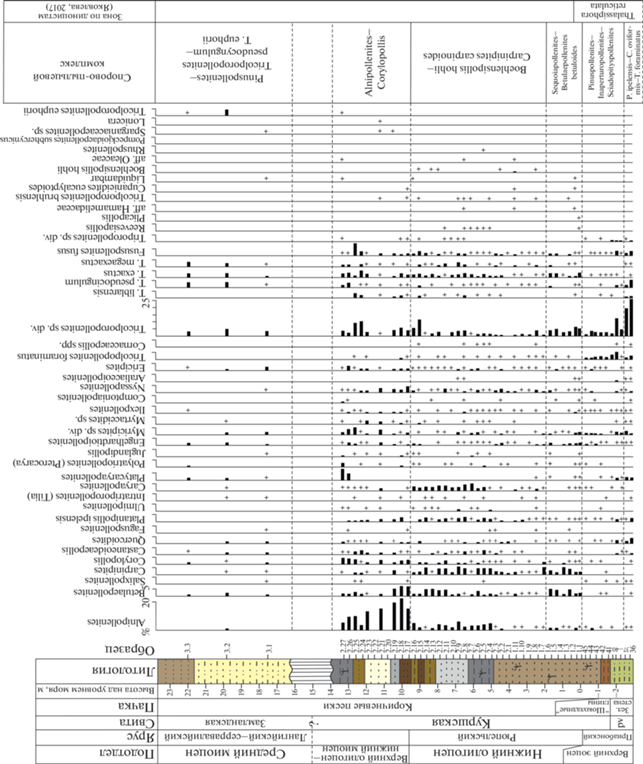
Количественное и качественное распределение спор и пыльцы по разрезу даны на рис. 5. Наиболее характерные виды спор и пыльцы наземных растений представлены на фототаблицах (табл. I, II ). Стратиграфическое распределение цист динофлагеллат представлено на рис. 6.
Рис. 6.
Стратиграфическое распределение цист динофлагеллат в верхах пальвеской свиты и в куршской свите в карьере Приморский. Числа в таблице – количество подсчитанных экземпляров. Экземпляры, обнаруженные после окончания подсчета, обозначены “0”.

Рис. 6.
Окончание
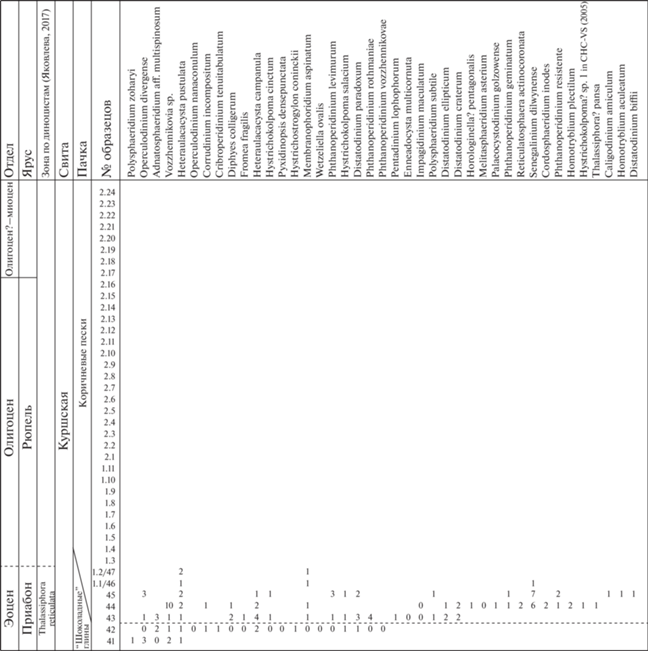
Таблица I . Пыльца голосеменных растений, споры мхов и папоротников из куршской свиты в карьере Приморский (Калининградская обл.). Длина масштабной линейки 20 мкм, для фиг. 44 – 10 мкм. 1 – Piceapollenites sp.; 2, 3 – Abiespollenites sp.; 4, 5 – Pinuspollenites s/g Haploxylon; 6–8 – Pinuspollenites s/g Diploxylon; 9–12 – Cathayapollis spp.; 13 – Podocarpidites libellus (Potonié) Krutzsch; 14 – Podocarpidites sp.; 15, 16 – Sciadopityspollenites sp.; 17–20 – Tsugaepollenites spp.; 21 – Glyptostrobus sp.; 22 – Inaperturopollenites sp.; 23, 24 – Cupressacites sp.; 25, 26 – Sequoiapollenites sp.; 27, 28 – Ephedripites sp.; 29 – Concavitriletes sp.; 30 – Triletes sp.; 31 – Retitriletes sp.; 32, 33 – Triletes spp.; 34 – Leiotriletes sp.; 35 – Selagosporis sp., 36 – Neogenisporis sp.; 37 – Neogenisporis neogenicus Krutzsch; 38 – Laevigatosporites sp.; 39, 40 – Osmundacidites spp.; 41, 42 – Triletes spp.; 43 – Stereisporites sp., 44 – Stereisporites small (Sphagnum).

Таблица II. Пыльца покрытосеменных растений из куршской свиты в карьере Приморский (Калининградская обл.). Длина масштабной линейки 20 мкм, для фиг. 7–14 и 56–74 – 10 мкм. 1, 2 – Faguspollenites sp.; 3, 4 – Quercoidites henrici (Potonié) Potonié, Thomson et Thiergart; 5–7 – Quercoidites spp.; 8 – Tricolporopollenites fallax (Potonié) Krutzsch; 9 – T. liblarensis (Thomson) Grabovska; 10, 11 – Quercoidites microhenrici (Potonié) Potonié, Thomson et Thiergart; 12–14 – Castaneoideaepollis oviformis (Potonié) Grabovska; 15–18 – Platanipollis ipelensis (Pacltová) Grabowska; 19 – Liquidambar sp.; 20, 21 – Comptoniapollenites sp.; 22 – Ulmipollenites sp.; 23 – Intratriporopollenites sp.; 24, 25 – Nyssapollenites sp.; 26–28 – Carpinipites carpinoides (Pflug) Nagy; 29 – Caryapollenites simplex (Potonié) Potonié; 30 – Caryapollenites sp.; 31 – Ilexpollenites propinquus (Potonié) Potonié; 32 – Ilexpollenites sp.; 33–35 – Ilexpollenites margaritatus (Potonié) Raatz; 36–38 – Ericipites spp.; 39 – Ericipites callidus (Potonié) Krutzsch; 40 – Rhuspollenites sp.; 41 – Cornaceaepollis sp.; 42 – Tricolporopollenites marcodurensis Pflug et Thomson; 43 – Tricolporopollenites dolium (Potonié) Thomson et Pflug; 44 – T. photinoides Skawińska; 45 – Tricolporopollenites pseudocingulum (Potonié) Thomson et Pflug; 46 – Tricolporopollenites euphorii (Potonié) Pflug et Thomson; 47 – Tricolporopollenites sp.; 48 – aff. Hamamelidaceae; 49, 50 – Tricolporopollenites microreticulatus Pflug et Thomson; 51 – T. foraminatus Manykin; 52, 53 – Tricolpopollenites foraminatus Manykin; 54 – Tricolporopollenites aff. villensis (Thomson) Thomson et Pflug; 55, 56 – Fususpollenites fusus (Potonié) Kedves; 57–59 – Araliaceoipollenites spp.; 60 – Tricolporopollenites aff. Hypericaceae; 61 – Castaneoideaepollis pusillus (Potonié) Grabovska; 62–65 – aff. Oleaceae; 66 – Engelhardtioipollenites punctatus (Potonié) Potonié; 67 – E. quietus (Potonié) Potonié; 68–71 – Tricolporopollenites exactus (Potonié) Grabovska; 72, 73 – Cupanieidites eucalyptoides Krutzsch; 74–77 – Myrtaceidites spp.; 78, 79 – aff. Hamamelidaceae; 80–82 – Tricolporopollenites megaexactus (Potonié) Thomson et Pflug; 83 – T. brühlensis (Thomson) Grabowska; 84 – Reevesiapollis sp.; 85 – Platycaryapollenites sp.; 86 – Triporopollenites plicoides Zaklinskaya; 87, 88 – Cornaceaepollis sp.; 89, 90 – Corylopollis spp.; 91 – Betulaepollenites betuloides (Pflug) Nagy; 92, 93 – Alnipollenites spp.; 94 – Myricipites bituites (Potonié) Nagy; 95, 96 – Myricipites rurensis (Pflug et Thomson) Nagy; 97, 104 – Juglandipollis spp.; 98 – Pompeckjoidaepollenites subhercynicus (Krutzsch) Krutzsch; 99–103 – Boehlensipollis hohli Krutzsch; 105 – Polyatriopollenites sp.

Цисты динофлагеллат. Выявленный в пачке “шоколадных” глин (обр. 41–42) и самых низах пачки коричневых песков (обр. 43–47) диноцистовый комплекс количественно немногочисленный, но содержит 64 вида. Абсолютным доминантом комплекса является Deflandrea phosphoritica (~36–24%), при этом в основании “шоколадных” глин заметно участие видов Spiniferites ramosus (14–16%) и Areosphaeridium diktyoplokum (8%). По таксономическому составу диноцистовый комплекс из низов куршской свиты очень близок к комплексу подстилающей пальвеской свиты позднеприабонского возраста, хотя и содержит несколько меньшее число таксонов, что явным образом связано с изменениями обстановки осадконакопления в условиях регрессии морского бассейна. Помимо прочего комплекс характеризуется присутствием видов Cerebrocysta bartonensis, Enneadocysta pectiniformis, Cordosphaerdium funiculatum, Glaphyrocysta semitecta, Distatodinium biffii. Согласно (Brinkhuis, Biffii, 1993), а также более поздним публикациям (Brinkhuis, 1994; Gedl, 2004; Egger et al., 2016), сама по себе граница эоцена/олигоцена не характеризуется каким-либо важным диноцистовым событием, однако упомянутые выше виды Areosphaeridium diktyoplokum, C. funiculatum, G. semitecta исчезают в самом начале рюпеля. Согласно (Heilmann-Clausen, Van Simaeys, 2005), последнее появление (highest occurrence) вида Cordosphaeridium funiculatum может быть использовано в качестве вторичного биособытия для определения границы эоцена/олигоцена в бассейне Северного моря. Исходя из присутствия видов A. diktyoplokum, G. semitecta, C. funiculatum, а также учитывая отсутствие раннеолигоценовых видов-индексов Wetzeliella gochtii и Chiropteridium galea, возраст низов куршской свиты соответствует, скорее всего, самому концу приабона вблизи границы эоцена/олигоцена.
Пыльца и споры растений. Подстилающую отложения куршской свиты пальвескую свиту (здесь обр. 36–37) характеризует спорово-пыльцевой комплекс (СПК) Platanipollis ipelensis–Castaneoideaepollis oviformis–Tricolpopollenites foraminatus с высоким содержанием мелкой пыльцы трехбороздно-порового строения. Подробно состав и структура комплекса описаны в работе (Кузьмина, Яковлева, 2023). Позднеэоценовый возраст отложений подтвержден данными изучения цист динофлагеллат (Iakovleva et al., 2021).
Из кровли пальвеской свиты (обр. 38–40), “шоколадных” глин и нижней части коричневых песков (обр. 41–45) куршской свиты в интервале –2.3…0.2 м выявлен СПК Pinuspollenites–Inaperturopollenites–Sciadopityspollenites. В нем стабильно доминирует пыльца голосеменных растений (до 89%), в этой группе преобладает пыльца различных Pinuspollenites (до 65%), причем пыльца обеих секций (Haploxylon и Diploxylon) присутствует примерно в равных количествах. Субдоминант в комплексе – пыльца сем. Cupressaceae (в сумме до 32.6%), при этом особенно весомой становится доля пыльцы Inaperturopollenites sp. (до 15%), в то время как пыльца родов Cupressacites и Glyptostrobus присутствует в небольших количествах (0.5–5%). Кроме того, комплекс отличается относительно высоким содержанием пыльцы Sciadopityspollenites (до 8%). В незначительных количествах (0.5–2.5%) отмечена пыльца Cathayapollis spp., еще реже (0.5–1%) встречается пыльца родов Piceapollenites, Podocarpidites, Pityosporites.
Пыльцы покрытосеменных в комплексе немного (10–17%), таксономическое разнообразие ее значительно снижается по сравнению с СПК пальвеской свиты (Кузьмина, Яковлева, 2023). Исключение составляет лишь спектр обр. 39, где доля такой пыльцы достигает 43%. Чаще всего в комплексе встречается пыльца различных Tricolporopollenites (до 14%), доля ее постепенно снижается вверх по разрезу (до 1–5%). Отмечены виды Tricolporopollenites exactus, T. megaexactus, T. microporites, T. liblarensis, T. cingulum, T. pseudocingulum. В кровле пальвеской свиты еще довольно часто встречается пыльца Tricolpopollenites foraminatus (до 5.5%), в нижней части куршской свиты доля такой пыльцы снижается до 0.5–1%. В незначительных количествах (0.5–2.5%) присутствует пыльца Fususpollenites fusus, Myricipites bituites, M. rurensis, Quercoidites microhenrici, Platanipollis ipelensis. Единично отмечены Alnipollenites sp., Engelhardtioipollenites punctatus, E. quietus, Castaneoideaepollis oviformis, C. pusillus, Caryapollenites simplex, Ilexpollenites sp., Ericipites callidus, Ericipites spp., Platycaryapollenites. Спорадически встречается пыльца Betulaepollenites betuloides, Salixpollenites, Corylopollis sp., Ulmipollenites, Polyatryopollenites sp., Nyssapollenites sp., Myrtaceidites sp., Comptoniapollenites sp., Araliaceoipollenites sp., Intratriporopollenites sp.
Спор папоротников и мхов в комплексе немного (0.5–2%), они принадлежат родам Cyathidites, Neogenisporis, Laevigatosporites, Osmundacidites.
Поскольку отложения в интервале образцов 38−45 представлены морскими и солоноватоводными фациями и содержат комплекс диноцист терминального приабона−перехода от эоцена к олигоцену, выявленный в карьере Приморский СПК Pinuspollenites–Inaperturopollenites−Sciadopityspollenites датируется нами самым концом эоцена−переходом к олигоцену. СПК Pinuspollenites–Inaperturopollenites−Sciadopityspollenites близок по составу и структуре к комплексу с Pinuspollenites, Inaperturopollenites hiatus, Sequoiapollenites, Cupressacites bockwitzensis, установленному в верхней части поморской свиты в Польше (Słodkowska, 2009). Помимо пыльцы наземных растений, палинокомплекс поморской свиты характеризуется присутствием микрофитопланктона Cordosphaeridium cf. funiculatum, Enneadocysta pectiniformis, Charlesdowniea (=Talladinium?) clathrata, Pentadinium laticinctum и Glaphyrocysta pastielsii, подтверждающего позднеприабонский возраст этих отложений (Słodkowska, 2009).
Коричневые пески куршской свиты в инт. ‒0.3…1.5 м (обр. 1.1−1.6) содержат СПК Sequoiapollenites–Betulaepollenites betuloides. Для него характерно резкое увеличение (до 28%) доли пыльцы Sequoiapollenites и также резкое снижение (до 1−2.5%) Inaperturopollenites sp.; в то же время доля пыльцы Cupressacites sp. несколько увеличивается (до 5%). Отметим, что в составе голосеменных по-прежнему преобладает пыльца различных Pinuspollenites, при этом чаще встречается пыльца Pinuspollenites s/g Haploxylon (до 25%), реже − Pinuspollenites s/g Diploxylon (до 15%). Увеличивается доля пыльцы Cathayapollis sp. (до 9%) и Podocarpidites sp. (до 2.5%). Постоянно отмечается пыльца Sciadopityspollenites sp. (2.2−7%), особенно она обильна (до 15%) в верхней части описываемого интервала. Реже, чем в предыдущем СПК, встречается пыльца Glyptostrobus sp. (0.5−1%). Спорадически присутствует пыльца Piceapollenites sp. В составе комплекса единично появляется пыльца Tsugaepollenites sp. и Ephedripites sp.
Таксономический состав пыльцы покрытосеменных СПК Sequoiapollenites–Betulaepollenites betuloides мало отличается от такового из предыдущего комплекса, за исключением более весомой роли пыльцы теплоумеренной флоры, особенно Betulaepollenites betuloides (до 4%) и Carpinipites carpinoides (до 4%); кроме того, единично отмечена пыльца Faguspollenites sp., Juglandipollis sp., Reevesiapollis sp., Plicapollis sp., Liquidambar sp., Tricolporopollenites brühlensis, Cupanieidites eucalyptoides, отсутствующая в нижележащих отложениях.
Спор папоротников и мхов в комплексе мало (0.5−4%), они принадлежат родам Cyathidites, Neogenisporis, Laevigatosporites, Osmundacidites, Stereisporites, Stereisporites small (Sphagnum).
СПК Sequoiapollenites–Betulaepollenites betuloides может быть сопоставлен с раннеолигоценовыми комплексами нижнемосинской свиты (Lower Mosina; Słodkowska, 2009) и угленосных слоев в скв. Łukowa-4 на юго-востоке Польши (Gedl et al., 2016). В нижнемосинской свите выявлен комплекс диноцист с Wetzeliella symmetrica раннеолигоценового возраста; спорово-пыльцевой комплекс этой свиты характеризуется преобладанием пыльцы голосеменных (Pinuspollenites, Sequoiapollenites, Sciadopityspollenites, Inaperturopolenites) и появлением стратиграфически важных раннеолигоценовых видов Boehlensipollis hohli и Cupanieidites eucalyptoides.
Пески и глины куршской свиты в инт. 2.3−9.5 м (обр. 1.7−2.16) характеризуются СПК Boehlensipollis hohli–Carpinipites carpinoides. В комплексе по-прежнему преобладает пыльца голосеменных (58−92.5%). В этой группе доминирует пыльца различных Pinuspollenites (до 50%), особенно много пыльцы Pinuspollenites s/g Haploxylon (до 30%). Постоянно в значительных количествах отмечается пыльца Cathayapollis (5−15%), содержание пыльцы Sequoiapollenites сильно варьирует (от 2.5 до 32%), увеличивается доля пыльцы Inaperturopollenites (до 11.7%). Постоянно, хотя и в незначительных количествах (0.5−4%), в комплексе встречается пыльца родов Glyptostrobus, Cupressacites, Podocarpidites. Пыльца Tsugaepollenites и Piceapollenites становится постоянным компонентом ассоциаций, в коричневых песках такой пыльцы еще немного, но выше по разрезу, в глинах, доля Tsugaepollenites увеличивается до 5.5%, а Piceapollenites − до 6.5%. Несколько сокращается (0.5−6%) количество пыльцы Sciadopityspollenites по сравнению с предыдущим СПК. Спорадически отмечена пыльца Abiespollenites sp., отсутствующая в нижележащих отложениях, а также Ephedripites sp.
Количественное и таксономическое разнообразие пыльцы покрытосеменных в СПК Boehlensipollis hohli–Carpinipites carpinoides определенно выше, чем в комплексах нижней части куршской свиты. Среди покрытосеменных появляется стратиграфически важный таксон Boehlensipollis hohli. Наиболее часто (до 10%) встречаемыми таксонами являются Caryapollenites simplex, Alnipollenites, Betulaepollenites betuloides, Carpinipites carpinoides. Несколько увеличивается (до 3%) доля пыльцы Platanipollis ipelensis. В незначительных количествах (0.5−2.5%), но постоянно встречается пыльца родов Corylopollis, Castaneoideaepollis (С. oviformis, C. pusillus), Quercoidites (Q. henrici, Q. microhenrici, Q. quisqualis), Engelhardtioipollenites (E. punctatus, E. quietus), Myricipites (M. bituites, M. rurensis), Nyssapollenites, Ericipites, Fususpollenites fusus. Содержание пыльцы формального рода Tricolporopollenites непостоянно и варьирует от 5 до 16% (суммарно), разнообразие такой пыльцы невысоко в коричневых песках, но повышается в глинах и песках верхней части описываемого интервала (обр. 2.4−2.15). Чаще всего встречаются Tricolporopollenites exactus, T. megaexactus, T. brühlensis, реже – T. retiformis, T. liblarensis, T. fallax, T. marcodurensis, T. microreticulatus, T. microporites, T. cingulum, T. pseudocingulum, T. leonensis, T. asper, T. dolium, T. theacoides. Спорадически встречается пыльца Juglandipollis sp., Ulmipollenites sp., Salixpollenites, Faguspollenites sp., Intratriporopllenites sp., Polyatryopollenites sp., Triporopollenites plicoides, Comptoniapollenites, Myrtaceidites, Ilexpollenites, Araliaceoipollenites, Cornaceaepollis, Reevesiapollis, Liquidambar, aff. Oleaceae, aff. Hamamelidaceae, Rhuspollenites sp., Cupanieidites eucalyptoides. Следует отметить, что практически выпадает из комплекса пыльца рода Platycaryapollenites, она лишь спорадически отмечена в низах интервала.
Спор в комплексе немного (0.5−5%), таксономический состав их несколько более разнообразный, чем в СПК нижней части куршской свиты. Споры принадлежат родам Laevigatosporites (встречается чаще всего, до 2.5%), Osmundacidites, Stereisporites, Stereisporites small (Sphagnum), Cyathidites, Neogenisporis, Echinosporis, Triletes, Retitriletes, Leiotriletes, Lygodium, Cryptogrammasporis, Concavisporites, Selaginellasporis.
Водные палиноморфы в комплексе редки, спорадически встречаются единичные диноцисты (Deflandrea phosphoritica, Operculodinium sp.), акритархи, зеленые водоросли Ovoidites sp., Pseudopediastrum boryanum.
Boehlensipollis hohli является видом-индексом нижнего олигоцена в Западной Европе (Châteauneuf, 1986), что позволяет нам отнести вмещающие отложения куршской свиты к нижнему олигоцену.
Из песков и глин куршской свиты в инт. 9.5–13.3 м выявлен СПК Alnipollenites–Corylopollis (обр. 2.17–2.27), в котором содержится примерно равное количество пыльцы голосеменных и покрытосеменных растений. В составе голосеменных сокращается доля пыльцы различных Pinuspollenites (в сумме до 30%), хотя в этой группе она все еще доминирует. Субдоминант – пыльца сем. Cupressaceae (суммарно до 25.5%). При этом определенно снижается доля пыльцы Sequoiapollenites (2.5–9%), в то время как содержание пыльцы Inaperturopollenites (5–11.5%), Cupressacites (1.5–8.5%) и Glyptostrobus (2.5–6%) незначительно повышается. Уменьшается доля пыльцы Cathayapollis (1.5–8.5%), Sciadopityspollenites (0.5–3%) и Piceapollenites (0.5–2.5%). Единично присутствует пыльца родов Tsugaepollenites, Podocarpidites.
Таксономическое разнообразие пыльцы покрытосеменных в СПК Alnipollenites–Corylopollis приблизительно такое же, как и в предыдущем комплексе. Отличие состоит в резком увеличении в первом пыльцы сережкоцветных Alnipollenites sp. (до 20%), Corylopollis sp. (до 4.5%) и Betulaepollenites betuloides (до 6%). Также стоит отметить несколько более высокое содержание в комплексе пыльцы Nyssapollenites (0.5–4.5%), Myrtaceidites (0.5–2.5%), Engelhardtioipollenites (0.5–2%). Напротив, заметно снижается доля пыльцы Caryapollenites simplex (0.5–2%).
В верхах описываемого интервала, в бурых песках и пластинчатых глинах (инт. 12.2–13.3 м, обр. 2.24–2.27) наблюдается резкий рост участия в комплексе мелкой трехбороздно-поровой пыльцы (суммарно до 12%), в том числе Fususpollenites fusus (до 8.6%), а также пыльцы формального рода Tricolporopollenites (T. exactus, T. megaexactus, T. liblarensis, T. pseudoexactus), Myricipites spp. (до 7%) и Platycaryapollenites sp. (до 7.7%). При этом доля пыльцы Alnipollenites снижается здесь до 5–10%.
Споры в СПК Alnipollenites–Corylopollis по-прежнему немногочисленны (1–4%), присутствуют Concavisporites, Laevigatosporites, Osmundacidites, Neogenisporis, Stereisporites, Stereisporites small (Sphagnum), Equisetum. Из водных палиноморф обнаружены единичные диноцисты (Deflandrea phosphoritica, Microdinium reticulatum), зеленые водоросли Pseudopediastrum boryanum, Zygnemataceae, а также остатки колоний зеленых водорослей Botryococcus (до 6.5% в палинокомплексе), устьица хвойных (Pinus-type, Picea-type) и споры грибов.
СПК Alnipollenites–Corylopollis из верхней части куршской свиты весьма близок по составу и структуре к комплексу Sm-5 из угленосных отложений в скв. 3 у дер. Смолярка Брестской области, датированному поздним олигоценом–ранним миоценом (Мурашко и др., 1998). Резкие изменения в структуре комплекса, характеризующего верхнюю часть коричневых песков куршской свиты, говорят о наличии стратиграфического перерыва на этом рубеже.
Перекрывающие отложения замландской свиты (инт. 16.2−23.5 м, обр. 3.1–3.3) характеризует СПК Pinuspollenites–Tricolporopollenites pseudocingulum–T. euphorii c преобладанием пыльцы голосеменных (до 80%). В составе голосеменных много пыльцы различных сосен – Pinuspollenites s/g Haploxylon (до 30%), Pinuspollenites s/g Diploxylon (до 15%), Pinuspollenites sp. (до 30%). От 3 до 10% приходится на долю пыльцы Cathayapollis sp., Sciadopityspollenites sp., Sequoiapollenites sp. Единично отмечена пыльца родов Inaperturopollenites, Cupressacites, Glyptostrobus, Tsugaepollenites, Piceapollenites, Podocarpidites.
Таксономическое разнообразие пыльцы покрытосеменных ниже, чем в СПК куршской свиты, вверх по разрезу оно еще больше снижается. Наиболее часто (до 16%) встречается пыльца формального рода Tricolporopollenites – T. pseudocingulum, T. euphorii, T. exactus, T. megaexactus, в незначительных количествах (1–3%) присутствует пыльца родов Betulaepollenites, Corylopollis, Quercoidites, Castaneoideaepollis, Engelhardtioipollenites, Myricipites, Ericipites. Единично обнаружена пыльца Salixpollenites, Carpinipites, Faguspollenites, Intratriporopollenites, Myrtaceidites, Caryapollenites, Juglandipollis, Ilexpollenites, Nyssapollenites, Liquidambar, Sparganiaceaepollenites.
Споры не играют существенной роли в комплексе (0.5–3%), наиболее разнообразно они представлены только в нижней части свиты (обр. 3.1): здесь присутствуют Cryptogrammasporis, Laevigatosporites, Triletes, Retitriletes, Stereisporites, Stereisporites small (Sphagnum). Выше по разрезу споры единичны (обр. 3.2) либо отсутствуют (обр. 3.3).
Водные палиноморфы представлены клетками зеленых водорослей Botryococcus (до 20% в комплексе), единичными Zygnemataceae, внутренними камерами фораминифер (обр. 3.1) и спорами грибов.
СПК Pinuspollenites–Tricolporopollenites pseudocingulum–T. euphorii замландской свиты отличается увеличением доли пыльцы различных сосен, в то время как кипарисовые в нем практически отсутствуют, за исключением пыльцы Sequoiapollenites. При этом таксономический состав покрытосеменных в комплексе становится гораздо беднее, чем в куршской свите. В этой группе заметную роль играют лишь Tricolporopollenites pseudocingulum, T. exactus, T. megaexactus, T. euphorii. По присутствию этих таксонов представляется возможным сопоставить данный комплекс с комплексами из среднемиоценовых отложений Польши (Piwocki, Ziembinska-Tworzydło, 1997).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате палинологического изучения стратотипа куршской свиты и низов стратотипа замландской свиты в карьере Приморский уточнен их стратиграфический возраст.
Так, верхняя часть пальвеской свиты, “шоколадные” глины и самая нижняя часть коричневых песков куршской свиты, охарактеризованные комплексом Pinuspollenites–Inaperturopollenites–Sciadopityspollenites, отнесены нами к терминальному приабону. Возраст этих отложений подтвержден данными изучения цист динофлагеллат (Iakovleva et al., 2021). Резкое увеличение доли пыльцы голосеменных, в том числе различных сосновых, кипарисовых, характерно для пограничных эоцен-олигоценовых (Kosmowska-Ceranowisz, Müller, 1985) и нижнеолигоценовых (Практическая…, 1990; Słodkowska, 2009; Запорожец, Ахметьев, 2017) отложений различных районов Евразии. Выявленный нами в Калининградской области СПК Pinuspollenites–Inaperturopollenites–Sciadopityspollenites весьма близок по составу и структуре к комплексу Sciadopitys verticillatiformis–Retitricolpites foraminatus из нижней части межигорской свиты Украины (Зосимович, Шевченко, 2015). В комплексе на Украине также преобладает пыльца голосеменных (до 91%), а именно Pinaceae, Cupressaceae, Sciadopityaceaе, его возраст по данным изучения диноцист – рюпель (интервал диноцистовой зоны DP 13 Wetzeliella gochtii) (Стотланд, 1984, 1986).
Нижняя часть коричневых песков куршской свиты содержит раннеолигоценовый комплекс Sequoiapollenites–Betulaepollenites betuloides. Данный комплекс сопоставляется с одновозрастными комплексами нижнемосинской свиты (Lower Mosina; Słodkowska, 2009) и угленосных слоев в скв. Łukowa-4 на юго-востоке Польши (Gedl et al., 2016). Увеличение доли сережкоцветных (Betulaepollenites betuloides, Carpinipites carpinoides), резкое снижение таксономического разнообразия теплолюбивых цветковых растений, а также появление пыльцы Tsugaepollenites в этом комплексе указывает на похолодание, которое характеризует начало олигоцена на глобальном уровне. Так, значительное увеличение пыльцы сосновых, кипарисовых, а также березовых (особенно граба), появление пыльцы ели и тсуги характерно для палинокомплекса нижнеолигоценовой (майкопской) пшехской свиты на Северном Кавказе (Запорожец, Ахметьев, 2017). Таким образом, коричневые пески куршской свиты с комплексом Sequoiapollenites–Betulaepollenites betuloides могут быть сопоставлены и с нижнеолигоценовыми отложениями Пери-Тетиса.
Средняя часть коричневых песков куршской свиты, содержащая комплекс Boehlensipollis hohli–Carpinipites carpinoides, формировалась в раннем олигоцене (рюпельский век). Boehlensipollis hohli является стратиграфическим маркером нижнего олигоцена во Франции (Châteauneuf, 1986) и в Центральном и Западном Паратетисе (Hochuli, 1978). Этот вид известен из нижнего олигоцена Германии (Nickel, 1996) и Великобритании (Boulter, Craig, 1979). Комплекс спор и пыльцы, включающий виды Boehlensipollis hohli и Cupanieidites eucalyptoides, характерен также для нижнеолигоценовых нижнемосинской (Lower Mosina) и чемпинской (Czempin) свит, распространенных на территории Польши (Piwocki, 2004). В Турции вид Boehlensipollis hohli известен из нижнего олигоцена, а его последнее появление совпадает с границей рюпеля и хатта (Akkiraz, Akgün, 2005; Akkiraz et al., 2011). Boehlensipollis hohli отмечен в остракодовых слоях нижнего олигоцена в стратотипическом разрезе на р. Белая в Предкавказье (Запорожец, 1999) и в нижнеолигоценовых курганских слоях на юге Западной Сибири (Ахметьев и др., 2001).
К позднему олигоцену–раннему миоцену отнесена верхняя часть коричневых песков куршской свиты с комплексом Alnipollenites–Corylopollis. Ограничить возраст этих отложений исключительно поздним олигоценом нельзя, так как позднеолигоценовые палинокомплексы Белоруссии и Польши имеют несколько иную структуру: в них пыльца сережкоцветных играет весьма скромную роль. Так, позднеолигоценовый комплекс из углесодержащей стародубской свиты Белоруссии характеризуется высоким содержанием пыльцы голосеменных и низким – пыльцы Alnus (Мурашко и др., 1998; Стратиграфические…, 2010). На территории Польши верхнеолигоценовые отложения сохранились фрагментарно (Słodkowska, 2004), поскольку в позднем олигоцене море отступило с севера Польской низменности, а позднее эти образования были разрушены процессами денудации. Те немногие палинологические данные, которые имеются для верхнего олигоцена Польши, указывают на то, что комплексы из этих отложений также содержат очень мало пыльцы арктотретичной флоры (Słodkowska, 2004). Сокращение пыльцы хвойных на фоне увеличения содержания пыльцы арктотретичной флоры отмечено в верхнеолигоценовых–нижнемиоценовых нерасчлененных отложениях Белоруссии. Так, выделенный нами комплекс Alnipollenites–Corylopollis из верхней части куршской свиты весьма близок по составу к комплексу Sm-5 из угленосных отложений в скв. 3 у дер. Смолярка Брестской области (Мурашко и др., 1998). Для последнего характерно резкое сокращение участия пыльцы хвойных, преобладание пыльцы покрытосеменных, особенно много ольхи (Alnus, до 20%). Отмечено, что значительное участие в этом комплексе пыльцы покрытосеменных, определяемой по искусственной систематике, сближает его с позднеолигоценовыми комплексами; в то же время высокая роль арктотретичных элементов позволяет сопоставлять вмещающие отложения и с нижним миоценом (Мурашко и др., 1998). Известно, что при переходе к позднему олигоцену состав европейских флор изменился незначительно, широкое распространение арктотретичные элементы получили только в конце позднего олигоцена–начале раннего миоцена: во флорах этого времени увеличивается влияние таких листопадных древесных, как Alnus, Corylus, Carpinus и др. (Буданцев, Свешникова, 1964; Ахметьев, 1978). Появление большого количества пыльцы Alnipollenites verus отмечено в низах миоцена Центральной и Северной Европы, где установлена одноименная палинозона (Piwoki, Ziembinska-Tworzydło, 1997), отличающаяся обеднением флоры, вызванным похолоданием климата на рубеже олигоцена и миоцена. На данном этапе исследований мы склонны относить верхнюю часть куршской свиты, содержащую СПК Alnipollenites–Corylopollis, к верхнему олигоцену–нижнему миоцену, поскольку резкое увеличение пыльцы Alnipollenites вполне может отражать похолодание на рубеже хаттского–аквитанского веков, а увеличение доли пыльцы теплолюбивых покрытосеменных, а также кипарисовых в верхней части интервала вполне может соответствовать началу климатического оптимума конца раннего–начала среднего миоцена. Более точно определить возраст верхней части куршской свиты станет возможно по мере накопления палинологических данных в регионе.
Таким образом, полученные нами палинологические данные позволили уточнить возраст стратотипа куршской свиты в карьере Приморский – “шоколадные глины” и нижняя часть коричневых песков имеют терминальноприабонский возраст, средняя часть коричневых песков формировалась в раннем олигоцене (рюпель), верхняя часть свиты предположительно соответствует верхнему олигоцену–нижнему миоцену.
Перекрывающая куршскую свиту замландская свита, содержащая комплекс Pinuspollenites–Tricolporopollenites pseudocingulum–T. euphorii, соответствует среднему миоцену. В настоящее время замландскую свиту сопоставляют с нижнемиоценовыми равичской (Rawicz), горзовской (Gorzow), сцинавской (Scinawa), краженской (Krajenska) и среднемиоценовыми адамовской (Adamow), павловицкой (Pavlowice) свитами Польской низменности (Piwocki, Ziembinska-Tworzydło, 1997). В этой связи следует отметить, что выявленный нами комплекс Pinuspollenites–Tricolporopollenites pseudocingulum–T. euphorii совершенно не дает оснований для его сопоставления с раннемиоценовыми комплексами Польши, поэтому корреляции замландской свиты с нижнемиоценовыми свитами Польской низменности сомнительны. В то же время довольно частое присутствие в комплексе замландской свиты пыльцы Tricolporopollenites exactus, T. megaexactus, T. brühlensis, а также Tricolporopollenites pseudocingulum и T. euphorii позволяет сопоставлять эти отложения с зонами VI (T. megaexactus) либо IX (T. pseudocingulum) среднего миоцена Польши (Piwocki, Ziembinska-Tworzydło, 1997). Более точная корреляция этой части разреза карьера Приморский станет возможной при более детальном изучении всего интервала замландской свиты.
Соотношение различных групп морских, водных и континентальных палиноморф в палинологических комплексах куршской свиты и перекрывающей ее нижней части замландской свиты, а также анализ таксономического состава спорово-пыльцевых комплексов позволяют нам сделать определенные выводы об условиях осадконакопления и изменениях растительного ландшафта Южной Прибалтики в результате постепенного изменения климата в позднем палеогене–начале неогена.
Так, сокращение содержания цист динофлагеллат (15–19%) по отношению к континентальным палиноморфам в палинокомплексе кровли пальвеской свиты и низах куршской свиты четко указывает на развитие регрессии морского бассейна в самом конце приабона вблизи границы эоцена/олигоцена. В это время по берегам пролива произрастали мезофитные смешанные хвойно-широколиственные леса с дубами, каштанами, платанами. Участки суши, освободившиеся от моря, были оккупированы болотной растительностью: в растительных сообществах в это время значительно увеличилось количество болотного кипариса, климат же оставался все еще достаточно теплым и влажным, близким к субтропическому.
Значительное похолодание климата на рубеже эоцена и олигоцена привело к тому, что в составе лесных сообществ увеличилось количество представителей теплоумеренной флоры (ольха, береза, граб). По-видимому, значительно сократились площади заболоченных участков на суше, поскольку в сообществах этого времени резко уменьшилась доля участия болотного кипариса (Inaperturopollenites). В прибрежной части бассейна широкое распространение получили секвойи, а в составе хвойных появились тсуги.
Во второй половине раннего олигоцена в составе хвойных увеличились доли катайи, ели, тсуги, сократилось количество зонтичной сосны. Климат постепенно становился более мягким по сравнению с самым началом олигоцена: на это указывает постоянное присутствие в составе широколиственных таких теплолюбивых представителей, как дуб, платан, каштан, орех, нисса, гикори, птерокария, энгельгардия, мирика. Тем не менее значительную роль в ландшафтах уже играли умеренно теплолюбивые сережкоцветные (ольха, береза, граб).
Верхняя часть коричневых песков куршской свиты формировалась в более суровых климатических условиях. Так, в позднем олигоцене–раннем миоцене представители умеренно теплолюбивой арктотретичной флоры существенно потеснили теплолюбивые широколиственные в лесных сообществах. Климат тем не менее оставался влажным, на что указывает растительная ассоциация, включающая растения, предпочитающие переувлажненный субстрат – ольха, нисса, мирика, глиптостробус, болотный кипарис. Пески и глины самой верхней части куршской свиты накапливались в более благоприятных климатических условиях, вероятно соответствующих потеплению на рубеже раннего– среднего миоцена, поскольку в сообществах увеличилось количество таких теплолюбивых растений, как миртовые, цирилловые, ореховые (Platycaryapollenites).
В среднем миоцене (замландское время) климат оставался все еще достаточно теплым, в составе смешанных мезофитных лесов присутствовали различные сосны, секвойя, катайя, зонтичная сосна, дубы, каштаны, мирики, энгельгардии, а также теплолюбивые представители сем. Cyrillaceae. Роль мелколиственной растительности была невелика. Тем не менее климатические показатели явно ухудшились, поскольку из состава лесной растительности практически исчезли такие породы, как гикори, платан, птерокария, орех, нисса, падуб. По-видимому, климат стал относительно более сухим, на что указывает отсутствие в составе пыльцы влаголюбивых ногоплодника, ели, глиптостробуса, болотного кипариса.
По таксономическому составу олигоценовая и ранне-среднемиоценовая палинофлоры Самбийского полуострова отличаются от одновозрастных палинофлор Европы отсутствием таких теплолюбивых элементов, как Palmae, Laurus, Cinnamonum, характерных для так называемой полтавской фитогеографической провинции. Обособленное положение этой флоры было отмечено ранее при анализе таксономического состава макроостатков растений из куршской свиты (Буданцев, Свешникова, 1964), что объяснялось тем, что флора Самбийского полуострова является самой северной из европейских флор и могла формироваться на границе двух фитогеографических зон – полтавской и тургайской. В целом южнобалтийская флора олигоцена–среднего миоцена оставалась субтропической, но испытывала на себе влияние тургайского типа растительности.
ЗАКЛЮЧЕНИЕ
В результате проведенного палинологического изучения отложений стратотипа куршской свиты и низов стратотипа замландской свиты в Приморском карьере удалось выявить один диноцистовый и пять спорово-пыльцевых комплексов, таксономический состав и количественная структура которых позволяют уточнить стратиграфический возраст вмещающих отложений, а также сопоставить эти толщи c отложениями сопредельных территорий. “Шоколадные глины” и нижняя часть коричневых песков куршской свиты имеют терминальноприабонский возраст, средняя часть коричневых песков формировалась в раннем олигоцене (рюпель), верхняя часть свиты предположительно соответствует верхнему олигоцену–нижнему миоцену. К среднему миоцену отнесена перекрывающая куршскую свиту замландская свита.
Реконструированы условия осадконакопления, выявлены изменения в растительных сообществах, происходившие в результате постепенного изменения климата в позднем палеогене–начале неогена в Южной Прибалтике. Показано, что олигоценовые и ранне-среднемиоценовые палинофлоры Самбийского полуострова не содержат типично тропических элементов, чем отличаются от одновозрастных европейских ископаемых флор. Вероятно, это произошло вследствие их формирования на границе двух фитогеографических зон – полтавской и тургайской.
Благодарности. Авторы выражают благодарность Генеральному директору ФГУП “Янтарный комбинат” М.И. Зацепину за доступ в Приморский карьер, а также главному геологу Янтарного комбината А.Е. Коркину и научному сотруднику Института Океанологии РАН (Калининград) В.И. Алексееву за помощь в отборе проб. Авторы признательны Л.Ф. Бобковой и Е.В. Тупицыной (ИНГГ СО РАН) за химическую обработку палинологических образцов, а также рецензентам Н.К. Лебедевой, О.В. Шурековой, М.Е. Былинской за ценные замечания и комментарии, послужившие улучшению качества статьи.
Финансирование работы. Настоящая работа выполнена при финансовой поддержке РФФИ и Калининградской области, проект РФФИ № 119-45-390001. Палинологические исследования проведены в рамках темы госзаданий “Микрофитопланктон и высшие растения позднего палеозоя–кайнозоя: проблемы палеофлористики, фитостратиграфии, палеофитогеографии и палеоклиматологии” ГИН РАН, FWZZ-2022-0004 (ИНГГ СО РАН) и FMWE-2021-0012 (ИО РАН).
Список литературы
Ахметьев М.А. Стратиграфия и флора позднего кайнозоя Исландии. М.: Наука, 1978. 188 с. (Тр. ГИН РАН СССР. Вып. 316).
Ахметьев М.А., Александрова Г.Н., Беньямовский В.Н., Васильева О.Н., Железко В.И., Запорожец Н.И. Курганские слои нижнего олигоцена Западной Сибири и Зауралья // Стратиграфия. Геол. корреляция. 2001. Т. 9. № 4. С. 65–79.
Балтакис В.И. Осадочные формации и литологические комплексы палеогена и неогена Южной Прибалтики // Литология и геология полезных ископаемых Южной Прибалтики. Тр. Института геологии (Вильнюс). 1966. Вып. 3. С. 277–323.
Буданцев Л.И., Свешникова И.Н. Третичная флора Калининградского полуострова. Ч. IV // Тр. Ботанического института АН СССР. 1964. Сер. 8. Палеоботаника. № 5. С. 83–112.
Веножинскене А.И. Палинологические комплексы палеогена Южной Прибалтики // Мезозой и кайнозой Южной Прибалтики и Белоруссии. Научн. сообщ. Ин-та геологии и географии АН ЛитССР, 1960. Т. 12. С. 41–47.
Геологические и биотические события позднего эоцена–раннего олигоцена на территории бывшего СССР. Ч. I. Региональная геология верхнего эоцена и нижнего олигоцена. Отв. ред. Крашенинников В.А., Ахметьев М.А. М.: ГЕОС, 1996. 314 с. (Тр. ГИН РАН. Вып. 489).
Запорожец Н.И. Палиностратиграфия и зональное расчленение по диноцистам среднеэоценовых–нижнемиоценовых отложений р. Белой (Северный Кавказ) // Стратиграфия. Геол. корреляция. 1999. Т. 7. № 2. С. 61–78.
Запорожец Н.И., Ахметьев М.А. Палеоботаническое изучение разреза майкопской серии олигоцена–нижнего миоцена по р. Белая выше г. Майкопа, Предкавказье // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 6. С. 61–83.
Зосимович В.Ю. Граница олигоцена и миоцена в бореальной провинции Восточно-Европейской платформы // Геол. журн. (Киев). 1991. № 3. С. 89–98.
Зосимович В.Ю., Шевченко Т.В. Палеогенові відклади Північноукраїнської палеоседиментаційної провінції // Палеоокеанографія та палеоседиментогенез. Збiрник наукових праць Iнституту геологiчних наук НАН Украïни. 2015. Т. 8. С. 68–121.
Кузьмина О.Б., Яковлева А.И. Новые данные о спорово-пыльцевой характеристике верхнеэоценовых отложений Самбийского полуострова, Калининградская область // Стратиграфия. Геол. корреляция. 2023. Т. 31. № 3. С. 99–115.
Мурашко Л.И., Рылова Т.Б., Якубовская Т.В. Поздний олигоцен Беларуси // Стратиграфия. Геол. корреляция. 1998. Т. 6. № 4. С. 67–80.
Практическая палиностратиграфия. Ред. Панова Л.А., Ошуркова М.В., Романовская Г.М. Л.: Недра, 1990. 348 с.
Стотланд А.Б. Микрофитофоссилии позднего эоцена–среднего миоцена Днепровско-Донецкой впадины и их стратиграфическое значение. Автореф. дисс. … канд. геол.-мин. наук. Киев, 1984. 26 с.
Стотланд А.Б. Корреляция олигоценовых отложений Днепровско-Донецкой впадины по динофлагеллатам // Актуальные вопросы современной палеоальгологии. Киев: Наук. думка, 1986. С. 60–65.
Стратиграфические схемы докембрийских и фанерозойских отложений Беларуси. Объяснительная записка. Минск: БелНИГРИ, 2010. 282 с.
Харин Г.С., Лукашина Н.П. Условия образования и корреляция янтареносной прусской свиты (верхний эоцен, Калининградская область) // Стратиграфия. Геол. корреляция. 2002. Т. 10. № 2. С. 93–99.
Яковлева А.И. Детализация эоценовой диноцистовой шкалы для восточного Перитетиса // Бюлл. Моск. о-ва испытателей природы. Отд. геол. 2017. Т. 92. Вып. 2. С. 32–48.
Akkiraz M.S., Akgün F. Palynology and age of the Early Oligocene units in Cardak-Tokca basin, southwest Anatolia: paleoecological implications // Geobios. 2005. V. 38. P. 283–299.
Akkiraz M.S., Akgün F., Örçen S. Stratigraphy and palaeoenvironment of the Lower–“middle” Oligocene units in the northern part of the Western Taurides (Incesu area, Isparta, Turkey) // J. Asian Earth Sci. 2011. V. 40. P. 452–474.
Boulter M.C., Craig D.L. A middle Oligocene pollen and spore assemblage from the Bristol channel // Rev. Paleobot. Palynol. 1979. V. 28. P. 259–272.
Brinkhuis H. Late Eocene to early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994. V. 107 (1–2). P. 121–163.
Brinkhuis H., Biffi U. Dinoflagellate cyst stratigraphy of the Eocene/Oligocene transition in central Italy // Marine Micropaleontol. 1993. V. 22. P. 131–183.
Châteauneuf J.J. Evolution of the microflora and dinocysts at the Eocene–Oligocene boundary in Western Europe // Terminal Eocene events. Eds. Pomerol C., Premoli-Silva I. Amsterdam: Elsevier Science Publishers B.V., 1986. P. 289–292.
Egger L.M., Sliwinska K.K., Van Peer T.E., Liebrand D., Lippert P.C., Friedrich O., Wilson P.A., Norris R.D., Pross J. Magnetostratigraphically-calibrated dinoflagellate cyst bioevents for the uppermost Eocene to lowermost Miocene of the western North Atlantic (IODP Expedition 342, Paleogene Newfoundland sediment drifts) // Rev. Palaeobot. Palynol. 2016. V. 234. P. 159–185.
Gedl P. Dinoflagellate cyst record of the Eocene–Oligocene boundary succession in flysch deposits at Leluchow, Carpathian Mountains, Poland // Geol. Soc. London. Spec. Publ. 2004. V. 230 (1). P. 309–324.
Gedl P., Worobiec E., Słodkowska B. Palynology of Lower Oligocene brown coal and lower most Middle Miocene sand deposits from the Łukowa-4 borehole (Carpathian Foredeep, SE Po land) – implications for palaeogeographical reconstructions // Geol. Quart. 2016. V. 60 (4). P. 943–958.
Göppert H.R., Berendt G.C. Der Bernstein und die in ihm befindlichen Pflanzenreste der Vorwelt. Berlin, 1845. 126 p.
Heilmann-Clausen C., Van Simaeys S. Dinoflagellate cysts from the Middle Eocene to ?Lowermost Oligocene succession in the Kysing Research borehole, central Danish Basin // Palynology. 2005. V. 29 (1). P. 143–204.
Hochuli P.A. Palynologische untersuchungen im Oligosen der zentralen und westlichen Paratethys // Beitrrage Palaontologie. 1978. Osterreich. V. 4. P. 1–132.
Iakovleva A.I., Aleksandrova G.N., Mychko E.V. Late Eocene (Priabonian) dinoflagellate cysts from Primorsky quarry, southeast Baltic coast, Kaliningrad Oblast, Russia // Palynology. 2021. https://doi.org/10.1080/01916122.2021.1980743
Kosmowska-Ceranowisz B., Müller C. Lithology and calcareous nannoplankton in amberbearing Tertiary sediments from boreholes Chlapovo // Bull. Ac. Pol. Terre. 1985. V. 33. P. 119–129.
Nickel B. Palynofazies und Palynostratigraphie der Pechelbronner Schichten im nordlichen Oberrheintalgraben // Palaeontographica. 1996. Abteilung B. Ionnides 240. P. 1–151.
Piwocki M. Paleogen // Budowa geologiczna Polski, 1: stratygrafia, 3a: kenozoik, paleogen, neogen. Eds. Peryt T.M., Piwocki M. Warszawa: Pañstwowy Instytut Geologiczny, 2004. P. 22–71 (in Polish).
Piwocki M., Ziembinska-Tworzydło M. Neogene of the Pollish Lowlands – lithostratigraphy and pollen-spore zones // Geol. Quart. 1997. V. 41. № 1. P. 21–40.
Schweigger A.F. Beobachtungen auf naturhistorischen Reisen. Berlin, 1819. 130 p.
Słodkowska B. Palynological studies of the Paleogene and Neogene deposits from the Pomeranian Lakeland Area (NW Poland) // Polish Geol. Instit. Spec. Pap. 2004. № 14. P. 1–116.
Słodkowska B. Palynology of the Paleogene and Neogene from the Warmia and Mazuri areas (NE Poland) // Geologos. 2009. V. 15. № 3–4. P. 219–234.
Thomas K. Die Bernsteinformation des Samlandes // Preußische Provinzial-Blätter. 1847. Bd. 3. P. 241–245.
Von Heer O. Miocene Baltische Flora // Beiträge zur Naturkunde Preussens. Königlichen physikalisch-öconomischen Gesellschaft zu Königsberg. Königsberg, 1869. 104 p.
Zaddach G. Űber die Bernstein- und Braunkohlenlager des Samlandes // Schriften der Physikalisch-Ökonomischen Gesellschaft zu Königsberg in Pr. 1860. Jg. 1. P. 1–44.
Zaddach G. Das Tertiärgebirge des Samlandes // Schriften der Physikalisch-Ökonomischen Gesellschaft zu Königsberg in Pr. 1868. J. 8. P. 85–197.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция