

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 2, стр. 3-38
Новые данные по биостратиграфии среднего девона Салаира, Юго-Западная Сибирь
Н. Г. Изох 1, *, Б. М. Попов 1, Е. С. Соболев 1, Т. А. Щербаненко 1
1 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
* E-mail: izokhng@ipgg.sbras.ru
Поступила в редакцию 17.06.2021
После доработки 25.10.2021
Принята к публикации 07.11.2021
- EDN: AWBTKJ
- DOI: 10.31857/S0869592X2202003X
Аннотация
Проведены ревизия и анализ биостратиграфических данных по брахиоподам, аммоноидеям, конодонтам и остракодам мамонтовского и заречного горизонтов Салаира. Уточнена стратиграфическая последовательность среднедевонских отложений Салаира и предложено выделение нового заречного горизонта, включающего акарачкинские, сафоновские и керлегешские слои. Установленная в верхней части акарачкинских слоев позднеэйфельская ассоциация аммоноидей генозоны Agoniatites является биостратиграфическим репером для глобальных корреляций. Сафоновские слои охарактеризованы позднеэйфельской–раннеживетской ассоциацией брахиопод с Indospirifer padaukpinensis (= I. pseudowilliamsi). В керлегешских слоях выявлены две среднеживетские брахиоподовые зоны: Rhynchospirifer hians (= Ilmenia subhians) и Subrensselandia? salairica (= Chascothyris salairica). Комплекс конодонтов акарачкинских слоев характеризует верхний эйфель. Верхняя часть сафоновских слоев содержит конодонты Icriodus difficilis Ziegler et Klapper, Polygnathus linguiformis klapperi Clausen, Leuteritz et Ziegler, Po. parawebbi Chatterton beta morphotype, Po. ovatinodosus Ziegler et Klapper, Po. pseudofoliatus Wittekind, характерные для среднего живета. В керлегешских слоях конодонты не обнаружены.
ВВЕДЕНИЕ
Нижне- и среднедевонские отложения Салаира играют ключевую роль в строении Алтае-Саянской складчатой области. Они представлены разнофациальными терригенно-карбонатными породами с богатой фауной (Решения…, 1982). Эти отложения наиболее полно вскрыты в карьерах и придорожных выемках вдоль восточного склона Салаира (рис. 1). В последние годы в ходе проведения биостратиграфических исследований получены важные результаты, которые позволили уточнить последовательность этих отложений. Для верхнеэйфельского и живетского интервала Салаира авторами настоящей статьи приводится комплексное обоснование выделения нового заречного горизонта, основанное на детальном изучении типовых выходов акарачкинских, сафоновских и керлегешских слоев (рис. 1).
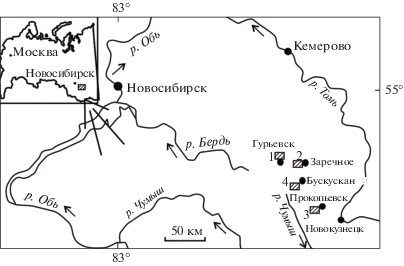
Рис. 1.
Местонахождение изученных разрезов среднего девона. 1 – окрестности г. Гурьевска, Малосалаиркинский карьер (разрезы Я-882, Б-8318) и правый берег р. Малый Бачат (разрез Б-8410); 2 – западная окраина с. Заречное, правый берег р. Большой Бачат (разрез Б-8333); 3 – придорожные карьеры на западной окраине г. Прокопьевска (разрезы БС-091, БС-092 и БС-093); 4 – Бускусканский карьер, юго-западнее ж/д ст. Бускускан (разрез Б-8170).
Полученные новые результаты были представлены на международных и российских конференциях и заседаниях девонской комиссии СибРМСК и МСК России (Бахарев и др., 2012; Bakharev et al., 2013; Изох, Язиков, 2014; Язиков, Изох, 2014а, 2014б; Язиков и др., 2014а, 2014б, 2015а, 2016 и др.).
ИСТОРИЯ ИЗУЧЕНИЯ
Изучение девонских отложений окраин Кузбасса имеет более чем полуторавековую историю и неразрывно связано с интенсивной эволюцией представлений об их биостратиграфическом расчленении (Петц, 1901; Лазуткин и др., 1936; Яворский, 1938; Карцева, Цырлина, 1956; Ржонсницкая, 1959; Фомичев, Алексеева, 1961; Елкин, 1964, 1968 и др.). Фактический материал, полученный при проведении детальных геолого-съемочных работ, был обобщен в монографии “Стратиграфия СССР. Девонская система” (Ржонсницкая, 1973). Этот материал был положен в основу схемы стратиграфии девона Сибири, которая была представлена на Межведомственном совещании в 1979 г. (Решения…, 1982) и до сих пор является официально действующей (табл. 1).
Таблица 1.
История развития региональной стратиграфической схемы среднедевонских отложений Салаира (западная часть Алтае-Саянской области)
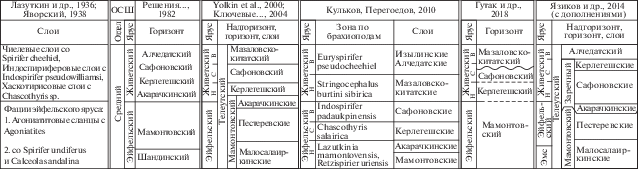
С начала 1980-х годов по настоящее время геологические исследования Салаира и окраин Кузнецкого бассейна связаны с детализацией изучения девонских отложений и различных групп фауны. Особое внимание было уделено изучению конодонтов, зональные виды-индексы которых являются маркерами нижних границ ярусных подразделений девона (Елкин и др., 1983; Елкин, Изох, 1986; Ржонсницкая и др., 1985; Гагиев и др., 1986, 1987 и др.). В начале 2000-х годов А.Ю. Язиковым была уточнена брахиоподовая зональная схема расчленения среднего девона складчатого обрамления Кузнецкого бассейна (Язиков, 2014в).
В изучении девонских отложений принимали участие исследователи из разных организаций: Е.А. Елкин, Р.Т. Грацианова, Н.К. Бахарев, В.М. Задорожный, А.Ю. Язиков, Н.Г. Изох, Б.М. Попов, Е.С. Соболев и Т.А. Щербаненко из ИГиГ СО АН СССР (ныне ИНГГ СО РАН), Новосибирск; В.А. Желтоногова, Я.М. Гутак и Л.В. Галенко из ЗСГУ, Новокузнецк; О.П. Мезенцева из НПИ, Новокузнецк; М.А. Ржонсницкая, Н.М. Петросян, В.Ф. Куликова, Т.Л. Модзалевская, Я.Н. Спасский, В.Л. Клишевич и О.Б. Тимофеева из ВСЕГЕИ, Ленинград; Г.Д. Исаев, В.И. Краснов, Н.П. Кульков, С.А. Степанов из СНИИГГиМС, Новосибирск; М.Х. Гагиев из СВКНИИ, Магадан; С.А. Родыгин из ТГУ, Томск; Н.М. Мирецкая из ТО СНИИГГиМС, Томск и др.
В результате анализа ранее опубликованных и полученных новых данных по био- и литостратиграфии девонских отложений Салаира и западных окраин Кузнецкого бассейна, авторы настоящей статьи пришли к выводу о необходимости пересмотра схемы стратиграфии среднего девона западной части Алтае-Саянской складчатой области и внесения в нее соответствующих коррективов. Такое решение было основано на выявлении несоответствия сложившихся представлений о стратиграфической последовательности региональных подразделений среднего девона Салаира (Решения…, 1982) и той последовательности, которая была установлена в последнее время в типовых разрезах акарачкинских, сафоновских и керлегешских слоев.
Развитие представлений о стратиграфическом расчленении среднего девона Салаира показано в табл. 1. На первом этапе были выделены слои с фауной, которые позднее переведены в ранг горизонтов (Лазуткин и др., 1936; Яворский, 1938; Ржонсницкая, 1952; Стратиграфический…, 1975; Решения…, 1982 и др.). В последние годы были приведены разные оценки возраста акарачкинского, керлегешского и сафоновского горизонтов (Кульков, Перегоедов, 2010; Гутак и др., 2018 и др.), которые отличаются от возраста этих подразделений в официально действующей схеме (табл. 1). Неоднозначность в понимании возраста этих отложений приводит к затруднению определения истинной стратиграфической последовательности среднего девона Салаира. После комплексного анализа полученных новых и опубликованных данных мы пришли к выводу о необходимости провести корректировку этой последовательности и считать керлегешские (хаскотирисовые) слои более молодыми по сравнению с сафоновскими (индоспириферовыми). Исходя из понимания горизонта как определенного трансгрессивно-регрессивного цикла в развитии палеобассейна, мы предлагаем выделить новый заречный горизонт для среднего девона Салаира, который включает акарачкинские, сафоновские и керлегешские слои (Язиков и др., 2014а) (рис. 2).
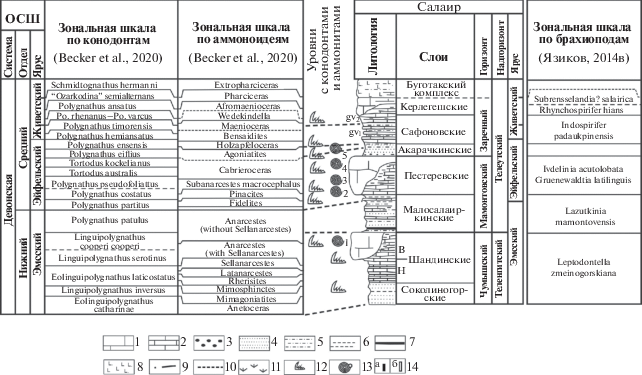
Рис. 2.
Региональная стратиграфическая схема нижнего и среднего девона Салаира (по Язиков и др., 2016, с уточнениями). Условные обозначения: 1 – известняки массивные; 2 – известняки слоистые; 3 – гравелиты; 4 – песчаники; 5 – алевролиты; 6 – аргиллиты; 7 – уголь; 8 – андезибазальты, трахибазальты, базальты; 9 – разломы; 10 – корреляционные линии; 11 – задернованный интервал; 12 – конодонты; 13 – аммоноидеи; 14а – определение брахиопод до вида; 14б – виды брахиопод, определенные с cf. Уровни с аммоноидеями: 1 – Mimagoniatites bohemicus (Barrande), шандинские слои; 2 – Fidelites fidelis (Barrande), Fidelites occultus (Barrande), Fidelites sp.; пестеревские слои (нижняя часть); 3 – Fidelites fidelis (Barrande), Fidelites bicanaliculatus (Sandberger et Sandberger), Fidelites sp., Subanarcestes aff. marhoumensis Göddertz; пестеревские слои (нижняя часть); 4 – Fidelites bicanaliculatus (Sandberger et Sandberger), Subanarcestes ex gr. marhoumensis Göddertz; пестеревские слои (верхняя часть); 5 – Agoniatites vanuxemi (Hall), Fidelites sp., Cabrieroceras salairicum Nikolaeva; акарачкинские слои (верхняя часть) (Язиков и др., 2015а, 2016).
Обоснованием для пересмотра схемы стратиграфии среднего девона послужили следующие данные:
1. Выявлен комплекс аммоноидей генозоны Agoniatites верхнего эйфеля, который может рассматриваться в качестве регионального и глобального биомаркера. В четырех изученных разрезах (Б-8410, БС-091, БС-093 и Б-8333) эти аммоноидеи прослежены в туфогенной пачке в кровле акарачкинских слоев.
2. Уточнена последовательность брахиопод в изученных разрезах среднего девона Салаира. Акарачкинские слои с аммоноидеями генозоны Agoniatites верхнего эйфеля согласно перекрываются сафоновскими слоями с комплексом брахиопод зоны Indospirifer padaukpinensis (= I. pseudowilliamsi) (разрезы БС-091 и БС-093), на которых залегают отложения с брахиоподами зон Rhynchospirifer hians (= Ilmenia subhians) и Subrensselandia? salairica (= Chascothyris salairica), характеризующими более высокие керлегешские слои (разрезы БС-092 и Б-8170).
3. Наконец, установленный в верхней части сафоновских слоев комплекс конодонтов характеризует зону Polygnathus rhenanus/Polygnathus varcus (= Icriodus difficilis) и низы зоны Polygnathus ansatus среднего живета.
МАТЕРИАЛ
Изученные разрезы расположены в типовой местности распространения акарачкинских, сафоновских и керлегешских слоев в окрестностях г. Гурьевска, г. Прокопьевска, с. Заречного и ж/д ст. Бускускан Кемеровской области (рис. 1). Основные сборы макрофауны и отбор образцов на микрофауну были сделаны при детальном описании опорных разрезов среднего девона в период с 1980-х годов до начала 2000-х годов и затем дополнены в последующие годы. Коллекция аммоноидей, насчитывающая около 300 экземпляров, была собрана из разрезов Б-8410, Е-837, БС-091, БС-093 и Б-8333 Е.А. Елкиным, Н.К. Бахаревым, О.П. Мезенцевой и Е.С. Соболевым в 1983, 1984 и 2005–2014 гг. На микрофауну (конодонты и остракоды) был отобран 201 образец весом от 2 до 8 кг: 10 образцов из разреза Б-8410, 61 образец из разреза БС-091, 70 образцов из разреза БС-093, 20 образцов из разреза Б-8333, 20 образцов из разреза БС-092 и 20 образцов из разреза Б-8170.
Коллекция брахиопод заречного горизонта насчитывает несколько тысяч экземпляров и характеризует комплекс из 73 таксонов, ревизованных А.Ю. Язиковым согласно современной систематике. Важные номенклатурные изменения коснулись в том числе видов, давших названия “индоспириферовые слои” и “хаскотирисовые слои”. Так, зональный вид сафоновских слоев Indospirifer pseudowilliamsi включен в синонимику вида Indospirifer padaukpinensis, а виды-индексы керлегешских слоев Ilmenia subhians и “Chascothyris” salairica отнесены к видам Rhynchospirifer hians и Subrebsselandia salairica соответственно (Язиков, 2014в).
Коллекции микрофауны получены с использованием традиционных методик химического препарирования карбонатных пород слабым раствором уксусной кислоты (5–7%) и плавлением с использованием гипосульфита натрия в лабораториях ИНГГ СО РАН. Конодонтовые элементы хорошей и удовлетворительной сохранности найдены только в 23 образцах из 201, при этом следует отметить, что в двух разрезах (Б-8170 и БС-092) они не были обнаружены. Раковины и разрозненные створки остракод хорошей и удовлетворительной сохранности встречены в 55 образцах из пяти разрезов, за исключением разреза Б-8410. Конодонты изучались Н.Г. Изох, остракоды – Н.К. Бахаревым и Б.М. Поповым, аммоноидеи – Н.К. Бахаревым, С.В. Николаевой, Е.С. Соболевым, Т. Беккером и К. Клюгом, брахиоподы – А.Ю. Язиковым и Т.А. Щербаненко. Фотографирование конодонтов и остракод было проведено в Аналитическом центре коллективного пользования ИГМ СО РАН на сканирующем электронном микроскопе TESCAN MIRA3. Изучение коллекций проводилось под бинокулярным микроскопом Stemi 2000 ZEISS. Коллекции фауны хранятся в Лаборатории палеонтологии и стратиграфии палеозоя, Лаборатории микропалеонтологии и Лаборатории палеонтологии и стратиграфии мезозоя, а также в Центре коллективного пользования (ЦКП) “Коллекция ГЕОХРОН” Института нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН в г. Новосибирске.
БИОСТРАТИГРАФИЧЕСКИЕ ИССЛЕДОВАНИЯ
Телеутский надгоризонт
Телеутский надгоризонт выделен Е.А. Елкиным для среднего девона западной части Алтае-Саянской складчатой области и включает мамонтовский, керлегешский, сафоновский и алчедатский горизонты (Елкин и др., 1983). В настоящее время мы предлагаем рассматривать в составе телеутского надгоризонта мамонтовский, заречный и алчедатский горизонты и сопоставляем его с верхней частью эмсского яруса, эйфельским и живетским ярусами Общей стратиграфической шкалы. Мамонтовский горизонт включает малосалаиркинские и пестеревские слои, а заречный горизонт – акарачкинские, сафоновские и керлегешские слои. Основные биостратиграфические результаты получены при изучении типовых выходов этих региональных подразделений с послойным отбором палеонтологических образцов и точной их привязкой в разрезах.
Мамонтовский горизонт
Название дано М.А. Ржонсницкой (1952) по дер. Мамонтово на юго-западной окраине Кузбасса. Стратотип установлен на правом берегу р. Малый Бачат, северо-восточнее Акарачкинского карьера в окрестностях г. Гурьевска. Мамонтовский горизонт отвечает верхней части эмсского яруса и большей части эйфельского яруса или, в конодонтовой биозональности, зонам Polygnathus patulus, Polygnathus partitus, Polygnathus costatus, Polygnathus pseudofoliatus, Tortodus australis и нижней части зоны Tortodus kockelianus (Ключевые…, 2004; Язиков и др., 2014а, 2014б, 2015а, 2016). Горизонт включает малосалаиркинские и пестеревские слои (рис. 2).
Малосалаиркинские слои. Впервые название было использовано М.А. Ржонсницкой (1968) для преимущественно терригенных отложений, перекрывающих верхнешандинские слои с Zdimir baschkiricus. Название дано по дер. Малая Салаирка вблизи г. Гурьевска. В качестве стратотипа был предложен разрез “на первом холме к востоку от дер. Малая Салаирка” (Ржонсницкая, 1968 с. 87), в настоящее время, по-видимому, попавший в контуры Малосалаиркинского карьера. Нижние стратиграфические уровни малосалаиркинских слоев и контакт с верхнешандинскими представлены в северо-восточном борту Акарачкинского карьера, а выходы их верхних уровней слагают южную и юго-западную части Малосалаиркинского карьера.
Малосалаиркинские слои по брахиоподам соответствуют биозоне Lazutkinia mamontovensis. Характерными видами слоев являются: Lazutkinia mamontovensis Ržonsnickaja, Retzia salairica Peetz, Beckmannia ex gr. pentagona (Kayser), Tetratomia? yavorskii Ržonsnickaja, Urella asiatica Ržonsnickaja и Eoreticularia aviceps (Kayser) (Язиков, 2014а, 2014в).
Малосалаиркинские слои конодонтами слабо охарактеризованы, встречены только Caudicriodus sp. A. Первые зональные эйфельские конодонты Polygnathus partitus Klapper, Ziegler et Mashkova и Po. costatus Klapper совместно установлены в нижней части пестеревских известняков. Таким образом, стратиграфический интервал малосалаиркинских слоев соответствует конодонтовым зонам Polygnathus patulus и Polygnathus partitus, т.е. пограничным отложениям эмса и эйфеля (рис. 2). Учитывая масштабную смену брахиоподовых ассоциаций в верхней части малосалаиркинских слоев, авторы считают целесообразным именно на этом уровне условное проведение границы эмсского и эйфельского ярусов (Изох, Язиков, 2014; Язиков, Изох, 2014а, 2014б).
Пестеревские слои. Название дано Г.Г. Петцем (1901) по дер. Новопестерево Гурьевского района. Стратотип был определен М.А. Ржонсницкой и рассматривался как выходы светло-серых биоморфных известняков, “слагающие посередине дер. Ново-Пестерево холм, на котором расположена школа” (Харин, 1958; Ржонсницкая, 1968 с. 88). В настоящее время пестеревские слои в полном объеме обнажены в центральной, северной и северо-восточной частях Малосалаиркинского карьера, где предлагается установить неостратотип.
В нижней части пестеревских известняков встречены первые зональные эйфельские конодонты Polygnathus partitus Klapper, Ziegler et Mashkova и Po. costatus Klapper, совместно с Icriodus regularicrescens Bultynck, Caudicriodus sp. A, а также аммоноидеи, характерные для нижнеэйфельской генозоны Pinacites (Ключевые…, 2004; Middle-Upper…, 2011).
В верхней части пестеревских слоев обнаружены конодонты нижней части зоны Tortodus kockelianus: Polygnathus costatus Klapper, Po. linguiformis pinguis Weddige, Po. linguiformis klapperi Clausen, Leuteritz et Ziegler, Po. angusticostatus Wittekindt, Po. robusticostatus Bischoff et Ziegler, Icriodus regularicrescens Bultynck, Tortodus cf. planus (Bischoff et Ziegler) (Ключевые…, 2004; Middle-Upper…, 2011). Перечисленные таксоны являются космополитными и широко распространены в разных регионах мира (Weddige, 1977; Klapper, Johnson, 1980 и др.). Polygnathus costatus Klapper, Po. linguiformis pinguis Weddige, Po. angusticostatus Wittekindt и Po. robusticostatus являются характерными видами эйфельского яруса. Подвид Po. linguiformis klapperi появляется с зоны Tortodus kockelianus верхнего эйфеля и проходит в живетский ярус до конодонтовой зоны “Ozarkodina” semialternans (Clausen et al., 1979; Walliser, Bultynck, 2011).
К числу характерных видов брахиопод рассматриваемых слоев относятся: Clorinda asinuata Sapelnikov, Gruenewaldtia latilinguis (Schnur), Corvinopugnax resupinata (Kulkov), Peratos pellax L. Mizens, Desquamatia (I.) zonata (Schnur), Isospinatrypa aspera subspinosa (Lazutkin) и Ivdelinia acutolobata (Sandberger) (Язиков, 2014а, 2014б, 2014в).
Пестеревские слои в типовых выходах в Малосалаиркинском карьере охарактеризованы конодонтами зоны Po. costatus и нижней части зоны Tortodus kockelianus, а по брахиоподам соответствуют биозонам Gruenewaldtia latilinguis и Ivdelinia acutolobata (рис. 2).
Мамонтовский горизонт отвечает единому трансгрессивному циклу и перекрывается акарачкинскими слоями заречного горизонта. Граница мамонтовского и заречного горизонтов представлена в северо-восточном борту Малосалаиркинского карьера. Несмотря на то что происходит резкая смена состава отложений (от известняков к тонкообломочным осадкам), граница между ними согласная.
Заречный горизонт
Название дано А.Ю. Язиковым и Н.Г. Изох (Язиков и др., 2014) по с. Заречное Беловского района Кемеровской области, где выходы этого горизонта широко распространены. Стратотип заречного горизонта является составным и представлен совокупностью стратотипов входящих в него слоев – акарачкинских, сафоновских и керлегешских, которые ранее рассматривались в ранге горизонтов (рис. 2) (Решения…, 1982). Эта последовательность представлена в серии разрезов на окраине г. Гурьевска (Я-986, Б-8410), г. Прокопьевска (БС-091, БС-093, БС-092), с. Заречное (Б-8333) и в Бускусканском карьере (разрез Б-8170) (рис. 1). При повторном изучении разрезов БС-091, БС-093, БС-092, Б-8333 и Б-8170 выяснено, что первоначально они были описаны в обратной последовательности (Язиков, 2014в).
Заречный горизонт отвечает интервалу верхней части эйфеля, нижней и средней частей живетского яруса, в конодонтовой зональной последовательности – зонам Tortodus kockelianus, Polygnathus eiflius, Polygnathus ensensis, Polygnathus hemiansatus, Polygnathus rhenanus–Polygnathus varcus, Polygnathus ansatus и Ozarkodina semialternans (рис. 2).
Акарачкинские слои. Название дано М.А. Ржонсницкой по пос. Акарачкино вблизи г. Гурьевска (Ржонсницкая, 1952). Стратотип акарачкинских слоев (разрез Б-8410) расположен на правобережье р. Малый Бачат, в 0.5 км восточнее окраины пос. Раздольный (рис. 1, 3, 4), однако контакты с нижележащими пестеревскими и с перекрывающими сафоновскими слоями здесь не вскрыты. По этой причине для акарачкинских слоев предложен составной неостратотип, в котором нижние уровни акарачкинских слоев представлены в северо-восточной стенке Малосалаиркинского карьера (Я-896), а более высокие уровни – на западной окраине г. Прокопьевска (разрезы БС-091 и БС-093) и на западной окраине с. Заречное (разрез Б-8333) (рис. 1, 5–10). Мощность акарачкинских слоев составляет около 130 м. Акарачкинские слои, ранее включаемые в состав мамонтовского горизонта (Ключевые…, 2004, Middle-Upper…, 2011), авторами настоящей статьи рассматриваются в ранге нижних слоев заречного горизонта (Язиков, 2014в; Язиков, Изох, 2014б; Язиков и др., 2014).
Рис. 3.
Литологическая колонка и распространение фауны в акарачкинских слоях (брахиоподы, конодонты, аммоноидеи) в разрезе Б-8410 по правому берегу р. Малый Бачат. Условные обозначения см. рис. 2.
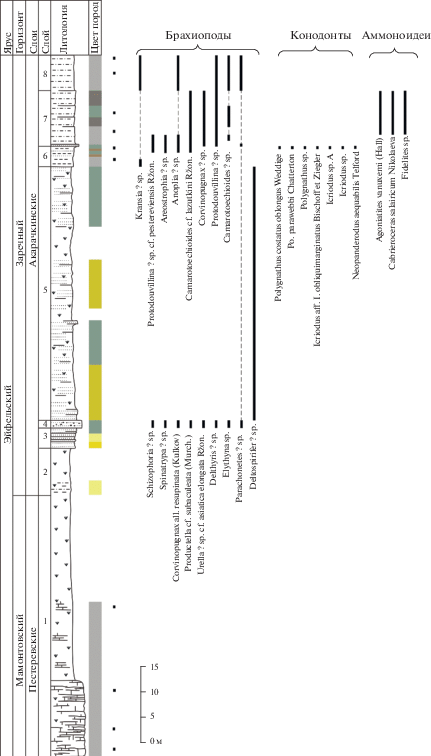
Рис. 4.
Фотопанорама верхней части разреза Б-8410, слой 7 с находками аммоноидей, окрестности г. Гурьевска, правый берег р. Малый Бачат.

Рис. 5.
Литологическая колонка и распространение фауны в акарачкинских, сафоновских и керлегешских слоях (брахиоподы, конодонты, остракоды, аммоноидеи) в разрезе БС-091 в придорожной выемке по левому берегу безымянного ручья – левого притока руч. Егос, окрестности г. Прокопьевска. Условные обозначения см. рис. 2.

Рис. 5.
Продолжение

Рис. 5.
Окончание

Рис. 6.
Фотопанорама разреза БС-091 в придорожной выемке по левому берегу безымянного ручья – левого притока руч. Егос, окрестности г. Прокопьевска.

Рис. 7.
Литологическая колонка и распространение фауны в акарачкинских, сафоновских и керлегешских слоях (брахиоподы, конодонты, аммоноидеи, остракоды) в разрезе БС-093 в придорожной выемке, окрестности г. Прокопьевска. Условные обозначения см. рис. 2.
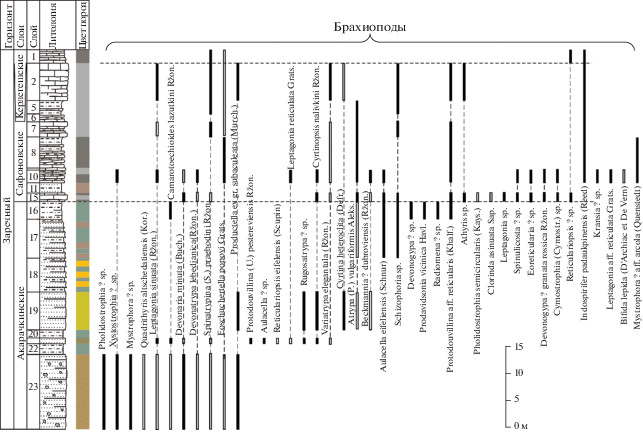
Рис. 7.
Окончание
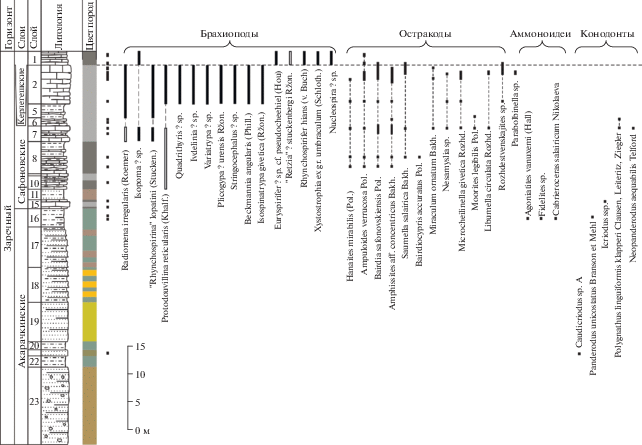

Рис. 9.
Литологическая колонка и распространение фауны в акарачкинских, сафоновских и керлегешских слоях (брахиоподы, аммоноидеи, остракоды) в разрезе Б-8333 (западная окраина с. Заречное). Условные обозначения см. рис. 2.

Акарачкинские слои в своей нижней части фаунистически слабо охарактеризованы, однако в верхней части встречены многочисленные брахиоподы, аммоноидеи и конодонты (рис. 3, 5, 7). Находки тентакулитов Nowakia procera (Maurer) известны в нижней части акарачкинских слоев Малосалаиркинского карьера; в верхней части слоев на правом берегу р. Малый Бачат обнаружены Viriatellina afoniana G. Ljaschenko (Клишевич, Мирецкая, 1986).
Брахиоподы. Ассоциация брахиопод включает 54 вида в составе 40 родов (23 таксона определены до рода). Характерные только для этих слоев виды отсутствуют; все обнаруженные брахиоподы встречены либо в нижележащих, либо в более молодых отложениях (рис. 3, 5, 7, 9). Однако в акарачкинских слоях в комплексе брахиопод намечается переход к новой ассоциации заречного горизонта и угасание большинства мамонтовских форм (Язиков, 2014а, 2014б, 2014в; Язиков и др., 2014).
Аммоноидеи. Региональным и глобальным биомаркером акарачкинских слоев является комплекс аммоноидей, выявленный в их кровле в четырех разрезах (рис. 3, 5, 7, 9): Б-8410 (окрестности г. Гурьевска), БС-091, БС-093 (окрестности г. Прокопьевска) и Б-8333 (окрестности с. Заречное, правобережье р. Большой Бачат) (Николаева, Бахарев, 2010; Bakharev, Sobolev, 2011; Язиков и др., 2014, 2015а). В известковистых алевролитах верхней части типового разреза акарачкинских слоев (разрез Б-8410, слой 7; рис. 3, 4) обнаружены: Agoniatites vanuxemi (Hall) (табл. I , фиг. 1–7), Fidelites sp. (табл. I , фиг. 9, 10), Cabrieroceras salairicum Nikolaeva (табл. I , фиг. 11, 12). Идентичные аммоноидеи обнаружены в двух разрезах (БС-091, БС-093) окрестностей г. Прокопьевска в туфогенной терригенно-карбонатной пачке (рис. 5, 7; табл. I , фиг. 8, 13; табл. II), которая ранее традиционно рассматривалась в составе сафоновской свиты (Ржонсницкая, 1968).
Рис. 11.
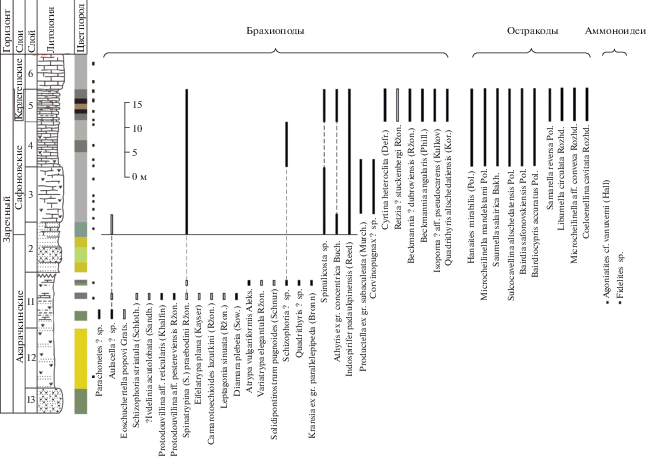
Литологическая колонка и распространение фауны в керлегешских слоях (брахиоподы, остракоды) разреза БС-092 в придорожной выемке по правому берегу безымянного ручья – левого притока руч. Егос, окрестности г. Прокопьевска. Условные обозначения см. рис. 2.

Рис. 12.
Фотопанорама разреза БС-092 в придорожной выемке по правому берегу безымянного ручья – левого притока руч. Егос, окрестности г. Прокопьевска.

Рис. 13.
Литологическая колонка и распространение фауны в сафоновских и керлегешских слоях (брахиоподы, остракоды) в разрезе Б-8170 в Бускусканском карьере (по Бахарев, 1985, с дополнениями). Условные обозначения см. рис. 2.
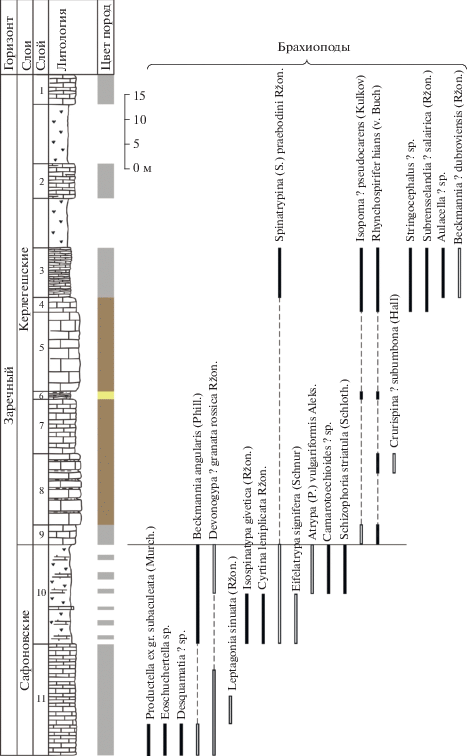
Рис. 13.
Окончание

Таблица I . Аммоноидеи среднего девона Салаира (верхний эйфель, генозона Agoniatites, заречный горизонт, акарачкинские слои; окрестности городов Гурьевска и Прокопьевска). Все изображения даны в натуральную величину. Во всех случаях: а – вид с латеральной стороны, б – вид с вентральной стороны, в – вид с устья. 1–8 – Agoniatites vanuxemi (Hall, 1879): 1 – экз. № Б-8410-7/113, вид на вентральную и латеральную стороны; окрестности города Гурьевска, разрез Б-8410, слой 7; 2 – экз. № Б-8410-7/100; окрестности г. Гурьевска, разрез Б-8410, слой 7; 3 – экз. № Б-8410-7/109; окрестности г. Гурьевска, разрез Б-8410, слой 7; 4 – экз. № Б-8410-7/116; окрестности г. Гурьевска, разрез Б-8410, слой 7; 5 – экз. № Б-8410-7/115; окрестности г. Гурьевска, разрез Б-8410, слой 7; 6 – экз. № Б-8410-7/117, окрестности г. Гурьевска, разрез Б-8410, слой 7; 7 – экз. № Б-8410-7/120; окрестности г. Гурьевска, разрез Б-8410, слой 7; 8 – экз. № БС-093-16/86; окрестности г. Прокопьевска, разрез БС-093, слой 16; 9, 10 – Fidelites sp.: 9 – экз. № Б-8410-7/95; окрестности г. Гурьевска, разрез Б-8410, слой 7; 10 – экз. № Б-8410-7/123; окрестности г. Гурьевска, разрез Б-8410, слой 7; 11–13 – Cabrieroceras salairicum Nikolaeva, 2010: 11 – экз. № Б-8410-7/124; окрестности г. Гурьевска, разрез Б-8410, слой 7; 12 – экз. № Б-8410-7/125; окрестности г. Гурьевска, разрез Б-8410, слой 7; 13 – экз. № БС-091-13/69; окрестности г. Прокопьевска, разрез БС-091, слой 13.

Таблица II. Аммоноидеи среднего девона Салаира (верхний эйфель, генозона Agoniatites, заречный горизонт, акарачкинские слои; окрестности г. Прокопьевска). Все изображения даны в натуральную величину. Во всех случаях: а – вид с латеральной стороны, б – вид с вентральной стороны, в – вид с устья. 1–3 – Agoniatites vanuxemi (Hall, 1879): 1 – экз. № БС-091-13/54; разрез БС-091, слой 13; 2 – экз. № БС-093-16/36; разрез БС 093, слой 16; 3 – экз. № БС-091-13/74; разрез БС-091, слой 13; 4, 5 – Cabrieroceras salairicum Nikolaeva, 2010: 4 – экз. № БС-091-13/77; разрез БС-091, слой 13; 5 – экз. № БС-093-16/85; разрез БС 093, слой 16; 6 – Fidelites sp., экз. № БС-091-13/55; разрез БС-091, слой 13.

Таблица III. Конодонты из акарачкинских и сафоновских слоев заречного горизонта; верхи эйфельского яруса и живетский ярус, разрез Б-8410 (восточная окраина г. Гурьевска), разрезы БС-091 и БС-093 (северо-западная и западная окраины г. Прокопьевска). Длина масштабной линейки 100 мкм. 1, 2 – Icriodus aff. I. obliquimarginathus Bischoff et Ziegler, 1957, акарачкинские слои, разрез Б-8410, обр. Б-8410-6/кр: 1 – экз. № Б-8410/1: 1а – вид сверху, 1б – вид сбоку, 1в – вид снизу, 1г – вид сбоку; 2 – экз. № Б-8410/2: 2а – вид сверху, 2б – вид снизу; 3 – Icriodus sp. A, экз. № Б-8410/4: 3а – вид сверху, 3б – вид снизу, 3в – вид сбоку; акарачкинские слои, разрез Б-8410, обр. Б-8410-6/кр; 4, 5 – Polygnathus parawebbi Chatterton, 1974, акарачкинские слои, разрез Б-8410, обр. Б-8410-6/кр: 4 – экз. № Б-8410/3: 4а – вид сверху, 4б – вид снизу; 5 – экз. № Б-8410/5, вид сверху; 6 – Polygnathus sp., ювенильный экземпляр, экз. № Б-8410/6: 6а – вид сверху, 6б – вид снизу; акарачкинские слои, разрез Б-8410, обр. Б-8410-6/кр; 7 – Polygnathus sp., экз. № БС-091/2, вид сверху, акарачкинские слои, разрез БС-091, обр. БС-091-13/2.8; 8 – Polygnathus sp., экз. № БС-091/3, вид сверху, акарачкинские слои, разрез БС-091, обр. Е-834-7/1 (=БС-091-13/2.8); 9, 10, 13 – Polygnathus linguiformis klapperi Clausen, Leuteritz et Ziegler, 1979: 9 – экз. № БС-091/1: 9а – вид сверху, 9б – вид снизу; сафоновские слои, разрез БС-091, обр. БС-091-7/2.6; 10 – экз. № БС-093/1, вид сверху, сафоновские слои, разрез БС-093, обр. БС-093-7/0.05 м; 13 – экз. № БС-091/4, вид сверху, акарачкинские слои, разрез БС-091, обр. Е-834-7/1 (=БС-091-13/2.8); 11, 12 – Caudicriodus sp.: 11 – экз. № БС-091/5: 11а – вид сверху, 11б – вид снизу, 11в – вид сбоку; сафоновские слои, разрез БС-091, обр. БС-091-7/2.6; 12 – экз. № БС-093/2: 12а – вид сверху, 12б – вид снизу, 12в – вид сбоку; акарачкинские слои, разрез БС-093, обр. БС-093-21/2.0 м; 14 – Polygnathus aff. pseudofoliatus Wittekindt, 1966, экз. № БС-091/11: 14а – вид сверху, 14б – вид сбоку, акарачкинские слои, обр. Е-834-7/1 (=БС-091-13/2.8); 15 – Tortodus cf. T. obliquus (Wittekindt, 1966), экз. № БС-091/12, вид сбоку, акарачкинские слои, разрез БС-091, обр. БС-091-13/2.8; 16 – Tortodus planus (Bischoff et Ziegler, 1957), экз. № БС-091/13: 16а – вид сбоку, 16б – вид сверху; сафоновские слои, разрез БС-091, обр. БС-091-7/2.6; 17 – Neopanderodus aequabilis Telford, 1975, экз. № БС-091/14, вид сбоку, сафоновские слои, разрез БС-091, обр. БС-091-5/1.0; 18 – Belodella sp. A, экз. № БС-091/9, вид сбоку, акарачкинские слои, разрез БС-091, обр. БС-091-14/2.0.
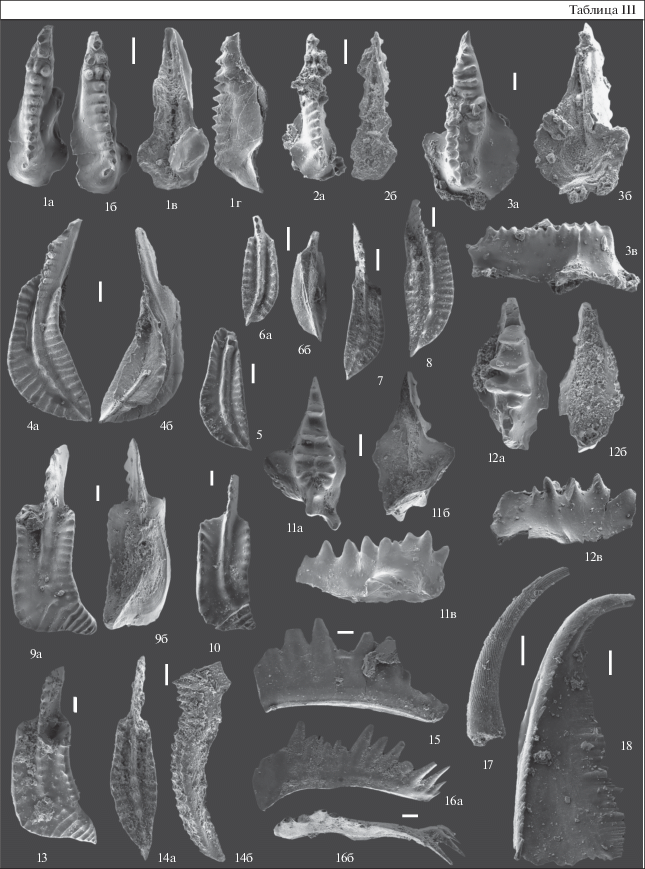
Таблица IV. Конодонты акарачкинских и сафоновских слоев заречного горизонта Салаира; верхи эйфельского яруса и живетский ярус, разрез БС-091 (западная окраина г. Прокопьевска). Длина масштабной линейки 100 мкм. 1 – Icriodus sp. C, экз. № БС-091/15: 1а – вид сверху, 1б – вид снизу, 1в – вид сбоку; акарачкинские слои, обр. Е-834-7/1 (=БС-091-13/2.8); 2 – Icriodus sp. D, экз. № БС-091/16: 2а – вид сверху, 2б – вид снизу, 2в – вид сбоку; акарачкинские слои, обр. Е-834-7/1 (=БС-091-13/2.8); 3, 5, 9 – Icriodus sp. E: 3 – экз. № БС-091/17: 3а – вид сверху, 3б – вид снизу, 3в – вид сбоку; акарачкинские слои, обр. Е-834-7/1 (=БС-091-13/2.8); 5 – экз. № БС-091/18: 5а – вид сверху, 5б – вид снизу, 5в – вид сбоку; сафоновские слои, обр. БС-091-7/0.9; 9 – экз. № БС-091/19: 9а – вид сверху, 9б – вид снизу, 9в – вид сбоку; сафоновские слои, обр. БС-091-3/3.7; 4 – Icriodus regularicrescens Bultynck, 1970, экз. № БС-091/20: 4а – вид сверху, 4б – вид снизу, 4в – вид сбоку; акарачкинские слои, обр. Е-834-7/1 (=БС-091-13/2.8); 6 – Icriodus sp. B, экз. № БС-091/21: 6а – вид сверху, 6б – вид сбоку; акарачкинские слои, обр. БС-091-14/1.0; 7, 8, 13 – Icriodus difficilis Ziegler, Klapper et Johnson, 1976: 7 – экз. № БС-091/8: 7а – вид сверху, 7б – вид снизу, 7в – вид сбоку; сафоновские слои, обр. БС-091-7/2.0; 8 – экз. № БС-091/33: 8а – вид сверху, 8б – вид снизу, 8в – вид сбоку; сафоновские слои, обр. БС-091-3/3.7; 13 – экз. № БС-091/22: 13а – вид сверху, 13б – вид снизу, 13в – вид сбоку; сафоновские слои, обр. БС-091-3/3.7; 10 – Icriodus eslaensis van Adrichem Boogaert, 1967, экз. № БС-091/23: 10а – вид сверху, 10б – вид снизу, 10в – вид сбоку; сафоновские слои, обр. БС-091-3/3.7; 11 – Icriodus brevis Stauffer, 1940, экз. № БС-091/24: 11а – вид сверху, 11б – вид снизу, 11в – вид сбоку; сафоновские слои, обр. БС-091-2/0.8; 12 – Icriodus aff. excavatus Weddige, 1984, экз. № БС-091/25: 12а – вид сверху, 12б – вид снизу, 12в – вид сбоку; сафоновские слои, обр. БС-091-3/3.7.
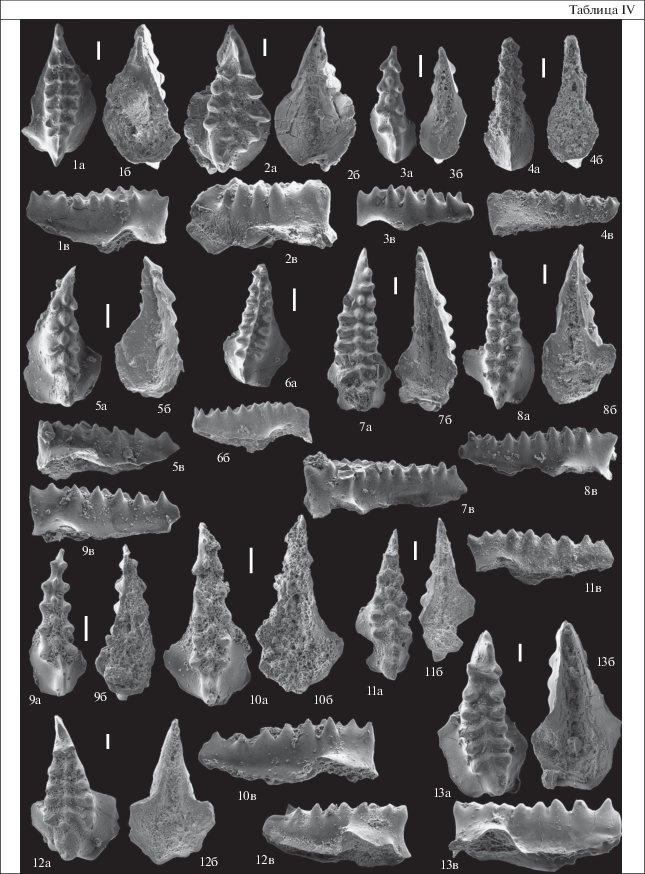
Таблица V. Остракоды сафоновских и керлегешских слоев заречного горизонта Салаира (местонахождения БС-092, БС-093, Б-8333, Б-8170). Длина масштабной линейки 100 мкм. 1, 2 – Ampuloides verrucosa Polenova, 1952, обр. Б-8170-11, сафоновские слои, ×50: 1 – экз. № 1/71, вид со стороны правой створки; 2 – экз. № 1/72, вид со стороны спинного края; 3, 4 – Libumella aff. discoides Rozhdestvenskaja, 1959, обр. Б-8170-3/2, керлегешские слои, ×30: 3 – экз. № 1/48, вид со стороны правой створки; 4 – экз. № 1/49, вид со стороны спинного края; 5, 6 – Bairdiocypris accuratus Polenova, 1960, обр. Б-8333-5/2, сафоновские слои, ×20: 5 – экз. № 1/1, вид со стороны правой створки; 6 – экз. № 1/2, вид со стороны спинного края; 7, 8 – Moorites legibilis Polenova, 1960, обр. БС-093-7/0.05 м, сафоновские слои, ×20: 7 – экз. № 1/42, вид со стороны левой створки; 8 – экз. №1/43, вид со стороны брюшного края; 9–12 – Bairdia safonovskiensis Polenova, 1960, обр. Б-8170-4/2, керлегешские слои, ×30: 9 – экз. № 1/3, вид со стороны левой створки; 10 – экз. № 1/4, вид со стороны правой створки; 11 – экз. № 1/5, вид со стороны спинного края; 12 – экз. № 1/6, вид со стороны брюшного края; 13–15 – Hanaites mirabilis Polenova, 1952, обр. Б-8170-3/2, керлегешские слои, ×50: 13 – экз. № 1/56, вид со стороны правой створки; 14 – экз. № 1/57, вид со стороны брюшного края; 15 – экз. № 1/58, вид со стороны правой створки; 16–18 – Sulcocavellina altschedatensis Polenova, 1960, обр. Б-8170-9, керлегешские слои, ×50: 16 – экз. № 1/39, вид со стороны левой створки; 17 – экз. № 1/41, вид со стороны спинного края; 18 – экз. № 1/40, вид со стороны брюшного края; 19–23 – Coeloenellina cavitata Rozhdestvenskaja, 1962, сафононовские слои, ×50: 19 – экз. № 1/36, обр. Б-8333-7, вид со стороны спинного края; 20 – экз. № 1/35, обр. Б-8333-5/3, вид со стороны спинного края; 21 – экз. № 1/34, обр. Б-8333-5/3, вид со стороны левой створки; 22 – экз. № 1/37, обр. Б-8333-5/3, вид со стороны брюшного края; 23 – экз. № 1/38, обр. Б-8333-5, вид со стороны левой створки; 24, 25 – Subligaculum aff. scrobiculatum Kesling et Mc Millan, 1951, обр. Б-8170-3/2, керлегешские слои, ×50: 24 – экз. № 1/62, вид со стороны левой створки; 25 – экз. № 1/63, вид со стороны брюшного края; 26, 27 – Miraculum ornatus Bakharev, 1985, обр. Б-8170-3/2, керлегешские слои, ×50: 26 – экз. № 1/16, вид со стороны правой створки; 27 – экз. № 1/27, вид со стороны левой створки; 28, 29 – Microcheilinella larionovae Polenova, 1955, обр. БС-092-2, керлегешские слои, ×30: 28 – экз. № 1/7, вид со стороны правой створки; 29 – экз. № 1/8, вид со стороны спинного края; 30, 31 – Microcheilinella givetica Rozhdestvenskaja, 1959, обр. Б-8170-9, керлегешские слои, ×50: 30 – экз. № 1/11, вид со стороны правой створки; 31 – экз. № 1/12, вид со стороны спинного края.

Таблица VI. Брахиоподы сафоновских и керлегешских слоев заречного горизонта Салаира. 1–3 – Indospirifer padaukpinensis (Reed, 1908): 1 – экз. № Я132/11: 1а – брюшная створка, 1б – спинная створка, 1в – замочный край, 1г – передний край, 1д – вид сбоку, все ×2.0; западная окраина Кузнецкого бассейна, с. Заречное Беловского района Кемеровской области; нижний живет, сафоновские слои, обр. Я-132; 2 – экз. № БС093-2z/2: 2а – брюшная створка, 2б – спинная створка, 2в – замочный край, 2г – передний край, 2д – вид сбоку, натуральная величина; западная окраина г. Прокопьевска, небольшой карьер у коттеджного поселка; нижний живет, сафоновские слои, обр. БС-093-2z; 3 – экз. № Я133/4, фрагмент раковины с микроскульптурой, ×2.0; западная окраина Кузнецкого бассейна, с. Заречное Беловского района Кемеровской области; нижний живет, сафоновские слои, обр. Я-133; 4 – Rhynchospirifer hians (v. Buch, 1837), экз. № Б8170-3/2: 4а – брюшная створка, 4б – спинная створка, 4в – замочный край, 4г – передний край, 4д – вид сбоку, все ×3.0; западная окраина Кузнецкого бассейна, карьер в 2 км южнее ст. Бускускан, живет, керлегешские слои, обр. Б-8170-3; 5 – Subrensselandia ? salairica (Ržonsnickaja, 1955), экз. № Б8170-3/5, целая раковина (разрушен передний край): 5а – брюшная створка, 5б – спинная створка, 5в – вид сбоку, 5г – замочный край, натуральная величина; западная окраина Кузнецкого бассейна, карьер в 2 км южнее ст. Бускускан, живет, керлегешские слои, обр. Б-8170-3.

Во время Международной полевой экскурсии в 2011 г. при показе “типовых” выходов сафоновских слоев (разрез Заречное, Б-8333, слой 11; рис. 9), в туфопесчаниках впервые были найдены позднеэйфельские аммоноидеи Agoniatites cf. vanuxemi (Hall) и Fidelites sp. По заключению ведущих специалистов по аммоноидеям девона, участвовавших в полевой экскурсии, Т. Беккера, К. Клюга, С.В. Николаевой, Е.С. Соболева и Н. К. Бахарева, вмещающие отложения указанных местонахождений аммоноидей (Б-8410, БС-091, БС-093 и Б-8333) следует относить к единому, довольно узкому биостратиграфическому интервалу генозоны Agoniatites верхнего эйфеля. Этот интервал отвечает верхней части акарачкинских слоев.
Выявленный комплекс аммоноидей акарачкинских слоев, как было отмечено, насчитывает около 300 экземпляров, которые равномерно распределены по всем четырем местонахождениям. Приведенный комплекс характеризует достаточно узкий биостратиграфический интервал генозоны Agoniatites верхнего эйфеля стандартной аммоноидной шкалы (MD I-F1), которая сопоставляется с конодонтовой зоной Polygnathus eiflius (Becker et al., 2020).
Конодонты. В типовом разрезе акарачкинских слоев ниже находок аммоноидей обнаружен позднеэйфельский комплекс конодонтов (рис. 3, табл. III). Этот комплекс включает таксоны Icriodus aff. obliquimarginatus Bischoff et Ziegler, Icriodus sp. A, Icriodus sp., Polygnathus costatus oblongus Weddige, Po. parawebbi Chatterton и Neopanderodus aequabilis Telford. Вид Po. costatus oblongus был описан из слоев Ahrdorf верхнего эйфеля Эйфельских гор и характеризует узкий интервал – конодонтовую зону Tortodus kockelianus (Weddige, 1977; Klapper, Johnson, 1980). Po. parawebbi появляется в зоне Tortodus kockelianus и проходит до нижней подзоны Polygnathus varcus (Klapper, Johnson, 1980 и др.). Экземпляры, отнесенные к Icriodus aff. obliquimarginatus Bischoff et Ziegler, по общему строению платформы наиболее близки к типовым экземплярам Icriodus obliquimarginatus, но отличаются от них скошенным назад задним краем платформы (табл. III, фиг. 1г). Для выделения нового таксона необходим дополнительный материал. Neopanderodus aequabilis известен в интервале от верхнего эмса до среднего живета Австралии (Telford, 1975; Anderson, 2003) и в нижнем–среднем живете Ирана (Adhamian, 2003; Ghobadi Pour et al., 2013).
Из этого же разреза М.Х. Гагиевым с соавторами (Ржонсницкая и др., 1985, 1990; Гагиев и др., 1986, 1987) был определен близкий комплекс конодонтов, включающий Polygnathus parawebbi Chaterton beta morphotype, Icriodus cymbiformis Branson et Mehl.
В разрезах, вскрывающих акарачкинские слои в придорожных карьерах (БС-091 и БС-093) в окрестностях г. Прокопьевска, конодонты представлены другими ассоциациями (рис. 5, 7, табл. 2, 3, табл. III, IV). В верхней части акарачкинских слоев (разрез БС-091), непосредственно в одном слое с позднеэйфельскими аммоноидеями, были найдены конодонты Icriodus regularicrescens Bultynck, Icriodus sp. B, Icriodus sp. C, Icriodus sp. D, Icriodus sp. E, Polygnathus linguiformis klapperi Clausen, Leuteritz et Ziegler, Po. aff. Po. pseudofoliatus Wittekindt, Neopanderodus aequabilis Telford, Tortodus cf. T. obliquus (Wittekindt), Belodella sp. A и др. В изученной коллекции преобладают представители рода Icriodus, среди которых выделены четыре группы: Icriodus sp. B, Icriodus sp. C, Icriodus sp. D. и Icriodus sp. E. Для отнесения их к новым видам необходимо получить дополнительный материал. Таксон Tortodus obliquus (Wittekindt) не выходит за пределы эйфельского яруса (Klapper, Johnson, 1980 и др.). Интервал распространения Polygnathus linguiformis klapperi Clausen, Leuteritz et Ziegler охватывает верхний эйфель и нижний–средний живет, от конодонтовой зоны Tortodus kockelianus до Ozarkodina semialternans (Clausen et al., 1979; Walliser, Bultynck, 2011). Polygnathus pseudofoliatus Wittekindt харктеризует верхнеэйфельские и живетские отложения (конодонтовые зоны от Tortodus australis до Polygnathus L. varcus) (Ziegler et al., 1976; Weddige, 1977; Klapper, Johnson, 1980; Барсков и др., 1991 и др.). Выявленный комплекс конодонтов акарачкинских слоев характеризует верхнюю часть конодонтовой зоны Tortodus kockelianus верхнего эйфеля.
Таблица 2.
Таксономический состав и количество конодонтовых элементов в акарачкинских и сафоновских слоях в разрезе БС-091
| Слои | Акарачкинские | Сафоновские | ИТОГО | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Виды | Образцы БС-091 | |||||||||||||||||||||||
| 21/3.0 | 14/2.0 | 14/1.2 | 14/1.0 | 13/3.0 | 13/2.8 | Е-834-7/1 (=13/2.8) | 13/2.0 | 12/0.85 | 10/5.2 | 9/4.0 | 7/3.4 | 7/2.6 | Е-834-7/4 (=7/2.6) | 7/2.0 | 7/0.9 | 5/1.0 | 5/0.1 | 3/3.8 | 3/3.7 | 3/1.5 | 2/4.7 | 2/0.8 | ||
| Belodella sp. A | 1 | 3 | 1 | 5 | ||||||||||||||||||||
| Panderodus unicostatus | 2 | 1 | 3 | |||||||||||||||||||||
| Icriodus regularicrescens | 1 | 1 | 2 | 4 | ||||||||||||||||||||
| Icriodus sp. B | 1 | 1 | 2 | |||||||||||||||||||||
| Neopanderodus aequabilis | 1 | 2 | 1 | 3 | 2 | 9 | ||||||||||||||||||
| Po. aff. P. pseudofoliatus | 3 | 3 | 6 | |||||||||||||||||||||
| Tortodus cf. T. obliquus | 1 | 1 | ||||||||||||||||||||||
| Polygnathus ssp. | 1 | 2 | 1 | 2 | 6 | |||||||||||||||||||
| Icriodus sp. D | 2 | 2 | ||||||||||||||||||||||
| Icriodus sp. C | 2 | 2 | ||||||||||||||||||||||
| Icriodus sp. E | 1 | 1 | 1 | 3 | ||||||||||||||||||||
| Polygnathus linguiformis klapperi | 2 | 2 | 1 | 1 | 6 | |||||||||||||||||||
| Tortodus planus | 1 | 1 | ||||||||||||||||||||||
| Caudicriodus sp. A | 2 | 4 | 6 | |||||||||||||||||||||
| Icriodus difficilis | 4 | 1 | 1 | 2 | 8 | |||||||||||||||||||
| Icriodus sp. | 1 | 1 | 2 | 1 | 5 | |||||||||||||||||||
| I. eslaensis | 4 | 3 | 7 | |||||||||||||||||||||
| I. aff. excavatus | 1 | 1 | ||||||||||||||||||||||
| I. brevis | 1 | 1 | ||||||||||||||||||||||
| Рамиформный элемент | 1 | 1 | 3 | 5 | ||||||||||||||||||||
| ИТОГО | 3 | 1 | 8 | 7 | 1 | 1 | 3 | 13 | 4 | 3 | 2 | 2 | 2 | 2 | 18 | 11 | 2 | 83 | ||||||
Таблица 3.
Таксономический состав и количество конодонтовых элементов в акарачкинских и сафоновских слоях в разрезе БС-093
| Слои | Акарачкин. | Сафоновские | ИТОГО | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Виды | Образцы БС-093 | |||||||||||||
| 21/2.0 | 21/1.0 | 16/1.3 | 15/1.6 | 15/0.1 | 15/1.6 | 10/1/2 | 8/0.5 | 7/0.05 | 6/1.5 | 6/0.08 | 4/3.8 | 2/2.0 | ||
| Caudicriodus sp. A | 1 | 1 | ||||||||||||
| Panderodus unicostatus | 1 | 1 | ||||||||||||
| Icriodus ssp. | 1 | 1 | ||||||||||||
| Neopanderodus aequabilis | 3 | 3 | ||||||||||||
| Polygnathus linguiformis klapperi | 2 | 1 | 3 | |||||||||||
| Рамиформный элемент | 1 | 1 | ||||||||||||
| ИТОГО | 1 | 1 | 1 | 1 | 5 | 1 | 10 | |||||||
В верхней части акарачкинских слоев (разрез БС-093), ниже находок позднеэйфельских аммоноидей, были найдены конодонты Caudicriodus sp. A (табл. 3; табл. III, фиг. 12).
В разрезе Заречное в толще конгломератов и туфогенных песчаников, относимых в настоящее время к акарачкинским слоям, известны находки конодонтов Polygnathus linguiformis linguiformis Hinde epsilon morphotype, Po. linguiformis linguiformis morphotype indet., Icriodus ex gr. expansus Branson et Mehl (Гагиев и др., 1987). Этот комплекс характеризует верхний эйфель (зона Polygnathus ensensis) и живетский ярус (до зоны Schmidtognathus hermanni).
Остракоды. В стратотипическом разрезе акарачкинских слоев (Б-8410, окраина г. Гурьевска) остракоды не обнаружены. Богатый комплекс остракод установлен в придорожном карьере на окраине г. Прокопьевска (разрез БС-091; рис. 3, табл. V). В верхней части акарачкинских слоев определены таксоны: Amphissites aff. concentricus Bakharev, Hanaites mirabilis (Polenova), Parabolbinella sp., Ampuloides verrucosa Polenova, Microcheilinella givetica Rozhdestvenskaya, Subligaculum aff. scrobiculatum Kesling at McMillan, Miraculum ornatus Bakharev, Bairdia safonovskiensis Polenova. Почти все установленные виды имеют широкое вертикальное и латеральное распространение, но есть и виды, имеющие только локальное распространение. Таксон Amphissites concentricus Bakharev известен в шандинском горизонте верхнего эмса Салаира (Бахарев, 1987). Miraculum ornatus Bakharev описан в керлегешских слоях среднего девона Салаира (Бахарев, 1986). Виды Bairdia safonovskiensis Polenova и Subligaculum aff. scrobiculatum Kesling at McMillan встречаются в сафоновских слоях Салаира (Поленова, 1960; Бахарев, 1985). Голотип Subligaculum scrobiculatum Kesling at McMillan описан из среднедевонских отложений штата Мичиган (США) (Kesling, McMillan, 1951).
Вид Hanaites mirabilis (Polenova) первоначально был обнаружен в старооскольском надгоризонте (живет) центральной части Восточно-Европейской платформы (ВЕП) (Поленова, 1952), а также в бийском (нижний эйфель) и афонинском (верхний эйфель) горизонтах Южного Урала (Рождественская, 1962). В Тимано-Печорской провинции этот вид встречен в бийском (нижний эйфель), кедровском и омринском (верхний эйфель) горизонтах, в верхней части возейшорской свиты (верхний эйфель) и в колвинской свите (верхний эйфель) Колвинского мегавала, а также в старооскольском надгоризонте (живет) южной части вала Сорокина (Москаленко, 2001). Данный таксон известен в верхнем живете Западной Померании Польши (Zbikowska, 1983). На Салаире ранее он был установлен в керлегешских слоях (Бахарев, 1985).
Вид Ampuloides verrucosa Polenova описан из старооскольского надгоризонта (живетский ярус) ВЕП (Поленова, 1952), а также встречен в изъельской свите (нижний и средний живет) Тимано-Печорской провинции (Москаленко, 2001) и в верхнем живете в Западной Померании Польши (Zbikowska, 1983). На Салаире этот вид ранее был описан из сафоновских слоев, однако найденные экземпляры отличаются от типовых присутствием более четкой борозды, ограничивающей заднюю часть раковины (Бахарев, 1985). Вид Microcheilinella givetica Rozhdestvenskaya встречен в нижней части старооскольского надгоризонта (живет) Западной Башкирии (Рождественская, 1959), в верхней части омринского горизонта и нижней части колвинской свиты (верхний эйфель) Тимано-Печорской провинции (Москаленко, 2001).
Установленный комплекс остракод акарачкинских слоев характеризует верхнеэйфельский интервал.
Сафоновские слои. Название стратиграфического подразделения дано М.А. Ржонсницкой (1952) по дер. Сафоново Кемеровской области. Стратотип сафоновского горизонта (слоев, свиты) располагался в южной водораздельной части гряды сопок правого берега р. Большой Бачат западной окраины с. Заречное Беловского района (Ржонсницкая, 1952). По результатам изучения типовых выходов сафоновского горизонта на восточном склоне Салаира у с. Заречное в 2009–2014 гг. было выявлено, что вскрытая здесь толща обнажена не полностью, в ней отмечаются зоны дробления и разломы, в связи с чем ее послойное достоверное описание не может быть составлено (Язиков и др., 2014). В качестве гипостратотипа сафоновских слоев предложен интервал слоев 12–2 разреза БС-091 западной окраины г. Прокопьевска (Язиков, 2014в; Язиков и др., 2014). Здесь представлен весь стратиграфический интервал сафоновских слоев, хорошо охарактеризованных фаунистически и имеющих нормальные нижнюю и верхнюю стратиграфические границы (рис. 5, 6). Мощность сафоновских слоев составляет около 40 м. Непосредственный контакт акарачкинских и сафоновских слоев фиксируется еще в двух разрезах восточного склона Салаира: в разрезе БС-093 на окраине г. Прокопьевска (рис. 7, 8) и в разрезе Б-8333 у с. Заречное (Middle-Upper…, 2011) (рис. 9, 10). Верхняя граница с керлегешскими слоями вскрыта также в разрезах БС-093, БС-092, Б-8333 и Б-8170 (рис. 11–14).
Рис. 14.
Схема корреляции опорных разрезов акарачкинских, сафоновских и керлегешских слоев заречного горизонта среднего девона восточного склона Салаира. Условные обозначения см. рис. 2.
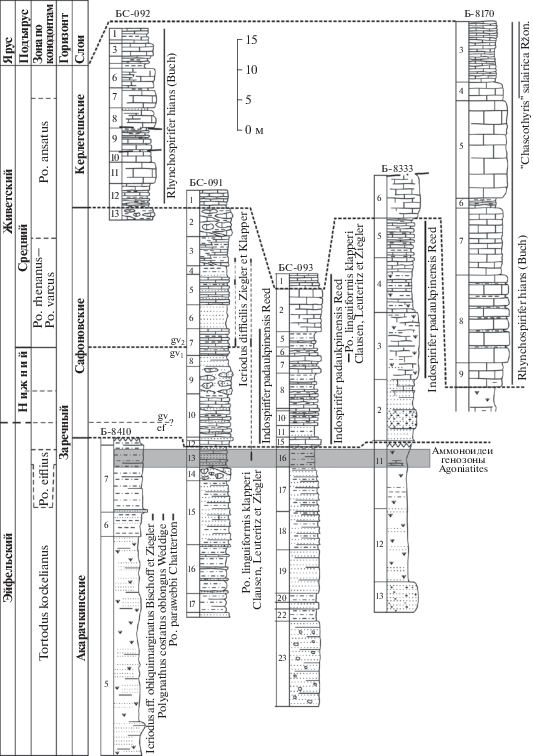
Ранее сафоновские слои рассматривались как перекрывающие керлегешские (Ржонсницкая, 1952, 1968; Решения…, 1982; Yolkin et al., 2000; Зональная…, 2006 и др.), но полученные в последнее время био- и литостратиграфические данные (Язиков, 2014а, 2014б, 2014в; Язиков и др., 2014, 2015а, 2016; Язиков, Изох, 2014а, 2014б) показали ошибочность таких представлений.
Брахиоподы. По брахиоподам сафоновские слои отвечают биозоне Indospirifer padaukpinensis (рис. 5, 7, 9, 13). Кроме I. padaukpinensis (Reed) (= Indospirifer pseudowilliamsi Ržonsnickaja) (табл. VI, фиг. 1–3) в разряд характерных для сафоновских слоев входят: “Rhynchospirina” lopatini (Stuckenberg), Radiomena irregularis (Roemer), Beckmannia angularis (Phillips) и Isospinatrypa givetica (Ržonsnickaja) (Ржонсницкая, 1952, 1968, 1975; Язиков, 2014а, 2014б, 2014в).
Сафоновская ассоциация брахиопод обнаруживает сходство с комплексом Падаукпинского известняка Мья́нмы (Бирма), описанным М. Андерсоном, А. Буко и Д. Джонсоном (Anderson et al., 1969). Общие элементы брахиоподовых ассоциаций: Aulacella eifeliensis (Verneuil), Xystostrophia umbraculum (Schlotheim), Radiomena irregularis (Roemer), Devonaria minuta (v. Buch), Plectospira ferita (v. Buch), Reticulariopsis eifelensis (Scupin), Indospirifer padaukpinensis (Reed), Bifida lepida (D’Archiac et De Vern), Cyrtina heteroclita (Defrance), а также морфологически близкие представители родов Schizophoria, Mystrophora, Productella, Variatrypa, Atrypa, Isospinatrypa, Athyris и др. – свидетельствуют об одновозрастности содержащих их отложений. Всесторонний анализ фауны Падаукпинского известняка привел М. Андерсона с соавторами (Anderson et al., 1969) к заключению о позднеэйфельском возрасте вмещающих отложений, что подтверждают и приводимые в работе конодонты Polygnathus linguiformis linguiformis Hinde, Po. sp. cf. Po. xylus sensu Bischoff et Ziegler, Belodella triangularis (Stauffer). Вместе с тем отмечалось, что в ряде местонахождений Южного Китая Indospirifer padaukpinensis (Reed) был обнаружен и в живетских отложениях в ассоциации со Stringocephalus sp. (Anderson et al., 1969, p. 119). Таким образом, можно сделать заключение о распространении Indospirifer padaukpinensis (Reed) в верхнем эйфеле и, по-видимому, нижнем живете Китая, Мья́нмы (Бирма) и Индии (Язиков, 2014в). Сходство комплекса брахиопод индоспириферового горизонта на Салаире с падаукпинскими слоями Бирмы и маоерхуанскими слоями Южного Китая также отмечали М.А. Ржонсницкая (1937), Н.П. Кульков и Л.Г. Перегоедов (2010). Этот вывод хорошо согласуется как с новым положением сафоновских слоев в сводном разрезе Салаира (в данной работе они рассматриваются в интервале от переходных слоев эйфеля и живета до середины среднего живета), так и с новыми данными по конодонтам (Язиков и др., 2014; Язиков, Изох, 2014а, 2014б).
Конодонты. Конодонты родов Polygnathus, Icriodus, Belodella и, вероятно, последних представителей рода Caudicriodus установлены только в верхней части сафоновских слоев (рис. 5, 7; табл. 2, 3; табл. III, IV). Ассоциация включает новые таксоны Belodella sp. A и Caudicriodus sp. A, а также Icriodus difficilis Ziegler et Klapper, I. eslaensis van Adrichem Boogaert, I. regularicrescens Bultynck, I. aff. excavatus Weddige, Icriodus sp. B, Icriodus sp. E, Neopanderodus aequabilis Telford, Panderodus unicostatus Branson et Mehl, Polygnathus linguiformis klapperi Clausen, Leuteritz et Ziegler, Polygnathus aff. pseudofoliatus Wittekindt, Polygnathus sp., Tortodus planus (Bischoff et Ziegler), Panderodus unicostatus (Branson et Mehl). Таксоны Caudicriodus sp. A, Belodella sp. A, Icriodus sp. E, Neopanderodus aequabilis, Polygnathus aff. pseudofoliatus Wittekindt и Polygnathus linguiformis klapperi проходят из нижележащих акарачкинских слоев.
Icriodus arkonensis встречается в широком интервале от верхнего эйфеля (зона Tortodus kockelianus) до верхнего живета (зона Schmidtognathus germanni) Канады, Марокко, Европы, России, Китая, Таджикистана и Австралии (Bardashev, Ziegler, 1985; Bultynck, 2003; Kononova, Kim, 2005; Narkiewicz, Bultynck, 2007 и др.). Icriodus eslaensis van Adrichem Boogaert известен в нижнем и среднем живете (зона Lower Polygnathus varcus) (Bultynck, 1987). П. Бултинк обосновал валидность этого таксона при изучении Icriodus brevis, I. lindensis и I. eslaensis из среднедевонских разрезов Марокко и Арденн (Bultynck, 1987).
Tortodus planus встречен в живетских отложениях Франции, конодонтовые зоны Polygnathus hemiansatus и Polygnathus ansatus (Brice et al., 1979; Narkiewicz, Bultynck, 2010).
Icriodus excavatus характеризует живетские (зона Polygnathus varcus) и франские отложения (зона Ancyrognathus triangularis) (Weddige, 1984).
Icriodus difficilis Ziegler et Klapper является видом-индексом одноименной зоны для мелководных фаций или зоны Polygnathus rhenanus–Polygnathus varcus, а также нижней границы среднего живета (Ziegler et al., 1976; Weddige, 1977; Bultynck, 2003; Bultynck, Gouwy, 2008). На Салаире он установлен в средней части сафоновских слоев (разрез БС-091, слой 7), и, соответственно, основание зоны Polygnathus rhenanus–Polygnathus varcus следует проводить внутри слоя 7 (обр. БС-091-7/2.6м).
В карбонатной пачке сафоновских слоев в разрезе Заречное, по данным М.Х. Гагиева и соавторов (Гагиев и др., 1986, 1987; Ржонсницкая и др., 1990), встречены космополитные таксоны конодонтов Polygnathus parawebbi Chatterton beta morphotype, Po. ovatinodosus Ziegler et Klapper, Po. pseudofoliatus Wittekind, Icriodus expansus Branson et Mehl и I. difficilis Ziegler et Klapper. В целом этот комплекс конодонтов характеризует среднюю подзону зоны varcus. Polygnathus pseudofoliatus известен в интервале верхнего эйфеля, нижнего и среднего живета, или конодонтовых зон kockelianus, ensensis, hemiansatus, нижней подзоны зоны varcus (Ziegler et al., 1976; Weddige, 1977; Klapper, Johnson, 1980; Барсков и др., 1991 и др.). Вид Po. ovatinodosus появляется внутри средней подзоны зоны varcus (= зона ansatus) и проходит в низы франского яруса (Ziegler et al., 1976; Klapper, Johnson, 1980; Аристов, 1988; Барсков и др., 1991 и др.). Icriodus difficilis Ziegler et Klapper известен в разрезах начиная со среднего живета (зона rhenanus/varcus) и проходит в нижний фран (Ziegler et al., 1976; Weddige, 1977; Klapper, Johnson, 1980; Bultynck, 2003). Вид Icriodus expansus Branson et Mehl более характерен для верхнего живета (Ziegler et al., 1976; Klapper, Johnson, 1980; Narkiewicz, Bultynck, 2007), но в то же время были отмечены находки этого вида в Центральной Канаде, начиная с зоны varcus (Sparling, 1995, 1999; Bultynck, 2003).
В настоящее время, после детального изучения сложно построенного в тектоническом отношении разреза в районе с. Заречное (рис. 10), мы пришли к выводу, что эта карбонатная пачка с конодонтами отвечает верхней части сафоновских слоев (Язиков, 2014в; Язиков и др., 2016).
Исходя из анализа имеющихся данных по конодонтам, сафоновские слои охватывают интервал от пограничных слоев эйфеля и живета до нижней части среднего живета или в конодонтовом биозональном стандарте соответствуют интервалу от зоны ensensis до Middle varcus (нижняя часть зоны ansatus).
Остракоды. Комплекс остракод сафоновских слоев был изучен на основе анализа коллекций из разрезов на окраине г. Прокопьевска (разрезы БС-091, БС-09), в Бускусканском карьере (разрез Б-8170) и на окраине с. Заречное (разрез Б-8333) (рис. 5, 7, 9, 13, табл. V). В разрезе БС-091 (рис. 5) комплекс остракод небольшой и представлен тремя таксонами Amphissites aff. concentricus Bakharev, Parabolbinella sp., Hanaites mirabilis (Polenova), проходящими из нижележащих акарачкинских слоев. В остальных разрезах остракоды встречены в верхней части сафоновских слоев.
Параллельный разрез БС-093 (слои 8–2; рис. 7, табл. V) содержит более богатый комплекс остракод: Ampuloides verrucosa Polenova, Hanaites mirabilis (Polenova), Bairdia safonovskiensis Polenova, Amphissites aff. concentricus Bakharev, Saumella salairica Bakharev, Bairdiocypris accuratus Polenova, Miraculum ornatus Bakharev, Nesamyslia sp., Microcheilinella givetica Rozhdestvenskaya, Rozhdestvenskajites sp., Libumella circulata Rozhdestvenskaya, Parabolbinella sp., Moorites legibilis Polenova (табл. V, фиг. 7, 8).
Из верхней части этого же разреза Н.И. Савиной были определены остракоды Bairdia aff. safonovskiensis Polenova (Мезенцева и др., 2007).
Стоит отметить, что большинство видов разреза БС-093 совпадает с видами, установленными в акарачкинских и сафоновских слоях разреза БС-091 (рис. 5, табл. V), но есть ряд таксонов, встреченных только в первом разрезе. Например, Saumella salairica Bakharev ранее был описан из сафоновского и керлегешского горизонтов Салаира (Бахарев, 1985). Bairdiocypris accuratus Polenova встречен в лебедянских слоях (верхний живет) северо-восточной окраины Кузнецкого бассейна (Поленова, 1960). Более широкое латеральное распространение имеет Libumella circulata Rozhdestvenskaya, известный из афонинского (верхний эйфель) горизонта Южного Урала (Рождественская, 1962), а также из кедровского и омринского горизонтов (верхний эйфель) Тимано-Печорской провинции (Москаленко, 2001). Moorites legibillis Polenova ранее встречен в керлегешских и сафоновских слоях Салаира, а также в лебедянских слоях северо-восточной окраины Кузнецкого бассейна и в бейской свите (верхний живет) Минусинской котловины (Поленова, 1960). Coeloenellina cavitata Rozhdestvenskaya впервые был описан из бийского горизонта (нижний эйфель) Южного Урала (Рождественская, 1962). Microcheilinella convexa Rozhdestvenskaya определен из афонинского горизонта (верхний эйфель) Южного Урала, но также встречен в старооскольском надгоризонте ВЕП (живет) (Рождественская, 1962) и кедровском и омринском горизонтах Тимано-Печорской провинции (Москаленко, 2001) (верхний эйфель).
В Бускусканском карьере (рис. 13, табл. V) в верхней части сафоновских слоев (слой 11) встречены виды: Bairdia plicatula Polenova, Microcheilinella givetica Rozhdestvenskaya, Libumella circulata Rozhdestvenskaya, L. aff. L. discoides Rozhdestvenskaya (табл. V, фиг. 3, 4), Evlanella gemella Moskalenko, Ampuloides verrucosa Polenova (табл. V, фиг. 1, 2), Clavofabellina aff. abunda Polenova, Knoxiella sp., Sulcocavellina altschedatensis Polenova, Saumella salairica Bakharev, Bairdia safonovskiensis Polenova, Bairdiocypris accuratus Polenova, Parabairdiacypris sp.
Вид Sulcocavellina altschedatensis Polenova известен в сафоновских и керлегешских слоях Салаира и лебедянских слоях северо-восточной окраины Кузнецкого бассейна (Поленова, 1960). Вид Clavofabellina abunda Polenova описан из вязовского горизонта (верхний эмс) Южного Урала (Поленова, 1955), а также из койвенского (верхний эмс), бийского (нижний эйфель), кедровского и омринского (верхний эйфель) горизонтов (Москаленко, 2001).
В этом комплексе встречены два вида, характерных для живетского яруса. Первый вид Elvanella gemella Moskalenko известен из верхней части колвинской свиты (нижний живет) Тимано-Печорской провинции (Москаленко, 2001). Второй вид Bairdia plicatula Polenova, описанный из старооскольского (живет) надгоризонта ВЕП (Поленова, 1952), встречается в чеславском (нижний и средний живет) горизонте Среднего Урала (Рождественская, 1962) и в изъельской свите (нижний и средний живет) Тимано-Печорской провинции (Москаленко, 2001).
В верхней части сафоновской свиты (окраина с. Заречное, разрез Б-8333; рис. 9) установлен следующий комплекс: Saumella salairica Bakharev, Bairdiocypris accuratus Polenova (табл. V, фиг. 5, 6), Libumella circulata Rozhdestvenskaya, Bairdia safonovskiensis Polenova, Sulcocavellina altschedatensis Polenova, Hanaites mirabilis (Polenova), Samarella reversa Polenova, Coeloenellina cavitata Rozhdestvenskaya (табл. V, фиг. 19–23 ), Microcheilinella mandelstami Polenova, Microcheilinella aff. convexa Rozhdestvenskaya. Первые четыре вида характерны для трех разрезов (БС-093, Б-8170, Б-8333), а пятый вид характерен для двух разрезов (Б-8170 и Б-8333).
Типовой вид Microcheilinella mandelstami Polenova ранее описан из старооскольского (живет) надгоризонта ВЕП (Поленова, 1952), а также встречен в изъельской (нижний и средний живет) свите Тимано-Печорской провинции (Москаленко, 2001).
На основании изученного комплекса остракод установлено, что сафоновские слои имеют раннесреднеживетский возраст.
Керлегешские слои. Название дано М.А. Ржонсницкой (1952) по дер. Большой Керлегеш Прокопьевского района Кемеровской области. Стратотип располагался у дер. Большой Керлегеш, по-видимому, “к северу от деревни по задернованному склону холма” (Мезенцева, 2009, с. 59). В настоящее время стратотип недоступен для изучения, в качестве неостратотипа предлагается интервал с 9 по 1 слой разреза Б-8170, расположенного в старом карьере в 2 км южнее ст. Бускускан (Бахарев, 1985; Язиков, 2014в) (рис. 13). Мощность керлегешских слоев составляет около 100 м. Керлегешские слои являются более молодыми, чем сафоновские, и соответствуют верхней половине живета, начиная с зоны Middle varcus (= ansatus). Контакт сафоновских и керлегешских слоев фиксируется в пяти разрезах: БС-091, БС-092, БС-093 (окрестности г. Прокопьевска), Б-8333 (с. Заречное) и Б-8170 (заброшенный карьер в 2 км южнее ст. Бускускан) (Middle-Upper…, 2011; Язиков, 2014в; настоящая работа, рис. 5, 7, 9, 11, 13, 14).
На восточном склоне Салаира контакт керлегешских слоев с отложениями более молодого возраста не наблюдается. В вышележащем интервале развиты эффузивно-интрузивные образования буготакско-митрофановского магматического комплекса. Керлегешские слои в меньшей степени охарактеризованы ископаемой фауной по сравнению с сафоновскими слоями. Кроме находок брахиопод, остракод, кораллов (Ржонсницкая, 1973 и др.), в них отмечены также и находки тентакулитов Heteroctenus aff. tagangaivi G. Ljaschenko (Клишевич, Мирецкая, 1986).
Брахиоподы. В керлегешских слоях установлены две брахиоподовые зоны: Rhynchospirifer hians (= Ilmenia subhians) и стратиграфически более высокая Subrensselandia? salairica (= “Chascothyris” salairica) (рис. 5, 7, 9, 11). Кроме видов-индексов (табл. VI, фиг. 4, 5) к числу характерных видов брахиопод по литературным данным относится ряд представителей близкородственных стрингоцефалид: Newberria (= Denckmannella) damesi (Holzapfel), N. circularis (Holzapfel) и Bornhardtina sp. (Ржонсницкая, 1968), однако они отсутствуют в изученной коллекции. Точная стратиграфическая привязка этих видов пока неясна.
Характерной чертой ассоциации брахиопод керлегешских слоев является ее крайне бедный видовой состав, обусловленный фациальными причинами. В ней присутствуют лишь виды-рифофилы, образующие банково-ракушняковые скопления (стрингоцефалиды) и псевдопланктонно-супербентосные (Грацианова, 1987) ринхоспириферы, рассеянные в осадках различного литологического состава. Все эти виды повсеместно в девоне указывают на живетское время, за исключением представителей Bornhardtina, находки которых известны и в эйфеле.
Остракоды. Остракоды керлегешских слоев получены из Бускусканского карьера (рис. 13, разрез Б-8170) и придорожного карьера на окраине г. Прокопьевска в правом борту безымянного ручья – левого притока руч. Егос (рис. 11, разрез БС-092).
В Бускусканском карьере (рис. 13) определен комплекс остракод: Knoxiella sp., Sulcocavellina altschedatensis Polenova (табл. V, фиг. 16–18 ), Saumella salairica Bakharev, Bairdia safonovskiensis Polenova (табл. V, фиг. 9–12), Bairdiocypris accuratus Polenova, Parabairdiacypris sp., Subligaculum aff. scrobiculatum Kesling et McMillan (табл. V, фиг. 24, 25 ), Coeloenellina sp., Hanaites mirabilis (Polenova) (табл. V, фиг. 13–15 ), Uchtovia abundans (Pokorny), Evanella sp., Cavellina sp., Paraschmidtella minussiensis Polenova, Microcheilinella mandelstami Polenova, M. givetica Rozhdestvenskaya (табл. V, фиг. 30, 31 ), Miraculum ornatus Bakharev (табл. V, фиг. 26, 27 ), Libumella aff. discoides Rozhdestvenskaya (табл. V, фиг. 3, 4).
В этом комплексе встречены виды, которые характерны только для живетского яруса. Первые семь видов проходят из нижележащих сафоновских слоев в этом разрезе. Uchtovia abundans (Pokorny) описан из мулинского (верхний живет) горизонта Западной Башкирии (Рождественская, 1959), также он встречается в Моравии (верхний живет), Чехия (Pokorny, 1950), в польской Померании (верхний живет) (Zbikowska, 1983) и в верхней части изъельской свиты (живет) Тимано-Печорской провинции (Москаленко, 2001). Вид Paraschmidtella minussiensis Polenova ранее был определен из бейской свиты (живет) Южно-Минусинской котловины (Поленова, 1960).
Экземпляры, отнесенные к виду Libumella aff. L. discoides Rozhdestvenskaya, близки к типовому виду Libumella discoides Rozhdestvenskaya (Бахарев, 1985), который изначально найден в бийском (нижний эйфель) горизонте Западной Башкирии (Рождественская, 1959) и Тимано-Печорской провинции (Москаленко, 2001).
В придорожном карьере на правобережье руч. Егос (рис. 11, разрез БС-092) комплекс остракод немного другой: Aparchitellina sp., Microcheilinella larionovae Polenova (табл. V, фиг. 28, 29 ), Orthocypris regula Polenova, Parabolbinella sp., Bairdiocypris accuratus Polenova, Hanaites mirabilis (Polenova), Sulcocavellina altschedatensis Polenova.
Для данного комплекса характерны остракоды, имеющие локальное распространение в пределах Салаира (Поленова, 1960). Вид Orthocypris regula Polenova имеет широкое вертикальное распространение внутри бассейна. Впервые он был определен из шандинских (верхний эмс) слоев, а также встречен в салаиркинских, мамонтовских, керлегешских и сафоновских слоях (Поленова, 1960).
Из широко распространенных видов установлены только два – Hanaites mirabilis (Polenova) и Microcheilinella larionovae Polenova. Сведения о Hanaites mirabilis даны при характеристике акарачкинских слоев, в которых он встречен. Таксон Microcheilinella larionovae Pol. первоначально описан в восточной части ВЕП в бийском (нижний эйфель) горизонте (Поленова, 1955), а позднее найден в койвенском (верхний эмс) горизонте Западной Башкирии (Рождественская, 1962), в Тимано-Печорской провинции он встречен в койвенском (верхний эмс) и бийском (нижний эйфель) горизонтах (Москаленко, 2001). Этот таксон определялся ранее и на Салаире в крековских (верхний лохков), салаиркинских (нижний эмс), шандинских (верхний эмс) слоях (Поленова, 1960).
Исходя из биостратиграфического анализа выделенного комплекса остракод, следует вывод о среднеживетском возрасте керлегешских слоев.
Позднеживетский комплекс остракод (слои с Bairdia carinata) ранее был установлен в нижней части изылинской свиты в бассейне р. Изылы дер. Вассино (Попов, 2019). В изылинский комплекс остракод из нижележащего керлегешского проходят три таксона: Parabolbinella sp., Bairdiocypris accuratus Pol. и Coeloenellina cavitata Rozh.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Проведенный анализ распространения аммоноидей, брахиопод, конодонтов и остракод в разрезах среднего девона Салаира позволил нам внести ряд изменений в существующую стратиграфическую последовательность местных подразделений. В серии разрезов пересмотрены взаимоотношения акарачкинских, сафоновских и керлегешских слоев (рис. 14). При переизучении и подготовке разрезов среднего девона Салаира к демонстрации участникам Международной полевой экскурсии в 2011 г., в предложенной рабочей стратиграфической схеме девона Салаира керлегешский горизонт был ошибочно сопоставлен с нижней частью мамонтовского горизонта, а сафоновский – с верхней частью этого горизонта, включая и акарачкинские слои (Middle-Upper…, 2011, fig. 5). Основанием для этого послужила корреляция отложений по одновозрастным ассоциациям аммоноидей, приведенных ранее в списках фауны акарачкинского и сафоновского горизонтов (Ржонсницкая, 1973; Николаева, Бахарев, 2010 и др.). Так, в известковистых алевролитах верхней части типового разреза акарачкинских слоев в районе Гурьевска (слой 7, разрез Б-8410) отмечались находки Agoniatites vanuxemi (Hall), Fidelites sp. и Cabrieroceras salairicum Nikolaeva (Middle-Upper…, 2011; Бахарев и др., 2012). Тот же комплекс аммоноидей был установлен в разрезах, относимых ранее к сафоновской свите, в окрестностях г. Прокопьевска: по левому берегу руч. Егос (рис. 5, разрез БС-091, слои 13, 16) и в небольшом карьере, расположенном на западной окраине города (рис. 7, разрез БС-093, слой 16) (Николаева, Бахарев, 2010; Middle-Upper…, 2011).
Во время Международной геологической экскурсии в 2011 г. при осмотре “типовых выходов” сафоновского горизонта у с. Заречное (Ржонсницкая, 1968), в туфогенной, терригенно-карбонатной пачке верхней части разреза (рис. 9, 10, Б-8333, пачка 11) впервые были найдены Agoniatites cf. vanuxemi (Hall) и Fidelites sp. Этот комплекс идентичен уже известным находкам аммоноидей в других местонахождениях акарачкинских слоев (разрезы Б-8410, БС-091, БС-093) и характеризует довольно узкий биостратиграфический интервал генозоны Agoniatites верхнего эйфеля. Новая находка аммоноидей позволила пересмотреть строение данного разреза, в котором кроме сафоновских присутствуют и акарачкинские слои (рис. 9, 10).
Непосредственный контакт акарачкинских и сафоновских слоев фиксируется в трех разрезах восточного склона Салаира: БС-091, БС-093 окрестностей Прокопьевска (рис. 5–8) и Б-8333 у с. Заречное (рис. 9, 10) (Middle-Upper…, 2011). Выше находок аммоноидей установлена ассоциация брахиопод, отвечающая зоне Indospirifer padaukpinensis и характеризующая сафоновские слои. Из этого следует, что сафоновские слои перекрывают акарачкинские слои и сопоставимы с интервалом от переходных слоев эйфеля к живету до нижней части среднего живета. Их стратиграфический объем соответствует конодонтовым зонам ensensis – Polygnathus rhenanus/ Polygnathus varcus) и низам зоны Polygnathus ansatus.
Контакт сафоновских и керлегешских слоев фиксируется в разрезах БС-091, БС-092 и БС-093 в окрестностях Прокопьевска (рис. 5–8, 11, 12), в центральной части разреза Б-8333 (рис. 9, 10) у с. Заречное и в разрезе Б-8170 (рис. 13), расположенном в карьере в 2 км южнее ст. Бускускан (Middle–Upper…, 2011 (разрезы приведены в обратной последовательности слоев); Язиков, 2014в; Язиков и др., 2014б). Керлегешским слоям свойственно массовое появление брахиопод Rhynchospirifer hians (v. Buch) (= Ilmenia subhians Ržonsnickaja) и Subrensselandia? salairica (Ržonsnickaja) (= “Chascothyris” salairica Ržonsnickaja). Керлегешские слои следует считать более молодыми, чем сафоновские, и они, вероятно, отвечают верхней части среднего живета, начиная с верхней части зоны Polygnathus ansatus (Язиков, Изох, 2014б; Язиков и др., 2014а).
В современном понимании телеутский надгоризонт включает мамонтовский и заречный горизонты и аналоги буготакского комплекса. Мамонтовский горизонт сопоставляется с интервалом верхнего эмса и большей части эйфельского яруса и отвечает конодонтовым зонам Polygnathus patulus, Polygnathus partitus, Polygnathus costatus и Tortodus kockelianus. Заречный горизонт коррелируется с верхней частью эйфельского яруса, нижней и средней частями живетского яруса или, по конодонтовой биозональности, с зонами Tortodus kockelianus (верхняя часть) Polygnathus eiflius, Polygnathus ensensis, Polygnathus hemiansatus, Polygnathus timorensis, Polygnathus rhenanus–Polygnathus varcus и Ozarkodina semialternans. По конодонтам положение верхней границы заречного горизонта в настоящее время точно не может быть определено из-за отсутствия видов-индексов зон, а также таксонов, характерных для этого интервала.
Контакт керлегешских слоев со стратиграфически более молодыми отложениями на восточном склоне Салаира не наблюдается. В вышележащем интервале развиты эффузивно-интрузивные образования буготакского магматического комплекса, в нижней части которого встречены прослои известняков. В этих известняках (с. Усть-Каменка) встречены конодонты Icriodus cf. expansus Branson et Mehl, характеризующие интервал от верхней части живетского яруса до франского яруса (зоны hermanni (верхний живет)–нижняя часть зоны punctata (базальные уровни среднего франа); Язиков и др., 2015б). Стратиграфический объем буготакской свиты с учетом полученных данных отвечает интервалу верхний живет–нижний фран (ориентировочно зоны hermanni–guanwushanensis) (с. Усть-Каменка, с. Вассино).
Изучение среднедевонских разрезов Салаира (окрестности с. Заречное и г. Прокопьевска) с отбором образцов мшанок, ругоз и аммоноидей были проведены О.П. Мезенцевой с соавторами (Мезенцева и др., 2007; Мезенцева, 2009; Гумерова и др., 2015 и др.). На основе биостратиграфического анализа комплексов фауны (мшанки, ругозы), а также данных по аммоноидеям (определения Н.К. Бахарева и С.В. Николаевой), эти авторы пришли к выводу о том, что нижняя граница живетского яруса может быть сопоставлена с основанием сафоновского горизонта, перекрывающего акарачкинский горизонт (Гумерова и др., 2015).
В период с 2005 по 2018 гг. на заседаниях СибРМСК по рассмотрению региональных стратиграфических схем девона Алтае-Саянской складчатой области нового поколения предлагались к обсуждению разные их варианты. В одном из них, представленном коллективом авторов (Гутак и др., 2018), средний девон Салаира включал мамонтовский, керлегешский и сафоновский горизонты. А ранее выделяемый акарачкинский горизонт авторы поместили в состав керлегешского горизонта, в качестве его “субаэрального аналога”, на основе находок в нем живетской флоры (Гутак и др., 2018, с. 7). С такой трактовкой нельзя согласиться ввиду того, что акарачкинские слои в их типовых выходах имеют достаточно обоснованную позднеэйфельскую фаунистическую характеристику (аммоноидеи, конодонты, брахиоподы) и занимают определенное положение в стратиграфической схеме девона Салаира.
Межрегиональная корреляция заречного горизонта, исходя из современных биостратиграфических данных, сводится к следующим вариантам. Заречный горизонт может быть скоррелирован с верхней частью мосоловского горизонта, черноярским горизонтом и старооскольским надгоризонтом Воронежской антеклизы Русской платформы, конодонтовая характеристика которых в последнее время была уточнена В.М. Назаровой и Л.И. Кононовой (2015); с верхней частью афонинского горизонта, с чусовским и чеславским горизонтами западного склона Урала; с верхней частью лангурского горизонта и высотинским горизонтом восточного склона Урала (Стратиграфические…, 1993; Sapel’nikov et al., 2000); со средней частью герасимовского горизонта Западно-Сибирской геосинеклизы (Стратиграфия…, 2001); частично с юктинским горизонтом Сибирской платформы (Матухин, 1995).
ВЫВОДЫ
На основании биостратиграфического анализа имеющихся коллекций макро- и микрофауны из типовых выходов акарачкинских, сафоновских и керлегешских слоев среднего девона Салаира была уточнена стратиграфическая последовательность региональных подразделений среднего девона Салаира. Результаты проведенных исследований свидетельствуют о более молодом возрасте керлегешских слоев, чем считалось ранее. Полученные результаты сводятся к следующим:
1. Выявленные комплексы аммоноидей в четырех изученных разрезах (Б-8410, окрестности г. Гурьевска; БС-091, БС-093, окрестности г. Прокопьевска; Б-8333, окрестности с. Заречное) являются одновозрастными и отвечают генозоне Agoniatites верхнего эйфеля. Этот комплекс является региональным и глобальным биомаркером и приурочен к верхней части туфогенной пачки в кровле акарачкинских слоев Салаира.
2. Установлена последовательность конодонтов среднего девона Салаира. В верхней части акарачкинских слоев ниже и вместе с находками аммоноидей (разрезы Б-8410, окрестности г. Гурьевска; БС-091, окрестности г. Прокопьевска) найдены позднеэйфельские комплексы конодонтов. В средней части сафоновских слоев (разрез БС-091, окрестности г. Прокопьевска) определены конодонты, характеризующие среднеживетский интервал, зону Polygnathus rhenanus–Polygnathus varcus (= Icriodus difficilis) и низы зоны Polygnathus ansatus. Керлегешские слои следует считать более молодыми, чем сафоновские, и они сопоставимы с верхней частью среднего живета, с верхней частью зоны Polygnathus ansatus (разрез Б-8333, окрестности с. Заречное).
3. Акарачкинские слои с аммоноидеями генозоны Agoniatites и ассоциацией конодонтов зоны Polygnathus eiflius верхнего эйфеля согласно перекрываются сафоновскими слоями с комплексом брахиопод зоны Indospirifer padaukpinensis (разрезы БС-091 и БС-093), а выше залегают керлегешские слои, охарактеризованные брахиоподами зоны Rhynchospirifer hians и стратиграфически более молодой зоны Subrensselandia? salairica (разрезы БС-092 и Б-8170).
4. Заречный горизонт включает акарачкинские, сафоновские и керлегешские слои верхней части эйфеля, нижней и средней частей живетского яруса и отвечает интервалу в конодонтовой зональной последовательности: зоны Tortodus kockelianus, Polygnathus eiflius, Polygnathus ensensis, Polygnathus hemiansatus, Polygnathus rhenanus–Polygnathus varcus, Polygnathus ansatus и Ozarkodina semialternans (рис. 2).
Полученные новые биостратиграфические данные показали, что действующая стратиграфическая схема расчленения среднего девона (Решения…, 1982) нуждается в пересмотре.
Благодарности. Авторы посвящают данную статью памяти своих учителей и коллег Е.А. Елкина, Н.К. Бахарева, Р.Т. Грациановой, В.А. Желтоноговой и А.Ю. Язикова.
Авторы благодарны Т. Беккеру (Германия), П. Бултинку (Бельгия), Я.М. Гутаку, К. Клюгу (Швейцария), Л.И. Кононовой, В.И. Краснову, Н.П. Кулькову, Т.Л. Модзалевской, В.М. Назаровой, К. Наркиевич (Польша), С.В. Николаевой, Л.Г. Перегоедову и др. за конструктивную критику, консультации и рациональные предложения при обсуждении полученных материалов и подготовке статьи. Выражаем также свою благодарность рецензентам Ю.А. Гатовскому, С.В. Николаевой и Т.Ю. Толмачевой за высказанные замечания и предложения.
Источники финансирования. Исследование выполнено в рамках проектов ФНИ №№ 0331-2019-0003, 0331-2019-0004 и 0331-2019-0005.
Список литературы
Аристов В.А. Девонские конодонты Центрального девонского поля (Русская платформа). М.: Наука, 1988. 120 с.
Барсков И.С., Воронцова Т.Н., Кононова Л.И., Кузьмин А.В. Определитель конодонтов девона и нижнего карбона. М.: Изд-во МГУ, 1991. 184 с.
Бахарев Н.К. Живетские остракоды Салаира // Микрофауна и биостратиграфия фанерозоя Сибири и смежных регионов. Новосибирск: Наука, 1985. С. 35–48.
Бахарев Н.К. Остракоды пограничных отложений нижнего и среднего девона Салаира. Автореф. дисс. … канд. геол.-мин. наук. Новосибирск: ИГиГ АН СССР, 1986. 26 с.
Бахарев Н.К. Новые виды остракод теленгитского надгоризонта (девон Салаира) // Микрофауна и биостратиграфия фанерозоя Сибири и смежных регионов. Новосибирск: Наука, 1987. С. 105–111.
Бахарев Н.К., Изох Н.Г., Язиков А.Ю. Новые данные и новые проблемы в стратиграфии нижнего и среднего девона Салаира // Материалы III Всероссийского совещания “Палеозой России: региональная стратиграфия, палеонтология, гео- и биособытия” (24–28 сентября 2012 г., СПб.). СПб.: Изд-во ВСЕГЕИ, 2012. С. 37–40.
Гагиев М.Х., Родыгин С.А., Тимофеева О.Б. Конодонтовые комплексы и зональное расчленение девонских отложений Салаира // Новые данные по стратиграфии докембрия и палеозоя Алтае-Саянской складчатой области. Новокузнецк: Запсибгеология, 1986. С. 49–51.
Гагиев М.Х., Ржонсницкая М.А., Родыгин С.А., Тимофеева О.Б. Конодонты и корреляция девонских отложений Салаира // Материалы по стратиграфии и палеонтологии Сибири. Томск: Изд-во ТГУ, 1987. С. 30–46.
Грацианова Р.Т. Уровни обитания брахиопод по отношению к дну бассейна // Среда и жизнь в геологическом прошлом. Палеоландшафты морей и континентов. Новосибирск: Наука, 1987. С. 41–58.
Гумерова Н.В., Мезенцева О.П., Забродина И.К. Ругозы и мшанки среднедевонских (раннеживетских) отложений северо-восточного Салаира // Международный журнал прикладных и фундаментальных исследований. Геол.-мин. науки. 2015. № 8. С. 924–928.
Гутак Я.М., Родыгин С.А., Перегоедов Л.Г., Макаренко С.Н., Антонова В.А. Региональная стратиграфическая шкала девона западной части Алтае-Саянской складчатой области // Геология и минерально-сырьевые ресурсы Сибири. 2018. № 1. С. 3–13.
Елкин Е.А. Типовые разрезы нижнего и среднего девона северо-восточного склона Салаира и их палеонтологическая характеристика. Новосибирск: ИГиГ, МСК, 1964. Ротапринт.
Елкин Е.А. Трилобиты (дехенеллиды) и стратиграфия нижнего и среднего девона юга Западной Сибири. М.: Наука, 1968. 154 с.
Елкин Е.А., Грацианова Р.Т., Бахарев Н.К., Желтоногова В.А. О ярусном расчленении среднего девона западной части Алтае-Саянской области // Средний девон СССР, его границы и ярусное расчленение. II Выездная сессия комиссии МСК по девонской системе на Урале. Уфа: БФАН СССР, 1983. С. 68.
Елкин Е.А., Изох Н.Г. Состав и особенности распределения конодонтов в стратотипических разрезах теленгитского надгоризонта среднего девона Салаира // Новые данные по стратиграфии докембрия и палеозоя Алтае-Саянской складчатой области. Новокузнецк: Запсибгеология, 1986. С. 51–52.
Зональная стратиграфия фанерозоя России. Научн. ред. Корень Т.Н. СПб.: Изд-во ВСЕГЕИ, 2006. 256 с.
Изох Н.Г., Язиков А.Ю. Схема расчленения нижнего девона Салаира // Корреляция алтаид и уралид: магматизм, метаморфизм, стратиграфия, геохронология, геодинамика и металлогеническое прогнозирование. Материалы Второго Российско-Казахстанского международного научного совещания, 1–4 апреля 2014 г., Новосибирск. Новосибирск: Изд-во СО РАН, 2014. С. 67–68.
Карцева Г.Н., Цырлина В.Б. Стратиграфия девонских отложений Кузнецкого бассейна // Тр. ВНИГРИ. Нов. сер. 1956. Вып. 95. Геол. сб. № 2. С. 147–164.
Клишевич В.Л., Мирецкая Е.М. Распространение тентакулитов в нижнем и среднем девоне Салаира и Алтая // Новые данные по стратиграфии докембрия и палеозоя Алтае-Саянской складчатой области. Тез. докл. к научно-технической конференции. Новокузнецк, 1986. С. 93–94.
Ключевые разрезы девона Рудного Алтая, Салаира и Кузбасса. Отв. редактор Елкин Е.А. Новосибирск: Изд-во СО РАН, 2004. 104 с.
Кульков Н.П., Перегоедов Л.Г. Новый взгляд на возраст керлегешских и сафоновских слоев среднего девона Салаира по брахиоподам // Эволюция жизни на Земле. Материалы Международного симпозиума, 10–12 ноября 2010 г. Томск: ТМЛ-Пресс, 2010. С. 261–265.
Лазуткин П.С., Наливкин Д.В., Ржонсницкая М.А., Яворский В.И. Девон Салаира // Проблемы советской геологии. 1936. № 8. С. 719–726.
Матухин Р.Г., Меннер В.В., Соколов П.Н., Решетняк Д.Р. Стратиграфическая основа девонской системы Сибирской платформы. Новосибирск: СНИИГГиМС, 1995. 82 с.
Мезенцева О.П. Новые данные о стратиграфии нижнеживетских отложений восточного склона Салаира // Природа и экономика Западной Сибири и сопредельных территорий. Том 1. Геология и палеонтология. Материалы Всероссийской конференции. Новокузнецк, 2009. С. 59–67.
Мезенцева О.П., Удодов В.П., Удодов Ю.В. Распространение трепостомид (мшанки) в нижнеживетских отложениях Салаира // Природа и экономика Кузбасса. 2007. Вып. 11. Т. 1. С. 38–49.
Москаленко М.Н. Среднедевонские остракоды Тимано-Печорской провинции. СПб.: ВСЕГЕИ, 2001. 230 с.
Назарова В.М., Кононова Л.И. Руководящие виды конодонтов среднего девона Воронежской антеклизы // Состояние стратиграфической базы центра и юго-востока Восточно-Европейской платформы. Материалы совещания. М.: ВНИГНИ, 2015. С. 51–55.
Николаева С.В., Бахарев Н.К. Находка рода Cabrieroceras (Werneroceratidae, Ammonoidea) на Салаире // Палеонтол. журн. 2010. № 3. С. 11–17.
Петц Г.Г. Материалы к познанию фауны девонских отложений окраин Кузнецкого угленосного бассейна. Санкт-Петербург, 1901. Т. 4. 393 с.
Поленова Е.Н. Остракоды верхней части живетского яруса Русской платформы // Микрофауна СССР. Сборник V. Л.: Гостоптехиздат, 1952. С. 65–156 (Тр. ВНИГРИ. Нов. сер. Вып. 60).
Поленова Е.Н. Остракоды девона Волго-Уральской области // Фораминиферы, радиолярии и остракоды девона Волго-Уральской области. Л.: Гостоптехиздат, 1955. С. 191–317 (Тр. ВНИГРИ. Вып. 87).
Поленова Е.Н. Девонские остракоды Кузнецкого бассейна и Минусинской котловины. Л.: Гостоптехиздат, 1960. 166 с. (Тр. ВНИГРИ. Вып. 152).
Попов Б.М. Биостратиграфические данные по остракодам из опорных разрезов среднего и верхнего девона окраин Кузнецкого бассейна (юго-восток Западной Сибири) // Геология и минерально-сырьевые ресурсы Сибири. 2019. № 2(38). С. 3–15.
Решения Всесоюзного совещания по разработке унифицированных стратиграфических схем докембрия, палеозоя и четвертичной системы Средней Сибири (Новосибирск, 1979 г.). Ч. II. Средний и верхний палеозой. Новосибирск: СНИИГГиМС, 1982. 129 с.
Ржонсницкая М.А. Брахиоподы индоспириферового горизонта (живетский ярус) юго-западной окраины Кузнецкого бассейна // Палеонтология и стратиграфия Кузбасса. Л.–М.: ЦНИГРИ, 1937. С. 97–138 (Тр. ЦНИГРИ. 1937. Вып. 97).
Ржонсницкая М.А. Спирифериды девонских отложений окраин Кузнецкого бассейна. М.: Госгеолиздат, 1952. 232 с.
Ржонсницкая М.А. К стратиграфии девонских отложений Кузнецкого бассейна // Сов. геол. 1959. № 9. С. 20–31.
Ржонсницкая М.А. Биостратиграфия девона окраин Кузнецкого бассейна. Стратиграфия. Л.: Недра, 1968. Т. 1. 288 с.
Ржонсницкая М.А. Кузнецкий прогиб и сопредельные районы Западной Сибири // Стратиграфия СССР. Девонская система. М.: Недра, 1973. Т. 2. С. 15–54.
Ржонсницкая М.А. Биостратиграфия девона окраин Кузнецкого бассейна. Описание брахиопод Pentamerida и Atrypida. Л.: Недра, 1975. 232 с. (Тр. ВСЕГЕИ. Т. 244).
Ржонсницкая М.А., Гагиев М.Х., Куликова В.Ф., Спасский Я.Н., Тимофеева О.Б. Граница эйфельского и живетского ярусов на Салаире // Средний девон СССР, его границы и ярусное расчленение. М.: Наука, 1985. С. 132–139.
Ржонсницкая М.А., Тимофеева О.Б., Гагиев М.Х. Конодонты среднего девона Салаира // Конодонты эйфельского яруса среднего девона. Казань: Изд-во КГУ, 1990. С. 46–57.
Рождественская А.А. Остракоды терригенной толщи девона западной Башкирии и их стратиграфическое значение // Материалы по палеонтологии и стратиграфии девонских и более древних отложений Башкирии. М.: Изд-во АН СССР, 1959. С. 117–180.
Рождественская А.А. Среднедевонские остракоды западного склона Южного Урала и платформенной части Башкирии // Брахиоподы, остракоды и споры среднего и верхнего девона Башкирии. М.: Изд-во АН СССР, 1962. С. 167–350.
Стратиграфические схемы Урала (докембрий, палеозой). Екатеринбург: ИГГ УрО РАН, Уралгеолком, 1993.
Стратиграфический словарь СССР. Л.: Недра, 1975. 621 с.
Стратиграфия нефтегазоносных бассейнов Сибири. Палеозой Западной Сибири. Редакторы Елкин Е.А., Краснов В.И. Новосибирск: Изд-во СО РАН, Филиал “Гео”, 2001. 165 с.
Фомичев В.Д., Алексеева Л.Э. Геологический очерк Салаира. М.: Госгеолтехиздат, 1961. 202 с. (Тр. ВСЕГЕИ. Нов. сер. Вып. 63).
Харин Г.С. Новые данные по стратиграфии девона восточного склона Салаира // Материалы по геологии Западной Сибири. М.: Госгеолтехиздат, 1958. Вып. 61. С. 74–85.
Яворский В.И. Девон юго-западной окраины Кузнецкого бассейна // Тр. ЦНИГРИ. 1938. Вып. 107. С. 2–36.
Язиков А.Ю. Характеристика брахиоподовых ассоциаций среднего девона Салаира // Интерэкспо ГЕО-Сибирь-2014. X Международный научный конгресс, 8–18 апреля 2014 г., Новосибирск. Международная научная конференция “Недропользование. Горное дело. Направления и технологии поиска, разведки и разработки месторождений полезных ископаемых. Геоэкология”. Сб. материалов в 4 т. Новосибирск: СГГА, 2014а. Т. 1. С. 236–240.
Язиков А.Ю. Филогения сибирских видов рода Protodouvillina (брахиоподы, девон) // Интерэкспо ГЕО-Сибирь-2014. X Международный научный конгресс, 8–18 апреля 2014 г., Новосибирск. Международная научная конференция “Недропользование. Горное дело. Направления и технологии поиска, разведки и разработки месторождений полезных ископаемых. Геоэкология”. Сб. материалов в 4 т. Новосибирск: СГГА, 2014б. Т. 1. С. 231–235.
Язиков А.Ю. Брахиоподы и биостратиграфия среднего девона складчатого обрамления Кузнецкого бассейна. Автореф. дисс. … канд. геол.-мин. наук. Новосибирск, 2014в. 19 с.
Язиков А.Ю., Изох Н.Г. Периодичность экосистемных перестроек в раннем и среднем девоне Салаира // Материалы LX сессии Палеонтологического общества при РАН (7–11 апреля 2014 г., Санкт-Петербург). СПб.: Изд-во ВСЕГЕИ, 2014а. С. 147–148.
Язиков А.Ю., Изох Н.Г. Биостратиграфия девона Салаира // Геология, геофизика и минеральное сырье Сибири. Материалы 1-й Научно-практической конференции. Новосибирск: СНИИГГиМС, 2014б. Т. 1. С. 254–258.
Язиков А.Ю., Изох Н.Г., Соболев Е.С. Взаимоотношения акарачкинского, сафоновского и керлегешского “горизонтов” среднего девона Салаира // Геология, геофизика и минеральное сырье Сибири. Материалы 1-й Научно-практической конференции. Новосибирск: СНИИГГиМС, 2014а. Т. 1. С. 250–254.
Язиков А.Ю., Изох Н.Г., Соболев Е.С. Схема расчленения среднего девона Салаира // Корреляция алтаид и уралид: магматизм, метаморфизм, стратиграфия, геохронология, геодинамика и металлогеническое прогнозирование. Материалы Второго Российско-Казахстанского международного научного совещания, 1–4 апреля 2014 г., Новосибирск, Россия. Новосибирск: Изд-во СО РАН, 2014б. С. 185–186.
Язиков А.Ю., Изох Н.Г., Соболев Е.С. Брахиоподы, конодонты и аммоноидеи в эмс-эйфельских стратотипических разрезах Салаира // Интерэкспо ГЕО-Сибирь-2015. XI Международный научный конгресс, 13–25 апреля 2015 г., Новосибирск. Международная научная конференция “Недропользование. Горное дело. Направления и технологии поиска, разведки и разработки месторождений полезных ископаемых. Геоэкология”. Сб. материалов в 3 т. Новосибирск: СГУГиТ, 2015а. Т. 1. С. 217–221.
Язиков А.Ю., Изох Н.Г., Широких В.А., Кутолин В.А. О возрасте буготакской свиты Колывань-Томской складчатой зоны по палеонтологическим данным // Интерэкспо ГЕО-Сибирь-2015. XI Международный научный конгресс, 13–25 апреля 2015 г., Новосибирск. Международная научная конференция “Недропользование. Горное дело. Направления и технологии поиска, разведки и разработки месторождений полезных ископаемых. Геоэкология”. Сб. материалов в 3 т. Новосибирск: СГУГиТ, 2015б. Т. 1. С. 212–216.
Язиков А.Ю., Изох Н.Г., Соболев Е.С. Эволюционные преобразования биот (брахиоподы, конодонты, аммоноидеи) в эмс-эйфельских отложениях Салаирского палеобассейна // Материалы LХII Сессии Палеонтологического общества. СПб.: Изд-во ВСЕГЕИ, 2016. С. 202–203.
Adhamian A. Middle Devonian (Givetian) conodont biostratigraphy in the Soh area, north of Esfahan, Iran // Cour. Forsch.-Inst. Senckenberg. 2003. V. 245. P. 183–193.
Anderson M.A. A study of conodont genera Belodella and Neopanderodus with emphasis on faunas from eastern Australia // Cour. Forsch.-Inst. Senckenberg. 2003. V. 245. P. 463–491.
Anderson M.M., Boucot A.J., Johnson J.G. Eifelian brachiopods from Padaukpin, northern Shan States, Burma // Brit. Mus. (Nat. Hist.) Bull. (Geology). 1969. V. 18. № 4. P. 105–163.
Bakharev N.K., Sobolev E.S. Ammonoidea and Middle Devonian biostratigraphy of the Salair // Biostratigraphy, paleogeography and events in Devonian and Lower Carboniferous (SDS/IGCP 596 joint field meeting). Contributions of Int. Conf. in memory of Evgeny A. Yolkin. Ufa, Novosibirsk, July 20–August 10, 2011. Novosibirsk: Publishing House of SB RAS, 2011. P. 27–33.
Bakharev N.K., Izokh N.G., Yazikov A.Yu. New Data on the stratigraphy of the Lower and Middle Devonian of Salair (West Siberia, Russia) // Int. Field Symp. “The Devonian and Lower Carboniferous of northern Gondwana” (Rabat, Morocco, 2013). Abstracts book. Eds. Hassani A.E., Becker R.T., Tahiri A. Documents de L’Institut Scientifique (Rabat, Morocco). 2013. № 26. P. 25–27.
Bardashev I.A., Ziegler W. Conodonts from a Middle Devonian section in Tadzhikistan (Kalagach Formation, Middle Asia, USSR) // Cour. Forsch.-Inst. Senckenberg. 1985. V. 75. P. 65–78.
Becker R.T., House M.R. Devonian ammonoid zones and their correlation with established series and stage boundaries // Cour. Forsch.-Inst. Senckenberg. 2000. V. 220. P. 113–151.
Becker R.T., Gradstein F.M., Hammer O. The Devonian Period // The Geologic Time Scale 2012. 2-volume set. Elsevier Science Ltd., 2012. P. 559–601.
Becker R.T., Marshall J.E.A., Da Silva A.-C., Agterberg F.P., Gradstein F.M., Ogg J.G. The Devonian Period // The Geological Time Scale 2020. Eds. Gradstein F., Ogg J.G., Schmitz M.D., Ogg G.M. Amsterdam: Elsevier, 2020. V. 2. P. 733–810.
Brice D., Bultynck P., Deunff J., Loboziak S., Streel M. Données biostratigraphiques nouvelles sur le Givétian et le Frasnien de Fergues (Boulonnais, France) // Ann. Soc. Geol. Nord. 1979. V. XCVIII. P. 325–344.
Bultynck P. Pelagic and neritic conodont successions from the Givetian of pre-Sahara Morocco and the Ardennes // Bull. Inst. R. Sci. Naturelles de Belgique, Sciences de la Terre. 1987. V. 57. P. 149–181.
Bultynck P. Devonian Icriodontidae: biostratigraphy, classification and remarks on paleoecology and dispersal // Rev. Española Micropaleontol. 2003. V. 35. P. 295–314.
Bultynck P., Gouwy S. Reference sections for the Middle Givetian substage // Newslett. SDS. 2008. № 23. P. 21–31.
Clausen C.D., Leuteritz K., Ziegler W. Biostratigraphie und Lithofazies am Sudrand der Elsper Mulde (hohes mittel-und tiefes Oberdevon: Sauerland, Rheinisches Schiefergebirge) // Geol. Jb. 1979. V. 51. S. 3–37.
Ghobadi Pour M., Popov L.E., Hosseini M., Adhamian A., Yazdi M. Late Devonian (Frasnian) trilobites and brachiopods from the Soh area, Central Iran // Mem. Assoc. Australasian Palaeontol. 2013. V. 44. P. 149–158.
Kesling R.V., McMillan G.W. Ostracods of the family Hollinidae from the Bell Shale of Michigan // Contrib. Mus. Paleontol. Univer. Michigan. 1951. V. IX. № 2. P. 45–81.
Klapper G., Johnson J.G. Endemism and dispersal of Devonian conodonts // J. Paleontol. 1980. V. 54. P. 400–455.
Kononova L.I., Kim S.-Y. Eifelian conodonts from the Central Russian Platform // Paleontol. J. 2005. V. 39. Supp. 2. 134 p.
Middle–Upper Devonian and Lower Carboniferous Biostratigraphy of Kuznetsk Basin. Field Excursion Guidebook. Int. Conf. “Biostratigraphy, paleogeography and events in Devonian and Lower Carboniferous” (SDS/IGCP 596 joint field meeting), Novosibirsk, July 20–August 10, 2011. Eds. Bakharev N.K., Izokh N.G., Obut O.T., Talent J.A. Novosibirsk: Publishing House of SB RAS, 2011. 98 p.
Narkiewicz K., Bultynck P. Conodont biostratigraphy of shallow marine Givetian deposits from the Radom-Lublin area, SE Poland // Geo. Quarterly. 2007. V. 51. № 4. P. 419–442.
Narkiewicz K., Bultynck P. The Upper Givetian (Middle Devonian) subterminus conodont Zone in North America, Europe and North Africa // J. Paleontol. 2010. V. 84. № 4. P. 588–625.
Pokorпý V. Ostracods of the Middle Devonian Red Corals Limestones of Celechovice // Sb. statn. geol. ust. Ceskosl. Repub. 1950. V. 17. P. 513–530.
Sapel’nikov V.S., Nasedkina V.A., Mizens L.I., Zenkova G.G. Devonian Stage boundaries of the Urals // Cour. Forsch.-Inst. Senckenberg. 2000. V. 225. P. 323–327.
Sparling D.R. Conodonts from the Middle Devonian Plum Brook Shale of north-central Ohio // J. Paleontol. 1995. V. 69. № 6. P. 1123–1139.
Sparling D.R. Conodonts from the Prout Dolomite of northcentral Ohio and Givetian (Upper Middle Devonian) correlation problems // J. Paleontol. 1999. V. 73. № 5. P. 892–907.
Telford P.G. Lower and Middle Devonian conodonts from the Broken River Embayment, North Queensland, Australia // Spec. Pap. Palaeontol. 1975. V. 15. P. iv + 96.
Walliser O., Bultynck P. Extinctions, survival and innovations of conodont species during Kacak Episode in South-Eastern Morocco // Bull. de l`Institut royal des Sciences naturelles de Belgique, Sciences de la Terre. 2011. V. 8. P. 5–25.
Weddige K. Die Conodonten der Eifel-Stufe im Typusgebiet und in benachbarten Faziesgebieten // Seckenbergiana Lethaea. 1977. V. 58. № 4/5. P. 271–419.
Weddige K. Zur Stratigraphie und Palaogeographie des Devons und Karbons von NE-Iran // Seckenbergiana Lethaea. 1984. V. 65. № 1/3. P. 179–223.
Yolkin E.A., Gratsianova R.T., Izokh N.G., Yazikov A.Y., Bakharev N.K., Alekseeva R.E., Erina M.V., Kim A.I., Shishkina G.R. Devonian standard boundaries within the shelf belt of the Siberian Old Continent (South of the Western Siberia, Mongolia, Russian Far East) and in the South Tien Shan // Cour. Forsch.-Inst. Senckenberg. 2000. V. 225. P. 303–318.
Zbikowska B. Middle to Upper Devonian ostracods from Northwestern Poland and their stratigraphic significance // Palaeontol. Polon. 1983. № 44. 108 p.
Ziegler W., Klapper G., Johnson J.G. Redefinition and subdivision of the varcus-Zone (Conodonts, Middle–?Upper Devonian) in Europe and North America // Geol. Palaeontol. 1976. V. 10. P. 109–140.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция