

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 4, стр. 76-99
Стратиграфическое положение и состав аянкинской флоры из верхнего мела Охотско-Чукотского вулканогенного пояса, Северо-Восток России
М. Г. Моисеева 1, *, А. Б. Герман 1, А. Б. Соколова 2
1 Геологический институт РАН
Москва, Россия
2 Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: masha.moiseeva@gmail.com
Поступила в редакцию 25.10.2021
После доработки 29.12.2021
Принята к публикации 30.01.2022
- EDN: NQLHMC
- DOI: 10.31857/S0869592X22040068
Аннотация
Впервые приводится детальное описание трех флористических комплексов, остатки растений которых были собраны в бассейне р. Большая Аянка (Северо-Восток России) в верхнемеловых вулканогенных отложениях Охотско-Чукотского вулканогенного пояса. В составе флоры установлено около 60 видов ископаемых растений, среди которых доминируют хвойные и покрытосеменные, менее разнообразны папоротники; также встречены единичные хвощовые, печеночники, гинкговые и цикадовые. Сравнение аянкинской флоры с другими ископаемыми флорами показало ее несомненное сходство с сантон-кампанскими флорами Северо-Востока России и Северной Аляски (с барыковской, позднебыстринской, усть-эмунеретской, в меньшей степени с ольской и Ранней Когосакрак), что позволило нам сделать вывод о ее наиболее вероятном сантон-кампанском возрасте. Флористические комплексы аянкинской флоры отражают как склоновую растительность, так и растительность пониженных участков рельефа – берегов реки или озера, причем в первой доминировали хвойные растения, а во второй – покрытосеменные. Эти комплексы аянкинской флоры представляют собой свидетельство внедрения растительных сообществ с доминированием продвинутых покрытосеменных из располагавшихся восточнее приморских низменностей Анадырско-Корякского субрегиона на территорию вулканического нагорья, где в позднем мелу часто встречались папоротники и хвойные и еще сохранились реликтовые растения раннего и начала позднего мела.
ВВЕДЕНИЕ
Наряду с радиоизотопными и палеомагнитными данными, сведения о возрасте ископаемых растений широко используются для стратиграфии субаэральных образований Охотско-Чукотского вулканогенного пояса (ОЧВП) и геологического картирования этой территории. Фитостратиграфические исследования сыграли, в частности, решающую роль в дискуссии о времени формирования ОЧВП – был ли он областью активного вулканизма лишь в первой половине сеномана (Белый, 1977), или же вулканическая деятельность продолжалась в нем до середины кампана включительно (Филатова, Лебедев, 1982; Лебедев, 1992). Наименее изученными в настоящее время остаются сантон-кампанские флоры данного региона, которые до недавних пор фигурировали в публикациях только в виде списков предварительных определений.
Остатки растений, обнаруженные М.Ю. Хотиным и В.А. Фараджевым на Чукотке в долине р. Обрывистая – притоке р. Большая Аянка (Филатова, Лебедев, 1982), собирали М.Ю. Хотин в 1969 г., Н.И. Филатова в 1970 г., Е.Л. Лебедев в 1977 г. и Е.Л. Лебедев, А.Б. Герман и Е.И. Костина в 1985 г. Этот комплекс растений впервые кратко охарактеризовали Филатова и Лебедев (1982); позже Лебедев (1987) выделил его как аянкинскую флору (флористический комплекс) сенонского возраста. Наиболее полный список растений аянкинской флоры, включающий 26 видов, приведен в работе Лебедева (1987, с. 72), который считал, что возраст этой флоры примерно соответствует раннему кампану (Лебедев, 1992). Следует отметить, что данный список, несомненно далеко не полный, был им составлен по результатам предварительных определений. Кроме этого, не был решен вопрос о положении флороносных образований бассейна р. Обрывистая в существующих схемах местных стратиграфических подразделений.
Следует отметить, что Лебедев (1987) ископаемую флору с р. Обрывистая называл “аянкинская флора” либо “аянкинский комплекс”. В настоящей статье под термином “тафофлора” или “комплекс” (“флористический комплекс”) мы понимаем совокупность ископаемых растений из одного или нескольких территориально и стратиграфически близких местонахождений, отражающую растительность определенной местности в определенный отрезок времени; составляющие тафофлору растения существовали совместно на ограниченной территории в течение небольшого интервала геологической истории. Термин “ископаемая флора” (или просто “флора”) используется нами как термин свободного пользования. Поскольку, как будет показано ниже, ископаемые растения из бассейна р. Обрывистая происходят из достаточно удаленных друг от друга местонахождений, их совокупность мы называем “аянкинская флора”, а растения из каждого из этих местонахождений – тафофлорой или флористическим комплексом.
Позднее М.Г. Моисеева с соавторами (2014) провели таксономическую ревизию аянкинской флоры, в результате чего в ней были определены 42 вида ископаемых растений. Однако в этой публикации был недостаточно обоснован вывод о возрасте флороносных слоев, а ископаемые растения не были охарактеризованы и изображены, т.е. остались малопригодными для целей фитостратиграфии и палеофлористики. В настоящей статье мы восполняем этот пробел и приводим детальное описание флористических комплексов из трех местонахождений бассейна р. Обрывистая, изображения всех основных ископаемых растений аянкинской флоры, а также обоснование возраста изученной флоры.
МАТЕРИАЛ И МЕТОДЫ
Изученные ископаемые растения были собраны Е.Л. Лебедевым, А.Б. Германом и Е.И. Костиной в 1985 г. из трех местонахождений в бассейне верхнего течения р. Обрывистая – левого притока р. Большая Аянка, протекающей на Чукотке (рис. 1а). Они получили номера (полевые обозначения) 700, 701 и 702.
Рис. 1.
Схематическая геологическая карта и положение трех местонахождений аянкинской флоры (а) и последовательности стратиграфических подразделений, выделяемых геологами объединения “Аэрогеология” и СВПГО “Севвостгеология” (б) района верхнего течения р. Обрывистая, составленные по материалам государственных геологических карт масштаба 1 : 200 000.
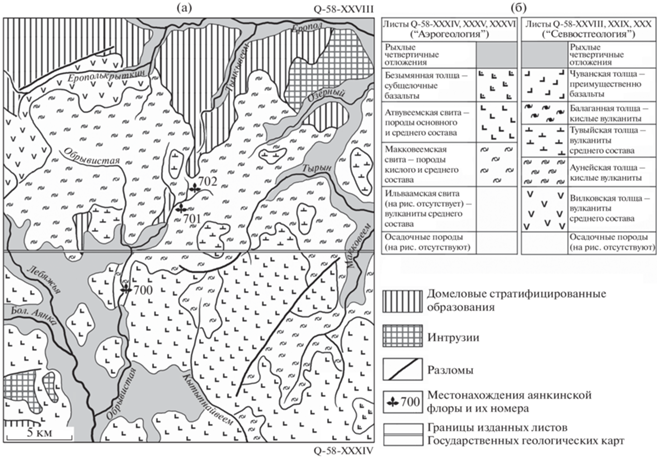
Местонахождение 700 включает флороносные слои двух близко расположенных обнажений на р. Обрывистая (рис. 1а) примерно в 20 км от ее устья. Наиболее крупное обнажение (рис. 2) находится на левом берегу реки приблизительно в 1700 м юго-западнее высоты 692 м. Здесь в береговом обрыве выходят серые и желтовато-серые туфы, туфопесчаники и туфоалевролиты видимой мощностью около 20–25 м. В южной части обнажения остатки растений хорошей сохранности встречены в нескольких прослоях тонкозернистых туфопесчаников и туфоалевролитов; здесь преобладают отпечатки хвойных Parataxodium Arnold et Lowther и Taxites Brongniart и менее многочисленных покрытосеменных, среди которых найдены единичные листья “Macclintockia” и Barykovia tchucotica (Abramova) Moiseeva. В северной части обнажения в грубо- и крупнозернистых туфопесчаниках остатки растений фрагментарны и представлены покрытосеменными и хвойными, обрывки которых располагаются в виде изолированных скоплений (“гнезд”). В средней, наиболее высокой части обнажения, остатки хвойных и покрытосеменных были найдены в тонкозернистых туфоалевролитах.
Рис. 2.
Местонахождение 700 на левом берегу р. Обрывистая. Значком в виде трилистника показаны места сбора ископаемых растений в 1985 г.

Второе, меньшее по мощности и протяженности обнажение расположено на правом берегу реки примерно в 200 м выше по течению от первого, приблизительно в 300–350 м восточнее высоты 658.9 м. Ископаемые растения здесь встречены в верхней части обнажения в прослое тонкозернистого туфопесчаника, выходящего среди более грубозернистых туфопесчаников. Оба обнажения точки 700 располагаются близ кровли флороносной толщи кислого состава под покровами лав основного–среднего состава (рис. 1), что позволяет считать эти обнажения одновозрастными.
Местонахождение 701 представляет собой склон левого берега ручья Звонкий (рис. 1а) – левого притока р. Обрывистая, примерно в 12 км от точки 700. Здесь ископаемые растения были собраны в основном из осыпей, в которых представлены туфоалевролиты и мелкозернистые туфопесчаники. Разнообразие ископаемых растений в этой точке небольшое, причем по количеству экземпляров наиболее многочисленны хвойные (остатки полиморфных побегов “Sequoia” sp.), и встречены редкие покрытосеменные “Macclintockia” и Quereuxia.
Местонахождение 702 расположено на прирусловом склоне правого берега ручья Звонкий (рис. 1а). Ископаемые растения были собраны из небольшой линзы (размером 1.5 × 0.5 м) плотных мелкозернистых туфопесчаников, обнажающихся среди более крупнозернистых туфопесчаников приблизительно в 10 м выше основания склона. Здесь преобладают отпечатки листьев разнообразных покрытосеменных растений.
Флороносные слои местонахождений 701 и 702, по нашим полевым наблюдениям, располагаются приблизительно на одном стратиграфическом уровне. Их соотношение с отложениями местонахождения 700 проследить на местности не представляется возможным, поскольку последняя закрыта таежной и тундровой растительностью. На геологических картах разрывных нарушений между местонахождениями не показано.
Несколько севернее местонахождения 700 проходит граница двух листов Государственной геологической карты масштаба 1 : 200 000 (ГГК-200), составленных разными организациями, использовавшими разные стратиграфические схемы и номенклатуру картируемых стратонов (рис. 1). По границе листов к выходам макковеемской свиты, к которой приурочено местонахождение 700, примыкают выходы аунейской толщи, в которой располагаются слои местонахождений 701 и 702. Поскольку разломы на картах в этом месте не обозначены, можно с уверенностью полагать, что это – образования одного стратона, названного на южном и северном листах карты по-разному. Исходя из сказанного, мы считаем все три местонахождения ископаемых растений приблизительно одновозрастными.
Материал представлен отпечатками листьев, облиственных побегов и репродуктивных органов (женских и мужских шишек хвойных и плодов покрытосеменных), фитолеймы на них не сохранились (табл. I–VI ). Эта коллекция состоит из 390 образцов (штуфов) с отпечатками растений хорошей сохранности. Коллекция № 3395 хранится в Геологическом институте (ГИН) РАН, г. Москва.
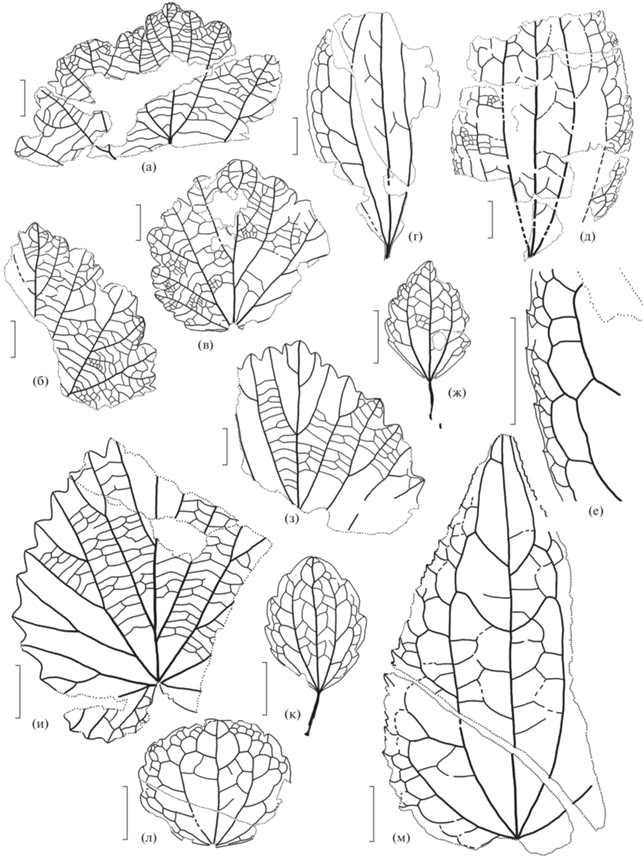
Таблица I . Ископаемые растения аянкинской флоры из местонахождения 700, р. Обрывистая. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 2, 6, 15, 17 ; для фиг. 2, 15 – 0.5 см; для фиг. 6 – 0.2 см; для фиг. 17 – 0.1 см. 1, 2 – Thallites sp. 1, экз. 3395/328: 1 – общий вид таллома, 2 – увеличенные детали; 3 – Thallites sp. 2, экз. 3395/329а; 4 – Arctopteris sp. 1, экз. 3395/323б-1; 5 – Arctopteris sp. 2, экз. 3395/326; 6 – женская шишка Cupressaceae тип 3 (Taxodioideae?), экз. 3395/13; 7 – Ginkgodium (?) sp., экз. 3395/383; 8 – Coniopteris tschuktschorum (Krysht.) Samyl., фрагмент фертильного пера, экз. 3395/357; 9 – Cladophlebis sp. 1, экз. 3395/295-5; 10 – Ginkgo ex gr. adiantoides Heer, экз. 3395/381-3; 11, 12 – Ditaxocladus sp.: 11 – экз. 3395/79, 12 – экз. 3395/85; 13 – Ginkgo ex gr. digitata Brong., экз. 3395/387; 14 – женская шишка Cupressaceae тип 3, экз. 3395/301; 15 – побег с мужскими шишками Cupressaceae тип 3, экз. 3395/116; 16, 17 – Monocotylophyllum sp.: 16 – экз. 3395/286-8, общий вид листа, 17 – экз. 3395/286-8, детали верхушки и жилкования.

Таблица II. Ископаемые растения аянкинской флоры из местонахождения 700, р. Обрывистая. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 2 (2 мм). 1, 2, 8 – Parataxodium cf. wigginsii Arnold et Lowther, облиственные побеги: 1 – экз. 3395/26а, побег с мужскими шишками, 2, 8 – экз. 3395/11, побег с двумя типами листорасположения – очередным и супротивным; 3 – Parataxodium sp., экз. 3395/30-1, облиственный побег; 4 – Pityocladus sp., побег с брахибластами, экз. 3395/114; 5 – женская шишка Cupressa-ceae тип 1 (подсем. Sequoioideae), экз. 3395/93а; 6 – Elatocladus sp. 1, облиственный побег, экз. 3395/16а; 7 – Metasequoia sp. 1, облиственный побег с супротивным листорасположением, экз. 3395/19; 9 – побег с мужскими шишками Cupressaceae тип 2, экз. 3395/113; 10 – Metasequoia sp. 2, побег с супротивным листорасположением, экз. 3395/1-3; 11 – Pityostrobus sp. 2, женская шишка, экз. 3395/96; 12 – Taxites sp., облиственный побег, экз. 3395/75; 13 – мужские шишки Cupressaceae тип 1, экз. 3395/96; 14 – Pityospermum sp., крылатое семя, экз. 3395/109а; 15 – Pityostrobus sp. 1, женская шишка, экз. 3395/97; 16 – Pityolepis sp., дисперсная шишечная чешуя, экз. 3395/47.

Таблица III. Ископаемые растения аянкинской флоры из местонахождения 700, р. Обрывистая. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 4 и 11 ; для фиг. 4 – 0.5 см, для фиг. 11 – 0.1 см. 1, 2, 4 – “Vitis” penzhinica Herman: 1 – экз. 3395/383б, 2 – экз. 3395/382а, 4 – экз. 3395/383б, детали строения краевых зубцов; 3 – Trochodendroides sp. 1, экз. 3395/297а; 5 – Cissites sp., экз. 3395/381-1; 6 – Celastrinites sp., экз. 3395/259; 7 – “Macclintockia” ochotica Vachrameev et Herman, экз. 3395/286а; 8, 13 – Trochodendroides notabilis Herman: 8 – экз. 3395/249а, детали строения зубцов, 13 – экз. 3395/249б, общий вид листа; 9 – Dicotylophyllum sp. 1, экз. 3395/285а; 10 – Dicotylophyllum sp. 2, экз. 3395/295-1а; 11, 12 – Viburniphyllum sp., экз. 3395/299-1: 11 – детали строения зубцов, 12 – общий вид листа; 14, 15 – Menispermites sp.: 14 – экз. 3395/247а, фрагмент края листа, 15 – экз. 3395/250, нижняя часть листа; 16 – фрагмент платаноидного листа, экз. 3395/281.

Таблица IV. Ископаемые растения аянкинской флоры из местонахождения 701, руч. Звонкий. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 2, 4, 9, 13 ; для фиг. 2 – 0.2 см, для фиг. 4, 9, 13 – 0.5 см. 1 – “Sequoia” sp., облиственный побег, экз. 3395/119; 2, 8 – Pagiophyllum sp., экз. 3395/122: 2 – увеличенный фрагмент, 8 – общий вид побега; 3 – Sphenopteris sp., экз. 3395/344; 4 – Equisetites sp., экз. 3395/379; 5, 13, 15 – Arctopteris sp. 2: 5 – общий вид фертильного листа, экз. 3395/339-1б; 13 – то же, увеличенный фрагмент; 15 – верхняя часть стерильного листа, экз. 3395/335-1; 6 – Coniopteris tschuktschorum (Krysht.) Samyl., экз. 3395/340; 7 – Cladophlebis sp. 2, экз. 3395/349; 9 – Cupressinocladus sp., обрывок побега, экз. 3395/148-2; 10 – Pityophyllum sp., напластование дисперсных листьев, экз. 3395/138; 11, 12 – женские шишки Cupressaceae тип 4 (подсем. Sequoioideae?): 11 – экз. 3395/138-1, 12 – экз. 3395/138-2; 14 – Ginkgo ex gr. digitata Brong., экз. 3395/388-1.

Таблица V. Ископаемые растения аянкинской флоры из местонахождения 701, руч. Звонкий. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 8, 10, 11 ; для фиг. 8, 10 – 0.5 см, для фиг. 11 – 0.1 см. 1, 2, 3 – “Sequoia” sp., облиственные побеги: 1 – экз. 3395/118, 2 – экз. 3395/134, 3 – экз. 3395/135, побег с мужскими шишками; 4 – Elatocladus sp. 2, побег с линейными листьями, экз. 3395/140; 5, 12 – Dicotylophyllum sp. 4, экз. 3395/260: 5 – детали строения края листа, 12 – общий вид листа; 6 – Metasequoia sp. 1, фрагмент облиственного побега, экз. 3395/143; 7 – Pityocladus sp., побег с брахибластами, экз. 3395/147; 8 – Nyssidium (?) sp., экз. 3395/196; 9, 10 – Quereuxia angulata (Newb.) Krysht.: 9 – экз. 3395/237-1б, 10 – экз. 3395/238; 11 – Carpolithes sp., экз. 3395/335-2; 13 – “Macclintockia” ochotica Vachrameev et Herman, экз. 3395/241а.

Таблица VI. Ископаемые растения аянкинской флоры из местонахождения 702, руч. Звонкий. Длина масштабной линейки 1 см для всех изображений, кроме фиг. 12 (0.1 см). 1 – Cladophlebis sp. 1, экз. 3395/370; 2 – Heilungia (?) sp., экз. 3395/373a; 3 – Metasequoia sp. 2, облиственный побег, экз. 3395/152; 4 – Parataxodium cf. wigginsii Arnold et Lowther, полиморфный побег, экз. 3395/150-1; 5, 13 – Сelastrophyllum sp.: 5 – экз. 3395/201a, 13 – экз. 3395/200; 6–8 – Trochodendroides sp. 2: 6 – экз. 3395/156б, 7 – экз. 3395/157, общий вид листа, 8 – то же, увеличенный фрагмент; 9 – Raphaelia (?) sp., экз. 3395/164-2б; 10 – Nyssidium (?) sp., экз. 3395/194; 11 – Metasequoia sp. 1, облиственный побег, экз. 3395/153; 12, 17 – Dicotylophyllum sp. 5, экз. 3395/208: 12 – детали строения края листа, 17 – общий вид листа; 14, 15, 18 – Trochodendroides sp. 1: 14 – экз. 3395/176, 15 – экз. 3395/165б, 18 – экз. 3395/171; 16 – Menispermites sp., экз. 3395/246; 19 – Dicotylophyllum sp. 6, экз. 3395/220-2; 20 – “Macclintockia” ochotica Vachrameev et Herman, экз. 3395/220-1.

Ископаемые растения после очистки и препарирования изучались при помощи налобной бинокулярной лупы и стереомикроскопов Zeiss Stemi 2000 C, оснащенного цифровой камерой Canon G10 Power Shot, и Leica M165C с цифровой камерой Leica DFC 420C. Растительные остатки фотографировали при помощи цифровой камеры Canon EOS 7D, оснащенной макрообъективом Canon EF 1:2.5/50 мм. Рисунки ископаемых растений были выполнены с помощью графического планшета Wacom Intuos3.
СИСТЕМАТИЧЕСКИЙ СОСТАВ АЯНКИНСКОЙ ФЛОРЫ
Как упоминалось выше, аянкинская флора никогда подробно не изучалась. В работе Е.Л. Лебедева (1987) приводился только список предварительных определений ископаемых растений (без изображений), который включал 26 видов: Thallites sp., Equisetites sp., Cladophlebis cf. arctica (Heer) Seward, Cladophlebis sp., Sphenopteris sp., Ginkgo ex gr. adiantoides (Unger) Heer, G. ex gr. sibirica Heer, Phoenicopsis sp., Sphenobaiera sp., Cephalotaxopsis heterophylla Hollick, C. intermedia Hollick, C. magnifolia Fontaine, Sequoia minuta Sveshnikova, Sequoia sp., Metasequoia cuneata (Knowlton) Chaney, Pityolepis sp., Pityospermum sp., Libocedrus catenulata (Bell) Kryshtofovich, Macclintockia ochotica Vachrameev et Herman, Quercus tchucotica Abramova, “Acer” aff. arcticum Heer, Viburnum sp., Pseudoprotophyllum sp., Trochodendroides ex gr. arctica (Heer) Berry, Cissites (?) sp., Dicotyledones spp.
В результате наших исследований состав аянкинской флоры значительно расширился и включает около 60 видов (табл. 1), что более чем в два раза превышает предыдущие оценки разнообразия этой флоры. В целом в ее составе преобладают хвойные и покрытосеменные растения, менее разнообразны папоротники; помимо них встречены единичные хвощовые, печеночники, гинкговые и цикадовые. Хвойные аянкинской флоры довольно разнообразны и многочисленны. Они включают 24 вида, представленных различными органами растений: облиственными побегами (12 видов), дисперсными листьями (1 вид), семенами (1 вид), семенными чешуями (1 вид), а также микро- (3 вида) и макростробилами (6 видов), некоторые из которых находятся в органической связи с побегами. Следует отметить, что многочисленность выделенных таксонов не отражает естественного разнообразия хвойных, так как включает таксоны, выделенные по морфологической классификации для различных органов, которые в сочетании могут принадлежать одному материнскому растению. Начатые детальные исследования хвойных с помощью компьютерной томографии и сканирующей электронной микроскопии показывают объемную сохранность некоторых форм, перспективных для подробных анатомических исследований. Среди покрытосеменных в аянкинской флоре определено 22 вида, представленных в основном отпечатками листьев (20 видов) и плодов (2 вида). В их составе также выявлены новые таксоны, детальное описание которых готовится к публикации. Так как состав комплексов в различных местонахождениях довольно существенно различается, мы рассматриваем их отдельно (табл. 1).
Таблица 1.
Распределение таксонов ископаемых растений по местонахождениям
| № п/п | Название ископаемых растений | Точка 700 | Точка 701 | Точка 702 |
|---|---|---|---|---|
| 1 | Thallites sp. 1 | + | ||
| 2 | Thallites sp. 2 | + | ||
| 3 | Equisetites sp. | + | ||
| 4 | Coniopteris tschuktschorum (Krysht.) Samyl. | + | + | |
| 5 | Sphenopteris sp. | + | ||
| 6 | Cladophlebis sp. 1 | + | + | |
| 7 | Cladophlebis sp. 2 | + | ||
| 8 | Arctopteris sp. 1 | + | ||
| 9 | Arctopteris sp. 2 | + | + | |
| 10 | Raphaelia (?) sp. | + | ||
| 11 | Ginkgo ex gr. adiantoides Heer | + | + | |
| 12 | Ginkgo ex gr. digitata Brong. | + | + | |
| 13 | Ginkgodium (?) sp. | + | ||
| 14 | Heilungia (?) sp. | + | ||
| 15 | Ditaxocladus sp. | + | ||
| 16 | Cupressinocladus sp. | + | ||
| 17 | Parataxodium sp. | + | + | |
| 18 | Parataxodium cf. wigginsii Arnold et Lowther | + | + | |
| 19 | Metasequoia sp. 1 | + | + | + |
| 20 | Metasequoia sp. 2 | + | + | |
| 21 | “Sequoia” sp. | + | ||
| 22 | Женские шишки Cupressaceae тип 1 (подсем. Sequoioideae) |
+ | ||
| 23 | Женские шишки Cupressaceae тип 2 (подсем. Taxodioideae ?) |
+ | ||
| 24 | Женские шишки Cupressaceae (?) тип 3 | + | ||
| 25 | Женские шишки Cupressaceae тип 4 (подсем. Sequoioideae?) |
+ | ||
| 26 | Мужские шишки Cupressaceae тип 1 | + | ||
| 27 | Мужские шишки Cupressaceae тип 2 | + | + | |
| 28 | Мужские шишки Cupressaceae тип 3 | + | ||
| 29 | Pityophyllum sp. | + | + | |
| 30 | Pityospermum sp. | + | ||
| 31 | Pityostrobus sp. 1 | + | ||
| 32 | Pityostrobus sp. 2 | + | ||
| 33 | Pityocladus sp. | + | + | |
| 34 | Pityolepis sp. | + | ||
| 35 | Taxites sp. | + | ||
| 36 | Pagiophyllum sp. | + | ||
| 37 | Elatocladus sp. 1 | + | ||
| 38 | Elatocladus sp. 2 | + | ||
| 39 | Monocotylophyllum sp. | + | ||
| 40 | “Vitis” penzhinica Herman | + | ||
| 41 | Trochodendroides notabilis Herman | + | ||
| 42 | Trochodendroides sp. 1 | + | + | |
| 43 | Trochodendroides sp. 2 | + | ||
| 44 | “Macclintockia” ochotica Vachr. et Herman | + | + | + |
| 45 | Cissites sp. | + | ||
| 46 | Menispermites sp. | + | + | |
| 47 | Celastrinites sp. | + | ||
| 48 | Celastrophyllum sp. | + | ||
| 49 | Viburniphyllum sp. | + | ||
| 50 | Фрагменты Platanoids (?) | + | ||
| 51 | Dicotylophyllum sp. 1 | + | ||
| 52 | Dicotylophyllum sp. 2 | + | ||
| 53 | Dicotylophyllum sp. 3 | + | ||
| 54 | Dicotylophyllum sp. 4 | + | ||
| 55 | Dicotylophyllum sp. 5 | + | ||
| 56 | Dicotylophyllum sp. 6 | + | ||
| 57 | Quereuxia angulata (Newb.) Krysht. | + | ||
| 58 | Phragmites sp. | + | ||
| 59 | Nyssidium sp. | + | + | |
| 60 | Carpolites sp. | + |
В местонахождении 700 наблюдается наибольшее разнообразие видов ископаемых растений (табл. 1). Следует отметить, что их остатки здесь были собраны из нескольких флороносных слоев (рис. 2) в двух отстоящих друг от друга обнажениях. Из этого местонахождения определены два вида печеночников, которые характеризуются довольно крупными ветвящимися талломами. Для Thallites sp. 1 (табл. I , фиг. 1, 2) характерно моноподиальное ветвление таллома, боковые ответвления длиной более 7 см, с неровным лопастным (?) краем, шириной до 15 мм. У Thallites sp. 2 талломы ветвятся дихотомически, края слоевища нелопастные, волнистые или зазубренные, ширина фрагментов около 1 см (табл. I , фиг. 3).
Рис. 3.
Ископаемые покрытосеменные из аянкинской флоры бассейна р. Обрывистая (местонахождения 700 и 702). Длина масштабной линейки 1 см. а–в – Cissites sp.: a – экз. 3395/270a, б – экз. 3395/380, в – экз. 3395/381-1; г–е – “Macclintockia” ochotica Vachrameev et Herman: г – экз. 3395/220-1, д – экз. 3395/286б, е – экз. 3395/220-1, увеличенный фрагмент; ж, к, л – Trochodendroides sp. 1: ж – экз. 3395/165б, к – экз. 3395/166, л – экз. 3395/167б; з, и – “Vitis” penzhinica Herman: з – экз. 3395/266, и – экз. 3395/382б; м – Trochodendroides sp. 2, экз. 3395/156б.
Папоротники представлены стерильными и фертильными перьями Coniopteris tschuktschorum (Kryshtofovich) Samylina (табл. I , фиг. 8 ), а также несколькими небольшими фрагментами стерильных перьев Arctopteris sp. 1 с довольно крупными перышками (табл. I , фиг. 4), которые сходны с видом A. ilirnensis, описанным из чаунской и ульинской флор ОЧВП (Головнева, 2018; Akinin et al., 2019). Похожие папоротники были встречены и в усть-эмунеретской флоре Центральной Чукотки и были отнесены нами ранее к Arctopteris aff. rarytkensis Vassilevskaja (Моисеева, Соколова, 2014). Фрагменты стерильных листьев с более мелкими перышками (табл. I , фиг. 5 ) отнесены к другому виду Arctopteris sp. 2. Кроме того, в местонахождении 700 единично встречаются фрагменты стерильных дваждыперистых листьев Cladophlebis sp. 1 (табл. I , фиг. 9 ).
Рис. 4.
Проникновение тополей по речным косам в местность с расчлененным рельефом, населенную преимущественно хвойной растительностью. Тибет, верховья р. Меконг (по Щепетов и др., 2019, с изменениями).

Немногочисленные гинкговые отнесены из-за отсутствия сохранившихся фитолейм к двум сборным видам: Ginkgo ex gr. adiantoides Heer (слабо рассеченные листья) (табл. I , фиг. 10 ) и Ginkgo ex gr. digitata Brongniart (сильно рассеченные листья) (табл. I , фиг. 13 ), а также к Ginkgodium (?) sp. (табл. I , фиг. 7 ).
Среди хвойных в точке 700 доминируют представители сем. Cupressaceae sensu lato. Преобладают многочисленные побеги Parataxodium cf. wigginsii Arnold et Lowther (табл. II, фиг. 1, 2, 8 ), сходные с побегами типового вида рода Parataxodium из сантон(?)- маастрихтских тафофлор Ранняя и Поздняя Когосакрак Северной Аляски (Arnold, Lowther, 1955). Объем этого вида был недавно пересмотрен (Rothwell et al., 2020). К этому же роду нами отнесены побеги Parataxodium sp. (табл. II, фиг. 3), также характеризующиеся сочетанием очередного и супротивного листорасположения, но отличающиеся от Parataxodium cf. wigginsii по форме листьев и частоте их расположения на побеге. В комплексе местонахождения 700 часто встречаются конечные побеги рода Metasequoia Miki ex Hu et Cheng со строго супротивным листорасположением двух типов: Metasequoia sp. 1 (табл. II, фиг. 7 ) и Metasequoia sp. 2 (табл. II, фиг. 10 ). Они различаются по форме листьев (ланцетовидные с заостренной верхушкой у первого и эллипсоидальные с широко-округлой верхушкой у второго вида) и углу их прикрепления к побегу (25°–45° у первого и 70°–90° у второго вида). Надо отметить, что, возможно, часть побегов первого типа может относиться к Parataxodium cf. wigginsii.
Особый интерес вызывают генеративные органы хвойных, встреченные в этой точке. Из кипарисовых можно отметить дисперсные женские шишки трех типов, первый из которых (табл. II, фиг. 5 ) определенно относится к подсемейству Sequoioideae, второй (табл. I , фиг. 6 ), криптомероидного типа, вероятно, принадлежит к подсемейству Taxodioideae, а третий (табл. I , фиг. 14 ) имеет неопределенное на данном этапе исследования положение в рамках семейства Cupressaceae sensu lato. Как принадлежащие семейству Cupressaceae s.l. мы также рассматриваем три разновидности фертильных побегов с мужскими шишками, условно названные тип 1 (табл. II, фиг. 13 ), тип 2 (табл. II, фиг. 9 ) и тип 3 (табл. I , фиг. 15 ). Они различаются по размеру мужских шишек, способу их прикрепления к побегам и морфологии листьев, расположенных на побегах. В местонахождении 700 нередко встречаются побеги семейства Cupressaceae s.s., отнесенные к Ditaxocladus sp. (табл. I , фиг. 11, 12 ).
Из хвойных сем. Pinaceae установлены крупные женские шишки двух видов, различающиеся по форме и отнесенные к Pityostrobus sp. 1 (табл. II, фиг. 15 ) и Pityostrobus sp. 2 (табл. II, фиг. 11 ), а также единичные дисперсные чешуи Pityolepis sp. (табл. II, фиг. 16 ), семена Pityospermum sp. (табл. II, фиг. 14 ) и побеги Pityocladus sp. (табл. II, фиг. 4).
Еще один характерный компонент тафофлоры местонахождения 700 – это крупные побеги с длинными линейно-ланцетными листьями, определенные формально как Taxites sp. (табл. II, фиг. 12 ). Такие побеги являются частыми компонентами меловых флор Северного полушария и в разных работах определяются в рамках ископаемых родов Cephalotaxopsis Fontaine (Ефимова, 1966; Филиппова, Абрамова, 1993), Elatocladus Halle (Bell, 1949), Taxites (Самылина, 1988) или в рамках современного рода Taxodium Richard (Головнева, Щепетов, 2011; Herman et al., 2016; Головнева, 2018; Щепетов и др., 2019). Мы воздерживаемся от отнесения изученных ископаемых хвойных к современным родам, так как для этого нет достаточных оснований: отсутствуют данные по строению эпидермы и/или фертильных органов, найденных в органической связи с побегами. Морфология этих побегов, помимо Cupressaceae sensu lato, вполне типична и для представителей других семейств хвойных, например Taxaceae. Поэтому мы считаем, что уместнее рассматривать остатки такой сохранности в рамках ископаемых родов без привязки к семействам современных хвойных (например, род Taxites). Для типового вида Cephalotaxopsis было изучено эпидермальное строение, указывающее на его принадлежность к семейству Taxaceae (Berry, 1911; Florin, 1958), поэтому данное название не подходит для изученных нами ископаемых побегов.
Из хвойных неясного систематического положения в комплексе встречены многочисленные длинные и узкие не полностью сохранившиеся листья Pityophyllum sp., которые образуют листовые скопления на плоскостях напластования. Такие листья являются частыми компонентами позднемеловых флор Северо-Востока Азии. Помимо них, единично представлены побеги с линейными листьями Elatocladus sp. 1 (табл. II, фиг. 6 ).
Наряду с хвойными, в точке 700 доминируют покрытосеменные растения, которые представлены 13 видами. В данном комплексе наиболее многочисленны отпечатки листьев “Vitis” penzhinica Herman (табл. III, фиг. 1, 2, 4; рис. 3з, 3и) – характерного вида для сантон-кампанских флор Северо-Западной Камчатки и бухты Угольной (Чукотка) (Герман, Лебедев, 1991; Моисеева, Соколова, 2011). Похожие листья также были описаны из позднемеловой (сантон?-маастрихтской) флоры Северной Аляски как Archaeampelos sp. (Rothwell et al., 2020). В захоронении доминируют отпечатки лопастных листьев Cissites sp. (табл. III, фиг. 5 , рис. 3а–3в), которые, вероятно, будут отнесены к новому виду. Вид “Macclintockia” ochotica Vachrameev et Herman (табл. III, фиг. 7 , рис. 3д) также является характерным представителем сантон-кампанских флор Северо-Востока России. Недавно Л.Б. Головнева с соавторами (Головнева и др., 2017) перевели этот вид в род Trochodendroides Berry, но, с нашей точки зрения, характеристика этого вида не вполне соответствует диагнозу данного рода (Berry, 1922; Crane, 1984). Родовая принадлежность этого вида требует дальнейшего уточнения, на данном этапе мы пока оставляем его первоначальное название, беря его в кавычки. Представители рода Trochodendroides в местонахождении 700 представлены двумя видами. К T. notabilis Herman отнесены листья с двоякогородчатым краем листа (табл. III, фиг. 8, 13 ). Кроме них, встречаются довольно мелкие листья с немногочисленными округлыми зубцами или с волнистым краем, которые определены как Trochodendroides sp. 1 (табл. III, фиг. 3). Вид T. notabilis изначально был описан из турон-коньякских отложений Северо-Западной Камчатки (Герман, Лебедев, 1991), но также встречается и в раннекампанской барыковской флоре бухты Угольной, Чукотка (Моисеева, Соколова, 2011).
Кроме перечисленных выше видов, в местонахождении 700 присутствуют фрагменты пельтатных листьев с волнистым краем Menispermites sp. (табл. III, фиг. 14, 15 ), цельнокрайние листья с рельефным брохидодромным жилкованием Celastrinites sp. (табл. III, фиг. 6 ), а также фрагменты листьев с двоякозубчатым краем и перистым семикраспедодромным жилкованием Viburniphyllum sp. (табл. III, фиг. 11, 12 ). Единично встречаются фрагменты крупных платаноидных листьев (табл. III, фиг. 16 ), а также отпечатки листьев двудольных неясного систематического положения Dicotylophyllum sp. 1–3 (табл. III, фиг. 9, 10 ). К однодольным Monocotylophyllum sp. отнесены узкие линейные листья с округлой верхушкой, довольно толстой средней жилкой первого порядка и многочисленными тонкими параллельными жилками второго порядка (табл. I , фиг. 15, 16 ). Необходимо отметить, что, судя по описанию аянкинского флористического комплекса в работах Лебедева (1987, 1992) и записям в полевом дневнике А.Б. Германа, в местонахождении 700 были найдены единичные листья Barykovia tchucotica (Abramova) Moiseeva, первоначально описанные как Quercus tchucotica Abramova (Абрамова, 1979); к сожалению, эти образцы были впоследствии утеряны.
Разнообразие ископаемых растений в местонахождении 701 небольшое (24 вида), причем в нем как по количеству экземпляров, так и по разнообразию преобладают хвойные. Среди хвощовых определены единичные Equisetites sp. (табл. IV, фиг. 4). Папоротники так же, как в точке 700, представлены видами Coniopteris tschuktschorum (табл. IV, фиг. 6 ) и Arctopteris sp. 2 (табл. IV, фиг. 5, 13, 15 ). Последний вид представлен в данном местонахождении фертильными и стерильными листьями. Единичные фрагменты конечных стерильных перьев отнесены к Sphenopteris sp. (табл. IV, фиг. 3) и Cladophlebis sp. 2 (табл. IV, фиг. 7 ); последний отличается от Cladophlebis sp. 1, встреченного в точке 700, более крупными перышками серповидной формы, а также их жилкованием: прямой средней жилкой и ветвящимися боковыми жилками. Из гинкговых так же, как в точке 700, обнаружены листья со слабо рассеченной пластинкой Ginkgo ex gr. adiantoides и сильно рассеченной пластинкой Ginkgo ex gr. digitata (табл. IV, фиг. 14 ).
Среди хвойных в роли абсолютных доминантов, встреченных только в местонахождении 701, выступают облиственные полиморфные побеги, определенные нами как “Sequoia” sp. (табл. IV, фиг. 1; табл. V, фиг. 1, 2). Взятие родового названия в кавычки в данном случае подразумевает, что мы не имеем в виду отнесение ископаемых остатков к современному роду Sequoia Endlicher, поскольку неоднократно высказывалось сомнение в существовании этого таксона в меловом периоде (Rothwell, Ohana, 2016; Sokolova et al., 2017). К этому же растению, вероятно, относится побег с мужскими шишками (табл. V, фиг. 3) и, возможно, женские шишки секвойевого (?) типа (табл. IV, фиг. 11, 12 ). Кроме того, из местонахождения 701 определены побеги Metasequoia sp. 1 (табл. V, фиг. 6 ). К Cupressaceae s.s. относится единственный побег Cupressinocladus sp. (табл. IV, фиг. 9 ). Как и в местонахождении 700, в точке 701 встречаются дисперсные линейные листья Pityophyllum sp. (табл. IV, фиг. 10 ) и побеги Pityocladus sp. (табл. V, фиг. 7 ). Из характерных таксонов только этого местонахождения надо отметить хвойное неясного систематического положения Pagiophyllum sp. (табл. IV, фиг. 2, 8 ). Единично встречаются побеги с линейными листьями Elatocladus sp. 2 (табл. V, фиг. 4), отличающиеся от побегов Elatocladus sp. 1 из точки 700 прямыми, а не изогнутыми линейными листьями с длинно-заостренной верхушкой треугольной формы, в отличие от коротко-заостренной верхушки у Elatocladus sp. 1.
Покрытосеменные в местонахождении 701 редки и фрагментарны. Они представлены отпечатками листьев водного растения Quereuxia angulata (Newberry) Kryshtofovich (табл. V, фиг. 9, 10 ), фрагментами листьев плохой сохранности “Macclintockia” ochotica (табл. V, фиг. 13 ), Dicotylophyllum sp. 4 с мелкопильчатым краем листа (табл. V, фиг. 5, 12 ) и Phragmites sp., а также мелкими плодами Nyssidium (?) sp. (табл. V, фиг. 8 ) и Carpolithes sp. (табл. V, фиг. 11 ).
В местонахождении 702 разнообразие тоже небольшое, но, в отличие от точки 701, здесь в этом флористическом комплексе преобладают покрытосеменные растения. Кроме них, встречаются только единичные фрагменты папоротников C-ladophlebis sp. 1 (табл. VI, фиг. 1) и Raphaelia (?) sp. (табл. VI, фиг. 9 ), листьев цикадовых Heilungia (?) sp. (табл. VI, фиг. 2) и единичные побеги хвойных Metasequoia sp. 1 (табл. VI, фиг. 11 ), Metasequoia sp. 2 (табл. VI, фиг. 3), Parataxodium cf. wigginsii (табл. VI, фиг. 4) и Parataxodium sp. Среди покрытосеменных доминируют полиморфные листья рода Trochodendroides. К Trochodendroides sp. 1 (табл. VI, фиг. 14, 15, 18 ; рис. 3ж, 3к, 3л) отнесены мелкие листья с изменчивой формой пластинки и краем листа, который варьирует от волнистого или нерегулярно-зубчатого до городчатого, с небольшим количеством относительно крупных зубцов. Листья Trochodendroides sp. 2 характеризуются мелкопильчатым краем с железистыми зубцами 2–3 порядков (табл. VI, фиг. 6–8 ; рис. 3м). Также в этом захоронении преобладают отпечатки листьев “Macclintockia” ochotica (табл. VI, фиг. 20 ; рис. 3г, 3е). Реже встречаются отпечатки небольших листьев Menispermites sp. (табл. VI, фиг. 16 ) и Сelastrophyllum sp. (табл. VI, фиг. 5, 13 ). Единично представлены плоды Nyssidium (?) sp. (табл. VI, фиг. 10 ), а также листья покрытосеменных неясного систематического положения Dicotylophyllum sp. 5 (табл. VI, фиг. 12, 17 ) и Dicotylophyllum sp. 6 (табл. VI, фиг. 19 ).
СРАВНЕНИЕ С ДРУГИМИ ПОЗДНЕМЕЛОВЫМИ ФЛОРАМИ И ВОЗРАСТ АЯНКИНСКОЙ ФЛОРЫ
Аянкинская флора по составу имеет наибольшее сходство с раннекампанской барыковской флорой района бухты Угольной (Моисеева, Соколова, 2011). Барыковская флора надежно датирована по соотношению флороносных отложений с морскими, содержащими остатки морских моллюсков (Пергамент, 1974; Герман, 2011). Сходство среди хвойных этих флор проявляется в наличии многочисленных побегов с длинными линейно-ланцетными листьями, определенных формально в данной работе как Taxites sp., а в барыковской флоре как Elatocladus gracillimus (Hollick) Sveshnikova. Кроме того, в обеих флорах встречается несколько морфологически сходных видов побегов метасеквойевого и секвойевого облика и характерные женские шишки подсемейства Sequoioideae. Также в аянкинской и барыковской флорах установлены женские шишки сосновых рода Pityostrobus Nathorst и побеги рода Cupressinocladus Seward. Из покрытосеменных в обоих комплексах встречаются следующие таксоны: “Vitis” penzhinica, “Macclintockia” ochotica, Trochodendroides notabilis, а также представители родов Barykovia Moiseeva, Cissites Debey in Capellini et Heer, Menispermites Lesquereux и Celastrinites Saporta. Следует отметить наличие в аянкинском и барыковском комплексах крупных платаноидных листьев, водных покрытосеменных Quereuxia angulata, цикадовых (представленных разными родами), гинкговых и папоротников Arctopteris.
Аянкинская флора отличается от барыковской в целом более бедным составом, менее разнообразными папоротниками и цикадовыми, разными доминантами в составе как хвойных (Parataxodium и “Sequoia” sp.), так и покрытосеменных. Среди последних в барыковской флоре преобладают крупнолистные платаноиды, Barykovia tchucotica, довольно много цельнокрайних и крупнолистных таксонов, тогда как в аянкинской флоре доминируют мелкие формы Trochodendroides, “Vitis” penzhinica и Cissites.
По составу покрытосеменных растений наблюдается сходство аянкинской флоры с позднебыстринской флорой Северо-Западной Камчатки (Герман, Лебедев, 1991; Герман, 2011). Возраст флороносных слоев верхнебыстринской подсвиты установлен как ранний кампан (возможно, включая поздний сантон и средний кампан), исходя из возраста подстилающих и перекрывающих ее морских образований (Герман, Лебедев, 1991; Герман, 2011). В обеих флорах встречаются “Vitis” penzhinica, “Macclintockia” ochotica, Quereuxia angulata, а также представители родов Barykovia, Trochodendroides, Cissites, Celastrinites, Viburniphyllum Nathorst, отмечается незначительное участие платаноидов. Папоротники довольно редки в позднебыстринской флоре, но так же, как и в аянкинской, среди них встречены представители рода Arctopteris. Хвойные растения из позднебыстринской флоры, к сожалению, до сих пор не были детально изучены, поэтому их сравнение с аянкинскими пока не проводилось. По предварительным определениям А.Б. Германа (2011), для позднебыстринской флоры характерно присутствие видов Cephalotaxopsis heterophylla Hollick, Sequoia sp., Metasequoia cuneata (Newberry) Chaney и Cupressinocladus cretaceus (Heer) Seward.
В целом аянкинская флора имеет сходство с усть-эмунеретской флорой из вулканогенных образований Центральной Чукотки (Моисеева, Соколова, 2014; Моисеева, 2015; Соколова, Моисеева, 2016; Nosova et al., 2020). Возраст усть-эмунеретской флоры определен по последним изотопным данным в пределах сантона–раннего кампана (Tikhomirov et al., 2012). Среди общих таксонов установлены печеночники Thallites sp. 1 (неопубликованные данные авторов), папоротники Arctopteris и Coniopteris tschuktschorum, гинкговые Ginkgo ex gr. adiantoides и Ginkgodium. Остатки хвойных довольно разнообразны в обоих комплексах, среди них определены семена Pityospermum Nathorst, чешуи Pityolepis Nathorst, листья Pityophyllum Nathorst, побеги Cupressinocladus, а также побеги “Sequoia” sp. Интересно наличие в обеих флорах побегов с мужскими шишками, которые отнесены к “Sequoia” sp. в аянкинской флоре и определены как Glyptostrobus comoxensis Bell в усть-эмунеретской. Побеги с мужскими шишками такого типа – частые компоненты позднемеловых флор Северной Азии и Северной Америки (Bell, 1956; Красилов, 1979). Представители рода Parataxodium также установлены в обеих флорах, но если для аянкинской – это основной из доминантов, то в усть-эмунеретской встречен единственный экземпляр. По составу покрытосеменных сходство проявляется в преобладании мелколистных видов Trochodendroides; характерным компонентом являются “Macclintockia” ochotica, Barykovia tchucotica и представители родов Cissites, Menispermites, Viburniphyllum; платаноиды отсутствуют в усть-эмунеретской и лишь единично встречаются в аянкинской флоре; в целом среди покрытосеменных обеих флор преобладают мелколистные таксоны.
Аянкинская флора отличается от усть-эмунеретской почти полным отсутствием реликтовых форм среди гинкговых и чекановскиевых (таких как Sphenobaiera ex gr. pulchella (Heer) Florin, S. ex gr. longifolia (Pomel) Florin, Ginkgo insperata E. Lebedev, Phoenicopsis chukotkensis Nosova), преобладанием хвойных с линейно-ланцетными листьями Taxites, Parataxodium, Metasequoia и Elatocladus. В усть-эмунеретской флоре, напротив, доминируют побеги хвойных Sequoiadendron tchucoticum A. Sokolova с листьями разнообразной формы – от чешуевидных до серповидных (Соколова, Моисеева, 2016), которые не встречены в аянкинской флоре. Также в аянкинской флоре не установлены характерные виды покрытосеменных усть-эмунеретской флоры: Cissites reticulatus Moiseeva, Viburniphyllum emuneretum Moiseeva и Dicotylophyllum stipulare Moiseeva (Моисеева, 2014).
Определенное сходство аянкинской флоры, особенно по составу хвойных и покрытосеменных из местонахождения 700, проявляется с флорой Ранняя Когосакрак из свиты Принс Крик Северной Аляски (Arnold, Lowther, 1955; Herman, 2013; Rothwell et al., 2020; http://arcticfossils.nsii.org.cn/). Сантон?-кампанский возраст данной флоры определен по взаимоотношению флороносных отложений с замещающими их по простиранию морскими слоями (Detterman, Bickel, 1963; Frederiksen, McIntyre, 2000; http://arcticfossils.nsii.org.cn/). Помимо основного компонента среди хвойных – побегов Parataxodium wigginsii – в обеих флорах есть сходные фертильные остатки хвойных, включающие несколько типов женских шишек секвойевого и криптомероидного облика и мужских шишек семейства Cupressaceae, а также дисперсные листья Pityophyllum. Покрытосеменные во флоре Ранняя Когосакрак встречаются крайне редко и в основном представлены фрагментарно, но следует отметить, что и среди них были установлены таксоны, сходные с аянкинскими. В частности, к ним относятся листья, определенные нами в аянкинской флоре как “Vitis” penzhinica, а во флоре Северной Аляски – как Archаeampelos sp. (Rothwell et al., 2020). Мелкий лист двудольного морфотипа 2 (Rothwell et al., 2020, fig. 6.5 ) с крупными железистыми зубцами и актинодромным (вероятно, акродромным) жилкованием имеет сходство с аянкинским Trochodendroides sp. 1. А фрагмент листа двудольного морфотипа 3 (Rothwell et al., 2020, fig. 7.3–7.5 ) очень похож на верхушку листа Barykovia tchucotica. Кроме того, во флоре Ранняя Когосакрак, как и в аянкинской, установлены Quereuxia angulata и остатки платаноидных листьев, отнесенные в аляскинской флоре к роду Paraprotophyllum Herman (Герман, 2011).
Интересно также было провести сравнение аянкинской и ольской флор (Самылина, 1988; Филиппова, Абрамова, 1993; Герман, 2011; Щепетов и др., 2019). Возраст последней по изотопным данным определен как кампан (80–83 млн лет; Акинин, Миллер, 2011). В составе ольской флоры, в отличие от аянкинской, преобладают разнообразные хвойные растения. Сравнение этих комплексов показывает их определенное сходство по морфологическим типам хвойных. Так, в обеих флорах встречены сходные представители семейства Pinaceae: шишечные чешуи рода Pityolepis, дисперсные семена Pityospermum, женские шишки Pityostrobus и побеги с брахибластами Pityocladus (Nathorst) Seward. Также в этих флорах установлены дисперсные листья Pityophyllum. Из кипарисовых общими являются побеги рода Metasequoia, и встречаются сходные женские шишки секвойевого типа (Sequoioideae), определенные в ольской флоре как Pityostrobus sp. (Щепетов и др., 2019, табл. 14 , фиг. 13, 14 ). Часть побегов Tollia cf. cunninghamoides Sveshnikova et Budantsev (Щепетов и др., 2019, табл. 16 , фиг. 1, 2) из ольской флоры напоминает аянкинские секвойеподобные побеги из точки 701 (табл. V, фиг. 1). Следует также отметить сходство крупных побегов с длинными линейными листьями, определенных нами как Taxites sp., а в ольской флоре – под названиями Taxodium amguemensis (Efimova) Golovneva и Elatocladus kukenliveemsis Philippova. В целом эти флоры различаются по основным доминантам: в аянкинской флоре преобладают Parataxodium cf. wigginsii и “Sequoia” sp., а в ольской – Taiwania cretacea Samylina, Tollia cf. cunninghamoides и Elatocladus communis Philippova. Кроме того, в аянкинской флоре не встречены характерные таксоны ольской флоры Cunninghamia orientalis (Philippova) Samylina, Pagiophyllum sp. и др. (Щепетов и др., 2019). Основным отличием сравниваемых флор также является преобладание разнообразных покрытосеменных в аянкинской флоре, среди которых в ольской установлены только Quereuxia angulata, мелкие листья Trochodendroides, Macclintockia beringiana Herman и немногочисленные фрагменты двудольных неясного систематического положения. Из папоротников встречен только один общий вид – Coniopteris tschuktschorum.
В гораздо меньшей степени наблюдается сходство аянкинской флоры с турон-коньякской арманской флорой Магаданской области (Herman et al., 2016). Среди общих таксонов хвойных можно отметить только Taxites sp., фигурирующий в арманской флоре под названием Taxodium amguemensis, а также побеги “Sequoia” sp. (которые доминируют в точке 701), отнесенные в арманской флоре к виду Sequoia armanensis Golovneva (Herman et al., 2016). Кроме того, в обеих флорах установлены сходные по морфологии женские шишки Pityostrobus sp., семена Pityospermum sp., дисперсные листья Pityophyllum. Среди покрытосеменных отдаленное сходство можно проследить только на родовом уровне: это роды Trochodendroides, Cissites, Menispermites, Celastrophyllum. Аянкинская флора существенно отличается по составу папоротников, в ней отсутствуют арманские реликтовые гинкговые (Sphenobaiera Florin), чекановскиевые (Czekanowskia Heer, Phoenicopsis Heer, Leptostrobus Heer), цикадовые (Nilssonia Brongniart) и беннетитовые (Pterophyllum Brongniart). Среди хвойных в арманской флоре также присутствуют реликтовые таксоны, такие как два вида рода Podozamites F. Braun и Elatocladus smittiana (Heer) Seward, не встреченные в аянкинской флоре.
Установлено лишь незначительное сходство при сравнении аянкинской флоры с чаунской, которая происходит из вулканогенных образований одноименной серии Пегтымельского прогиба Центральной Чукотки (Щепетов, 1991; Головнева, 2018). Возраст чаунской флоры определен по последним изотопным данным как коньякский (около 88 млн лет; Kelley et al., 1999). Для чаунской флоры характерно значительное разнообразие среди папоротников, из которых общими c аянкинскими являются лишь Coniopteris tschuktschorum и Arctopteris. Характерными элементами чаунской флоры являются папоротники Tchaunia tchaunensis Samylina et Philippova, T. lobifolia Philippova, Kolymella raevskii Samylina et Philippova, которые не установлены в аянкинской флоре. Цикадовые также более разнообразны в чаунской флоре (несколько видов Heilungia Prynada и Ctenis Lindley et Hutton), тогда как в аянкинской встречены лишь единичные фрагменты Heilungia. При сравнении хвойных обращает на себя внимание сходство на родовом уровне представителей семейства Pinaceae (Pityostrobus, Pityolepis, Pityospermum), а также дисперсных линейных листьев Pityophyllum, часто образующих в обоих местонахождениях скопления. Следует отметить, что сосновые в чаунской флоре значительно более разнообразны по видовому составу по сравнению с аянкинскими. Общими для двух флористических комплексов также являются побеги, определенные нами как Taxites sp. (Taxodium amguemensis в чаунской флоре), а также побеги Metasequoia и секвойеподобные женские шишки (сем. Sequoioideae), отнесенные в чаунской флоре к Sequoia ochotica Yudova et Golovneva (Головнева, 2018). Покрытосеменные очень редки в чаунской флоре, они в основном представлены мелкими листьями Trochodendroides microphylla Philippova и Quereuxia angulata. В отличие от аянкинской флоры, в чаунской присутствуют многочисленные реликты: Phoenicopsis, Leptostrobus, Sphenobaiera, Podozamites, а также характерные хвойные родов Araucarites Presl, Elatocladus и Pagiophyllum Heer, формирующие более древний облик этой флоры.
Аунейский флористический комплекс, который происходит из вулканогенных отложений аунейской толщи в среднем течении р. Еропол (бассейн р. Анадырь) (Головнева, Щепетов, 2013), близок по составу к чаунской флоре и имеет мало общего с аянкинской флорой. Последняя существенно отличается от аунейской отсутствием среди папоротников характерных родов Tchaunia Samylina et Philippova и Kolymella Samylina et Philippova, а также реликтовых таксонов хвойных Podozamites, Elatocladus zheltovskii Philippova и Araucarites subacutensis Philippova; доминированием среди хвойных Parataxodium cf. wigginsii и разнообразных фертильных побегов кипарисовых; преобладанием покрытосеменных растений. Из общих таксонов в обеих флорах установлены сходные представители сосновых и секвоид, а также покрытосеменные Quereuxia angulata и мелкие листья Trochodendroides, один из которых (Головнева, Щепетов, 2013, табл. I , фиг. 5 ) имеет сходство с Trochodendroides sp. 1 из аянкинской флоры.
Также нами было проведено сравнение аянкинской флоры с ульинской флорой ОЧВП из амкинской свиты Ульинского прогиба, возраст которой по последним изотопным данным определен как самый конец коньяка (Akinin et al., 2019). Среди папоротников близким таксоном является только Arctopteris. Из общих таксонов хвойных установлены Ditaxocladus S.X. Guo et Z.H. Sun и Cupressinocladus, сходные побеги метасеквойевого и секвойевого типов, а также женские шишки сосновых Pityostrobus, чешуи Pityolepis и дисперсные листья Pityophyllum. Кроме того, в местонахождениях присутствуют крупные побеги с длинными линейными листьями Taxites sp. (Taxodium amguemensis в ульинской флоре). Среди покрытосеменных общими являются лишь Quereuxia angulata и различные виды рода Trochodendroides. Так же как чаунская, аунейская и арманская флоры, ульинская флора включает целый ряд реликтовых таксонов (Hausmannia Dunker, Podozamites, Phoenicopsis, Baiera Braun, Sphenobaiera). Характерные виды ульинской флоры, такие как Tchaunia, Sagenopteris Presl, Araucarites sheikashoviae Golovneva, Elatocladus amkensis Golovneva, Elatocladus gyrbykensis Golovneva и Pagiophyllum umitbaevii Golovneva, в аянкинской флоре не встречены.
Таким образом, детальное сравнение аянкинской флоры показало ее несомненное сходство с сантон-кампанскими флорами Северо-Востока России и Северной Аляски (с барыковской, позднебыстринской, усть-эмунеретской, в меньшей степени с ольской и Ранней Когосакрак), что позволяет нам сделать вывод о наиболее вероятном сантон-кампанском возрасте аянкинской флоры.
РАСТИТЕЛЬНЫЕ СООБЩЕСТВА АЯНКИНСКОЙ ФЛОРЫ
Флороносные отложения изученных местонахождений аянкинской флоры примерно одновозрастны, однако составы ископаемых растений из них заметно различаются (табл. 1): во всех трех местонахождениях встречаются лишь виды Metasequoia sp. 1 и “Macclintockia” ochotica, еще 13 видов были встречены в двух местонахождениях. Наиболее существенные различия наблюдаются в составе растений из точек 701 и 702: общими для них являются лишь четыре вида, причем в местонахождении 701 по разнообразию и количеству экземпляров абсолютно преобладают хвойные, а находки покрытосеменных единичны, тогда как в местонахождении 702, наоборот, преобладают покрытосеменные растения. Интересно при этом отметить, что в составе тафофлоры точки 701 больше, чем в составе тафофлоры точки 702, относительно древних растений, более характерных для флор раннего и начала позднего мела. К ним можно отнести Coniopteris tschuktschorum, Ginkgo ex gr. digitata, представителей родов Arctopteris, Pagiophyllum и Elatocladus.
Вероятно, в условиях расчлененного рельефа ОЧВП одновременно существовали растительные сообщества, значительно различавшиеся составом входивших в них растений и населявшие разные местообитания: как и во многих вариантах современной растительности (рис. 4), в долинах рек или по берегам озер преобладали покрытосеменные растения, а на склонах и возвышенностях доминировали хвойные. Флористический комплекс местонахождения 701, скорее всего, отражает склоновую растительность, тогда как растения, остатки которых были встречены в местонахождении 702, вероятно, населяли локально пониженные участки рельефа – берега реки или озера.
Объяснение большего участия в тафофлоре местонахождения 701 относительно древних растений мы видим в следующем. Ведущая роль в позднемеловом флорогенезе Северной Пацифики принадлежала новым кайнофитным сообществам с доминированием покрытосеменных, постепенно проникавших в мезофитные флоры (Herman, 2002; Герман, 2011; Щепетов и др., 2019). В течение альба эти сообщества, очевидно, заселили нарушенные прирусловые местообитания приморских низменностей Северо-Востока Азии и Аляски, в пределах которых растительный покров периодически уничтожался морскими трансгрессиями, миграциями русел меандрирующих рек и латеральной эрозией. В течение позднемеловой эпохи кайнофитные сообщества проникали вдоль речных долин и других нестабильных местообитаний вглубь Азиатского континента – на вулканическое нагорье ОЧВП и во внутриконтинентальные районы Азии – и постепенно вытесняли существовавшую там преимущественно хвойно-папоротниковую растительность.
Большинство ископаемых флор из меловых образований ОЧВП по систематическому составу существенно отличается от одновозрастных флор приморских низменностей. Вполне вероятно, что растительный покров этой обширной вулканической области восстанавливался после массовых извержений в основном за счет пула местных видов, а отсутствие конкуренции со стороны покрытосеменных способствовало длительному сохранению в составе таких флор относительно древних групп растений и появлению на их основе некоторых новых таксонов (Щепетов и др., 2019). Аянкинская флора представляет собой яркое свидетельство внедрения из прилегающих с востока приморских низменностей растительных сообществ с доминированием продвинутых покрытосеменных на территорию вулканического нагорья, населенную в позднем мелу сообществами, в которых доминировали папоротники и хвойные и еще сохранились растения, характерные для флор раннего и начала позднего мела.
В отличие от местонахождений 701 и 702, в которых ископаемые растения были собраны из небольших по мощности и протяженности обнажений, растительные остатки из местонахождения 700 происходят из нескольких флороносных слоев в двух отстоящих друг от друга обнажениях. Этим объясняется и более разнообразный состав ископаемых растений местонахождения 700, и отсутствие явно выраженной доминирующей группы. Со значительной долей уверенности можно предположить, что флористический комплекс этого местонахождения включает остатки растений, слагавших разные (два или более) естественные растительные сообщества.
СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ ФЛОРОНОСНЫХ ОТЛОЖЕНИЙ
Как было отмечено выше, вопрос о стратиграфическом положении флороносных отложений аянкинского комплекса возник из-за того, что местонахождения ископаемых растений попадают на два смежных листа ГГК-200: Q-58-XXXIV на юге и Q-58-XXVIII на севере, составленных разными геологическими организациями, использовавшими при этом разные стратиграфические схемы и номенклатуру картируемых стратонов (Белый, 1977; Филатова и др., 1977).
При характеристике аянкинского комплекса Лебедев (1987) указал, что он происходит с р. Обрывистая (левый приток р. Большая Аянка) примерно в 20 км от устья, а также по руч. Звонкий. На листе ГГК-200 Q-58-XXXIV, составленном объединением “Аэрогеология” в 1987 г., ручей Звонкий не обозначен, а на р. Обрывистая указано единственное местонахождение растительных остатков (Алексеев, 1987; http://www.geolkarta.ru/ list_200.php?idlist=Q-58-XXXIV). Оно приурочено к макковеемской свите близ северной границы листа (рис. 1а). Судя по расстоянию от устья реки и по содержанию стратиграфической колонки к листу, это и есть точка 700, о которой идет речь в работе Лебедева (1987). В работе А.Б. Германа (2011, с. 237) ошибочно указано, что аянкинская флора происходит из отложений вышележащей атвувеемской свиты.
Примерно в 3 километрах к северу от т. 700 проходит южная граница листа ГГК-200 Q-58-XXVIII, составленного СВПГО “Севвостгеология” в 1987 г. (Евстафьев, 1987; http://www.geolkarta.ru/ list_200.php?idlist=Q-58-XXVIII). По границе листов к выходам кислых образований макковеемской свиты примыкает поле распространения кислых пород аунейской толщи (рис. 1а), где находятся местонахождения 701 и 702. Никаких разломов на картах здесь не обозначено – это, безусловно, образования одного стратона, названного по-разному. То же наблюдается и в отношении перекрывающих флороносную толщу выходов атвувеемской свиты (на юге) и тувыйской толщи (на севере): на границе листов ГГК-200 поля их распространения примыкают друг к другу и их нижние границы совпадают (рис. 1а).
Однако, согласно стратиграфической колонке для листов ГГК-200 Q-58-XXVIII, XXIX, XXX, аунейская толща должна считаться первой (нижней) вулканогенной толщей в гомодромной последовательности вулканизма внешней зоны ОЧВП (Акинин, Миллер, 2011): ее перекрывает тувыйская толща вулканитов среднего состава, еще одна кислая балаганная толща и завершающие разрез оливин-пироксеновые базальты чуванской толщи (рис. 1б).
Примерно в восьмидесяти километрах к востоку от т. 700 в аунейской толще был установлен аунейский флористический комплекс (Головнева, Щепетов, 2013), имеющий значительное сходство с флорой Tchaunia-Kolymella из чаунской серии Пегтымельского прогиба (Самылина, 1974; Белый, 1977; Щепетов, 1991, 2020; Щепетов, Нешатаева, 2021): из 21 вида аунейского комплекса 14 видов – общие с чаунской флорой, в том числе и несколько видов-эндемиков этой флоры. Согласно сравнительно недавно полученным результатам 40Ar/39Ar датирования и новым палеоботаническим данным (Kelley et al., 1999; Головнева, Щепетов, 2013), возраст флоры Tchaunia-Kolymella оценивается как турон-коньякский (Герман, 2011) либо коньякский (Головнева, Щепетов, 2013), исходя из чего возраст аунейского флористического комплекса и аунейской толщи Л.Б. Головнева и С.В. Щепетов (2013) определили как коньякский. Позже, однако, Щепетов изменил свое мнение о возрасте чаунской флоры и полагает, что она “не имеет четкой латеральной и стратиграфической приуроченности” (Щепетов, Нешатаева, 2021, с. 55) и существовала с туронского по кампанский века. С этим, однако, трудно согласиться. Так или иначе, аянкинская флора (сантон–кампан) моложе, чем аунейский комплекс (турон–коньяк или коньяк), и, вероятнее всего, происходит из верхней части флороносной толщи, закартированной как макковеемская свита и аунейская толща (рис. 1а).
Лебедев (1987, с. 72) отмечает: “Аянкинская флора в бассейне р. Большая Аянка залегает под “верхними” базальтоидами пояса…”. Однако на карте (рис. 1) флороносные отложения перекрывает атвувеемская свита, сложенная андезибазальтами, базальтами и андезитами, которые едва ли можно считать “верхними базальтоидами пояса”. Таковыми, вероятно, является вышележащая безымянная толща субщелочных базальтов, датированная палеогеном (рис. 1б). Так или иначе, оба обнажения точки 700 действительно располагаются близ кровли кислой толщи под покровами лав основного-среднего состава.
Можно предположить, что атвувеемская свита и перекрывающие ее субщелочные базальты вместе являются стратиграфическим аналогом чуванской толщи, выделяемой на северных листах ГГК-200 геологами “Севвостгеология”. Однако там, где образования атвувеемской свиты переходят границу листов, они были идентифицированы как тувыйская толща (рис. 1а), причем на территории, расположенной в 50–60 км восточнее (на листе ГГК-200 Q-58-XXX: Куклев, 1980; http://www.geolkarta.ru/list_200.php?idlist=Q-58-XXX), тувыйская толща отделена от “верхних базальтоидов пояса” (в этом районе – чуванской толщи) еще одной (балаганной) кислой толщей. Балаганная толща либо ее аналоги, как и чуванская толща, отсутствуют на листах, где были собраны остатки растений аянкинской флоры (рис. 1а).
ЗАКЛЮЧЕНИЕ
Проведенное впервые детальное изучение аянкинской флоры, содержащей около 60 видов ископаемых растений, которые составляют три тафофлоры из трех местонахождений, позволяет датировать эту флору сантонским и кампанским веками позднего мела. Более детальное ее датирование – ранним кампаном (Лебедев, 1987, 1992) – представляется нам недостаточно обоснованным.
Флористические комплексы аянкинской флоры отражают склоновую растительность, в которой доминировали хвойные (тафофлора точки 701), и растительность пониженных участков рельефа – берегов реки или озера, где преобладали покрытосеменные (тафофлора точки 702). Таким образом, сонахождение этих тафофлор, по всей видимости, отражает процесс постепенного внедрения кайнофитных растительных сообществ на территорию вулканического нагорья, в позднем мелу еще занятую сообществами мезофита.
Относительно стратиграфического положения флороносных отложений аянкинской флоры можно заключить, что (1) аянкинская флора происходит из верхней части флороносной толщи, на разных листах ГГК-200 закартированной как макковеемская свита и аунейская толща; (2) аунейский флористический комплекс древнее аянкинской флоры и, вероятнее всего, происходит из стратиграфически более низкой части флороносных отложений; (3) можно предположить, что вулканиты балаганной толщи отражают локальный эпизод кислого вулканизма, предшествовавший излиянию “верхних” базальтов ОЧВП – чуванской толщи и ее аналогов – и не проявившийся в прилегающих с запада районах. Последнее предположение может быть подтверждено либо опровергнуто лишь при дополнительных полевых исследованиях.
Меловой флорогенез на Северо-Востоке Азии не был простым “линейным” процессом замещения одних флор другими, более продвинутыми. Приведенные данные о составе и возрасте аянкинской флоры показывают, что определение возраста верхнемеловых флороносных отложений ОЧВП по ископаемым растительным остаткам – дело непростое и нетривиальное.
Благодарности. Авторы искренне признательны Е.И. Костиной (Геологический институт РАН, Москва), в 1985 г. принимавшей участие в сборе ископаемых растений из местонахождения 700, и С.В. Щепетову (Ботанический институт РАН, Санкт-Петербург) за ценные советы и консультации по стратиграфии и геологическому строению рассматриваемого в статье района. Мы благодарны рецензентам нашей статьи – Е.В. Бугдаевой (ФНЦ Биоразнообразия ДВО РАН, Владивосток), Н.В. Носовой (Ботанический институт РАН, Санкт-Петербург), Ю.Б. Гладенкову и М.А. Рогову (оба – ГИН РАН, Москва), конструктивные замечания и советы которых помогли нам существенно улучшить данную статью.
Источники финансирования. Работа выполнена в рамках тем государственного задания Геологического института РАН (г. Москва) и Палеонтологического института РАН (г. Москва) и при поддержке Российского фонда фундаментальных исследований, грант № 19-05-00121.
Список литературы
Абрамова Л.Н. Новый вид дуба из верхнего мела Корякского нагорья // Палеонтол. журн. 1979. № 1. С. 144–147.
Акинин В.В., Миллер Э.Л. Эволюция известково-щелочных магм Охотско-Чукотского вулканогенного пояса // Петрология. 2011. Т. 19. № 3. С. 249–290.
Алексеев А.А. Государственная геологическая карта СССР масштаба 1 : 200000, лист Q-58-XXXIV. М.: Аэрогеология, 1987.
Белый В.Ф. Стратиграфия и структуры Охотско-Чукотского вулканогенного пояса. М.: Недра, 1977.
Герман А.Б. Альбская–палеоценовая флора Северной Пацифики. М.: ГЕОС, 2011. (Тр. ГИН РАН. Вып. 592).
Герман А.Б., Лебедев Е.Л. Стратиграфия и флора меловых отложений Северо-Западной Камчатки. М.: Наука, 1991. (Тр. ГИН АН СССР. Вып. 468).
Головнева Л.Б. Чаунская флора Охотско-Чукотского вулканогенного пояса. СПб.: Марафон, 2018.
Головнева Л.Б., Щепетов С.В. Карамкенский флористический комплекс из позднемеловых образований Охотско-Чукотского вулканогенного пояса // Палеоботаника. 2011. Т. 2. С. 100–113.
Головнева Л.Б., Щепетов С.В. Аунейский флористический комплекс из верхнемеловых отложений Охотско-Чукотского вулканогенного пояса // Палеоботаника. 2013. Т. 4. С. 96–115.
Головнева Л.Б., Алексеев П.И., Гниловская А.А., Юдова Д.А. Род Trochodendroides (Cercidiphyllaceae) в позднемеловых флорах Северо-Востока России // Палеоботаника. 2017. Т. 8. С. 122–179.
Евстафьев Ю.И. Государственная геологическая карта СССР масштаба 1 : 200 000, лист Q-58-XXVII, XXVIII. Магадан: СВПГО “Севвостокгеология”, 1987.
Ефимова А. Ф. Верхнемеловая флора бассейна р. Амгуэмы // Материалы по геологии и полезным ископаемым Северо-Востока СССР. 1966. Вып. 19. С. 172–188.
Красилов В.А. Меловая флора Сахалина. М.: Наука, 1979.
Куклев В.П. Государственная геологическая карта СССР масштаба 1:200000, лист Q-58-XXIX, XXX. Магадан: СВПГО “Севвостокгеология”, 1980.
Лебедев Е.Л. Стратиграфия и возраст Охотско-Чукотского вулканогенного пояса. М.: Наука, 1987 (Тр. ГИН АН СССР. Вып. 421).
Лебедев Е.Л. Меловые флоры Северо-Востока Азии // Изв. АН СССР. Сер. геол. 1992. № 4. С. 85–96.
Моисеева М.Г. Новые покрытосеменные из позднемеловой усть-эмунэрэтской флоры Центральной Чукотки // Палеонтол. журн. 2014. № 6. С. 102–111.
Моисеева М.Г. Условия произрастания усть-эмунэрэтской флоры и сантон-кампанский климат Чукотки // Стратиграфия. Геол. корреляция. 2015. Т. 23. № 4. С. 64–75.
Моисеева М.Г., Соколова А.Б. Кампанская флора района бухты Угольной (Северо-Восток России): систематический состав и палеоклимат // Чтения памяти А.Н. Криштофовича. Ред. Головнева Л.Б. Вып. 7. СПб.: Марафон, 2011. С. 62–87.
Моисеева М.Г., Соколова А.Б. Новые данные о составе и возрасте усть-эмунэрэтской флоры бассейна р. Энмываам (Центральная Чукотка) // Стратиграфия. Г-еол. корреляция. 2014. Т. 22. № 3. С. 45–63.
Моисеева М.Г., Соколова А.Б., Герман А.Б. Позднемеловая аянкинская флора Охотско-Чукотского вулканогенного пояса: новые данные // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Материалы Седьмого Всероссийского совещания, 10–15 сентября 2014 г., г. Владивосток. Ред. Барабошкин Е.Ю. и др. Владивосток: Дальнаука, 2014. С. 223–226.
Пергамент М.А. Биостратиграфия и иноцерамы сенона (сантон–маастрихт) тихоокеанских районов СССР. М.: Наука, 1974. (Тр. ГИН АН СССР. Вып. 260).
Самылина В.А. Раннемеловые флоры Северо-Востока СССР (К проблеме становления флор кайнофита) // ХХVII Комаровские чтения. Л.: Наука, 1974.
Самылина В.А. Аркагалинская стратофлора Северо-Востока Азии. Л.: Наука, 1988.
Соколова А.Б., Моисеева М.Г. Новый вид рода Sequoiadendron Buchholz (Cupressaceae) из верхнемеловых отложений бассейна р. Энмываам (Центральная Чукотка) // Палеонтол. журн. 2016. № 1. С. 91–101.
Филатова Н.И., Лебедев Е.Л. Сенонские отложения Охотско-Чукотского вулканогенного пояса // Изв. АН СССР. Сер. геол. 1982. № 10. С. 111–114.
Филатова Н.И., Дворянкин А.И., Лебедев Е.Л., Цукерник А.Б. Стратиграфия меловых отложений Охотско-Чукотского вулканического пояса (бассейны рек Пенжины, Еропола, Анадыря) // Бюлл. МОИП. Отд. геол. 1977. Т. 52. № 2. С. 67–82.
Филиппова Г.Г., Абрамова Л.Н. Позднемеловая флора Северо-Востока России. М.: Недра, 1993.
Щепетов С.В. Среднемеловая флора чаунской серии (Центральная Чукотка). Магадан: СВКНИИ ДВО АН СССР, 1991.
Щепетов С.В. О стратиграфическом положении меловых отложений с гребенкинской флорой, Северо-Восток России // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 1. С. 1–8.
Щепетов С.В., Герман А.Б., Нешатаева В.Ю. Формирование палеофлор и меловой вулканизм на Северо-Востоке Азии. СПб.: Марафон, 2019.
Щепетов С.В., Нешатаева В.Ю. К вопросу о чаунской палеофлоре из неморского мела Чукотки // Тихоокеанская геология. 2021. Т. 40. № 2. С. 55–66.
Akinin V.V., Golovneva L.B., Salnikova E.B., Anisimova I.V., Shczepetov S.V., Nosova N.V. The composition and age of the Ul’ya flora (Okhotsk-Chukotka volcanic belt, North-East of Russia): paleobotanical and geochronological constraints // Acta Palaeobot. 2019. V. 59. № 2. P. 251–276.
Arnold C.A., Lowther J.S. A new Cretaceous conifer from Northern Alaska // Am. J. Botany. 1955. V. 42. № 6. P. 522–528.
Bell W.A. Uppermost Cretaceous and Paleocene floras of Western Alberta // Geol. Surv. Bull. 1949. V. 285. P. 1–231.
Bell W.A. Lower Cretaceous floras of western Canada // Geol. Surv. Can. Mem. 1956. V. 285. P. 1–331.
Berry E.W. Systematic palaeontology of the Lower Cretaceous deposits of Maryland: Pteridophyta; Cycadophytae; Gymnospermae; Monocotyledonae; Dicotyledonae. Baltimore: Johns Hopkins Press, 1911. P. 1–729.
Berry E.W. The flora of Woodbine Sand at Arthur Bluff, Texas // U.S. Geol. Surv. Prof. Pap. 1922. № 129-G. P. 153–180.
Crane P.R. A re-evaluation of Cercidiphyllum-like plant fossils from the British Early Tertiary // Bot. J. Linn. Soc. 1984. V. 89. P. 199–230.
Detterman R.L., Bickel R.S. Geology of the Chandler River Region, Alaska // U.S. Geol. Surv. Prof. Pap. 1963. V. 303E. P. 223–324.
Florin R. On Jurassic taxads and conifers from north-western Europe and eastern Greenland // Acta Horti Bergiani. 1958. V. 17. P. 257–402.
Frederiksen N.O., McIntyre D.J. Palynomorph Biostratigraphy of Mid(?)-Campanian to Upper Maastrichtian Strata along the Colville River, North Slope of Alaska // U.S. Geol. Surv. Open File Report. 2000. V. 00–493. P. 1–36.
Herman A.B. Late Early–Late Cretaceous floras of the North Pacific Region: florogenesis and early angiosperm invasion // Rev. Palaeobot. Palynol. 2002. V. 122. № 1–2. P. 1–11.
Herman A.B. Albian–Paleocene flora of the North Pacific: systematic composition, palaeofloristics and phytostratigraphy // Stratigr. Geol. Correl. 2013. V. 21. № 7. P. 689–747.
Herman A.B., Golovneva L.B., Shczepetov S.V., Grabovsky A.A. The Late Cretaceous Arman Flora of Magadan Oblast, North-Eastern Russia // Stratigr. Geol. Correl. 2016. V. 24. № 7. P. 651–760.
Kelley S., Spicer R.A., Herman A.B. New 40Ar/39Ar dates for Cretaceous Chauna Group tefra, Northeastern Russia, and their implications for the geologic history and floral evolution of the North Pacific region // Cretaceous Res. 1999. V. 20. № 1. P. 97–106.
Nosova N., Herman A., Grabovskiy A., Kostina E. Cretaceous Phoenicopsis Heer (Leptostrobales): a new species and a geological history of the genus // Cretaceous Res. 2020. V. 116. P. 104578.
Rothwell G.W., Ohana T. Stockeystrobus gen. nov. (Cupressaceae), and the evolutionary diversification of sequoioid conifer seed cones // Botany. 2016. V. 94. № 9. P. 847–861.
Rothwell G.W., Stockey R.A., Smith S.Y. Revisiting the Late Cretaceous Parataxodium wigginsii flora from the North Slope of Alaska, a high-latitude temperate forest // Cretaceous Res. 2020. V. 116. P. 104592.
Sokolova A., Gordenko N., Zavialova N. The most ancient member of the Sequoioideae – the new genus Krassilovidendron Sokolova, Gordenko et Zavialova (Cupressaceae s.l.) from the Albian–Cenomanian of Western Siberia (Russia) // Cretaceous Res. 2017. V. 77. P. 1–27.
Tikhomirov P.L., Kalinina E.A., Moriguti T., Makishima A., Kobayashi K., Cherepanova I.Yu., Nakamura E. The Cretaceous Okhotsk–Chukotka Volcanic Belt (NE Russia): geology, geochronology, magma output rates, and implications on the genesis of silicic LIPs // J. Volcanol. Geothermal Res. 2012. V. 221–222. P. 14–32.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция