

Стратиграфия. Геологическая корреляция, 2023, T. 31, № 1, стр. 52-84
Таксономия и биостратиграфическое значение тоарских двустворчатых моллюсков рода Meleagrinella Whitfield, 1885
О. А. Лутиков 1, *, Г. Арп 2, **
1 Геологический институт, Российская академия наук
Москва, Россия
2 Геонаучный центр Гёттингенского университета им. Георга-Августа
Гёттинген, Германия
* E-mail: niipss@mail.ru
** E-mail: garp@gwdg.de
Поступила в редакцию 24.04.2022
После доработки 12.06.2022
Принята к публикации 21.06.2022
- EDN: JOAZPS
- DOI: 10.31857/S0869592X23010040
Аннотация
Исследована морфология лигаментного блока и наружных частей раковины тоарских двустворчатых моллюсков рода Meleagrinella (семейство Oxytomidae Ichikawa, 1958). На основе выводов по эволюции онтогенезов лигаментной ямки, сделанных в результате изучения обширного материала из Восточной Сибири и Германии, и с учетом данных по микроскульптуре остракума реконструирован филогенез Meleagrinella и Arctotis в тоаре. Осуществлена ревизия вида “Monotis” substriata (Münster, 1831), широко распространенного в тоарских отложениях Европы, Азии и Северной Америки. П-утем сравнения выборок с разных стратиграфических уровней нижнего тоара Восточной Сибири, Северо-Востока России и Германии обосновано выделение трех видов: Meleagrinella (Praemeleagrinella?) golberti sp. nov., M. (Clathrolima) substriata (Münster) и M. (Meleagrinella) prima sp. nov. Приведены изображения аммонита Tiltoniceras sp. ind., верхнеплинсбах-тоарских двустворчатых моллюсков рода Meleagrinella и верхнетоарских двустворчатых моллюсков рода Arctotis. Впервые изображена микроскульптура остракума тоарских представителей родов Arctotis и Meleagrinella. В верхнем плинсбахе–нижнем тоаре установлена последовательность таксонов окситомид и на основании бореальной аммонитовой шкалы дана оценка их биостратиграфического значения. Последовательность представлена четырьмя видами: (1) Meleagrinella (Praemeleagrinella) deleta (верхний плинсбах), (2) M. (Praemeleagrinella ?) golberti (зоны Tiltoniceras antiquum, Harpoceras falcifer), (3) M. (Clathrolima) substriata (зона Dactylioceras commune), (4) M. (Meleagrinella) prima (зоны Zugodactylites braunianus, Pseudolioceras compactile).
ВВЕДЕНИЕ
Двустворчатые моллюски рода Meleagrinella Whitfield, 1885 (семейство Oxytomidae Ichikawa, 1958) являются одной из наиболее важных групп фауны для биостратиграфии бореального тоара. В тоаре эта группа окситомид характеризуется маленькими размерами, тонкой ребристой радиальной скульптурой и относится большинством современных специалистов к виду Meleagrinella substriata (Münster). Вид “Monotis” substriata впервые был описан Г. Мюнстером из нижней юры Германии (Münster, 1831). Рисунки нескольких экземпляров этого таксона были приведены в монографии Г.А. Гольдфусса (Goldfuss, 1835, p. 138, tab. XX , figs. 7a–7f ). При описании вида утверждалось, что образцы происходят из разных местонахождений (Münster, 1831, p. 407; Goldfuss, 1835, p. 138). Их находки указывались из таких мест, как Банц, Вайс-Майн, Кульмбах, Хайлигенштадт, Мистельгау, Эшенау, Амберг, Альтдорф, Берг, Эллинген, где вскрывается нижнетоарская формация “Posidonienschiefer”, а образцы из местности Вассеральфинген происходят из верхнеааленской формации “Eisensandstein”. Согласно представлениям Ф.А. Квенштедта, вид “Monotis” substriata в Германии приурочен к уровню распространения Ammonites (=Dactylioceras) anguinus (Reinecke) и встречается стратиграфически ниже уровня с Ammonites (=Catacoeloceras) crassus (Young et Bird) (Quenstedt, 1858). Позднее этот таксон указывался во всех трех аммонитовых зонах нижнего тоара Германии – Dactylioceras tenuicostatum (Hoffmann, Martin, 1960), Harpoceras falcifer и Hildoceras bifrons (Urlichs, 1971). Широко распространенных в нижнем тоаре России и Германии “монотисов” с середины ХХ в. сначала относили к Pseudomonotis substriata (Атлас…, 1947; Бодылевский, Шульгина, 1958), затем к Pseudomonotis (Meleagrinella) substriata (Hoffmann, Martin, 1960) либо к Meleagrinella substriata (Полевой…, 1968; Urlichs, 1971; Стратиграфия…, 1976; Князев и др., 1991).
В России Pseudomonotis cf. substriata впервые были определены В.И. Бодылевским по сборам С.В. Обручева (колымская экспедиция 1929–1930 гг.) из тоарских отложений по р. Мунугуджак (приток р. Омолон) (Атлас…, 1947). Позднее эти отложения датировались как позднетоарские (А.А. Дагис, А.С. Дагис, 1965; Дагис, 1968). Из этого же разреза И.В. Полуботко описала вид Meleagrinella substriata (Полевой…, 1968). Мелеагринелловидные формы со слабовыраженной радиальной скульптурой из средней и верхней частей тоарского яруса (разрезы рр. Токур-Юрях, Левый Кедон, Вилига) И.В. Полуботко отнесла к виду M. faminaestriata Polubotko (Полевой…, 1968). Согласно современным представлениям, к M. ex gr. substriata и M. substriata относятся мелеагринеллы из зон Tiltoniceras antiquum и Harpoceras falcifer на Северо-Востоке России, а “мелеагринеллы” из зон Dactylioceras commune, Zugodactylites braunianus и Pseudolioceras rosenkrantzi рассматриваются в составе Meleagrinella faminaestriata (Репин, Полуботко, 2004; Решения…, 2009).
В Западной Сибири вид Pseudomonotis substriata впервые был определен В.И. Бодылевским из керна скважины 3-р Усть-Енисейского района (Бодылевский, Шульгина, 1958). Виды M. cf. substriata и M. substriata приводились Б.Н. Шурыгиным из керна еще двух скважин – Стахановской-390 (междуречье Пур–Обь) и Верхне-Толькинской-5 (междуречье Пур–Енисей) (Шурыгин и др., 2000). В нижней части тоара Западной Сибири выделены слои с Dacryomya inflata, Tancredia bicarinata, в характерном комплексе которых указана Meleagrinella sp., а в средней части тоара выделены слои с M. faminaestriata (Решение…, 2004).
В Восточной Сибири вид M. cf. substriata указывался в подошве тоарских отложений Анабарской губы (Стратиграфия…, 1976). Эта пачка в дальнейшем была включена в верхи плинсбаха (Шурыгин и др., 2000). В разрезе р. Келимяр вид M. substriata отмечался в зоне Harpoceras falcifer (Князев и др., 1984). В разрезах рр. Марха, Тюнг и Анабарской губы из зон Dactylioceras commune, Zugodactylites braunianus приводился вид M. faminaestriata (Князев и др., 1991, 2003). На Дальнем Востоке вид M. ex gr. substriata указывался из зоны Zugodactylites braunianus (Решения…, 1994).
При проведении ревизии юрских окситомид (Лутиков, Шурыгин, 2010) виды Avicula substriata (Zieten, 1830) из синемюра Германии и Monotis substriata (Münster, 1831) из тоара Германии предположительно рассматривались в составе единой геттанг-раннетоарской группы мелеагринелл – Meleagrinella (Praemeleagrinella) Lutikov et Schurygin. Поскольку название “substriata” было преоккупировано синемюрским таксоном, все раннетоарские экземпляры предварительно были отнесены к Praemeleagrinella aff. substriata (Zieten). Восточносибирские окситомиды из зоны Zugodactylites monestieri на основании строения лигаментного и биссусного блоков были отнесены к виду Arctotis (Praearctotis) milovae, выделенного Т.М. Окуневой (2002) на материале из верхнетоарских отложений Восточного Забайкалья. При переизучении типовой коллекции Meleagrinella faminaestriata, хранящейся в отделе стратиграфии и палеонтологии ВСЕГЕИ, и собственного материала О.А. Лутикова, происходящего из типовой местности (Северо-Восток России, бассейн р. Левый Кедон), был сделан вывод, что данный вид не относится к роду Meleagrinella (Лутиков, Шурыгин, 2010). По расположению лигаментных площадок в плоскости смыкания створок вид “Meleagrinella” faminaestriata более близок к роду Praebuchia Zakharov, 1981.
В последнее десятилетие сделаны новые находки M. substriata из тоара Западной Европы, как в типовой местности – в Баварии, Германия (Teichert, Nützel, 2015; Arp, Gropengiesser, 2016), так и в других районах Европы – в Йоркшире, Англия (Caswell et al., 2009; Morris et al., 2019) и Венгрии (Szente, 2015). Мофологически близкие мелеагринеллы распространены в нижнем тоаре Западной Канады (Martindale, Aberhan, 2017).
Таким образом, согласно существовавшим в ХХ в. взглядам, мелеагринеллы из зон Tiltoniceras antiquum, Harpoceras falcifer и низов верхнего тоара на Северо-Востоке России относились к виду M. substriata. Мелеагринеллы из зон Dactylioceras commune, Zugodactylites braunianus и Pseudolioceras rosenkrantzi рассматривались в составе вида M. faminaestriata. В Германии к виду M. substriata относили все мелеагринеллы из зон Dactylioceras tenuicostatum, Harpoceras falcifer и Hildoceras bifrons.
МАТЕРИАЛ
Основным материалом для исследований послужили коллекции О.А. Лутикова, собранные в период с 1980 по 1987 гг. в естественных выходах тоарских отложений на Омолонском массиве (бассейн р. Левый Кедон), в Приверхоянском прогибе (р. Моторчуна), Вилюйской синеклизе (рр. Марха, Вилюй), Лено-Анабарском прогибе (р. Келимяр), Енисей-Хатангском прогибе (Анабарская губа, р. Анабар, Восточный Таймыр) (рис. 1), и коллекции Г. Арпа, собранные в период с 2010 по 2014 гг. в местностях Дёрльбах (Канал Людвига) и Берг (Южная Германия) (рис. 2).
Рис. 1.
Обзорная карта изученных разрезов тоара Восточной Сибири и Северо-Востока России. 1 – Северо-Восток России: бассейн р. Левый Кедон (рр. Сатурн, Старт, Бродная, Астрономическая); 2–10 – Восточная Сибирь: 2 – р. Моторчуна, 3 – мыс Цветкова (Восточный Таймыр), 4 – Анабарская губа, 5 – р. Анабар, 6 – р. Келимяр, 7 – р. Марха, 8 – р. Вилюй, 9 – р. Тюнг, 10 – рр. Сюнгюдэ, Молодо; 11–14 – участки бурения: 11 – Тенкеляхский (междуречье Тюкян–Марха), 12 – Правобережный (междуречье Марха–Вилюй), 13 – Серки-Линденский (междуречье Тюнг–Лена), 14 – Оттурский (междуречье Марха–Вилюй).
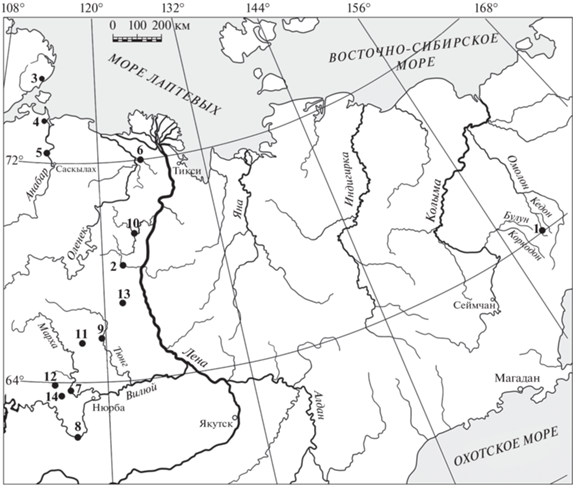
Рис. 2.
Карта расположения местонахождений разрезов тоара Германии, из которых происходят изученные коллекции. 1–5 – Южная Германия: 1 – Банц, 2 – Мистельгау, 3 – Альтдорф, 4 –Дёрльбах, 5 – Берг; 6, 7 – Северная Германия: 6 – Аденштедт, 7 – Веллерсен.

Дополнительно были изучены большие коллекции, любезно предоставленные коллегами: В.Г. Князевым (р. Тюнг), Б.Н. Шурыгиным (Анабарская губа), И.В. Будниковым, В.П. Девятовым и И.С. Павлухиным (Тенкеляхский, Правобережный, Серки-Линденский, Оттурский участки разведочного бурения), С. Сеппельтом (S. Seppelt) (временный строительный карьер в местности Аденштедт, Северная Германия).
Были изучены синтипы вида Monotis substriata из коллекции Г. Мюнстера, происходящие из местностей Банц и Альтдорф, хранящиеся в музее Г. Гольдфусса (Goldfuss-Museum) (Бонн, Германия). Образцы изучались по фотографиям, любезно предоставленным коллегами из Института наук о Земле и палеонтологии (Institute for Geosciences, Palaeontology) (Бонн, Германия) – доктором Г. Хойманном (G. Heumann) и фотографом Г. Олешински (G. Oleschinski). Также по фотографиям были изучены синтипы типовой серии вида Monotis substriata из коллекции Г. Мюнстера, происходящие из местности Мистельгау, хранящиеся в Баварском государственном хранилище палеонтологических и геологических коллекций (Bayerische Staatssammlung für Paläontologie und Geologie) (Мюнхен, Германия). Фотографии любезно предоставил доктор В. Вернер (W. Werner). Часть изученной коллекции хранится в Государственном геологическом музее (ГГМ) им. В.И. Вернадского (Москва).
В коллекциях двустворчатых моллюсков, относящихся к роду Meleagrinella, собранных в нижнем тоаре, насчитывается около 200 экземпляров из четырех разрезов Северо-Востока России, около 800 экземпляров из тринадцати разрезов Восточной Сибири, около 20 экземпляров из шести разрезов Германии.
МЕТОДЫ
Методы систематики. В своих исследованиях авторы руководствовались основными принципами, привлекаемыми для филогенетических реконструкций: хронологическим, хорологическим, онтогенетическим, гомологическим и основного звена (Руженцев, 1960). Для установления хронологии эволюционных преобразований в пределах изучаемой группы использовалась аммонитовая шкала, разработанная для Северо-Западной Европы (Howarth, 1992; Page, 2003), а также бореальный аммонитовый стандарт (Князев и др., 2003). Выборки из локальных популяций привязывались к уровням с аммонитами. Онтогенез изучался по сопоставлению очертаний элементов лигаментного блока у раковин на разных стадиях их индивидуального развития и по линиям роста элементов замочных структур на отдельных раковинах взрослых особей. Выявление изменений в строении лигаментного и биссусного блоков внутри группы проводилось путем сравнительного анализа положения гомологичных частей раковин в выборках из ископаемых популяций. Для распределения таксонов по группам за основное звено развития были приняты изменения относительного положения краев лигаментной ямки, определяющих ее форму.
На основании рабочих гипотез, базирующихся на визуальной оценке изменений гомологичных частей в лигаментном блоке во времени – морфогенетических трендов, была проведена классификация надвидовых таксонов. Выводы по эволюции надвидовых групп внутри семейства основывались на трансформациях лигаментного блока в ископаемых популяциях, отделенных друг от друга стратиграфическими интервалами.
Методы таксономии. При выборе методов таксономии авторы следовали эволюционному подходу Дж. Г. Симпсона (Симпсон, 2006) и Э. Майра (Майр, 1971), согласно которому систематика строилась на основе объединения в группы по взвешенным признакам без привлечения филогенетических гипотез и затем интерпретировалась на основе эволюционного подхода. Морфология таксонов изучалась в три этапа:
1) проводилось всестороннее исследование фоссилий по всем возможным морфологическим признакам;
2) проводилось апостериорное взвешивание признаков, основанное на опыте оценки количества филогенетически значимой информации, содержащейся в выбранном признаке, и выбирались таксономически значимые признаки;
3) на основе выбранных признаков и изучения морфогенеза изученные экземпляры объединялись в видовые и надвидовые таксоны; выводы о родстве делались на основании морфологического сходства гомологичных элементов.
Морфология раковин. В литературе, посвященной двустворчатым моллюскам семейства Oxytomidae (Борисяк, 1909, 1914; Кошелкина, 1963; Treatise…, 1969; Невесская и др., 2013), некоторые морфологические элементы не определены достаточно точно. С целью обозначения границ морфологических элементов применялось зонирование створок с помощью геометрических фигур. Фотографию створки, сделанную из положения сверху, вырезали по контуру створки в программе Photoshop и затем переносили в программу Corel DRAW. С помощью инструментов программы Corel DRAW внешний контур створки вписывали в большой прямоугольник I. Горизонтальные стороны прямоугольника I ориентировали параллельно линии замочного края. Вертикальные стороны наружного прямоугольника I проводили касательно к выступающим краям раковины. Фотографию створки, сделанную со стороны переднего края створки, вписывали в малый прямоугольник II. Двумерную проекцию раковины разделяли на участки с помощью внутреннего прямоугольника III. Верхнюю горизонтальную сторону прямоугольника III ориентировали параллельно замочному краю. Одну вертикальную сторону прямоугольника III проводили перпендикулярно замочному краю по касательной к переднему концу раковины, вторую вертикальную сторону проводили перпендикулярно замочному краю по касательной к вырезу заднего края до пересечения с контуром створки. Нижнюю горизонтальную сторону прямоугольника III проводили от точки пересечения с контуром створки и ориентировали параллельно замочному краю (рис. 3). Этот подход позволил определить относительные границы частей створки, сформулировать термины для обозначения признаков и провести количественную оценку признаков и их состояний, используемых при описании таксонов.
Рис. 3.
Схема морфологических элементов раковины Meleagrinella. a – вид левой створки изнутри; б – вид раковины со стороны переднего края; в, г – вид левой створки снаружи. Лс – левая створка, Пс – правая створка, М – макушка, У – умбо, П – продиссоконх, ПКл – передний киль створки, ЗЗП – заднее залигаментное поле, ЗКр – заднее крыло, ПКр – переднее крыло, ЦО – центральная ось, ЗМК – замочный край, ВК – верхний (спинной) конец створки, ПК – передний конец створки, ЗК – задний конец створки, НК – нижний (брюшной) конец створки, МВпЛ – максимальная выпуклость левой створки, МВпП - максимальная выпуклость правой створки, ВКС – верхний край створки (отрезок 1–2), ЗКС – задний край створки (отрезок 2–3), НКС – нижний край створки (отрезок 3–4), ПКС – передний край створки (отрезок 4–1), ЗУ – заднее ушко, ОЗУ – окончание заднего ушка, ОПУ – окончание переднего ушка, ДпЗУ – депрессия заднего ушка, ДпЗКр – депрессия заднего крыла, ВЗК– вырез заднего края, НОЗКр – нижнее окончание заднего крыла, ВЗКр – вершина заднего крыла, Х – хорда заднего крыла, Р – ребра, ПСС – плоскость смыкания створок.
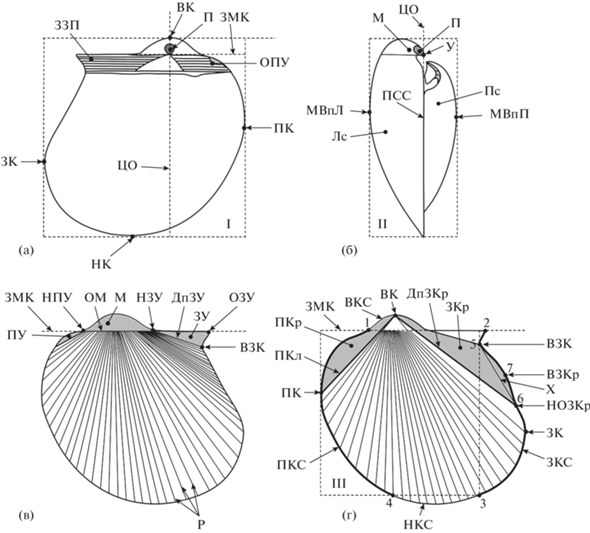
Наружные признаки. Точки пересечения внутреннего прямоугольника III с контуром раковины условно приняты за точки границ между краями створки (рис. 3):
задний край створки (ЗКС) – участок контура раковины от окончания заднего ушка до точки пересечения с прямоугольником III (между точками 2–3);
нижний (брюшной) край створки (НКС) – участок контура раковины между двумя точками пересечения внутреннего прямоугольника III с внешним краем (между точками 3–4);
передний край створки (ПКС) – участок контура раковины между двумя точками пересечения большого внутреннего прямоугольника с внешним краем (между точками 4–1);
верхний (спинной) край створки (ВКС) – участок контура раковины между двумя точками пересечения большого внутреннего прямоугольника с внешним краем (между точками 1–2).
Наиболее выступающие точки переднего, заднего, брюшного и спинного краев створки обозначены как передний конец створки (ПК), задний конец створки (ЗК), нижний конец створки (НК) и верхний конец створки (ВК).
Другие признаки обозначены следующим образом:
замочный край (ЗМК) – прямая линия, параллельная верхнему краю заднего ушка;
умбо (У) – клювообразный конец левой створки, в апикальной точке которого находится первичная раковина велигера – продиссоконх (П);
центральная ось створки (ЦО) – прямая линия, проходящая через умбо (У) и нижний край створки (НКС), перпендикулярно к замочному краю;
макушка (М) – часть раковины, выступающая над замочным краем; макушка имеет вершину, совпадающую с верхним концом створки (ВК), и основание макушки (ОМ);
основание макушки (ОМ) – средняя часть замочного края, ограниченная верхним краем створки;
начало заднего ушка (НЗУ) – точка пересечения замочного края створки с основанием макушки, расположенная ближе к заднему краю створки;
начало переднего ушка (НПУ) – точка пересечения замочного края створки с основанием макушки, расположенная ближе к переднему краю створки;
окончание заднего ушка (ОЗУ) – остроугольный изгиб (точка 2), образованный пересечением верхнего и заднего краев створки;
вырез заднего края (ВЗК) – выемка на заднем крае створки ниже окончания заднего ушка;
заднее ушко (ЗУ) – треугольная область в задней части створки вблизи замочного края, отделенная от остальной части раковины депрессией; форма ушка определяется точкой начала заднего ушка, точкой окончания ушка и точкой (точка 5) максимального выреза заднего края;
депрессия заднего ушка (ДпЗУ) – линия прогиба поверхности створки, соединяющая начало заднего ушка (НЗУ) и вырез заднего края (ВЗК) и отделяющая заднее ушко со стороны верхнего края створки от заднего крыла;
заднее крыло (ЗКр) – треугольная волнообразная складка в задней части створки, занимающая положение между задним ушком и основной частью створки, отделенная сверху депрессией заднего ушка и снизу депрессией заднего крыла; поверхность заднего крыла отделена от остальной поверхности створки большей плотностью ребристости;
передний киль (ПКл) – линия перегиба поверхности створки, соединяющая верхний конец створки и передний конец створки;
депрессия заднего крыла (ДпЗКр) – линия прогиба наружной поверхности створки, отделяющая складку заднего крыла створки от большей части створки со стороны нижнего края;
нижнее окончание заднего крыла (НОЗКр) – выемка (точка 6) на заднем крае створки в месте пересечения депрессии заднего крыла (ДпЗКр) и заднего края створки (ЗКС); нижнее окончание заднего крыла левой створки может быть расположено либо выше заднего конца створки, либо ниже его;
вершина заднего крыла (ВЗКр) – точка (7) максимального изгиба дуги заднего крыла;
хорда заднего крыла (Х) – линия, соединяющая вырез заднего края (ВЗК) с нижним окончанием заднего крыла (НОЗКр);
точка максимальной выпуклости левой створки (МВпЛ) – точка максимальной кривизны наружной поверхности левой створки;
точка максимальной выпуклости правой створки (МВпП) – точка максимальной кривизны наружной поверхности правой створки.
Внутренние признаки. Замок у изученных форм лишен зубов и относится к “птеринеоидному типу” (Невесская и др., 1971). Элементы замка, связанные в основном с функционированием лигамента, рассматриваются в составе лигаментного блока, а элементы, большей частью связанные с функционированием биссуса, в составе биссусного блока. В лигаментный блок рода Meleagrinella (рис. 4), как и у морфологически близкого к нему рода Arctotis, входят: (1) элементы лигаментной площадки – лигаментная ямка (ЛЯ), переднее и заднее залигаментные поля (ПЗП, ЛЗП), лигаментные бороздки (ЛБ), передний и задний лигаментные желобки (ПЛЖ и ЗЛЖ), лигаментный валик (ЛВ); (2) элементы псевдосвязочной площадки – латеральный валик (ЛТВ), протуберанец (ПР). В биссусный блок входят: биссусное ушко (БУ), биссусный желобок (БЖ) и биссусный вырез (БВ). Термины использовались в понимании, принятом автором ранее (Лутиков, Шурыгин, 2010; Лутиков, 2021).
Рис. 4.
Схема строения замка Meleagrinella: (а) – левой створки; (б, в) – правой створки (а, б – лигаментный блок, в – биссусный блок). ЛЯ – лигаментная ямка, ПЛЖ – передний лигаментный желобок, ЗЛЖ – задний лигаментный желобок, ЛВ – лигаментный валик, ЛБ – лигаментные бороздки, НКЯ – нижний край лигаментной ямки, ЗКЯ – задний край лигаментной ямки, ПКЯ – передний край лигаментной ямки, БУ – биссусное ушко, БЖ – биссусный желобок, БВ – биссусный вырез, ЛТВ – латеральный валик, ПР – протуберанец, ЗЗП – заднее залигаментное поле, ПЗП – переднее залигаментное поле; ПХ – продиссоконх, ПП – псевдосвязочная площадка.
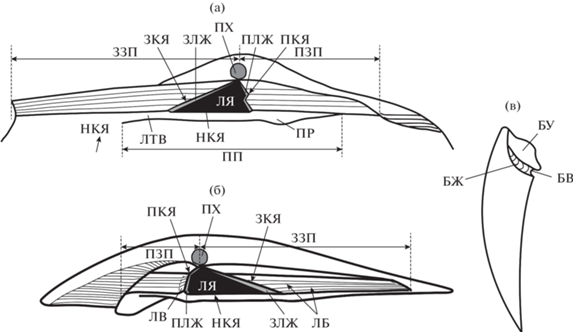
Новыми терминами являются:
нижний край лигаментной ямки (НКЯ) – кромка лигаментной ямки в месте соединения лигаментной площадки и псевдосвязочной площадки;
задний край лигаментной ямки (ЗКЯ) – кромка лигаментной ямки, образованная пересечением заднего залигаментного поля и заднего лигаментного желобка;
передний край лигаментной ямки (ПКЯ) – кромка лигаментной ямки, образованная пересечением переднего залигаментного поля и переднего лигаментного желобка.
Измерения и количественная оценка признаков. При проведении замеров с помощью штангенциркуля измерялся только один параметр. В зависимости от сохранности материала использовались значения высоты или длины створки. В целях уменьшения погрешности остальные параметры определялись по фотографиям раковин, трансформированным в векторную форму, с помощью инструментов программы Corel DRAW. Для количественной оценки признаков лигаментного блока проводились измерения элементов лигаментной ямки и лигаментной площадки по трем признакам (рис. 5а):
Рис. 5.
Схема замеров раковин Meleagrinella. a – вид левой створки изнутри; б – вид раковины со стороны переднего края; в, г – вид левой створки снаружи. Морфологические элементы: НОЗКр – нижнее окончание заднего крыла, ВЗК – вырез заднего края, ВЗКр – вершина заднего крыла, ЗМК – замочный край, ПКл – передний киль, ДпЗКр – депрессия заднего крыла, Х – хорда заднего крыла, МВпЛ – точка максимальной выпуклости левой створки, МВпП – точка макимальной выпуклости правой створки. Метрические внутренние признаки: ДЗП – длина заднего залигаментного поля, ДНЛ – длина нижнего края лигаментной ямки, ДЗЛ – длина проекции заднего края лигаментной ямки. Метрические наружные признаки: ВЛс – высота левой створки, ВПс – высота правой створки, Д – длина, ВпЛ – выпуклость левой створки, ВпП – выпуклость правой створки, ДЗЧ – длина задней части створки, ДПЧ – длина передней части створки, УЗКр – угол заднего крыла, УЗУ – угол заднего ушка, ДЗКр – длина заднего крыла, ШЗКр – ширина заднего крыла, ШР – ширина ребра, ШП – ширина межреберного промежутка.
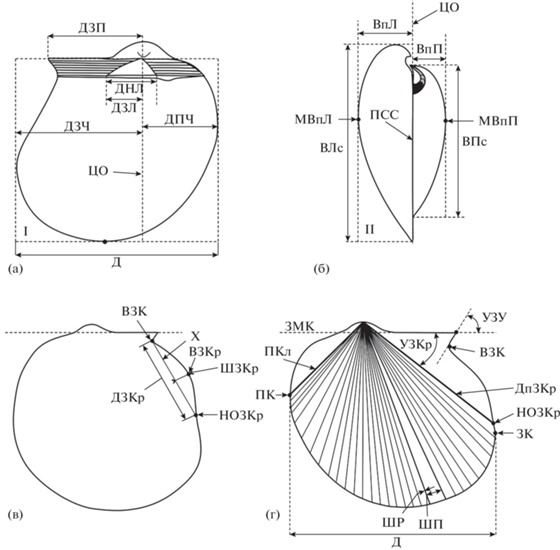
длина нижнего края лигаментной ямки (ДНЛ) – расстояние между точками пересечения боковых сторон лигаментной ямки с нижним краем лигаментной площадки;
длина проекции заднего края лигаментной ямки (ДЗЛ) – расстояние между двумя параллельными прямыми, одна из которых перпендикулярно проходит через точку пересечения боковых сторон лигаментной ямки, а другая через точку пересечения задней стороны лигаментной ямки с нижним краем лигаментной площадки;
длина заднего залигаментного поля (ДЗП) – расстояние от центральной оси створки до окончания заднего ушка.
Для характеристики наружных морфологических элементов раковин Meleagrinella производились замеры по 12 признакам (рис. 5а–5г):
высота створки (В) – расстояние между апикальным выступом верхнего края и апикальным выступом нижнего заднего края;
длина створки (Д) – расстояние между апикальным выступом переднего края и апикальным выступом заднего края;
длина задней части створки (ДЗЧ) – расстояние между центральной осью створки и касательной к апикальному выступу заднего края;
длина передней части створки (ДПЧ) – расстояние между центральной осью створки и апикальным выступом переднего края;
угол заднего крыла (УЗКр) – угол между замочным краем (ЗМК) и депрессией заднего крыла (ДпЗКр);
угол заднего ушка (УЗУ) – угол между замочным краем (ЗМК) и линией, соединяющей окончание заднего ушка (ОЗУ) с вырезом заднего края (ВЗК);
ширина заднего крыла (ШЗКр) – расстояние между хордой заднего крыла (Х) и вершиной заднего крыла (ВЗКр);
длина заднего крыла (ДЗКр) – расстояние между вырезом заднего края (ВЗК) и нижним окончанием заднего крыла (НОЗКр);
выпуклость створки (Вп) – расстояние между точкой максимальной выпуклости створки (МВпЛ или МВпП) и центральной осью, совпадающей у Meleagrinella с плоскостью смыкания створок (ПСС);
ребристость створки (Р) – количество ребер на участке раковины в промежутке между передним килем створки и депрессией заднего крыла створки;
ширина ребер (ШР) – расстояние между краями ребер;
ширина межреберных промежутков (ШП) – расстояние между соседними ребрами.
Количественная оценка признаков проводилась на основе системы градаций, примененной при изучении рода Arctotis (Лутиков, 2021), с дополнениями. Для характеристики таксонов на основании количественной оценки признаков использовалось 12 размеров и отношений, переведенных в условные градации (табл. 1).
Таблица 1.
Градации раковин Meleagrinella по особенностям формы и скульптуры
| Величина В, мм |
Градации по размеру створки (1) |
Значение В/Д |
Градации по контуру (2) | Значение ДНЛ/ДЗЛ |
Градации по форме лигаментной ямки (3) |
|---|---|---|---|---|---|
| менее 10 | очень маленькая | менее 0.95 | очень низкая | менее 0.85 | клиновидная |
| 10.01–30 | маленькая | 0.95–0.97 | низкая | 0.85–0.99 | остроугольная |
| 30.01–50 | средняя | 0.98–1.02 | равновеликая | 1.00–1.04 | субпрямоугольная |
| 50.01–70 | большая | 1.03–1.05 | высокая | 1.05–1.50 | широкоугольная |
| более 70 | очень большая | более 1.05 | очень высокая | более 1.50 | субсимметричная |
| Значение ДПЧ/Д |
Градации по симметрии створки (4) | Значение Вп/В |
Градации по степени выпуклости створки (5) | Значение ДЗП/ДЗЧ |
Градации по длине заднего залигаментного поля левой створки (6) |
| менее 0.35 | сильно неравносторонняя | менее 0.20 | слабовыпуклая | менее 0.6 | очень короткое |
| 0.35–0.40 | неравносторонняя | 0.20–0.30 | умеренно-выпуклая | 0.60–0.70 | короткое |
| 0.41–0.45 | умеренно неравносторонняя | 0.31–0.40 | сильновыпуклая | 0.71–0.80 | длинное |
| 0.45–0.50 | равносторонняя | более 0.40 | вздутая | более 0.80 | очень длинное |
| Угол заднего крыла (УЗКр) |
Градации по ширине заднего крыла левой створки (7) | Значение ШЗКр/ДЗКр |
Градации по форме заднего крыла (8) |
Значение ВПс/ВЛс |
Градации по соотношению размеров створок (9) |
| менее 35° | очень узкое | менее 0.05 | прямое | менее 0.80 | сильно неравностворчатые |
| 35°–40° | узкое | 0.05–0.10 | слабовыгнутое | 0.90–0.80 | умеренно неравностворчатые |
| 41°–45° | широкое | 0.11–0.15 | умеренно-выгнутое | более 0.90 | слабо неравностворчатые |
| более 45° | очень широкое | более 0.15 | сильновыгнутое | ||
| Угол заднего ушка (УЗУ) | Градации по форме заднего ушка (10) |
Значение Р/Д |
Градации по плотности ребристости створки (11) | Значение ШП/ШР |
Градации по ширине межреберных промежутков (12) |
| менее 85° | остроугольное | менее 1.00 | редкоребристые | менее 1.50 | очень узкие |
| 85°–105° | субпрямоугольное | 1.00–2.00 | умеренно-ребристые | 1.5–2.00 | узкие |
| более 105° | тупоугольное | 2.01–3.00 | среднеребристые | 2.01–2.50 | умерено широкие |
| 3.01–4.00 | часторебристые | 2.51–3.00 | широкие | ||
| более 4.00 | густоребристые | более 3.00 | очень широкие |
1. По величине высоты створки (В) проводилась размерная градация раковин.
2. Отношение высоты к длине (В/Д) применялось для градации раковин по контуру.
3. Отношение длины нижнего края лигаментной ямки к длине проекции заднего края лигаментной ямки (ДНЛ/ДЗЛ) применялось для градации по форме лигаментной ямки.
4. Отношение длины передней части к длине (ДПЧ/Д) применялось для градации раковин по симметрии.
5. Отношение выпуклости к высоте створки (Вп/В) применялось для градации по степени выпуклости створки.
6. Отношение длины заднего залигаментного поля к длине задней части створки (ДЗП/ДЗЧ) применялось для градации по длине заднего залигаментного поля левой створки.
7. По величине угла заднего крыла (УЗКр) проводилась градация по ширине заднего крыла левой створки.
8. Отношение ширины заднего крыла к длине заднего крыла (ШЗКр/ДЗКр) применялось для градации раковин по форме заднего крыла.
9. По отношению высоты правой створки к высоте левой створки (ВПс/ВЛс) проведена градация по соотношению размеров створок.
10. По величине угла заднего ушка (УЗУ) проведена градация по форме заднего ушка.
11. Отношение числа ребер на участке между передним и задним концом створки к длине створки (Р/Д) использовалось для оценки плотности ребристости створок.
12. Отношение ширины межреберных промежутков к ширине ребер (ШП/ШР) применялось для градации раковин по ширине межреберных промежутков.
Таксономическая значимость признаков. Из-за индивидуальной и возрастной изменчивости наружных морфологических признаков границы между таксонами сложно однозначно определить по малому числу признаков. Объединение в таксоны проводилось методом апостериорного взвешивания признаков, основанным на оценке количества содержащейся филетической информации в выбранном признаке (Майр, 1971). Диагностика надвидовых таксонов осуществлялась с помощью качественной и количественной оценки состояний признаков с большим и средним весом. Для установления степени дискретности видовых таксонов определялся набор любых наружных признаков со средним и малым весом, который был необходим и достаточен для описания вида как замкнутой системы. Полученные в результате таксономических исследований данные сопоставлялись с данными по изучению филогенеза (Лутиков и др., 2010).
К признакам с большим весом отнесены:
а) Форма лигаментной ямки на трех онтогенетических стадиях.
На ювенильной стадии: субпрямоугольная лигаментная ямка характерна для Oxytoma; широкоугольная лигаментная ямка – для Praemeleagrinella; субсимметричная лигаментная ямка – для Meleagrinella s.str. и Arctotis.
На юношеской стадии: сильноскошенная лигаментная ямка характерна для Oxytoma; широкоугольная лигаментная ямка – для Arctotis и Meleagrinella s.str.; субпрямоугольная лигаментная ямка – для Praemeleagrinella; остроугольная лигаментная ямка – для Clathrolima;
На взрослой стадии: клиновидная лигаментная ямка характерна для Oxytoma; остроугольная лигаментная ямка – для Clathrolima; широкоугольная лигаментная ямка – для Arctotis и Meleagrinella s.str. У Praemeleagrinella лигаментная ямка широкоугольная или субпрямоугольная.
б) Длина заднего залигаментного поля. Длинное заднее залигаментное поле характерно для Meleagrinella, короткое – для Arctotis.
в) Соотношение размеров створок. Сильно неравностворчатые раковины характерны для Meleagrinella, слабо неравностворчатые – для Arctotis.
К признакам со средним весом отнесены:
г) Микроскульптура остракума правой створки. Микроскульптура остракума в средней части правой створки у Meleagrinella и Oxytoma представлена четырехугольными призмами, расположенными параллельно радиальным ребрам. Микроскульптура остракума правой створки у Arctotis представлена шестиугольными призмами, расположенными мозаично.
д) Форма заднего ушка. Остроугольное заднее ушко характерно для Praemeleagrinella и среднеюрских Clathrolima; субпрямоугольное – для тоарских и верхнеюрских Meleagrinella s.str.; тупоугольное заднее ушко характерно для тоарских Clathrolima, среднеюрских Arctotis s.str.
е) Форма заднего крыла. Прямое заднее крыло левой створки характерно для Praemeleagrinella и среднеюрских Clathrolima; выгнутое заднее крыло левой створки характерно для тоарских Praemeleagrinella, Clathrolima и Praearctotis. Нижнее окончание заднего крыла левой створки у Clathrolima и Praearctotis расположено выше заднего конца створки, у Meleagrinella s.str. и Arctotis s.str. – ниже.
К признакам с малым весом отнесены признаки, подверженные возрастной или незакономерной изменчивости: размер, контур, симметрия, выпуклость, плотность ребристости. К этой же категории отнесена ширина межреберных промежутков, которую трудно формализовать ввиду нечеткости границ между ребрами и межреберными промежутками.
ОПИСАНИЕ ТАКСОНОВ
СЕМЕЙСТВО OXYTOMIDAE ICHIKAWA, 1958
ПОДСЕМЕЙСТВО OXYTOMINAE ICHIKAWA, 1958
Род Meleagrinella Whitfield, 1885
Подрод Praemeleagrinella Lutikov et Schurygin, 2010
Meleagrinella (Praemeleagrinella ?) golberti Lutikov et Arp sp. nov.
Табл. I , фиг. 1–12
Рис. 6.
Реконструкция филогенеза родов Meleagrinella и Arctotis. Широкими вертикальными линиями обозначен стратиграфический диапазон подродов, штриховыми линиями показаны родственные связи между подродами, штрихпунктирной линией – между родами.
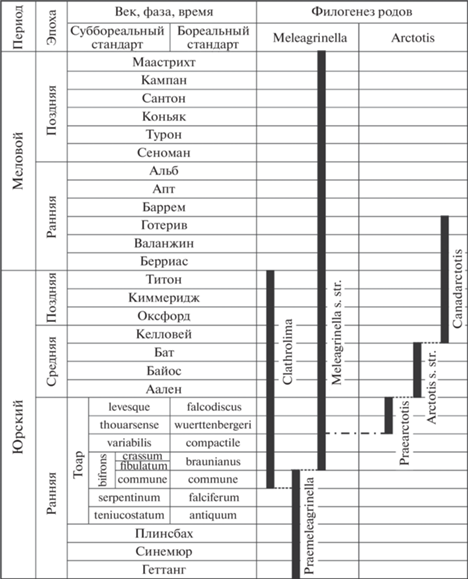
Pseudomonotis substriata: Бодылевский, Шульгина, 1958, с. 48–49.
Meleagrinella substriata: Caswell et al., 2009, text-fig. 5C (здесь табл. I , фиг. 12 ); Teichert, Nützel, 2015, text-fig. 4B (здесь табл. I , фиг. 4); Arp, Gropengiesser, 2016, text-fig. 2f (здесь табл. I , фиг. 1), 2g; Morris et al., 2019, text-fig. 9.6H, I.
Таблица I . Фиг. 1–13 . Meleagrinella (Praemeleagrinella ?) golberti Lutikov et Arp sp. nov. Фиг. 1. Голотип, экз. № GZG.INV.70490 (из Arp, Gropengiesser, 2016, text-fig. 2f): а – левая створка, ×1, б – то же ×2; Дёрльбах, Канал Людвига (Dorlbach Ludwigskanal), зона Harpoceras serpentinum (подзона Cleviceras elegans). Фиг. 2. Экз. № GZG.INV.70640, а – левая створка, ×1, б – то же, ×4; местонахождение то же. Фиг. 3. Экз. № GZG.INV.70641, а – левая створка, ×1, б – то же, ×3; местонахождение то же. Фиг. 4. Экз. № BSPG 2008 XXIX 1d (из Teichert, Nutzel, 2015, text-fig. 4В), а – левая створка, ×1, б – то же, ×3; Альтдорф, Германия, нижний тоар, зона Harpoceras serpentinum (подзона Cleviceras elegans). Фиг. 5. Экз. № GZG.INV.70650, а – правая створка, ×1, б – то же, ×3; Берг, Германия, нижний тоар, зона Harpoceras serpentinum (подзона Cleviceras elegans). Фиг. 6. Экз. № GZV.INV.70649, а – левая створка, ×1, б – то же, ×2.5; Аденштедт, Германия, зона Harpoceras serpentinum (подзона Cleviceras exaratum). Фиг. 7 . Экз. № К-1098/1, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×2.5, в – вид сбоку со стороны переднего края, × 2.5; р. Келимяр, обн. 16, сл. 3, уровень 0.7–0.8 м, обр. № 1098, келимярская свита, курунгская подсвита, нижний тоар, зона Tiltoniceras antiquum. Фиг. 8 . Экз. № К-1097/2, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×5; р. Келимяр, обн. 16, сл. 3, уровень 1.0–1.2 м; келимярская свита, курунгская подсвита, нижний тоар, зона Harpoceras falciferum, подзоны Harpoceras (=Cleviceras) exaratum–Harpoceras falciferum. Фиг. 9 . Экз. № К-1069/1, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×5; р. Келимяр, обн. 11б, келимярская свита, курунгская подсвита, нижний тоар. Фиг. 10 . Экз. № АС-72/1, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×5; р. Астрономическая, обн. 2, сл. 14, обр. № 72, основание, стартинская свита, нижняя подсвита, нижний тоар, зона Harpoceras falciferum, подзона Harpoceras (=Cleviceras) exaratum. Фиг. 11 . Экз. № МТ-213/3, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×5; р. Моторчуна, обн. 3, сл. 2, обр. № 213, уровень 4.0 м, сунтарская свита, нижний тоар. Фиг. 12 . Экз. № BM PI MB 991 (из Caswell et al., 2009, text-fig. 5C), а – левая створка, ×1, б – то же, ×2; Port Mulgrave, Англия, нижний тоар, зона Harpoceras serpentinum (подзона Cleviceras exaratum). Фиг. 13 . Экз. № L2430, TMP 2015.051.0096, а – слепок левой створки, ×1, б – то же, ×2 (из Martindale, Aberhan, 2017, text-fig. 4O); Альберта, Западная Канада, нижний тоар, подзона Harpoceras serpentinum. Фиг. 14, 15 . Meleagrinella (Praemeleagrinella ?) aff. golberti Lutikov et Arp sp. nov. Фиг. 14 . Экз. № БР-152/2, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×3; р. Бродная, обн. 3, сл. 18, уровень 6.5 м, обр. № 152, стартинская свита, нижняя подсвита, нижний тоар, зона Harpoceras falciferum (подзона Harpoceras falciferum). Фиг. 15 . Экз. № МТ-213/4, а – ядро левой створки с остатками раковинного слоя, ×1, б – то же, ×3, в – вид сбоку со стороны переднего края, ×3; Моторчуна, обн. 3, сл. 2, обр. № 213, уровень 4.0 м, нижний тоар, сунтарская свита, зона Harpoceras falciferum (?).
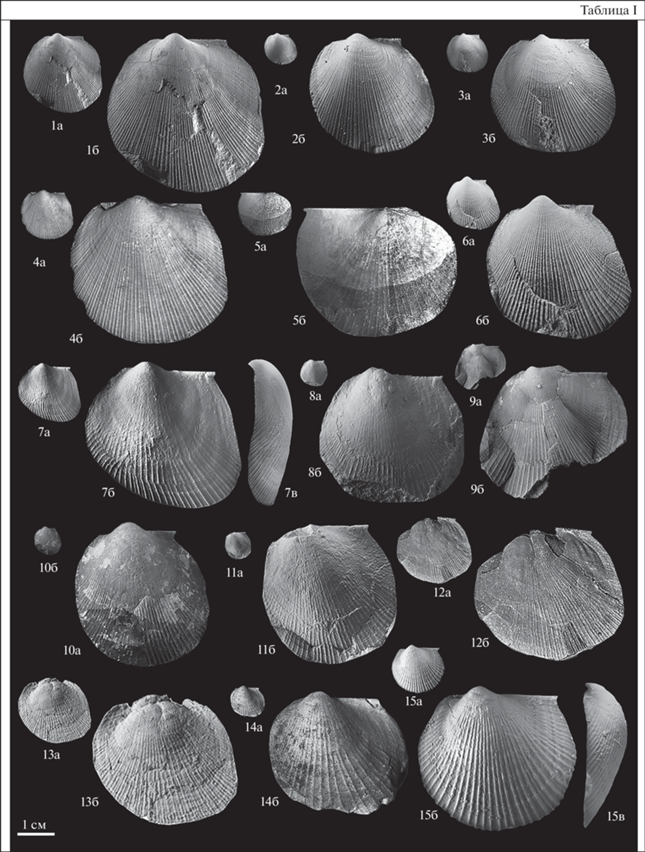
Таблица II. Фиг. 1–11 . Meleagrinella (Clathrolima) substriata (Münster, 1831). Фиг. 1. Синтип (Goldfuss, 1835, p. 138, tab. CXX , figs. 7 а, 7b), а – левая створка, ×1, б – то же, ×3; формация Posidonienschiefer, Банц, Германия. Фиг. 2. Лектотип, экз. № IGPB-Goldfuss-729а/1, а – левая створка, ×1, б – то же, ×3; местонахождение то же. Фиг. 3. Синтип, экз. № IGPB-Goldfuss-729a/3, ядро левой створки, вид заднего ушка и заднего крыла, ×7; местонахождение то же. Фиг. 4. Синтип, экз. № AS-VII-2087 a, a – левая створка, ×1, б – вид лигаментной площадки, ×12; Мистельгау, Германия. Фиг. 5. Синтип, экз. № AS-VII-2087 b, a – правая створка, ×1, б – то же, ×2.5; местонахождение то же. Фиг. 6. Экз. № GZG.INV.70646, а – левая створка, ×1, б – то же, ×4; Дёрльбах, Канал Людвига (Ludwigskanal), Германия, формация Posidonienschiefer, пачка 14 (“Monotis-Bank”), зона Hildoceras bifrons (подзона Dactylioceras commune). Фиг. 7 . Экз. № GZG.INV.70644, а – левая створка, ×1, б – то же, 3; местонахождение то же. Фиг. 8 . Экз. № GZG.INV.70643, а – левая створка, ×1, б – то же, ×3; местонахождение то же. Фиг. 9 . Экз. № АС-77/1, а – левая створка, ×1, б – то же, ×4, в – то же, вид сбоку, ×4; р. Астрономическая, обн. 2, сл. 17; обр. № АС-77, стартинская свита, верхняя подсвита, нижний тоар, зона Dactylioceras commune. Фиг. 10 . Экз. № Т-254/6, а – ядро левой створки, ×1, б – то же, ×3; р. Тюнг, обн. 15а, обр. № 254, осыпь слоев 3–4, сунтарская свита, вторая пачка, нижний тоар. Фиг. 11 . Экз. № Т-254/17, а – ядро левой створки, ×1, б – то же, ×3; местонахождение то же. Фиг. 12 . Meleagrinella (Clathrolima) sp., экз. № П-470-521/1, а – левая створка, ×1, б – то же, ×5, в – вид лигаментной площадки, ×22; Правобережный участок, линия 470, скв. 521, глубина 21.9 м, сунтарская свита, третья пачка, нижний тоар. Фиг. 13 . Otapiria substriata (Zieten), экз. GZG.INV.70639, а – ядро левой створки, ×1, б – то же, ×5; Веллерсен, Германия, нижний синемюр.
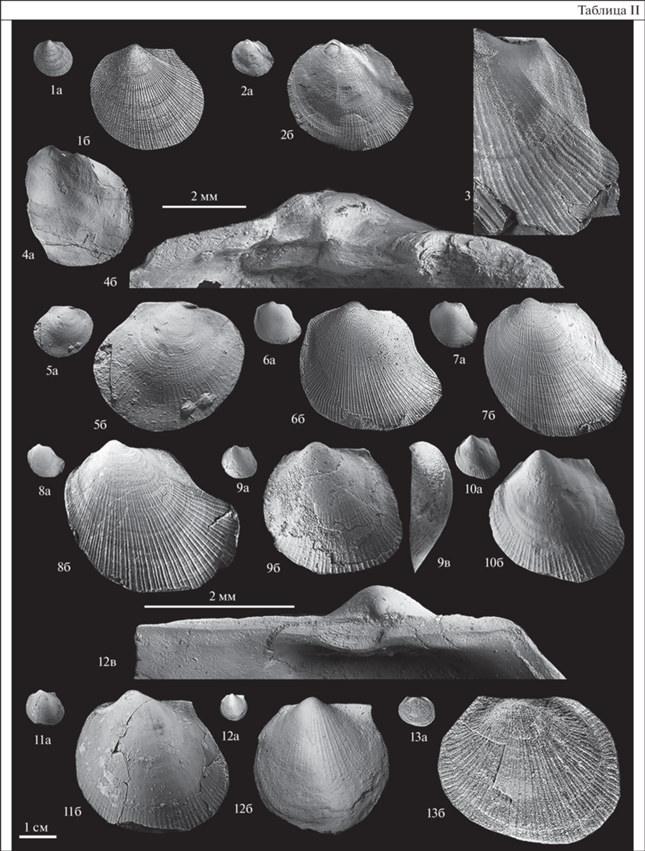
Таблица III. Фиг. 1–11 . Meleagrinella (Meleagrinella) prima Lutikov sp. nov. Фиг. 1. Голотип, экз. № МХ-237/5, а – левая створка, ×1, б – то же, ×4; Марха, обн. 10, слой 9, обр. № МХ-237-Б, сунтарская свита, третья пачка, нижний тоар, зона Zugodactylites braunianus. Фиг. 2. Экз. № МХ-237/4, а – левая створка, ×1, б – то же, ×4; местонахождение то же. Фиг. 3. Экз. № МХ-237/3, а – левая створка, ×1, б – то же, ×4; местонахождение то же. Фиг. 4. Экз. № МХ-309/17, а – ядро левой створки с внешним отпечатком правой створки, ×1, б – то же, × 4; Марха, обн. 8, слой 7, обр. № МХ-309, нижний тоар, зона Zugodactylites braunianus. Фиг. 5. Экз. № МХ-229/1, a – левая створка, ×1, б – то же, ×3, в – вид заднего крыла левой створки, ×7; Марха, обн. 6, слой 3, обр. № МХ-229, сунтарская свита, нижний тоар, зона Zugodactylites braunianus. Фиг. 6. Экз. № АГ-103/4, а – левая створка, ×1, б – то же, ×4; Анабарская губа, западный берег, обн. 5, слой 76, обр. № АГ-103, эренская свита, нижний тоар, зона Zugodactylites braunianus. Фиг. 7 . Экз. № ТХ-1/3, а – левая створка, ×1, б – то же, ×5; в – то же, вид сбоку со стороны переднего края, ×5, нижний тоар, третья пачка сунтарской свиты, Тенкеляхский участок, линия 1060, скв. 350, глуб. 31 м. Фиг. 8 . Экз. № ТХ-1/1, а – правая створка, ×1, б – то же, ×5; местонахождение то же. Фиг. 9 . Экз. № АГ-103/1, лигаментный блок левой створки ювенильной особи (широкоугольный тип лигаментной ямки), ×40; Анабарская губа, западный берег, обн. 5, эренская свита, слой 76, обр. № АГ-103, нижний тоар, зона Zugodactylites braunianus. Фиг. 10 . № ТХ-1/10, лигаментный блок левой створки юношеской особи (сужающийся вид онтогенеза), ×30; Тенкеляхский участок, линия 1060, скв. 350, глуб. 31 м, обр. № ТХ-1, нижний тоар, сунтарская свита, третья пачка. Фиг. 11 . Экз. № АГ-103/2, лигаментный блок левой створки взрослой особи, ×11.0, Анабарская губа, западный берег, обн. 5, эренская свита, слой 76, нижний тоар, зона Zugodactylites braunianus. Фиг. 12, 13 . Meleagrinella (Meleagrinella) aff. prima Lutikov sp. nov. Фиг. 12 . Экз. № 24/12566, ЦНИГР музей им. акад. Ф.Г. Чернышева (Санкт-Петербург), левая створка, ×1; р. Мунугуджак, сборы Б.А. Сняткова, Ю.С. Репина, верхний тоар. Фиг. 13 . Экз. № Ст-1/1, а – ядро левой створки, б – то же, ×2.5; в – вид ядра левой створки сбоку, ×2.5; р. Старт, стартинская свита, верхняя подсвита, осыпь слоя 2, обнажение 5, зона Pseudolioceras compactile. Фиг. 14, 15 . Meleagrinella (Meleagrinella?) milovae Okuneva, 2002. Фиг. 14 . Экз. № 44/12919, ЦНИГР музей им. акад. Ф.Г. Чернышева (Санкт-Петербург), а – отпечаток левой створки, ×1; б – то же, ×4; бассейн р. Газимур, пядь Большая Кулинда, сборы Т.М. Окуневой, верхний тоар, слои с Meleagrinella milovae. Фиг. 15 . Экз. № 46/12919, ЦНИГР музей им. акад. Ф.Г. Чернышева (Санкт-Петербург), ракушняковое скопление ядер левых и правых створок; местонахождение то же.
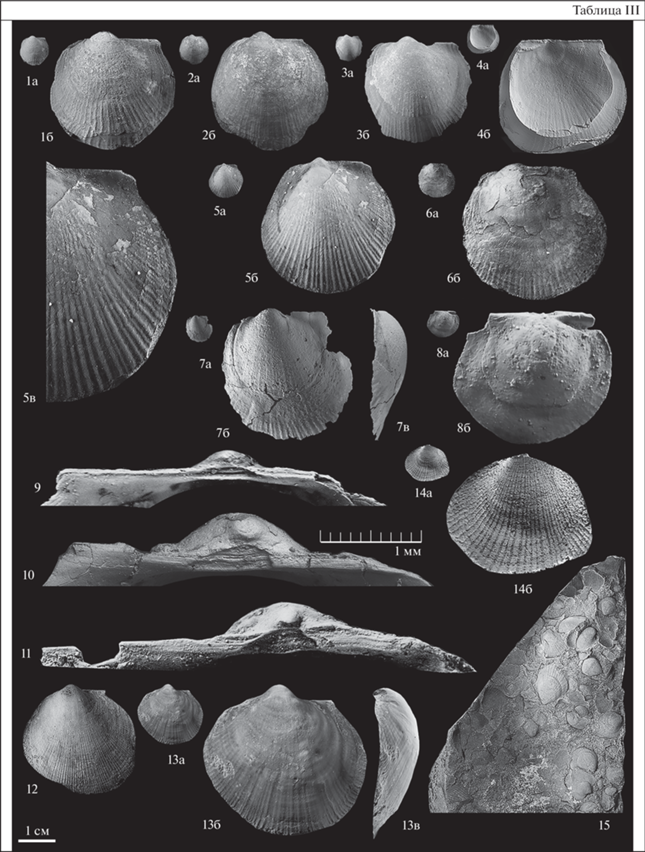
Таблица IV. Фиг. 1–3. Meleagrinella (Praemeleagrinella) deleta (Dumortier, 1869). Фиг. 1. Синтип, а – левая створка, ×1, б – то же, ×5 (Dumortier, 1869, pl. XXХV, fig. 5), верхний плинсбах, Живерди, Бургундия, Франция. Фиг. 2. Экз. № Пр-21п/23, а – левая створка, ×1, б – то же, ×5, в – лигаментная площадка, ×20; Правобрежная площадь, линия 524, скв. 546.5, глуб. 18.6 м, тюнгская свита, верхний плинсбах. Фиг. 3. Экз. № Пр-21п/26, а – левая створка, ×1, б – то же, ×5, в – лигаментная площадка, ×20; местонахождение то же. Фиг. 4, 5. Arctotis (Praearctotis) marchaensis (Petrova, 1947).Фиг. 4. Экз. № ТХ-1/5, а – левая створка, ×1, б – то же, × 2; Тенкеляхский участок, линия 1080, скв. 350, гл. 35 м, обр. 1, сунтарская свита, четвертая пачка, верхний тоар. Фиг. 5. Экз. № ТХ-1/1, а – правая створка, ×1, б – то же, ×3, в – часть поверхности призматического слоя, длина масштабного отрезка 0.5 мм; местонахождение то же. Фиг. 6. Tiltoniceras sp. ind., экз. № К-1097/А, а – сдавленный экземпляр, ×1, б – то же, ×4; р. Келимяр, обн. 16, сл. 3, уровень 1.0 м, обр. № 1097, келимярская свита, курунгская подсвита, нижний тоар, зона Tiltoniceras antiquum. Фиг. 7 . Meleagrinella (Meleagrinella) prima Lutikov sp. nov., экз. № МХ-237/10, часть поверхности призматического слоя правой створки, длина масштабного отрезка 0.2 мм; р. Марха, обн. 10, слой 9, обр. 237-Б, сунтарская свита, третья пачка, нижний тоар.

Meleagrinella (Praemeleagrinella) aff. substriata: Лутиков, Шурыгин, 2010, табл. III, фиг. 10 (здесь табл. I , фиг. 7 ).
Meleagrinella sp.: Martindale, Aberhan, 2017, text-fig. 4O (здесь табл. I , фиг. 13 ), 4P, 4Q, 4R; Muscente et al., 2019, text-fig. 3H.
Meleagrinella (Praemeleagrinella) sp. 1: Лутиков, Арп, 2020, табл. I , фиг. 1–4.
Arctotis (Praearctotis) sp. 1: Лутиков, Арп, 2020, табл. I , фиг. 5–8 .
Название. Название дано в память о российском исследователе мезозойских отложений, геологе Аркадии Викторовиче Гольберте.
Голотип: Геологический музей Гёттингенского университета, экз. № GZG.INV.70490, левая створка. Нижний тоар, зона Harpoceras serpentinum (подзона Cleviceras elegans), Дёрльбах, Германия, Канал Людвига, пачка “Laibstein II”. Изображен: табл. I , фиг. 1.
Материал. Около 20 левых и правых створок из нижнего тоара разреза Канала Людвига в местности Дёрльбах (Южная Германия), фотографии образцов из нижнего тоара разреза местности Альтдорф (Южная Германия), одна левая створка из строительного котлована в местности Аденштедт (Северная Германия), одна правая створка из местности Берг (Южная Германия). Около 15 ядер левых и правых створок из конкреций курунгской пачки келимярской свиты разреза р. Келимяр, более 20 деформированных ядер левых и правых створок с остатками раковинного слоя из глин курунгской пачки келимярской свиты р. Келимяр; 1 деформированное ядро левой створки из разреза нижнего тоара р. Моторчуна, около 10 ядер и отпечатков левых и правых створок из конкреций стартинской свиты разреза нижнего тоара рр. Сатурн и Астрономическая. Внутреннее ядро левой створки из скважины 3-р Усть-Енисейского района из коллекции В.И. Бодылевского, хранящееся в Палеонтолого-стратиграфическом музее Горного университета (Санкт-Петербург).
Диагноз. Раковина низкая, умеренно неравносторонняя. Заднее крыло широкое, умеренно-выгнутое. Нижнее окончание заднего крыла левой створки расположено выше заднего конца створки. Заднее ушко обеих створок остроугольное.
Описание. Раковина маленькая, до 22 мм в высоту. Левая створка низкая, неравносторонняя, умеренно-выпуклая. Заднее крыло широкое, умеренно-выгнутое. Заднее залигаментное поле левой створки короткое. Заднее ушко остроугольное или субпрямоугольное. Левая створка густоребристая. Участок между выступами переднего и заднего концов створки покрыт радиальными ребрами двух порядков (от 42 до 60), незакономерно распределенными по створке, с различающимися по ширине межреберными промежутками. Ребра второго порядка незакономерно вклиниваются в межреберные промежутки. На заднем крыле количество радиальных ребер до 24. На переднем крыле их до 10. На ядрах вблизи переднего и заднего краев ребра слабо выражены. Межреберные промежутки в разных частях левой створки различаются по ширине от умеренно широких до широких, редко превышают ширину ребер более чем в 2 раза.
Правая створка очень низкая, умеренно неравностороняя. Заднее залигаментное поле правой створки очень длинное. Заднее крыло правой створки слабовыгнутое. Заднее ушко правой створки остроугольное. Радиальная скульптура слабовыраженная. Межреберные промежутки по нижнему краю очень широкие.
Размеры, мм и отношения. См. табл. 2.
Таблица 2.
Размеры в мм и отношения
| Колл. №, створка |
Местонахождение | Д | В | В/Д | Вп | Вп/В | ДПЧ | ДПЧ/Д | ДЗЧ | ДЗЧ/В | ДЗП | ДЗП/ДЗЧ | УЗКр | Р | Р/Д | ШП/ШР | УЗУ | ШЗКр/ДЗКр |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Meleagrinella (Praemeleagrinella ?) golberti | ||||||||||||||||||
| GZG.INV.70490, голотип, левая | Дёрльбах, Германия, Ludwigskanal, пачка “Laibstein II”, зона Harpoceras serpentinum (подзона Cleviceras elegans) | 21.78 | 21.5 | 0.99 | – | – | 8.99 | 0.41 | 12.79 | 0.59 | 7.01 | 0.55 | 46° | 66 | 3.03 | 2.88 | 96° | 0.16 |
| GZG.INV.70640, левая | То же | 8.81 | 8.57 | 0.97 | – | – | 3.48 | 0.40 | 3 | 0.62 | 3.15 | 0.59 | 45° | 54? | 6.35 | 2.66 | 88° | 0.10 |
| GZG.INV.70641, левая | То же | 11.67 | 10.6 | 0.97 | – | – | 4.45 | 0.38 | 7.22 | 0.68 | 3.91 | 0.54 | 42° | 61 | 5.23 | 2.86 | – | 0.13 |
| GZG.INV.70642, левая | То же | 12.83 | 13.0 | 1.01 | – | – | 4.4 | 0.34 | 8.43 | 0.65 | 4.95 | 0.59 | 44° | 83 | 6.47 | 2.05 | 81° | 0.14 |
| GZG.INV.70650, правая | Берг, Германия, пачка “Laibstein II”, зона Harpoceras serpentinum (подзона Cleviceras elegans) | 14.44 | 11.69 | 0.81 | – | – | 6.36 | 0.44 | 8.08 | 0.69 | 7.50 | 0.93 | 42° | 28 | 1.92 | 4.17 | 24° | 0.06 |
| BSPG 2008 XXIX 1d, левая |
Альтдорф, Германия, зона Harpoceras serpentinum (подзона Cleviceras elegans), | 14.2 | 13.33 | 0.94 | – | – | 6.14 | 0.43 | 8.06 | 0.60 | 5.71 | 0.71 | 48° | 67 | 4.72 | 2.23 | 93° | 0.10 |
| GZV.INV.70649 левая |
Аденштедт, Германия, зона Harpoceras serpentinum (подзона Cleviceras exaratum) | 14.44 | 14.34 | 0.99 | – | – | 5.58 | 0.38 | 8.86 | 0.62 | 5.12 | 0.58 | 43° | 52 | 3.60 | 3.41 | 75° | 0.09 |
| К-1098/1, левая |
р. Келимяр, келимярская свита, курунгская пачка, обн. 16, сл. 3, уровень 0.7–0.8 м, обр. №1098, зона Tiltoniceras antiquum | 17.45 | 15.8 | 0.91 | 4.74 | 0.30 | 6.28 | 0.36 | 11.17 | 0.71 | 7.75 | 0.76 | 42° | 48 | 2.75 | 2.47 | 62° | 0.06 |
| К-1097/1, левая |
р. Келимяр, обн. 16, сл. 3, уровень 0.1–1.2 м, обр. №1097, зона Harpoceras falciferum (подзона Cleviceras exaratum) | 10.96 | 8.8 | 0.81 | – | – | 3.88? | 0.35 | 7.08? | 0.80? | 5.25 | 0.74? | 43° | 35? | 3.19 | 2.8 | 69° | – |
| К-1097/2, левая |
То же | 7.67 | 7.2 | 0.94 | 2.19 | 0.28 | 3.25 | 0.42 | 4.42 | 0.61 | 3.32 | 0.75 | 45° | 47 | 6.40 | 2.92 | 61° | 0.10 |
| К-1069/1, левая |
р. Келимяр, келимярская свита, курунгская пачка, обн. 11б | 13.83 | 12.4 | 0.90 | – | – | – | – | – | – | – | – | – | 50? | 3.61 | 3.50 | 69° | – |
| МТ-213/3, левая |
р. Моторчуна, обн. 3, сл. 2, обр. 213, сунтарская свита |
6.98? | 7.3 | 1.02? | – | – | 2.46 | 0.35 | 4.52 | 0.62 | 3.17 | 0.70? | 42° | 42 | 4.63 | 2.76 | 67° | 0.10 |
| АС-72/1 левая |
р. Астрономическая, обн. 2, сл. 14, основание, обр. 72, зона Harpoceras falciferum (подзона Harpoceras falciferum) | 7.40 | 7.8 | 1.05 | – | – | 3.00 | 0.41 | 4.40 | 0.56 | 2.51 | 0.57 | 48° | 52 | 7.01 | 2.08 | 68° | 0.12 |
| Meleagrinella (Praemeleagrinella ?) aff. Golberti | ||||||||||||||||||
| БР-152/2 левая |
р. Бродная, обн. 3, сл. 18, уровень 6.5 м, обр. № 152, зона Harpoceras falciferum (подзона Harpoceras falciferum) | 8.21 | 8.3 | 1.01 | – | – | 3.51 | 0.43 | 4.7 | 0.57 | 3.58 | 0.76 | 46° | 20 | 2.44 | 5.69 | 96° | 0.16 |
| МТ-213/4, левая |
р. Моторчуна, обн. 3, сл. 2, обр. 213, уровень 4.0 м, сунтарская свита | 14.16 | 13.1 | 0.92 | 3.40 | 0.30 | 5.41 | 0.38 | 8.75 | 0.67 | 7.10 | 0.81 | 34° | 32 | 2.26 | 3.22 | 74° | 0.03 |
Сравнение. От Meleagrinella (Praemeleagrinella) deleta (Dumortier, 1869) из верхнего плинсбаха Франции и Восточной Сибири (табл. IV, фиг. 1–3) отличается большими размерами, низким контуром левой створки, умеренно-выгнутым задним крылом, незакономерной дифференциацией ребер на два порядка, густоребристой левой створкой.
По контуру левой створки, симметрии раковины и форме заднего крыла левой створки наиболее близок к Meleagrinella oxytomaeformis Polub. из верхнего плинсбаха Северо-Востока России (Полевой…, 1968, табл. 22 , фиг. 1–5). Новый вид отличается незакономерной дифференциацией ребер на два порядка.
В Восточной Сибири к новому виду относятся экземпляры из курунгской пачки келимярской свиты разреза р. Келимяр (табл. I , фиг. 7–9 ). Вид распространен в низах сунтарской свиты в разрезах р. Моторчуна (табл. I , фиг. 11 ), р. Тюнг, в бассейне р. Ыгыатта (Оттурский участок бурения).
На Северо-Востоке России к новому виду относятся экземпляры из ракушняков нижней подсвиты стартинской свиты (подзона Harpoceras exaratum) в разрезах рр. Астрономическая (табл. I , фиг. 10 ), Сатурн, Бродная.
В Западной Сибири в китербютской свите распространены двустворки, которые В.И. Бодылевский относил к Pseudomonotis substriata (Бодылевский, Шульгина, 1958, с. 48–49), а позднее они были отнесены к Praearctotis milovae (Лутиков, Шурыгин, 2010). Образец № 166/238 из скважины 3-р Усть-Енисейского района с глубины 1029.1–1030.7 м, хранящийся в Палеонтолого-стратиграфическом музее Горного университета (Санкт-Петербург), был переизучен О.А. Лутиковым в 2020 г. с использованием новой системы градации признаков. Левая створка характеризуется очень маленькими раковинами (до 6 мм), густой скульптурой всей поверхности (до 60 ребер, включая переднее и заднее крыло), выгнутым задним крылом. Нижнее окончание заднего крыла левой створки расположено выше заднего конца створки. По этим признакам экземпляр относится к новому виду.
В Германии к M. (P.) golberti sp. nov. принадлежат экземпляры из местности Дёрльбах (Канал Людвига, пачка “Laibstein II”, зона Harpoceras serpentinum, подзона Cleviceras elegans) (Arp, Gropengiesser, 2016, text-fig. 2f; здесь табл. I , фиг. 13 ), относившиеся ранее к “Meleagrinella substriata”, а также экземпляры из местности Альтдорф (зона Harpoceras serpentinum, подзона Cleviceras elegans) (Teichert, Nützel, 2015, text-fig. 4B; здесь табл. I , фиг. 4), экземпляр из местности Берг (зона Harpoceras serpentinum, подзона Cleviceras elegans) (табл. I , фиг. 5) и экземпляр из строительного котлована в местности Аденштедт, встреченный в одном образце с Hildaites murleyi (Moxon) (здесь табл. I , фиг. 6).
В Англии к новому виду относятся экземпляры из Йоркшира (Порт Малгрейв, формация Whitby Mudstone, зона Harpoceras serpentinum, подзона Cleviceras exaratum) (Caswell et al., 2009, text-fig. 5C, здесь табл. I , фиг. 12 ; Morris et al., 2019, text-figs. 9.6 H, I).
В Западной Канаде к новому виду принадлежит “Meleagrinella sp.” из местности Альберта (р. Скальп Крик, формация Fernie, подзона Harpoceras serpentinum) (Martindale, Aberhan, 2017, text-figs. 4O (здесь табл. I , фиг. 13 ), 4P, 4Q, 4R; Muscente et al., 2019, text-fig. 3H).
Замечания. Строение замка неизвестно. Принадлежность к подроду Praemeleagrinella предположительна.
На Северо-Востоке России (р. Бродная) в подзоне Harpoceras falciferum встречены Meleagrinella (Praemeleagrinella ?) aff. golberti sp. nov., которые характеризуются более широкими межреберными промежутками (табл. I , фиг. 14 ). В Восточной Сибири подобные мелеагринеллы обнаружены в ракушняках в нижней части сунтарской свиты на р. Моторчуна (табл. I , фиг. 15 ), где вместе с ними встречены несомненные M. (P.?) golberti.
Фациальная приуроченность и тафономическая характеристика. На р. Келимяр ядра отдельных створок с остатками раковинного слоя найдены в карбонатных конкрециях и в сланцеватых тонкоплитчатых глинах курунгской пачки келимярской свиты. Мелеагринеллы образуют ракушняковые скопления в глинах совместно с аммонитами и кедонеллами. Ядра отдельных створок встречаются в известковых конкрециях совместно с устрицами вблизи основания курунгской пачки. Тип ископаемого ценоза – слабо перемещенный танатоценоз.
На р. Моторчуна отдельные створки встречаются в конкрециях совместно с кедонеллами. Тип ископаемого ценоза – слабо перемещенный танатоценоз.
На р. Астрономическая в алевритах нижней подсвиты стартинской свиты целые отдельные створки образуют гнездовидные ракушняковые скопления и захоронены совместно с аммонитами, кедонеллами, лимами и энтолиумами. Тип ископаемого ценоза – слабо перемещенный танатоценоз.
Образ жизни и условия обитания. Представители рассматриваемого вида могли вести псевдопланктонный образ жизни, прикрепляясь к плавающим водорослям или раковинам живых аммонитов. Эту точку зрения поддерживают некоторые исследователи тоарских битуминозных сланцев Германии (Röhl, 1998; Teichert, Nützel, 2015). Их относили к фильтраторам, устойчивым к условиям с пониженным содержанием кислорода (Caswell et al., 2009). В случае отрыва от места прикрепления и падения на дно, не имея возможности продолжительное время существовать в беcкислородных условиях на грунте, они, вероятно, могли перемещаться к месту повторного прикрепления за счет схлопывания створок. Псевдопланктонный образ жизни с возможностью активного перемещения был необходимой адаптацией мелеагринелл к условиям дефицита кислорода в раннетоарских морях.
Геологический возраст и географическое распространение. Нижнетоарский подъярус, зоны Tiltoniceras antiquum, Harpoceras falciferum бореальной шкалы; зоны Dactylioceras tenuicostatum, Harpoceras serpentinum суббореальной шкалы.
Западная Сибирь: Усть-Енисейский район, китербютская свита. Восточная Сибирь: р. Келимяр, келимярская свита, нижний тоар; Анабарская губа, р. Анабар, китербютская свита; р. Моторчуна, сунтарская свита; р. Тюнг, Оттурский участок (бассейн р. Ыгыатта), сунтарская свита. Северо-Восток России: рр. Сатурн, Астрономическая, Бродная, стартинская свита (нижняя подсвита). Германия: Дёрльбах, Берг, Альтдорф, Аденштедт, формация Posidonienschiefer. Англия: Йоркшир, формация Whitby Mudstone. Западная Канада: Альберта, формация Fernie.
Подрод Clathrolima (Cossman, 1908)
Meleagrinella (Clathrolima) substriata (Münster, 1831)
Табл. II, фиг. 1–11
Monotis substriata: Münster, 1831, p. 406.
Monotis substriata: Goldfuss, 1835, p. 138, Table CXX , figs. 7a–7f .
Arctotis (Praearctotis) substriata: Лутиков, Арп, 2020, табл. I , фиг. 9–12 .
Meleagrinella (Clathrolima) substriata: Rogov, Lutikov, 2022, Figs. 4I–4N.
Голотип вида не был установлен. Типовая серия была частично изображена в работе Г.А. Гольдфусса (Goldfuss, 1835). Авторами в качестве лектотипа выбран экземпляр из типовой серии № IGPB-Goldfuss-729 a/1. Изображен здесь – табл. II, фиг. 2, хранится в Институте наук о Земле (Германия, Бонн). Формация Posidonienschiefer, подзона Dactylioceras commune, местонахождение Банц, Германия.
Материал. Более 20 ядер и отдельных створок из тоарских отложений разрезов Анабарской губы, рек Астрономическая, Сатурн, Тюнг, Вилюй. 23 отпечатка и отдельных створок, около 150 фрагментов из тоарских отложений Канала Людвига (Дёрльбах) и местности Берг, Германия. Фотографии синтипов из типовой коллекции Г. Мюнстера, происходящей из формации Posidonienschiefer, подзоны Dactylioceras commune (Банц, Мистельгау, Германия). 1 экземпляр с сохранившейся раковиной, 6 ядер и отпечатков створок из тоарских отлоложений разреза бухты Агард, Шпицберген.
Диагноз. Раковина очень низкая, сильно неравносторонняя. Заднее крыло узкое, умеренно-выгнутое. Задний край образует S-образный изгиб. Заднее ушко обеих створок тупоугольное, без выреза.
Описание. Раковина маленькая, преимущественно до 15 мм в высоту, один синтип имеет высоту до 33 мм. Левая створка очень низкая, неравносторонняя, сильновыпуклая. Заднее крыло левой створки узкое, умеренно-выгнутое. Задний край образует S-образный изгиб. Нижнее окончание заднего крыла левой створки расположено выше заднего конца створки. Заднее залигаментное поле левой створки очень короткое. Заднее ушко тупоугольное, без выреза. Левая створка густоребристая. Участок между концами переднего и заднего краев покрыт радиальными ребрами одного порядка в количестве от 35 до 60, незакономерно распределенными по створке, с различающимися по ширине межреберными промежутками. Ребра второго порядка незакономерно вклиниваются в межреберные промежутки. На заднем крыле количество радиальных ребер более 20. На переднем крыле количество радиальных ребер более 10. На ядре левой створки в области макушки, вблизи переднего и заднего краев ребра слабо выражены. Межреберные промежутки по нижнему краю умеренно широкие.
Правая створка низкая. Заднее залигаментное поле правой створки очень длинное. Заднее ушко правой створки остроугольное. Заднее крыло узкое, умеренно-выгнутое в направлении заднего конца створки. Правая створка умеренно-ребристая. Межреберные промежутки по нижнему краю умеренно широкие.
Размеры, мм и отношения. См. табл. 3.
Таблица 3.
Размеры, мм и отношения
| Колл. №, створка | Местонахождение | Д | В | В/Д | Вп | Вп/В | ДПЧ | ДПЧ/Д | ДЗЧ | ДЗЧ/В | ДЗП | ДЗП/ДЗЧ | УЗКр | Р | Р/Д | ШП/ШР | УЗУ | ШЗКр/ДЗКр | ДНЛ/ДЗЛ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Meleagrinella (Clathrolima) substriata | |||||||||||||||||||
| IGPB-Goldfuss-729 a/1, лектотип, левая | Банц, Германия, формация Posidonienschiefer | 11.5 | 10.09 | 0.88 | – | – | 4.31 | 0.37 | 7.19 | 0.71 | 3.28 | 0.46 | 34° | 60 | 5.22 | 1.76 | 134° | 0.10 | – |
| № AS-VII-2087a синтип, правая |
Мистельгау, Германия, формация Posidonienschiefer |
16.26 | 14.03 | 0.86 | – | – | 7.24 | 0.45 | 9.02 | 0.64 | 4.11 | 0.46 | 35° | 31 | 1.91 | 2.20 | 131° | 0.11 | – |
| № AS-VII-2087a синтип, левая |
То же | – | 32.84 | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 0.90 |
| GZG. INV.70643, левая | Канал Людвига, Дёрльбах, Германия,
формация Posidonienschiefer, пачка 14, зона Hildoceras bifrons, подзона Dactylioceras commune |
13.30 | 12.0 | 0.90 | – | – | 4.95 | 0.37 | 8.35 | 0.70 | 3.31 | 0.40 | 34° | 46 | 3.46 | 3.22 | 143° | 0.12 | – |
| GZG. INV.70644, левая | То же | 12.33 | 12.0 | 0.97 | – | – | 4.00 | 0.32 | 8.33 | 0.69 | 4.04 | 0.48 | 32° | 65 | 5.27 | 3.5 | 144° | 0.09 | – |
| GZG.INV.70646, левая | То же | 9.96 | 9.0 | 0.90 | – | – | 3.15 | 0.32 | 6.81 | 0.76 | 2.49 | 0.37 | 25° | 65 | 6.53 | 2.13 | – | 0.18 | – |
| АК-77/1, левая | р. Астрономическая обн. 2, сл. 17; зона Dactylioceras commune, стартинская свита, верхняя подсвита | 9.49 | 9.06 | 0.95 | 2.96 | 0.33 | 3.47 | 0.36 | 5.93 | 0.65 | 3.37 | 0.57 | 39° | 35? | 3.69 | 1.91 | 114° | 0.09 | – |
| Т-254/6, левая | р. Тюнг, обн. 15а, осыпь слоев 3–4, обр. 254, зона Dactylioceras commune, сунтарская свита, вторая пачка | 11.77 | 12.1 | 1.03 | – | – | 4.34 | 0.37 | 7.43 | 0.61 | 4.28 | 0.57 | 40° | 48 | 4.08 | 2.31 | 100° | 0.06 | – |
| Т-254/17, левая | То же | 10.38 | 9.8 | 0.89 | – | – | 3.97 | 0.38 | 6.61 | 0.65 | 3.94 | 0.60 | 35° | 58 | 5.59 | 1.87 | 112° | 0.07 | – |
| Т-254/30, левая | То же | 10.98 | 11.1 | 1.01 | – | – | 4.21 | 0.38 | 6.77 | 0.61 | 4.13 | 0.61 | 38° | 50 | 4.55 | 2.35 | – | 0.08 | – |
| Meleagrinella (Clathrolima) sp. | |||||||||||||||||||
| П-470-521/1, левая | Правобережный участок, лин. 470, скв. 521, гл. 21.9 м, сунтарская свита, третья пачка | 7.40 | 7.53 | 1.02 | – | – | 3.24 | 0.44 | 4.16 | 0.55 | 3.26 | 0.79 | 49° | 30 | 4.05 | 1.17 | 67° | 0.12 | 0.97 |
Индивидуальная изменчивость. Контур левой створки варьирует от очень низкого до высокого. Симметрия левых створок изменяется от умеренно неравносторонней до сильно неравносторонней. Заднее залигаментное поле левой створки варьирует от очень короткого до длинного. Радиальная скульптура левой створки изменяется от часторебристой до густоребристой.
Сравнение. От Meleagrinella (Clathrolima) sp. из третьей пачки сунтарской свиты Правобережного участка (табл. II, фиг. 12 ) отличается более неравносторонними раковинами, S-образным задним краем левой створки, положением окончания заднего крыла левой створки выше заднего конца створки, тупоугольным задним ушком без выреза.
Замечания. От Meleagrinella (Praemeleagrinella?) golberti sp. nov. из зон Tiltoniceras antiquum и Harpoceras falciferum отличается более неравносторонними раковинами, S-образным задним краем левой створки, тупоугольным задним ушком без выреза.
Вид “Avicula” substriata (Zieten, 1830), происходящий из формации Arietenkalk (нижний синемюр, район Штутгарта, Южная Германия), предположительно рассматривался в составе подрода Meleagrinella (Praemeleagrinella) (Лутиков, Шурыгин, 2010). В одновозрастных отложениях формации Arietenton в Северной Германии Г. Арпом были обнаружены экземпляры, принадлежащие к этому виду (глиняный карьер в местности Веллерсен, синклиналь Маркольдендорф) (табл. II, рис. 13 ). По очертаниям переднего ушка и коленообразно изгибающимся радиальным ребрам у нижнего края синемюрский вид “Avicula” substriata принадлежит к роду Otapiria (Marwick, 1935), поэтому для тоарского вида название Meleagrinella substriata является валидным.
По фотографиям нескольких экземпляров из типовой коллекции вида Monotis substriata, хранящейся в музее им. Г. Гольдфусса (Бонн, Германия), в качестве лектотипа вида выбран экземпляр, происходящий из местности Банц (формация Posidonienschiefer, пачка Dactylioceras-Monotis-Bed, зона Hildoceras bifrons, подзона Dactylioceras commune) (табл. II, фиг. 2). Экземпляр, изображенный на рисунках в монографии Г. Гольдфусса (Goldfuss, 1835, p. 138, tab. CXX , figs. 7a, 7b ; здесь табл. II, фиг. 1), очень похож на лектотип. Синтипы, изображенные на других рисунках в монографии Г. Гольдфусса (Goldfuss, 1835, p. 138, tab. CXX , figs. 7c, 7e , здесь табл. II, фиг. 4, 5), происходят из местности Мистельгау (формации Posidonienschiefer, Lias epsilon) и хранятся в Музее Bayerische Staatssammlung für Paläontologie und Geologie (Мюнхен, Германия) под № AS-VII-2087. Лигаментная ямка левой створки синтипа № AS-VII-2087а имеет остроугольные очертания (ДНЛ/ДЗЛ = = 0.90) (табл. II, фиг. 4б). По этому признаку вид Г. Мюнстера относится к подроду Meleagrinella (Clathrolima). Правая створка синтипа № AS-VII-2087б имеет тупоугольную форму заднего ушка (табл. II, фиг. 5).
Экземпляры, происходящие из пачки Dactylioceras-Monotis-Bed разреза Канала Людвига в местности Дёрльбах (табл. II, фиг. 6–8 ) и из зоны Dactylioceras commune разрезов р. Астрономическая (табл. II, фиг. 9 ) и р. Тюнг (табл. II, фиг. 10–11 ), имеют тупоугольные очертания заднего ушка левой створки, S-образный изгиб заднего края, очень короткое заднее залигаментное поле. По этим признакам они относятся к виду Meleagrinella (Clathrolima) substriata.
Фациальная приуроченность и тафономическая характеристика. На р. Тюнг экземпляры представлены целыми раковинами с двумя створками в конкрециях с ракушняками. Мелеагринеллы захоронены совместно с аммонитами, дакриомиями, лиострями, окситомами, леноцерамусами, модиолусами и танкредиями. Тип ископаемого ценоза – слабо перемещенный танатоценоз. Захоронение происходило вблизи мест обитания. На рр. Астрономическая и Сатурн раковины образуют гнездовидные ракушняковые скопления в массивных конкрециях. Мелеагринеллы представлены разрозненными створками плохой сохранности, захоронены с аммонитами, белемнитами, астартами, энтолиумами, кукуллеями, редкими пропеамуссиумами, окситомами, лиостреями, гониомиями. Тип ископаемого ценоза – перемещенный танатоценоз.
Образ жизни и условия обитания. Обитали на небольших глубинах на удалении от берега (рр. Тюнг, Астрономическая, Сатурн, Канал Людвига). Селились колониями. Прикреплялись биссусом к водорослям и корягам. Могли образовывать “бентосные острова”, прикрепляясь к твердым частям раковин отмерших других организмов, в том числе к аммонитам (Schmidt-Rohl, Rohl, 2003). На приближенных к берегу участках (р. Вилюй, Анабарская губа) и на глубоководных площадях (Мыс Цветкова) поселения не образовывали.
Геологический возраст и географическое распространение. Нижнетоарский подъярус, зона Dactylioceras commune – бореальный стандарт; зона Hildoceras bifrons, подзона Dactylioceras commune – стандарт Северо-Западной Европы.
Восточная Сибирь: рр. Тюнг, Вилюй, сунтарская свита; Анабарская губа, эренская свита. Северо-Восток России: рр. Сатурн, Астрономическая, Бродная, стартинская свита (верхняя подсвита). Южная Германия: Банц, Мистельгау, Дёрльбах, формация Posidonienschiefer; Англия; Шпицберген; Иран; Япония.
Подрод Meleagrinella Whitfield, 1885
Meleagrinella (Meleagrinella) prima Lutikov sp. nov.
Табл. III, фиг. 1–12
Meleagrinella substriata: Полевой…, 1968, с. 42, табл. 39 , фиг. 1–2.
Meleagrinella faminaestriata: Князев и др., 2003, с. 90, табл. 6 , фиг. 1.
Arctotis (Praearctotis) milovae: Лутиков, Шурыгин, 2010, табл. III, фиг. 12–14 (part.); Лутиков и др., 2010, табл. I , фиг. 9 , табл. II, фиг. 2.
Arctotis (Praearctotis) substriata: Лутиков, Арп, 2020, табл. I , фиг. 15–17 (part.).
Название от лат. prima – первая.
Голотип – ГГМ им. В.И. Вернадского, г. Москва, экз. № МХ-237/5, р. Марха, обн. 10, слой 9, обр. № МХ-237-Б, нижний тоар, зона Zugodactylites braunianus.
Материал. Более 700 отдельных створок, ядер с остатками раковинного слоя из разрезов рр. Астрономическая, Сатурн, Старт, Марха, Анабарской губы, Тенкеляхского участка разведочного бурения (междуречье Хання–Тюкян).
Диагноз. Раковина округлая, умеренно неравносторонняя. Заднее крыло очень широкое, сильновыгнутое в направлении заднего конца створки. Нижнее окончание заднего крыла левой створки расположено ниже заднего конца створки. Заднее ушко обеих створок субпрямоугольное, с вырезом. Биссусное ушко трапециевидное, с коротким биссусным вырезом.
Описание. Раковина от очень маленькой до маленькой. Левая створка округлая, умеренно неравносторонняя, умеренно-выпуклая. Заднее крыло левой створки очень широкое, сильновыгнутое в направлении заднего конца створки. Нижнее окончание заднего крыла левой створки расположено ниже заднего конца створки. Заднее залигаментное поле левой створки короткое. Заднее ушко левой створки субпрямоугольное, с вырезом. Левая створка густоребристая. Участок между апикальными выступами переднего и заднего краев покрыт радиальными ребрами одного порядка в количестве от 37 до 38, незакономерно распределенными по створке, с различающимися по ширине межреберными промежутками. На заднем крыле количество радиальных ребер более 12. На переднем крыле количество радиальных ребер более 10. На ядре левой створки в области макушки, вблизи переднего и заднего краев ребра слабо выражены. На некоторых экземплярах в межреберных промежутках есть ребра второго порядка, отличающиеся по степени выраженности от ребер первого порядка. Межреберные промежутки по нижнему краю узкие. Правая створка низкая, неравносторонняя. Заднее залигаментное поле правой створки очень длинное. Заднее ушко правой створки остроугольное. Заднее крыло очень широкое, сильновыгнутое в направлении заднего конца створки. Правая створка умеренно-ребристая.
Таблица 4.
Размеры, мм и отношения
| Колл. №, створка | Местонахождение | Д | В | В/Д | ВПс/ ВЛс | Вп | Вп/В | ДПЧ | ДПЧ/Д | ДЗЧ | ДЗЧ/В | ДЗП | ДЗП/ДЗЧ | УЗКр | Р | Р/Д | ШП/ШР | УЗУ | ШЗКр/ДЗКр | ДНЛ/ДЗЛ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Meleagrinella (Meleagrinella) prima | ||||||||||||||||||||
| МХ-237/5,
голотип, левая |
р. Марха, обн. 10, слой 9, обр. № МХ-237-Б, сунтарская свита, нижний тоар, зона Zugodactylites braunianus | 7.89 | 8.1 | 1.03 | – | – | – | 3.42 | 0.43 | 4.47 | 0.55 | 3.56 | 0.78 | 56° | 37? | 4.69 | 2.00 | 90° | 0.16 | – |
| МХ-237/4, левая | То же | 8.00 | 8.1 | 1.01 | – | – | – | 3.49 | 0.44 | 4.51 | 0.56 | 3.03? | 0.67? | – | – | – | – | – | 0.24 | – |
| МХ-237/3, левая | То же | 7.38 | 7.3 | 0.99 | – | – | – | 2.96 | 0.40 | 4.42 | 0.61 | 3.51 | 0.79 | – | – | – | – | – | – | – |
| М-309/16, левая | р. Марха, обн. 8, слой 7, сунтарская свита, нижний тоар, зона Zugodactylites braunianus | 8.84 | 8.80 | 1.00 | - | 2.55 | 0.29 | 3.42 | 0.38 | 5.42 | 0.62 | 3.49 | 0.64 | 52° | 38 | 4.27 | 1.68 | - | 0.17 | – |
| М-309/17, левая правая |
То же | 8.68 7.00 |
7.80 6.72 |
0.90 0.96 |
0.86 | – – |
– – |
3.58 2.81 |
0.41 0.40 |
5.10 4.19 |
0.65 0.62 |
3.48 3.48 |
0.68 0.83 |
50° 50° |
– 19 |
– 2.71 |
– – |
– – |
0.17 0.19 |
– |
| МХ-229/19, левая | Марха, обн. 6, слой 3, обр. № МХ-229, сунтарская свита нижний тоар, зона Zugodactylites monestieri | 9.37 | 9.4 | 1.00 | – | – | – | 3.92 | 0.42 | 5.45 | 0.58 | 3.49 | 0.64 | 49° | 38 | 4.06 | 2.00 | – | 0.16 | – |
| АГ-103/4, левая | Анабарская губа, западный берег,эренская свита, обн. 5, слой 76; зона Zugodactylites braunianus | 9.93 | 9.30 | 0.94 | – | 2.67 | 0.29 | 3.66 | 0.37 | 6.22 | 0.67 | 4.04 | 0.65 | – | – | – | 1.95 | – | – | – |
| АГ-103/2, левая | То же, юношеская стадия, взрослая стадия |
– | 9.50 | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 1.451.55 |
| АГ-103/1, левая | То же ювенильная стадия |
– | 2.50 | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | 1.87 |
| ТХ-1/3, левая | Тенкеляхский участок, линия 1060, скв. 350, глуб. 31 м; нижний тоар, сунтарская свита | 7.21 | 7.20 | 0.99 | – | – | – | 2.75 | 0.38 | 4.52 | 0.63 | 3.01 | 0.67 | – | – | – | 1.58 | – | – | – |
| ТХ-1/1, правая | То же | 8.83 | 7.20 | 0.81 | – | 1.86 | 0.21 | 3.62 | 0.40 | 5.21 | 0.72 | – | – | – | – | – | – | – | – | – |
| ТХ-1/10, левая | То же, ювенильная стадия, юношеская стадия |
3.10 | 2.50 | 0.80 | – | – | – | 1.26 | 0.41 | 1.84 | 0.74 | 0.93 | 0.51 | – | – | – | – | – | – | 1.70 1.43 |
| Meleagrinella (Meleagrinella) aff. prima | ||||||||||||||||||||
| Ст-1/1, левая | р. Старт, обн. 5, стартинская свита, осыпь слоя 2, зона Pseudolioceras compactile | 17.77 | 16.5 | 0.93 | – | 5.08 | 0.31 | 8.30 | 0.47 | 9.47 | 0.57 | 5.67 | 0.60 | 53° | 56 | 3.15 | 1.81 | – | 0.17 | – |
Размеры, мм и отношения. См. табл. 4.
Индивидуальная изменчивость. Размеры раковины от очень маленьких (менее 10 мм) до маленьких (до 30 мм). Контур левой створки варьирует от очень низкого до высокого. Симметрия левых створок изменяется от неравносторонней до умеренно неравносторонней. Заднее залигаментное поле левой створки варьирует от очень короткого до длинного. Плотность ребристости левой створки изменяется от часторебристой до густоребристой.
Сравнение. От Meleagrinella (Clathrolima) substriata (Münster) из зоны Hildoceras bifrons (подзона Dactylioceras commune) отличается субпрямоугольным задним ушком и вырезом под задним ушком.
Замечания. Мелеагринеллы, выделенные в новый вид, ранее О.А. Лутиковым относились к виду Arctotis (Praearctotis) milovae (Okuneva) (Лутиков, Шурыгин, 2010; Лутиков, 2021). Очень маленькие размеры (до 10 мм), субпрямоугольное заднее ушко и густая ребристость сближают представителей нового вида из зоны Zugodactylites braunianus (р. Марха, Анабарская губа) с видом Meleagrinella milovae из верхнего тоара Восточного Забайкалья (Окунева, 2002, с. 379, табл. CXXIV , фиг. 15–18 ). В 2022 г. О.А. Лутиковым была переизучена типовая коллекция Т.М. Окуневой, хранящаяся в ЦНИГР музее им. Ф.Г. Чернышева (Санкт-Петербург), с использованием новой системы градации признаков. Левые створки у представителей дальневосточной популяции характеризуются менее выгнутым задним крылом, нижнее окончание заднего крыла левой створки расположено выше заднего конца створки (табл. III, фиг. 14 ). Точную родовую принадлежность таксона установить невозможно ввиду плохой сохранности ископаемого материала. Типовая выборка представлена ракушняком, состоящим из разрозненных очень маленьких створок плохой сохранности примерно одного размера (табл. III, фиг. 15 ). С учетом данных по существованию протяженного участка суши, разделявшего в тоарском веке восточносибирские и дальневосточные моря (Девятов и др., 2011), можно предположить, что популяция из бассейна р. Газимур была изолирована и относится к самостоятельному виду. Эта локальная популяция занимает в разрезе стратиграфический уровень выше, чем слои с Pseudolioceras rosenkrantzi, и может относиться к виду Arctotis (Praearctotis) marchaensis (Petrova), но это предположение требует проверки и дополнительных сборов.
У представителей описываемого таксона в выборке из третьей пачки сунтарской свиты (р. Марха, Тенкеляхский участок) форма лигаментной ямки на ювенильной стадии – широкоугольная (ДНЛ/ДЗЛ более 1.5) (табл. III, фиг. 9, 10 ). По этому признаку эта филетическая группа не относится к подроду Paemeleagrinella, имеющему прямоугольную форму лигаментной ямки на ювенильной стадии (ДНЛ/ДЗЛ менее 1.5). Микроскульптура остракума в нижней части правой створки представлена в основном четырехугольными призмами, которые расположены параллельно радиальным ребрам от макушки в направлении нижнего края (табл. IV, фиг. 9 ). У представителей Arctotis (Praearctotis) marchaensis в выборке из четвертой пачки сунтарской свиты остракум правых створок характеризуется шестиугольными призмами, расположенными неупорядоченно (табл. IV, фиг. 5в). По этому признаку обе популяции принадлежат к разным надвидовым группам. Популяции c тупоугольной лигаментной ямкой на ювенильной стадии из зоны Zugodactylites braunianus относятся к Meleagrinella s.str., а популяции из зоны Pseudolioceras wuerttenbergeri относятся к подроду Arctotis (Praearctotis) Lutikov et Schurygin.
На Северо-Востоке России (рр. Мунугуджак, Старт) в зоне Pseudolioceras compactile встречены мелеагринеллы, близкие по морфологии к описываемому виду, но отличающиеся крупными размерами (до 30 мм в высоту) (Полевой…, 1968, табл. 39 , фиг. 1–2; здесь табл. III, фиг. 12, 13 ). С учетом более высокого стратиграфического положения и размеров эти экземпляры отнесены к Meleagrinella (Meleagrinella) aff. prima sp. nov.
Фациальная приуроченность и тафономическая характеристика. На р. Марха и в Анабарской губе раковины образуют ракушняковые скопления. Раковины не сортированы, но радиальные ребра имеют следы потертости. Мелеагринеллы встречаются совместно с аммонитами, дакриомиями, лиострями, окситомами, леноцерамусами, модиолусами и танкредиями. Тип ископаемого ценоза – слабо перемещенный танатоценоз. Захоронение происходило вблизи места обитания. На рр. Астрономическая и Сатурн раковины образуют гнездовидные ракушняковые скопления в массивных конкрециях. Разрозненные створки плохой сохранности захоронены с аммонитами, белемнитами, астартами, энтолиумами, кукуллеями, пропеамуссиумами, окситомами, лиостреями, гониомиями. Тип ископаемого ценоза – перемещенный танатоценоз. В междуречье Хання–Тюкян (Тенкеляхский участок) раковины образуют ракушняковые скопления из целых раковин и отдельных створок хорошей сохранности, представлены как взрослыми, так и ювенильными особями. Захоронение происходило на месте обитания. Тип ископаемого ценоза – автохтонный танатоценоз.
Образ жизни и условия обитания. Обитали на мелкопесчаных грунтах, на небольших глубинах вблизи берега. Прикреплялись биссусом к твердому субстрату. Селились колониями.
Геологический возраст и географическое распространение. Восточная Сибирь, Анабарская губа, р. Марха, междуречье Хання–Тюкян. Северо-Восток России, рр. Астрономическая, Сатурн, Старт; нижний тоар, зоны Zugodactylites braunianus, Pseudolioceras compactile.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
На основе ранее предложенной методики изучения морфогенеза замочных структур у двустворчатых моллюсков семейства Oxytomidae Ichikawa, 1958 (Лутиков и др., 2010) осуществлена ревизия представителей Meleagrinella из тоара России и Германии. Предложенная классификация основана на наборе взвешенных признаков. Для распределения таксонов по группам за основное звено развития были приняты изменения относительного положения краев лигаментной ямки, определяющих ее форму. Общей закономерностью онтогенезов лигаментной ямки юрских окситомид является резкое изменение формы, которое случалось дважды, иногда трижды по мере увеличения лигаментного блока у раковины моллюска. По первым трем точкам излома переднего края выделены три стадии состояния лигаментного блока, условно сопоставляемые с тремя возрастными периодами развития организмов – ювенильным, юношеским и взрослым. Тип лигаментной ямки определялся относительным положением всех ее сторон в исходном состоянии, которое наблюдалось у самых мелких раковин (менее 3 мм) или у взрослых раковин (менее 10 мм) вблизи продисоконха. В зависимости от угла наклона переднего края лигаментной ямки к ее нижнему краю, форма лигаментной ямки у окситомид может меняться в онтогенезе и становиться клиновидной, остроугольной, субпрямоугольной, широкоугольной или субсимметричной. Путем измерения отношения длины нижнего края ямки к длине проекции заднего края ямки (ДНЛ/ДЗЛ) была проведена количественная оценка этого признака. Проведенные замеры ДНЛ/ДЗЛ лигаментной ямки на трех стадиях состояния лигаментного блока в выборках из разных стратиграфических уровней позволили установить отличия плинсбахских Meleagrinella (Praemeleagrinella) от тоарских Meleagrinella s.str.
Эволюция онтогенезов лигаментной ямки плинсбах-тоарских Meleagrinella и Arctotis принята за основу реконструкции филогенеза окситомид в раннем тоаре (рис. 6). В качестве исходной формы в ряду преемственных филогенетических таксонов рассматривался вид Meleagrinella (Praemeleagrinella) deleta (Dumortier) из верхней зоны плинсбаха Франции (табл. I , фиг. 1). К этому таксону ранее были отнесены мелкие мелеагринеллы, распространенные в терминальных слоях верхнеплинсбахских толщ Восточной Сибири (Восточный Таймыр, Анабарская губа, р. Келимяр) (Лутиков и др., 2022). Для оценки начального состояния лигаментного блока в плинсбах-тоарской хроноклине была исследована небольшая выборка (около 20 экземпляров) из керна скважины в районе междуречья Марха–Вилюй (Правобережный участок бурения). Выборка происходит из тюнгской свиты. Измерения проводились по двум левым створкам, имевшим наибольшие различия в строении лигаментного блока (табл. I , фиг. 2–3). Форма лигаментной ямки у обоих экземпляров на ювенильной стадии до первого перелома в онтогенезе широкоугольная. Отношение ДНЛ/ДЗЛ на ювенильной стадии у обеих раковин варьирует от 1.35 до 1.44.
Форма лигаментной ямки на второй (юношеской) стадии изменяется – передний край ямки коленообразно изгибается, а его нижнее окончание перемещается в направлении заднего края. При этом задний край ямки остается прямым, как на ювенильной стадии, и удлиняется в направлении заднего края раковины. Отношение ДНЛ/ДЗЛ у обеих раковин на юношеской стадии колеблется от 1.15 до 1.22, что свидетельствует о сужении ямки. Уменьшение относительной длины нижнего края лигаментной ямки придает ей слабоскошенный вид. Нижний край лигаментной ямки у обоих экземпляров прямой, на псевдосвязочной площадке левой створки вблизи переднего края лигаментной ямки развит эллипсоидальный протуберанец, лигаментная площадка левой створки лежит в плоскости смыкания створок. По этим признакам локальная популяция относится к подроду Praemeleagrinella. Форма лигаментной ямки на взрослой стадии онтогенеза в выборке имеет две вариации. У одного экземпляра передний лигаментный желобок изгибается по направлению к заднему краю и образует с нижним краем лигаментной площадки тупой угол (табл. I , фиг. 3а). Уменьшение длины нижнего края лигаментной ямки по отношению к длине проекции заднего края лигаментной ямки на второй и третьей стадиях состояния этого признака определяют “скошенно-сужающийся” подвид онтогенеза лигаментной ямки. У второго экземпляра передний лигаментный желобок изгибается по направлению к переднему концу раковины и образует с нижним краем лигаментной площадки острый угол (табл. I , фиг. 3в). Уменьшение длины нижнего края лигаментной ямки по отношению к длине проекции заднего края лигаментной ямки на второй стадии и увеличение длины нижнего края лигаментной ямки по отношению к длине проекции заднего края лигаментной ямки на третьей стадии состояния этого признака определяют “скошенно-расширяющийся” подвид онтогенеза лигаментного блока.
Наиболее ранним представителем рода Meleagrinella в тоаре является вид Meleagrinella (Praemeleagrinella?) golberti, появившийся в фазе Dactylioceras tenuicostatum (=Tiltoniceras antiquum). Обоснованием присутствия этого вида в нижней зоне тоара являются совместные его находки с аммонитом Tiltoniceras sp. ind. (табл. IV, фиг. 6) в основании курунской пачки на р. Келимяр. Присутствие возрастных аналогов нижней зоны тоара на р. Келимяр предполагалось ранее (Никитенко, 2009; Девятов и др., 2010). Находка представителя рода Tiltoniceras впервые доказывает присутствие зоны Tiltoniceras antiquum в Восточной Сибири. Вид Meleagrinella (Praemeleagrinella?) golberti широко распространен во второй зоне нижнего тоара. На р. Келимяр этот таксон встречен совместно с Harpoceras exaratum (Young et Bird), Harpoceras falciferum (Sowerby) (Князев и др., 1984, 2003). В разрезе Канала Людвига с этим видом встречены аммониты Cleviceras elegans (Sowerby), Cleviceras cf. exaratum (Young et Bird), Phylloceras heterophyllum (Sowerby), Harpoceras serpentinum (Schlotheim), “Peronoceras” desplacei (d’Orbigny), Nodicoeloceras crassoides (Simpson), Dactylioceras semiannulatum Howarth, Dactylioceras anguinum (Reinecke) (Arp et al., 2021).
Вид Meleagrinella (Praemeleagrinella?) golberti отличается от Meleagrinella (Praemeleagrinella) deleta выгнутым задним крылом левой створки. Закрепление этого фена естественным отбором происходило в локальных популяциях на территории Восточной Сибири, Северо-Востока России, Германии, Англии и Канады в фазе Harpoceras serpentinum (=Harpoceras falciferum). В условиях дефицита кислорода в придонных водах вид, вероятно, вел псевдопланктонный образ жизни, прикрепляясь к плавающим водорослям или раковинам живых аммонитов. Биозона вида охватывает аммонитовые зоны Dactylioceras tenuicostatum и Harpoceras serpentinum суббореальной шкалы (Page, 2003) и одновозрастные зоны Tiltoniceras antiquum и Harpoceras falciferum бореальной шкалы (Захаров и др., 1997; Князев и др., 2003; Шурыгин и др., 2011).
Вторым в тоарской последовательности является вид Meleagrinella (Clathrolima) substriata, появившийся в фазе Hildoceras bifrons (=Dactylioceras commune). На р. Тюнг с этим видом встречены аммониты Dactylioceras commune (Sowerby), D. crassifactum (Simpson), D. athleticum (Simpson), D. suntarense Krimholz, D. amplum А. Dagis, Catacoeloceras crassum (Young et Bird) (Князев и др., 2003). На р. Астрономическая и на Шпицбергене этот вид найден совместно с аммонитами Dactylioceras commune (Sowerby), Dactylioceras kopiki Rogov (Князев и др., 2003; Rogov, Lutikov, 2022). В разрезе Канала Людвига (Дёрльбах, Германия) вид встречен с аммонитами Dactylioceras commune (Sowerby), Dactylioceras athleticum (Simpson), Hildoceras cf. lusitanicum Meister (Arp et al., 2021). Этот таксон отличается от Meleagrinella (Praemeleagrinella?) golberti тупоугольным ушком и S-образным задним крылом. Закрепление этого фена естественным отбором происходило в локальных популяциях на территории Восточной Сибири, Северо-Востока России, Шпицбергена и Германии в фазе Hildoceras bifrons (=Dactylioceras commune). Единственный крупный экземпляр (около 32 мм) с сохранившимся лигаментным блоком из типовой выборки Г. Мюнстера характеризуется остроугольными очертаниями лигаментной ямки на взрослой стадии (ДНЛ/ДЗЛ = 0.90) и наличием протуберанца (табл. III, фиг. 4б). По этим признакам этот вид принадлежит к подроду Clathrolima. К виду относятся мелеагринеллы, распространенные в верхней подсвите стартинской свиты (р. Астрономическая; зона Dactylioceras commune), во второй пачке сунтарской свиты (рр. Тюнг, Марха; зона Dactylioceras commune) и в нижней части эренской свиты (Анабарская губа; зона Dactylioceras commune). Биозона Meleagrinella (Clathrolima) substriata охватывает нижнюю часть зоны Hildoceras bifrons (подзона Dactylioceras commune) суббореального стандарта и зону Dactylioceras commune (=Harpoceras subplanatum) бореального стандарта.
Третьим в тоарской последовательности является вид Meleagrinella (Meleagrinella) prima, появившийся в фазе Zugodactylites braunianus. На р. Марха с этим видом встречены аммониты Zugodactylites braunianus, Catacoeloceras crassum (Князев и др., 2003), на Анабарской губе – аммониты Zugodactylites braunianus (d’Orbigny), Pseudolioceras lythense (Young et Bird).
На рр. Астрономическая и Сатурн с этим видом встречены аммониты Zugodactylites braunianus, Pseudolioceras lythense, Pseudolioceras compactile (Simpson), Poroceras vortex (Simpson), Collina gemma Bonarelli, Catacoeloceras confectum (Buckman) (Князев и др., 2003). Этот таксон отличается от Meleagrinella (Clathrolima) substriata сильновыгнутым задним крылом, нижнее окончание которого расположено ниже заднего конца левой створки. Закрепление этого фена естественным отбором происходило в локальных популяциях на территории Восточной Сибири и Северо-Востока России в фазе Zugodactylites braunianus. Для количественной оценки состояний лигаментного блока у этого таксона были исследованы две выборки в образцах из обнажений западного берега Анабарской губы (около 20 экземпляров) и из керна скважин в междуречье Тюкян–Марха (Тенкеляхский участок) (около 20 экземпляров). Выборка из обнажения западного берега Анабарской губы из эренской свиты, выборка из керна сважины Тенкеляхского участка происходят из сунтарской свиты. Измерения отношения ДНЛ/ДЗЛ на ювенильной стадии проводились по двум левым створкам, имеющим ширину менее 3 мм, с сохранившимися лигаментными площадками. Оба экземпляра на ювенильной стадии характеризуются субсимметричной лигаментной ямкой. У анабарского экземпляра отношение ДНЛ/ДЗЛ = 1.87 (табл. III, фиг. 9 ). У тенкеляхского экземпляра отношение ДНЛ/ДЗЛ = 1.70 (табл. III, фиг. 10 ). Отсутствие протуберанца и субсимметричная форма лигаментной ямки на ювенильной стадии характерны для подрода Meleagrinella s.str. Форма лигаментной ямки у тенкеляхского экземпляра на второй (юношеской) стадии перелома онтогенеза широкоугольная (ДНЛ/ДЗЛ = 1.43). Уменьшение относительной длины нижнего края лигаментной ямки на юношеской стадии свидетельствует о ее сужении, как и у Praemeleagrinella. У крупного (около 10 мм) анабарского экземпляра форма лигаментной ямки на второй стадии перелома онтогенеза также широкоугольная (ДНЛ/ДЗЛ = 1.45), а на третьей (взрослой) стадии – субсимметричная (ДНЛ/ДЗЛ = 1.55) (табл. III, фиг. 11 ). Увеличение относительной длины нижнего края лигаментной ямки свидетельствует о ее расширении на взрослой стадии. Эта особенность присутствовала у всех тенкеляхских экземпляров. К этому таксону относятся мелеагринеллы, распространенные в верхней подсвите стартинской свиты (рр. Астрономическая, Сатурн), в третьей пачке сунтарской свиты (р. Марха, Тенкеляхский участок), в верхней части эренской свиты (Анабарская губа). В Европе этот таксон достоверно не установлен.
Биозона Meleagrinella (Meleagrinella) prima охватывает аммонитовые зоны Zugodactylites monestieri (=Pseudolioceras lythense) и Pseudolioceras compactile бореальной шкалы (Захаров и др., 1997; Князев и др., 2003; Шурыгин и др., 2011). Одновременно с Meleagrinella (Meleagrinella) prima в параллельной филетической линии образовался вид Meleagrinella (Clathrolima) sp. Этот таксон встречен в третьей пачке сунтарской свиты (Правобережный участок). Единственный экземпляр с сохранившимся лигаментным блоком из типовой выборки характеризуется остроугольными очертаниями лигаментной ямки на взрослой стадии (ДНЛ/ДЗЛ = 0.97) и наличием плохо выраженного эллипсоидального протуберанца.
Сменяющие друг друга в последовательных слоях тоара таксоны демонстрируют разнонаправленное изменение формы лигаментной ямки. В ряду Meleagrinella (Praemeleagrinella) deleta–Meleagrinella (Praemeleagrinella ?) golberti–Meleagrinella (Clathrolima) substriata–Meleagrinella (Clathrolima) sp. изменения формы лигаментной ямки происходили в направлении образования у нее остроугольной формы. У взрослых особей в этой хроноклине значение отношения длины нижнего края лигаментной ямки к длине проекции заднего края лигаментной ямки уменьшается от 1.10 до 0.90. Скошенная раковина и остроугольная лигаментная ямка, характерные для Meleagrinella (Clathrolima) substriata, являются адаптациями к биссусному способу прикрепления к гибким, возвышающимся над дном субстратам (преимущественно водорослям). Эти приспособления, вероятно, были конкурентно оправданны при обитании в условиях активного гидродинамического режима и были закреплены естественным отбором в начале фазы Hildoceras bifrons c образованием подрода Clathrolima.
В ряду в Meleagrinella (Praemeleagrinella) deleta–Meleagrinella (Praemeleagrinella ?) golberti–Meleagrinella (Meleagrinella) prima изменения формы лигаментной ямки происходили в направлении расширения в длину и затем расширения в ширину у Arctotis (Praearctotis) marchaensis. У взрослых особей в этой хроноклине значение отношения длины нижнего края лигаментной ямки к длине проекции заднего края лигаментной ямки (ДНЛ/ДЗЛ) увеличивалось от 1.10 до 1.62.
“Скошенно-расширяющийся” подвид онтогенеза лигаментной ямки, возникший уже в конце плинсбахского века у предковой группы Meleagrinella (Praemeleagrinella) deleta, мог являться мутацией (табл. IV, фиг. 3в). Изменение в направлении расширения площади лигаментной ямки, необходимое для усиления волокнистой связки, было эффективной адаптацией для обитания в приближенных к берегу мелководно-морских обстановках с повышенной гидродинамикой, что было закреплено естественным отбором в фазе Zugodactylites braunianus. Триггером появления первых Meleagrinella s.str. стал результат разрастания в ширину внутренней связки. Округлая форма и слабовыраженная скульптура верхней примакушечной части правой створки у Meleagrinella (Meleagrinella) prima и Arctotis (Praearctotis) marchaensis свидетельствуют о близком прилегании раковин к субстрату (табл. III, фиг. 8; табл. IV, фиг. 5).
Если предположить, что предковым видом для Meleagrinella (Meleagrinella) prima был вид Meleagrinella (Clathrolima) substriata, то морфогенез лигаментной ямки происходил бы в направлении расширения относительно оси створки скачкообразно. Значение отношения длины нижнего края лигаментной ямки к длине проекции заднего края лигаментной ямки (ДНЛ/ДЗЛ) увеличивалось от 0.95 до 1.62. В этом случае приходится признать сальтационный механизм возникновения Meleagrinella (Meleagrinella) prima от предкового вида Meleagrinella (Clathrolima) substriata. Процесс надвидовой перестройки в подобном случае должен был проходить довольно быстро в пределах одной аммонитовой фазы.
Более вероятным представляется происхождение Meleagrinella (Meleagrinella) prima от таксона, относящегося к подроду Meleagrinella (Praemeleagrinella) (рис. 6). Предположительно это был вид Meleagrinella (Praemeleagrinella ?) aff. golberti, находки которого встречены с аммонитами Harpoceras falciferum (Sowerby) на р. Бродная (Князев и др., 2003). Однако нахождение этого таксона в зоне Dactylioceras commune достоверно не установлено. В этом случае дивергенция, вероятно, случилась либо в фазе Harpoceras falciferum, либо в фазе Zugodactylites braunianus (=Peronoceras fibulatum–Catacoeloceras crassum).
Эволюция окситомид в условиях мелководных морей Восточной Сибири происходила в направлении расширения лигаментной ямки. Эволюция окситомид в условиях более глубоких морей Южной Германии происходила в направлении сужения лигаментной ямки. При этом сценарии обе филетические линии – Clathrolima и Meleagrinella s.str., отходящие от общего предкового ствола Praemeleagrinella, в тоаре эволюционировали параллельно. В пользу этого варианта филогении свидетельствуют находки мелеагринелл со смешанными признаками в верхней части сунтарской свиты Правобережного участка (табл. III, фиг. 12). У вида Meleagrinella (Clathrolima) sp. устройство лигаментного блока соответствует Meleagrinella (Clathrolima), все остальные наружные признаки, включая видоспецифические – вырез под задним ушком и сильно выгнутое заднее крыло, соответствуют Meleagrinella (Meleagrinella) prima. Благодаря гомологичному мутированию и параллелизму линий Clathrolima и Meleagrinella s.str., признаки нового вида Meleagrinella (Meleagrinella) prima появились не в одной, а в нескольких популяциях (Анабарского, Тенкеляхского и Прибрежного участков). Появление в массовом количестве определенных фенов в параллельных близкородственных филолиниях может выглядеть как мгновенное расселение видов (Красилов, 1977).
Для раннего тоара (фазы Harpoceras falciferum, Dactylioceras commune) при высоком положении уровня Мирового океана объяснение параллельной последовательности видов рода Meleagrinella в различных разрезах возможно и с помощью концепции Дарвина–Хаксли, согласно которой доминирующие прогрессивные виды быстро расселялись из центра происхождения и повсеместно вытесняли более примитивные предковые виды (Дарвин, 1991). В фазах Zugodactylites braunianus и Pseudolioceras compactile одновременное конкурентное исключение предковых таксонов дочерними может вызывать возражения. Маловероятно, что в Арктической и Бореально-Атлантической палеобиогеографических областях существовали хорошие связи между бассейнами, поскольку комплексы бентосной фауны Восточной Сибири, Северо-Востока России и Германии имеют мало общего.
У представителей популяции Meleagrinella (Meleagrinella) prima из третьей пачки сунтарской свиты (р. Марха, Тенкеляхский участок) микроскульптура остракума в средней части правой створки в основном состоит из четырехугольных призм, расположенных параллельно радиальным ребрам по направлению от макушки к нижнему краю (табл. IV, фиг. 7). У представителей популяции Arctotis (Praearctotis) marchaensis из четвертой пачки сунтарской свиты микроскульптура остракума правых створок характеризуется шестиугольными призмами, расположенными неупорядоченно (табл. IV, фиг. 5в). Случайно возникшее изменение микроскульптуры остракума было закреплено естественным отбором в фазе Grammoceras thouarsense (=Pseudolioceras wuerttenbergeri), с отделением филетической линии Arctotis. Начальная стадия формирования нового адаптивного типа носила квантовый характер и отличалась высокими темпами эволюции. Это приспособление, вероятно, было конкурентно оправданно при обитании в условиях охлаждения морских бассейнов. К уровню появления первых преарктотисов приурочены прослои с глендонитами – индикаторами холодноводности (Rogov et al., 2021). Группы Meleagrinella s.str. и Arctotis имеют субсимметричную лигаментную ямку на ювенильной стадии и образуют одну вертикальную граду, эволюционирующую параллельно группе Clathrolima (рис. 6).
ЗАКЛЮЧЕНИЕ
Среди многообразия позднеплинсбахских и раннетоарских мелеагринелл установлено три морфологических типа, принадлежащих трем надвидовым таксонам: Praemeleagrinella, Clathrolima, Meleagrinella s.str. (рис. 6), чья дискретность определялась по признакам лигаментного блока.
Полученные в результате проведенных исследований сведения существенно меняют представления об объеме вида Monotis substriata (Münster). С помощью предложенной системы градаций наружных признаков определены границы дискретности видов. В результате проведенной ревизии мелеагринелл в нижнем тоаре Германии установлено два последовательно сменяющих друг друга вида: Meleagrinella (Praemeleagrinella ?) golberti sp. nov. и Meleagrinella (Clathrolima) substriata (Münster). В тоаре Восточной Сибири и Северо-Востока России установлено три последовательно сменяющих друг друга вида: Meleagrinella (Praemeleagrinella ?) golberti sp. nov., Meleagrinella (Clathrolima) substriata (Münster) и Meleagrinella (Meleagrinella) prima sp. nov. Каждому из этих таксонов соответствует определенный набор признаков. Ввиду недостаточного материала для обоснования выделения новых видов, три таксона обозначены в открытой номенклатуре: Meleagrinella (Praemeleagrinella ?) aff. golberti sp. nov., Meleagrinella (Meleagrinella) aff. prima sp. nov., Meleagrinella (Clathrolima) sp.
Формирование Meleagrinella (Praemeleagrinella?) golberti произошло в фазе Dactylioceras tenuicostatum и было связано с появлением нового фена – выгнутого заднего крыла. Биозона вида охватывает аммонитовые зоны Dactylioceras tenuicostatum–Harpoceras serpentinum суббореального стандарта и одновозрастные им зоны Tiltoniceras antiquum–Harpoceras falciferum бореального стандарта. Вид наиболее широко распространен в тоаре Восточной Сибири, Северо-Востока России, Германии, Англии (Йоркшир) и Западной Канады.
Формирование Meleagrinella (Clathrolima) substriata произошло в фазе Hildoceras bifrons и связано с появлением новых фенов – остроугольной лигаментной ямки и тупоугольного заднего ушка. Биозона вида охватывает аммонитовую подзону Dactylioceras commune суббореального стандарта и одновозрастную ей зону Dactylioceras commune бореального стандарта. Вид широко распространен в тоаре Восточной Сибири, Северо-Востока России, Шпицбергена и Германии.
Формирование Meleagrinella (Meleagrinella) prima произошло в фазе Hildoceras bifrons и связано с появлением нового фена – субсимметричной лигаментной ямки и сильновыгнутого заднего крыла левой створки. Биозона вида охватывает зону Zugodactylites braunianus и зону Pseudolioceras compactile бореального стандарта. Вид распространен в тоаре Восточной Сибири и Северо-Востока России.
Морфологическое обособление рода Arctotis в позднем тоаре Восточной Сибири фиксируется резким изменением типа микроскульптуры остракума правой створки. Предположительно предковой формой Arctotis (Praearctotis) marchaensis является вид Meleagrinella (Meleagrinella) prima, у которого имеются все морфологические признаки в лигаментном блоке, присущие Arctotis (Praearctotis). Наличие у ювенильных форм Meleagrinella (Meleagrinella) prima субсимметричной ямки, как у Arctotis s.str., свидетельствует о значении внутренней организации подрода Meleagrinella s.str. как причины канализации филогенеза (рис. 6).
Вид Meleagrinella (Praemeleagrinella ?) golberti распространен по всей Панбореальной палеобиогеографической надобласти. Географический ареал распространения вида Meleagrinella (Clathrolima) substriata ограничен Бореально-Атлантической и Арктической палеобиогеографическими областями. Географический ареал распространения вида Meleagrinella (Meleagrinella) prima ограничен Арктической палеобиогеографической областью. В Бореально-Атлантической области этот вид не установлен. Анализ ареалов распространения изученных таксонов был проведен с использованием собственных наблюдений авторов, а также опубликованных данных (Князев и др., 1984, 1991, 2003; Решения…, 2009; Arp et al., 2021; Rogov, Lutikov, 2022).
Благодарности. При подготовке рукописи авторы получали значимые консультации у В.П. Девятова (СНИИГГиМС, Новосибирск), М.А. Рогова (ГИН РАН, Москва), Ю.С. Репина (ВНИГРИ, Санкт-Петербург), В. Вернера (Bavarian State Collection for paleontology and geology, Munchen). Помощь в ознакомлении с типовой коллекцией, изображенной в монографии Г. Гольдфусса, оказали Г. Хойманн (Georg Heumann) (Goldfuss-Museum; Institute for Geosciences, Sec. Palaeontology, Bonn) и В. Вернер (Winfried Werner). Фотографии оригинальной коллекции Г. Мюнстера и Г. Гольдфусса выполнили Г. Олешински (Georg Oleschinski) (Institute for Geosciences, Sec. Palaeontology, Bonn) и В. Вернер. Фотографии микроскульптуры выполнил А.А. Мироненко (ГИН РАН, Москва). Ряд ценных замечаний и рекомендаций, способствовавших улучшению статьи, получены от С.В. Попова (ПИН РАН, Москва) и Б.Н. Шурыгина (ИНГГ СО РАН, Новосибирск). Всем указанным специалистам авторы выражают искреннюю благодарность. Отдельную благодарность выражаем В.А. Захарову (ГИН РАН, Москва) за советы при проведении исследования и оформлении данной статьи.
Источники финансирования. Работа выполнена по теме госзадания ГИН РАН при поддержке гранта РФФИ и Национального центра научных исследований Франции в рамках научного проекта № 21-55-15015.
Список литературы
Атлас руководящих форм ископаемых фаун СССР. Т. 8. Нижний и средний отделы юрской системы. М.–Л.: Госгеолиздат, 1947. 278 с.
Бодылевский В.И., Шульгина Н.И. Юрские и меловые фауны низовьев Енисея. М.: Госгеолтехиздат, 1958. 196 с.
Борисяк A.A. Pelecypoda юрских отложений Европейской России. Вып. IV. Aviculidae. CПб.: Изд-во Геол. комитета, 1909. 26 с.
Борисяк А.А. О Pseudomonotis (Eumorphotis) lenaensis Lah. Sp. (=Hinnites lenaensis Lah.) // Тр. Геол. музея им. Петра Великого Императорской Академии наук. Петроград: Изд-во Императорской Академии наук, 1915. Т. VIII. С. 141–153.
Дагис А.А. Тоарские аммониты (Dactylioceratidae) севера Сибири // Тр. ИГиГ СО АН СССР. 1968. Вып. 40. 108 с.
Дагис А.А., Дагис А.С. О зональном расчленении тоарских отложений на Северо-Востоке СССР // Стратиграфия и палеонтология мезозойских отложений севера Сибири. М.: Наука, 1965. С. 15–26.
Дарвин Ч. Происхождение видов путем естественного отбора. СПб.: Наука, 1991. 546 с.
Девятов В.П., Князев В.Г., Никитенко Б.Л., Мельник О.А., Глинских Л.А. Граница плинсбаха и тоара севера Восточной Сибири и стратиграфическое положение курунгской пачки келимярской свиты (р. Келимяр, бассейн р. Оленёк) // Отечеств. геология. 2010. № 5. С. 105–112.
Девятов В.П., Никитенко Б.Л., Шурыгин Б.Н. Палеогеография Сибири в юрском периоде на этапах основных перестроек // Новости палеонтологии и стратиграфии. Вып. 16–17. Приложение к журн. “Геология и геофизика”. 2011. Т. 52. С. 87–101.
Захаров В.А., Богомолов Ю.И., Ильина В.И., Константинов А.Г., Курушин Н.И., Лебедева Н.К., Меледина С.В., Никитенко Б.Л., Соболев Е.С., Шурыгин Б.Н. Бореальный зональный стандарт и биостратиграфия мезозоя Сибири // Геология и геофизика. 1997. Т. 38. № 5. С. 927–956.
Князев В.Г., Девятов В.П., Лутиков О.А. Тоарский ярус, его зональное деление и граница нижней и средней юры на востоке Сибирской платформы // Проблемы ярусного расчленения систем фанерозоя Сибири. Новосибирск: Изд-во СНИИГГиМС, 1984. С. 58–66.
Князев В.Г., Девятов В.П., Шурыгин Б.Н. Стратиграфия и палеогеография ранней юры востока Сибирской платформы. Якутск: ЯНЦ СО АН СССР, 1991. 100 с.
Князев В.Г., Кутыгин Р.В., Девятов В.П., Никитенко Б.Л., Шурыгин Б.Н. Зональный стандарт тоарского яруса Северо-Востока азиатской части России. Якутск: Изд-во СО РАН, 2003. 103 с.
Кошелкина З.В. Стратиграфия и двустворчатые моллюски юрских отложений Вилюйской синеклизы и Приверхоянского краевого прогиба. Магадан, 1963. 274 с.
Красилов В.А. Эволюция и биостратиграфия. М.: Наука, 1977. 256 с.
Лутиков О.А. Биохронологическая шкала верхнего тоара–нижнего аалена Восточной Сибири по двустворчатым моллюскам рода Arctotis Bodylevsky, 1960. Стратиграфия и геологическая корреляция // Стратиграфия. Геол. корреляция. 2021. Т. 29. № 6. С. 54–83. https://doi.org/10.31857/S0869592X21060065
Лутиков О.А., Арп Г. Ревизия Monotis substriata (Münster, 1831) и новые виды двустворчатых моллюсков в нижнем тоаре на севере России и юге Германии (семейство Oxytomidae Ichikawa, 1958) // Юрская система России: проблемы стратиграфии и палеогеографии. Материалы VIII Всероссийского совещания с международным участием. Онлайн-конференция, 7–10 сентября 2020 г. Отв. ред. Захаров В.А. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 126–131.
Лутиков О.А., Шурыгин Б.Н. Новые данные по систематике юрских и меловых двустворчатых моллюсков семейства Oxytomidae Ichikawa, 1958 // Новости палеонтологии и стратиграфии. Вып. 14. Приложение к журн. “Геология и геофизика”. 2010. Т. 51. С. 111–140.
Лутиков О.А., Тёмкин И.Е., Шурыгин Б.Н. Эволюция онтогенезов и филогения некоторых представителей семейcтва Oxytomidae Ichikawa, 1958 (Mollusca: Bivalvia) // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 4. С. 28–44.
Лутиков О.А., Шурыгин Б.Н., Сапьяник В.В., Алейников А.Н., Алифиров А.С. Новые данные по стратиграфии юрских (плинсбах-ааленских) отложений района мыса Цветкова, Восточный Таймыр // Стратиграфия. Геол. корреляция. 2022. Т. 30. № 1. С. 69–93. https://doi.org/10.31857/S0869592X22010033
Майр Э. Принципы зоологической систематики. М.: Мир, 1971. 455 с.
Невесская Л.А., Скарлато О.А., Старобогатов Я.И. Эберзин А.Г. Новые представления о системе двустворчатых моллюсков // Палеонтол. журн. 1971. № 2. С. 3–20.
Невесская Л.А., Попов С.В., Гончарова И.А., Гужов А.В., Янин Б.Т., Полуботко И.В., Бяков А.С., Гаврилова В.А. Двустворчатые моллюски России и сопредельных стран в фанерозое // Тр. ПИН РАН. 2013. Т. 294. 524 с.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Окунева Т.М. Юрская система. Нижний и средний отделы. Разделы: Морские отложения и Моллюски // Атлас фауны и флоры палеозоя и мезозоя Забайкалья. Новосибирск: Наука, 2002. С. 354–387.
Полевой атлас юрской фауны и флоры Северо-Востока СССР. Магадан: Магаданское книжн. изд-во, 1968. 379 с.
Репин Ю.С., Полуботко И.В. Биохронология тоара Арктической палеозоохории // Стратиграфия нефтегазоносных бассейнов России. СПб.: Недра, 2004. С. 93–124.
Решение 6-го Межведомственного стратиграфического совещания по рассмотрению и принятию уточненных стратиграфических схем мезозойских отложений Западной Сибири (Новосибирск, 2003 г.). Новосибирск: СНИИГГиМС, 2004. 114 с.
Решения 4-го Межведомственного регионального стратиграфического совещания по докембрию и фанерозою юга Дальнего Востока и Восточного Забайкалья. Хабаровск: ХГГГП, 1994. 124 с.
Решения 3-го Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России (Санкт-Петербург, 2002). СПб.: Изд-во ВСЕГЕИ, 2009. 266 с.
Руженцев В.Е. Принципы систематики, система и филогения палеозойских аммоноидей // Тр. Палеонтол. ин-та АН СССР. 1960. Т. 83. 331 с.
Симпсон Д.Г. Принципы таксономии животных. М.: Товарищество научных изданий КМК, 2006. 293 с.
Стратиграфия юрской системы севера СССР. М.: Наука, 1976. 436 с.
Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П., Ильина В.И., Меледина С.В., Гайдебурова Е.А., Дзюба О.С., Казаков А.М., Могучева Н.К. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск: Изд-во СО РАН, филиал “Гео”, 2000. 480 с.
Шурыгин Б.Н., Никитенко Б.Л., Меледина С.В., Дзюба О.С., Князев В.Г. Комплексные зональные шкалы юры Сибири и их значение для циркумарктических корреляций // Геология и геофизика. 2011. Т. 52. № 8. С. 1051–1074.
Arp G., Gropengiesser S. The Monotis–Dactylioceras Bed in the Posidonienschiefer Formation (Toarcian, southern Germany): condensed section, tempestite, or tsunami-generated deposit? // Paläontologische Zeitschrift. 2016. V. 90. P. 271–286. https://doi.org/10.1007/s12542-015-0271-7
Arp G., Gropengiesser S., Schulbert C., Jung D., Reimer A. Biostratigraphy and sequence stratigraphy of the Toarcian Ludwigskanal section (Franconian Alb, Southern Germany) // Zitteliana. 2021. V. 95. P. 57–94. https://doi.org/10.34194/geusb.v1.4646
Caswell B.A., Coe A.L., Cohen A.S. New range data for marine invertebrate species across the early Toarcian (Early Jurassic) mass extinction // J. Geol. Soc. 2009. V. 166 (5). P. 859–872. https://doi.org/10.1144/0016-76492008-0831
Cossman M. Description de quelques Pélécypodes jurassiques recueillis en France. Association française pour l’Avancement des Sciences. Congrès de Reims. 1908. P. 292–305.
Dumortier E. Etudes paleontologiques sur les depots jurassiques du bassin du Rhone. 3 Partie (Lias moyen). Paris, 1869. 348 p.
Goldfuss G.A. Petrefacta Germaniae tam ea, quae in museo universitatis regiae Borussicae Friedericiae Wilhelminae Rhenanae servantur quam alia quaecunque in museis Hoeninghusiano Muensteriano aliisque extant, iconibus et descriptionibus illustrata: Abbildungen und Beschreibungen der Petrefacten Deutschlands und der angränzenden Länder unter Mitwirkung des Herrn Grafen Georg zu Münster. Düsseldorf, 1835. V. 2. P. 69–140.
Hoffmann K., Martin G. Die Zone des Dactylioceras tenuicostatum (Toarcien, Lias) in NW- und SW-Deutschland // Paläontologische Zeitschrift, Stuttgart. 1960. V. 34. P. 103–149.
Howarth M. The Ammonite family Hildoceratidae in the Lower Jurassic of Britain // Monograph of the Palaeontographical Society. 1992. V. 145. № 586. P. 1–106.
Martindale R.C., Aberhan M. Response of macrobenthic communities to the Toarcian Oceanic Anoxic Event in northeastern Panthalassa (Ya Ha Tinda, Alberta, Canada) // Palaeogeogr. Palaeoclimat. Palaeoecol. 2017. V. 478. P. 103–120. https://doi.org/10.1016/j.palaeo.2017.01.009
Morris N.J., Knight R.I., Little C.S., Atkinson J.W. Mollusca – Bivalves. Fossils from the Lias of the Yorkshire Coast // Palaeontol. Assoc. Field Guide to Fossils. 2019. № 15. P. 105–157.
Münster G. Uber das geognostische Vorkommen einiger zu Monotis gehorenden Versteinerungen // Jahrbuch fur Mineralogie, Geognosie, Geologie und Petrefaktenkunde. Heidelberg, 1831. P. 403–408.
Muscente A.D., Martindale R.C., Schiffbauer J.D., Creighton A.L., Bogan B.A. Taphonomy of the Lower Jurassic konservat – lagerstaette at Ya Ha Tinda (Alberta, Canada) and its significance for exceptional fossil preservation during oceanic anoxic events // Palaios. 2019. V. 34. P. 515–541. https://doi.org/10.2110/palo.2019.050
Page K.N. The Lower Jurassic of Europe – its subdivision and correlation // The Jurassic of Denmark and Greenland. Eds. Ineson J., Surlyk F. Geol. Surv. Denmark Greenland Bull. 2003. V. 1. P. 23–59. https://doi.org/10.34194/geusb.v1.4646
Quenstedt F.A. Der Jura. Tubingen, 1858. 842 p.
Röhl H.-J. Hochauflösende palökölogische und sedimentologische Untersuchungen im Posidonienschiefer (Lias epsilon) von SW-Deutschland // Tübinger geowissenschaftliche Arbeiten. 1998. V. 47. P. 1–170.
Rogov M.A., Lutikov O.A. Dactylioceras–Meleagrinella (Clathrolima) assemblage from the Agardhbukta (eastern coast of Western Spitsbergen): a first in situ Toarcian molluscan occurrence from Svalbard providing interregional correlation // Norwegian J. Geol. 2022. V. 102. https://doi.org/10.17850/njg102-1-2
Rogov M., Ershova V., Vereshchagin O., Vasileva K., Mikhailova K., Krylov A. Database of global glendonite and ikaite records throughout the Phanerozoic // Earth System Science Data. 2021. V. 13. Iss. 2. P. 343–356. https://doi.org/10.5194/essd-13-343-2021
Schmidt-Rohl A., Rohl H.-J. Overgrowth on ammonite conchs: environmental implications for the Lower Toarcian Posidonia Shale // Palaeontology. 2003. V. 46. P. 339–352. https://doi.org/10.1111/1475-4983.00302
Szente I. Some macrofossils, chiefly bivalves, from the Lower Jurassic black shale of the Mecsek Mts (Hungary) // Hantkeniana. Budapest. 2015. № 10. P. 55–58.
Teichert S., Nützel A. Early Jurassic anoxia triggered the evolution of the oldest holoplanktonic gastropod Coelodiscus minutus by means of heterochrony // Acta Palaeontol. Polon. 2015. V. 60(2). P. 269–276. https://doi.org/10.4202/app.00145.2014
Treatise on Invertebrate Paleontology. Ed Moore R.C. Pt. N. Mollusca 6, Bivalvia. N.Y.: Geological Society of America, 1969. V. 1–3. 1224 p.
Urlichs M. Alter und Genese des Belemnitenschlachtfeldes im Toarcium von Franken // Geologische Blätter für Nordost-Bayern. 1971. № 21. P. 65–83.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция