

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 1, стр. 94-114
Развитие природной среды на мысе Малый Чукочий (Колымская низменность) в среднем–позднем неоплейстоцене: реконструкция по микропалеонтологическим данным
О. Г. Занина 1, *, Д. А. Лопатина 2, **, Я. С. Овсепян 2, С. А. Кузьмина 3, А. Ю. Степанова 4, Е. Е. Талденкова 5, Т. А. Вишнивецкая 1, 6, Е. М. Ривкина 1
1 Институт физико-химических и биологических проблем почвоведения РАН
Пущино, Московская область, Россия
2 Геологический институт РАН
Москва, Россия
3 Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
4 Texas A&M University
College Station, USA
5 Московский государственный университет им. М.В. Ломоносова, географический факультет
Москва, Россия
6 University of Tennessee
37996 Knoxville, TN, USA
* E-mail: oksanochka_zet@mail.ru
** E-mail: dalopat@mail.ru
Поступила в редакцию 11.11.2020
После доработки 19.05.2021
Принята к публикации 06.07.2021
- EDN: USOUOZ
- DOI: 10.31857/S0869592X22010082
Аннотация
Представлены результаты микропалеонтологического изучения многолетнемерзлых отложений, вскрытых скважиной CHUK 1/17 на мысе Малый Чукочий, на побережье Восточно-Сибирского моря (Колымская низменность). Микропалеонтологический анализ с учетом литологического строения и основных физико-химических характеристик позволил оценить особенности формирования отложений в условиях смены прибрежно-морских обстановок на континентальные в среднем и позднем неоплейстоцене. Для данного региона получено свидетельство существования тундростепного биома, что доказывает его значительное продвижение на север. Настоящие исследования на примере рассматриваемой территории показывают важность изучения микрофоссилий для понимания истории региона в четвертичное время и представляют собой основу для дальнейшего микробиологического и метагеномного изучения отложений, вскрытых скважиной.
ВВЕДЕНИЕ
В статье приведены результаты изучения микрофоссилий из отложений, вскрытых скважиной CHUK 1/17, пробуренной в прибрежно-морских отложениях на мысе Малый Чукочий (Колымская низменность) (рис. 1). Исследование видимой части разреза мыса Малый Чукочий, состоящего из двух обнажений: Речное в устье р. Б. Чукочья и Морское на берегу Колымской губы, проводилось А.А. Архангеловым с соавторами (1979). Обнажения представляют собой два останца ледового комплекса высотой 25–30 м, протяженностью около 1 км, разделенных аласной котловиной. В разрезе выделяется верхняя толща коричневых алевритов (10–13 м); ниже залегают буровато-желтые пески (8–9, до 12 м) с подстилающими их опесчаненными алевритами с ледяными жилами (2–4 м), в основании разреза – толща сильнольдистых желто-зеленых алевритов с линзами и прослоями торфа (до 4–5 м). По радиоуглеродным данным для верхней толщи алевритов получена дата 33 200 ± 200 лет (МАГ 392), для песков – 39 000 лет (МАГ 390), для озерных толщ аласа – 8400 ± 100 лет (Кузнецов, 1979). В работе А.А. Архангелова с соавторами (1979) сделан вывод об озерно-аллювиальном генезисе исследованных толщ, приводятся данные по фауне млекопитающих, изученной А.В. Шером и относимой к позднему плейстоцену, а также результаты изучения спор и пыльцы Г.Г. Карташовой. Отмечена низкая насыщенность проб из этого разреза палиноморфами, а на основании анализа палиноспектров проб из нижнего, среднего и верхнего горизонтов обнажения Речное делается вывод о существовании тундровых ландшафтов со значительным участием луговых и степных ценозов. Т.К. Свиридовой и Л.Ф. Орловой (Каплина, 1979) описаны спектры с заметным содержанием пыльцы деревьев и кустарников (до 40%) из верхней части разреза мощностью 10 м. А.К. Васильчук (2007) приводит материалы изучения этого разреза Л.Л. Козачихиной, выделившей локальные палинозоны (ЛПЧ): ЛПЧ-1 Poaceae–Caryophyllaceae (26–33 м), ЛПЧ-2 Betula sect. Nanae–Alnaster (26–14 м), ЛПЧ-3 Bryales–Poaceae (10–14 м), ЛПЧ-4 Selaginella sibirica–Poaceae (3–8 м). Здесь также приводится датировка 13 850 ± ± 450 лет (Ri-262), полученная В.Г. Миллером с глубины 12.5 м.
Палеопедологический анализ разреза мыса Малый Чукочий показал наличие профилей трех погребенных почв в толщах обнажения, представленных ледовым комплексом (Губин, Занина, 2014). В публикациях Е.М. Ривкиной с соавторами (Ривкина, Гиличинский, 1996; Ривкина и др., 2006) приведены данные по содержанию и закономерностям распределения метана в кернах скважин, пробуренных как через обнажение, так и под урез реки. Коллективом авторов во главе с Д.А. Гиличинским (Gilichinsky et al., 2003) проводились микробиологические исследования как самой толщи, так и находящихся в ней криопэгов. Криопэги – линзы природных незамерзших высокоминерализованных растворов (свободных хлоридно-натриевых вод с минерализацией 60–300 г/л), имеющих отрицательную температуру (около –9°С). На Колымской низменности криопэги залегают на глубине от нескольких метров до нескольких десятков метров в мерзлых толщах морского происхождения возрастом около 120 тыс. лет (Gilichinsky et al., 2003; Abramov et al., 2021).
Результаты исследования образований ледового комплекса и аласа разреза мыса Малый Чукочий по кернам бурения в 1991 г. опубликованы Д.Г. Шмелевым (2015). Им изучен гранулометрический состав отложений, описана цикличность строения аласных толщ разреза, обусловленная колебаниями уровня водоема (спуск и наполнение термокарстового озера). В близлежащем районе, на левом берегу р. Б. Чукочья, примерно в 10 км выше по течению от устья, была пробурена 8-метровая скважина 2Д (Евсеев, Недешева, 1983). В цитированной работе приводятся результаты изучения химических свойств отложений, фауны фораминифер и остракод и сделан вывод о формировании рассматриваемых толщ в зоне осушки, сходной с современной зоной лайды, и мелководной части моря.
Цель данной работы – детальное микропалеонтологическое изучение отложений скважины CHUK 1/17 для уточнения генезиса отложений, реконструкции условий осадконакопления, ландшафтов и растительного покрова времени их формирования. Полученные данные послужат основой для дальнейшего изучения истории развития шельфа арктических морей в плейстоцене и влияния уровня моря на климат прилегающих территорий. В будущем планируются микробиологические и метагеномные исследования этих отложений с целью мониторинга поведения законсервированной в них биоты при повышении температуры мерзлоты и ее деградации. Это является актуальным в связи с прогнозами изменения климата Арктики в ближайшем будущем.
МАТЕРИАЛ И МЕТОДЫ
Скважина CHUK 1/17 на мысе Малый Чукочий (69°52.752′ с.ш., 159°40.153′ в.д.) пробурена в 2017 г. в ходе полевых исследований в зоне осушки Восточно-Сибирского моря (рис. 1). Буровые работы проводились сотрудниками лаборатории криологии почв Института физико-химических и биологических проблем почвоведения РАН (ИФХ-иБПП РАН) Т.А. Вишнивецкой, О.Г. Заниной, В.А. Сороковиковым. Поскольку целью экспедиции был поиск и отбор образцов морских отложений и связанных с ними криопэгов, бурение проводилось под урез моря. Высота устья скважины над урезом 2 м. Колонковым бурением установкой УКБ 12/25 было опробовано более 21 м. В результате бурения были вскрыты отложения четвертичного возраста, прочно сцементированные льдом, а также отложения сезонно-талого слоя и современная почва. Бурение проводили без промывки и продувки, что обеспечило сохранение мерзлого состояния кернов как для стерильного отбора образцов для микробиологических и метагеномных исследований, так и для их корректного литологического описания. Керны поднимали на поверхность через каждые 30–50 см, затем, после литологического описания и описания криотекстур, часть кернов в мерзлом виде помещали в морозильную камеру, а часть отбирали на влажность. Остаток образцов высушивали до воздушно-сухого состояния и затем использовали для анализа водной вытяжки, pH и других определений.
Аналитические исследования минеральной части (измерение актуальной кислотности, удельной электропроводности, содержание плотного остатка, катионов и анионов) проведены в Центре коллективного пользования ИФХиБПП РАН (Пущино) с использованием стандартных методик (Воробьева, 2006). Актуальную кислотность (рН водный) определяли потенциометрически, измеряя рН водных суспензий, полученных при соотношении массы грунта и объема воды 1 : 2.5. Анализ водной вытяжки проводили по общепринятым для почв методикам: удельную электрическую проводимость определяли кондуктометрически (кондуктометр Эксперт-002, Россия), содержание катионов Na+ – методом пламенной фотометрии (фотометр BWB-XP Perfomance Plus, BWB-Technologies, Великобритания), содержание катионов Mg2+ – комплексонометрическим методом, анионы ${\text{SO}}_{4}^{{2 - }}$ извлекали 0.2N раствором НСl и определяли гравиметрически, Cl– – аргентометрическим титрованием (по Мору).
Из отложений, вскрытых скважиной, детально изучены споры, пыльца, фитолиты, остатки насекомых, остракоды и бентосные фораминиферы. Обработку образцов для анализа палиноморф и фитолитов осуществляли с использованием сепарационного метода В.П. Гричука (Пыльцевой…, 1950; Палеопалинология, 1966). Подсчет фитолитов проводили в 5 вертикальных рядах стекла 24 × 24 мм. Подсчет палиноморф проводили до 200 зерен. При анализе мацератов фиксировалось количественное содержание диагностов повышенного увлажнения – диатомовых водорослей (морских и пресноводных форм) и спикул губок, а также проводилась качественная оценка остатков растительного детрита.
Изучение распределения по скважине бентосных фораминифер, остракод и остатков насекомых изначально не планировалось, но было принято решение провести исследование на материале, оставшемся после основных анализов, описанных выше. Так как не была использована стандартная методика отбора проб на эти группы микрофауны, анализ в данной работе следует считать качественным. Вес образцов и интервал отбора сильно различаются по разрезу, а малое количество фоссилий (например, для фораминифер менее 100 раковин) исключает возможность подсчета процентного соотношения видов в образцах. В основном изучались пробы весом около 1 кг, отобранные по керну из средней части интервалов мощностью от 20 до 50 см, а также пробы, оставшиеся после измерения влажности, весом около 100 г (рис. 2). Промывку осуществляли через сито с размером ячеек 100 мкм до светлых вод. Образцы высушивали и разбирали под бинокулярным микроскопом вручную.
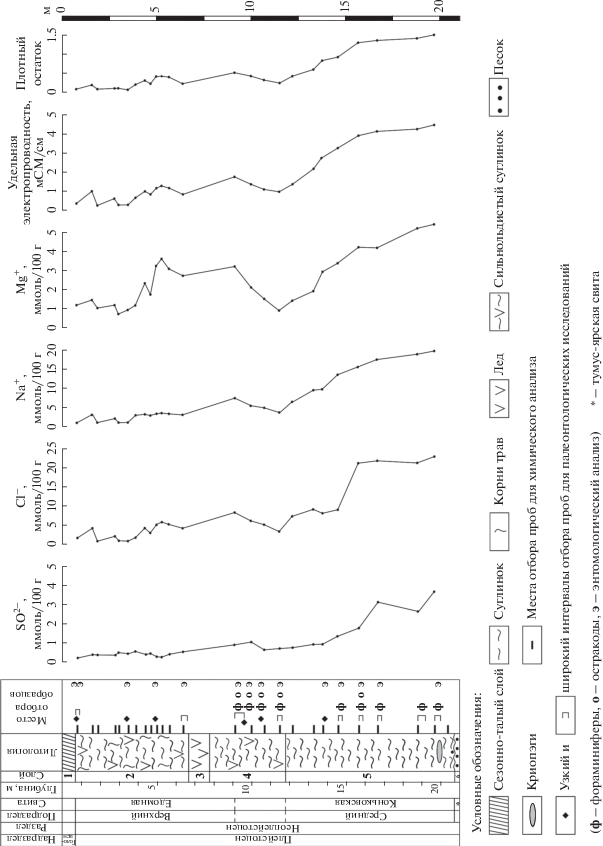
Рис. 2.
Схема строения разреза скважины CHUK 1/17 и некоторые физико-химические характеристики его отложений.
Биогенную фракцию образца исследовали и фотографировали в сканирующем электронном микроскопе Vega 3 Tescan в режиме высокого вакуума с использованием анализатора BSE. Электронные микрофотографии фитолитов, спор и пыльцы выполнены в Центре коллективного пользования ИФХиБПП РАН (Пущино), раковины фораминифер сняты в ГИН РАН. Створки остракод фотографировали при помощи светового микроскопа в университете A&M Texas, College Station.
РЕЗУЛЬТАТЫ
На основании первичного описания кернов скважины CHUK 1/17 вскрытая толща имеет следующее строение (рис. 2).
Слой 1 (0–0.7 м). Сезонно-талый слой (современная почва – криозем), представленный в верхней части (до 5 см) мертвым моховым опадом, ниже – серовато-бурым, хорошо оструктуренным суглинком, пронизанным корнями.
Вниз по разрезу идут мерзлые отложения:
Слой 2 (0.7–6.7 м). Сизый, с синевато-белесым отливом, сильнольдистый суглинок с тонкошлировой структурой, пронизанный тонкими корнями трав, с органическими включениями, охристыми пятнами и прожилками.
Слой 3 (6.7–7.8 м). Непрозрачный серый жильный лед с хаотично расположенными пузырьками воздуха и косыми прослойками органики.
Слой 4 (7.8.–11.8 м). Темно-сизый сильнольдистый суглинок.
Слой 5 (11.8–20.8 м). Темный до черного малольдистый пластичный суглинок, в интервалах 12.8–13.1, 19.2–19.7 м с запахом сероводорода. На глубине около 20 м обнаружены криопэги.
Слой 6 (20.8–21.0 м). Серый песок.
Химический анализ водной вытяжки показал, что состав водно-растворимых солей в отложениях преимущественно хлоридный и хлоридно-натриевый. В пробах до глубины 7.8 м содержание анионов ${\text{SO}}_{4}^{{2 - }}$ не превышает 0.5 ммоль/100 г, анионов Сl– колеблется в пределах 3–5 ммоль/100 г, катионов Na+ и Mg2+ в пределах 1–5 ммоль/100 г и 1–3 ммоль/100 г соответственно. Незначительное увеличение показателей Сl–, Na+ и Mg2+ зафиксировано на глубине 9 м. Начиная с отметки 14 м, содержание Сl– и Na+ увеличивается с достижением максимальных значений 21–24 ммоль/100 г в интервале 15.5–19.5 м. Для анионов ${\text{SO}}_{4}^{{2 - }}$ и катионов Mg2+ в этом интервале также фиксируется повышение значений. Низкие значения показателей удельной электрической проводимости и содержания плотного остатка отмечены в интервале до 7.8 м, затем следует их небольшое увеличение на 9 м и резкое возрастание начиная с 14 м, с максимальными отметками в интервале 15.5–19.5 м (рис. 2).
Значения рН отложений, вскрытых скважиной, колеблются в пределах 7.05–7.88 и являются нейтральными.
Микропалеонтологический анализ
Фитолиты. Являются единственными из всех микрофоссилий, которые определены во всех пробах скважины. Заметным содержанием фитолитов (150–200 экземпляров) характеризуются пробы из интервала разреза 9.6–20.5 м. Начиная с глубины 9.1 м, их количество сокращается почти в три раза и составляет в среднем 50 экземпляров. Исключением являются пробы с глубин 1.35 и 1 м, в которых зафиксировано максимальное для всего разреза содержание фитолитов (более 200 экземпляров). Процентное содержание всех описанных в керне скважины форм фитолитов приведено на рис. 3. Для удобства представления результатов фитолиты были разбиты на несколько групп по форме без учета размеров: длинные (гладкие цилиндрические, цилиндрические иглистые, пластинчатые, с волнистым краем), ланцетные (с широким основанием, с шипом), пластинки, округлые и трапециевидные (в том числе разнообразные лопастные симметричные и несимметричные формы), рондели, веерные (табл. I ).
Рис. 3.
Содержание кремнистых микробиоморф в разрезе скважины CHUK 1/17. Условные обозначения см. на рис. 2.

Таблица I . Фитолиты из скважины CHUK 1/17 на мысе Малый Чукочий, Колымская низменность. 1–4 – ланцетные формы; 5 – рондель; 6, 10, 11 – удлиненные формы; 7–9, 12–14 – веерообразные формы; 15, 19, 20 – округлые формы; 16, 21, 22 – трапециевидные лопастные формы, 11, 17, 18 – сильно корродированные формы.

Таблица II. Раковины фораминифер из скважины CHUK 1/17 на мысе Малый Чукочий, Колымская низменность. Длина масштабной линейки 100 мкм. 1 – Elphidium williamsoni Haynes, 1973: 1а – экз. 2, обр. 9.1-9.6, 1б – экз. 1, обр. 9.1–9.6, 1в – экз. 4, обр. 9.1–9.6, 1г – экз. 37, обр. 19.7–20; 2 – Elphidium subarcticum Cushman, 1944: 2a – экз. 18, обр. 9.6, 2б – экз. 3, обр. 9.1–9.6; 3 – Elphidium clavatum Cushman, 1930: 3а – экз. 15, обр. 9.6, 3б – экз. 39, обр. 19.7–20; 4 – Elphidium incertum (Williamson, 1858): 4а – экз. 19, обр. 9.6, 4б – экз. 23, обр. 10.5, 4в – вид со стороны устья, экз. 34, обр. 19.7–20, 4г – экз. 32, обр. 19.7–20; 5 – Haynesina orbiculare (Brady, 1881): 5а – экз. 14, обр. 9.6, 5б – экз. 9, обр. 9.1-9.6, 5в – вид со стороны устья, экз. 35, обр. 19.7–20; 6 – Elphidiella groenlandica (Cushman, 1933): 6а – экз. 26, обр. 10.5, 6б – экз. 17, обр. 10.5; 7 – Guttulina lactea (Walker et Jacob, 1798), экз. 10, обр. 9.1–9.6; 8 – Guttulina dawsoni Cushman et Ozawa, 1930, экз. 12, обр. 9.1–9.6; 9 – Quinqueloculina seminulum (Linnaeus, 1758), экз. 33, обр. 19.7–20; 10 – Reophax sp., экз. 22, обр. 10.5.
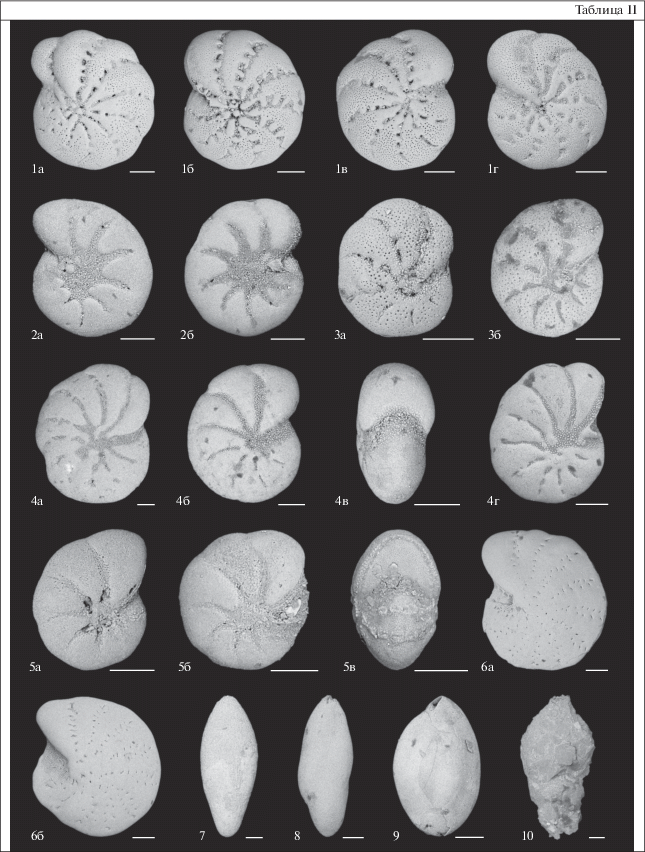
Таблица III. Створки остракод из скважины CHUK 1/17 на мысе Малый Чукочий, Колымская низменность. Длина масштабной линейки 100 мкм. 1 – Heterocyprideis sorbyana (Jones, 1856): 1а – левая створка, экз. 52, обр. 10.5, 1б –левая створка, экз. 53, обр. 11.4, 1в – карапакс, правая сторона, экз. 54, обр. 11.4; 2 – Paracyprideis pseudopunctillata Swain, 1963, правая створка, экз. 55, обр. 11.4; 3 – Roundstonia macchesneyi (Brady et Crosskey, 1871): 3а – правая створка, экз. 56, обр. 9.1, 3б – карапакс, левая сторона, экз. 57, обр. 9.6; 4 – Loxoconcha sp., левая створка, экз. 58, обр. 11.4; 5 – Cytherissa lacustris (Sars, 1863), правая створка, экз. 59, обр. 10.5; 6 – Limnocythere sanctipatricii (Brady et Norman, 1869), левая створка, экз. 60, обр. 10.5; 7 – Candona sp., левая створка, экз. 61, обр. 10.5; 8 – Pontocythere sp., карапакс, правая сторона, экз. 62, обр. 10.5.
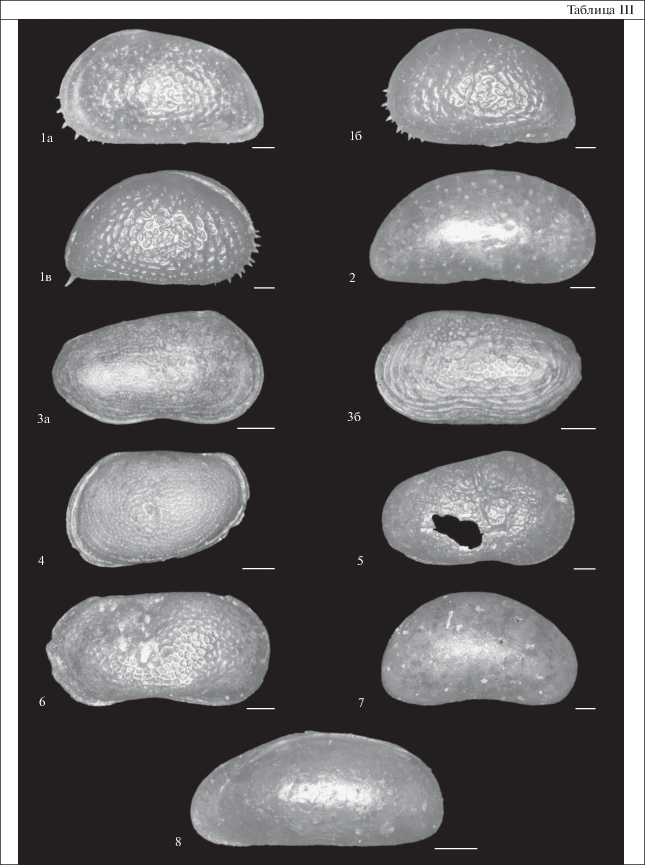
Таблица IV. Некоторые насекомые из скважины CHUK 1/17 на мысе Малый Чукочий, Колымская низменность. Длина масштабной линейки равна 1 мм. 1 – Stephanocleonus eruditus, голова, обр. Ch17-0 (инт. 0.7–0.9 м); 2 – Ceutorhynchus sp., левое надкрылье, обр. Ch17-6 (инт. 6.3–6.6 м); 3 – Euracmaeops angusticollis, основание левого надкрылья, обр. Ch17-10 (гл. 10.5 м); 4 – Morychus viridis, правое надкрылье, обр. Ch17-11 (инт. 11.4–11.6 м); 5, 6 – Micralymma brevilingue, переднеспинка и левое надкрылье, обр. Ch17-4 (инт. 4.9–5 м); 7 – Tachinus arcticus, правое надкрылье, обр. Ch17-11 (инт. 11.4–11.6 м); 8, 9 – Stenus sp., голова и правое надкрылье, обр. Ch17-4 (инт. 4.9–5 м); 10, 11, 12 – Aleocharinae gen. indet., голова, обр. Ch17-3 (инт. 3.4–3.5 м), переднеспинка, обр. Ch17-6 (инт. 6.3–6.6 м), левое надкрылье, обр. Ch17-11 (инт. 11.4–11.6 м); 13 – Gymmusa sp., правое надкрылье, обр. Ch17-10 (гл. 10.5 м); 14 – Curtonotus alpinus, переднеспинка, обр. Ch17-4 (инт. 4.9–5 м); 15 – Euura herbaceae, голова, обр. Ch17-3 (инт. 3.4–3.5 м).

В отложениях, вскрытых скважиной, прослеживается смена качественно различных фитолитных комплексов (ФК). Самый нижний ФК-I описан из слоя серого песка. Бóльшая часть фитолитов из этого комплекса сильно корродирована, их разнообразие невелико.
ФК-II выделен из темных пластичных малольдистых суглинков (пробы из интервала 14.4–20.4 м). Содержание фитолитов и разнообразие их морфотипов в этом комплексе достаточно высокие. В нем отмечены как хорошо сохранившиеся, так и сильно корродированные экземпляры, преобладают ланцентные формы с широким основанием, субдоминантами являются ланцетные с шипом, длинные гладкие цилиндрические и пластинчатые формы, а также округлые.
ФК-III описан из интервала разреза 4.2–13.1 м. Он выделяется по наличию и по преобладанию в большинстве проб веерообразных фитолитов, характерных для растений, произрастающих по берегам водоемов, по значительному содержанию ланцетных форм с широким основанием и округлых. Длинные цилиндрические и пластинчатые формы – субдоминанты и представлены приблизительно в таком же количестве, как и в ФК-II. Обращает на себя внимание заметное содержание разнообразных трапециевидных форм, преимущественно округлых и многолопастных симметричных.
ФК-IV выделен из верхней части слоя сильнольдистых суглинков с корнями трав в верхней части разреза. По сравнению с ФК из отложений, залегающих ниже, в данном комплексе доминируют длинные формы фитолитов – гладкие пластинчатые, цилиндрические и формы с волнистым краем. Установлено разнообразие трапециевидных форм, но их содержание ниже, чем в ФК-III. По сравнению с ФК-II и ФК-III здесь значительно снижается количество округлых фитолитов и ланцетных форм с широким основанием.
Диатомовые водоросли. Зафиксированы в небольшом количестве в большинстве проб из рассматриваемого разреза. Отмечено три уровня с их заметным содержанием. Первый установлен в интервале разреза 16.4–21 м; здесь определено значительное количество морских форм, преимущественно обломанных. Второй уровень с максимальным количеством панцирей пресноводных диатомей зафиксирован в интервале 1–1.35 м, третий уровень – поверхностные пробы, в которых присутствуют пресноводные формы (рис. 3).
Спикулы губок. Выявлены практически во всех пробах рассматриваемого разреза с заметным содержанием в интервале 13.1–21 м.
Растительный детрит. В пробах из нижнего интервала разреза (с глубин 16.4 и 19.2 м) содержится в значительном количестве, но бóльшая его часть бесструктурна, основу составляют проводящие ткани стеблей и корни, отмечены характерные ткани мхов. В пробе с глубины 10.5 м в составе растительного детрита отмечены остатки эпидермиса однодольных трав и ткани мхов, с глубины 9.1 м – проводящие ткани корней, а также специфические формы, характерные для древесины лиственницы. В пробах с глубин 4.2 и 3.2 м определен растительный детрит преимущественно однодольных трав и мхов, с глубины 1.35 м – детрит сфагновых мхов, с глубины 0.4 м – остатки тканей злаков и осок, в поверхностных пробах – разнообразные ткани мхов.
Споры и пыльца. Насыщенность спорами и пыльцой всех препаратов высокая, за исключением проб из нескольких интервалов. Палиноморфы не были обнаружены в самой нижней пробе из серого песка с глубины 21 м. В интервале из темно-сизых сильнольдистых суглинков и верхней части слоя темных пластичных малольдистых суглинков (9.7–12.4 м), а также в пробе с глубины 4.2 м из сильнольдистого суглинка с корнями трав обнаружены лишь единичные экземпляры спор и пыльцы. Из отложений, вскрытых скважиной, по количественному содержанию палиноморф выделены три спорово-пыльцевых комплекса (СПК) (рис. 4).
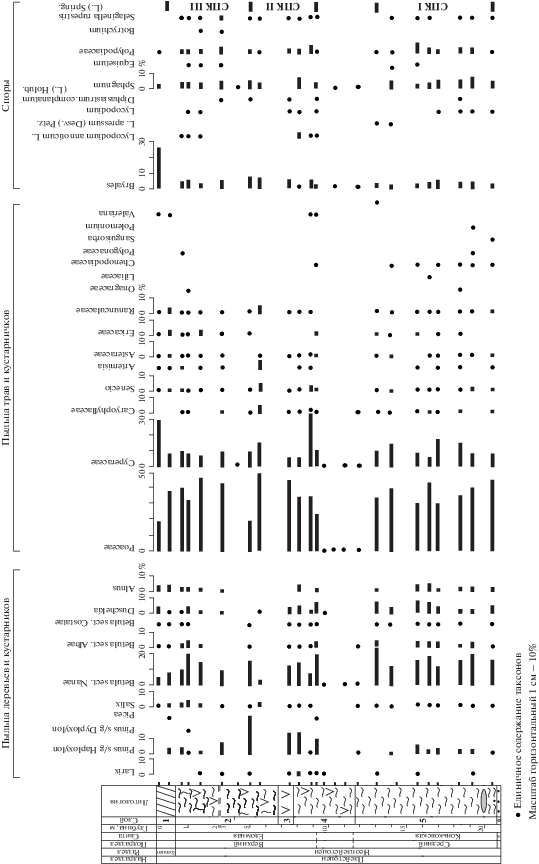
В самом нижнем, СПК I, из темного пластичного малольдистого суглинка (13.1–20.5 м) более половины от суммы всей пыльцы составляет пыльца трав и кустарничков с доминированием Poaceae (более 40%). Содержание пыльцы деревьев и кустарников 27–34.5% с преобладанием Betula sect. Nanae, спор в среднем 10%.
СПК II выделен из нижней части слоя сизого суглинка с тонкошлировой структурой, из слоя льда и темно-сизого сильнольдистого суглинка (5.0–9.1 м). Он отличается от СПК I бóльшим содержанием пыльцы деревьев и кустарников (34.5–46%). Количество пыльцы Betula sect. Nanae и Duschekia здесь примерно такое же, как и в СПК I, но стабильно присутствует единичная пыльца Larix, выше содержание Pinus s/g Haploxylon (10–25%). В группе трав и кустарничков (42–52%) доминируют Poaceae (23–44%). Среди спор в заметном количестве определены Bryales, Sphagnum и Lycopodium. Состав спектров из проб, отобранных с глубин 5.6 и 8.9 м, несколько “выпадает” из рассматриваемого СПК и, по-видимому, носит локальный характер. В них отмечено максимальное для данного разреза содержание пыльцы трав и кустарничков с преобладанием осоковых (проба с глубины 8.9 м) и злаковых (проба с глубины 5.6 м).
СПК III описан из верхней части разреза (0.4–3.2 м) и объединяет спектры близкого состава из сизого сильнольдистого суглинка с корнями трав и нижней части сезонно-талого слоя. СПК III характеризуется доминированием трав и кустарничков, среди которых преобладают Poaceae (порядка 40%), в меньшем количестве (до 10%) определены Cyperaceae, остальные семейства (Asteraceae, Ranunculaceae, Caryophyllaceae, Ericaceae) представлены единичными зернами. Содержание пыльцы деревьев и кустарников в среднем 20–27%, заметным количеством характеризуется пыльца Betula sect. Nanae (7.5–14%), остальные таксоны определены в количестве 2–5%. В группе спор преобладают споры Bryales и Sphagnum.
Во всех трех СПК в незначительном количестве (5–10 экземпляров) зафиксированы обрывки колоний зеленых водорослей рода Botryococcus.
В спектре из поверхностной пробы более половины составляет пыльца трав и кустарничков с доминированием Cyperaceae (32%) и Poaceae (18%); значительным содержанием характеризуются Bryales (26.5%).
Бентосные фораминиферы. Определены в девяти пробах из интервала разреза 9.1–20 м. Они представлены 13 видами, относящимися в основном к сем. Elphidiidae (табл. 1). Практически все фораминиферы с секреционно-известковой стенкой, за исключением одной агглютинированной раковины Reophax sp. с глубины 10.5 м (табл. II). Эльфидииды являются типичными представителями окраинных морей Российской Арктики. Виды Elphidium clavatum Cushman, 1930, Elphidium incertum (Williamson, 1858), Haynesina orbiculare (Brady, 1881), Elphidiella groenlandica (Cushman, 1933) и представители сем. Polymorphinidae (роды Guttulina и Polymorphina) приурочены в основном к опресненным областям в непосредственной близости от устьев рек, а также к самим эстуариям (Polyak et al., 2002).
Таблица 1.
Распределение бентосных фораминифер в осадках скважины Ch1/17
| Интервал, м | Cornuspira involvens | Elphidium williamsoni | Elphidium clavatum | Elphidium subarcticum | Elphidium incertum | Elphidiella groenlandica | Stainforthia loeblichi | Haynesina orbiculare | Quinqueloculina seminulum | Polymorphina sp. | Guttulina lactea | Guttulina dawsoni | Reophax sp. | Общее количество раковин/образец | Количество видов/образец | % Elphidium williamsoni | % Elphidium clavatum | % Elphidium subarcticum | % Elphidium incertum | % Elphidiella groenlandica | % Haynesina orbiculare |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 9.1–9.6 | 141 | 8 | 17 | 5 | 1 | 172 | 5 | 82 | 4.7 | 9.9 | |||||||||||
| 9.6 | 182 | 20 | 3 | 20 | 1 | 226 | 5 | 81 | 8.8 | 1.3 | 8.8 | ||||||||||
| 10.5 | 10 | 10 | 16 | 1 | 7 | 1 | 2 | 1 | 48 | 8 | 21 | 21 | 33 | 15 | |||||||
| 11.4–11.6 | 16 | 4 | 1 | 6 | 19 | 2 | 1 | 49 | 7 | 33 | 8.2 | 2 | 12 | 39 | |||||||
| 14.6–14.8 | 1 | 1 | 1 | ||||||||||||||||||
| 15.7–15.9 | 2 | 1 | 3 | 2 | 1 | 9 | 5 | ||||||||||||||
| 16.69–16.89 | 1 | 1 | 17 | 2 | 3 | 24 | 5 | ||||||||||||||
| 18.8–19.2 | 1 | 1 | 2 | 1 | 5 | 4 | |||||||||||||||
| 19.7–20.0 | 1 | 57 | 9 | 11 | 1 | 9 | 2 | 1 | 91 | 8 | 63 | 9.9 | 12 | 1.1 | 9.9 |
Данный обедненный комплекс ископаемых бентосных фораминифер по преобладанию видов, переносящих опреснение (Polyak et al., 2002), напоминает современные сообщества из эстуариев Оби и Енисея. Но он также схож с комплексами литорали Баренцева и Белого морей, в которых многочислен вид Elphidium williamsoni Haynes, 1973 (Корсун и др., 1994; Korsun et al., 2014). В Белом море E. williamsoni встречается на литорали и в нижней зоне заплеска на соленых болотах (маршах) при солености поверхностных вод 26–27‰ (Korsun et al., 2014). В Баренцевом море он найден на мягких грунтах литорали вдали от мест впадения рек, где соленость близка к нормально-морской (Корсун и др., 1994; Корсун, 1995). Однако в Балтийском и Северном морях, а также в проливе Скагеррак вид обитает в мелководных опресненных обстановках с пониженной соленостью от 0 до 25‰ (Murray, 2006).
Остракоды. Определены из интервала разреза 9.1–15.9 м (табл. III). Сравнительно большое количество ювенильных створок и карапаксов, составляющих более половины всех экземпляров, свидетельствует об отсутствии переотложения и накоплении их in situ. В комплексе доминируют эвригалинные виды Heterocyprideis sorbyana (Jones, 1856) и Paracyprideis pseudopunctillata Swain, 1963 и солоноватоводный вид Roundstonia (Cytheromorpha) macchesneyi (Brady et Crosskey, 1871). В небольшом количестве присутствуют мелководные морские виды рода Rabilimis. В пробе с глубины 10.5 м определены представители родов Pontocythere и Loxoconcha, которые встречаются преимущественно на опресненных прибрежных мелководьях, а также виды родов Limnocythere, Cytherissa и Candona, обитающие в пресных или слабосоленых водоемах (Wetterich et al., 2005; Frenzel et al., 2010) (табл. 2). Как и бентосные фораминиферы, остракоды, определенные в отложениях скважины CHUK 1/17, в основном представлены современными видами, характерными для прибрежных мелководий арктических морей, опресненных речным стоком (Cronin et al., 1991; Brouwers et al., 2000; Stepanova et al., 2003, 2007, 2019).
Таблица 2.
Распределение остракод в осадках скважины Ch1/17
| Интервал, м | Heterocyprideis sorbyana | Paracyprideis pseudopunctillata | Roundstonia macchesneyi | Rabilimis septentrionalis | Rabilimis sp. | Pontocythere sp. | Loxoconcha sp. | Limnocythere sp. | Cytheriissa sp. | Candona sp. | Общее количество створок/образец | Количество ювенильных створок/образец | Количество видов/образец |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 9.1–9.6 | 6с8юс 1юк |
7с 1к1юк |
27 | 12 | 2 | ||||||||
| 9.6 | 2к2юк | 1юс | 9 | 5 | 2 | ||||||||
| 10.5 | 18с23юс 3к1юк |
3юк | 1к | 1юс | 2с | 1с | 2с 1юк |
64 | 34 | 7 | |||
| 11.4–11.6 | 17с24юс 7юк |
5с2юс 1юк |
4юс 5юк |
3юс | 81 | 59 | 4 | ||||||
| 15.7–15.9 | 1к | 2 | 0 | 1 |
Палеоэнтомологический анализ. Для данного анализа из отложений скважины отобрано 15 проб, насекомые обнаружены во всех пробах за исключением одной – из интервала 18.8–19.2 м. В интервале разреза 14.6–21 м они представлены единичными фрагментами (табл. 3, табл. IV).
Таблица 3.
Распределение насекомых, пресноводных и наземных беспозвоночных в осадках скважины CHUK 1/17 на мысе Малый Чукочий, Колымская низменность
| Таксон | Eco_code | Глубина, м | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 19.7–20 | 16.69–16.89 | 15.7–15.95 | 14.6–14.8 | 13.8–13.9 | 11.4–11.6 | 10.5 | 9.6 | 9.1–9.6 | 6.3–6.6 | 4.9–5 | 3.4–3.5 | 0.7–0.9 | 0.7–0.8 | ||
| Phyllum Arthropoda, subphylum Hexapoda, class Insecta, ord. Coleoptera | |||||||||||||||
| Fam. Carabidae | |||||||||||||||
| Pterostichus (Cryobius) sp. | mt | 1 | 1 | 1 | |||||||||||
| Curtonotus alpinus Payk. | dt | 1 | 1 | ||||||||||||
| Harpalinae gen. indet | no | 1 | 1 | 1 | |||||||||||
| Carabidae gen. indet. | no | 1 | |||||||||||||
| Fam. Dytiscidae | |||||||||||||||
| Agabus moestus (Curt.) | aq | 1 | |||||||||||||
| Agabus sp. | aq | 1 | |||||||||||||
| Fam. Leiodidae | |||||||||||||||
| Cholevinus sibiricus (Jean.) | mt | 1 | |||||||||||||
| Fam. Staphylinidae | |||||||||||||||
| Olophrum sp. | pl | 1 | |||||||||||||
| Micralymma brevilingue Schtd. | dt | 1 | 1 | 1 | 1 | ||||||||||
| Tachinus arcticus Motsch. | mt | 1 | 2 | ||||||||||||
| T. brevipennis Sahlb. | mt | 2 | 1 | ||||||||||||
| Tachinus spp. (T. arcticus + T. brevipennis) | mt | 2 | 1 | 1 | 1 | ||||||||||
| Gymmusa sp. | pl | 1 | |||||||||||||
| Aleocharinae gen. indet. | pl | 1 | |||||||||||||
| Stenus sp. | ri | 1 | |||||||||||||
| Fam. Byrrhidae | |||||||||||||||
| Morychus viridis Kuzm. et Korot. | ss | 1 | 1 | 2 | 2 | 1 | 4 | 1 | 3 | 2 | 2 | ||||
| Simplocaria sp. | dt | 1 | |||||||||||||
| Fam. Cerambicidae | |||||||||||||||
| Euracmaeops angusticollis (Gebl.?) | me | 1 | |||||||||||||
| Fam. Chrysomelidae | |||||||||||||||
| Chrysolina sp. | st? | 1 | |||||||||||||
| Fam. Brachyceridae | |||||||||||||||
| Notaris bimaculatus F. | ri | 1 | |||||||||||||
| Fam. Curculionidae | |||||||||||||||
| Ceutorhynchus sp. | xe | 1 | |||||||||||||
| Hypera sp. | dt? | 1 | |||||||||||||
| Stephanocleonus eruditus Faust | st | 3 | 1 | ||||||||||||
| Cleonini gen. indet. | ms | 1 | |||||||||||||
| Ord. Hemiptera | |||||||||||||||
| Cicadellidae gen. indet. | me | 1 | 1 | ||||||||||||
| Ord. Hymenoptera, subord. Symphyta, fam. Tenthredinidae | |||||||||||||||
| Euura herbaceae (Cam.) | me | ||||||||||||||
| subord. Parasitica | |||||||||||||||
| Hymenoptera (Parasitica) gen. indet. | oth | 1 | 1 | 2 | |||||||||||
| Ord. Diptera | |||||||||||||||
| Chironomidae gen. indet. (larvae) | aq | 1 | 1 | ||||||||||||
| Diptera gen. indet. (puparia) | oth | 2 | 1 | ||||||||||||
| Ord. Trichoptera | |||||||||||||||
| Trichoptera gen. indet. (larvae) | aq | 1 | |||||||||||||
| Class Arachnida | |||||||||||||||
| spider chelicera | oth | 1 | |||||||||||||
| oribatid mites | oth | 2 | 2 | 3 | 42 | 1 | 5 | 2 | 1 | ||||||
| Subphylum Crustacea, class Branchiopoda, ord. Cladocera, fam. Daphniidae | |||||||||||||||
| Daphnia sp. winther eggs | aq | 1 | |||||||||||||
| Phylum Platyhelminthes, class Turbellaria | |||||||||||||||
| Turbellaria eggs | aq | 1 | 3 | 2 | 2 | ||||||||||
| Phylum Ectoprocta (Bryozoa), class Phylactolaema | |||||||||||||||
| Phylactolaema gen. indet. | aq | 1 | 2 | 2 | 3 | 2 | |||||||||
| Сумма | 4 | 2 | 4 | 4 | 1 | 11 | 12 | 8 | 52 | 5 | 11 | 19 | 9 | 9 | |
Примечание. Число указано в MNI (Minimum number of individuals), посчитанное с учетом того, что одна особь насекомого при захоронении разлагается на несколько фрагментов. Eco_code (экологический код): mt – тундровые мезогигрофилы, dt – тундровые ксерофилы, no – нет кода, aq – водные, pl – растительная подстилка, гниющие органические вещества и грибы, ri – прибрежные, ss – насекомые гемикриофитных степей, me – луговые, st – насекомые термофитных степей, xe – ксерофилы, ms – лугостепные, oth – прочие.
В самой нижней пробе (19.7–21 м) отмечен один неопределимый фрагмент ноги жужелицы и, кроме этого, три остатка пауков и мелкие панцири почвенных клещей отряда Oribatida.
В пробе из интервала 16.69–16.89 м найдено крыло цикадки (сем. Cicadellidae). Цикадки обычно приурочены к луговой растительности.
Начиная с глубины 14.6–14.8 м, количество и разнообразие насекомых постепенно увеличивается. Появляются тундровые стафилиниды рода Tachinus (T. arcticus Motsch. или T. brevipennis Sahlb., по надкрыльям трудно различимые), зимние яйца дафний – индикаторы пресноводных водоемов, четкие остатки тундростепного жука пилюльщика Morychus viridis Kuzm. et Korot.
С глубины 11.4–11.6 м насекомых становится заметно больше. Встречаются остатки типичных тундровых видов, таких как жужелицы Pterostichus (Cryobius) sp., стафилиниды Micralymma brevilingue Schtd., T. arcticus, остатки пресноводных организмов, таких как жуки плавунцы, статобласты мшанок, яйца планарий. Доминирует в комплексе тундростепной вид M. viridis.
В образце с глубины 10.5 м также доминирует M. viridis и присутствуют тундровые виды, но отмечен также фрагмент жука усача, скорее всего Euracmaeops angusticollis Gebl. Это луговой вид, который часто встречается в бореальной зоне, но в ископаемом состоянии пока не был отмечен.
Доминирование M. viridis продолжается до интервала 9.1–9.6 м. Выше, в интервале 6.3–6.6 м он исчезает, остаются единичные тундровые стафилиниды, пресноводные жуки и беспозвоночные.
В интервале 4.9–5 м вновь появляется M. viridis, но уже в меньшем количестве. Комплекс насекомых достаточно разнообразен, он включает тундровые жужелицы Curtonotus alpinus Payk., стафилиниды M. brevilingue, T. brevipennis, некоторые прибрежные (например, стафилинида Stenus sp.) и пресноводные организмы, почвенные клещи орибатиды.
В интервале 3.4–3.5 м и выше также имеются тундровые виды, заметно участие M. viridis, присутствуют остатки степных долгоносиков Stephanocleonus eruditus Faust. Кроме того, в пробе обнаружен пилильщик Euura herbaceae Cameron. Эти насекомые образуют галлы на карликовых видах ивы.
Состав энтомофауны по всему профилю позволяет реконструировать тундростепные условия (Киселев, 1981; Kiselev, Nazarov, 2009; Kuzmina, 2015). Роль ксерофильных насекомых возрастает снизу вверх. В нижних пробах, где заметно участие морских беспозвоночных, остатки насекомых единичны и мало информативны, но уже с глубины 15 м отмечается стабильное присутствие пилюльщика M. viridis. Этого жука можно назвать “лицом” плейстоценовой тундростепной фауны северо-востока Сибири и Чукотки (Киселев, 1981; Kuzmina, 2015). Он присутствует, часто доминирует практически в любом комплексе насекомых, а иногда его участие достигает 90%. Вместе с тем в современной фауне данный вид встречается лишь локально, на участках реликтовой ксерофильной растительности.
В верхней части разреза возрастает численность и разнообразие насекомых. Замечательным маркером остепнения служит появление долгоносика S. eruditus с глубины 3 м. Этот жук является индикатором термофильной вариации плейстоценовой тундростепи; в настоящее время его ареал находится значительно южнее, чем в плейстоцене (самые северные точки его распространения сейчас – реликтовые степи центральной Якутии). Жук отлично переносит низкие зимние температуры, но летом требователен к прогреву почвы. Данные условия обеспечивались высокой континентальностью климата в холодные периоды плейстоцена. Для проб характерно заметное участие водных насекомых и беспозвоночных. Мы предполагаем наличие внутриполигональных луж, столь присущих ландшафтам тундры.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В настоящей статье впервые был проделан большой объем комплексных исследований в целях реконструкции палеоусловий в конце среднего неоплейстоцена–позднем неоплейстоцене на мысе Малый Чукочий. В работе представлен новый материал, являющийся основой для дальнейших исследований района устья р. Б. Чукочья, в том числе и стратиграфических, поэтому заключения о возрасте исследуемых отложений следует считать предварительными.
Опираясь на представления о стратиграфическом расчленении отложений Колымской низменности по литературным данным (Решения…, 1987; Гиличинский, 2002; Gilichinsky et al., 2003, 2005; Ривкина и др., 2006; Abramov et al., 2021), литологические критерии, результаты химического анализа и данные по микрофауне, можно заключить, что слой 6 (серый песок, 20.8–21 м) относится к отложениям тумус-ярской свиты среднего неоплейстоцена (для данного интервала в настоящей работе не приведены реконструкции палеоусловий из-за отсутствия достаточного количества микропалеонтологических данных), слой 5 (темный, почти черный пластичный суглинок, 11.8–20.8 м), вероятнее всего, относится к морским отложениям коньковской свиты конца среднего неоплейстоцена, вскрытым скважинами по побережью Восточно-Сибирского моря между устьями рек Коньковая и Чукочья. На высоких элементах рельефа эта свита залегает под едомной свитой, на низких – под современными морскими террасами. Накопление отложений коньковской свиты мощностью до 20 м, представленной легкими суглинками и мелкозернистыми песками с древесным детритом, панцирями жуков, фрагментами губок, панцирями диатомей и фораминиферами, происходило во время последней обширной трансгрессии конца среднего неоплейстоцена, в прибрежной зоне моря, возможно в прибрежно-дельтовых или лагунных обстановках (Гиличинский, 2002). Рассматриваемая свита маркирует южную границу моря перед падением уровня Полярного бассейна в позднем плейстоцене и его отступлением на 300–400 км к северу. Сильнольдистые суглинки слоев 2 и 4 и лед слоя 3 (0.7–11.8 м) относятся к едомной свите верхнего плейстоцена, слагающей основные элементы рельефа Колымской низменности, слой 1 (0–0.7 м) – к голоцену.
Морские отложения из соседнего разреза, вскрытые скважиной 2Д, пробуренной в 10 км выше по течению от устья р. Б. Чукочья, были отнесены к постледниковой трансгрессии (поздний плейстоцен–ранний голоцен; Евсеев, Недешева, 1983). В нашем случае о морском генезисе отложений нижней части разреза (коньковская свита) свидетельствуют находки мелководных фораминифер и эвригалинных остракод. Единственную новую гипотезу об уточнении возраста можно выдвинуть для интервала 9.1–11.6 м (слой 4). Присутствие современных видов, также характерных для эпохи последнего межледниковья, увеличение биоразнообразия и обилие раковин фораминифер вида E. williamsoni (в более ранней литературе указан как Elphidium boreale) позволяют сделать предположение о возможной корреляции комплекса микрофауны из интервала 9.1–11.6 м скважины на Колымской низменности с комплексами микрофауны казанцевских отложений Западной Сибири (Гудина, 1969), эемских отложений северо-востока Белого моря (Grøsfjeld et al., 2006) и севера Европы (Feyling-Hanssen et al., 1971). Наличие E. williamsoni, которого относят к бореальной фауне (Гудина, 1969; Feyling-Hanssen et al., 1971), можно интерпретировать как свидетельство более теплых условий, что отличало моря во время трансгрессии в начале позднего неоплейстоцена. Вывод о потеплении климата подтверждается результатами спорово-пыльцевого анализа. В спектре из пробы с глубины 9.1 м фиксируется увеличение Betula sect. Nanae, Pinus s/g Haploxylon, появление единичных зерен Larix. В ФК-III из этого интервала определены фитолиты, характерные для крупномерной водной растительности, развитие которой происходило при более благоприятных климатических условиях.
Результаты микропалеонтологического анализа с учетом данных литологического строения и химического анализа исследованных толщ позволяют сделать выводы относительно условий накопления отложений, вскрытых данной скважиной. Сравнительно высокие содержания анионов ${\text{SO}}_{4}^{{2 - }},$ Сl–, катионов Na+, Mg2+, значения удельной электрической проводимости и содержания плотного остатка в пробах из темных пластичных малольдистых суглинков слоя 5, вероятнее всего относимых к коньковской свите верхов среднего неоплейстоцена, свидетельствуют о накоплении рассматриваемой толщи в морских условиях. Значения рН среды 7.05–7.8, присутствие в пробах обрывков колоний пресноводных зеленых водорослей и остатков пресноводных организмов (мшанки, яйца дафнии) позволяют предположить мелководные, сильно опресненные прибрежные условия. Данный вывод подтверждается результатами изучения микрофауны. В составе бентосных фораминифер и остракод доминируют типичные арктические виды, обитающие в прибрежных районах с сильным опреснением, такие как фораминиферы из группы “river-proximal species” (Polyak et al., 2002) и остракоды, характерные для внутреннего шельфа морей Лаптевых и Карского, включая эстуарии Оби и Енисея (Stepanova et al., 2003, 2007). О сильном опреснении палеобассейна говорит и отсутствие видов, типичных для арктических сообществ из районов с нормальной морской соленостью, например бентосных фораминифер семейства Cassidulinidae или остракод рода Cytheropteron. В самом основании изученного интервала (обр. 19.7–20.0 м) достаточно богатый в видовом отношении комплекс фораминифер свидетельствует о начальном затоплении территории и образовании мелководного приустьевого палеобассейна, вероятно, глубиной до 20 м.
Присутствие в рассматриваемом интервале разреза корродированных форм фитолитов с чешуйчатым типом коррозии, характерным для разрушения в водной среде на фоне значительного количества спикул губок (индикаторов холодных проточных вод), позволяет сделать вывод об аккумулятивном характере их осаждения при водном переносе. Не исключено, что часть фитолитов образовались не in situ, а были привнесены водами вместе со спикулами, что типично для гидрогенных слоев.
Можно предположить, что во время прижимных нагонных ветров происходило периодическое затопление мелководным морем прилегающих участков береговой суши. Не исключено, что здесь существовала лайда (разновидность соляных маршей), затопляемая во время морских приливов и высыхающая при отливах. Результаты анализа морфотипов фитолитов, на основании которого реконструируется луговая растительность, произрастающая на влажных почвах, подтверждают в-ывод о существовании лайды или периодически затопляемого заболоченного луга. В комплексе фитолитов доминируют ланцетные формы с широким основанием крупного (до 200 мкм) размера, характерные для травянистой высокорослой растительности влажных местообитаний, значительно содержание цилиндрических и длинных гладких пластинчатых форм, свойственных разнотравью, а также присутствуют ткани сфагновых мхов. В пробе из интервала 16.4–17.5 м содержание фитолитов увеличивается, что, вероятно, свидетельствует о более развитом травянистом покрове лайды. Широкое распространение травянистой растительности подтверждается результатами палинологического анализа. Состав СПК I с преобладанием пыльцы трав и кустарничков с доминированием злаковых отражает растительность, произраставшую в условиях холодного климата.
Выше по разрезу скважины фиксируется постепенное понижение показателей солености. На основании изучения микрофауны можно сделать вывод о том, что низы темно-сизых сильнольдистых суглинков слоя 4 (9.1–11.6 м), вероятнее всего, формировались в специфических условиях прибрежной зоны, для которой характерна периодическая смена морских и пресноводных условий. Учитывая низменный характер рельефа побережья в исследуемом районе, можно предположить, что появление остракод родов Limnocythere, Cytherissa и Candona, обитающих в пресных или слабосоленых водоемах, и увеличение процентного содержания “river-proximal” фораминифер (E. incertum, E. groenlandica) в пробе с глубины 10.5 м явились результатом перераспределения наносов, которые привели к образованию лагуны, впоследствии размытой. В образцах из интервала 9.1–9.6 м доля видов, связанных с сильным опреснением, падает, в то же время фиксируется небольшое повышение содержания анионов и катионов легкорастворимых солей, свидетельствующее об усилении влияния моря. Выше отметки 9.1 м морские беспозвоночные в разрезе скважины не встречаются. Возрастная принадлежность отложений из интервала 9.1–11.6 м остается дискуссионной. Находки в них морских микроорганизмов позволяют отнести их к образованиям коньковской свиты, которые были трансформированы эпикриогенетическими процессами и приобрели признаки едомной свиты верхнего неоплейстоцена.
Отличительной особенностью комплекса ФК-III из рассматриваемого интервала разреза является относительно высокое содержание, а в ряде проб преобладание веерообразных форм фитолитов, являющихся индикаторными для водной растительности, и остатков влаголюбивых мхов, вероятно образовывавших сплавины по берегам. Субдоминантами в этом комплексе являются ланцентные формы с широким основанием, являющиеся характерными для растений влажных местообитаний, а также длинные цилиндрические и гладкие пластинчатые формы, типичные для разнотравья. Таким образом, фитолитный анализ позволяет сделать вывод о существовании заозеренной лайды, заболоченного луга с травянисто-злаковой растительностью или пресноводного водоема с богатой прибрежной растительностью и развитым моховым покровом. Озера располагались вдоль берега моря и соединялись с ним протоками, по которым во время приливов в них поступала морская вода. Вывод о существовании пресноводного водоема подтверждается находками беспозвоночных – водных и прибрежных видов жуков, статобласт мшанок и яиц планарий.
В пробах из верхней части слоя 5 и нижней части слоя 4 (интервал 9.7–12.4 м) обнаружены лишь единичные экземпляры спор и пыльцы. Можно предположить, что в реконструируемом водоеме, для которого характерна смена прибрежно-морских и пресноводных условий, в определенные периоды происходила активная деструкция органического вещества.
Из верхней части слоя 4, льда слоя 3 и нижней части слоя 2, относимых к едомной свите, выделен СПК II с преобладанием злаковых и заметным содержанием пыльцы деревьев и кустарников, среди которых наряду с преобладанием карликовой березки, характерной для большей части толщи, в заметном количестве представлена пыльца Pinus s/g Haploxylon, вероятно принадлежащая кедровому стланику. Данный вид чувствителен к сезонному промерзанию и оттаиванию и является показателем наличия глубокого снежного покрова, хотя и не исключается усиление ветрового переноса пыльцы этого вида из более южных районов. В рассматриваемом СПК определена пыльца Larix в количестве трех-пяти зерен на препарат. Это число считается незначительным, когда речь идет о пыльце многих родов деревьев. Но пыльца лиственницы очень плохо сохраняется, поэтому данное количество позволяет фиксировать ее присутствие в составе реконструируемой растительности. Это подтверждается находками в составе растительного детрита специфических форм, характерных для древесины лиственницы, в пробе с глубины 9.1 м. Присутствие пыльцы лиственницы может косвенно свидетельствовать об озерно-болотном генезисе рассматриваемых отложений, в прибрежно-морской зоне она бы просто разрушилась. Состав данного комплекса позволяет сделать вывод о некотором потеплении климата и распространении на фоне господства тундровых ландшафтов островков лиственничников. Специфический состав спектров с глубин 5.6 и 8.9 м, в которых в значительном количестве присутствуют злаковые и осоковые, носит локальный характер и позволяет реконструировать растительность, произраставшую по влажным берегам водоема или заболоченного луга, что совпадает с результатами анализа фитолитов. Этот интервал слабо охарактеризован насекомыми, но в пробе 6.3–6.6 м найдены остатки жука плавунца Agabus moestus Curtis, типичного обитателя небольших стоячих тундровых водоемов.
Засоленность залегающих выше суглинков с включениями органики с корнями трав слоя 2, относимых к едомной свите верхнего неоплейстоцена (0.7–3.2 м), характеризуется минимальными значениями. Анализ микрофоссилий этих толщ показал присутствие в незначительном количестве спикул губок, заметное содержание панцирей пресноводных диатомей, достигающее максимума в пробах с глубин 1 и 1.35 м. Обилие диатомовых водорослей при небольшом количестве спикул губок свидетельствует о наличии органических веществ в воде, поскольку они способствуют развитию диатомей и подавляют развитие губок. В пробах с глубин 1 и 1.35 м также зафиксировано максимальное для данного разреза содержание фитолитов. Увеличение их количества свидетельствует о существовании более разнообразной растительности. Кроме того, высокое содержание фитолитов может являться показателем лучшей гумификации растительного материала в процессе диагенеза, в результате которого происходит их выделение из тканей. Процентное соотношение форм фитолитов во всех пробах слоя сходно. Преобладают разнообразные формы удлиненных фитолитов – цилиндрические, с волнистым краем, пластинчатые, являющиеся показателем значительного участия разнотравья в растительном покрове. Подавляющая часть удлиненных форм имеет сглаженные края, что при отсутствии игольчатых форм и ланцетных форм с длинным шипом указывает на гумидные условия. В незначительном количестве, но разнообразно представлены трапециевидные лопастные фитолиты, характерные, как и удлиненные фитолиты с волнистым краем, для растений влажных местообитаний. На основании палинологического анализа реконструируется травянистая растительность с доминированием злаковых и незначительным участием карликовой березки. Палеоэнтомологические данные указывают на постоянное господство в регионе в рассматриваемый продолжительный период времени тундростепных ландшафтов (Киселев, 1981; Kiselev, Nazarov, 2009; Kuzmina, 2015), при этом некоторые таксоны насекомых являются водными или околоводными, что подтверждает другие данные о том, что большую часть времени осадки формировались в субаквальных или локально переувлажненных субаэральных условиях, хотя в целом местность прилегала к обширной суше с преимущественно открытыми ландшафтами. Некоторое противоречие данных ботанического и энтомологического анализа часто имеет место при попытках реконструкции природной обстановки на северо-востоке Азии (Kuzmina, 2015). Как правило, насекомые показывают более значимое участие ксерофильных элементов биоты, чем палиноморфы и семена. Это связано как с тафономическими особенностями (семена водных растений более крупные и плотные, поэтому лучше сохраняются, пыльца лесных растений хорошо переносится ветром), так и с детальностью определения таксонов. Например, присутствие пилюльщика M. viridis указывает на то, что в его биотопе произрастали ксерофильные виды осоки, такие как Carex argunensis (Turcz. ex Trev.) Worosch., тогда как по пыльце осоки, как группы видов, принято делать реконструкции влажных условий. Но на основании присутствия насекомых нельзя отрицать наличие в ландшафте водоемов с комплексом прибрежной растительности, а также островков леса в большей или меньшей удаленности от местонахождения. Эти элементы ландшафта вписываются в понятие тундростепи. Кроме того, высокое содержание в ряде проб пресноводных диатомей, находки водных беспозвоночных, присутствие в ФК в заметном количестве характерных для гумидных условий форм являются показателем наличия переувлажненных участков понижений рельефа, возможно луж над головами ледяных жил, характерных для тундровых ландшафтов.
В поверхностных пробах отмечено относительно высокое количество пресноводных диатомей, что связано с сезонным переувлажнением горизонтов. Содержание спикул губок при этом единичное, что может служить показателем значительного содержания органики в водных растворах. В спектре из поверхностной пробы отмечено преобладание пыльцы осоковых, злаковых и зеленых мхов, что в целом отражает состав современной растительности.
ВЫВОДЫ
На основании результатов комплексных микропалеонтологических исследований, значительная часть которых проводилась для данного района впервые (изучение фораминифер, остракод, энтомофауны и фитолитов), и данных химического анализа отложений реконструированы условия осадконакопления и история развития ландшафтов и растительности мыса Малый Чукочий в среднем и позднем неоплейстоцене. Полученный материал по реконструкции палеоусловий будет являться основой для проведения микробиологических и метагеномных исследований.
Толща черных пластичных малольдистых суглинков (11.8–20.8 м) предположительно относится к коньковской свите верхов среднего неоплейстоцена, которая формировалась в мелководном морском бассейне. На это указывает как повышенная соленость рассматриваемых толщ, так и доминирование в составе бентосных фораминифер и остракод типичных ныне живущих арктических видов, обитающих в прибрежных районах, а также присутствие в отложениях обломков панцирей морских диатомей. Результаты фитолитного анализа позволяют предположить наличие на изучаемой территории лайды, заболоченного луга на прибрежной равнине, затопляемого во время приливов. На основании палинологического анализа реконструирована тундровая травянистая растительность с незначительным участием карликовой березки.
Отложения, вскрытые скважиной в интервале 9.1–11.6 м, формировались в специфических условиях прибрежной мелководной зоны, для которой была характерна периодическая смена морских и пресноводных условий. На основании присутствия в значительном количестве индикаторных фитолитов водной растительности, влажных местообитаний и мхов можно предположить, что здесь существовала сильно заозеренная лайда, где озера располагались вдоль берега моря и соединялись с ним протоками.
Отложения из интервала 4.2–9.1 м, относящиеся к едомной свите, формировались в локальных условиях пресноводного водоема, уже не имевшего связи с морем, возможно заросшего болота или озера с богатой прибрежной растительностью, с развитым моховым покровом или заболоченного луга с травянисто-злаковой растительностью. Состав СПК позволяет сделать вывод о некотором потеплении климата и распространении на фоне господства тундровых ландшафтов лиственничных редколесий.
Образование суглинков едомной свиты верхней части разреза (0.7–3.2 м) происходило в условиях тундровой равнины, на которой произрастали злаковые и злаково-разнотравные ассоциации. Обилие пресноводных диатомей в ряде проб, присутствие фитолитов, характерных для растений влажных местообитаний, находки пресноводных беспозвоночных являются показателем присутствия переувлажненных участков над головами ледяных жил, характерных для тундр.
В данной статье впервые приведены фотографические изображения микрофоссилий из плейстоценовых отложений Колымской низменности, их комплексное исследование позволило получить более полную и объективную характеристику условий формирования отложений. Так, анализ бентосных фораминифер и остракод позволяет сделать вывод о существовании морского бассейна, в той или иной мере опресненного. На основании содержания панцирей диатомовых водорослей и спикул губок можно установить характер и специфику обводнения территории. Результаты палинологического анализа позволяют выявить тенденции развития региональной растительности, а результаты фитолитного анализа – тенденции развития локальной растительности. Важным преимуществом фитолитного метода является возможность фиксировать степень увлажненности территории. На основании результатов палеоэнтомологического анализа для данного региона получено свидетельство существования тундростепного биома, что доказывает его значительное продвижение на север, а также сделано заключение о существовании пресноводного бассейна. Результаты комплексного анализа микрофоссилий частично перекрываются и дополняют друг друга, что сделало возможным получение новой дополнительной информации о генезисе исследуемых толщ и позволило более полно оценить эволюцию ландшафта, изменения растительности в целом и ее локальные особенности.
Источники финансирования. Экспедиционные исследования поддержаны National Science Foundation (grant NSF DEB-1442262). Аналитические исследования образцов проводились в рамках проекта РФФИ 19-29-05003 мк. Фитолитные исследования проводились в рамках темы государственного задания АААА-А18-118013190181-6 ИФХиБПП РАН и по проекту РФФИ № 20-05-00559 А, насекомые изучались по проекту РФФИ № 20-04-00165 А. По госбюджетной теме ГИН РАН проводилось исследование спор и пыльцы Д.А. Лопатиной и бентосных фораминифер Я.С. Овсепяном.
Список литературы
Архангелов А.А., Кузнецова Т.П., Карташова Г.Г., Коняхин М.А. Генезис и условия формирования верхнеплейстоценовых льдистых алевритов Колымской низменности (на примере Чукочьего Яра) // Проблемы криолитологии. 1979. Вып. VIII. С. 110–135.
Васильчук А.К. Палинология и хронология полигонально-жильных комплексов в криолитозоне России. М.: Изд-во МГУ, 2007. 488 с.
Воробьева Л.А. Теория и практика химического анализа почв. М.: ГЕОС, 2006. 400 с.
Гиличинский Д.А. Криобиосфера позднего кайнозоя: вечная мерзлота как среда сохранения жизнеспособных микроорганизмов. Дисс. … докт. геол.-мин. наук. 2002. 210 с.
Гудина В.И. Морской плейстоцен сибирских равнин. Фораминиферы Енисейского севера. М.: Наука, 1969. 80 с.
Евсеев В.П., Недешева Г.Н. Особенности формирования прибрежно-морских отложений Колымской низменности // Вестник МГУ. Сер. географ. 1983. № 5. С. 37–44.
Каплина Т.Н. Спорово-пыльцевые спектры осадков “ледового комплекса” приморских низменностей Якутии // Изв. АН СССР. Сер. географ. 1979. № 2. С. 85–93.
Киселев С.В. Позднекайнозойские жесткокрылые Северо-Востока Сибири. М.: Наука, 1981. 116 с.
Корсун С.А. Закономерности распределения бентосных фораминифер в Баренцевом море. Автореферат дисс. … канд. биол. наук. М.: ИО РАН, 1995. 24 с.
Корсун С.А., Погодина И.А., Тарасов Г.А., Матишов Г.Г. Фораминиферы Баренцева моря (гидробиология и четвертичная палеоэкология). Апатиты: Изд-во КНЦ РАН, 1994. 136 с.
Кузнецов Ю.В. О вещественном составе верхнеплейстоценовых и голоценовых отложений в обнажении Чукочьем // Проблемы криолитологии. 1979. Вып. VIII. С. 136–144.
Палеопалинология. Т. I. Ред. Покровская И.М. Л.: Недра, 1966. 352 с.
Пыльцевой анализ. Ред. Криштофович А.Н. М.: Госгеолиздат, 1950. 571 с.
Решения Междуведомственного стратиграфического совещания по четвертичной системе Востока СССР. Магадан: СВКНИИ ДВО АН СССР, 1987. 239 с.
Ривкина Е.М., Гиличинский Д.А. Метан как палеоиндикатор динамики мерзлых толщ // Литология и полезн. ископаемые. 1996. № 4. С. 445–448.
Ривкина Е.М., Краев Г.Н., Кривушин К.В., Лауринавичюс К.С., Федоров-Давыдов Д.Г., Холодов А.Л., Щербакова В.А., Гиличинский Д.А. Метан в вечномерзлых отложениях северо-восточного сектора Арктики // Криосфера Земли. 2006. № 3. С. 23–41.
Шмелев Д.Г. Роль криогенеза в формировании состава позднечетвертичных мерзлых отложений оазисов Антарктиды и северо-востока Якутии // Криосфера Земли. 2015. Т. XIX. № 1. С. 41–57.
Abramov A., Vishnivetskaya T., Rivkina E. Are permafrost microorganisms as old as permafrost? // FEMS Microbiol. Ecol. 2021. V. 97. № 2. P. 1–12.
Brouwers E.M., Cronin T.M., Horne D.J., Lord A.R. Recent shallow marine ostracods from high latitudes: implications for late Pliocene and Quaternary palaeoclimatology // Boreas. 2000. V. 29. P. 127–142.
Cronin T.M., Briggs W.M., Jr., Brouwers E.M., Whatley R.C., Wood A., Cotton M. Modern Arctic Podocopid database // USGS Open-File Report 91-385. 1991.
Feyling-Hanssen R.W., Jørgensen J.A., Knudsen K.L., Andersen A.L. Late Quaternary foraminifera from Vendsyssel, Denmark and Sandnes, Norway // Bull. Geol. Soc. Den. 1971. V. 21. P. 67–317.
Frenzel P., Keyser D., Viehberg F.A. An illustrated key and (paleo)ecological primer for Postglacial to Recent Ostracoda (Crustacea) of the Baltic Sea // Boreas. 2010. V. 39. P. 567–575.
Gilichinsky D., Rivkina E., Shcherbakova V., Laurinavichius K., Tiedje J. Supercooled water brines within permafrost – an unknown ecological niche for microorganisms: a model for astrobiology // Astrobiology. 2003. V. 3. № 2. P. 331–341.
Gilichinsky D., Rivkina E., Bakermans C., Shcherbakova V., Petrovskaya L., Ozerskaya S., Ivanushkina N., Kochkina G., Laurinavichius K., Pecheritsina S., Fattakhova R., Tiedje J. Biodiversity of cryopegs in permafrost // FEMS Microbiol. Ecol. 2005. V. 53. P. 117–128.
Grøsfjeld K., Funder S., Seidenkrantz M.-S., Glaister C. Last Interglacial marine environments in the White Sea region, northwestern Russia // Boreas. 2006. V. 35. P. 493–520.
Kiselev S.V., Nazarov V.I. Late Cenozoic insects of Northern Eurasia // Paleontol. J. Suppl. 2009. V. 43. № 7. P. 1–128.
Korsun S., Hald M., Golikova E., Yudina A., Kuznetsov I., Mikhailov D., Knyazeva O. Intertidal foraminiferal fauna and the distribution of Elphidiidae at Chupa Inlet, western White Sea // Marine Biology Res. 2014. V. 10. № 2. P. 153–166.
Kuzmina S.A. Quaternary insects and environment of the Northeastern Asia // Paleontol. J. Suppl. 2015. V. 49. № 7. P. 1–189.
Murray J. Ecology and application of benthic foraminifera. N.Y.: Cambridge Univ. Press, 2006. 426 p.
Polyak L., Korsun S., Febo L., Stanovoy V., Khusid T., Hald M., Paulsen B.E., Lubinski D.A. Benthic foraminiferal assemblages from the southern Kara Sea, a river-influenced Arctic marine environment // J. Foraminiferal Res. 2002. V. 32. № 3. P. 252–273.
Stepanova A., Taldenkova E., Bauch H.A. Recent Ostracoda of the Laptev Sea (Arctic Siberia): taxonomic composition and some environmental implications // Marine Micropalaeontol. 2003. V. 48(1–2). P. 23–48.
Stepanova A., Taldenkova E., Simstich J., Bauch H.A. Comparison study of the modern ostracod associations in the Kara and Laptev seas: ecological aspects // Marine Micropaleontol. 2007. V. 63(3–4). P. 111–142.
Stepanova A., Obrochta S., Quintana Krupinski N.B., Hyttinen O., Kotilainen A., Andrén T. Late Weichselian to Holocene history of the Baltic Sea as reflected in ostracod assemblages // Boreas. 2019. V. 48(3). P. 761–778.
Wetterich S., Schirrmeister L., Pietrzeniuk E. Freshwater ostracods in Quaternary deposits in the Siberian Arctic // J. Paleolimnol. 2005. V. 34. P. 363–376.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция