

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 6, стр. 54-83
Биохронологическая шкала верхнего тоара–нижнего аалена Восточной Сибири по двустворчатым моллюскам рода Arctotis Bodylevsky, 1960
Геологический институт, Российская академия наук
Москва, Россия
* E-mail: niipss@mail.ru
Поступила в редакцию 09.03.2021
После доработки 02.04.2021
Принята к публикации 20.04.2021
Аннотация
На основе изучения морфогенеза раковин уточнена классификация тоар-ааленских представителей рода Arctotis Bodylevsky, 1960. В опорных разрезах Анабарской губы и мыса Цветкова (Восточный Таймыр) установлена последовательность видов, которая прослежена в серии разрезов тоара и аалена Восточной Сибири, и сопоставлена с бореальной аммонитовой шкалой. Для верхнего тоара–нижнего аалена сконструирована биохронологическая шкала по двустворчатым моллюскам рода Arctotis (семейство Oxytomidae Ichikawa, 1958), имеющая собственную хронологическую периодизацию, в которой выделены четыре так называемые оксито-зоны. С помощью шкалы проведена межрегиональная корреляция разрезов верхнего тоара–нижнего аалена Восточной Сибири (Анабарская губа, мыс Цветкова, рр. Марха, Тюнг, Келимяр, Молодо, Моторчуна, скважины Вилюйской синеклизы) и Северо-Востока России (бассейн р. Левый Кедон). Хроностратиграфические объемы оксито-зон определены с детальностью, соответствующей зонам аммонитовой шкалы бореального стандарта: оксито-зона Praearctotis milovae соответствует аммонитовым зонам Zugodactylites braunianus и Pseudolioceras compactile, оксито-зона Praearctotis marchaensis – зоне Pseudolioceras wuerttenbergeri, оксито-зона Praearctotis similis – зоне Pseudolioceras falcodiscus, оксито-зона Arctotis tabagensis – зоне Pseudolioceras maclintocki.
ВВЕДЕНИЕ
Основным методом параллелизации региональных горизонтов юры Севера России со стратотипами ярусов Международной стратиграфической шкалы (МСШ) является корреляция аммонитовых зон (Зоны…, 1982). Для геологической корреляции тоар-ааленских отложений Северо-Восточной Азии в конце ХХ–начале ХХI в. сибирскими и санкт-петебургскими специалистами были разработаны две аммонитовые шкалы (Князев, 1991; Князев и др., 2003; Полуботко, Репин, 1994; Захаров и др., 1997; Шурыгин и др., 2011; Репин, 2016, 2017). Комплекс позднетоарских бореальных аммоноидей по сравнению с западноевропейским обеднен, хотя в нем есть ряд общих с Западной Европой родов и видов (Меледина, 2000). Тем не менее вопросы проведения границы между нижним и средним отделами юры и глобальной корреляции тоар-ааленских разрезов с подразделениями МСШ и на севере России остаются актуальными. В последних публикациях на эту тему нижнюю границу аалена в Восточной Сибири предлагается проводить в основании зоны Pseudolioceras maclintocki (Князев и др., 2007а), а на Северо-Востоке России – в основании слоев с Pseudolioceras beyrichi orientale (Репин, 2016). Нет общепринятой корреляции тоара и аалена по комплексам двустворчатых моллюсков в разрезах на территории Севера России, поскольку считается, что комплексы верхов тоара и низов аалена этих регионов эндемичны и разнятся между собой (Полуботко, Репин, 1992, 1994; Репин, 2020). Из-за редкой встречаемости представителей зональных видов аммонитов в тоар-ааленских отложениях, непосредственно пользоваться общей стратиграфической шкалой при расчленении и корреляции как естественных обнажений, так и разрезов, вскрытых скважинами, сложно (Шурыгин и др., 2001). Пограничные отложения нижнего и среднего отделов юрской системы на севере Сибири принято датировать тоаром–ааленом (Шурыгин и др., 2000).
Предлагаемая биохронологическая шкала по окситомидам разрабатывалась на одних и тех же разрезах севера Сибири (Анабарская губа, мыс Цветкова, рр. Моторчуна, Марха) и Северо-Востока России (бассейн р. Левый Кедон), как и аммонитовая зональная шкала тоара–аалена (Князев и др., 2003; Захаров и др., 1997; Шурыгин и др., 2011), поэтому параллелизация шкалы по двустворкам выполнена относительно сибирской аммонитовой шкалы (рис. 1).
Рис. 1.
Схема сопоставления биохронологической шкалы по окситомидам с международной стратиграфической шкалой и со стандартными зональными шкалами верхнего тоара–аалена северо-запада Западной Европы и Северо-Восточной Азии.
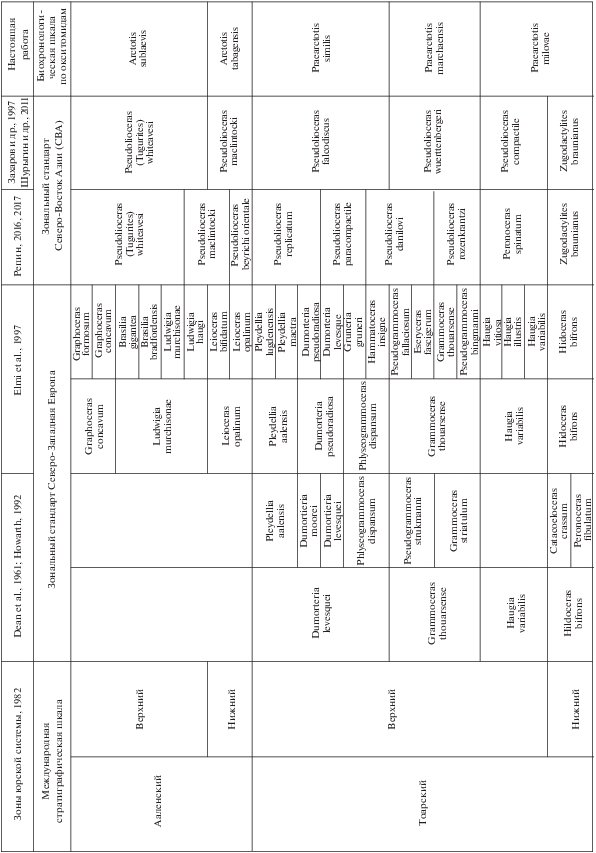
Главное назначение шкалы – периодизация отложений, вмещающих зональные комплексы, и корреляция в различной степени удаленных и разнофациальных разрезов нижней–средней юры.
Первая зональная схема юры севера Сибири была создана В.И. Бодылевским на основе изучения материалов из Анабаро-Хатангского района. В схеме для тоара и аалена наряду с аммонитами в качестве руководящих форм были указаны двустворчатые моллюски, отнесенные к двум видам – “Pseudomonotis vai sp. nov.” для верхнего лейаса (=тоара) и “Pseudomonotis lenaensis (Lahusen)”11 для аалена (Бодылевский, 1939).
В докладе на Межведомственном стратиграфическом совещании по разработке унифицированных стратиграфических схем Сибири в 1956 г. В.И. Бодылевский высказал соображение, что представители данной родовой группы для севера Сибири могут формировать определенный генетический ряд от тоара до валанжина (Бодылевский, 1957). Исследованиями английских и аргентинских специалистов установлено (биполярное) распространение рода Arctotis в юре и мелу (Crame, 1985; Crame et al., 1993; Damborenea, 1994; Damborenea et al., 2013).
Стандартные автономные шкалы по двустворчатым моллюскам, включенные в стратиграфические схемы Сибири и Северо-Востока России и являющиеся базовыми для геологических работ (Решение…, 2004; Решения…, 2009), основаны на таксонах, относящихся к разным семействам (Шурыгин, 1987; Репин, Полуботко, 2004). С появлением новых данных по распределению в разрезах двустворчатых моллюсков и аммоноидей стандартные шкалы по двустворчатым моллюскам продолжают совершенствоваться (Шурыгин и др., 2011; Репин, Полуботко, 2015). Так, в Региональной стратиграфической схеме нижней и средней юры Средней Сибири, принятой на 3-м Межведомственном региональном стратиграфическом совещании по мезозою и кайнозою Средней Сибири в 1978 г., слои с Propeamussium olenekense, Arctotis marchaensis соответствовали нижнему аалену (Решения…, 1981). Согласно современным представлениям, b-зона Arctotis marchaensis соответствует двум верхним а-зонам тоара и части нижнего аалена бореального аммонитового стандарта, b-зона Arctotis lenaensis соответствует верхней части нижнего аалена, верхнему аалену и нижнему байосу (Шурыгин и др., 2011). В Региональной стратиграфической схеме нижней и средней юры Западной Сибири, принятой на 6-м Межведомственном стратиграфическом совещании по рассмотрению и принятию уточненных стратиграфических схем мезозозойских отложений Западной Сибири в 2003 г., слои с Arctotis marchaensis соответствуют верхней части надояхского горизонта и коррелируются с двумя верхними а-зонами бореального аммонитового стандарта тоара и нижней частью аалена. Слои с Arctotis lenaensis соответствуют вымскому горизонту и верхней части лайдинского горизонта (Решение…, 2004). В Региональной стратиграфической схеме юрских отложений Северо-Востока России, принятой на 3-м Межведомственном региональном стратиграфическом совещании по докембрию, палеозою и мезозою Северо-Востока России в 2002 г., слои с Arctotis marchaensis соответствуют низам ясчанского горизонта (аален–низы байоса) (Решения…, 2009). Согласно современной корреляции (Репин, 2016), толща алевролитов с “Arctotis cf. marchaensis (Petr.)” на р. Вилига отвечает терминальной зоне тоара (Репин, Полуботко, 2015). В Региональной стратиграфической схеме юрских отложений Дальнего Востока, принятой на 4-м Межведомственном региональном стратиграфическом совещании по докембрию и фанерозою юга Дальнего Востока и Восточного Забайкалья в 1990 г., виды “Arctotis marchaensis (Petr.)”, “Arctotis aff. similis Velikzh.” отмечены в толще песчаников и алевролитов Южно-Верхоянской структурно-фациальной зоны (басс. рр. Акачан, Аллах-Юнь), где они указаны совместно с ранеааленским Pseudolioceras maclintocki (Haught). Вид “Arctotis lenaensis (Lah.)” характеризует тымагерскую свиту (аален) и усманковскую свиту (бат) Верхне-Амурской структурно-фациальной зоны (низовья рр. Шилка, Аргунь, Амазар) (Решения…, 1994).
За пределами России в стратиграфической схеме юрских отложений Аргентины представители вида Arctotis frenguellii (Damborenea) приводились при характеристике слоев с Meleagrinella, которые соответствуют части верхнего тоара и аалена (Damborenea, 1994; Riccardi et al., 2011).
Изучение коллекций двустворчатых моллюсков семейства Oxytomidae Ichikawa, 1958 из тоар-ааленских отложений севера Восточной Сибири и Северо-Востока России, начатое автором еще в конце 80-х годов ХХ в., позволило установить закономерности эволюции некоторых юрско-меловых представителей семейства Oxytomidae Ichikawa, 1958 (Лутиков и др., 2010) и выполнить предварительную классификацию таксонов (Лутиков, Шурыгин, 2010).
Целью настоящей работы является создание биохронологической шкалы верхнего тоара–нижнего аалена по двустворчатым моллюскам, относящимся к роду Arctotis Bodylevsky, 1960 (семейство Oxytomidae Ichikawa, 1958), и оценка шкалы для стратиграфических корреляций. Поводом для разработки шкалы явилось широкое распространение арктотисов в разрезах верхнего тоара и аалена и относительно высокие темпы морфогенеза у представителей этой группы двустворчатых моллюсков. В качестве основной задачи рассматривалась разработка методики видовой классификации.
С помощью шкалы проведена корреляция основных разрезов тоара и нижнего аалена Восточной Сибири и Северо-Востока России посредством прослеживания биостратонов, характеризующихся зональными видами или зональными комплексами.
МАТЕРИАЛ
В период с 1980 по 1987 г. автор участвовал в комплексных лито- и биостратиграфических исследованиях основных разрезов нижней и средней юры Восточной Сибири и Северо-Востока России (Анабарская губа, Восточный Таймыр, рр. Анабар, Келимяр, Моторчуна, Марха, Вилюй, Тюнг, Молодо, Сюнгюдэ, притоки р. Левый Кедон). Работы проводились в составе группы специалистов СНИИГГиМС (Новосибирск), изучавших литологию – В.П. Девятов, аммоноидеи – В.Г. Князев, двустворчатые моллюски – О.А. Лутиков, фораминиферы – В.В. Сапьяник. В период с 1990 по 2008 г. автор исследовал материалы из скважин разведочного бурения Вилюйской синеклизы, любезно предоставленные коллегами И.В. Будниковым и И.С. Павлухиным (рис. 2).
Рис. 2.
Обзорная карта изученных разрезов верхнего тоара и нижнего аалена Восточной Сибири и Северо-Востока России. Районы сборов коллекций: 1 – басс. р. Левый Кедон (рр. Сатурн, Старт, Бродная, Астрономическая), 2 – р. Моторчуна, 3 – мыс Цветкова (Восточный Таймыр), 4 – Анабарская губа, 5 – р. Анабар, 6 – р. Келимяр, 7 – р. Марха, 8 – р. Вилюй, 9 – р. Тюнг, 10 – рр. Сюнгюдэ, Молодо. Участки бурения: 11 – Тенкеляхский, 12 – Правобережный, 13 – Серки-Линденский.
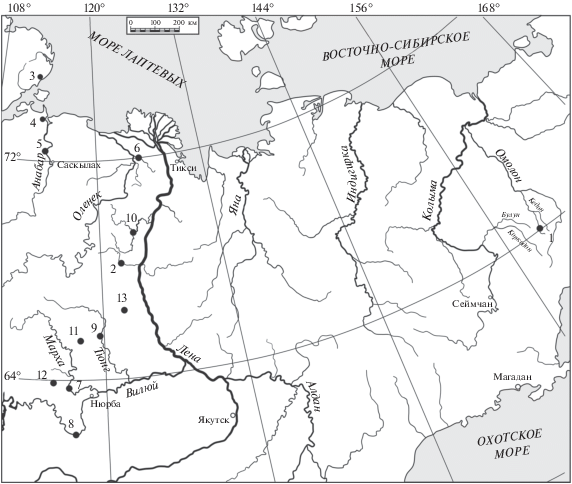
Основными объектами изучения являлись коллекции, собранные автором и коллегами в естественных выходах тоар-ааленских отложений и из керна скважин разведочного бурения, хранящиеся в департаменте стратиграфии и региональной геологии СНИИГГиМС (Новосибирск). Для уточнения систематической принадлежности и стратиграфического положения некоторых таксонов были изучены: коллекции И.В. Полуботко и Ю.С. Репина из разрезов р. Летняя и притоков р. Левый Кедон, хранящиеся в отделе стратиграфии и палеонтологии ВСЕГЕИ (Санкт-Петербург); коллекция А.Г. Ржонсницкого из разреза р. Марха, коллекция Г.А. Иванова из разреза р. Лена и коллекция Т.М. Окуневой из разреза р. Газимур, хранящиеся в музее ЦНИГР им. акад. Чернышева (Санкт-Петербург); коллекция Т.И. Кириной из разреза р. Вилюй, хранящаяся в музее ВНИГРИ (Санкт-Петербург); коллекция В.И. Бодылевского из разреза п-ова Юрюнг-Тумус, хранящаяся в музее Санкт-Петербургского горного университета; коллекции Б.Н. Шурыгина из разрезов п-ова Юрюнг-Тумус и Анабарской губы, хранящиеся в лаборатории палеонтологии и стратиграфии мезозоя и кайнозоя ИНГиГ СО РАН (Новосибирск). В коллекциях автора насчитывается более 1000 экземпляров окситомид, относящихся к роду Arctotis и представляющих более 60 выборок из 15 местонахождений. Список изученных образцов представлен ранее (Лутиков, Шурыгин, 2010) и дополнен в разделе “Описание зональных видов” в настоящей работе.
МЕТОДЫ
Полевые исследования. Стратиграфическое расчленение естественных выходов юрских отложений на севере Восточной Сибири и Северо-Востоке России проводилось на основе принципов послойной корреляции разрезов ритмичных терригенных толщ (Захаров, Юдовный, 1967). Образцы с двустворчатыми моллюсками привязывались к нижней границе слоев и к уровням находок аммоноидей. При описании разрезов предварительно определялась видовая и родовая принадлежность всех сопутствующих характерных комплексов двустворчатых моллюсков. Образцы из керна скважин привязывались к основанию литологических тел, установленных с помощью геофизических методов.
Методы систематики. В настоящем исследовании при выборе методов систематики автор следовал рекомендациям В.Е. Руженцева (Руженцев, 1960). Использовались онтогенетический принцип, принцип гомологий, принцип основного звена. Онтогенез изучался по сопоставлению очертаний элементов замочных структур у раковин разного размера в выборках из ископаемых популяций (Захаров, 1975) и по линиям роста элементов замочных структур на отдельных раковинах.
Изучение новообразований внутри группы проводилось путем сравнительного анализа гомологичных частей лигаментного и биссусного блоков раковин в выборках из разобщенных во времени и пространстве ископаемых популяций. При проведении систематики таксонов за основное звено развития окситомид приняты изменения относительного положения частей в лигаментном и биссусном блоках. В эволюции окситомид эмпирически установлены хроноклины. В результате апостериорного взвешивания признаков были определены морфологические признаки, характеризующие надвидовые таксоны. На фоне гипотетических эволюционных трендов, основанных на визуальной оценке изменений гомологичных частей в лигаментном и биссусном блоках во времени, была проведена первичная классификация таксонов (Лутиков, Шурыгин, 2010). Выводы по филогенезу основывались на сравнении типов и подтипов онтогенезов лигаментных ямок у юрских и меловых форм, относящихся к различным стратиграфическим уровням. С помощью кладистического анализа установлены филогенетические связи между родственными группами окситомид (Лутиков и др., 2010).
Методы классификации. При выборе методов классификации двустворчатых моллюсков автор следовал рекомендациям Э. Майра (Майр, 1971). В хронологической последовательности видовых групп из подродов Arctotis (Praearctotis) и Arctotis (Arctotis) дискретность видов определялась методом апостериорного взвешивания признаков, основанного на опыте оценки количества филогенетически значимой информации, содержащейся в выбранном признаке. Классификация и видовая диагностика таксонов в настоящем исследовании проводились с помощью количественной оценки состояний морфологических признаков в комплексе с данными по изучению морфогенеза.
К признакам с высоким таксономическим весом отнесены:
а) Форма переднего крыла. Тупоугольное переднее крыло присутствует у тоарских Praearctotis, субпрямоугольное переднее крыло появляется у ааленских Arctotis.
б) Ширина переднего крыла. Узкое крыло характерно для большинства тоарских представителей подрода Praearctotis. Умеренно широкое переднее крыло свойственно терминальным тоарским Praearctotis. Широкое переднее крыло характеризует аален-байосские таксоны подрода Arctotis s.str.
в) Соотношение выпуклости створок. Двояковыпуклые створки характерны для таксонов тоарского подрода Praearctotis, выпуклые левые створки и плоские правые створки свойственны раннеааленским Arctotis s.str. Выпукло-вогнутые створки характерны для позднеаален-байосских представителей подрода Arctotis s.str.
К признакам с низким таксономическим весом отнесены признаки, подверженные возрастной или индивидуальной изменчивости: размер, контур, симметрия, скошенность, длина переднего и заднего залигаментного поля, плотность ребристости. К этой же категории отнесены признаки, которые трудно определять, такие как относительная ширина межреберных промежутков и дифференцированность ребер по ширине и высоте на порядки.
При определении дискретности видов принята концепция эволюционного вида Дж. Симпсона, согласно которой “Эволюционный вид – это филетическая линия (последовательность популяций, связанных отношением предок–потомок), которая эволюирует независимо от других и обладает своей собственной единой эволюционной ролью и эволюционными тенденциями” (Симпсон, 2006, с. 178).
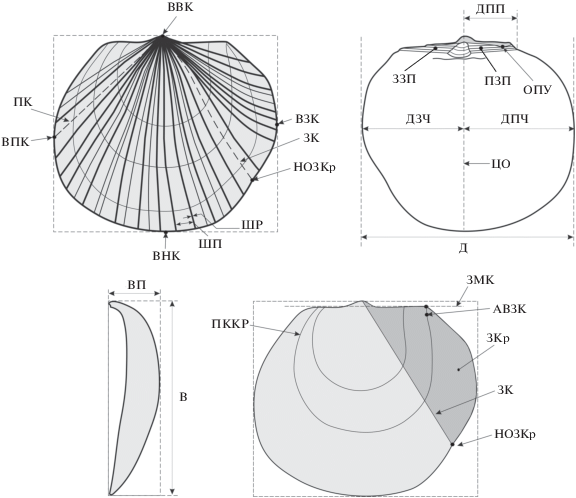
Морфология раковины и используемые измерения. Понятия таких морфологических признаков, как длина, высота, радиальные ребра, передний край, задний край, замочный край, переднее ушко, заднее ушко, передняя часть раковины, задняя часть раковины, выпуклость створки, лигаментная ямка, залигаментное поле, лигаментная площадка, протуберанец, известны в литературе по двустворчатым моллюскам с прямым замочным краем (Кошелкина, 1963; Лутиков, Шурыгин, 2010) (рис. 3).
Рис. 3.
Морфологические элементы левой створки арктотисов. ВВК – апикальный выступ верхнего края, ВПК – апикальный выступ переднего края, ВЗК – апикальный выступ заднего края, ВНК – апикальный выступ нижнего края, ПК – передний киль створки, ЗК – задний киль створки, ЦО – центральная ось створки, ЗЗП – заднее залигаментное поле, ПЗП – переднее залигаментное поле, ДПП – длина переднего залигаментного поля, ОПУ – окончание переднего ушка, ЗКр – заднее крыло створки, ЗМК – замочный край, ПККР – передний край концентра роста, НОЗКр – нижнее окончание заднего крыла створки, АВЗК – апикальный вырез заднего края концентра роста, ШР – ширина ребер, ШП – ширина межреберных промежутков, ДЗЧ – длина задней части, ДПЧ – длина передней части, Д – длина, ВП – выпуклость, В – высота.
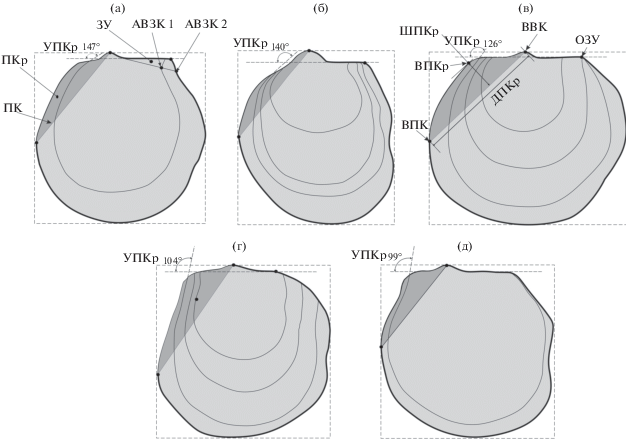
При описании формы переднего крыла левой створки использовались новые признаки, которым даны унифицированные понятия: угол переднего крыла (УПКр) – угол между замочным краем (ЗМК) и передним краем последнего видимого концентра роста; передний киль (ПК) – линия, соединяющая ВВК и ВПК; апикальный выступ внешнего края переднего крыла левой створки (ВПКр) – точка максимального удаления переднего края створки от киля; длина изгиба внешнего крыла правой створки (ДПКр) – расстояние между ВВК и ВПК; ширина изгиба внешнего крыла правой створки (ШПКр) – расстояние между килем и ВПКр; передний край концентра роста (ПККР) – дуга концентра роста, соединяющаяся с краем створки. При описании формы заднего крыла левой створки использовались признаки: ЗК – задний киль, ЗКр – заднее крыло, ЗУ – заднее ушко (дифференцировано только у раннетоарских форм), НОЗКр – нижнее окончание заднего крыла, АВЗК 1 – апикальный вырез заднего края концентра роста, АВЗК 2 – апикальный вырез внешнего края заднего крыла (рис. 4).
Рис. 4.
Морфологические элементы переднего и заднего крыла левой створки арктотисов. (а) – Arctotis (Praearctotis) milovae, (б) – Arctotis (Praearctotis) marchaensis, (в) – Arctotis (Praearctotis) similis, (г) – Arctotis (Arctotis) tabagensis, (д) – Arctotis (Arctotis) sublaevis. Морфологические элементы: ПКр – переднее крыло, ПК – передний киль, УПКр – угол переднего крыла, ДПКр – длина изгиба внешнего крыла правой створки, ШПКр – ширина изгиба внешнего крыла правой створки, ВВК – апикальный выступ верхнего края, ЗУ – заднее ушко, ОЗУ – окончание заднего ушка, АВЗК 1 – апикальный вырез заднего края концентра роста, АВЗК 2 – апикальный вырез внешнего края заднего крыла, ВПК – апикальный выступ переднего края, ВПКр – апикальный выступ внешнего края переднего крыла левой створки, ВВК – апикальный выступ верхнего края.
Для характеристики морфологических элементов раковин производились замеры по 9 признакам: 1) длина (Д), 2) высота (В), 3) выпуклость (Вп), 4) длина передней части (ДПЧ), 5) длина задней части (ДЗЧ), 6) длина переднего залигаментного поля (ДПП), 7) ребристость (Р) – количество ребер на участке раковины в промежутке между передним килем створки и задним килем створки, 8) ширина межреберных промежутков (ШП) – расстояние между параллельными ребрами, 9) ширина ребер (ШР) – расстояние между краями ребер (рис. 3). При проведении линейных замеров по признакам 1–6 измерялся только один параметр (высота или длина створки в зависимости от сохранности материала) с помощью штангельциркуля. Остальные параметры определялись по фотографиям раковин с помощью инструментов программы Photoshop. Замеры по признакам 7–9 производились с помощью инструментов программы Photoshop и CorelDRAW. Замеры по признакам УПКр, ДПКр, ШПКр производились с помощью инструментов программы CorelDRAW.
Количественная оценка признаков разработана на основе методики изучения формы раковин с прямым замочным краем (Лутиков, Шурыгин, 2010). Количественная оценка признаков проводилась по 9 градациям (рис. 5).
Рис. 5.
Морфологическая характеристика раковин окситомид на основании количественной оценки признаков.
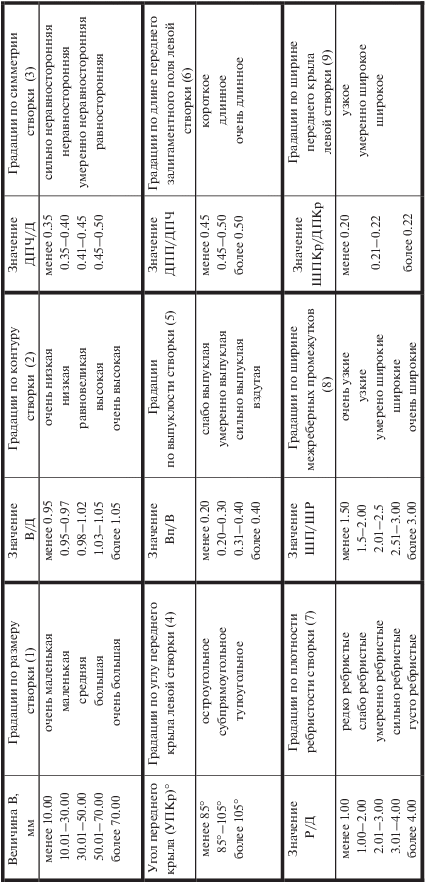
1. Градации по размеру раковины определялись высотой створок (В).
2. Градации по характеру контура раковины определялись отношением высоты раковины к длине (В/Д).
3. Градации по симметрии створок определялись отношением длины передней части к длине створки (ДПЧ/Д).
4. Градации по углу переднего крыла (УПКр) створок определялись углом между замочным краем створки и касательной к переднему краю последнего видимого концентра роста.
5. Градации по выпуклости левой створки определялись отношением выпуклости левой створки к высоте (ВпЛ/В).
6. Градации по длине переднего залигаментного поля левой створки определялись отношением длины переднего залигаментного поля левой створки к длине передней части створки (ДПП/ДПЧ).
7. Градации по плотности ребристости створок определялись отношением количества ребер по нижнему краю раковины на участке между апикальными выступами переднего и заднего краев к длине раковины (Р/Д).
8. Градации по ширине межреберных промежутков определялись отношением (ШП/ШР) ширины межреберного промежутка между соседними ребрами (ШП) к ширине смежного ребра (ШР). Поскольку ширина межреберных промежутков не равномерна, для замера выбирался наиболее широкий промежуток между ребрами по нижнему или заднему краю левых створок.
9. Градации по ширине переднего крыла левой створки определялись отношением ширины переднего крыла к длине переднего крыла левой створки (ШПКр/ДПКр).
Методология создания биохронологической шкалы. Биостратиграфическое расчленение и корреляции изученных разрезов проводились на основе эволюционного метода (Степанов, Месежников, 1979). Представленная зональная шкала верхнего тоара–нижнего аалена базируется на филогенетической последовательности видов рода Arctotis (семейство Oxytomidae Ichikawa, 1958). В качестве методологической основы для разработки шкалы была применена концепция биохронологических шкал (БХШ), предложенная В.В. Черных (Черных, 2016), апробированная на окситомидах нижнего тоара (Лутиков, Арп, 2020).
Эволюционные изменения состояний признаков, установленные в результате изучения морфогенеза лигаментного блока у раковин окситомид, происходящих из разных стратиграфических толщ, приняты за филогенетическую хроноклину при конструировании биохронологической шкалы. Различные сочетания состояний наружных признаков раковин лежат в основе периодизации шкалы. Соответственно, таксоны описывались на основании признаков, возникших в результате филогенеза, по состояниям которых определялись границы таксона. Относительная дискретность состояний признаков объясняется неполнотой геологической летописи (Дарвин, 1991).
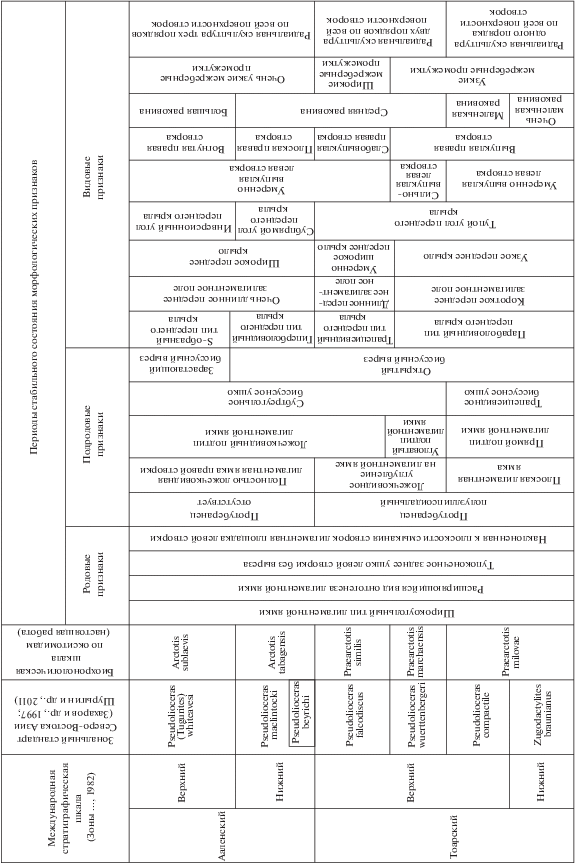
Дистанции с определенными состояниями морфологических признаков соответствуют времени существования видов. Из-за индивидуальной изменчивости признаков, границы между таксонами динамические (имеют интервалы неопределенности). Дискретность таксонов определялась в процессе изучения морфогенеза методом оценки количества филетической информации, содержащейся в эмпирически выбранном признаке (Майр, 1971). В результате установлена хронологическая последовательность состояний таксономически значимых признаков для тоара и аалена, калиброванная с помощью независимой бореальной аммонитовой шкалы (рис. 6). С одной стороны, генетические дистанции между таксонами не соответствуют хронологическим дистанциям соответствующих филозон, так как филозоны кратны элементарным подразделениям стандартной шкалы – аммонитовым зонам. С другой стороны, “если в эволюции направленность сочетается с периодичностью, то она обладает собственным временем и может служить часами, хотя и несовершенными” (Красилов, 1977, с. 29). Поэтому биохронологическая шкала по окситомидам, кроме выполнения функции стратиграфической корреляции, является инструментом датировки геологических событий. Поскольку события определяют время, одновременность может рассматриваться как сосуществование событий, для которых отношения “раньше-позже” неразличимы и определяются с точностью до фазы. Одновременность образования изолированных географически палеопопуляций определяется их принадлежностью к одному из классов эволюционных событий в морфогенезе лигаменного блока правой створки, которым соответствуют фаза образования прямого подтипа лигаментной ямки, фаза образования угловатого подтипа лигаментной ямки, фаза образования ложечковидного подтипа лигаментной ямки, фаза образования ложечки (полностью вогнутой лигаментной ямки).
Рис. 6.
Периодизация стабильных состояний морфологических признаков относительно подразделений международной стратиграфической шкалы и бореальной аммонитовой шкалы.
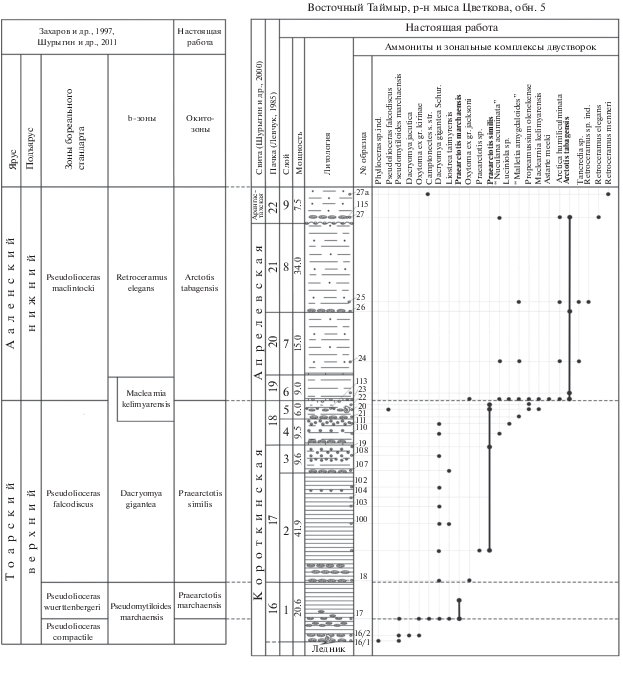
Для определения стратиграфического положения слоев с двустворчатыми моллюсками в общей последовательности зональных биостратонов в качестве опорных были выбраны разрезы тоара–аалена Анабарской губы и мыса Цветкова, которые были изучены во время полевых работ в 1985 и 1987 гг. Для оценки корреляционного потенциала шкалы последовательность видов, установленная в опорных разрезах, была прослежена в серии разрезов тоара–аалена Восточной Сибири (рр. Моторчуна, Молодо, Сюнгюдэ, Марха, Тюнг, Вилюй, скважины Вилюйской синеклизы) и Северо-Востока России (бассейн р. Левый Кедон).
МОРФОГЕНЕЗ РАКОВИНЫ РОДА ARCTOTIS BODYLEVSKY, 1960 В ПОЗДНЕМ ТОАРЕ–РАННЕМ ААЛЕНЕ
В процессе изучения закономерностей морфогенеза лигаментного блока у тоар-ааленских представителей семейства Oxytomidae Ichikawa, 1958 было установлено, что в хроноклине филогенетической линии Arctotis в раннем тоаре (фаза Zugodactylites braunianus) в лигаментном блоке возник прямой подтип лигаментной ямки, в позднем тоаре (фаза Pseudolioceras wuerttenbergeri) – угловатый подтип лигаментной ямки. В фазу Pseudolioceras falcodiscus появился ложечковидный подтип лигаментной ямки. В раннем аалене (фаза Pseudolioceras maclintocki) изменилось структурное положение элементов лигаментного блока правой створки – образовалась полностью вогнутая лигаментная ямка, уменьшилась длина лигаментной площадки относительно длины правой створки, увеличился наклон лигаментной ямки левой створки относительно плоскости смыкания створок (Лутиков и др., 2010).
Изменение типа переднего крыла левой створки, характеризующего внешний облик раковины, положено в основу периодизации морфогенеза раковин в позднем тоаре–позднем аалене. Тип переднего крыла определялся комбинацией трех признаков. У арктотисов установлено четыре типа переднего крыла. “Параболовидный” тип характеризуется коротким залигаментным полем (ДПП/ДПЧ менее 0.45), узким передним крылом (ШПКр/ДПКр менее 0.15) и тупоугольным передним крылом (УПКр более 105°). “Трапециевидный” тип характеризуется длинным залигаментным полем (ДПП/ДПЧ 0.45–0.50), умеренно широким передним крылом (ШПКр/ДПКр 0.15–0.20) и тупоугольным передним крылом (УПКр более 105°). “Гиперболовидному” типу свойственны очень длинное залигаментное поле (ДПП/ДПЧ более 0.50), широкое переднее крыло (ШПКр/ДПКр более 0.20) и субпрямоугольное (УПКр 85°–105°) или остроугольное переднее крыло (УПКр менее 85°). “S-образный” тип характеризуется очень длинным залигаментным полем (ДПП/ДПЧ более 0.50), широким передним крылом (ШПКр/ДПКр более 0.20) и инверсионным передним крылом – субпрямоугольным (УПКр 85°–105°) на ранних стадиях роста и тупоугольным (УПКр более 105°) на поздних стадиях роста.
В результате сравнения типов переднего крыла левой створки у форм из отложений, относящихся к разным стратиграфическим уровням, установлено, что морфогенез в хроноклине Arctotis (Praearctotis) milovae–Arctotis (Praearctotis) marchaensis (Petrova)–Arctotis (Praearctotis) similis Velikzhanina–Arctotis (Arctotis) tabagaensis (Petrova)–Arctotis (Arctotis) sublaevis (Bodylevsky) от фазы Zugodactylites braunianus до фазы Pseudolioceras whiteavesi проходил в направлении от форм с “параболовидным” типом переднего крыла к формам с “трапециевидным” типом переднего крыла, далее к формам с “гиперболовидным” и с “S-образным” типами переднего крыла. Морфогенез переднего крыла левой створки проходил от форм с тупоугольным крылом в тоаре к формам с субпрямоугольным крылом в аалене. Морфогенез заднего ушка левой створки проходил от форм с остроконечным окончанием заднего ушка на юношеских стадиях в раннем тоаре к формам с тупоугольным задним ушком на юношеских стадиях в позднем тоаре.
В представленной хронофилогенетической последовательности видов из подродов Prearctotis и Arctotis дискретность таксонов видового ранга определялась количественным состоянием признаков, установленных на основе изучения морфогенеза переднего крыла и заднего ушка левой створки. К виду Arctotis (Praearctotis) milovae (Okuneva), 2002, характерному для зоны Zugodactylites braunianus и зоны Pseudolioceras compactile верхнего тоара Восточной Сибири, Северо-Востока России и Дальнего Востока, отнесены формы с “параболовидным” типом переднего крыла, с остроугольным задним ушком на юношеских стадиях (табл. I , фиг. 1). К виду Arctotis (Praearctotis) marchaensis (Petrova), 1947, характерному для зоны Pseudoloceras wuerttenbergeri верхнего тоара Восточной Сибири, отнесены формы с “параболовидным” типом переднего крыла, с субпрямоугольным задним ушком на юношеских стадиях (табл. I , фиг. 2). К виду Arctotis (Praearctotis) similis Velikzhanina, 1966, характерному для зоны Pseudoloceras falcodiscus верхнего тоара Восточной Сибири, отнесены формы с “трапециевидным” типом переднего крыла и тупоугольным задним ушком на юношеских стадиях (табл. I , фиг. 3). К виду Arctotis (Arctotis) tabagensis (Petrova), 1953, характерному для зоны Pseudolioceras maclintocki нижнего аалена Восточной Сибири, отнесены формы с “гиперболовидным” типом переднего крыла и тупоугольным задним ушком на юношеских стадиях (табл. I , фиг. 4). К виду Arctotis (Arctotis) sublaevis (Bodylevsky), 1958, характерному для зоны Pseudoloceras whiteavesi, отнесены формы с “S-образным” типом переднего крыла и тупоугольным задним ушком на юношеских стадиях (табл. I , фиг. 5).
Таблица I . Фиг. 1. Arctotis (Praearctotis) milovae (Okuneva). а – экз. № МХ-309/19, левая створка, ×1, б – то же, ×3, Марха, обн. 6, слой 3, обр. № МХ-229, нижний тоар, зона Zugodactylites braunianus, в – экз. № АГ-103/2, лигаментная ямка левой створки, ×14, Анабарская губа, западный берег, обн. 5, слой 76, нижний тоар, зона Zugodactylites braunianus, г – экз. ТХ-1/1, прямой подтип лигаментной ямки правой створки, Тенкеляхская площадь, линия 1060, скв. 350, глуб. 31 м, нижний тоар. Фиг. 2. Arctotis (Praearctotis) marchaensis (Petrova). а – экз. № МХ-233/1, левая створка, ×1; р. Марха, обн. 10, сл. 8, верхний тоар, зона Pseudolioceras wuerttenbergeri, б – экз. № МХ/111-2; лигаментная ямка левой створки, ×3.5; р. Mapxa, обн. 4, осыпь слоев 2–3, верхний тоар, зона Pseudolioceras wuerttenbergeri, в – типовой экз. №574/5393-2, ЦНИГР музей им. акад. Ф.Н. Чернышова (Санкт-Петербург), угловатый подтип лигаментной ямки правой створки, лигаментная ямка без ложечковидного углубления; р. Марха, верхний тоар, колл. Г.Т. Петровой (Пчелинцевой), г – экз. № МХ/111-1, угловатый подтип лигаментной ямки правой створки, лигаментная ямка с узким ложечковидным углублением, р. Марха, обн. 4, осыпь слоев 2–3, ×3.5; верхний тоар, зона Pseudolioceras wuerttenbergeri. Фиг. 3. Arctotis (Praearctotis) similis Velikzhanina. а – экз. № ТР-19/1, левая створка, ×1; Восточный Таймыр, мыс Цветкова, обн. 5, слой 3, обр. ТФ-19, нижняя юра, верхний тоар, зона Pseudolioceras falcodiscus, б – экз. № МН-216/1, лигаментная ямка левой створки, ×4, р. Моторчуна, обн. 4, слой 2, обр. 216, верхний тоар, зона Pseudolioceras falcodiscus, в – экз. № ТР-19/1, ложечковидный подтип лигаментной ямки правой створки, лигаментная ямка с широким ложечковидным углублением, ×4, Восточный Таймыр, мыс Цветкова, обн. 5, слой 3, обр. ТФ-19, нижняя юра, верхний тоар, зона Pseudolioceras falcodiscus. Фиг. 4. Arctotis (Arctotis) tabagensis (Petrova). а – экз. № АГ-116/1-л, левая створка, ×1, б – экз. № АГ-116/1-п, вогнутая ложечковидная лигаментная ямка правой створки, ×2.5, Анабарская губа, западный берег, обн. 5, слой 92; средняя юра, конденсированный слой, нижний–верхний аален (?). Фиг. 5. Arctotis (Arctotis) sublaevis (Bodylevsky). а – экз. № АН-18/1, левая створка, ×1, р. Анабар, обн. 3, слой 8, обр. 18, средняя юра, конденсированный слой, верхний аален–нижний байос (?), б – сильно наклоненная лигаментная ямка того же экземпляра, ×2, в – экз. № АН-18/1-п, вогнутая ложечковидная, лигаментная ямка правой створки, ×2, р. Анабар, обн. 3, слой 8, обр. 18, средняя юра, конденсированный слой, верхний аален–нижний байос (?).
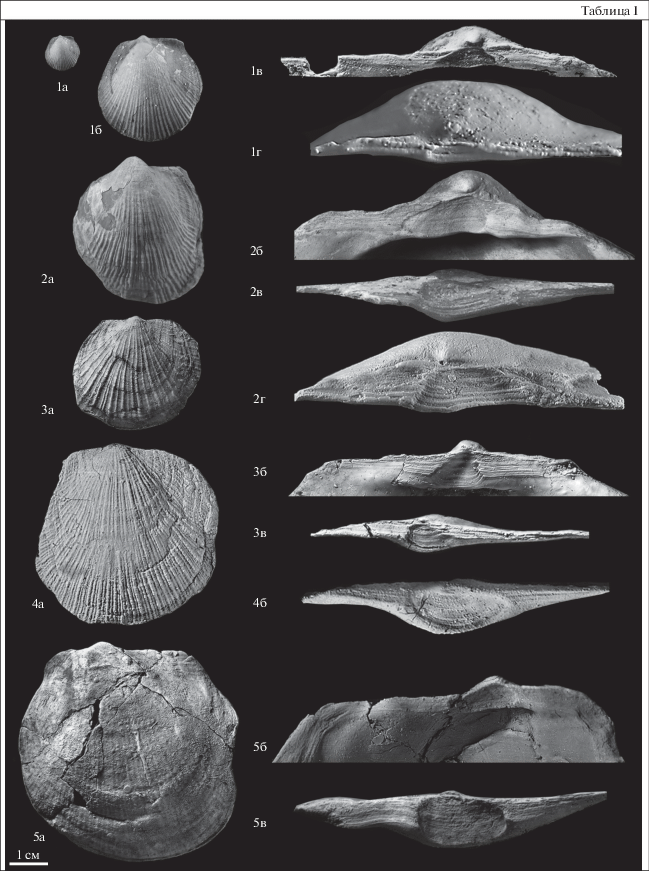
Таблица II. Фиг. 1–13 . Arctotis (Praearctotis) marchaensis (Petrova, 1947). 1 – голотип, экз. № 574/5393-1, а – вид левой створки снаружи, б – вид левой створки сбоку со стороны переднего края, ×1, в – паратип, экз. № 574/5393-2, вид правой створки изнутри; р. Марха, верхний тоар (коллекция А.Г. Ржонсницкого, ЦНИГР музей им. акад. Ф.Н. Чернышева); 2 – экз. № МХ-111/2, вид левой створки снаружи, ×1; р. Марха, обн. 4, осыпь слоев 2–3, обр. 111, верхний тоар; 3 – экз. № МХ-111/3, а – левая створка, б – вид со стороны переднего края; р. Марха, обн. 4, осыпь, верхний тоар; 4 – экз. № МХ-220/6 л, а – левая створка, б – вид со стороны переднего края; р. Марха, обн. 5, осыпь, обр. 220, верхний тоар; 5 – экз. № МХ-233/2, а – левая створка, ×1, б – то же, вид сбоку со стороны переднего края, ×1, в – вид со стороны замочного края, ×2.5; р. Марха, обн. 10, сл. 8, верхний тоар; 6 – экз. № МХ-233/1, вид левой створки снаружи, ×1; р. Марха, обн. 10, сл. 8, верхний тоар; 7 – экз. № ТХ-1/5, а – вид юношеского экземпляра левой створки снаружи, б – то же, ×2; Тенкеляхская площадь, линия 1080, скв. 350, гл. 35 м, верхний тоар; 8 – экз. № ТХ-1/2, а – вид юношеского экземпляра правой створки снаружи, б – то же ,×2; Тенкеляхский участок, линия 1080, скв. 350, гл. 35 м, верхний тоар; 9 – экз. № АГ-37/1, вид деформированной левой створки снаружи, ×1, Анабарская губа, обн. 4, сл. 18, верхний тоар; 10 – экз. № АГ-37/2, вид правой створки снаружи, ×1, Анабарская губа, обн. 4, сл. 18, верхний тоар; 11 – экз. № 7/306, голотип “Arctotis vai” Bodylevsky, а – вид левой створки снаружи, ×1, б – вид сбоку со стороны переднего края, ×1; п-ов Юрюнг-Тумус, верхний тоар, коллекция В.И. Бодылевского, Палеонтолого-стратиграфический музей Горного университета (Санкт-Петербург); 12 – экз. № 8/306, вид правой створки изнутри, ×1, п-ов Юрюнг-Тумус, верхний тоар, коллекция В.И. Бодылевского, Палеонтолого-стратиграфический музей Горного университета (Санкт-Петербург); 13 – экз. № Н-73/ 2913/1 л, а – ядро левой створки, ×1, б – вид раковины ядра сбоку со стороны переднего края, ×1, в – ядро правой створки, ×1; п-ов Юрюнг-Тумус, верхний тоар, коллекция Б.Н. Шурыгина (ИНГиГ, Новосибирск).

Таблица III. Фиг. 1. Arctotis (Praearctotis) marchaensis (Petrova, 1947), экз. № 20/653 (голотип “Arctotis viluiensis” Velikzhanina), музей ВНИГРИ, колл. Т.И. Кириной, ядро левой створки, ×1; р. Вилюй, обн. 54–56, обр. 1394, верхний тоар. Фиг. 2–8. Arctotis (Praearctotis) similis Velikzhanina, 1966. 2 – голотип, экз. № 14/653, музей ВНИГРИ, колл. Т.И. Кириной, левая створка, ×1; р. Вилюй, обн. 54, обр. 1953, верхний тоар; 3 – экз. № ТР-19/1, вид левой створки снаружи, ×1; Восточный Таймыр, мыс Цветкова, обн. 5, слой 3, обр. ТФ-19, нижняя юра, верхний тоар; 4 – экз. № ф-113/1, вид левой створки снаружи, ×1; Восточный Таймыр, мыс Цветкова, обн. 5, слой 5, обр. ф-113, нижняя юра, верхний тоар; 5 – экз. № МН-216/1, а – вид левой створки снаружи, ×1, б – вид левой створки сбоку, ×1; р. Моторчуна, обн. 4, сл. 2, нижняя юра, верхний тоар; 6 – экз. № МН-216/2, вид правой створки снаружи, ×1; там же, возраст тот же; 7 – экз. № АГ-114/1, ядро левой створки снаружи, ×1; Анабарская губа, западный берег, обн. 5, сл. 89; средняя юра, верхний тоар; 8 – экз. № АГ-114/2, отпечаток правой створки, ×1, там же, возраст тот же. Фиг. 9–14 . Arctotis (Arctotis) tabagensis (Petrova, 1953). 9 – голотип, экз. № 602/5393, ядро левой створки, ×1; р. Лена, Табагинский мыс (Большой Кангалласский Камень), средняя юра, аален; коллекция Г.А. Иванова, ЦНИГР музей им. акад. Ф.Н. Чернышева (Санкт-Петербург); 10 – экз. № ТФ-26/1, ядро левой створки, ×1; Восточный Таймыр, мыс Цветкова, обн. 5, сл. 8, обр. ТФ-26, средняя юра, нижний аален; 11 – экз. № 82 ф/1, а – ядро левой створки, ×1; б – отпечаток этой же створки, ×1; Восточный Таймыр, мыс Цветкова, обн. 7/1, сл. 3, кровля, обр. 82ф, средняя юра, нижний аален; 12 – экз. № АГ-116/1, вид левой створки снаружи, ×1, Анабарская губа, западный берег, обн. 5, сл. 92, средняя юра, нижний аален; 13 – экз. № АГ-116/2, вид правой створки снаружи, ×1, там же, возраст тот же; 14 – экз. ТФ-26/2, ядро левой створки, ×1; там же, возраст тот же; 14 – экз. ТХ-36/1, вид ядра левой створки, ×1, Тенкеляхский участок, линия 160, скв. 36, гл. 26 м.

Проведенные во время полевых исследований тафономические наблюдения указывают на сходные условия существования популяций каждого вида – все они обитали в условиях мелководья и активной гидродинамики. Следовательно, морфологические изменения в популяциях могут быть объяснены лишь как результат стабилизирующего действия отбора, т.е. эволюционными причинами.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На основе изучения морфогенеза раковин уточнена классификация тоар-ааленских представителей рода Arctotis Bodylevsky, 1960. Систематическая принадлежность надвидовых таксонов, выделенных в результате ревизии рода Arctotis Bodylevsky, 1960 (Лутиков, Шурыгин, 2010), определена с учетом кладистического анализа системы семейства Oxytomidae Ichikawa, 1958 (Лутиков и др., 2010). Систематическая принадлежность таксонов, относящихся к другим семействам и входящих в зональные комплексы оксито-зон, определена на основе проведенных ранее описаний и монографических исследований двустворчатых моллюсков (Крымгольц и др., 1953; Полевой…, 1968; Захаров, Шурыгин, 1978; Полуботко, 1992). Анализ стратиграфического положения таксонов, приведенных при характеристике разрезов, проведен с использованием собственных полевых наблюдений, а также сведений по литостратиграфии и зональной стратиграфии, полученных из публикаций коллег (Князев и др., 1991, 2003; Шурыгин и др., 2000 и др.).
В опорных разрезах Анабарской губы и мыса Цветкова (Восточный Таймыр) установлена эволюционная последовательнось видов рода Arctotis Bodylevsky, 1960, которая прослежена в серии разрезов тоара и аалена Восточной Сибири, и сопоставлена с бореальной аммонитовой шкалой (рис. 7, 8). Для верхнего тоара–нижнего аалена сконструирована биохронологическая шкала по двустворчатым моллюскам рода Arctotis.
БИОХРОНОЛОГИЧЕСКИЕ ПОДРАЗДЕЛЕНИЯ ПОЗДНЕГО ТОАРА–РАННЕГО ААЛЕНА ПО ОКСИТОМИДАМ
Биохронологическая шкала создана на основе изучения морфогенеза раковин двустворчатых моллюсков и сукцессии видов, принадлежащих одной филогенетической линии семейства Oxytomidae Ichikawa, 1958 – роду Arctotis Bodylevsky, 1960. Элементарными подразделениями шкалы являются “оксито-зоны”. По палеонтологическому и стратиграфическому критериям оксито-зоны представляют собой филозоны – слои, в которых распространены виды-индексы, представляющие собой отрезки филогенетической линии рода Arctotis. Морфогенез рода Arctotis, с одной стороны, имеет направленность, которая запечатлена в последовательности сменяющих друг друга неповторяющихся состояний лигаментного и биссусного блоков, с другой стороны, обладает периодичностью, которая обнаруживается в периодах относительно стабильного состояния различных наружных морфологических признаков, повторяющихся на разных стратиграфических уровнях. Различные сочетания состояний наружных морфологических признаков, характеризующих виды, позволяют обнаружить периодизацию филогенетической линии. Направленная эволюция рода Arctotis в сочетании с ее периодичностью обладает собственным временем, а соответствующая отрезкам филогенетической линии рода шкала может рассматриваться как биохронологическая. Время образования оксито-зон соответствует фазам существования видов-индексов.
В конкретных разрезах оксито-зоны выделяются по присутствию в них видов-индексов или условно – по комплексам зональных видов, определенных по их совместному нахождению с видом-индексом в стратотипах. На основании характеристик оксито-зон проведена детальная корреляция разрезов на севере Восточной Сибири.
Оксито-зона Praearctotis milovae
Номенклатура. В установленном хроностратиграфическом объеме выделяется впервые. В качестве зонального подразделения Восточной Сибири по двустворкам в объеме верхней части зоны Dactylioceras athleticum и зоны Zugodactylites monestieri ранее выделялась подзона Meleagrinella faminaestriata (Шурыгин, 1987). Формы из восточносибирских разрезов, относившиеся к виду “Meleagrinella faminaestriata”, в результате ревизии были переопределены как Arctotis (Praearctotis) milovae (Okuneva) (Лутиков, Шурыгин, 2010). Оксито-зона Praearctotis milovae соответствует зонам Zugodactylites braunianus и Pseudolioceras compactile бореального стандарта.
Вид-индекс: Arctotis (Praearctotis) milovae (Okuneva), 2002.
Стратотип оксито-зоны: Восточная Сибирь, Анабарская губа, обн. 5, слои 75–79, эренская свита, мощность 31.8 м.
Зональный комплекс оксито-зоны составляют виды двустворчатых моллюсков: Liostrea taimyrensis Zakh. et Schur., Dacryomya jacutica (Petr.), Modiolus numismalis Opp., Tancredia securiformis Dunk., Camptonectes s.str., Pseudomytiloides oviformis (Khudyaev in Petrova, 1953), P. marchaensis (Petr.).
Принцип выделения границ и обоснование возраста. Нижняя граница окито-зоны Praearctotis milovae устанавливается по появлению вида-индекса. Верхняя граница проводится по появлению Arctotis (Praearctotis) marchaensis (Petr.).
Возраст оксито-зоны относительно аммонитовой шкалы определяется суммой тельзон вида-индекса во всех известных разрезах. В разрезе Анабарской губы Arctotis (Praearctotis) milovae (Okuneva) встречаются совместно с Zugodactylites braunianus (Orb.) и Pseudolioceras lythense (Y. et B.), в разрезе р. Марха вид встречается совместно с Zugodactylites braunianus (Orb.). На Северо-Востоке России в разрезе р. Старт близкие к Arctotis (Praearctotis) milovae (Okuneva) формы обнаружены автором в осыпи слоев с Pseudolioceras compactile (Simps.), Porpoceras vortex (Simps.), Collina gemma Bonarelli. В Восточном Забайкалье в бассейне р. Газимур Arctotis (Praearctotis) milovae (Okuneva) приводятся из отложений выше уровня с Pseudolioceras rosenkrantzi A. Dagis (Окунева, 2002). Поскольку местная зона Peronoceras spinatum на Северо-Востоке России приравнена к зоне бореального стандарта Pseudolioceras compactile (Репин, 2016, 2017), а положение зоны Pseudolioceras rosenkrantzi относительно зоны Pseudolioceras compactile не является общепринятым (Князев и др., 2003; Репин, 2016), оксито-зона Praearctotis milovae достоверно отвечает зонам Zugodactylites braunianus, Pseudolioceras compactile бореального стандарта (Шурыгин и др., 2011) (рис. 1).
Корреляция. Оксито-зона Praearctotis milovae соответствует верхней части зоны Dacryomya inflata, Tancredia bicarinata и зоне Pseudomytiloides marchaensis зональной шкалы бореального стандарта по двустворчатым моллюскам (Шурыгин и др., 2011) и по находкам вида-индекса коррелируется с местной зоной Pseudolioceras rosenkrantzi Дальнего Востока (Окунева, 2002).
Оксито-зона Praearctotis marchaensis
Номенклатура. В установленном хроностратиграфическом объеме выделяется впервые. В качестве зонального подразделения Восточной Сибири зона Arctotis marchaensis была установлена в объеме нижней части нижнего аалена (Шурыгин, 1987). Согласно аналитической сводке по стратиграфии юры Сибири, зона Arctotis marchaensis охватывает часть верхнего тоара и нижнюю часть аалена, т.е. верхняя граница тоара (граница нижней и средней юры) проходит внутри зоны (Шурыгин и др., 2000). В настоящей работе оксито-зона Praearctotis marchaensis сопоставляется с зоной Pseudolioceras wuerttenbergeri бореального стандарта. Ввиду ревизии вида Arctotis marchaensis (Petrova) объем оксито-зоны Arctotis marchaensis изменен по сравнению с объемом стандартной b-зоны Arctotis marchaensis (Шурыгин и др., 2000).
Вид-индекс: Arctotis (Praearctotis) marchaensis (Petrova, 1947).
Стратотип оксито-зоны: Восточная Сибирь, Анабарская губа, обн. 5, слои 80–88, эренская свита, мощность 27.4 м.
Зональный комплекс двустворчатых моллюсков: Oxytoma ex gr. jacksoni Pomp., Luciniola sp.
Принцип выделения границ и обоснование возраста. Нижняя граница окито-зоны Praearctotis marchaensis устанавливается по появлению вида-индекса. Верхняя граница устанавливается по появлению вида Arctotis (Praearctotis) similis Velikzh.
Возраст оксито-зоны относительно аммонитовой шкалы определяется суммой тельзон вида-индекса во всех известных разрезах. В разрезе Анабарской губы вид-индекс занимает положение в средней части эренской свиты выше уровня с Zugodactylites braunianus (Orb.) и ниже уровня с Pseudolioceras falcodiscus (Quenstedt) (Князев и др., 1991, 2003). В разрезе р. Марха вид Arctotis (Praearctotis) marchaensis (Petr.) занимает положение в разрезе ниже и выше уровня с Pseudolioceras wuerttenbergeri (Denckmann). Достоверно оксито-зона Praearctotis marchaensis отвечает только зоне Pseudolioceras wuerttenbergeri бореального стандарта (Шурыгин и др., 2011) (рис. 1).
Корреляция. Оксито-зона Praearctotis marchaensis соответствует нижней части b-зоны Arctotis marchaensis зональной шкалы бореального стандарта по двустворчатым моллюскам (Шурыгин и др., 2011). На Северо-Востоке России в разрезе р. Летняя из отложений, отнесенных к нижней половине зоны Pseudolioceras danilovi, приводились Arctotis aff. marchaensis (Petг.), Lenoceramus sp. и Camptonectes sp. (Репин, Полуботко, 1993; Полуботко, Репин, 1994). Оксито-зона Praearctotis marchaensis условно отвечает нижней половине местной аммонитовой зоны Pseudolioceras danilovi (Репин, 2017) (рис. 1).
Оксито-зона Praearctotis similis
Номенклатура. Выделяется впервые.
Вид-индекс: Arctotis (Praearctotis) similis Velikzhanina, 1966.
Стратотип оксито-зоны: Восточная Сибирь, мыс Цветкова, обн. 5, слои 2–5, короткинская свита, мощность 67 м.
Зональный комплекс оксито-зоны составляют виды двустворчатых моллюсков: Propeamussium olenekense (Bodyl.), Dacryomya gigantea Zakh. et Schur., Mclearnia kelimyarensis Zakh. et Schur., “Malletia amygdaloides (Sow.)”, “Nuculana acuminata (Goldf.)”, Luciniola sp.
Принцип выделения границ и обоснование возраста. Нижняя граница оксито-зоны Praearctotis similis устанавливается по появлению вида-индекса, верхняя граница – по появлению вида-индекса вышележащей оксито-зоны – Arctotis (Arctotis) tabagensis (Petr.).
Возраст оксито-зоны относительно аммонитовой шкалы определяется суммой тельзон вида-индекса во всех известных разрезах. В разрезе Анабарской губы Arctotis (Praearctotis) similis распространен в хоргонской свите, в осыпи слоев которой был найден аммонит Pseudolioceras sp. (cf. maclintocki Haugh.) (Стратиграфия…, 1976), переопределенный как Pseudolioceras falcodiscus (Quenstedt) (Князев, 1991). В разрезе мыса Цветкова (Восточный Таймыр), по данным автора, вид-индекс занимает положение ниже и выше уровня с Pseudolioceras cf. falcodiscus (Quenstedt). В разрезе р. Моторчуна представители Arctotis (Praearctotis) similis Velikzh. распространены в верхах сунтарской свиты на уровне находок Pseudolioceras falcodiscus (Quenstedt) (Князев и др., 1991), Pseudolioceras motortschunense Repin (Репин, 2017) и ниже уровня с Pseudolioceras maclintocki (Haught.) (Стратиграфия…, 1976). Достоверно оксито-зона Praearctotis similis отвечает зоне Pseudolioceras falcodiscus бореального стандарта (Шурыгин и др., 2011) (рис. 1).
Корреляция. Оксито-зона Praearctotis similis соответствует верхней части b-зоны Arctotis marchaensis и нижней части b-зоны Mclearnia kelymiarensis зональной шкалы бореального стандарта по двустворчатым моллюскам (Шурыгин и др., 2011). На Северо-Востоке России в разрезе р. Вилига из отложений с Pseudolioceras replicatum приводились Arctotis cf. marchaensis (Petг.), Propeamussium olenekense (Bodyl.), Camptonectes sp., Malletia ex gr. amygdaloides (Sow.) (Репин, Полуботко, 2015). Оксито-зона условно соответствует верхам местной аммонитовой зоны Pseudolioceras danilovi, местным аммонитовым зонам Pseudolioceras paracompactile и Pseudolioceras replicatum Северо-Востока России (Репин, 2017) (рис. 1).
Оксито-зона Arctotis tabagensis
Номенклатура. Выделяется впервые.
Вид-индекс: Arctotis (Arctotis) tabagensis (Petrova, 1953).
Стратотип оксито-зоны: Восточная Сибирь, мыс Цветкова, обн. 5, слои 6–8; апрелевская свита, слой 9–арангастахская свита, мощность 67 м.
Зональный комплекс оксито-зоны составляют виды двустворчатых моллюсков: Arctica humiliculminata Schur., Astarte meeki Stant., Musculus sp., Retroceramus elegans Kosch., Tancredia sp.
Принцип выделения границ и обоснование возраста. Нижняя граница оксито-зоны Arctotis tabagensis устанавливается по появлению вида-индекса, верхняя граница – по появлению вида Arctotis (Arctotis) sublaevis (Bodyl.) – индекса вышележащих слоев. Возраст окито-зоны относительно аммонитовой шкалы определяется суммой тельзон вида-индекса во всех известных разрезах.
На мысе Цветкова (Восточный Таймыр) Arctotis (Arctotis) tabagensis (Petr.) раcпространен выше уровня с Pseudolioceras cf. falcodiscus. В разрезе Анабарской губы этот вид встречен в конденсированном слое в основании арангастахской свиты соместно с Retroceramus elegans Kosch. и Retroceramus jurensis Kosch. ниже уровня с Pseudolioceras (Tugurites) whiteavesi (White) и P. (T.) fastigatum (Меледина, Шурыгин, 2000). В разрезе р. Моторчуна он встречен в основании сюнгюдинской свиты на уровне с Pseudolioceras maclintocki (Haught.) (Стратиграфия…, 1976). Оксито-зона Arctotis tabagensis достоверно сопоставляется с зоной Pseudolioceras maclintocki бореального стандарта (Шурыгин и др., 2011) (рис. 1).
Корреляция. Оксито-зона Arctotis tabagensis соответствует верхней части b-зоны Maclearnia kelymiarensis и нижней части b-зоны Retroceramus elegans зональной шкалы бореального стандарта по двустворчатым моллюскам (Шурыгин и др., 2011). На Северо-Востоке России в разрезе рр. Акачан, Аллах-Юнь из толщи песчаников и алевролитов с ранеааленским Pseudolioceras maclintocki (Haught) приводились “Arctotis marchaensis (Petr.)”, “Arctotis aff. similis Velikzh.”, где они указаны совместно с Propeamussium olenekense (Bodyl.), “Mytiloceramus priscus Sey” (Решения…, 1994). Оксито-зона условно соответствует местной зоне Pseudolioceras beyrichi orientale и нижней части местной зоны Pseudolioceras maclintocki Северо-Востока России (Репин, 2017) (рис. 1).
ОПИСАНИЕ ЗОНАЛЬНЫХ ВИДОВ
Arctotis (Praearctotis) marchaensis (Petгova, 1947)
Табл. II, фиг. 1–13
Pseudomonotis (Eumorphotis) marchaensis Petrova: Атлас…, 1947, с. 123, табл. XII , фиг. 13–15 .
Pseudomonotis (Eumorphotis) vai Bodylevsky: Атлас…, 1947, с. 124, табл. XII , фиг. 20–23 .
Eumorphotis marchaensis: Крымгольц и др., 1953, с. 40, табл. IV , фиг. 1–4.
Eumorphotis sparsicosta: Крымгольц и др., 1953, табл. V , фиг. 7а, 7б, 7в, 8 (только).
Arctotis marchaensis: Кошелкина, 1963, с. 119, табл. II, фиг. 4а–4в; Великжанина, 1966, с. 86, табл. I , фиг. 15 , табл. II, фиг. 1–8; Полевой…, 1968, табл. 40 , фиг. 5; Князев и др., 2003, с. 90, табл. 6 , фиг. 5, 7, 10 .
Meleagrinella buschinskii Koschelkina: Кошелкина, 1963, с. 116, табл. II, фиг. 3а, 3б.
Arctotis viluiensis Velikzhanina: Великжанина, 1966, с. 89–90, табл. III, фиг. 1–6 (только).
Голотип: ЦНИГР музей им. акад. Ф.Н. Чернышева, Санкт-Петербург, экз. № 574/5393, коллекция А.Г. Ржонсницкого (1915 г.), изображен в работах: Атлас…, 1947, с. 123, табл. XII , фиг. 14 ; Крымгольц и др., 1953, с. 40, табл. IV , фиг. 2; настоящая работа, табл. II, фиг. 1; Восточная Сибирь, р. Марха. Нижняя юра, верхний тоар.
Материал. Более 100 целых раковин и отдельных створок из сунтарской свиты разреза р. Марха; типовая коллекция вида “Pseudomonotis (Eumorphotis) marchaensis ” с р. Марха, около 100 деформированных раковин и отдельных створок из эренской свиты Анабарского залива; ядра левых створок из верхнего тоара р. Сатурн; около 10 левых и правых створок хорошей сохранности из скважин Тенкеляхского участка; типовая коллекция вида “Pseudomonotis (Eumorphotis) vai” с п‑ова Юрюнг-Тумус В.И. Бодылевского; коллекция Б.Н.Шурыгина – около 20 ядер из разреза п‑ова Юрюнг-Тумус; типовая коллекция вида “Arctotis viluiensis” с р. Вилюй Т.И. Кириной.
Диагноз. Раковина чаще средней величины. Тип переднего крыла “параболовидный”. Переднее крыло тупоугольное, узкое или очень узкое. Переднее залигаментное поле левой створки короткое. Левая створка сильновыпуклая или вздутая, правая створка слабовыпуклая. Радиальные ребра двух порядков в количестве до 45, межреберные промежутки узкие. Заднее крыло левой створки покрыто тонкими радиальными ребрами в количестве до 15. Лигаментная площадка левой створки расположена под небольшим наклоном к плоскости смыкания створок. Угловатый подтип лигаментной площадки правой створки. Ложечковидное углубление занимает меньшую часть лигаментной ямки.
Размеры, мм и отношения: см. табл. 1.
Таблица 1.
Размеры, мм и отношения
| Колл. №, створка | Местонахождение | Д | В | В/Д | Вп | Вп/В | ДПЧ | ДПЧ/ Д |
ДПП | ДПП/ ДПЧ |
УПКр | Р | Р/Д | ШП/ ШР |
ШПКр/ ДПКр |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 574/5393 л, голотип |
р. Марха, верхний тоар, музей ЦНИГР, СПб., колл. А.Г. Ржонсницкого | 24.07 | 26.0 | 1.08 | 9.36 | 0.36 | 10.63 | 0.44 | 3.68 | 0.35 | 132° | 32 | 1.33 | 1.10 | 0.17 |
| 574/5393 п, типовой экземпляр |
То же | 21.2 | 19.0 | 0.90 | – | – | 11.9 | 0.56 | 6.9 | 0.58 | – | – | – | – | – |
| МХ-111/2 л | р. Марха, обн. 4, осыпь слоев 2–3, обр. 111, верхний тоар | 20.70 | 19.6? | 0.95? | 6.0 | 0.31 | 10.20 | 0.49 | 3.60 | 0.34 | 126° | 26 | 1.26 | 1.06 | 0.15 |
| МХ-111/3 л | То же | 30.41 | 31.6 | 1.09 | 13.17 | 0.42 | 14.52 | 0.48 | – | – | 121° | 45 | 1.55 | 1.23 | 0.16 |
| МХ-220/6 л | р. Марха, обн. 5, осыпь, обр. 220, верхний тоар | 38.02 | 34.1 | 0.90 | 13.22 | 0.39 | 16.38 | 0.43 | 6.52 | 0.40 | 127° | 41 | 1.10 | 1.64 | 0.09 |
| МХ-233/1 л | р. Марха, обн. 10, слой 8, обр. 233, верхний тоар | 35.00 | 39.1 | 1.12 | – | – | 16.3 | 0.47 | – | – | 130° | 43 | 1.23 | 0.92 | 0.15 |
| МХ-233/2 л | То же | 38.25 | 40.5 | 1.06 | 18.97 | 0.47 | 17.57 | 0.46 | 7.28 | 0.41 | 119° | 44 | 1.17 | 0.53 | 0.15 |
| ТХ-1/5 л | Тенкеляхская площадь, линия 1080, скважина 350, глубина 35 м | 21.8 | 22.6 | 1.04 | 6.5 | 0.29 | 9.5 | 0.44 | 3.81 | 0.40 | 144° | 37 | 1.70 | 1.49 | 0.17 |
| ТХ-1/2 п | То же | 20.55 | 18.9 | 0.92 | 2.8 | 0.15 | 9.02 | 0.50 | 5.04 | 0.56 | – | 33 | 1.62 | 1.50 | – |
| АГ-37/1 л | Анабарская губа, обнажение 4, слой 18, верхний тоар | 39.08 | 35.7 | 0.95 | – | – | 16.72 | 0.43 | – | – | 135° | 34 | 0.90 | 1.28 | 0.20 |
| АГ-37/2 п | То же | 36.27 | 34.7 | 0.96 | – | – | 16.30 | 0.45 | – | – | – | 28 | 0.77 | – | – |
| 7/306 л, голотип “Pseudomonotis (Eumorphotis) vai” |
п-ов Юрюнг-Тумус, верхний тоар, музей Горного института, СПб., колл. В.И. Бодылевского | 55.48 | 59.2 | 1.06 | 17.36 | 0.29 | 21.21 | 0.38 | – | – | 119° | 30 | 0.87 | 1.03 | 0.13 |
| 8/306 п, типовой экземпляр | То же | 43.52 | 40.18 | 0.92 | – | – | 21.46 | 0.49 | 12.57 | 0.59 | – | – | – | – | – |
| Н-73/ 2913/1 л | п-ов Юрюнг-Тумус, верхний тоар, музей ИНГиГ, Новосибирск, колл. Б.Н. Шурыгина | 42.06 | 46.3 | 1.10 | 14.27 | 0.31 | 18.84 | 0.45 | 7.66 | 0.41 | 144° | 22 | – | 2.14 | 0.15 |
| Н-73/ 2913/1 п | То же | 37.80 | 36.20 | 0.96 | 4.11 | 0.11 | 17.48 | 0.46 | – | – | – | – | – | – | – |
| №20/653, голотип “Arctotis viluiensis” | р. Вилюй, обн. 54-56, обр. 1394, верхний тоар, музей ВНИГРИ, колл. Т.И. Кириной | 39.33 | 36.2 | 0.92 | 13.89 | 0.38 | 18.33 | 0.47 | 6.78 | 0.37 | 135° | 35 | 0.89 | 1.13 | 0.17 |
Изменчивость. Размеры экземпляров из разрезов р. Марха, Анабарской губы, скважин Тенкеляхской площади преимущественно средние, встречаются маленькие (юношеские) экземпляры менее 30 мм. Высота варьирует от 19.6 до 40.5 мм. У голотипа вида “Praearctotis vai” (табл. II, фиг. 11 ), происходящего из разреза п-ова Юрюнг-Тумус, размеры большие – высота 59.2 мм, что не является характерным для описываемого таксона. В выборках из типового местонахождения вида “Praearctotis vai” преобладают раковины средних размеров до 46.0 мм, близкие по размерам к мархинским. Возможно, только отдельные экземпляры достигали больших размеров, их высота превышала пределы диапазона изменчивости этого признака.
Контуры левых створок изменчивы от очень низких до очень высоких. Коэффициент В/Д варьирует от 0.90 до 1.12. Относительно оси симметрии левая створка изменчива от умеренно неравносторонней до равносторонней. Коэффициент ДПЧ/Д варьирует от 0.43 до 0.48. На контуры и симметрию раковин, предположительно, влияла плотность колоний двустворчатых моллюсков на занимаемых ими локальных участках. Контур и симметрия раковины вариабельны и не рассматриваются как видоспецифичные признаки.
Левые створки изучавшихся экземпляров из разрезов р. Марха, Анабарской губы, скважин Тенкеляхской площади слабо ребристые. Коэффициент Р/Д варьирует от 1.06 до 1.90. У голотипа вида Praearctotis vai коэффициент Р/Д равен 1.03. По данному значению признака эти экземпляры близки. Плотность ребристости раковины коррелятивно связана с составом отложений. Предположительно, состояние признака зависит от удаленности обитания организмов относительно берега и глубины. У раннетоарских форм из более глинистых частей разреза – Arctotis (Praearctotis) subsriata (Muenster), Arctotis (Praearctotis) milovae (Okuneva) – коэффициент ребристости выше, чем у описываемого таксона.
Ширина межреберных промежутков коррелятивно связана с толщиной ребер и степенью ребристости. Коэффициент ШП/ШР у экземпляров из разрезов р. Марха, Анабарской губы, скважин Тенкеляхской площади варьирует от 0.53 до 1.64. Для таксона характерны узкие межреберные промежутки и широкие радиальные ребра. У голотипа вида “Praearctotis vai” коэффициент ШП/ШР равен 1.03. По данному значению признака эти экземпляры близки. У некоторых экземпляров из типового местонахождения вида “Praearctotis vai” на ядрах межреберные промежутки умеренно широкие, коэффициент ШП/ШР равен 2.14, что, вероятно, объясняется меньшей плотностью радиальных ребер на внутренних ядрах. Предположительно, состояние признака зависит от физических свойств грунта. У описываемого таксона, обитавшего на глинисто-алевритовых грунтах, межреберные промежутки узкие.
Левая створка сильновыпуклая или вздутая. Коэффициент Вп/В варьирует от 0.31 до 0.47. Выпуклость левых створок у таксонов, относящихся к филетической линии рода Arctotis, периодически меняется. Кроме описываемого таксона, сильновыпуклые или вздутые левые створки характерны для раковин батского Arctotis (Arctotis) lenaensis (Lahuzen) и волжского Arctotis (Canadarctotis) intermedia (Bodylevsky). Предположительно, сильновыпуклой или вздутой раковиной обладают формы, живущие в условиях сильного волнового воздействия. Слабая плотность ребристости и сильновыпуклая левая створка являются специфичными для таксонов видового ранга, обитавших в прибрежных мелководных обстановках.
Переднее залигаментное поле левой створки короткое. Коэффициент ДПП/ДПЧ варьирует в пределах 0.35–0.41. Переднее крыло левой створки узкое. Коэффициент ШПКр/ДПКр от 0.09 до 0.20. Переднее крыло тупоугольное. Угол переднего крыла левой створки (УПКр) варьирует от 121° до 144°. Короткое залигаментное поле, узкое переднее крыло и тупой угол переднего крыла левой створки характеризуют “параболовидный” тип переднего крыла. “Параболовидный” тип специфичен для группы позднетоарских видов и для описываемого таксона.
Сравнение. По очертаниям, симметрии, углу переднего крыла левой створки вид наиболее близок к голотипу Arctotis (Praearctotis) simisis Velikzhanina, 1966 (Великжанина, 1966, с. 90–92, табл. II, фиг. 9 ). Отличается более узким передним крылом, коротким передним залигаментным полем левой створки, угловатым подтипом лигаментной ямки, более узкими межреберными промежутками.
Замечания. По типу переднего крыла, угловатому подтипу лигаментной площадки голотип Prearctotis marchaensis из верхнего тоара Лено-Вилюйского района (р. Марха) (Атлас…, 1947, с. 123, табл. XII , фиг. 14 ; настоящая работа, табл. II, фиг. 1) и голотип “Pseudomonotis (Eumorphotis) vai” из верхнего тоара Нордвикского района (п‑ов Юрюнг-Тумус) (Атлас…, 1947, с. 124, табл. XII , фиг. 20 ; настоящая работа, табл. II, фиг. 11 ) относятся к одной группе. Вид В.И. Бодылевского предварительно рассматривался как самостоятельный вид в составе подрода Arctotis (Praearctotis) (Лутиков, Шурыгин, 2010). Поскольку размеры и ширина межреберных промежутков на левой створке вариабельны в выборках и не являются видоспецифичными, “Arctotis (Praearctotis) vai” в настоящей работе рассматривается как младший синоним Arctotis (Praearctotis) marchaensis.
По типу переднего крыла, выпуклости левой створки голотип Arctotis (Praearctotis) marchaensis из верхнего тоара Лено-Вилюйского района (р. Марха) и голотип “Arctotis viluiensis ” из верхнего тоара Лено-Вилюйского района (р. Вилюй) (Великжанина, 1966, с. 89–90, табл. III, фиг. 1; настоящая работа, табл. III, фиг. 1) относятся к одной группе. Вид Л.С. Великжаниной предварительно рассматривался как самостоятельный вид (Лутиков, Шурыгин, 2010). Поскольку значения выпуклости и длины переднего залигаментного поля левой створки у голотипов видов Г.Т. Петровой и Л.С. Великжаниной почти идентичны, вид “Arctotis viluiensis” рассматривается как младший синоним Arctotis (Praearctotis) marchaensis.
Из Лено-Вилюйского района (р. Марха) известен вид Meleagrinella buschinskii (Кошелкина, 1963, с. 38, табл. II, фиг. 3). Типовая коллекция Г.И. Бушинского, включая голотип, происходит из ожелезненных песчаников сунтарской свиты с Arctotis (Praearctotis) marchaensis. По типу переднего крыла, очертаниям створок, выпуклости и скульптуре вид З.В. Кошелкиной близок к Arctotis (Praearctotis) marchaensis и, очевидно, является его младшим синонимом.
Геологический возраст и географическое распространение. Верхний т-оар, оксито-зона Praearctotis marchaensis. Западная и Восточная Сибирь, Северо-Восток России.
Arctotis (Praearctotis) similis Velikzhanina, 1966
Табл. III, фиг. 1–8
Arctotis similis Velikzhanina: Великжанина, 1966, с. 89, табл. II, фиг. 9–10 (только).
Arctotis viluiensis Velikzhanina: Великжанина, 1966, с. 89, табл. III, фиг. 7 (только).
Pseudomonotis aff. elegans: Пчелинцев, 1933, с. 47, табл. III, рис. 45 .
Meleagrinella cf. echinata (Smith): Wierzbowski, 1981, p. 219, pl. 8, figs. 6–8.
Arctotis (Praearctotis) viluiensis: Лутиков, Шурыгин, 2010, табл. I , фиг. 8; табл. II, фиг. 4; табл. III, фиг. 23–26 .
Praearctotis viluiensis: Лутиков, Тёмкин, Шурыгин, 2010, табл. I , фиг. 12 .
Голотип: экз. № 14/653, левая створка. Музей ВНИГРИ, Санкт-Петербург, коллекция Т.И. Кириной (1961 г.). Изображен: Великжанина, 1966, табл. II, фиг. 9 ; настоящая работа, табл. III, фиг. 2; Восточная Сибирь, р. Вилюй. Нижняя юра, верхний тоар.
Материал. Около 10 левых и правых створок хорошей сокранности из верхней пачки короткинской свиты (пачка 18) разреза мыса Цветкова (Восточный Таймыр); 5 левых, 2 правых створки, несколько ядер и отпечатков левых и правых створок из ракушников хоргонской свиты разреза Анабарской губы; около 10 левых и правых створок хорошей сохранности из верхней части сунтарской свиты р. Моторчуны; типовая коллекция вида “Arctotis similis” из сунтарской свиты разреза р. Вилюй Т.И. Кириной.
Диагноз. Раковина чаще средней величины. Тип переднего крыла “трапециевидный”. Переднее крыло тупоугольное, умеренно широкое. Переднее залигаментное поле левой створки длинное или очень длинное. Левая створка умеренно выпуклая, правая створка плоская. Радиальные ребра двух порядков в количестве от 25 до 40, межреберные промежутки чаще очень широкие. Заднее крыло левой створки покрыто тонкими радиальными ребрами в количестве до 16. Лигаментная площадка левой створки расположена под большим наклоном к плоскости смыкания створок. Ложечковидный подтип лигаментной ямки. Ложечковидное углубление занимает бóльшую часть лигаментной ямки. Лигаментная площадка правой створки длинная.
Размеры, мм и отношения: см. табл. 2.
Таблица 2.
Размеры, мм и отношения
| Колл. №, створка | Местонахождение | Д | В | В/Д | Вп | Вп/В | ДПЧ | ДПЧ/ Д |
ДПП | ДПП/ ДПЧ |
УПКр | Р | Р/Д | ШП/ ШР |
ШПКр/ ДПКр |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| МН-216/1 л | р. Моторчуна, обн. 4, слой 2, верхний тоар | 32.81 | 31.5 | 0.96 | 8.0 | 0.25 | 14.52 | 0.44 | 6.35 | 0.44 | 146° | 26 | 0.79 | 2.02 | 0.16 |
| МН-216/2 п | То же | 30.69 | 24.6 | 0.80 | 2.0 | 0.08 | 15.5 | 0.50 | 8.3 | 0.54 | 115° | 25 | 0.81 | 3.28 | 0.20 |
| ТР-19/1 л | Восточный Таймыр, обн. 5, слой 3, нижняя юра, верхний тоар | 34.71 | 30.6 | 0.88 | – | – | 17.02 | 0.49 | 9.10 | 0.53 | 136° | 25 | 0.72 | 4.79 | 0.22 |
| ф-113/1 л | Восточный Таймыр, обн. 5, слой 5, обр. № ф-113, нижняя юра, верхний тоар | 40.00 | – | – | – | – | – | – | – | – | 115° | – | – | 1.14 | – |
| АГ-114/1 л | Анабарская губа, западный берег, обн. 5, слой 89, верхний тоар | 31.28 | 28.4 | 0.91 | – | – | 13.25 | 0.42 | 6.27 | 0.47 | 112° | 40 | 1.28 | 1.24 | 0.16 |
| АГ-114/2 п | То же | 33.95 | 29.0 | 0.93 | – | – | 13.0 | 0.42 | 5.88 | 0.45 | – | 40 | 1.28 | – | – |
| № 14/653, голотип | р. Вилюй, обн. 54, обр. 1953, верхний тоар, музей ВНИГРИ, колл. Т.И. Кириной | 42.96 | 41.6 | 0.97 | 12.54 | 0.30 | 18.84 | 0.44 | 10.20 | 0.54 | 131° | 40 | 0.93 | 2.03 | 0.20 |
Изменчивость. Размеры левых створок у экземпляров из разрезов р. Моторчуны, мыса Цветкова (Восточный Таймыр) и Анабарской губы маленькие или средние. Высота левой створки варьирует от 28.4 до 31.5 мм.
Контуры левых створок изменчивы от очень низких до равновеликих. Коэффициент В/Д варьирует от 0.88 до 0.96. У экземпляров из разреза р. Вилюй (колл. Т.И. Кириной) коэффициент В/Д равен 0.99. Относительно оси симметрии левая створка изменчива от умеренно неравносторонней до равносторонней. Коэффициент ДПЧ/Д варьирует от 0.42 до 0.49.
Левые створки у экземпляров из разрезов р. Моторчуны и р. Вилюй, мыса Цветкова (средняя часть короткинской свиты) редко ребристые. Коэффициент ребристости (Р/Д) варьирует от 0.72 до 0.99. У экземпляров из разреза Анабарской губы (хоргонская свита) (табл. III, фиг. 7, 8) плотность ребристости выше – до 1.28. Межреберные промежутки у экземпляров из разрезов р. Моторчуны, мыса Цветкова и р. Вилюй от умеренно широких до очень широких. Коэффициент ШП/ШР варьирует от 2.02 до 4.79. У экземпляров из разреза Анабарской губы (хоргонская свита) и одного экземпляра из разреза мыса Цветкова (верхняя часть короткинской свиты) (табл. III, фиг. 4) межреберные промежутки узкие, коэффициент ШП/ШР варьирует от 1.14 до 1.24. По густоте ребристости и ширине межреберных промежутков экземпляры из разреза Анабарской губы и экземпляр из верхней пачки короткинской свиты разреза мыса Цветкова близки к Arctotis (Arctotis) tabagensis, но отличаются типом переднего крыла. Возможно, при дальнейших исследованиях эти формы будут обособлены в пределах одной филогенетической линии, как таксон, занимающий промежуточное положение между Arctotis (Praearctotis) similis и Arctotis (Arctotis) tabagensis.
Левая створка умеренно выпуклая. Коэффициент выпуклости (Вп/В) измерялся только у одного экземпляра и составляет 0.25.
Переднее залигаментное поле левой створки длинное. Коэффициент ДПП/ДПЧ варьирует в пределах 0.44–0.54. Переднее крыло левой створки изменчиво от узкого до умеренно широкого. Коэффициент ШПКр/ДПКр варьирует от 0.16 до 0.22. Переднее крыло тупоугольное. Угол переднего крыла левой створки (УПКр) варьирует в пределах 112°–146°.
Длинное залигаментное поле, узкое или умеренно широкое переднее крыло, тупой угол переднего крыла левой створки характеризуют “трапециевидный” тип переднего крыла. “Трапециевидный” тип специфичен только для описываемого таксона.
Сравнение. По ложечковидному подтипу лигаментной ямки правой створки, выпуклости левой створки описываемый вид наиболее близок к Arctotis (Arctotis) tabagensis (Petrova, 1966) (Крымгольц и др., 1953, с. 89–90, табл. IV , фиг. 6–7). Отличается более длинной лигаментной площадкой правой створки, тупоугольным передним крылом, широкими межреберными промежутками. От Arctotis frenguellii (Damborenea et al., 2013, p. 124, fig. 6.1 p ) из тоар-ааленских отложений Аргентины отличается узким передним крылом (у аргентинского вида ШПКр/ДПКр 0.26), значительно меньшим количеством ребер.
Замечания. Л.С. Великжаниной при описании вида similis в качестве голотипа был выбран экземпляр, представленный левой створкой (Великжанина, 1966, табл. II, фиг. 9 ), происходящий из пограничных тоар-ааленских отложений (сунтарской свиты) р. Вилюй. Левые створки наших экземпляров из верхних частей тоара Жиганского, Лено-Анабарского и Восточно-Таймырского районов относятся к виду similis на основании внешнего сходства с голотипом по очертаниям переднего крыла.
По наличию слабовыраженного протуберанца на левой створке, установленного на экземплярах из короткинской свиты разреза мыса Цветкова (табл. I , фиг. 8) и из сунтарской свиты разреза р. Моторчуны, группу similis следует относить к подроду Praearctotis. По типу переднего крыла представители группы занимают промежуточное положение между позднетоарской группой marchaensis с узким крылом и ааленской группой tabagensis с широким крылом.
Из пачки песков и железистых песчаников с маломощным конгломератом в основании, расположенной в верхней части сунтарской свиты разреза р. Вилюй, приводились Arctotis similis Ve1ikzh. (Великжанина, 1966, с. 89, табл. II, фиг. 9–11 ) и A. viluiensis Vе1ikzh. (Великжанина, 1966, с. 89, табл. III, фиг. 1–7). Голотип вида Arctotis similis привязан к обнажению 54 на р. Вилюй. Голотип вида “Arctotis viluiensis” указан из обнажений 54–56 на р. Вилюй (Великжанина, 1966; Кирина, 1966). Предварительно оба вида рассматривались как синонимы (Лутиков, Шурыгин, 2010). При изучении оригинальной коллекции Т.И. Кириной, хранящейся в музее ВНИГРИ, установлено, что у голотипа Arctotis similis левая створка умеренно выпуклая, очень длинное переднее залигаментное поле (табл. III, фиг. 2). У голотипа “Arctotis viluiensis” левая створка сильно выпуклая, переднее залигаментное поле короткое (табл. III, фиг. 1).
Согласно описанию, основным отличием этих двух видов Л.С. Великжанина считала различия в скульптуре: у “A. viluiensis” ребра первого порядка очень сильные, четкие, почти в три раза тоньше широких межреберных промежутков, в которых помещаются ребра второго порядка. По этому признаку некоторые наши экземпляры из разрезов Восточного Таймыра и р. Моторчуны (табл. III, фиг. 3, 5) близки к экземпляру “Arctotis viluiensis” с р. Марха (Велижанина, 1966, табл. III, фиг. 7). Другим признаком отличия видов считались волнообразные морщины роста, характерные для Arctotis similis. По этому признаку все наши экземпляры из короткинской свиты Восточного Таймыра, хоргонской свиты Анабарской губы и сунтарской свиты р. Моторчуны (табл. III, фиг. 3–8) близки к Arctotis similis в описании Л.С. Великжаниной (Велижанина, 1966, табл. II, фиг. 9–11 ). У некоторых форм из разреза Восточного Таймыра, происходящих из верхней половины пачки 18, скульптура представлена большим числом радиальных ребер (табл. III, фиг. 4). В разрезе Анабарской губы в хоргонской свите встречаются экземпляры с часто расположенными ребрами (табл. III, фиг. 7, 8). Эти экземпляры имеют узкие межреберные промежутки и предварительно относились к tabagensis (Лутиков, Шурыгин, 2010, табл. IV , фиг. 1–2). Поскольку степень ребристости может зависеть от фаций, этот признак не рассматривается как видоспецифичный. Все морфологические разновидности с тупоугольным и умеренно широким передним крылом левой створки, длинным или очень длинным передним залигаментным полем отнесены к Arctotis (Praearctotis) similis Vе1ikzh. Вид “Arctotis viliuensis” (Великжанина, 1966) включен в синонимику Arctotis (Praearctotis) marchaensis (Атлас…, 1947).
Геологический возраст и географическое распространение. Нижняя юра, верхний тоар, зона Pseudolioceras falcodiscus, оксито-зона Praearctotis similis. Верхний тоар Восточной Сибири, Шпицбергена, Дальнего Востока России, Кавказа.
Arctotis (Arctotis) tabagensis (Petrova, 1953)
Табл. III, фиг. 9–14
Eumorphotis tabagensis Petrova: Крымгольц и др., 1953, с. 89–90, табл. IV , фиг. 6–7.
Arctotis (Arctotis) tabagensis: Лутиков, Шурыгин, 2010, табл. IV , фиг. 3–7.
Голотип: ЦНИГР музей им. акад. Ф.Н. Чернышева, Санкт-Петербург, экз. № 602/5393, ядро левой створки, коллекция Г.А. Иванова (1925 г.). Изображен: Крымгольц и др., 1953, табл. IV , фиг. 6; настоящая работа, табл. III, фиг. 9 ; Восточная Сибирь, р. Лена, Табагинский утес (Верхний Кангаласский Камень). Средняя юра, нижний аален.
Материал. Около десятка целых раковин и несколько ядер левых створок из кыстатымской свиты разрезов рек Сюнгюдэ, Молодо, правая створка и отпечаток левой створки из алевролитов сюнгюдинской свиты разреза р. Моторчуны; около 10 раковин из низов арангастахской свиты Анабарской губы, около 15 ядер из апрелевской свиты Восточного Таймыра, типовая коллекция Г.Т. Петровой с р. Лена, хранящаяся в ЦНИГР музее.
Диагноз. Раковина от средних до очень больших размеров. Тип переднего крыла “гиперболовидный”. Переднее крыло субпрямоугольное, широкое. Переднее залигаментное поле левой створки очень длинное. Левая створка умеренно выпуклая, правая створка плоская. Радиальные ребра трех порядков в количестве от 50 до 60, межреберные промежутки очень узкие. Заднее крыло левой створки покрыто радиальными ребрами в количестве до 20. Лигаментная площадка левой створки расположена под большим наклоном к плоскости смыкания створок. Ложечковидный подтип лигаментной площадки правой створки. Ложечковидное углубление полностью занимает лигаментную ямку. Лигаментная площадка правой створки короткая.
Размеры, мм и отношения: см. табл. 3.
Таблица 3.
Размеры, мм и отношения
| Колл. №, створка | Местонахождение | Д | В | В/Д | Вп | Вп/В | ДПЧ | ДПЧ/ Д |
ДПП | ДПП/ ДПЧ |
УПКр | Р | Р/Д | ШП/ ШР |
ШПКр/ ДПКр |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 602/5393 л голотип | р. Лена, Верхний Кангалласский камень, нижний аален | 49.50 | 48.85 | 0.98 | – | – | 22.42 | 0.45 | – | – | 97° | 60 | 1.28 | – | – |
| АГ-116/1 л | Анабарская губа, западный берег, обн. 5, слой 92, основание, обр. 116, нижний–верхний (?) аален | 46.14 | 47.8 | 1.04 | – | – | 19.68 | 0.43 | 12.62 | 0.64 | 104° | 68 | 1.47 | 1.14 | 0.26 |
| АГ-116/2 п | То же | 40.49 | 38.2 | 0.94 | – | – | 21.91 | 0.54 | 10.20 | 0.47 | – | 38 | 0.94 | 1.20 | – |
| ТХ/160/36-26/1 л | Тенкеляхская площадь, линия 160, скв. 36, гл. 26.0 м, нижний аален | – | 73.2 | – | – | – | 36.37 | – | 19.43 | 0.53 | 90° | 46 | – | 1.25 | 0.23 |
| ф/82-26/1 л ядро | Восточный Таймыр, обн. 7/1, сл. 3, обр. 82, нижний аален | 38.62 | 36.8 | 0.95 | – | – | 17.82 | 0.46 | 9.02 | 0.51 | 89° | 63 | 1.63 | 1.05 | 0.20 |
| ф/82-26/2 л отпечаток | То же | 38.87 | – | – | – | – | – | – | – | – | 89° | 72 | 1.85 | 1.22 | 0.24 |
Изменчивость. Размеры левых створок изучавшихся экземпляров из разрезов р. Лена, Анабарской губы и мыса Цветкова преимущественно маленькие или средние, варьируют в пределах 30.0–50.0 мм. У некоторых экземпляров и скважин Тенкеляхской площади (табл. III, фиг. 14 ) размеры очень большие – более 70.0 мм. Возможно, только отдельные экземпляры достигали больших размеров, их высота превышала пределы диапазона изменчивости этого признака. Формы из скважин Тенкеляхской площади по размерам близки к ааленскому–байосскому таксону Arctotis (Arctotis) sublaevis, но отличаются типом переднего крыла. Возможно, при дальнейших исследованиях эти формы будут обособлены в пределах одной филогенетической линии, как таксон, занимающий промежуточное положение между Arctotis (Praearctotis) similis и Arctotis (Arctotis) sublaevis.
Контуры левых створок изменчивы от очень низких до высоких. Коэффициент В/Д варьирует от 0.95 до 1.04.
Относительно оси симметрии левая створка изменчива от умеренно неравносторонней до равносторонней. Коэффициент ДПЧ/Д варьирует от 0.43 до 0.46.
Левые створки слабо ребристые. Коэффициент ребристости (Р/Д) варьирует от 1.28 до 1.85. Для этого таксона характерно появление интеркалирующих ребер третьего порядка в промежутках между ребрами второго порядка по всей поверхности раковины. Межреберные промежутки у экземпляров из разрезов р. Лена, мыса Цветкова и Тенкеляхского участка очень узкие. Коэффициент ШП/ШР варьирует от 1.05 до 1.25.
Левая створка умеренно выпуклая. Коэффициент выпуклости (Вп/В) измерялся только у одного экземпляра и составляет 0.26.
Переднее залигаментное поле левой створки очень длинное. Коэффициент ДПП/ДПЧ варьирует в пределах 0.51–0.64. Переднее крыло левой створки широкое. Коэффициент ШПКр/ДПКр варьирует от 0.20 до 0.26. Переднее крыло субпрямоугольное. Угол переднего крыла левой створки (УПКр) варьирует в пределах 89°–104°.
Очень длинное залигаментное поле, широкое переднее крыло, субпрямоугольное переднее крыло левой створки характеризуют “гиперболовидный” тип переднего крыла. “Гиперболовидный” тип специфичен только для описываемого таксона.
Сравнение. От Arctotis (Arctotis) sublaevis из верхнего аалена и нижнего байоса Нордвикского, Жиганского, Лено-Анабарского и Восточно-Таймырского районов описываемый вид отличается субпрямоугольным передним крылом левой створки на всех стадиях роста (у вида sublaevis переднее крыло на поздних стадиях роста тупоугольное), плоской правой створкой (у вида sublaevis правая створка вогнутая), открытым биссусным желобком на всех возрастных стадиях (у вида sublaevis биссусный желобок на поздних стадиях роста у крупных раковин зарастает). От Arctotis frenguellii (Damborenea et al., 2013, p. 124, fig. 6.1 p ) из тоар-ааленских отложений Аргентины отличается гиперболовидным типом переднего крыла левой створки (у аргентинского вида – трапециевидный тип переднего крыла).
Замечания. Голотип вида tabagensis представлен ядром левой створки и происходит из якутской свиты ааленского возраста (табл. III, фиг. 9 ). По субпрямоугольной форме переднего крыла наши экземпляры из ааленских отложений Анабарской губы (табл. III, фиг. 10, 12 ) и Восточного Таймыра (табл. III, фиг. 13, 15 ) тождественны с голотипом.
Геологический возраст и географическое распространение. Ааленский ярус, нижний подъярус, зона Pseudolioceras maclintocki, оксито-зона Praearctotis tabagensis, Западная Сибирь, Восточная Сибирь: Анабарская губа, Восточный Таймыр, бассейны рр. Лена, Молодо, Моторчуна, Вилюй, Марха, Тюнг.
ВЫВОДЫ
На основании монографического изучения коллекций двустворчатых моллюсков из тоар-ааленских отложений Восточной Сибири проведена ревизия таксонов двустворчатых моллюсков рода Arctotis Bodylevsky, 1960. Классификация видовых таксонов разработана на основании впервые предложенного автором подхода к классификации окситомид. Классификация таксонов проведена на основе изучения морфогенеза лигаментного и биссусного блоков обеих створок раковины и оценки таксономической значимости признаков наружной морфологии. Установлено, что на фоне постепенного образования морфотипа с развитой ложечкой в филогенетическом ряду Arctotis (Praearctotis)–Arctotis (Arctotis) изменялись состояния других уникальных признаков – длины переднего залигаментного поля, угла и ширины переднего крыла, соотношения выпуклости створок, числа порядков ребристости. В интервале от раннего тоара до раннего аалена постепенно изменялся тип переднего крыла левой створки от “параболовидного” типа к “трапециевидному” типу и затем к “гиперболовидному” типу. Морфогенез лигаментного блока раковины и переднего крыла левой створки у представителей рода Arctotis обнаружил относительно высокие темпы эволюции, приблизительно отвечающие аммонитовым фазам.
Морфогенетическая последовательность видов рода Arctotis Bodylevsky, 1960, установленная в опорных разрезах верхнего тоара–нижнего аалена Анабарской губы и мыса Цветкова (Восточный Таймыр), прослежена и в других разрезах Восточной Сибири (на рр. Моторчуна, Молодо, Марха, Тюнг, Вилюй, в скважинах Вилюйской синеклизы). На основе этой последовательности предложена биохронологическая шкала по окситомидам для верхнего тоара–нижнего аалена. Виды-индексы, представляющие собой отрезки филогенетической линии рода Arctotis Bodylevsky, 1960, характеризуют подразделения шкалы – оксито-зоны. Для разрезов тоара–нижнего аалена выделены четыре оксито-зоны, которые могут быть использованы для биостратиграфической корреляции и хронологической индексации геологических событий наряду с аммонитами: оксито-зона Praearctotis milovae (=зоны Zugodactylites braunianus и Pseudolioceras compactile), оксито-зона Praearctotis marchaensis (=зона Pseudolioceras wuerttenbergeri), оксито-зона Praearctotis similis (=зона Pseudolioceras falcodiscus), оксито-зона Arctotis tabagensis (=зона Pseudolioceras maclintocki). С помощью шкалы проведена региональная корреляция тоар-ааленских разрезов Восточной Сибири, Северо-Востока России и Дальнего Востока.
Проведено соотнесение шкалы с уже введенными в практику аммонитовыми шкалами (Решениe…, 2004; Решения…, 2009). Шкала является параллельной существующим региональным шкалам по двустворчатым моллюскам (Репин, Полуботко, 2004; Шурыгин и др., 2011).
Благодарности. Автор выражает признательность коллегам по совместным полевым работам В.П. Девятову (СНИИГГиМС, Новосибирск), В.Г. Князеву (ИГАБМ СО РАН, Якутск) и В.В. Сапьянику (СНИИГГиМС, Новосибирск), любезно предоставившим дополнительную информацию по литологической и палеонтологической характеристике разрезов. При подготовке данной работы автор получал значимые консультации от Ю.С. Репина (ВНИГРИ, Санкт-Петербург) и М.А. Рогова (ГИН РАН, Москва). Ряд ценных замечаний и рекомендаций, способствовавших улучшению статьи, получены от Б.Н. Шурыгина (ИНГГ СО РАН, Новосибирск), А.С. Бякова (СКВНИИ ДВО РАН, Магадан), В.С. Вишневской (ГИН РАН, Москва) и Ю.Б. Гладенкова (ГИН РАН, Москва). Всем перечисленным специалистам автор выражает искреннюю благодарность за помощь в подготовке статьи. Выражаю особую благодарность моему научному руководителю В.А. Захарову (ГИН РАН, Москва) за важнейшие советы при проведении исследования и оформлении данной статьи.
Источники финансирования. Работа выполнена по теме госзадания ГИН РАН при поддержке гранта РФФИ и Национального центра научных исследований Франции в рамках научного проекта № 21-55-15015.
Список литературы
Атлас руководящих форм ископаемых фаун СССР. Т. 8. Нижний и средний отделы юрской системы. М.–Л.: Госгеолиздат, 1947. 278 с.
Бодылевский В.И. К стратиграфии мезозойских отложений Анабарско-Хатангского района // Проблемы Арктики. 1939. № 10–11. С. 65–67.
Бодылевский В.И. Спорные вопросы стратиграфии юрских и меловых отложений Советской Арктики // Труды Межведомственного совещания по разработке унифицированных стратиграфических схем Сибири 1956 г. Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1957. С. 92–98.
Бодылевский В.И. Новый позднеюрский представитель авикулопектинид Таймыра // Новые виды древних растений и беспозвоночных СССР. Ч. 2. М.: Госгеолтехиздат, 1960. С. 44–45.
Великжанина Л.С. Представители родов Meleagrinella и Arctotis из нижней юры Западной Якутии // Геология и нефтегазоносность Западной Якутии. Л.: Недра, 1966. С. 80–120.
Дарвин Ч. Происхождение видов путем естественного отбора или сохранение благоприятных рас в борьбе за жизнь. Перевод с шестого издания (Лондон, 1872). Санкт-Петербург: Наука, 1991. 546 с.
Захаров В.А. Понятие популяции в палеонтологии морских беспозвоночных // Биология моря. 1975. № 3. С. 3–12.
Захаров В.А., Шурыгин Б.Н. Биогеография, фации и стратиграфия средней юры Советской Арктики (по двустворчатым моллюскам). Новосибирск: Наука, 1978. 206 с.
Захаров В.А., Юдовный Е.Г. Принципы послойной корреляции разрезов ритмичных терригенных толщ (на примере опорного разреза неокома на р. Боярке, Хатангская впадина) // Проблемы палеонтологического обоснования детальной стратиграфии мезозоя Сибири и Дальнего Востока. К Международному коллоквиуму по юрской системе (Люксембург, июль, 1967 г.). Л.: Наука, 1967. С. 28–40.
Захаров В.А., Богомолов Ю.И., Ильина В.И., Константинов А.Г., Курушин Н.И., Лебедева Н.К., Меледина С.В., Никитенко Б.Л., Соболев Е.С., Шурыгин Б.Н. Бореальный зональный стратотип и биостратиграфия мезозоя Сибири // Геология и геофизика. 1997. Т. 38(5). С. 927–956.
Зоны юрской системы в СССР. Отв. ред. Крымгольц Г.Я. Л.: Наука, 1982. 192 с.
Кирина Т.И. Стратиграфия нижнеюрских отложений западной части Вилюйской синеклизы. Л.: Недра, 1966. С. 18–71 (Тр. ВНИГРИ. Вып. 249).
Князев В.Г. Тоарские Harpoceratinae севера азиатской части СССР // Детальная стратиграфия и палеонтология юры и мела Сибири. Новосибирск: Наука, 1991. С. 37–46.
Князев В.Г., Девятов В.П., Шурыгин Б.Н. Стратиграфия и палеогеография ранней юры востока Сибирской платформы. Якутск: ЯНЦ СО АН СССР, 1991. 100 с.
Князев В.Г., Кутыгин Р.В., Девятов В.П., Никитенко Б.Л., Шурыгин Б.Н. Зональный стандарт тоарского яруса Северо-Востока азиатской части России. Якутск: Изд-во СО РАН, 2003. 103 с.
Князев В.Г., Девятов В.П., Мельник О.А. Опорный разрез ааленского и нижней части байосского ярусов Восточной Сибири (бассейн р. Молодо) // Отечественная геология. 2007а. № 5. С. 39–42.
Князев В.Г., Кутыгин Р.Т., Мельник А.О. Биохронологическая шкала тоара–раннего байоса северо-востока Азии по харпоцератинам (Ammonoidea) // Вопросы стратиграфии, палеонтологии и палеогеографии (посвящается 100-летию со дня рождения Г.Я. Крымгольца). СПб.: Изд-во СПб. ун-та, 2007б. С. 77–91.
Кошелкина З.В. Стратиграфия и двустворчатые моллюски юрских отложений Вилюйской синеклизы и Приверхоянского краевого прогиба // Тр. СВКНИИ. 1963. Вып. 5. 219 с.
Красилов В.А. Эволюция и биостратиграфия. М.: Наука, 1977. 256 с.
Крымгольц Г.Я., Петрова Г.Т., Пчелинцев В.Ф. Стратиграфия и фауна морских мезозойских отложений Северной Сибири. М.–Л.: Изд-во Главсевморпути, 1953. 134 с. (Тр. НИИГА. Т. XLV).
Лутиков О.А., Арп Г. Биохронологическая шкала нижнего тоара по двустворчатым моллюскам семейства Oxytomidae Ichikawa, 1958 // Юрская система России: проблемы стратиграфии и палеогеографии. Материалы VIII Всероссийского совещания с международным участием. Онлайн-конференция, 7–10 сентября 2020 г. Отв. ред. Захаров В.А. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 132–141.
Лутиков О.А., Шурыгин Б.Н. Новые данные по систематике юрских и меловых двустворчатых моллюсков семейства Oxytomidae Ichikawa, 1958 // Новости палеонтологии и стратиграфии. 2010. Вып. 14. С. 111–140.
Лутиков О.А., Тёмкин И.Е., Шурыгин Б.Н. Эволюция онтогенезов и филогения некоторых представителей семейства Oxytomidae Ichikawa, 1958 (Mollusca: Bivalvia) // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 4. С. 28–44.
Майр Э. Принципы зоологической систематики. М.: Мир, 1971. 455 с.
Меледина С.В. О зональной шкале тоарского яруса Северной Сибири // Геология и геофизика. 2000. Т. 41. № 7. С. 952–960.
Меледина С.В., Шурыгин Б.Н. Ааленский ярус (средняя юра) Восточной Сибири: зональное расчленение, характерные двустворки и аммоноидеи // Геология и геофизика. 2000. Т. 41. Вып. 2–3. С. 73–85.
Окунева Т.М. Юрская система. Нижний и средний отделы. Разделы: Морские отложения и Моллюски // Атлас фауны и флоры палеозоя и мезозоя Забайкалья. Новосибирск: Наука, Сиб. издат. фирма РАН, 2002. С. 354–387.
Полевой атлас юрской фауны и флоры Северо-Востока СССР. Ефимова А.Ф., Кинасов В.П., Паракецов К.В. и др. Магадан: Магаданское книжное изд-во, 1968. 379 с.
Полуботко И.В. Иноцерамовые двустворки нижней и средней юры Северо-Востока СССР и севера Сибири // Атлас руководящих групп фауны мезозоя Юга и Востока СССР. СПб.: Недра, 1992. 376 с.
Полуботко И.В., Репин Ю.С. Проблема верхнего тоара на Северо-Востоке Азии // Геологическая история Арктики в мезозое и кайнозое. СПб.: ВНИИОкеанология, 1992. С. 41–48.
Полуботко И.В., Репин Ю.С. Зональное расчленение и корреляция тоарских и ааленских отложений Северной Сибири и северо-востока России // Зональные подразделения и межрегиональная корреляция палеозойских и мезозойских отложений России и сопредельных территорий. Кн. 2. Мезозой. СПб.: ВСЕГЕИ, 1994. С. 51–78.
Пчелинцев В.Ф. Фауна верхнего лейаса Кавказа // Тр. Всесоюзного геолого-разведочного общества. 1933. Вып. 253. 35 с.
Репин Ю.С. Аммонитовая шкала нижней юры Северо-Востока Азии // Нефтегазовая геология. Теория и практика. 2016. Т. 11. № 4. С. 1–45.
Репин Ю.С. Род Pseudolioceras Buckman (Ammonoidea) в арктической юре // Нефтегазовая геология. Теория и практика. 2017. Т. 11. № 4. С. 1–46.
Репин Ю.С. Естественные границы отделов юры // Нефтегазовая геология. Теория и практика. 2020. Т. 15. № 1. С. 1–33.
Репин Ю.С., Полуботко И.В. Зональное расчленение верхнего тоара на Северо-Востоке России // Стратиграфия. Геол. корреляция. 1993. Т. 1. № 1. С. 109–117.
Репин Ю.С., Полуботко И.В. Биохронология тоара Арктической палеозоохории // Стратиграфия нефтегазоносных бассейнов России. СПб.: Недра, 2004. С. 93–124.
Репин Ю.С., Полуботко И.В. Стратотипический разрез юры Северного Приохотья: стратиграфия, литология // Нефтегазовая геология. Теория и практика. 2015. Т. 10. № 4. С. 1–42.
Решение 6-го Межведомственного стратиграфического совещания по рассмотрению и принятию уточненных стратиграфических схем мезозойских отложений Западной Сибири. Ред. Гурари Ф.Г. Новосибирск: СНИИГГиМС, 2004. 114 с.
Решения и труды 3-го Межведомственного регионального стратиграфического совещания по мезозою и кайнозою Средней Сибири. Новосибирск: СНИИГГиМС, 1981. 91 с.
Решения 4-го Межведомственного регионального стратиграфического совещания по докембрию и фанерозою юга Дальнего Востока и Восточного Забайкалья. Ред. Турбин М.Т. Хабаровск: ХГГГП, 1994. 124 с.
Решения 3-го Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России, Санкт-Петербург, 2002. СПб.: ВСЕГЕИ, 2009. 266 с.
Руженцев В.Е. Принципы систематики, система и филогения палеозойских аммоноидей // Тр. Палеонтол. ин-та АН СССР. 1960. Т. 83. 331 с.
Симпсон Дж.Г. Принципы таксономии животных. Пер. с англ. под ред. Шаталкина A.M. М.: Товарищество научных изданий КМК, 2006. 293 с.
Стратиграфия юрской системы севера СССР. М.: Наука, 1976. 436 с.
Степанов Д.Л., Месежников М.С. Общая стратиграфия (Принципы и методы стратиграфических исследований). Л.: Недра, 1979. 423 с.
Черных В.В. Основы зональной биохронологии. Екатеринбург: ИГГ УрО РАН, 2016. 268 с.
Шурыгин Б.Н. Стратиграфический объем и положение в шкале нижней и средней юры севера Сибири зон по двустворкам // Геология и геофизика. 1987. № 11. С. 3–11.
Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П., Ильина В.И., Меледина С.В., Гайдебурова Е.А., Дзюба О.С., Казаков А.М., Могучева Н.К. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск: Изд-во СО РАН, 2000. 476 с.
Шурыгин Б.Н., Девятов В.П., Захаров В.А., Князев В.Г., Ильина В.И., Меледина С.В., Никитенко Б.Л., Гриненко В.С. Стратиграфия юры Восточной Сибири (состояние изученности, основные проблемы и способы их решения) // Вестник Госкомгеологии: Материалы по геологии и полезным ископаемым Республики Саха (Якутия). 2001. № 1. С. 112–139.
Шурыгин Б.Н., Никитенко Б.Л., Меледина С.В., Дзюба О.С., Князев В.Г. Комплексные зональные шкалы юры Сибири и их значение для циркумарктических корреляций // Геология и геофизика. 2011. Т. 52. № 8. С. 1051–1074.
Crame J.A. New Late Jurassic oxytomid bivalves from the Antarctic Peninsula region // British Antarctic Surv. Bull. 1985. № 69. P. 35–55.
Crame J.A., Pirrie D., Crampton J. P., Duante A.M. Stratigraphy and regional significance of the Upper Jurassic–Lower Cretaceous Byers Group, Livingston Island, Antarctica // J. Geol. Soc. London. 1993. V. 50. P. 1075–1087.
Damborenea S.E. Circum-Pacific correlation of Argentine Early and Middle Jurassic bivalve zones // 3rd. Int. Symp. Jurassic Stratigr., Poitiers, 1991. Eds. Cariou E., Hanztpercgue P. Geobios. 1994. V. 27. P. 141–147.
Damborenea S.E., Echevarría J., Ros-Franch S. Southern Hemisphere Palaeobiogeography of Triassic-Jurassic Marine Bivalves. Dordrecht, Heidelberg, N.Y., London: Springer, 2013. 139 p.
Dean W., Donovan D., Howarth M. Liassic ammonite zones and subzones of the North-West province // Bull. Brit. Mus. (Natur. History) Geol. 1961. V. 4. № 10. P. 438–498.
Elmi S., Rulleau I., Gabilly I., Mouterde R. Toarcien. Biostratigraphie du Jurassiquc oucstcuropecn et mediterraneen: zonations parall eles et distribution des invertebres et microfossiles // Bulletin du Centre Recherches Elf Exploration Production. 1997. Mem. 17. P. 25–36.
Howarth M. The Ammonite family Hildoceratidae in the Lower Jurassic of Britain. London: Monograph of the Palaeontographical Society, 1992. 200 p.
Riccardi A.C., Damborenea S.E., Mancenido M.O., Leanza H.A. Megainvertebrados del Jurásico y su importancia geobiológica // XVIII Congreso Geológico Argentino. Geología y Recursos Minerales de la Provincia del Neuquén. Eds. Leanza H.A. et al. 2011. P. 441–464.
Wierzbowski A., Kulicki C., Pugaczewska H. Fauna and stratigraphy of the uppermost Triassic and the Toarcian and Aalenian deposits in the Sassenfjorden, Spitsbergen // Acta Palaeontol. Polon. 1981. V. 26. P. 195–241.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция