

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 1, стр. 55-68
Представители миаохенской биоты из дошурамских отложений эдиакария (венда) Патомского нагорья Сибири
П. Ю. Петров 1, *, Н. Г. Воробьева 1
1 Геологический институт РАН
Москва, Россия
* E-mail: petrov-geo-home@rambler.ru
Поступила в редакцию 27.04.2021
После доработки 09.07.2021
Принята к публикации 08.09.2021
- EDN: XGKBLJ
- DOI: 10.31857/S0869592X22010069
Аннотация
В нижневендских (среднеэдиакарских, около 580 млн лет) отложениях уринской свиты Патомского бассейна Сибири впервые обнаружены углефицированные макрофоссилии, относящиеся к многоклеточным водорослям и ранее известные в составе более молодой (<570 млн лет) миаохенской биоты. Уринские макрофоссилии представлены фитолеймами неветвящихся лентовидных слоевищ, сохраненных на поверхностях алевритистых аргиллитов. Среди диагностируемых таксонов определены виды Liulingjitaenia alloplecta и Jiuqunaoella simplicis. Рассмотрены особенности их строения. Проведены реконструкции прижизненных форм организмов и их колоний. Показано, что эти органические макрофоссилии являлись заметным детритообразующим компонентом Уринского бассейна. Уринские макрофиты сосуществовали с раннеэдиакарской микробиотой, составляя вместе с ней единую экосистему палеобассейна, которая была образована незадолго до Шурамского события и была существенно преобразована после него.
ВВЕДЕНИЕ
Самая крупная в истории Земли отрицательная аномалия δ13С Шурам (EN3), приуроченная приблизительно к середине эдиакарского периода, известна из осадочных последовательностей многих палеоконтинентов того времени (Grotzinger et al., 2011). Несмотря на противоречивые оценки ее точного возраста и длительности (Gong et al., 2017; Witkosky, Wernicke, 2018; Xiao, Narbonne, 2020), наиболее вероятно, что это событие произошло в интервале от 571 до 562 млн лет и длилось 8–9 млн лет (Minguez et al., 2015; Gong et al., 2017; Canfield et al., 2020). В палеонтологической летописи эдиакария аномалия Шурам, а точнее ее основание, представляет главный палеоэкологический рубеж, разделяющий закат эпохи господства раннеэдиакарских одноклеточных эукариот и появление первых многоклеточных животных позднего эдиакария. Предполагается, что наблюдаемые эволюционные новации и экосистемные перестройки на этом рубеже были связаны с процессом оксигенизации океана того времени (Canfield et al., 2007; Narbonne, 2010; Sahoo et al., 2016; Zhang et al., 2019; Fan et al., 2020).
Очевидно, что событие Шурам явилось переломным моментом в эволюции и других групп организмов, остатки которых обычно сохранены на поверхностях пород в виде макроскопических углефицированных компрессий – фитолейм (Burgess Shale-type preservation) и отпечатков, большая часть которых ассоциируется с макроводорослями. Остатки таких организмов известны как миаохенский тип ископаемых макробиот, наиболее полно представленный в постшурамских отложениях эдиакария Южного Китая (Xiao et al., 2002; Ye et al., 2017). Таксономически обедненные ассоциации этих макробиот также известны и за пределами Китая: в Западной Монголии (Dornbos et al., 2016), США (Rowland, Rodriguez, 2014), Австралии (Xiao et al., 2020), а также в Сибири (Grazhdankin et al., 2008) и на Урале (Гражданкин и др., 2007; Марусин и др., 2011). Как и эдиакарские “мягкотелые”, характерные представители миаохенской биоты приурочены к постшурамским отложениям, в которых отсутствуют ассоциации раннеэдиакарских акантоморф. В этой статье мы сообщаем о новом местонахождении некоторых таксонов миаохенской биоты в дошурамских отложениях уринской свиты Сибири, где эти макрофоссилии находятся в тесной ассоциации с широко известной уринской акантоморфной биотой доушаньтуо-пертататакского типа (Sergeev et al., 2011; Moczydłowska, Nagovitsin, 2012; Воробьева, Сергеев, 2018).
ПАТОМСКИЙ БАССЕЙН И ВОЗРАСТ ЕГО ОТЛОЖЕНИЙ
Патомский бассейн, обрамляющий южную окраину Сибирского кратона (рис. 1а), был сформирован после распада суперконтинента Родиния в конце неопротерозоя (Sovetov, 2002; Метелкин и др., 2012; Powerman et al., 2015). Нижняя часть терригенно-карбонатных отложений этого бассейна выделена в мощный (более 6 км) патомский комплекс, который включает снизу вверх: баллаганахскую, дальнетайгинскую и жуинскую серии (Чумаков и др., 2007). В основании дальнетайгинской серии, без признаков длительного стратиграфического перерыва, залегает большепатомский гляциогоризонт, представленный мощной (около 1 км) толщей диамиктитов, которые традиционно сопоставляются с оледенением Марино (635 млн лет) (Чумаков и др., 2013; Чумаков, 2015). Однако возраст большепатомских диамиктитов, как и всей дальнетайгинской серии, в настоящее время остается дискуссионным (Рудько и др., 2020). Данные бассейнового анализа (Петров, 2018а, 2018б) в сумме с палеонтологическими данными (Леонов, Рудько, 2012; Воробьева, Петров, 2020) и отчасти с данными δ13С (Рудько и др., 2017; Петров, Покровский, 2020) предполагают более молодой (<580 млн лет) возраст дальнетайгинских отложений и вместе с тем вендский (эдиакарский) возраст всего патомского комплекса. Недавно полученные Pb–Pb изохронные датировки по высокостронциевым известнякам дальнетайгинской серии, равные 581 ± 16 и 575 ± 20 млн лет (баракунская и каланчевская свиты соответственно), не противоречат такому предположению (Rud’ko et al., 2021). В этой интерпретации мощные толщи баллаганахской и дальнетайгинской серий составляют единый терригенно-карбонатный трансгрессивно-регрессивный цикл, последовательность которого во время наиболее интенсивного погружения ложа бассейна была нарушена эвстатическим падением уровня моря, связанным с большепатомским ледниковым эпизодом. Во время ледникового максимума эрозия затронула лишь внешние области Патомского бассейна, тогда как во внутренней глубоководной области осадконакопление, очевидно, оставалось непрерывным.
Рис. 1.
Регион изучения (а) и местоположение анализируемых ископаемых организмов в вендских (эдиакарских) отложениях Уринского поднятия (б). Изотопно-геохимические данные приведены по (Pelechaty, 1998; Покровский, Буякайте, 2015; Рудько и др., 2017; Петров, Покровский, 2020), стратиграфия по (Чумаков и др., 2013; Петров, 2018а, 2018б), палеонтологические данные по (Sergeev et al., 2011; Moczydłowska, Nagovitsin, 2012; Леонов, Рудько, 2012; Воробьева, Петров, 2020; Хоментовский и др., 2004; Кочнев, Карлова, 2010). Принятые сокращения: серии: Bl – баллаганахская, Dt – дальнетайгинская, Zu – жуинская, Tr – трехверстная; свиты: bp – большепатомская, br – баракунская, ur – уринская, kl – каланчевская, vl – валюхтинская, nk – никольская, cn – ченченская, zr – жербинская, tn – тинновская, nh – нохтуйская; Є – кембрий, О – ордовик. EN3 – отрицательная аномалия δ13С Шурам. Anabarites trisulcatus и Nochoroicyathus sunnaginicus (Aldanocyathus sunnaginicus): зоны немакит-далдынского яруса венда и нижнего кембрия (по Российской и Международной стратиграфическим шкалам соответственно).
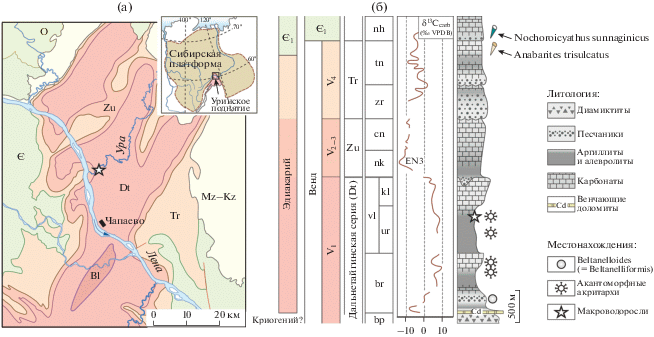
Постгляциальные отложения дальнетайгинской серии на Уринском поднятии представлены последовательностью баракунской, уринской и каланчевской свит (Чумаков и др., 2013). В этой последовательности мощностью более 2 км выделяются два терригенно-карбонатных цикла второго порядка: нижний баракунский и верхний уринско-каланчевский (Петров, 2018б). В более мористой части бассейна последний замещается относительно однородной и существенно более глинистой толщей валюхтинской свиты (рис. 1б). В песчаниках нижней части баракунской свиты присутствуют отпечатки вендских ископаемых Beltanelloides sorichevae Sokolov (Леонов, Рудько, 2012), по современной систематике относимые к Beltanelliformis brunsae Menner (Ivantsov et al., 2014). В составе акантомофных биот уринской и баракунской свит содержатся таксоны, характерные для верхних двух зональных комплексов Австралии и Китая (см. Воробьева, Петров, 2020).
Дальнетайгинская серия с размывом перекрывается глинисто-карбонатной толщей жуинской серии, в последовательности которой наблюдается значительный по амплитуде (до –11.9‰) и продолжительности (более 800 м разреза) отрицательный экскурс δ13С (EN3, рис. 1б), соответствующий глобальной аномалии Шурам (Melezhik et al., 2005, 2009; Покровский и др., 2006а, 2006б; Покровский, Буякайте, 2015), которая, по последним данным, началась сразу после 571 млн лет (Canfield et al., 2020) или 574 ± 4.7 млн лет (Rooney et al., 2020). Эта аномалия также прослеживается во внутренних районах Сибирской платформы в верхней части непского горизонта (Кочнев и др., 2018). Несогласие в основании жуинской серии является главной секвентной границей Патомского бассейна (Петров, 2018б) и представляет собой эрозионную поверхность без признаков углового несогласия, но с хорошо выраженными эрозионными врезами амплитудой до 100 м. Эрозионный рельеф в самом начале жуинского времени был нивелирован песчаными осадками, которые сейчас представляют локально распространенную базальную пачку никольской свиты (Чумаков и др., 2013). Эта пачка мощностью до 80 м ранее некоторыми исследователями выделялась в куллекинскую свиту (Бобров, 1964, 1979; Колосов, 1975). Очевидно, эрозия и последующее заполнение рельефа осадками явилось следствием кратковременного, но значительного по амплитуде падения и подъема уровня моря. Такие флуктуации могли быть вызваны тектонической перестройкой бассейна (цикличность 3-го порядка, длительностью 0.5–3.0 млн лет; Cloetingh et al., 1985) или имели гляциоэвстатическую природу, сходную по амплитуде и продолжительности с последним ледниковым событием позднего плейстоцена (Lambeck et al., 2014). В кровле жуинская серия ограничена подобной эрозионной поверхностью и перекрыта терригенно-карбонатными отложениями трехверстной серии в составе жербинской и тинновской свит (рис. 1б) (Чумаков и др., 2013). Глинисто-карбонатные породы верхней части тинновской свиты содержат мелкораковинную фауну зоны Anabarites trisulcatus немакит-далдынского яруса верхнего венда (Хоментовский и др., 2004), а в вышележащей нохтуйской свите появляется многочисленная фауна зоны Nochoroicyathus sunnaginicus томмотского яруса нижнего кембрия (Хоментовский и др., 2004; Кочнев, Карлова, 2010). Таким образом, в осадочной последовательности всего патомского комплекса, по крайней мере во внутренних частях бассейна, не наблюдается каких-либо стратиграфически значимых перерывов, а возраст палеонтологически охарактеризованных отложений дальнетайгинской серии ограничен интервалом 580–570 млн лет.
МАТЕРИАЛ И МЕТОДЫ
Основной палеонтологический материал получен из верхней части уринской свиты, вскрытой на правом берегу р. Ура в 5.5 км выше устья (GPS: 60.26753° с.ш., 117.10835° в.д.) (рис. 1б). Небольшая часть образцов была отобрана из средней части разреза свиты, расположенного в верхнем течении р. Ура, выше устья р. Улахан-Илигир (GPS: 60.40280° с.ш., 117.35112° в.д.). Особенностью уринских алевро-аргиллитов является повсеместно выраженная концентрическая и неупорядоченная линейная отдельность, ориентированная под разными углами к осадочным поверхностям, что создавало серьезные трудности при препарировании макрофоссилий. Так, во многих случаях экземпляры макрофоссилий были потеряны в результате самопроизвольного (без механического воздействия) разрушения образца при его повторном смачивании. С целью предотвращения разрушения образцов, особенно при изготовлении шлифов, использовалась горячая пропитка эпоксидной смолой.
Препарирование образцов и изучение макрофоссилий проводили под бинокулярным микроскопом МБС-9. Макросъемку фоссилий осуществляли при обработке поверхностей пород глицерином. Изучение микроструктур и микросъемку выполняли в проходящем и отраженном свете, а также при УФ-флюоресценции с помощью микроскопа Zeiss REM 5 и микроскопа Zeiss Axio Imager A1, оснащенного цифровой камерой Axio-CamMRc-5, Carl Zeiss, Германия. Метод флюоресценции был использован при изучении органических (высокоуглеродистых) микроостатков в шлифах, изготовленных с применением канадского бальзама. При возбуждении ультрафиолетовым излучением канадский бальзам и кварц-алюмосиликатный матрикс алевритистых аргиллитов флюоресцируют равномерным свечением, благодаря чему на их фоне становятся видны нефлюоресцирующие органические остатки и их тонкая микроструктура, не различимая в проходящем свете. Палеонтологический материал хранится в Геологическом институте РАН (г. Москва), коллекция № 14801.
ФАЦИАЛЬНАЯ ХАРАКТЕРИСТИКА ОТЛОЖЕНИЙ И ОРГАНИЧЕСКИЕ МАКРОФОССИЛИИ УРИНСКОЙ СВИТЫ
Обстановки осадконакопления Уринского бассейна. Существенно алеврит-глинистые осадки уринской свиты накапливались в глубоководных (ниже базиса штормовых волн) обстановках на пологом (около 1°) гомоклинальном рампе с повсеместным развитием поверхностных (не более первых десятков сантиметров мощности) оползней и грязевых потоков, инициированных сейсмическим воздействием (Петров, 2018б). В разрезе количество деформированных осадочных слоев нередко достигает 90%. Предполагается, что такой необычный для подобных очень пологих глинистых рампов потоково-оползневой характер седиментации был связан с интенсивными процессами метаногенеза в донных слоях. Такой вывод подтверждается С-изотопными данными для раннедиагенетического аутигенного карбоната (Петров, Покровский, 2020). Значительная часть тонкозернистой силикокластики в уринской свите, очевидно, имеет эоловое происхождение, что согласуется с характерным гранулометрическим спектром, а также с текстурными и минералогическими особенностями уринских осадков (Петров, 2018б). Заметное количество эолового материала, в свою очередь, предполагает аридизацию климата того времени. Небольшая доля аллотигенного карбоната в алевритистых аргиллитах уринской свиты была связана с волновой эрозией карбонатной платформы во внешней части бассейна, которая в каланчевское время распространилась по всей территории Уринского поднятия. К проградационному фронту этой платформы приурочен наиболее богатый тафоценоз уринских акантоморф, хотя обедненные ассоциации этой микробиоты были встречены на различных уровнях разреза в дистальных бассейновых фациях (Воробьева, Петров, 2020).
Органический детрит в уринских отложениях. Помимо микрофоссилий, в уринских аргиллитах широко распространены органические макроостатки в виде мелкоразмерного (1–10 мм) детрита и протяженных (до 10 см), иногда структурированных фрагментов (пленок) (рис. 2а, 2б). Такие фрагменты сохраняли целостность при переносе внутри оползневых и даже потоковых слоев (Петров, 2018б). Подобные в различной степени деформированные, но сохранявшие до определенной степени гибкость и пластичность пленки известны из отложений различного возраста и обычно интерпретируются как остатки бентосных микробных матов (Schieber, 1999, 2007; Simonson, Carney, 1999). Однако в отличие от бесформенных и обычно бесструктурных фрагментов микробных матов, некоторые уринские фрагменты имеют определенные повторяющиеся формы с четкими границами и в различной степени выраженное внутреннее строение, отличное от микробиальных структур. Среди мелкоразмерного детрита распространены пленки овальной или конусовидной формы, а среди крупных фрагментов – протяженные ленты с ровными краями. Внутренние структуры некоторых фрагментов, наблюдаемые в шлифах, также заметно отличаются от типичных сгустковых или гребенчато-слоистых микробиальных структур. Наблюдаемые структуры состоят из прямых четко обособленных, параллельно ориентированных пленок либо переплетающихся нитей или волокон микронной размерности (рис. 2в–2д). В косом сечении некоторых наименее деградировавших лентовидных фрагментов наблюдаются хорошо выдержанные волокнистоподобные структуры (рис. 2е). Структуры сильно деградировавших фрагментов состоят из серий сближенных пленок, схожих в поперечном сечении со структурами микробных матов, однако в отличие от них они имеют выдержанную толщину и более четкие внешние границы. Очевидно, бóльшая часть захороненного детрита подверглась значительной анаэробной бактериальной деструкции с образованием сингенетичного фрамбоидального пирита, количество которого в целом пропорционально степени деградации наблюдаемых биогенных структур (рис. 2ж, 2з).
Рис. 2.
Форма и структура крупноразмерного органического детрита в алевро-аргиллитах уринской свиты. а, б – сечения крупных обрывков пленок на поверхности оползневых слоев; в – поперечное сечение оползневого слоя (нижняя часть фотографии) и грязевого потока (верхняя часть); г, д – структуры детрита, захороненного внутри оползневого слоя; е – прерывисто-штриховатая волокнистоподобная структура лентовидного фрагмента Liulingjitaenia alloplecta в косом продольном сечении; ж, з – деградировавшие фрагменты детрита с выделениями сингенетичного фрамбоидального пирита. Шлифы в проходящем свете (в, г) и флуоресцентном свечении при возбуждении ультрафиолетовым излучением (е–з).

Таким образом, заметная часть органического детрита в уринской свите принадлежала неким организмам или колониальным образованиям, которые по составу и своей организации отличались от матформирующих микробных сообществ. Остатки таких организмов являлись детритобразующим компонентом в тонкозернистых уринских осадках.
Макрофоссилии уринской свиты. В верхней части свиты, в слоях с хорошо известной уринской акантоморфной микробиотой (Sergeev et al., 2011; Moczydłowska, Nagovitsin, 2012) были обнаружены макроскопические представители миаохенской биоты (табл. I, II ). В тафоценозе доминируют закрученные по спирали лентовидные образования Liulingjitaenia alloplecta Chen et Xiao. Прямые, узкие (обычно 2–3 мм) и строго выдержанные по ширине ленты не имеют ни начала, ни конца. В типовом местонахождении – миаохенской биоте Китая – максимальная протяженность таких лент достигает 15–20 см (Xiao et al., 2002). Максимальная длина лент в нашем материале составляет около 8 см. Обычно наблюдаются многочисленные скопления из ориентированных в одном направлении экземпляров (табл. I , фиг. 1–6 ). Морфологическое строение анализируемых макрофоссилий дискуссионно. По мнению одних исследователей, эти формы представляли собой плотный пучок нитей, закрученный по спирали (Chen, Xiao, 1992; Ye et al., 2017; Xiao et al., 2020). Наглядным примером внешнего вида таких форм является обычная толстая веревка. В иной интерпретации они имели форму полой многократно закрученной трубки с множеством спиралевидных складок смятия, которые лишь имитировали нитчатое строение стенки трубки (Steiner, 1994; Xiao et al., 2002). Однако из анализа уринских фитолейм, сохраненных одновременно на подошве и кровле одного и того же слоя (табл. I , фиг. 1), следует, что эти организмы имели скорее форму свернутой в объемную спираль ленты, чем пучка спирально закрученных нитей или спирально закрученной цилиндрической трубки. В максимально закрученных экземплярах витки спирали могли смыкаться, образуя подобие закрученного пучка нитей, а стыки витков могли имитировать полую сильно закрученную трубку. Вместе с тем в пределах одного и того же экземпляра нередко наблюдается изменение направления завивания ленты. Различные участки одной и той же ленты были завернуты в разные стороны (рис. 3а, 3б). Кроме того, отдельные части спирали могли быть развернуты в плоскую синусоидально изгибающуюся “меандрирующую” ленту (табл. I , фиг. 2, верхняя часть фотографии). Среди скоплений уринских Liulingjitaenia можно наблюдать различные формы этих организмов в зависимости от характера и степени закручивания лент. Однако в изученном материале заметно преобладают ленты, закрученные преимущественно в одном направлении, либо вправо, либо влево. Ленты Liulingjitaenia alloplecta имеют неясно выраженную волокнистую микроструктуру, на фоне которой выделяются четко обособленные нити различной толщины от 10 до 50 мкм (табл. I , фиг. 5, 7, 8 ). Некоторые нити образованы цепочками шаровидных агрегатов, представляющих вторичные выделения фрамбоидального пирита (табл. I , фиг. 9 ). Все нити изгибаются по спирали, подчеркивая каркас всей спиралевидной конструкции.
Рис. 3.
Реконструкция фрагмента колонии Liulingjitaenia alloplecta (а) и фрагмент отпечатка ленты, иллюстрирующий изменение направления завивания (б). Прямыми стрелками показаны участки, на которых происходило изменение направления завивания ленты (с правого на левое, с левого на правое).

Таблица I . Liulingjitaenia alloplecta M. Chen et Xiao, 1992. 1, 2, 8 – экз. ГИН № 14801-20024-1: 2, 8 – увеличенные фрагменты; 3–5, 7, 9 – экз. ГИН № 14801-20009-1: 4, 5, 7, 9 – увеличенные фрагменты; 6 – экз. ГИН № 14801-20017-1. Все экземпляры происходят из уринской свиты.

Таблица II. Макрофоссилии уринской свиты. 1–3 – Gen. et sp. indet. 1: 1, 2 – экз. ГИН № 14801-20012-1; 3 –экз. ГИН № 14801-20012-2; 4, 5 – Gen. et sp. indet. 2, экз. ГИН № 14801-20018-1; 6 – Jiuqunaoella simplicis Chen, 1991: экз. ГИН № 14801-20014-1; 7 – углефицированная пленка конусовидной формы, экз. ГИН № 14801-17140-1; 8–10 – Tawuia dalensis Hofmann, 1979: 8 – экз. ГИН № 14801-17148-1; 9 – экз. ГИН № 14801-17148-2; 10 – экз. ГИН № 14801-20019-1. Все экземпляры происходят из уринской свиты.

Предложенная нами интерпретация следует из наблюдения упомянутых нитевидных структур в парных отпечатках. При механическом разделении слоя компрессия ископаемого организма может быть разделена на две половины – нижнюю и верхнюю. В нашем случае нитевидные структуры в парных отпечатках оказываются зеркально симметричными, а значит, на подошве и кровле слоя наблюдаются разделенные слои нитей и нижней, и верхней частей лентовидной спирали. Поперечные или косые сечения лент, состоящих из нескольких слоев нитей, также наблюдаются в шлифах (рис. 2е). Косвенным подтверждением такой изначальной формы Liulingjitaenia alloplecta является хорошо выдержанная линейность фитолейм на всем их протяжении. Лента, закрученная в спираль, как и трубка, является наиболее устойчивой к изгибам по всем направлениям. Однако в отличие от трубки такая лента является гибкой, причем не только на изгиб, но и на растяжение. После снятия нагрузки лента без деформаций возвращалась в исходное состояние. Вероятно, такое строение способствовало усилению конструкции узкого и тонкого, но очень протяженного тела, что могло быть востребовано в условиях турбулентной среды.
Многочисленные одномоментно захороненные скопления L. alloplecta свидетельствуют о колониальном образе жизни этого вида. В скоплениях-колониях отдельные индивидуумы росли в одном направлении и пересекались друг с другом под очень небольшим (менее 5°) углом, образуя рыхлые сплетения (рис. 3). Края соприкасающихся спиралей фиксировали конструкцию колонии. По этой причине при переносе они не распадались, а перенесенные и захороненные их фрагменты представляли прижизненную организацию таких колоний. В ископаемом материале не выявлены органы прикрепления этих организмов, что может указывать на свободно плавающий или свободно стелящийся по дну стиль их обитания. Отсутствие определимых остатков Liulingjitaenia в глубоководных дистальных частях бассейна могло быть связано как с прибрежными (сублиторальными) обстановками обитания, так и с различными условиями их захоронения. Приуроченность микро- и макроскопических ископаемых организмов к одним и тем же узким фациальным зонам Уринского бассейна указывает на приуроченность тех и других к одной и той же биофации и в то же время предполагает ведущую роль тафономии в фациальном распределении тех и других ископаемых сообществ (Воробьева, Петров, 2020).
Другим представителем миаохенской биоты является Jiuqunaoella simplicis Chen (табл. II, фиг. 6 ). Эти червеподобные организмы были встречены только на одной поверхности, хотя их предполагаемые небольшие (первые миллиметры) фрагменты иногда присутствуют среди обильного углистого детрита в различных слоях разреза вместе с богатыми ассоциациями уринских акантоморфных акритарх. Обнаруженные экземпляры представляют собой компрессии трубчатых организмов длиной менее 1 см с замкнутыми, обычно завернутыми окончаниями и редкими поперечными складками вероятного смятия.
Помимо Liulingjitaenia и Jiuqunaoella, в уринских отложениях были обнаружены разнообразные “сегментированные” лентовидные отпечатки, описанные нами как Gen. et sp. indet. 1 (табл. II, фиг. 1–3) и Gen. et sp. indet. 2 (табл. II, фиг. 4, 5 ). По форме и размерным характеристикам их нельзя отнести к каким-либо известным таксонам, а количество и качество материала не позволяют описывать новые таксоны. Представители Gen. et sp. indet. 1 имеют некоторое отдаленное сходство с Calyptrina Sokolov (Соколов, 1965; Xiao et al., 2002; Ye et al., 2017), а Gen. et sp. indet. 2 с более древними протерозойскими ископаемыми Protoarenicola Wang (Wang, 1982). И, наконец, в уринских отложениях, среди обильного органического детрита на поверхностях слоев, широко распространены углистые пленки правильных овальных очертаний, отнесенные к Tawuia dalensis Hofmann (табл. II, фиг. 8–10 ), а также компрессии конусовидной формы (табл. II, фиг. 7 ). Эти фоссилии, в отличие от описанных выше, встречаются по всему разрезу уринской свиты.
Морфологическое подобие и предполагаемая биологическая природа Liulingjitaenia и Jiuqunaoella. Отсутствие клеточных микроструктур и данных по биомаркерам позволяют интерпретировать биологическое родство этих макроскопических ископаемых организмов лишь на основе общей сравнительной морфологии. Впервые описавшие род Liulingjitaenia M. Chen и Z. Xiao (1992) сопоставили анализированные формы с цианобактериями. Этой точки зрения также придерживался M. Steiner (1994). Он относил эти фоссилии к очень большим колониям цианобактерий. Позднее, S. Xiao с соавторами (2002) предположили, что вероятными морфологическими аналогами Liulingjitaenia являются сифоновые зеленые водоросли. В настоящее время род Liulingjitaenia относят к макрофитам (Xiao et al., 2020), хотя некоторые авторы не исключают их цианобактериальной природы (Ye et al., 2017). Действительно, колонии цианобактерий способны к самоорганизации, иногда продуцируя упорядоченные макроструктуры (Sim et al., 2012; Сумина, Сумин, 2013). Однако трихомы цианобактерий, особенно когда они находятся в скоплениях, за редкими исключениями, окружены плотным общим чехлом из полисахаридной слизи. Такой чехол выполняет защитную функцию как для клеток отдельно взятого трихома, так и для всей колонии, при этом его сохранность в ископаемом состоянии заметно выше, чем у самих клеток. У Liulingjitaenia каких-либо признаков существования такого чехла нет; наблюдаются лишь остатки параллельно ориентированных нитей среди небольшого количества диффузного органического вещества между ними. Сказанное не позволяет рассматривать эти ископаемые как вероятные остатки аномально крупных колоний нитчатых цианобактерий, которые не продуцировали внеклеточные полимеры и вместе с тем имели нехарактерный для цианобактерий весьма сложный уровень организации.
Liulingjitaenia alloplecta имеет морфологическое сходство с некоторыми позднеэдиакарскими и раннепалеозойскими формами, в частности с поздневендским–раннекембрийским таксоном Harlaniella Sokolov. А. Иванцов (2013) указывает на морфологическое сходство между Liulingjitaenia alloplecta и выделенным им видом Harlaniella ingriana Ivantsov, предполагая, что L. alloplecta представляет собой лишь тафономическую разновидность не заполненной осадком трубки H. ingriana.
Наиболее близким по морфологии и размерам к Liulingjitaenia является кембрийский монотиповой род Fuxianospira Chen et Zhou. Хотя этот таксон описывается как единое спирально-закрученное тело, у нескольких продемонстрированных экземпляров Fuxianospira gyrata видны фрагменты “закрученных” нитевидных структур (например, Wang et al., 2021, figs. 4-C, 5-A (S1)). Такие структуры аналогичны L. alloplecta. Первоначально Fuxianospira была описана как макроводоросль (Chen, Zhou, 1997), а позже была интерпретирована как возможный копролит (Steiner et al., 2005). Последующий детальный анализ этих фоссилий не выявил каких-либо характерных признаков копролита и позволил отнести Fuxianospira, как и Liulingjitaenia, к сифоновым водорослям (LoDuca et al., 2015; Wang et al., 2021). Цитированные авторы сравнивают Fuxianospira с кладофоровой зеленой водорослью Chaetomorpha, “волокнисто-скрученная” структура которой образована рядами спирально расположенных микрофибрилл внутри целлюлозной клеточной стенки.
Макрофоссилии другого эдиакарского рода Jiuqunaoella изначально были интерпретированы как остатки червеподобных животных (Chen, Xiao, 1991; Ding et al., 1996), а позже были также отнесены к макрофитам (Xiao et al., 2002). Слабо выраженная поперечная фрагментация на отпечатках Jiuqunaoella представляет собой складки сжатия цилиндрического тела. Эти макрофоссилии были интерпретированы как остатки трубкообразного организма, по морфологии близкого к некоторым видам ценоцитных зеленых водорослей (Xiao et al., 2002). Таким образом, идентифицированные углеродистые компрессии уринской свиты имеют макроводорослевую природу. Эти фоссилии также имеют своих морфологических аналогов и в вышележащих постшурамских и нижнекембрийских слоях.
ПАЛЕОНТОЛОГИЧЕСКОЕ ОПИСАНИЕ
Род Jiuqunaoella Chen in M. Chen et Xiao, 1991 emend. Xiao, Yuan, Steiner et Knoll, 2002
Jiuqunaoella simplicis Chen in M. Chen et Xiao, 1991 emend. Xiao, Yuan, Steiner et Knoll, 2002
Табл. II, фиг. 6
Jiuqunaoella simplicis Chen: Chen, Xiao, 1991, p. 320, pl. 5, fig. 6 , pl. 6, figs. 1, 2; Chen et al., 1994, p. 396–397, pl. 4, figs. 7–9 ; Ding et al., 1996, p. 112, pl. 32, fig. 5 ; pl. 33, figs. 1, 4, 10 ; pl. 34, fig. 11 ; Xiao et al., 2002, p. 363, figs. 7.2–7.4, 8.1 ; Yuan et al., 2002, p. 61, fig. 69 ; Гражданкин и др., 2007, с. 75, фиг. 2ж, 2з; Ye et al., 2017, p. 31, figs. 19A–19E ; Bykova et al., 2020, p. 3, fig. 1A.
Vendotaenia sp.: Yuan et al., 1995, p. 98, pl. 2, fig. 9 .
Jiuqunaoella convoluta Li: Ding et al., 1996, p. 112, pl. 33, figs. 2, 3.
Jiuqunaoella minia Li: Ding et al., 1996, p. 112–113, pl. 32, fig. 6 .
Jiuqunaoella sp.: Ding et al., 1996, pl. 34, fig. 6 .
Описание. Углефицированные лентовидные структуры, сохранившиеся на поверхности напластования пород. Ленты длинные, гладкие, изогнутые, замкнутые с одного конца. Окончания вытянуто-закругленные, с редкими поперечными складками смятия. Наблюдаемая длина лент до 35 мм. Ширина лент постоянная по всей длине (1.1–1.2 мм), сужается лишь к окончаниям до 0.5–0.7 мм.
Распространение. Китай, верхняя часть формации Доушаньтуо, пачка Миаохе, эдиакарий. Россия: Восточно-Европейская платформа, лямицкая свита, верхний венд; Сибирская платформа, хатыспытская свита, верхний венд; Байкало-Патомское нагорье, уринская свита, венд.
Материал. Два неполных экземпляра.
Род Liulingjitaenia M. Chen et Xiao, 1992
Liulingjitaenia alloplecta M. Chen et Xiao, 1992
Табл. I , фиг. 1–9
Liulingjitaenia alloplecta Chen et Xiao: 1992, p. 515, pl. 1, figs. 1–4 ; Гражданкин и др., 2007, с. 76, фиг. 3е; Wang еt al., 2007, p. 833, pl. 2, figs. 2–5 ; Марусин и др., 2011, с. 661, фиг. 2б; Wang еt al., 2014, p. 971, figs. 4g, 4h ; Ye еt al., 2017, p. 35, figs. 21A–21H .
Eoscytosiphon longitubulosum Ding: Ding et al., 1996, p. 88, pl. 22, figs. 3, 6 .
Liulingjitaenia alloplecta Chen et Xiao, emend. Steiner: Xiao еt al., 2002, p. 362, figs. 6.7–6.9, 6.12–6.13 (и синонимика в этой роботе); Yuan еt al., 2002, p. 62, figs. 71, 72 .
Problematicum type B: Hofmann, Mountjoy, 2010, p. 1315, fig. 7E .
Liulingjitaenia alloplecta: An еt al., 2015, p. 248, fig. 4h .
Описание. Длинные узкие ленты, сохранившиеся на поверхности напластования в виде углеродистых компрессий. Ленты прямые или слегка изогнутые, скрученные в косо-поперечном направлении. Скручивание ленты плотное, до 8 витков на 1 см длины. Ширина лент от 1 до 3.5 мм, выдержанная по всей длине. Окончание лент не наблюдается. Длина фрагментов до 8 см. Ленты состоят из продольно ориентированных тонких (10–30 мкм) нитей (табл. I , фиг. 5, 7–9 ).
Замечания. В первоначальном диагнозе M. Chen и Z. Xiao (1992) описывали Liulingjitaenia alloplecta как длинное слоевище, имеющее спиралевидную форму и состоящее из продольных нитей. Позже диагноз был исправлен, и L. alloplecta описывалась как узкие ленточные отпечатки изначально трубчатых, мембранных организмов или колоний организмов со спиральными складками, образованными скручиванием (Steiner, 1994; Xiao et al., 2002). Впоследствии большинство исследователей вернулись к оригинальному диагнозу M. Chen и Z. Xiao (1992) (Hofmann, Mountjoy, 2010; Xiao et al., 2013, 2020; Ye et al., 2017). Наш материал показывает, что организм представлял собой не закрученный цилиндр, а ленту, завернутую в объемную спираль. Лента образована тонкими нитями-волокнами, ориентированными вдоль линии роста. Такое строение тела наиболее близко к первоначальному диагнозу Liulingjitaenia (Chen, Xiao, 1992), в котором пучки нитей, а не складки трубки создают характерную текстуру фитолейм.
Распространение. Южный Китай: провинция Хубэй, верхняя часть формации Доушаньтуо, пачка Миаохе, эдиакарий; провинция Гуйчжоу, верхняя часть формации Доушаньтуо, биота Венхуэй, эдиакарий. Канада, Британская Колумбия, верхняя часть группы Миетте, эдиакарий. Россия: Средний Урал, перевалокская свита, верхний венд; Сибирская платформа, хатыспытская свита, верхний венд; Байкало-Патомское нагорье, уринская свита, венд.
Материал. Более 30 экземпляров.
Gen. et sp. indet. 1
Табл. II, фиг. 1–3
Описание. Фрагменты изогнутых лент, сохранившихся в виде рельефных отпечатков узких выпуклых гребней, образованных на месте глубоких складок тела организма, и органического вещества между ними. Боковая кромка ленты извилистая. Длина неполных фрагментов до 30 мм. Ленты выдержаны по ширине от 2 до 2.5 мм и лишь у окончаний сужаются до 1 мм. Гребни ланцетовидной формы, шириной в средней части 0.7–1 мм. Они не всегда пересекают ленту по всей ее ширине и часто достигают лишь трети или половины ширины отпечатка. Относительно линейно вытянутого организма гребни расположены под углом 90°. Интервалы между гребнями 0.4–0.9 мм.
Сравнение. От рода Calyptrina Sokolov, 1967 они отличаются меньшими размерами, отсутствием характерной для этого рода структуры трубок в виде сложенных друг в друга воронок.
Распространение. Россия, Байкало-Патомское нагорье, уринская свита, венд.
Материал. Два неполных экземпляра.
Gen. et sp. indet. 2
Табл. II, фиг. 4, 5
Описание. Фрагменты организма, сохранившиеся в виде сегментированных углеродистых лентовидных компрессий на поверхности напластования пород. Ленты гладкостенные, жесткие, прямые, плавно закругленные с одного конца. Наблюдаемая длина фрагментов 15 и 25 мм, ширина постоянная по всей длине (1.2–1.5 мм) и сужается только к окончаниям до 0.5–0.9 мм. Длина сегментов неравномерная от 0.8 до 2.2 мм. В местах соединения сегментов внешний контур лент слегка пережат. Межсегментные перетяжки прямые и четкие.
Сравнение. От рода Protoarenicola Wang, 1982, emend. Dong et al., 2008 описываемые экземпляры отличаются более редкой сегментацией и отсутствием терминальных дисков.
Распространение. Россия, Байкало-Патомское нагорье, уринская свита, венд.
Материал. Два неполных экземпляра.
ЗАКЛЮЧЕНИЕ
В нижневендских (дошурамских, 580–570 млн лет) тонкозернистых силикокластических отложениях уринской свиты, вместе с богатыми ассоциациями акантоморфных акритарх, были обнаружены углефицированные макрофоссилии, характерные для более молодой постшурамской (<570 млн лет) миаохенской биоты. Среди уринских макрофоссилий определены виды Liulingjitaenia alloplecta и Jiuqunaoella simplicis, наряду с которыми были обнаружены неидентифицированные “сегментированные” лентовидные отпечатки, а также углистые компрессии, соответствующие диагнозу Tawuia dalensis.
В уринской свите наиболее распространены протяженные лентовидные остатки L. alloplecta, которые при жизни имели форму спирально свернутых лент. Такие организмы формировали крупные плавающие или свободно стелящиеся по дну колонии, состоящие из ориентированных в одном направлении индивидов. Большая часть уринских макрофоссилий была представлена макрофитами, которые являлись заметным детритообразующим компонентом палеобассейна. Вместе с эукариотным микрофитопланктоном этот детритообразующим основную часть захороненного в осадках органического вещества. Обнаруженные в уринской свите макрофиты сосуществовали с раннеэдиакарской микробиотой, очевидно составляя вместе с ней единую экосистему, которая была образована незадолго до Шурамского события и была существенно преобразована после него.
Благодарности. Авторы благодарны Д.В. Гражданкину, Е.Ю. Голубковой и М.А. Федонкину за ценные замечания и предложения, позволившие улучшить статью, а также Г.С. Феофанову за всестороннюю помощь в подготовке и проведении полевых работ.
Источники финансирования. Исследования проведены при поддержке гранта РФФИ 19-05-00155 и РНФ 20-77-10066.
Конфликт интересов. Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Бобров А.К. Геология Предбайкальского краевого прогиба. М.: Наука, 1964. 228 с.
Бобров А.К. Стратиграфия и палеогеография отложений верхнего докембрия Южной Якутии. Якутск: Якутское книжное издательство, 1979. 128 с.
Воробьева Н.Г., Петров П.Ю. Микробиота баракунской свиты и биостратиграфическая характеристика дальнетайгинской серии: ранний венд Уринского поднятия Сибири // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 4. С. 26–42. https://doi.org/10.31857/S0869592X20040109
Воробьева Н.Г., Сергеев В.Н. Stellarossica gen. nov. и инфрагруппа Keltmiides infragr. nov.: аномально крупные акантоморфные акритархи венда Сибири и Восточно-Европейской платформы // Палеонтол. журн. 2018. № 5. С. 91–100. https://doi.org/10.1134/S0031031X18040141
Гражданкин Д.В., Наговицин К.Е., Маслов А.В. Миаохенская экологическая ассоциация позднего венда Восточно-Европейской платформы // Докл. АН. 2007. Т. 417. № 1. С. 73–78.
Иванцов А.Ю. Новые данные о поздневендских проблематических ископаемых рода Harlaniella // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 6. С. 25–34. https://doi.org/10.7868/S0869592X13060069
Колосов П.Н. Стратиграфия верхнего докембрия юга Якутии. Новосибирск: Наука, 1975. 156 с.
Кочнев Б.Б., Карлова Г.А. Новые данные по биостратиграфии немакит-далдынского яруса венда юга Сибирской платформы // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 5. С. 28–41.
Кочнев Б.Б., Покровский Б.Г., Кузнецов А.Б., Марусин В.В. С- и Sr-изотопная хемостратиграфия карбонатных отложений венда–нижнего кембрия центральных районов Сибирской платформы // Геология и геофизика. 2018. Т. 59. № 6. С. 731–755.
Леонов М.В., Рудько С.В. Находка вендских фоссилий в отложениях дальнетайгинской серии Патомского нагорья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 5. С. 96–99. https://doi.org/10.31857/S0024497X20030052
Марусин В.В., Гражданкин Д.В., Маслов А.В. Редкинский этап эволюции вендских макрофитов // Докл. АН. 2011. Т. 436. № 5. С. 658–664.
Метелкин Д.В., Верниковский В.А., Казанский А.Ю. Тектоническая эволюция Сибирского палеоконтинента от неопротерозоя до позднего мезозоя: палеомагнитная запись и реконструкции // Геология и геофизика. 2012. Т. 53. № 7. С. 883–899. https://doi.org/10.1016/j.rgg.2012.05.006
Петров П.Ю. Постледниковые отложения дальнетайгинской серии: ранний венд Уринского поднятия Сибири. Сообщение 1. Баракунская свита // Литология и полезн. ископаемые. 2018а. № 5. С. 459–472. https://doi.org/10.31857/S0869592X20040109
Петров П.Ю. Постледниковые отложения дальнетайгинской серии: ранний венд Уринского поднятия Сибири. Cообщение 2. Уринская и каланчевская свиты и история бассейна // Литология и полезн. ископаемые. 2018б. № 6. С. 521–538. https://doi.org/10.1134/S0024497X18060083
Покровский Б.Г., Буякайте М.И. Геохимия изотопов C, O и Sr в неопротерозойских карбонатах юго-западной части Патомского палеобассейна, юг Средней Сибири // Литология и полезн. ископаемые. 2015. № 2. С. 159–186. https://doi.org/10.7868/S0024497X15010048
Петров П.Ю., Покровский Б.Г. С-изотопные свидетельства метаногенеза в осадках дальнетайгинской серии (нижний венд Патомского бассейна Сибири) // Литология и полезн. ископаемые. 2020. № 2. С. 99–112. https://doi.org/10.31857/s0024497x20020068
Покровский Б.Г., Мележик В.А., Буякайте М.И. Изотопный состав С, O, Sr и S в позднедокембрийских отложениях патомского комплекса, Центральная Сибирь. Сообщение 1. Результаты, изотопная стратиграфия и проблемы датирования // Литология и полезн. ископаемые. 2006а. № 5. С. 505–530.
Покровский Б.Г., Мележик В.А., Буякайте М.И. Изотопный состав С, O, Sr и S в позднедокембрийских отложениях патомского комплекса, Центральная Сибирь. Сообщение 2. Природа карбонатов с ультранизкими и ультравысокими значениями δ13С // Литология и полезн. ископаемые. 2006б. № 6. С. 642–654.
Рудько С.В., Петров П.Ю., Кузнецов А.Б., Шацилло А.В., Петров О.Л. Уточненный тренд δ13С в дальнетайгинской серии Уринского поднятия (венд, юг Средней Сибири) // Докл. АН. 2017. Т. 477. № 5. С. 590–594. https://doi.org/10.7868/S0869565217350183
Рудько С.В., Кузнецов А.Б., Петров П.Ю. Изотопный состав Sr в известняках дальнетайгинской серии Патомского бассейна: опорный разрез венда Сибири // Литология и полезн. ископаемые. 2020. № 3. С. 243–256. https://doi.org/10.31857/S0024497X20030052
Соколов Б.С. Древнейшие отложения раннего кембрия и сабеллидитиды // Всесоюзный симпозиум по палеонтологии докембрия и раннего кембрия. Новосибирск, 1965. С. 78–91.
Сумина Е.Л., Сумин Д.Л. Морфогенез в сообществе нитчатых цианобактерий // Онтогенез. 2013. Т. 44. № 3. С. 203–220. https://doi.org/10.7868/S0475145013030087
Хоментовский В.В., Постников А.А., Карлова Г.А., Кочнев Б.Б., Якшин М.С., Пономарчук В.А. Венд Байкало-Патомского нагорья (Сибирь) // Геология и геофизика. 2004. Т. 45. № 4. С. 465–484.
Чумаков Н.М. Оледенения Земли: история, стратиграфическое значение и роль в биосфере. М.: ГЕОС, 2015. 160 с. (Труды ГИН РАН. Вып. 611).
Чумаков Н.М., Покровский Б.Г., Мележик В.А. Геологическая история патомского комплекса, поздний докембрий, Средняя Сибирь // Докл. АН. 2007. Т. 413. № 3. С. 379–383.
Чумаков Н.М., Семихатов М.А., Сергеев В.Н. Опорный разрез вендских отложений юга Средней Сибири // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 4. С. 26–51. https://doi.org/10.7868/S0869592X13040029
An Z., Jiang G., Tong J., Tian L., Ye Q., Song H., Song H. Stratigraphic position of the Ediacaran Miaohe biota and its constraints on the age of the upper Doushantuo δ13C anomaly in the Yangtze Gorges area, South China // Precambrian Res. 2015. V. 271. P. 243–253. https://doi.org/10.1016/j.precamres.2015.10.007
Bykova N., LoDuca S.T., Ye Q., Marusin V., Grazhdankin D., Xiao S. Seaweeds through time: morphological and ecological analysis of Proterozoic and early Paleozoic benthic macroalgae // Precambrian Res. 2020. V. 350. https://doi.org/10.1016/j.precamres.2020.105875
Canfield D.E., Poulton S.W., Narbonne G.M. Late Neoproterozoic deep-ocean oxygenation and the rise of animal life // Science. 2007. V. 315. P. 92–95. https://doi.org/10.1126/science.1135013
Canfield D.E., Knoll A.H., Poulton S.W., Narbonne G.M., Dunning G.R. Carbon isotopes in clastic rocks and the Neoproterozoic carbon cycle // Am. J. Sci. 2020. V. 320. P. 97–124. https://doi.org/10.2475/02.2020.01
Chen M., Xiao Z. Discovery of the macrofossils in the Upper Sinian Doushantuo Formation at Miaohe, eastern Yangtze Gorges // Scientia Geol. Sinica. 1991. № 4. P. 317–324.
Chen M., Xiao Z. Macrofossil biota from upper Doushantuo Formation in eastern Yangtze Gorges, China // Acta Palaeontol. Sin. 1992. V. 31. P. 513–529.
Chen J.Y., Zhou G.Q. Biology of the Chengjiang fauna // Bull. Nat. Mus. Sci. 1997. V. 10. P. 11–106.
Chen M., Xiao Z., Yuan X. A new assemblage of megafossils – Miaohe biota from Upper Sinian Doushantuo Formation, Yangtze Gorges // Acta Palaeontol. Sin. 1994. V. 33. P. 391–403.
Cloetingh S., McQueen H., Lambeck K. On a tectonic mechanism for regional sea level variations // Earth Planet. Sci. Lett. 1985. V. 75. P. 157–166. https://doi.org/10.1016/0012-821X(85)90098-6
Ding L., Li Y., Hu X., Xiao Y., Su C., Huang J. Sinian Miaohe Biota. Beijing: Geol. Publ. House, 1996. 221 p.
Dong L., Xiao S., Shen B., Yuan X., Yan X., Peng Y. Restudy of the worm-like carbonaceous compression fossils Protoarenicola, Pararenicola, and Sinosabellidites from early Neoproterozoic successions in North China // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008. V. 258. P. 138–161. https://doi.org/10.1016/j.palaeo.2007.05.019
Dornbos S.Q., Oji T., Kanayama A., Gonchigdorj S. A new Burgess shale-type deposit from the Ediacaran of western Mongolia // Scientific Rep. 2016. V. 6. P. 1–5. https://doi.org/10.1038/srep23438
Fan H., Nielsen S.G., Owens J.D., Auro M., Shu Y., Hardisty D.S., Horner T.J., Bowman C.N., Young S.A., Wen H. Constraining oceanic oxygenation during the Shuram excursion in South China using thallium isotopes // Geobiology. 2020. V. 18. P. 348–365. https://doi.org/10.1111/gbi.12379
Gong Z., Kodama K.P., Li Y.-X. Rock magnetic cyclostratigraphy of the Doushantuo Formation, South China, and its implications for the duration of the Shuram carbon isotope excursion // Precambrian Res. 2017. V. 289. P. 62–74. https://doi.org/10.1016/j.precamres.2016.12.002
Grazhdankin D.V., Balthasar U., Nagovitsin K.E., Kochnev B.B. Carbonate-hosted Avalon-type fossils in Arctic Siberia // Geology. 2008. V. 36. P. 803–806. https://doi.org/10.1130/G24946A.1
Grotzinger J.P., Fike D.A., Fischer W.W. Enigmatic origin of the largest-known carbon isotope excursion in Earth’s history // Nature Geosci. 2011. V. 4. P. 285–292. https://doi.org/10.1038/ngeo1138
Hofmann H.J., Mountjoy E.W. Ediacaran body and trace fossils in Miette Group (Windermere Supergroup) near Salient Mountain, British Columbia, Canada // Can. J. Earth Sci. 2010. V. 47. P. 1305–1325. https://doi.org/10.1139/e10-070
Ivantsov A.Yu., Gritsenko V.P., Konstantinenko L.I., Zakrevskaya M.A. Revision of the problematic Vendian microfossil Beltanelliformis (=Beltanelloides, Nemiana) // Paleontol. J. 2014. V. 48. № 13. P. 1423–1448. https://doi.org/10.1134/S0031030114130036
Lambeck K., Rouby H., Purcell A., Sun Y., Sambridge M. Sea level and global ice volumes from the Last Glacial Maximum to the Holocene // PNAS. 2014. V. 111. P. 15296–15303. www.pnas.org/cgi/doi/10.1073/pnas.1411762111
LoDuca S.T., Wu M.Y., Zhao Y.L., Xiao S.H., Schiffbauer J.D., Caron J.-B., Babcock L.E. Reexamination of Yuknessia from the Cambrian of China and first report of Fuxianospira from North America // J. Paleontol. 2015. V. 89. P. 899–911. https://doi.org/10.1017/jpa.2016.3
Melezhik V.A., Fallick A.E., Pokrovsky B.G. Enigmatic nature of thick sedimentary carbonates depleted in 13C beyond the canonical mantle value: the challenges to our understanding of the terrestrial carbon cycle // Precambrian Res. 2005. V. 137. P. 131–165. https://doi.org/10.1016/j.precamres.2005.03.010
Melezhik V.A., Pokrovsky B.G., Fallick A.E., Kuznetsov A.B., Bujakaite M.I. Constraints on 87Sr/86Sr of Late Ediacaran seawater: insight from Siberian high-Sr limestones // J. Geol. Soc. 2009. V. 166. P. 183–191. https://doi.org/10.1144/0016-76492007-171
Minguez D., Kodama K.P., Hillhouse J.W. Paleomagnetic and cyclostratigraphic constraints on the synchroneity and duration of the Shuram carbon isotope excursion, Johnnie Formation, Death Valley Region, CA // Precambrian Res. 2015. V. 266. P. 395–408. https://doi.org/10.1016/j.precamres.2015.05.033
Moczydłowska M., Nagovitsin K.E. Ediacaran radiation of organic-walled microbiota recorded in the Ura Formation, Patom Uplift, East Siberia // Precambrian Res. 2012. V. 198–199. P. 1–24. https://doi.org/10.1016/j.precamres.2011.12.010
Narbonne G.M. Neoproterozoic oceans and early animals // Science. 2010. V. 328. P. 53–54. https://doi.org/10.1126/science.1188688
Pelechaty S.M. Integrated chronostratigraphy of the Vendian System of Siberia: implications for a global stratigraphy // J. Geol. Soc. 1998. V. 155. P. 957–973. https://doi.org/10.1144/gsjgs.155.6.0957
Powerman V., Shatsillo A., Chumakov N., Kapitonov I., Hourigan J. Interaction between the Central Asian Orogenic Belt (CAOB) and the Siberian Craton as recorded by detrital zircon suites from Transbaikalia // Precambrian Res. 2015. V. 267. P. 39–71. https://doi.org/10.1016/j.precamres.2015.05.015
Rooney A.D., Cantine M.D., Bergmann K.D., Gómez-Pérez I., Al Baloushi B., Boag T.H., Busch J.F., Sperling E.A., Strauss J.V. Calibrating the coevolution of Ediacaran life and environment // Proc. Natl. Acad. Sci. 2020. V. 117(29). P. 16824–16830. https://doi.org/10.1073/pnas.2002918117
Rowland S.M., Rodriguez M.G. A multicellular alga with exceptional preservation from the Ediacaran of Nevada // J. Paleontol. 2014. V. 88. P. 263–268. https://doi.org/10.1666/13-075
Rud’ko S.V., Kuznetsov A.V., Petrov P.Yu., Sitkina D.R., Kaurova O.K. Pb–Pb dating of the Dal’nyaya Taiga Group in the Ura uplift of southern Siberia: implication of C-isotopic and biotic events in the Ediacaran // Precambrian Res. 2021. V. 362. https://doi.org/10.1016/j.precamres.2021.106285
Sahoo S.K., Planavsky N.J., Jiang G., Kendall B., Owens J.D., Wang X., Shi X., Anbar A.D., Lyons T.W. Oceanic oxygenation events in the anoxic Ediacaran ocean // Geobiology. 2016. V. 14. P. 457–468. https://doi.org/10.1111/gbi.12182
Schieber J. Microbial mats in terrigenous clastics: the challenge of identification in the rock record // Palaios. 1999. V. 14. P. 3–12. https://doi.org/10.2307/3515357
Schieber J. Microbial mats on muddy substrates – examples of possible sedimentary features and underlying processes // Atlas of Microbial Mat Features Preserved within the Siliciclastic Rock Record, Atlases in Geoscience 2. Eds. Schieber J., Bose P.K., Eriksson P.G., Banerjee S., Sarkar S., Altermann W., Catuneanu O. Elsevier, 2007. P. 117–134.
Sergeev V.N., Knoll A.H., Vorob’eva N.G. Ediacaran microfossils from the Ura Formation, Baikal-Patom Uplift, Siberia: taxonomy and biostratigraphic significance // J. Paleontol. 2011. V. 85. P. 987–1011. https://doi.org/10.1666/11-022.1
Sim M.S, Liang B., Petroff A., Evans A., Klepac-Ceraj V., Flannery D.T., Walter M.R., Bosak T. Oxygen-dependent morphogenesis of modern clumped photosynthetic mats and implications for the Archean stromatolite record // Geosciences. 2012. V. 2. P. 235–259. https://doi.org/10.3390/geosciences2040235
Simonson B.M., Carney K.E. Roll-up structures: evidence of in situ microbial mats in Late Archaean deep shelf environments // Palaios. 1999. V. 14. P. 13–24. https://doi.org/10.2307/3515358
Sovetov J.K. Vendian foreland basin of the Siberian cratonic margin: Paleopangean accretionary phases // Rus. J. Earth Sci. 2002. V. 4. № 5. P. 363–387. https://doi.org/10.2205/2002ES000092
Steiner M. Die neoproterozoischen Megaalgen Sudchinas // Berliner geowissenschaftliche Abhandlungen. 1994. R. (E). B. 15. S. 1–146.
Steiner M., Zhu M., Zhao Y., Erdtmann B.D. Lower Cambrian Burgess shale-type fossil associations of South China // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005. V. 220. P. 129–152. https://doi.org/10.1016/j.palaeo.2003.06.001
Wang G. Late Precambrian Annelida and Pogonophora from the Huainan of Anhui Province // Bull. Tianjin Inst. Geol. Miner. Resour. 1982. V. 6. P. 9–22.
Wang P.-L., LoDuca S.T., Wu M.-Y., Tang Y.-G., Sun Z.-X. Benthic primary producers in exceptionally preserved Cambrian biotas of North China // Palaeoworld. 2021. https://doi.org/10.1016/j.palwor.2021.01.009
Wang Y., Wang X., Huang Y. Macroscopic algae from the Ediacaran Doushantuo Formation in northeast Guizhou, South China // Earth Sci. J. China Univ. Geosci. 2007. V. 32. P. 828–844.
Wang Y., Wang Y., Du D., Wang X. The correlation between macroscopic algae and metazoans in the Ediacaran: a case study on the Wenghui biota in northeastern Guizhou, South China // J. Aust. Earth Sci. 2014. V. 61. P. 967–977. https://doi.org/10.1080/08120099.2014.956231
Witkosky R., Wernicke B.P. Subsidence history of the Ediacaran Johnnie Formation and related strata of southwest Laurentia: implications for the age and duration of the Shuram isotopic excursion and animal evolution // Geosphere. 2018. V. 14. P. 2245–2276. https://doi.org/10.1130/GES01678.1
Xiao S., Narbonne G.M. The Ediacaran Period // Geologic Time Scale. Eds. Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Oxford: Elsevier, 2020. V. 1. P. 521–561. https://www.doi.org/10.1016/B978-0-12-824360-2.00018-8
Xiao S., Yuan X., Steiner M., Knoll A.H. Macroscopic carbonaceous compressions in a terminal Proterozoic shale: a systematic reassessment of the Miaohe biota, South China // J. Paleontol. 2002. V. 76. P. 347–376. https://doi.org/10.1017/S0022336000041743
Xiao Sh., Droser M., Gehling J.G., Hughes I.V., Wan B., Chen Z., Yuan X. Affirming life aquatic for the Ediacaran biota in China and Australia // Geology. 2013. V. 41. P. 1095–1098. https://doi.org/10.1130/G34691.1
Xiao S., Gehling J.G., Evans S.D., Hughes I.V., Droser M.L. Probable benthic macroalgae from the Ediacaran Member, South Australia // Precambrian Res. 2020. V. 350. https://doi.org/10.1016/j.precamres.2020.105903
Ye Q., Tong J., An Z., Hu J., Tian L., Guan K., Xiao S. A systematic description of new macrofossil material from the upper Ediacaran Miaohe Member in South China // J. Syst. Palaeontol. 2017. V. 17. P. 183–238. https://doi.org/10.1080/14772019.2017.1404499
Yuan X., Li J., Chen M. Development and their fossil records of metaphytes from late Precambrian // Acta Palaeontol. Sin. 1995. V. 34. P. 90–102.
Yuan X., Xiao S., Yin L., Knoll A.H., Zhou C., Mu X. Doushantuo fossils: life on the eve of animal radiation. Hefei: China University of Science and Technology Press, 2002. 171 p.
Zhang F., Xiao S., Romaniello S.J., Hardisty D., Li C., Melezhik V., Pokrovsky B., Cheng M., Shi W., Lenton T.M., Anbar A.D. Global marine redox changes drove the rise and fall of early animals // Geobiology. 2019. V. 17. P. 594–610. https://doi.org/10.1111/gbi.12359
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция