

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 6, стр. 67-86
Изменения фаунистических и флористических ассоциаций Печорского Урала с конца среднего валдая до позднего голоцена
И. В. Кряжева 1, *, Д. В. Пономарев 1, Ю. В. Голубева 1
1 Институт геологии им. акад. Н.П. Юшкина Коми научного центра Уральского отделения
Российской академии наук
Сыктывкар, Россия
* E-mail: innageologi@mail.ru
Поступила в редакцию 11.02.2021
После доработки 02.02.2022
Принята к публикации 26.03.2022
- EDN: LNFIGZ
- DOI: 10.31857/S0869592X22060084
Аннотация
Изучены микротериофауна и палиноспектры двух местонахождений пещерного типа в логе Иорданского на Северном Урале (Печоро-Илычский биосферный заповедник): навеса Студеного и ниши Филина. Реконструированы условия накопления рыхлых отложений по палеофаунистическим и палинологическим данным. Видовые комплексы остатков мелких млекопитающих и спорово-пыльцевые спектры свидетельствуют о распространении кустарниковой заболоченной тундры с лесными островками в районе Верхней Печоры в конце среднего валдая (бызовской интерстадиал, МИС 3) и тундростепных ландшафтов в условиях прогрессирующей сухости климата в эпоху максимума последнего оледенения. В позднеледниковое время преобладали тундроподобные ерниковые заросли и климат был значительно холоднее современного. В фауне грызунов доминировали тундровые и тундростепные виды. В раннем голоцене отмечалось потепление климата, в связи с которым роль ксерофитных кустарников заметно снизилась, их место заняли березово-елово-сосновые леса. В фауне грызунов доля тундровых и тундростепных видов сократилась более чем в три раза. В позднем голоцене получили широкое распространение темнохвойные леса с примесью пихты и кедра. Ассоциации грызунов приобрели типичный лесной облик, где на долю обитателей леса приходилось более половины от всех остатков (60–80%), а тундровые виды отсутствовали.
ВВЕДЕНИЕ
Для реконструкции палеоклиматов и палеоландшафтов большое значение имеют как палинологические данные, так и анализ фауны мелких растительноядных млекопитающих, обитавших на территории изучаемого района. Это связано с приспособленностью грызунов к определенным природным зонам и биотопам, что позволяет реконструировать параметры окружающей среды. Массовые скопления костных остатков мелких млекопитающих, образовавшиеся в результате питания хищных птиц и четвероногих хищников, использующих карстовые образования в качестве убежищ и мест гнездования, являются одним из наиболее доступных и всеобъемлющих объектов для этой цели. Несмотря на существенную избирательность накопления костных остатков в таком типе захоронений, способность хищников тщательно облавливать территорию вокруг временного жилища/убежища позволяет точно воссоздать животное население прошлых лет района исследований.
Кроме того, пещерные отложения являются естественной ловушкой для спор и пыльцы растений, занесенных в пещеру на шерсти (перьях) и лапах животных, водой, просачивающейся с поверхности по трещинам горной породы, и ветром. Спорово-пыльцевые спектры из спелеогенных осадков более ценны для реконструкций палеосред, чем спектры из аллювиальных отложений, в которых содержание переотложенных зерен очень велико, что зачастую приводит к неверным выводам при воссоздании палеоландшафтов.
Несмотря на продолжительную историю изучения эволюции природной среды и климата в позднем плейстоцене и голоцене на обширной территории крайнего северо-востока Европы, в данном направлении исследований все еще остается много интересных нерешенных вопросов. Так, из отложений среднего валдая (= бызовского интерстадиала = МИС 3) описаны палинокомплексы, по которым реконструируются контрастные климатические события – потепления и похолодания (Дурягина, Коноваленко, 1993), однако хронометрические данные для подтверждения средневалдайского возраста палинокомплексов и привязки отдельных фаз динамики растительности пока получены только в единичных разрезах (Андреичева и др., 2015; Астахов, 2021). Другой проблемой является отсутствие до настоящего времени палинологических данных для реконструкции растительности последнего ледникового максимума в данном регионе. До сих пор основными источниками сведений для реконструкции природной среды максимума последнего оледенения были палеофаунистические материалы из местонахождений Пымвашор на Полярном Урале (Golovachov, Smirnov, 2009), Медвежьей пещеры и навеса Студеного на Северном Урале (Гуслицер и др.,1989; Кочев, 1993; Смирнов, 1996; Кряжева, Пономарев, 2019), а также геологические данные – наличие эоловых перигляциальных отложений в покровной формации плейстоцена региона (Astakhov, 2014).
В настоящей работе делается попытка реконструировать изменения климата и природной среды на западном склоне Северного Урала с конца бызовского (средневалдайского = МИС 3) времени до последнего ледникового максимума и с позднеледниковья до позднего голоцена по данным изучения спор и пыльцы, а также костных остатков мелких млекопитающих из отложений двух местонахождений пещерного типа – навеса Студеного и ниши Филина. Регион предгорий Урала (с севера на юг протягиваются Полярный, Приполярный и Северный Урал), где развиты карстующиеся породы и откуда известны пещерные местонахождения остатков позднеплейстоценовых и голоценовых млекопитающих, имеет большое значение для исследования четвертичной истории териофауны крайнего северо-востока Европы. Наиболее широкое распространение пещеры и другие карстовые образования имеют на Северном Урале в бассейне реки Печора. Самые известные и относительно крупные пещеры расположены в карстовом логу Иорданского: Медвежья, Туфовая, Ледяная и навес Студеный. Ранее в разрушенной пещере Студеной шурфовые работы проводились в 1961 и 1984 гг. параллельно с раскопками в Медвежьей пещере, которая многие годы служит опорным местонахождением при исследовании позднего плейстоцена Урала (Верещагин, Кузьмина, 1962; Гуслицер, Канивец, 1965; Гуслицер, Павлов, 1988; Гуслицер и др., 1989; Кузьмина, 1971; Кочев, 1991,1993; Смирнов, 1996).
Отложения навеса Студеного интересны тем, что они содержат самые древние (конец среднего валдая = бызовского интерстадиала = МИС 3) ископаемые комплексы пещерного тафономического типа в Приуральской Субарктике. Кроме того, здесь найдены также комплексы остатков микротериофауны последнего ледникового максимума (Смирнов, 1996), которые долгое время оставались самым достоверным источником сведений о характеристиках природной среды этого интервала на северо-востоке Европы при отсутствии надежно подтвержденных радиоуглеродным методом палинологических данных. В результате проведенных авторами полевых исследований в 2016 г. в логе Иорданского были получены новые остеологические и палинологические материалы из навеса Студеного и ниши Филина. Вместе с геохронометрическими данными о возрасте отложений навеса Студеного, появившимися в последние годы (Lagerholm et al., 2014; Palkopoulou et al., 2016), новые материалы позволяют впервые, причем не только для западного склона Северного Урала, но и для обширного региона – крайнего северо-востока Европы, реконструировать растительность последнего ледникового максимума по палинологическим данным, а также уточнить характеристики и возраст растительных ассоциаций конца среднего валдая. Кроме того, новые материалы дают возможность объединить палинологические и палеонтологические данные и провести комплексный анализ развития природной среды западного склона Северного Урала в конце среднего и позднем валдае, в позднеледниковье и голоцене.
ОПИСАНИЕ МЕСТОНАХОЖДЕНИЙ
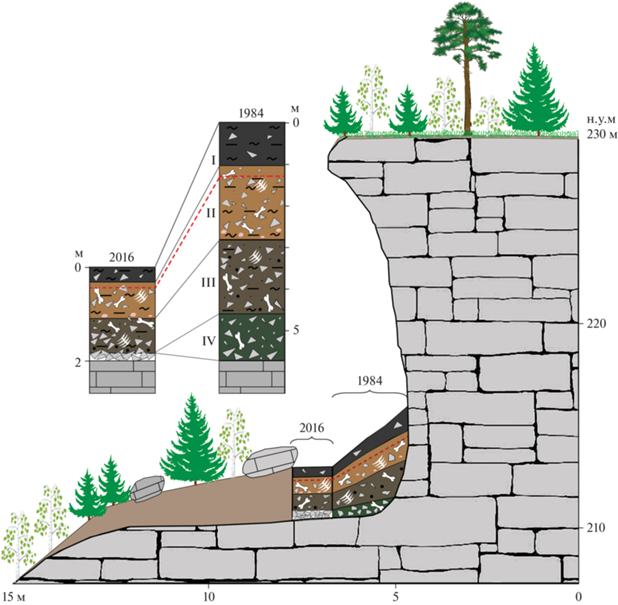
Навес Студеный расположен в предгорьях западного склона Северного Урала (62°00.766′ с.ш., 58°38.865′ в.д., 230 м над уровнем моря (н.у.м.)) на правом берегу р. Печора, в 17 км выше устья р. Большой Шежим, в 100 м к западу от приустьевой части лога Иорданского (рис. 1), на высоте 18–20 м над урезом реки и представляет собой крупный скальный навес высотой 14 м и шириной 30 м, предположительно оставшийся после обрушения свода крупной пещеры. В западной части навеса имеется хорошо сохранившийся шурф 1984 г., в котором максимальная мощность рыхлых отложений составляет более 5 м. Западная стенка этого шурфа примыкает к известняковому массиву, рыхлые отложения залегают под небольшим уклоном в юго-восточном направлении, площадь раскопа составляет около 3 м2. По описанию Б.И. Гуслицера и др. (1989) в разрезе было выделено 4 слоя (сверху вниз):
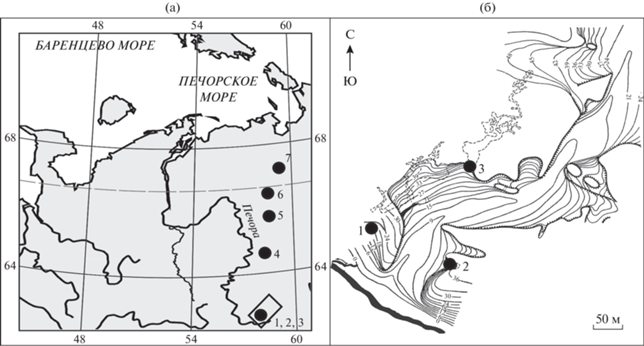
Рис. 1.
Географическое положение местонахождений. (а) – карта-схема, (б) – схема лога Иорданского (Гуслицер, Канивец, 1965). 1 – Навес Студеный, 2 – Ниша Филина, 3 – Медвежья пещера, 4 –Кожим-1, 5 –Щугер-4, 6 – Уса-2, 7 – Пымвашор.
Слой 1. Гумус с большим содержанием щебня известняка и растительных остатков, неслоистый. Мощность 1.2 м.
Слой 2. Суглинок бурый, с крупными глыбами и угловатым щебнем известняка. Мощность 1.6 м. На глубине 1.6 м от поверхности прослежен пятисантиметровый прослой, сложенный исключительно костями мелких млекопитающих.
Слой 3. Глина коричнево-серая, сильно алевритистая, с небольшим содержанием полуразложившегося щебня известняка, мелких угольков и железистых конкреций. Порода разбита ожелезненными трещинами. Мощность 1.6 м.
Слой 4. Щебень известняка с прослоями гумуса. Мощность 1.4 м.
В 2016 г. от восточной стенки данного раскопа был заложен шурф площадью 0.8 м2. Мощность четвертичных отложений составила менее 2 м (рис. 2). В разрезе рыхлых отложений навеса были вскрыты три слоя разного литологического состава, представленные песчано-глинистыми алевритами (сверху вниз):
Рис. 2.
Схематическое изображение навеса Студеного и разрезов рыхлых отложений в нем по данным раскопок 2016 г. и 1984 г. Нумерация слоев показана римскими цифрами. Костеносная прослойка выделена штрихпунктирной линией. I – черные гумусированные глинистые отложения, II – бурый глинистый алеврит с неокатанными обломками известняка, III – буро-серый песчано-глинистый алеврит, IV – щебень известняка с прослоями гумуса.
Слой 1. Черный гумусированный сильно глинистый алеврит (горно-лесной почвенный горизонт) с растительными остатками и щебенкой известняка. Мощность слоя 0.25 м. Граница с нижележащим слоем четкая по цвету.
Слой 2. Бурый глинистый алеврит с неокатанными обломками известняка. На глубине 0.4 м прослежена маломощная прослойка, состоящая практически полностью из костных остатков мелких млекопитающих. В основании слоя содержится небольшое количество обломков известняка со следами ожелезнения. Граница с нижележащим слоем нечеткая (по цвету). Мощность слоя 0.75 м.
Слой 3. Буро-серый песчано-глинистый алеврит с большим содержанием крупного щебня известняка. В основании слоя встречены мелкие угольки и неокатанные обломки сильно выветрелого кварца. Мощность слоя 0.8 м.
Вскрытые новым шурфом слои хорошо сопоставляются со слоями в раскопе 1984 г. по литологическим признакам. Строение разрезов в целом совпало, за исключением самой нижней части, которая отсутствует в новом раскопе. Различия в мощности отложений, скорее всего, связаны с неравномерным поступлением грубообломочного материала с обрушавшегося скального козырька навеса.
При определении возраста отложений навеса Студеного Б.И. Гуслицер с коллегами (Гуслицер и др., 1989) и В.А. Кочев (1993) опирались на методику, разработанную В.А. Кочевым (1984), которая заключалась в оценке эволюционного уровня первого и второго верхних коренных зубов копытных леммингов путем расчета показателя эволюционного уровня (ПЭУ). Они сравнивали ПЭУ копытных леммингов из отложений навеса Студеного и бурого суглинка Медвежьей пещеры. При этом радиоуглеродные датировки горизонта бурого суглинка (две для нижней части слоя (бурый суглинок Б) 16 300 ± 150 (ЛЕ-3060) и 17 980 ± ± 200 лет и одна для верхней части слоя (бурый суглинок А) 12 230 ± 100 лет (ЛЕ-3059) (Гуслицер, Павлов, 1988)) считались омоложенными. По их мнению, верхняя часть разреза (слой 2) имеет средневалдайский возраст и является одновозрастной с бурым суглинком Медвежьей пещеры на основании близости значений ПЭУ копытных леммингов из этих местонахождений. Возраст нижней части разреза Студеного (слой 3) принимался как ранневалдайский. Отсутствие более молодых валдайских отложений и залегание голоценовых осадков непосредственно на средневалдайских объяснялось солифлюкцией. В дальнейшем Н.Г. Смирновым (1996) было показано, что радиоуглеродные датировки отложений Медвежьей пещеры хорошо сопоставляются с особенностями фауны, климата и ландшафтов. По его мнению, возраст верхних горизонтов навеса Студеного (слой 2), где преобладают остатки копытного лемминга (криоксерофильного вида), – последний ледниковый максимум, а нижняя часть разреза (слой 3) с обилием остатков сибирского лемминга (криогидрофильного вида) отлагалась в конце раннего валдая–среднем валдае. В дальнейшем были сделаны новые радиоуглеродные датировки костей крупных млекопитающих из отложений Медвежьей пещеры, которые позволяют сопоставлять время формирования бурого суглинка с многочисленными остатками копытного лемминга с последним ледниковым максимумом (около 22 тыс. кал. лет назад; Ponomarev et al., 2013).
Несколько лет назад появились новые данные, позволяющие намного точнее оценивать возраст отложений навеса Студеного. Так, были опубликованы пять радиоуглеродных датировок остатков копытного лемминга из отложений навеса Студеного (Palkopoulou et al., 2016), варьирующих от 22 380 ± 170 лет (OxA-30034) до 24 790 ± 220 лет (OxA-30032), калиброванные значения которых находятся в диапазоне 26 430–29 070 лет. Кроме того, есть также оценки абсолютного возраста костных остатков сибирского лемминга из навеса Студеного – 30–35 тыс. календарных (кал.) лет, которые получены по степени расхождения генетических линий в программе BEAST (Lagerholm et al., 2014). Таким образом, фауна с преобладанием сибирского лемминга датируется концом бызовского интерстадиала, а фауна с доминированием копытного лемминга – последним ледниковым максимумом, как и предполагалось Н.Г. Смирновым (1996). Соответственно, в разрезе отсутствуют отложения позднеледниковья, которые были размыты во время завершающейся стадии дегляциации ледникового покрова. В целом данные оценки возраста горизонтов навеса Студеного находятся в соответствии с реконструкциями растительных ассоциаций Восточной Европы (Гричук, 1989; Дурягина, Коноваленко, 1993; Кольфсхотен и др., 2008; Андреичева и др., 2015) и фауны грызунов.
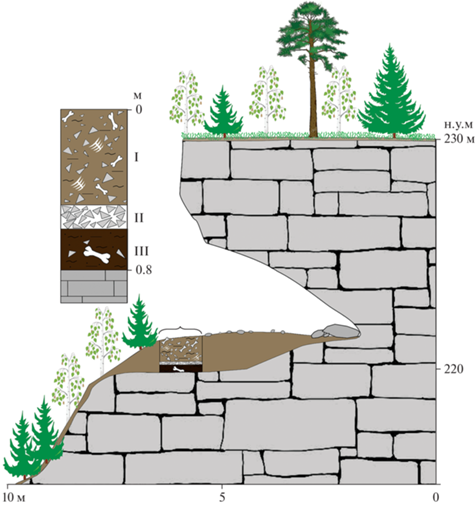
Местонахождение ниша Филина (62°00.714′ с.ш., 58°38.965′ в.д., 230 м н.у.м.) располагается в 100 м юго-восточнее навеса Студеного, в левом борту лога, в 55 м от его устья на высоте 30 м от уреза реки и представляет собой небольшую слабо выработанную нишу длиной 5 м и шириной 13.5 м (рис. 3). В разрезе рыхлых отложений ниши были вскрыты три слоя, представленных глинистыми алевритами и песчано-алевритовыми глинами (сверху вниз):
Рис. 3.
Схематическое изображение ниши Филина и разреза рыхлых отложений. Нумерация слоев показана римскими цифрами. I – коричневый глинистый алеврит, II –щебень известняка без заполнителя, III – темно-коричневая песчано-алевритовая глина.
Слой 1. Коричневый глинистый алеврит с большим содержанием крупного и мелкого щебня известняка. Мощность 0.55 м.
Слой 2. Щебень известняка без заполнителя, костных остатков не содержит. Мощность слоя 0.1 м.
Слой 3. Темно-коричневая песчано-алевритовая глина с мелкой щебенкой известняка. Мощность 0.15 м.
О возрасте горизонтов рыхлых отложений ниши Филина можно судить по положению в разрезе, составу и структуре фауны. Мы предполагаем, что слой 3 сформировался в один из интервалов позднеледниковья (или поздних этапов дегляциации), а слой 1 – в голоцене. Накопление костных остатков и растительности было прервано сильным отслоением потолка грота, скорее всего связанным с потепление климата и повышением влажности, в результате чего образовался плотный стерильный слой из щебня и крошки известняка без палеонтологических и палинологических остатков (слой 2).
В комплексе остатков слоя 3 преобладают три тундровых вида (копытный и сибирский лемминги с узкочерепной полевкой) при почти равном соотношении долей остатков этих видов между собой. Схожая структура фауны, в которой наблюдается содоминирование остатков тундровых видов, известна в датированных по 14C комплексах из горизонта А бурого суглинка Медвежьей пещеры, расположенной в 150 м к северу от ниши Филина, а также в слоях 2 и 3 грота Щугер-4, который находится в 250 км к северу на Приполярном Урале (Кряжева и др., 2012).
Комплекс остатков из слоя 1 ниши Филина условно поделен на верхнюю и нижнюю части. Фауна грызунов из верхней части состоит из видов, типичных для современной таежной фауны Северного Урала, а в фауне из нижней части помимо таежных элементов фауны присутствует значительное количество тундровых и тундростепных видов. Исходя из данных по составу и структуре фауны и по положению в разрезе, предполагается, что верхняя часть слоя 1 сформировалась в позднем голоцене, а нижняя – вероятно, в раннем голоцене.
Тафономические особенности. Кости черепа и посткраниального скелета крупных млекопитающих (Lepus timidus L, 1758, Mustela sp., Vulpes sp., Ursus sp., Rangifer tarandus L, 1758) из всех местонахождений сильно фрагментированы, некоторые имеют следы пищеварительной коррозии. Пригодными для видового и родового определения костных остатков крупных млекопитающих были только изолированные зубы, фрагменты нижних челюстей и фрагменты лицевой части черепа с зубами. Такой характер фрагментации костных остатков указывает на их накопление в результате деятельности хищных млекопитающих, которые использовали местонахождение в качестве логова.
Ископаемые остатки насекомоядных млекопитающих (Eulipotyphla) представлены фрагментами нижней и верхней челюсти и изолированными зубами. Ископаемые остатки грызунов являются самыми многочисленными в отложениях и представлены всеми частями скелета.
Для идентификации таксономической принадлежности ископаемых костных остатков видов отрядов Rodentia и Lagomorpha (семейство Ochotonidae) было использовано 9994 нижних и верхних щечных зуба, по которым было определенно 16 видов (табл. 1): красно-серая полевка (Craseomys rufocanus Sundervall, 1846), рыжая полевка (Clethrionomys glareolus Schreber, 1780), красная полевка (Clethrionomys rutilus Pallas, 1779), копытный (Dicrostonyx gulielmi Sanford, 1870) и сибирский (Lemmus sibiricus Kerr, 1792) лемминги, узкочерепная полевка (Lasiopodomys gregalis Pallas, 1779), пищуха (Ochotona pusilla Pallas, 1768), водяная полевка (Arvicola amphibius Linnaeus, 1758), полевка-экономка (Alexandromys oeconomus Pallas, 1778), полевка Миддендорфа (Alexandromys middendorffii Poljakov, 1881), темная полевка (Microtus agrestis Linnaeus, 1761), белка (Sciurus vulgaris Linnaeus, 1758), бурундук (Tamias sibiricus Laxmann, 1769), летяга (Pteromys volans Linnaeus, 1758), лесная мышовка (Sicista betulina Pallas, 1778), лесной лемминг (Myopus schisticolor Lilljeborg, 1844).
Таблица 1.
Соотношение остатков и общее число зубов мелких растительноядных млекопитающих (%) из местонахождений лога Иорданского
| Местонахождение | Навес Студеный | Навес СтуденыйГ | Медвежья пещера | Ниша Филина | Биом | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Таксон |
Слой | ||||||||||||||||||
| Сл.1 | Сл.2 в | Сл.2 н | Сл.3 | Сл.2 в | Сл.2 н | Сл.3 | Сл.4 | Зел. супесь | Гум. почва | Бур. сугл. А | Бур. сугл. Б | Сл.1 в | Сл.1 н | Сл.3 | |||||
| с | т | л | и | ||||||||||||||||
| Ochotona pusilla – степная пищуха | – | – | – | – | 1 | – | – | 24 | ед. | ед. | ед. | ед. | – | – | 1 | + | – | – | – |
| Pteromys volans – обыкновенная летяга | – | – | – | – | – | – | – | – | – | – | – | – | 0.5 | – | – | – | – | + | – |
| Sciurus vulgaris – обыкновенная белка | ед. | – | – | – | – | – | – | – | 0.8 | – | – | – | 36.8 | 0.7 | – | – | – | + | – |
| Tamias sibiricus – азиатский бурундук | ед. | – | – | – | – | – | – | – | – | – | – | – | 2.5 | – | – | – | – | + | – |
| Sicista betulina – лесная мышовка | – | – | – | – | – | – | – | – | – | – | – | – | 0.2 | – | – | – | – | + | – |
| Craseomys rufocanus – красно-серая полевка | ед. | – | – | 0.9 | – | 1 | 1 | 5 | 11.4 | 16.7 | 1 | 0.3 | 4.5 | 7 | 3 | – | – | + | – |
| Clethrionomys glareolus – рыжая полевка | – | – | 2.1 | 17.1 | 16.7 | 1.5 | 0.5 | 1.7 | 10.3 | – | – | – | + | – | |||||
| Clethrionomys rutilus – красная полевка | – | – | 3 | 2 | 11.7 | 1.5 | – | – | + | – | |||||||||
| Dicrostonyx gulielmi – копытный лемминг | – | 70.3 | 51.9 | 2.9 | 63 | 50 | 11 | 30 | 8.9 | 8.3 | 17 | 73.9 | – | – | 28.5 | – | + | – | – |
| Lemmus sibiricus – сибирский лемминг | – | 9.9 | 26.9 | 69.3 | 11 | 24 | 71 | 9 | 19.5 | 16.7 | 30 | 10.4 | – | 7.9 | 22.3 | – | + | – | – |
| Myopus schisticolor – лесной лемминг | – | – | – | – | – | – | – | – | 4.9 | – | 1 | 3.5 | 0.7 | 3 | – | – | – | + | – |
| Arvicola amphibius – водяная полевка | ед. | – | – | 2.1 | – | – | – | 4 | 8.1 | – | 1 | – | 38.8 | 9.4 | 8 | – | – | – | + |
| Microtus agrestis – темная полевка | ед. | 0.1 | – | 3.8 | – | – | 2 | 5 | 9.8 | 8.3 | 7 | – | 8.2 | 22.8 | 10.7 | – | – | + | – |
| Lasiopodomys gregalis – узкочерепная полевка | – | 18.8 | 20.9 | 1 | 21 | 22 | 7 | 4 | 13 | 20.8 | 32.5 | 13.7 | – | 15.8 | 25 | + | + | – | – |
| Alexandromys middendorffii – полевка Миддендорфа | – | – | – | 1 | – | – | – | 6 | – | – | – | 0.3 | – | – | – | – | + | – | – |
| Alexandromys oeconomus – полевка-экономка | ед. | 0.9 | 0.3 | 13.8 | 3 | 2 | 7 | 10 | 6.5 | 12.5 | 6.5 | 0 | 4.1 | 11.4 | – | – | – | – | + |
| Всего | – | 2339 | 1039 | 660 | 7978 | 1572 | 849 | 48 | 62ос | 12ос | 100ос | 197ос | 4910 | 710 | 336 | ||||
Коллекции ископаемых остатков грызунов хранятся в Институте геологии Коми НЦ УрО РАН.
Сведения об экологии видов грызунов. Для детального анализа динамики фауны необходимо классифицировать виды грызунов в соответствии с их биологическими и экологическими требованиями: биотопами, климатическими предпочтениями (температура и влажность) и их распространением в современных природных зонах. Эта информация взята из различных источников (Огнев, 1950; Громов, Поляков, 1977; Громов, Ербаева, 1995; Ануфриев, 1994a, 1994б, 1994в; Ануфриев, Бобрецов, 1994; Петров, 1994a, 1994б, 1994в, 2007; Пыстин, 1994).
Исходя из основных климатических факторов, таких как температура и влажность, грызуны объединены следующим образом: 1) криоксерофильные виды – копытный лемминг (Dicrostonyx); 2) мезотермно-ксерофильные – узкочерепная полевка (L. gregalis) и степная пищуха (Ochotona pusilla); 3) мезофильные виды, предпочитающие умеренные условия, – виды Craseomys и Clethrionomys, темная полевка (Microtus agrestis), белка (Sciurus vulgaris), бурундук (Tamias sibiricus), летяга (Pteromys volans) и лесная мышовка (Sicista betulina); 4) мезотермно-гидрофильные полевка-экономка (Alexandromys oeconomus) и лесной лемминг (Myopus schisticolor); 5) эвритермно-гидрофильный вид – водяная полевка (Arvicola amphibius); 6) криогидрофильные виды – сибирский лемминг (Lemmus sibiricus) и полевка Миддендорфа (Alexandromys middendorffii).
Одно из важнейших экологических свойств – это предпочитаемый биотоп. Грызуны предпочитают определенные биотопы, и по этому параметру их можно разделить на шесть групп: 1) типичные тундровые биотопы, за исключением влажных местообитаний – копытный лемминг (Dicrostonyx) и частично узкочерепная полевка (L. gregalis), которая также обильна на лугах, в кустарниках и поймах рек; 2) влажные тундровые биотопы – сибирский лемминг (Lemmus sibiricus) и полевка Миддендорфа (Alexandromys middendorffii); 3) полуводные (прибрежные) местообитания – водяная полевка (Arvicola amphibius); 4) разные лесные местообитания – все полевки Craseomys-Clethrionomys, белка, бурундук, летяга и лесная мышовка; 5) различные лесные и луговые местообитания – темная полевка (Microtus agrestis); 6) разные степные биотопы – степная пищуха (Ochotona pusilla). В тундре темная полевка обитает на участках с ивами, в кустарниковой тундре и на поймах рек. Водяная полевка Arvicola заселяет прибрежные местообитания в основном летом, а зимой занимает возвышенности вдали от водоемов. Полевка-экономка (Alexandromys oeconomus) в какой-то мере также может быть отнесена к третьей группе, так как предпочитает кустарниковую высокопродуктивную пышную прибрежную растительность по берегам небольших рек и родников. Лесная мышовка обитает в насаждениях с богатым травяным ярусом, а белке и летяге необходимы леса с сомкнутым древостоем.
По предпочтительной природной зоне (степь, тундра, лес) все выявленные виды были сгруппированы следующим образом: степные зональные виды (пищуха), тундро-степные зональные виды (узкочерепная полевка), тундровые зональные виды (копытный и сибирский лемминги, полевка Миддендорфа), лесные зональные виды (белка, бурундук, лесной лемминг, красная, красно-серая, рыжая и темная полевки, лесная мышовка, летяга) и интразональные виды (полевка-экономка, водяная полевка). Такие виды, как лесные полевки (Craseomys–Clethrionomys), темная полевка, лесная мышовка проникают в тундровую зону, а красная полевка даже достигает там высокой численности (Петров, 2007).
МЕТОДЫ ИССЛЕДОВАНИЙ
Рыхлые отложения вскрывали условными горизонтами мощностью не более 10 см. Извлечение костных остатков животных производилось при помощи ручной промывки вмещающей породы на ситах с размером ячеи 0.8 мм. Полученную смесь костей и обломков породы высушивали, и из нее вручную отбирали остатки позвоночных.
Диагностика морфологически сходных видов полевковых, таких как полевка Миддендорфа–темная полевка и сибирский лемминг–лесной лемминг, проводилась по методике Н.Г. Смирнова с соавторами (Смирнов и др., 1997). Видовую принадлежность остатков узкочерепной полевки, полевки Миддендорфа, полевки экономки и темной полевки определяли по первому нижнему коренному зубу. Все остальные моляры группы родов Microtus распределяли по видам в соответствии с распределением первых нижних коренных. Видовую принадлежность лесных полевок (род Clethrionomys) проводили по методике А.В. Бородина с соавторами (Бородин и др., 2005).
Для исследования эволюционных морфологических особенностей моляров копытного лемминга и узкочерепной полевки из навеса Студеного зубы из условных горизонтов объединили в несколько групп, чтобы получить достаточное количество материала для анализа. У копытного лемминга изучали форму первого и второго коренных зубов по методике, предложенной Н.Г. Смирновым с соавторами (1997) с небольшими изменениями (Ponomarev, Puzachenko, 2015). По строению М1 и М2 выделяли три морфотипа – simplicior, henseli и torquatus, а далее по соотношению этих морфотипов выборки относили к одной из стадий развития зубной системы, сменяющим друг друга по степени сложности – уменьшению доли “архаичных” (simplicior), возрастанию доли промежуточных (henseli) и “продвинутых” (torquatus) морфотипов. Все изученные выборки относили к одному из хроновидов: Dicrostonyx simplicior, D. gulielmi или D. torquatus. В пределах каждого вида были выделены по три морфы, а кроме видов различались промежуточные, переходные формы неуказанного таксономического ранга (Смирнов др., 1997).
У узкочерепной полевки исследовали форму передней непарной петли (параконида) и общие размеры первого нижнего коренного зуба (Большаков и др., 1980; Головачев и др., 2001; Ponomarev, Puzachenko, 2017). Эволюционный уровень полевок оценивается по соотношению морфотипов, со временем увеличивается доля продвинутых, сложных зубов (грегалоидно-микротидный и микротидный морфотипы) и уменьшается относительное количество архаичных, простых моляров (грегалоидный морфотип).
В работе используется систематика полевковых, предложенная Н.И. Абрамсон и А.А. Лисовским (2012), за исключением красной и рыжей полевок, для которых валидным названием следует считать Clethrionomys (Kryštufek et al., 2020).
Образцы для спорово-пыльцевого анализа отбирали из зачищенной стенки обнажений сплошной колонкой от скального ложа до поверхности с учетом изменения литологических особенностей отложений. Лабораторную обработку и анализ образцов под микроскопом проводили по стандартным методикам (Гричук, Заклинская, 1948). Палиноморфы изучались под микроскопом Motic BA210 при увеличении ×400. При определениях использовались опубликованные определители и атласы спор и пыльцы, базы данных на сайтах и коллекция спор и пыльцы современных травянистых растений западного склона Северного Урала (Институт геологии ФИЦ Коми НЦ УрО РАН). Пыльца и споры определялись до семейства, рода, в оптимальном случае – до вида. Пыльцевые зерна рода Betula подразделялись на три секции в зависимости от типа поровой камеры (Ошуркова, 1959). Секция Betula sect. Albae содержит пыльцу древесных видов, к секциям Betula sect. Fruticosae и Betula sect. Nanae отнесена пыльца кустарниковых видов. Спорово-пыльцевые диаграммы построены с помощью компьютерной программы TILIA (Grimm, 1990). Процентные соотношения спор и пыльцы были подсчитаны групповым способом.
Временные интервалы, используемые в работе (кал. лет): 1) окончание среднего валдая (бызовское время), 35 000–28 000; 2) последний ледниковый максимум, понимаемый в широком смысле от начала МИС 2 до начала периода дегляциации 29 000–20 000; 3) позднеледниковье (в широком смысле) или период дегляциации, 20 000–11 700, 4) ранний голоцен, 11700–8200; 5) средний голоцен, 8200–4200; 6) поздний голоцен, 4200–современность.
Радиоуглеродные датировки калибровали с использованием рекомендованной калибровочной кривой IntCal20 (Reimer et al., 2020).
РЕЗУЛЬТАТЫ
Фаунистические данные
В нижней части навеса Студеного (слой 3) микротериофауна представлена сибирским леммингом (69.3%), полевкой-экономкой (13.8%), лесными полевками (6%), копытным леммингом (2.9%), темной полевкой (3.8%), водяной полевкой (2.1%), полевкой Миддендорфа (1%), узкочерепной полевкой (1%). Состав комплекса остатков грызунов слоя 2 обеднен. В нижней части слоя 2 (0.6–1 м) найдены копытный лемминг (51.9%), сибирский лемминг (26.9%), узкочерепная полевка (20.9%) и полевка-экономка (0.3%), а в верхней части этого слоя (0.25–0.6 м) обнаружены копытный лемминг (70.3%), сибирский лемминг (9.9%), узкочерепная полевка (18.8%), полевка-экономка (0.9%) и темная полевка (0.1%).
Микротериофауна слоя 3 ниши Филина на 76.8% представлена тундровыми и степными видами: копытным леммингом (28.5%), узкочерепной полевкой (25%), сибирским леммингом (22.3%), пищухой (1%), на 15.2% лесными видами – темной полевкой (10.7%), лесными полевками (4.5%) и на 8% интразональным видом – водяной полевкой (8%). Микротериофауна в нижней части слоя 1 (0.4–0.55 м) состоит на 55.5% из лесных видов: лесные полевки (29%), темная полевка (22.8%), лесной лемминг (3%) и белка (единичные остатки), на 23.7% из тундровых (сибирский лемминг – 7.9%) и тундростепных (узкочерепная полевка – 15.8%) видов и на 20.8% из интразональных видов (полевка-экономка – 11.4%, водяная полевка – 9.4%). В верхней части слоя 1 ниши Филина (0–0.4 м) фауна грызунов представлена лесными видами: летяга (0.5%), белка (36.8%), бурундук (2.5%), лесная мышовка (0.2%), лесные полевки (8.2%), лесной лемминг (0.7%) и темная полевка (8.2%) – и интразональными видами: водяная полевка (38.8%) и полевка-экономка (4.1%).
Особенности морфологии копытного лемминга и узкочерепной полевки
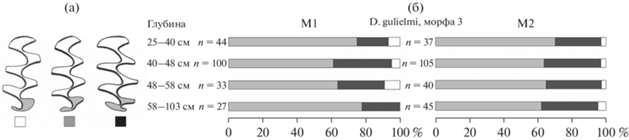
По своему эволюционному уровню остатки копытных леммингов из слоя 2 (0.25–1.0 м) навеса Студеного, датируемые последним ледниковым максимумом, сопоставляются с одной стадией развития зубной системы, представленной морфой 3 D. gulielmi. Выборки зубов из отдельных условных горизонтов внутри слоя 2 не показывают различий в морфологии (рис. 4). Для этой морфы характерно преобладание моляров морфотипа henseli, на втором месте torquatus, а на третьем – simplicior на обоих зубах (рис. 4). Животные, относящиеся к этой эволюционной стадии, обитали в регионе примерно от 30 до 11 кал. тыс. лет назад, т.е. от конца бызовского времени до начала голоцена (Ponomarev, Puzachenko, 2015). К этой же стадии относятся лемминги из таких местонахождений, как Пижма-1, Щугер-4 (слои 1 и 2), Уньинская пещера, Серчейю-6, Рябово и Нижний Двойник.
Рис. 4.
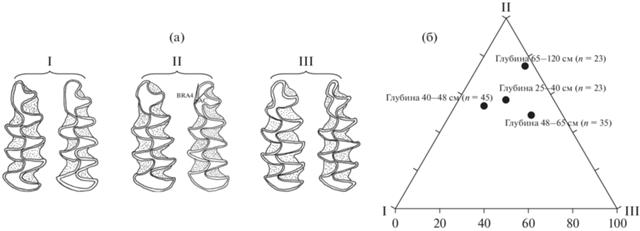
Морфотипы моляров копытных леммингов (а) и их соотношение среди М1 и М2 у леммингов из отложений навеса Студеного (б). Белым цветом показан морфотип simplicior, серым – henseli, черным – torquatus.
Изменчивость размерных и морфотипических характеристик первого нижнего коренного зуба узкочерепной полевки из отложений навеса Студеного по отдельным условным горизонтам сравнительно низкая (рис. 5). Средние значения размеров m1 в разных условных горизонтах разреза навеса Студеного: длина 2.53–2.63 мм, ширина 0.93–0.98 мм. При этом средние размеры m1 современных узкочерепных полевок трех выборок с северо-востока европейской части России: длина от 2.64 до 2.68 мм, ширина от 1.0 до 1.02 мм, а средние размеры m1 позднеплейстоценовых узкочерепных полевок варьируют от 2.52 до 2.81 мм в длину и от 0.9 до 1.0 мм в ширину (Ponomarev, Puzachenko, 2017). Узкочерепные полевки из навеса Студеного по морфологии m1 схожи с другими поздненеоплейстоценовыми полевками региона (Ponomarev, Puzachenko, 2017). Их отличительными чертами являются относительно мелкие зубы со сравнительно архаичной формой жевательной поверхности – среди моляров преобладают экземпляры грегалоидно-микротидного морфотипа, а доля моляров микротидной морфы заметно меньше, хотя размах изменчивости этих параметров как в современных, так и в позднеплейстоценовых выборках очень велик (Ponomarev, Puzachenko, 2017).
Рис. 5.
Морфотипы m1 узкочерепной полевки (а) и их соотношение у полевок из отложений навеса Студеного (б). Показана схема измерения угла BRA4 на антерокониде (AC) m1 (Смирнов и др., 1990; Головачев и др., 2001). BRA – buccalre-entrantangle (буккальный входящий угол), AC – anteriorcap (передняя непарная петля) (Meulen, 1973). I – грегалоидный морфотип; II –грегалоидно-микротидный морфотип; III – микротидный морфотип.
Палинологическая характеристика
По данным палинологического изучения рыхлых отложений навеса Студеного выделены три палинозоны (ПЗ) (рис. 6).
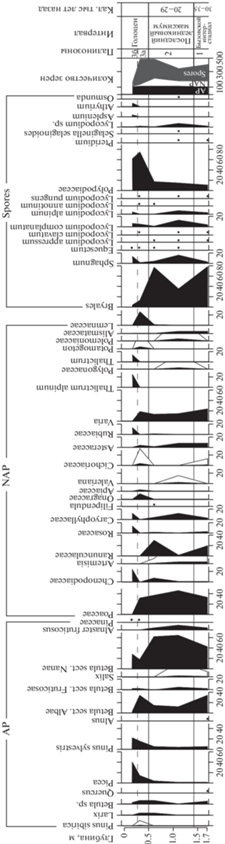
ПЗ 1 (слой 3, гл. 1.7 м). В общем составе спорово-пыльцевого спектра (СПС) преобладают споры (54%). На долю пыльцы древесных и травянистых растений приходится 26 и 20% соответственно. Среди пыльцы древесных доминирует пыльца берез (Betula sect. Nanae – 44% и Betula sect. Albae – 35%). Группу пыльцы травянистых растений образуют преимущественно злаки (33%) и разнотравье (58%) из лютиковых, астровых, розоцветных, гвоздичных и цикориевых. Встречена пыльца ксерофильных трав (Artemisia sp. – 4%, Chenopodiaceae – единичные зерна). Из спор наряду с преобладающими зелеными мхами (78%) присутствуют папоротники (11%), а также в единичном количестве сфагновые мхи, таежные (Lycopodium clavatum, L. complanatum) и тундровые аркто-альпийские виды (L. alpinum, L. appressum, L. pungens) плаунов.
ПЗ 2 (слой 2, гл. 1.1–0.6 м) выделена на основании увеличения содержания пыльцы тундрового кустарника карликовой березы до 65% и сокращения участия пыльцы древовидной березы (12–14%). В группе трав продолжает доминировать пыльца злаков (38–46%) и разнотравья (46–53%), среди которого появляются Rubiaceae, Filipendula sp., Valeriana sp., Polygonaceae и Polemoniaceae. Отличительной особенностью группы споровых является присутствие единичных спор Osmunda sp.
ПЗ 3 (слой 1, гл. 0.3–0.15 м) подразделяется на две подзоны. Подзона 3а характеризуется уменьшением содержания пыльцы трав (7%) при увеличении количества спор (до 70%). Пыльца деревьев и кустарников составляет 23% и в основном представлена Betula sect. Albae (35%), Pinus sylvestris (16%) и Picea sp. (15%). Встречены единичные зерна Pinus sibirica. Содержание пыльцы криофильных кустарников в СПС, напротив, значительно сокращается до 18%. Среди пыльцы травянистых растений по-прежнему доминирует пыльца злаков и разнотравья. Присутствие пыльцы водных растений (рдеста, ряски) указывает на существование в окрестностях навеса Студеного водоемов со стоячей или медленнотекущей водой. В группе спор резко увеличивается содержание спор папоротников за счет сокращения спор зеленых мхов. Подзона 3б отличается от предыдущей подзоны значительным увеличением доли пыльцы древесной растительности (до 65%) и сокращением участия спор (до 30%). В группе пыльцы древесных растений максимальных значений достигает количество пыльцы хвойных пород (Pinus sylvestris – 22% и Picea sp. – 41%). Существенную долю составляет пыльца Betula sect. Nanae (29%), что указывает на распространение ерниковых зарослей, приуроченных к заболоченным участкам. В группе пыльцы травянистых растений обнаружены незначительные содержания или единичные зерна маревых, розоцветных, гвоздичных, мареновых и василистника альпийского, а злаки и водные растения исчезают. В составе спор сохраняется доминирование папоротников (62%), среди которых появляются кочедыжник и костенец.
По результатам палинологических исследований рыхлых отложений ниши Филина на диаграмме выделены две палинозоны (рис. 7).
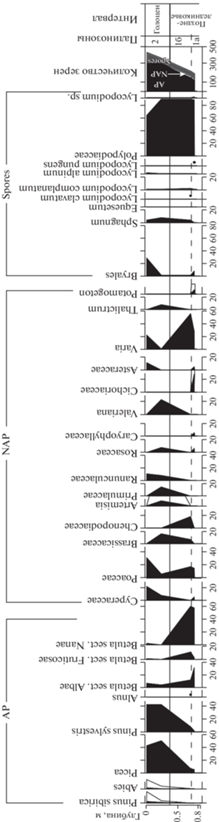
ПЗ 1 (слой 3, гл. 0.8–0.65 м). В общем составе СПС распределение между тремя группами растений следующее: древесно-кустарниковая группа – 61–68%, группа трав – 7–10%, споры – 25–28%. В первой группе доминирует пыльца кустарников Betula sect. Nanae (58–61%), Betula sect. Fruticosae (до 11%), встречаются единичные зерна Alnaster fruticosus. Пыльца древесных пород составляет меньшую долю: Betula sect. Albae –13–31%, Picea sp. – 5–7% и Pinus sylvestris – ед. зерна. В составе пыльцы травянистых растений преобладают Poaceae (17–19%) и разнотравье (62–72%) из Cichoriaceae (27%), Asteraceae, Brassicaceae, Rosaceae и Caryophyllaceae (по 6%). Среди споровых доминирует Polypodiaceae (88–91%). Также присутствуют споры зеленых мхов (2–7%) и плаунов (ед. зерна Lycopodium complanatum, L. clavatum и L. pungens). Подразделение палинозоны на две подзоны проведено на основании следующих изменений в составе СПС вверх по разрезу: сокращения участия древовидной березы и обеднения состава пыльцы трав. К тому же отличительной особенностью для подзоны 1а является пик пыльцы цикориевых, характерных для слабо задернованных грунтов.
В составе СПС ПЗ 2 (слой 1, гл. 0.3–0 м) сохраняются пропорции групп спор и пыльцы, но происходят значительные изменения процентных содержаний внутри групп. Намечается четкая тенденция к увеличению содержания пыльцы деревьев Picea sp. (43–51%), Pinus sylvestris (43–44%), Betula sect. Albae (4–7%), появляются единичные зерна Abies sp. и Pinus sibirica. Состав пыльцы трав и спор становится разнообразнее. Отличия касаются появления в группе трав пыльцы Brassicaceae, Rosaceae, Primulaceae и др., а среди спор – Equisetum sp. и Sphagnum sp.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Конец бызовского интерстадиала (среднего валдая)
Самый древний комплекс микромаммалий, датируемый концом бызовского (средневалдайского) интерстадиала (35–30 тыс. кал. лет назад), найден в нижней части разреза навеса Студеного (слой 3). Фауна на 74% состоит из обитателей тундры, на 16% из интразональных видов, и 10% приходится на лесных полевок (рис. 8). Доминирование в составе фауны криогидрофильного вида – сибирского лемминга, предпочитающего увлажненные тундры с обилием мхов, свидетельствует о преобладании ландшафтов влажной моховой тундры, а присутствие лесных полевок говорит о наличии лесных островков или галерейных лесов в верховьях Печоры.
Палинологические спектры из этой части разреза указывают на произрастание в данном районе березовых редколесий по кустарниковой тундре. Наряду с кустарниковыми зарослями из березы, ольховника и ивы, повсеместное распространение также имели злаковые и маревые. Среди споровых растений доминировали зеленые мхи. Спектр отражает лесотундровый тип растительности и климат значительно холоднее, чем в настоящее время.
Климат среднего валдая был значительно холоднее, чем в более ранние межледниковые эпохи, например во время сулинского (микулинского) межледниковья. Для европейской части России в бызовское (средневалдайское) время выделяют несколько фаз изменения растительности. На северо-западе и северо-востоке в периоды потепления отмечалось развитие елово-березовых, елово-сосновых и березовых лесных формаций с участием широколиственных пород, а холодные периоды характеризовались развитием березовых редколесий с элементами ксерофитной перигляциальной флоры. В северо-восточной части наряду с ксерофитными ассоциациями были широко распространены болотно-тундровые формации, которые на северо-западе появились только в завершающей стадии средневалдайского интерстадиала. Также для северо-востока отмечается отсутствие климатических оптимумов, которые зафиксированы в северо-западной части (Арсланов и др., 1981; Спиридонова, 1983; Дурягина, Коноваленко, 1993; Марченко-Вагапова, 2012; Андреичева и др., 2015).
Последний ледниковый максимум
Фауна грызунов из слоя 2 навеса Студеного на 99% состоит из тундровых и тундростепных видов. От основания слоя вверх по разрезу наблюдается заметное изменение в соотношении остатков представленных видов (снизу вверх): доля копытного лемминга увеличивается с 52 до 70%, в то время как доля сибирского лемминга, наоборот, уменьшается с 27 до 10%. Доля остатков зубов узкочерепной полевки остается значительной (21 и 22%).
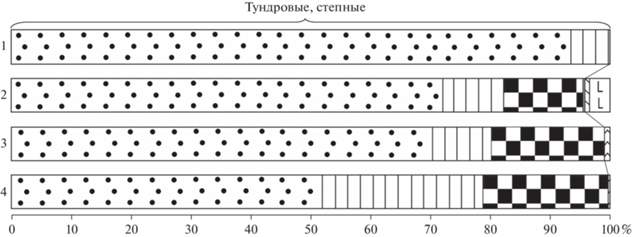
Фауна верхней части слоя 2 по составу и структуре схожа с фауной из бурого суглинка Б Медвежьей пещеры (рис. 9), которая датируется максимальной холодной стадией позднего валдая (примерно 22 тыс. кал. лет назад). Здесь на копытного лемминга приходится 74%, на узкочерепную полевку – 14%, на сибирского лемминга – 10%, на полевку Миддендорфа – менее 1% и на лесные виды – менее 2% (табл. 1) (Смирнов, 1996). Похожая картина наблюдается и в слое 6 Пымвашора (рис. 9), расположенного в 600 км к северу от лога Иорданского. Возрастной диапазон разных горизонтов слоя 6 варьирует от 13 до 21 14С тыс. лет (Смирнов и др., 1999; Svendsen et al., 2010), а по последним опубликованным радиоуглеродным датировкам по костям копытного лемминга варьирует от 15408–15946 до 25707–26807 кал. лет назад (Palkopoulou et al., 2016). Здесь на копытного лемминга приходится 89%, на сибирского лемминга – 9% и на узкочерепную полевку – 1%. Несмотря на то что местонахождение расположено в заполярном районе, где и в настоящее время также доминируют тундровые виды (неопубликованные данные сборов погадок в 1979 г. Б.И. Гуслицера и В.И. Кочева на р. Адьзва и сборы И.В. Кряжевой в урочище Пымвашор в 2018 г.), в заключительную фазу валдайского оледенения микротериофауна западного склона Северного Урала и Большеземельской тундры имели общие черты. Качественные и количественные характеристики комплексов сравниваемых местонахождений схожи. Во всех случаях остатки копытных леммингов резко доминируют, причем зубы из всех сравниваемых выборок относятся к одной стадии развития зубной системы, представленной морфой 3 Dicrostonyx gulielmi (Смирнов и др., 1999; Ponomarev, Puzachenko, 2015; Кряжева, Пономарев, 2019). Животные, относящиеся к этой эволюционной стадии, обитали в регионе примерно от 30 до 11 кал. тыс. лет назад. На втором месте по обилию остатков в отложениях Северного Урала находится узкочерепная полевка. Это животное практически индифферентно к низким температурам, однако предпочитает достаточно аридные условия и в настоящее время является обычным обитателем степей и тундр, причем в тундровой зоне она избирательно заселяет сухие местообитания. Высокая доля обитателей сухих тундр указывает на более холодный и сухой климат в верховьях Печоры во время седиментации этой части слоя 2 по сравнению с условиями формирования нижележащего слоя 3 навеса Студеного, где 69% остатков принадлежат сибирскому леммингу – обитателю заболоченных тундр.
Рис. 9.
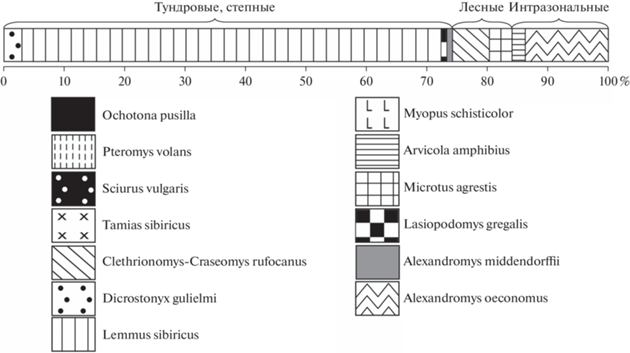
Состав фауны грызунов (%) последнего ледникового максимума (LGM) из местонахождений Тимано-Североуральского региона. 1 – Пымвашор (слой 6), 2 – Медвежья пещера (бурый суглинок Б), 3 – Навес Студеный (слой 2, верхняя часть), 4 – Навес Студеный (слой 2, нижняя часть). Условные обозначения см. рис. 8.
Фауна грызунов из нижней части слоя 2 отражает переходную фазу развития природной среды от умеренно влажных холодных условий завершающейся стадии бызовского интерстадиала к более суровым – холодным и аридным условиям максимума последнего оледенения. Это выразилось в постепенной смене вида доминанта с сибирского лемминга на копытного, при этом общая доля тундровых и тундростепных видов в фауне грызунов не изменилась. Из чего можно заключить, что климат с окончания среднего валдая и на протяжении последнего ледникового максимума был холодным, меняясь от холодного влажного в конце бызовского интерстадиала к умеренно влажному арктическому и далее переходя в арктический сухой.
Палинологическими спектрами из этой части разреза зафиксированы наиболее холодные климатические условия. Ухудшение климатических условий вызвало исчезновение лесных растительных ассоциаций и распространение гипоарктических и арктических видов (Betula nana, Alnaster fruticosus, Salix sp., Lycopodium alpinum, L. pungens, Selaginella selaginoides). Среди трав господствующее положение занимали разнотравно-злаковые ассоциации, произрастали маревые и полыни. Спектры отражают развитие тундровых арктических формаций и холодных степей в условиях сурового и сухого климата.
Сочетание перигляциальных тундровых и криофитных степных растительных ассоциаций было характерным для эпохи последнего оледенения на всей территории Восточной Европы, когда на огромных пространствах установилось господство холодного и сухого климата (Болиховская и др., 2001; Кольфсхотен и др., 2008; Гричук, Борисова, 2009; Андреичева и др., 2015; Величко и др., 2017). Так, в это время в растительном покрове центральной и южной частей Русской равнины отмечается абсолютное господство в спектрах пыльцы травянистых растений – злаков, полыней, маревых, что характеризует своеобразные ландшафты аридно-перигляциальных степей, не имеющих аналогов в современном растительном покрове Русской равнины (Болиховская и др., 2001; Величко и др., 2017). На севере и северо-востоке Восточно-Европейской равнины по немногочисленным палинологическим данным в эпоху максимального распространения материковых льдов доминировал перигляциально-тундровый тип растительности в сочетании со степными ассоциациями/группировкам. По объединенным палеобиологическим данным (териологическим и флористическим) для региона в последний ледниковый максимум реконструируются кустарниковая тундра и перигляциальная тундро-лесостепь (Гричук, 1989; Кольфсхотен и др., 2008; Барышников, Маркова, 2009; Гричук, Борисова, 2009).
Позднеледниковье
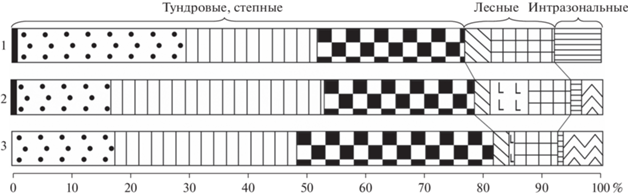
Фауна грызунов из слоя 3 ниши Филина на 77% представлена тундровыми и тундростепными видами, на 15% лесными и на 8% интразональными. Аналогичные состав и структуру демонстрируют датированные позднеледниковые ассоциации грызунов из бурого суглинка А Медвежьей пещеры (14.2 кал. тыс. лет назад) и грота Щугер-4 на Приполярном Урале (13.7 кал. тыс. лет назад) (рис. 10). В комплексе остатков из бурого суглинка А Медвежьей пещеры 79% приходится на тундровые и тундростепные виды, 11% на лесные и 8% на интразональные (Гуслицер, Павлов, 1988). В микротериофауне из Щугер-4 77% приходится на тундровые и тундростепные виды, 17% на лесные и 5% на интразональные (Кряжева и др., 2012).
Рис. 10.
Состав фауны грызунов (%) из позднеледниковых местонахождений Северного и Приполярного Урала. 1 – Ниша Филина (слой 3), 2 – Щугер-4 (слой 2 и 3), 3 – Медвежья пещера (бурый суглинок А). Условные обозначения см. рис. 8.
Палинологические спектры из этой части разреза ниши Филина указывают на преобладание в растительном покрове ерников – кустарниковых зарослей березы, с березовыми редколесьями и единичными деревьями ели. В травяном покрове преобладали злаковые и маревые. Среди споровых доминировали папоротниковые. Климатические условия были холоднее современных.
По данным авторов, на западном склоне Приполярного Урала в это время также произрастали тундроподобные ерниковые березовые заросли и травянистые ассоциации из разнотравья, злаков и маревых. Климат был значительно холоднее, чем сейчас, и, вероятно, напоминал современные климатические условия в пределах предгорных ерниковых тундр Полярного Урала (средние температуры июля 8–10°С, января (–20)°С, года ниже (–7)°С, сумма годовых осадков 400–500 мм; Голубева, Кряжева, 2020).
В это время на севере Восточной Европы были широко распространены перигляциальная кустарниковая тундра (севернее 61°–63° с.ш.) и перигляциальная лесотундра (59°–62° с.ш.) с участками тундростепи. Здесь произрастали ассоциации с Betula nana, Salix, Ericales, Hippophae rhamnoides, Juniperus, Rubus, Helianthemum, Armeria, Sphagnum, Selaginella при участии сосново-березовых и еловых лесов (Арсланов и др., 1981; Гричук, 1982; Никифорова, 1982; Borisova, Zelikson, 1995; Velichko et al., 1997, 2002; Симакова, Пузаченко, 2008). Таким образом, состав и структура ископаемого комплекса остатков грызунов, палинологическая характеристика и положение в разрезе костеносного слоя, свидетельствуют о том, что накопление рыхлых отложений в основании разреза ниши Филина происходило в позднеледниковое время.
Голоцен
Состав и структура микротериофауны нижней и верхней частей слоя 1 ниши Филина существенно различаются, несмотря на то, что по литологическим признакам рыхлые отложения не имеют явных различий. В нижней части фауна на 55% представлена лесными видами, на 24% тундровыми и тундростепными и на 21% интразональными, в то время как в верхней части в ее составе присутствуют только лесные (57%) и интразональные виды (43%).
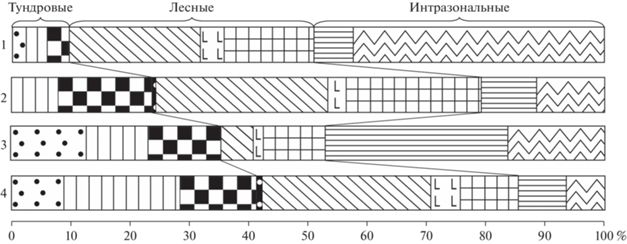
По составу и структуре фауна грызунов из нижней части слоя не похожа ни на один известный голоценовый комплекс мелких млекопитающих Тимано-Североуральского региона. В начале раннего голоцена (около 10.6 кал. тыс. лет назад) в фауне грызунов региона наблюдалась очень высокая доля тундровых и тундростепных видов (от 35 до 41%), затем около 9.5 кал. тыс. лет назад их доля сократилась до 10% (рис. 11) (Смирнов, 1996; Смирнов и др., 1999; Кряжева и др., 2012). Учитывая высокую долю тундровых и тундростепных видов в фауне грызунов из нижней части слоя 1 ниши Филина и положение в разрезе костеносного горизонта, можно предположить, что формирование нижней части слоя 1 ниши Филина происходило в один из периодов раннего голоцена, при переходе от тундроподобных ландшафтов к лесным.
Рис. 11.
Состав фауны грызунов (%) из раннеголоценовых местонахождений Тимано-Североуральского региона. 1 – Уса-2 (слой 2), 2 – Ниша Филина (слой 1, нижняя часть), 3 – Кожим-1 (слой 2), 4 – Медвежья пещера (слой зеленоватой супеси). Условные обозначения см. рис. 8.
Фауна грызунов из верхней части слоя 1 ниши Филина по составу и структуре имеет типичный лесной облик, характерный для многих позднеголоценовых и современных ассоциаций грызунов лесной зоны Урала и Тимана.
Микротериофауна слоя 1 навеса Студеного также состоит из лесных и интразональных видов грызунов и схожа со всеми известными современными комплексами грызунов верховьев р. Печора (Смирнов, Садыкова, 2003). Это типичная фауна темнохвойной средней тайги.
Палинологический спектр из нижней части разреза слоя 1 навеса Студеного указывает на произрастание в данном районе березово-еловых лесов с примесью сосны и лиственницы, с заболоченными участками и водоемами со стоячей или медленнотекущей водой. В покрове лесов значительная роль принадлежала папоротникам.
Палинологические спектры из верхней части разреза ниши Филина и слоя 1 навеса Студеного очень похожи и указывают на четкую тенденцию к увеличению содержания пыльцы хвойных деревьев (ели, сосны), отмечается появление кедра и пихты. Состав пыльцы трав и спор становится разнообразнее, водные растения исчезают. Эти изменения в составе спектров позволяют сделать вывод о значительном потеплении климата, которое способствовало развитию хвойных сосново-еловых лесов с примесью кедра и пихты.
Надежно подтвержденные радиоуглеродным датированием палинологические данные по голоцену верховьев р. Печора получены при комплексном исследовании двух скважин Yak2 и Yak3, заложенных в районе пос. Якша, расположенного в 100 км к западу от лога Иорданского (Chéïma Barhoumi et al., 2020). Согласно этим палинологическим записям, в начале голоцена 11700–8200 кал. лет назад в Северном Предуралье была развита светлохвойная тайга. В составе древостоев преобладали береза и сосна/ель. В середине голоцена, 8200–4200 кал. лет назад, в составе лесной растительности произошло перераспределение среди древесных пород: увеличилось участие ели за счет сокращения долей сосны и березы, появились кедр и пихта, что указывает на распространение темнохвойной тайги. С 4200 по 100 кал. лет назад в древостоях сосна вытесняла ель и кедр; постепенно увеличивались площади, занимаемые светлохвойной тайгой вследствие участившихся пожаров. В период 100 кал. лет назад по настоящее время резко увеличилось количество березы, свидетельствующее о значительном влиянии человеческой деятельности (лесозаготовок, сжигания лесов при расчистке территории) на окружающую среду.
ВЫВОДЫ
Комплексное палеозоологическое и палинологическое изучение спелеогенных отложений двух местонахождений пещерного типа в карстовом логе Иорданского в верховьях р. Печора (западный склон Северного Урала) – навеса Студеный и ниши Филина – позволило реконструировать специфические особенности животных и растительных ассоциаций в верховьях Печоры в конце среднего валдая, последнем ледниковом максимуме, в позднеледниковье и в голоцене (рис. 12).
Рис. 12.
Изменения фаунистических и флористических ассоциаций на западном склоне Северного Урала (верховья Печоры) с конца среднего валдая до позднего голоцена. 1 – тундровые и тундростепные виды, 2 – лесные виды, 3 – интразональные виды, 4 – копытный лемминг, 5 – сибирский лемминг, 6 – узкочерепная полевка, 7 – полевка Миддендорфа, 8 – лиственные деревья, 9 – хвойные деревья, 10 – кустарники, 11 – степная растительность, 12 – разнотравье, 13 – влажность, 14 – температурная кривая. * Для построения диаграммы были использованы данные по среднему голоцену Приполярного Урала (грот Соколиный).
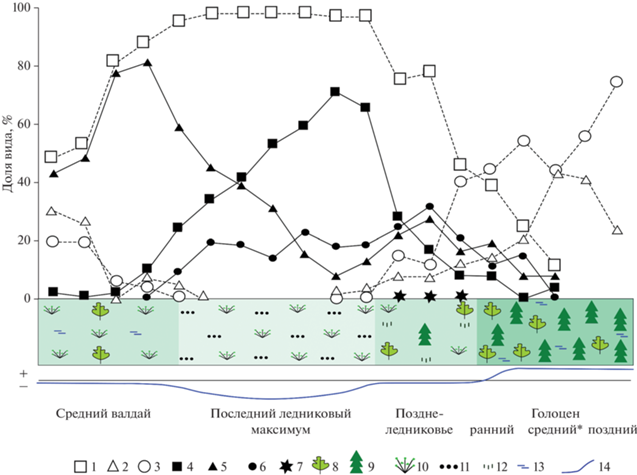
В конце среднего валдая (35–30 тыс. кал. лет назад) на территории Печорского Урала были развиты ландшафты влажных моховых тундр с березовыми редколесьями, климат был значительно холоднее, чем в настоящее время. В составе микротериофауны доминировал обитатель заболоченной тундры – сибирский лемминг. Лесные полевки населяли лесные островки в тундре и участки леса в долине р. Печора и ее притоках.
В переходный период от интерстадиала к максимуму последнего оледенения климат оставался холодным и умерено влажным. В фауне грызунов произошла постепенная смена вида доминанта с сибирского лемминга на копытного.
В период максимума последнего оледенения произошло ухудшение климатических условий, которое выразилось в сильном похолодании и аридизации климата, что привело к процессу остепнения тундровых ландшафтов. Исчезли лесные ассоциации и распространились гипоарктические и арктические виды растений. Расцвет получили злаковые. В фауне грызунов доминировал экстремально криоксерофильный вид – копытный лемминг и узкочерепная полевка – вид, предпочитающий холодные и сухие условия специфического плейстоценового биома – тундростепи.
В позднеледниковое время были распространены тундроподобные ерниковые заросли березы с березовыми редколесьями и единичными деревьями ели. Климат был значительно холоднее современного и, вероятно, напоминал современные климатические условия в пределах предгорных ерниковых тундр Полярного Урала. В фауне грызунов доминировали тундровые и тундростепные виды.
В начале голоцена в Северном Предуралье была развита светлохвойная тайга. В составе древостоев преобладали береза и сосна/ель. В фауне грызунов преобладали лесные виды. Доля тундровых и тундростепных видов была весьма значительной (24%).
В среднем и позднем голоцене на территории были распространены хвойные сосново-еловых леса с примесью кедра и пихты. Фауна грызунов состояла из лесных и интразональных видов.
Источники финансирования. Работа выполнена в рамках темы НИР “Эволюция биоты и среды ее обитания как основа расчленения и геологической корреляции осадочного чехла Печорской плиты и ее складчатого обрамления” № 1021062311460-9.
Список литературы
Абрамсон Н.И., Лисовский А.А. Полевки // Млекопитающие России: систематико-географический справочник. Ред. Павлинов И.Я., Лисовский А.А.М.: Товарищество научных изданий КМК, 2012. С. 220–276 (Сборник трудов Зоологического музея МГУ. Т. 52).
Андреичева Л.Н., Марченко-Вагапова Т.И., Буравская М.Н., Голубева Ю.В. Природная среда неоплейстоцена и голоцена на Европейском Северо-Востоке России. М.: ГЕОС, 2015. 224 с.
Ануфриев В.М. Рыжая (европейская) полевка // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Часть 1. СПб.: Наука, 1994a. С. 173–185.
Ануфриев В.М. Полевка-экономка // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Часть 1. СПб.: Наука, 1994б. С. 242–251.
Ануфриев В.М. Темная (пашенная) полевка // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Часть 1. СПб.: Наука, 1994в. С. 252–262.
Ануфриев В.М., Бобрецов А.В. Красно-серая полевка // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Часть 1. СПб.: Наука, 1994. С. 167–173.
Арсланов Х.А., Лавров А.С., Никифорова Л.Д. О стратиграфии, геохронологии и изменении климата среднего и позднего плейстоцена и голоцена на северо-востоке Русской равнины. Плейстоценовые оледенения Восточно-Европейской равнины. М.: Изд-во АН СССР, 1981. С. 45–54.
Астахов В.И. О датировании палеоландшафтов среднего валдая // Пути эволюционной географии – 2021. Материалы II Всероссийской научной конференции, посвященной памяти профессора А.А. Величко (Москва, 22–25 ноября 2021 г.). М.: Институт географии РАН, 2021.С. 752–757.
Барышников Г.А., Маркова А.К. Глава 10. Основные териокомплексы в холодную эпоху позднего плейстоцена (карта 23) // Палеоклиматы и палеоландшафты внетропического пространства Северного полушария. Поздний плейстоцен–голоцен. Атлас-монография. М.: ГЕОС, 2009. С. 79–85.
Болиховская Н.С., Гунова В.С., Соболев В.М. Основные этапы развития перигляциальной растительности центра и юга Русской равнины в период существования мамонтовой фауны // Мамонт и его окружение: 200 лет изучения. М.: ГЕОС, 2001. С. 168–187.
Большаков В.Н., Васильева И.А., Малеева А.Г. Морфотипическая изменчивость зубов полевок. М.: Наука, 1980. 140 с.
Бородин А.В., Коурова Т.П., Маркова Е.А. Размерные характеристики щечных зубов лесных полевок Clethrionomus (Craseomys) rufocanus, Cl. (Clethrionomus) glareolus, Cl. (Cl.) rutilus (Arvicolinae, Rodentia) и их использование для видовой идентификации // Зоологич. журн. 2005. Т. 84. № 2. С. 236–244.
Величко А.А., Борисова О.К., Захаров А.Л., Кононов Ю.М., Константинов Е.А., Курбанов Р.Н., Морозова Т.Д., Панин П.Г., Тимирева С.Н. Смена ландшафтных обстановок на юге Русской равнины в позднем плейстоцене по результатам исследования лёссово-почвенной серии Приазовья // Изв. РАН. Сер. географ. 2017. № 1. С. 74–83.
Верещагин Н.К., Кузьмина И.Е. Раскопки в пещерах Северного Урала // Природа. 1962. № 3. С. 76–78.
Головачев И.Б., Смирнов Н.Г., Добышева Э.В., Пономарев Д.В. К истории современных подвидов узкочерепной полевки // Современные проблемы популяционной, исторической и прикладной экологии. Материалы конференции молодых ученых. Екатеринбург, 2001. В. 2. С. 49–57.
Голубева Ю.В., Кряжева И.В. Развитие растительности и микротериофауны в позднеледниковье и голоцене на территории национального парка “Югыд-ва” // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 3. С. 148–160.
Гричук В.П. Растительность Европы в позднем плейстоцене // Палеогеография Европы за последние сто тысяч лет. М.: Наука, 1982. С. 92–109.
Гричук В.П. История флоры и растительности Русской равнины в плейстоцене. М.: Наука, 1989. 183 с.
Гричук В.П., Борисова О.К. Глава 9. Растительный покров. 9А. Поздний плейстоцен (карты 20–21) // Палеоклиматы и палеоландшафты внетропического пространства Северного полушария. Поздний плейстоцен–голоцен. Атлас-монография. М.: ГЕОС, 2009. С. 70–74.
Гричук В.П., Заклинская Е.Д. Анализ ископаемых пыльцы и спор и его применение в палеогеографии. М.: ОГИЗ ГЕОГРАФГИЗ, 1948. 223 с.
Громов И.М., Ербаева М.А. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб.: ЗИН РАН, 1995. 522 с.
Громов И.М., Поляков И.Я. Фауна СССР. Млекопитающие. Полевки (Microtinae). Т. 3. В. 8. Л.: Наука, 1977. 504 с.
Гуслицер Б.И., Канивец В.И. Пещеры Печорского Урала. М.: Наука, 1965. 134 с.
Гуслицер Б.И., Павлов П.Ю. Верхнепалеолитическая стоянка Медвежья пещера // Памятники эпохи камня и металла Северного Приуралья. Материалы по археологии Европейского Северо-Востока. 1988. Вып. 11. С. 5–18.
Гуслицер Б.И., Павлов П.Ю., Панюкова Н.Н. Биостратиграфия и возраст отложений пещеры Студеной на верхней Печоре // Тр. Ин-та геологии Коми НЦ УрО АН СССР. 1989. Вып. 73. С. 92–100.
Дурягина Д.А., Коноваленко Л.А. Палинология плейстоцена северо-востока европейской части России. СПб.: Наука, 1993. 124 с.
Кольфсхотен Т.ван, Маркова А.К., Симакова А.Н., Пузаченко А.Ю. Экосистемы максимального похолодания последнего оледенения (LGM) (<24–>=17 тыс. л. н.) // Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н). М.: Товарищество научн. изд. КМК, 2008. С. 459–465.
Кочев В.А. Определение возраста четвертичных отложений по ископаемым остаткам копытных леммингов. Сыктывкар: Коми филиал АН СССР, 1984. 16 с. (Сер. препринтов “Новые научные методики”. Коми филиал АН СССР. Вып. 12).
Кочев В.А. Развитие фауны грызунов плейстоцена Северо-Востока европейской части СССР. Сыктывкар: Геонаука, 1991. 83 с.
Кочев В.А. Плейстоценовые грызуны северо-востока Европейской части России и их стратиграфическое значение. СПб.: Наука, 1993. 112 с.
Кряжева И.В., Пономарев Д.В. История фауны грызунов Северного Урала в позднем неоплейстоцене (данные палеонтологического исследования отложений навеса Студеного) // Вестник ИГ Коми НЦ УрО РАН. 2019. № 12. С. 25–33. https://doi.org/10.19110/2221-1381-2019-12-25-33
Кряжева И.В., Пономарев Д.В., Кольфсхотен Т.ван, ван дер Плихт Й. История формирования современных сообществ микромаммалий Приполярного Урала // Экология. 2012. № 6. С. 434–441.
Кузьмина И.Е. Формирование териофауны Северного Урала в позднем антропогене // Тр. Зоологич. ин-та АН СССР. 1971. Т. 49. С. 44–122.
Марченко-Вагапова Т.И. Палинологическая характеристика средневалдайского интерстадиала (поздний плейстоцен) на Европейском северо-востоке России // Литосфера. 2012. № 1. С. 38–53.
Никифорова Л.Д. Динамика ландшафтных зон голоцена Северо-Востока Европейской части СССР // Развитие природы территории СССР в позднем плейстоцене и голоцене. М.: Наука, 1982. С. 154–162.
Огнев С.И. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). Т. 7. Microtinae. М.–Л.: Изд-во АН СССР, 1950. 380 с.
Ошуркова М.Н. Описание пыльцы некоторых видов берез, произрастающих на территории СССР // Проблемы ботаники. В. 4. М.–Л.: Изд-во АН СССР, 1959. С. 68–91.
Петров А.Н. Копытный лемминг // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Ч. 1. СПб.: Наука, 1994а. С. 202–212.
Петров А.Н. Обский, или сибирский, лемминг // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Ч. 1. СПб.: Наука, 1994б. С. 213–220.
Петров А.Н. Узкочерепная полевка // Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Ч. 1. СПб.: Наука, 1994в. С. 236–241.
Петров А.Н. Мелкие млекопитающие (Insectivora, Rodentia) трансформированных и ненарушенных территорий восточноевропейских тундр. СПб.: Наука, 2007. 178 с.
Пыстин А.Н. Красная (сибирская) полевка. Фауна европейского Северо-Востока России. Млекопитающие. Т. II. Ч. 1. СПб.: Наука, 1994. С. 186–202.
Симакова А.Н., Пузаченко А.Ю. Растительность в период межстадиальных потеплений беллинг–аллеред (<12.4–≥10.9 тыс. л. н.) // Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24–8 тыс. л. н.). М.: Товарищество научн. изд. КМК, 2008. С. 369–395.
Смирнов Н.Г. Разнообразие мелких млекопитающих Северного Урала в позднем плейстоцене и голоцене // Материалы и исследования по истории современной фауны Урала. Екатеринбург: Изд-во “Екатеринбург”, 1996. С. 39–83.
Смирнов Н.Г., Садыкова Н.О. Источники погрешностей при фаунистических реконструкциях в четвертичной палеозоологии // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Урал. ун-та, 2003. С. 98–115.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А., Панова Н.К., Коробейников Ю.И., Ольшванг В.Н., Ерохин Н.Г., Быкова Г.В. Историческая экология животных гор Южного Урала. Свердловск: УрО АН СССР, 1990. 244 с.
Смирнов Н.Г., Головачев И.Б., Бачура О.П., Кузнецова И.А., Чепраков М.И. Сложные случаи определения зубов грызунов из отложений позднего плейстоцена и голоцена тундровых районов Северной Евразии // Материалы по истории и современному состоянию фауны севера Западной Сибири. Челябинск: Рифей, 1997. С. 60–90.
Смирнов Н.Г., Андреичева Л.Н., Корона О.М., Зиновьев Е.В., Головачев И.Б., Павлов П.Ю., Хуфтхаммер А.К. Материалы к характеристике биоты Приуральской Субарктики в голоценовом оптимуме // Биота Приуральской Субарктики в позднем плейстоцене и голоцене. Екатеринбург: Изд-во “Екатеринбург”, 1999. С. 23–60.
Спиридонова Е.А. Палинологическая характеристика средневалдайского мегаинтерстадиала и ее значение для восстановления истории развития флоры и растительности Русской равнины // Бюлл. Комиссии по изучению четвертичного периода. 1983. № 52. С. 42–57.
Astakhov V.I. The postglacial Pleistocene of the northern Russian mainland // Quaternary Sci. Rev. 2014. V. 92. P. 388–408.
Borisova O.K., Zelikson E.M. Vegetation and climate of Eastern Europe during the Late Glacial Climate and environment changes of East Europe during Holocene and Late–Middle Pleistocene // Preprint of research materials for IGU 46 Conference “Global Changes and Geography”. M.: Institute of Geography RAS, 1995. P. 14–19.
Chéïma Barhoumi, Adam A. Ali, Odile Peyron, Lucas Dugerdil, Olga Borisova, Yulia Golubeva, Dmitri Subetto, Alexander Kryshen, Igor Drobyshev, Sébastien Joannin. Did long-term fire control the coniferous boreal forest composition of northern Ural region (Komi Republic, Russia)? // J. Biogeography. 2020. V. 47. Iss. 11. P. 2426–2441. https://doi.org/10.1111/jbi.13922
Golovachov I.B., Smirnov N.G. The Late Pleistocene and Holocene rodents of the Pre-Urals Subarctic // Quaternary Int. 2009. V. 201. P. 37–42.
Grimm E.C. TILIA and TILIA GRAPH.PC spreadsheet and graphics software for pollen data // INQUA. Working Group on Data-Handling Methods. Newsletter. 1990. V. 4. P. 5–7.
Kryštufek B., Tesakov A.S., Lebedev V.S., Bannikova A.A., Abramson N.I., Shenbrot G. Back to the future: the proper name for red-backed voles is Clethrionomys Tilesius and not Myodes Pallas // Mammalia. 2020. V. 84. № 2. P. 214–217.
Lagerholm V.K., Sandoval-Castellanos E., Ehrich D., Abramson N.I., Nadachowski A., Kalthoff D.C., Germonpré M., Angerbjörn A., Stewart J.R., Dalén L. On the origin of the Norwegian lemming // Molecular Ecology. 2014. V. 23. № 8. P. 2060–2071.
Meulen A.J. van der. Middle Pleistocene smaller mammals from the Monte Pegalia (Orvieto, Italy) with special reference to the phylogeny of Microtus (Arvicolidae, Rodentia) // Quaternaria. 1973. V. 17. P. 1–144.
Palkopoulou E., Baca M., Abramson N.I., Sablin M., Socha P., Nadachowski A., Prost S., Germonpré M., Kosintsev P., Smirnov N.G., Vartanyan S., Ponomarev D., Nyström J., Nikolskiy P., Jass C.N., Litvinov Y.N., Kalthoff D.C., Grigoriev S., Fadeeva T., Douka A., Higham T.F.G., Ersmark E., Pitulko V., Pavlova E., Stewart J.R., Węgleński P., Stankovic A., Dalén L. Synchronous genetic turnovers across Western Eurasia in Late Pleistocene collared lemmings // Global Change Biology. 2016. V. 22. Iss. 5. P. 1710–1721.
Ponomarev D., Puzachenko A. Evolution of occlusal shape of the first and second upper molars of Middle-Late Pleistocene collared lemmings (Dicrostonyx, Arvicolinae, Rodentia) in northeast European Russia // Boreas. 2015. V. 44. P. 741–759.
Ponomarev D., Puzachenko A. Changes in the morphology and morphological diversity of the first lower molar of narrow-headed voles (Microtus gregalis, Arvicolinae, Rodentia) from northeastern European Russia since the Late Pleistocene // Quaternary Int. 2017. V. 436. P. 239–252.
Ponomarev D., Puzachenko A., Bachura O., Kosintsev P., van der Plicht J. Mammal fauna during the Late Pleistocene and Holocene in the far northeast of Europe // Boreas. 2013. V. 42. Iss. 3. P. 779–797.
Reimer P.J., Austin W.E.N., Bard E., Bayliss A., Blackwell P.G., Bronk Ramsey C., Butzin M., Cheng H., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P., Hajdas I., Heaton T.J., Hogg A.G., Hughen K.A., Kromer B., Manning S.W., Muscheler R., Palmer J.G., Pearson C., van der Plicht J., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Turney C.S.M., Wacker L., Adolphi F., Büntgen U., Capano M., Fahrni S.M., Fogtmann-Schulz A., Friedrich R., Köhler P., Kudsk S., Miyake F., Olsen J., Reinig F., Sakamoto M., Sookdeo A., Talamo S. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 calkBP) // Radiocarbon. 2020. V. 62. P. 725–757. https://doi.org/10.1017/RDC.2020.41
Svendsen J.I., Heggen H.P., Hufthammer A.K., Mangerud J., Pavlov P., Roebroeks W. Geo-archaeological investigations of Palaeolithic sites along the Ural Mountains – on the northern presence of humans during the last Ice Age // Quaternary Sci. Rev. 2010. V. 29. P. 3138–3156.
Velichko A.A., Andrev A.A., Klimanov V.A. Climate and vegetation dynamics in the Tundra and Forest Zone during the Late glacial and Holocene // Quaternary Int. 1997. V. 41/42. P. 71–96.
Velichko A.A., Catto N., Drenova A.N., Klimanov V.A., Kremenetski K.V., Nechaev V.P. Climate changes in East E-urope and Siberia at the Late glacial-holocene transition // Quaternary Int. 2002. V. 91. P. 75–99.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция