

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 5, стр. 113-120
О стратиграфии пограничных отложений байоса и бата (средняя юра) бассейна р. Ижма (европейский север России)
1 Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
2 Череповецкий государственный университет
Череповец, Россия
* E-mail: mitta@paleo.ru
Поступила в редакцию 02.10.2020
После доработки 16.11.2020
Принята к публикации 19.01.2021
Аннотация
Обсуждаются недавно опубликованные В.А. Захаровым, Д.Н. Киселевым, А.П. Ипполитовым и др. данные по руководящим ископаемым и стратиграфии пограничных отложений байоса и бата севера европейской части России. Критически рассматривается предложенная этими авторами корреляция ключевых для стратиграфии средней юры Печорского Севера разрезов по р. Дрещанка (приток р. Ижма), в результате которой вдвое завышается видимая мощность отложений. Новые находки аммонитов Arctocephalites arcticus (Newton) подтверждают биостратиграфические данные, полученные В.В. Митта и др. ранее. Установленный впервые В.А. Захаровым и др. интервал с Mezounio zwerkovi и Retroceramus lungershauseni располагается в видимом основании сысольской свиты непосредственно ниже интервала находок Arctocephalites arcticus и, вероятнее всего, должен сопоставляться с верхним, а не с нижним байосом. Предложение С.В. Мелединой о переносе зон Borealis, Indistinctus и Pompeckji бореальной шкалы из верхнего байоса в нижний байос рассматривается как необоснованное.
ВВЕДЕНИЕ
В последние годы в журнале “Стратиграфия. Геологическая корреляция” опубликованы статьи, в которых рассматриваются вопросы стратиграфии пограничных отложений байоса и бата Печорского Севера (р. Ижма) и ископаемые, важные для биостратиграфического расчленения этих отложений (Киселев, 2020; Захаров и др., 2020). Ранее в тезисной форме были изложены результаты изучения обнажений байоса–бата на речке Дрещанка, правом притоке р. Ижма (Ипполитов и др., 2019), где в новом свете представлена их корреляция; недавно эти данные были дополнены (Киселев, Ипполитов, 2020).
Ниже приводится критическое рассмотрение некоторых результатов исследований, приведенных в указанных выше публикациях и в статье С.В. Мелединой (2014).
ОБСУЖДЕНИЕ
В статье В.А. Захарова с соавторами (2020) описаны 7 видов двустворок байоса, в том числе 1 новый род и 2 новых вида, относящихся к семействам Retroceramidae и Unionidae. Несомненно, это важный вклад в наши познания по систематическому составу этих групп ископаемых и уточнение зональных шкал по ретроцерамам. Вопросы вызывают сопоставление разрезов и корреляция некоторых биостратиграфических подразделений.
Сопоставление разрезов
В обсуждаемых статьях приведены колонки разрезов, охарактеризованных в тексте очень кратко, и их сопоставление между собой (Захаров и др., 2020, рис. 2; Киселев, Ипполитов, 2020, рис. 1). Среди разрезов (прежде всего р. Дрещанка) есть как известные (Кравец и др., 1976; Меледина, 1987; Митта, 2006, 2009; Митта и др., 2013, 2014; Mitta et al., 2015), так и прежде не публиковавшиеся. Данные по строению разрезов и их мощности, представленные А.П. Ипполитовым (Ипполитов и др., 2019; Захаров и др., 2020; Киселев, Ипполитов, 2020), не противоречат в целом опубликованным ранее мною в указанных выше работах. Отличается сопоставление, в результате которого видимая мощность разреза сысольской свиты на р. Дрещанка, по нашим данным составляющая около 7 м, увеличивается в два раза, до 13 м (Захаров и др., 2020, рис. 5). Естественно, такое расхождение в корреляции обнажений и мощности сводного разреза нуждается в дополнительном разборе.
Рис. 1.
Схема расположения обнажений байоса и бата на р. Дрещанка (основа из Google Earth). Номера с литерой D (Дрещанка) – авторская нумерация обнажений, частично приведенная в (Mitta et al., 2015); в квадратных скобках приведены номера по А.П. Ипполитову с соавторами (Захаров и др., 2020; Киселев, Ипполитов, 2020).
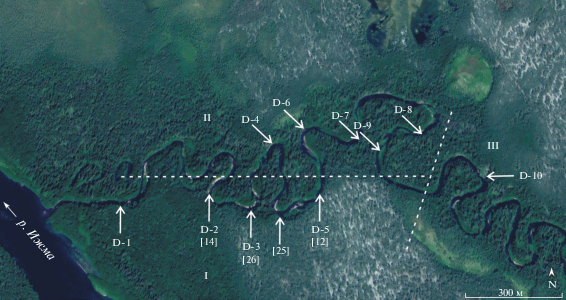
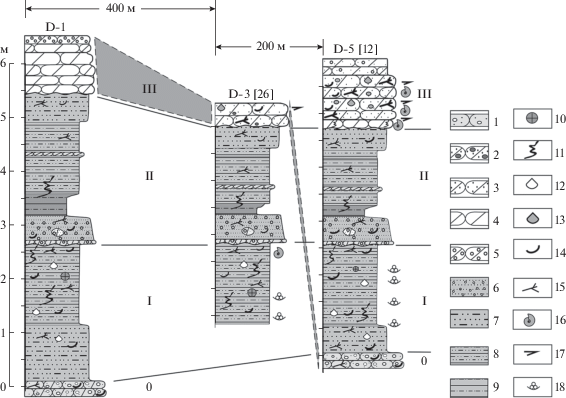
Рис. 2.
Ключевые разрезы левобережья р. Дрещанка группы I и их сопоставление. Сплошными линиями показано сопоставление границ пачек 0–III по (Mitta et al., 2015), заливкой в пунктире – сопоставление “пласта B” по А.П. Ипполитову с соавторами (Ипполитов и др., 2019; Захаров и др., 2020). Условные обозначения: 1 – песчаник галечно-гравийный, 2 – песчаник галечно-гравийный известковый, 3 – песчаник гравийный известковый, 4 – песчаник известковый, 5 – песчаник известковый с железистыми оолитами, 6 – песок глинистый с железистыми оолитами, 7 – песок глинистый, 8 – глина алеврито-песчаная, 9 – глина алевритистая, 10 – пиритовые стяжения, 11 – биотурбации, 12 – целые створки тонкостенных раковин, 13 – целые створки толстостенных раковин, 14 – раковинный детрит, 15 – растительный детрит, 16 – аммониты, 17 – белемниты, 18 – фораминиферы.
На рис. 1 представлен участок нижнего течения р. Дрещанка, с указанием расположения обсуждаемых обнажений: номера с литерой D (Дрещанка) согласно моей нумерации (Mitta et al., 2015), в квадратных скобках приведены номера обнажений из статьи оппонентов (Захаров и др., 2020). Следует оговорить, что в нашей статье по комплексному изучению литологии и макро- и микрофоссилий (Mitta et al., 2015) были представлены лишь наиболее важные разрезы, здесь же показано расположение всех изучавшихся нами в 2006, 2012 и 2013 гг. обнажений по р. Дрещанка; по полноте и строению они распадаются на три группы.
К первой группе (I) относятся обнажения верхнего байоса–нижнего бата по левому берегу: D-1, D-2 [14 ] 11, D-3 [26], [25 ] , D-5 [12 ] , в которых насчитывается до трех уровней с песчаниками, различающимися по литологической характеристике и мощности. В подошве наиболее полных разрезов, над урезом воды (обн. D-1) или в русле (обн. D-5 [12 ] ), располагается песчаник галечно-гравийный, с прослоями песка, видимой мощностью 0.3–0.5 м (пачка 0 в (Mitta et al., 2015)). В средней части залегающей выше песчано-глинисто-алевритовой толщи мощностью около 5.5 м прослеживается прослой песчаника среднезернистого, с железистыми оолитами, по подошве которого проведена граница пачек I и II. Венчает разрез пачка III песчаников песчано-гравийных разнозернистых (обн. D-5 [12 ] ), песчаников гравийных известковых (обн. D-3 [26]) или песчаников среднезернистых с косой слоистостью, в кровле мелкозернистых, с железистыми оолитами (обн. D-1) (рис. 2). В кровле пачки III залегают четвертичные галечники, в той или иной мере срезающие ее верхнюю часть в низовьях р. Дрещанка.
В песчанике пачки 0 обнажений D-5 [12 ] и D-1 найдены только мелкоразмерные двустворки22 плохой сохранности. В осыпи пачки I обнажения D-3 [26] обнаружено пиритизированное ядро аммонита Arctocephalites arcticus (Newton). В пачке III обнажения D-5 [12 ] встречены аммониты Arcticoceras ishmae (von Keyserling), A. harlandi Rawson, здесь же найдены многочисленные двустворки и ростры белемнитов, редкие гастроподы. В осыпи этого обнажения обнаружена раковина аммонита Greencephalites freboldi (Spath), особенности матрикса которой указывают на происхождение из подошвы пачки III на контакте с пачкой II. К этому же виду, по-видимому, относится найденный в осыпи обнажения D-5 [12 ] небольшой фрагмокон, выполненный песчаником пачки III. В пачке III обнажения D-3 [26] встречены лишь редкие двустворки плохой сохранности, единичные ростры белемнитов и остатки древесины; в пачке III обнажения D-1 ископаемые не обнаружены.
Ко второй группе (II) относятся обнажения к северу от горизонтальной пунктирной линии (рис. 1), расположенные как по правому (D-4, D-6, D-8), так и по левому (D-7, D-9) берегу р. Дрещанка. Эти разрезы представляют собой выходы песчаников пачки III нижнего бата непосредственно над урезом воды, видимой мощностью от 1.1 м в западных обнажениях и до 0.1 м выше по реке. В разрезе D-4 найдены аммониты Arcticoceras harlandi, ростры белемнитов, многочисленные и разнообразные двустворки, редкие брахиоподы Boreiothyris. В обнажениях, расположенных восточнее, ископаемые встречаются реже; в разрезах D-6, D-7, D-9 найдены Arcticoceras ishmae.
Третью группу обнажений (III) на р. Дрещанка представляет единственный разрез D-10, преимущественно глинистый, большей частью принадлежащий к чуркинской свите (Унифицированная…, 2012), отнесенный предположительно к среднему бату и сопоставленный с аналогичным разрезом у порогов Грива на р. Ижма. Из макрофоссилий здесь найдены лишь редкие ростры белемнитов.
Этот краткий обзор данных по строению обнажений верхнего байоса и бата на р. Дрещанка, полученных в результате многолетних исследований и в основном приведенных в нашей итоговой статье (Mitta et al., 2015), необходим для обсуждения их биостратиграфического расчленения и сопоставления.
Разрезы байоса–бата группы I располагаются практически на одной субширотной линии (рис. 1). Согласно их сопоставлению А.П. Ипполитовым с коллегами (Захаров и др. 2020, рис. 2), весь разрез D-5 [12 ] мощностью ~6 м на расстоянии ~300 м к западу срезается квартером, и в обнажении D-2 [14 ] экспонируется еще ~6 м отложений, кровля которых сопоставляется с подошвой разреза D-5 (12). Это было бы возможно при падении отложений с запада на восток. Но, согласно коллегам (Захаров и др., 2020, с. 74), средняя юра залегает здесь с падением слоев на СВ под углом ~0.5°.
В статье В.А. Захарова с соавторами (2020) не рассматривается разрез D-1 (Mitta et al., 2015, рис. 2; настоящая работа, рис. 2), расположенный западнее, в ~300 м от обнажения D-2 [14 ] и в ~400 м от D-3 [26]; но он упоминается в более ранней публикации (Ипполитов и др., 2019, с. 32). В схематичном виде корреляция А.П. Ипполитова с коллегами, с учетом данных из обеих указанных публикаций, показана на рис. 3 (горизонтальный масштаб соответствует таковому на рис. 1). Согласно коллегам, “песчанистый пласт B” разрезов D-1, D-2 [14 ] и D-3 [26] на протяжении ~400 м завершает юрский разрез, а еще через ~200 м далее к востоку “ныряет” в его подошву.
Рис. 3.
Схематическое отображение интервалов с песчаниками в разрезах группы I на левобережье р. Дрещанка и взглядов А.П. Ипполитова с соавторами (Ипполитов и др., 2019; Захаров и др., 2020; Киселев, Ипполитов, 2020) на сопоставление так называемого “пласта B” (показано заливкой в пунктире).
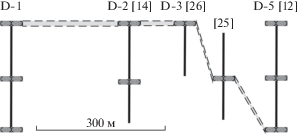
Иными словами, песчаники в кровле юры обнажений D-1 и D-3 [26]), отстоящих друг от друга на ~400 м (рис. 2), А.П. Ипполитовым с коллегами сочтены изохронными. Согласно коллегам, еще в ~200 м к востоку им соответствуют уже песчаники в подошве обнажения D-5 [12 ] , выше которых располагается разрез, идентичный западным разрезам по строению и мощности слоев.
Перепад абсолютных высот от обнажения D-5 [12 ] по руслу р. Дрещанка к ее устью незначителен, в пределах 1 м; дно речки почти плоское, с неглубокими фарватерами только на излучинах. Тектонических сбросов непосредственно к западу от обнажения D-5 [12 ] и в обсуждаемом районе в целом не установлено.
На литологических колонках коллег (Захаров и др., 2020, рис. 2) видно, что при сопоставлении основания пласта C обн. D-5 [12 ] с основанием пласта B обнажения D-2 [14 ] слой 2 обнажения D-5 [12 ] хорошо коррелирует с прослоем песчаника в верхней части сл. 1а обнажения D-2 [14 ] , в том числе по мощности залегающего между ними интервала.
Соответственно, я считаю наше сопоставление разрезов р. Дрещанка (Митта, 2006, 2009; Mitta et al., 2015) правильным и вполне обоснованным. Дополнительным обоснованием нашей корреляции служат разрезы правобережья р. Дрещанка, отстоящие от левобережных на 200–300 м на север и северо-восток (D-4, D-6, D-7), представленные “арктикоцерасовым” песчаником (Захаров и др., 2020) нижнебатской пачки III (Mitta et al., 2015) сначала в его нижней (D-4, слои с Arcticoceras harlandi), а выше по речке и в верхней части (D-6, D-7, слои с Arcticoceras ishmae). Слои с A. ishmae, обнажающиеся над урезом воды далее на восток выше по течению (разрезы D-8, D-9) и, соответственно, выше гипсометрически, являются верхами пачки III. Это вполне согласуется с упомянутым выше падением слоев сысольской свиты на СВ: пачки I и II обнажений группы I на протяжении первых сотен метров на СВ погружаются, и в разрезах группы II экспонированы лишь “арктикоцерасовые” песчаники пачки III, выше по речке также уходящие под урез воды, уже в связи с современной геоморфологией. Еще выше по р. Дрещанка обнажаются более высокие слои пачки IV (разрез D-10, предположительно средний бат).
Ожелезнение отдельных прослоев песчаника, по-видимому поставленное оппонентами в основу корреляции, является, скорее всего, вторичным. Гораздо информативнее железистые оолиты несомненно синседиментационного генезиса, обнаруженные нами в песчаниках и песках в подошве пачки II и выше, в том числе в отдельных прослоях пачки III и низах пачки IV (Митта и др., 2013; Mitta et al., 2015). Наличие железистых оолитов позволяет предполагать существование поблизости устья крупной реки (Холодов и др., 2012). О близости береговой линии свидетельствует и большое количество крупных фрагментов древесины в пачках II и III. Кроме того, это объясняет полное отсутствие находок аммонитов и фораминифер в пачке II – в авандельте, в результате ее распреснения, не могли обитать стеногалинные морские животные. Совершенно “немые” песчаники пачки III обнажения D-1 сменяются восточнее песчаниками обнажения D-3 [26], уже с двустворками и растительными остатками и единичными рострами белемнитов, а еще дальше на восток в них появляется более разнообразная морская фауна, включая аммониты и фораминиферы. Соответственно, дельта “пра-Ижмы” располагалась в конце байоса–начале бата вблизи устья р. Дрещанка или еще далее к западу.
Сопоставление линз песчаника (по А.П. Ипполитову с соавторами, “огромных блинообразных тел конкреционной природы, или мегаконкреций”), обнажающихся в русле устья р. Дрещанка, и песчаников по р. Ижма непосредственно ниже и выше устья Дрещанки (Захаров и др., 2020) считаю абсолютно верным. С учетом падения слоев и современного рельефа, разные интервалы этой пачки 0 (Mitta et al., 2015), переслаивающиеся и переходящие в пески по латерали, обнажаются в основании юрского разреза и выше по р. Дрещанка, вплоть до обнажения D-5 (12). В то же время отнесение этого интервала к нижнему байосу, даже условно, считаю необоснованным.
Бореально-тетическая корреляция
Предположение В.А. Захарова с соавторами (Захаров и др., 2020) о возможной принадлежности слоев с Mezounio zwerkovi и Retroceramus lungershauseni к нижнему байосу опираются на недавние выводы С.В. Мелединой (2014) о соответствии зоны Arctocephalites arcticus бореальной шкалы всему верхнему байосу, а располагающихся ниже трех других бореальных зон – самым верхам нижнего байоса (рис. 4).
Рис. 4.
Взгляды С.В. Мелединой (2014) на сопоставление пограничных подразделений нижнего и верхнего байоса (фрагмент).
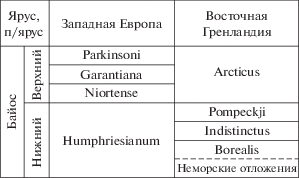
В предложенном С.В. Мелединой (2014) варианте корреляции три зоны верхнего байоса стандартной западноевропейской шкалы, каждая из которых подразделена на три подзоны и многочисленные фаунистические горизонты, соответствуют одной бореальной зоне Arcticus, понимаемой в Восточной Гренландии в объеме двух фаунистических горизонтов по аммонитам (Callomon, 1993), а в Северной Сибири в объеме двух подзон (Меледина, 1994, 2014). Напротив, верхняя зона нижнего байоса западноевропейской шкалы сопоставлена с тремя зонами бореальной шкалы, представленными, по последним данным (Callomon et al., 2015), более чем 20 биогоризонтами. Логические построения С.В. Мелединой основаны преимущественно на сходстве раковин восточногренландских и северосибирских Arctocephalites Spath и американских Megasphaeroceras Imlay и дальневосточных Umaltites Kalacheva.
Несколько десятилетий назад (Callomon, 1984, 1985) была предложена схема филогении сем. Cardioceratidae, принятая в дальнейшем всеми исследователями, в том числе С.В. Мелединой (1994), и не претерпевшая особых изменений до наших дней (Howarth, 2017). Согласно этой схеме, первые представители Cardioceratidae (род Cranocephalites, подсем. Arctocephalitinae) происходят от рода Defonticeras (подсем. Sphaeroceratinae, одноименного семейства), из верхней зоны нижнего байоса. Это дало Дж. Калломону логичные основания предположить, что интервал первого появления Cranocephalites – зона Cranocephalites borealis – находится непосредственно выше, в базальном верхнем байосе.
Рис. 5.
Схема сопоставления зональных шкал средней юры Западной Европы (перитетический стандарт) и Восточной Гренландии (бореальный стандарт) (Callomon, 1993, с изменениями и уточнениями по (Morton et al., 2020)).
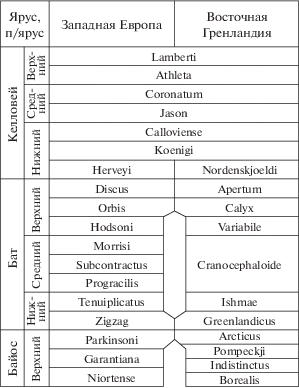
Род Megasphaeroceras (и Umaltites, как его младший синоним) (подсем. Eurycephalitinae Thierry, сем. Sphaeroceratidae), согласно схеме Дж. Калломона, тоже отделился от основного ствола развития сфероцератид, и тоже в начале позднего байоса. Поэтому общее сходство некоторых представителей арктоцефалитин и еврицефалитин, относительно недавно разошедшихся по разным эволюционным линиям, вполне объяснимо. Впрочем, сравнение Arctocephalites и Megasphaeroceras показывает, что для раковин последних характерны тонкие густые ребра, что является обычным для ранних сфероцератид; это хорошо видно и на фотографиях, приведенных С.В. Мелединой (2014, табл. I ). В то же время раковины Defonticeras и Cranocephalites (Callomon, 1985, рис. 7 ) чрезвычайно сходны как по размерам и форме раковины, так и по скульптуре, представленной редкими, заметно более грубыми ребрами.
Семейство Cardioceratidae являлось в байосе, бате, келловее и затем в поздней юре (вплоть до вымирания в конце кимериджа) одной из наиболее быстро эволюционировавших групп аммонитов; со временем оно распространилось из высокобореальных районов далеко на юг и не утратило при этом значения для детальной биостратиграфии (Callomon, 1985). Трудно представить, что в фазу Arcticus скорость эволюции Arctocephalitinae вдруг замедлилась в разы, по сравнению с таковой до и после, и что интервал существования двух последовательных видов в бореальных районах (Arcocephalites delicatus и A. arcticus) может соответствовать интервалу объемом в 9 подзон по аммонитам на северо-западе Тетис.
В недавней монографии по аммонитам и биостратиграфии байоса Восточной Гренландии (Callomon et al., 2015) зона Arcticus отнесена к базальному бату и предположительно соответствует узкому стратиграфическому интервалу нижней подзоны (Convergens) зоны Zigzag. Ниже в зональной шкале Восточной Гренландии, как и в обновленной версии Бореального стандарта (Меледина и др., 2011; Шурыгин и др., 2011; Никитенко и др., 2013), располагаются еще три зоны по аммонитам, отнесенные авторами упомянутых работ к верхнему байосу. Впрочем, некоторые другие исследователи юры Гренландии полностью принимают мои с соавторами выводы, располагая весь интервал от зоны Borealis до зоны Arcticus включительно в верхнем байосе (Kelly et al., 2015).
Действительно, зона Arcticus сопоставляется нами (Mitta et al., 2014, 2015 и более ранние публикации) с самыми верхами байоса только по положению в разрезе и теоретически может относиться к самым низам бата. Песчаники и пески пачки 0 (интервал с “мегаконкрециями”, или слои с Mezounio zwerkovi и Retroceramus lungershauseni) могут относиться как к низам зоны Arcticus, так и к зоне Pompeckji, располагающейся в бореальной шкале непосредственно ниже (Callomon, 1993; Callomon et al., 2015). По моему мнению, это безусловно верхний байос (рис. 5).
Аммониты и инфразональные подразделения нижнего бата
В статье Д.Н. Киселева (2020) рассматриваются представители подсем. Arctocephalitinae из зоны Ishmae и смежных интервалов, прежде всего бассейна р. Печора. Впрочем, к обсуждению привлечены опубликованные таксоны Arcticoceras всей Панбореальной надобласти, включая Шпицберген, бассейны рек Печора и Волга, Северную и Восточную Сибирь, Восточную Гренландию и Арктическую Канаду. В результате Д.Н. Киселев приходит к выводу, что большинство пригодных видовых названий, предложенных различными авторами в роде Arcticoceras, не являются валидными. Для вида A. ishmae он выделяет три различные “вариации” (морфы): A. ishmae (Keys.) morpha ishmae (Keys.), A. ishmae (Keys.) morpha stepankovi Tuchkov, A. ishmae (Keys.) morpha pseudishmae (Spath). Далее в этом же виде обозначаются три “хроновариации” (уже как подвиды): A. ishmae (Keys.) subsp. A, A. ishmae (Keys.) subsp. B и A. ishmae ishmae (Keys.).
Статья Д.Н. Киселева сопровождается многочисленными и разнообразными графиками. На первый взгляд, прекрасным обоснованием выводов автора служит иллюстрация рядов изменчивости (Киселев, 2020, табл. I ). Однако бóльшая часть аммонитов на этой фототаблице происходит из старых музейных коллекций, собранных в различных местонахождениях, то есть их точная стратиграфическая приуроченность неизвестна33. Соответственно, есть большая вероятность происхождения этих аммонитов из разных интервалов зоны Ishmae, и не только. Три экземпляра из изображенных на этой таблице происходят из сборов Д.Н. Киселева, при этом одна раковина (Киселев, 2020, табл. I , фиг. 4) явно отличается от двух других (Киселев, 2020, табл. 1 , фиг. 1, 2) сильно вздутыми оборотами с заметно более широким пупком и меньшим коэффициентом ветвления ребер, что характерно для Greencephalites ex gr. freboldi (Spath).
Следует учитывать и то, что морфотип предкового вида какое-то время может проявляться в видах-потомках. Впрочем, и в рассмотренной выше статье с участием Д.Н. Киселева допускается, пусть по отношению к двустворкам, что “нередко смена видов, характерных для смежных зон, происходит внутри некоего стратиграфического интервала с переходными формами” (Захаров и др., 2020, с. 90).
Субъективность выделения и понимания палеонтологических видов общеизвестна. Наиболее объективным критерием разделения родственных родов и видов, относящихся к одному таксону ранга семейства и найденных на одном стратиграфическом уровне, является отсутствие переходных форм. В настоящее время не установлено никаких переходов от A. ishmae к аммонитам нижнего бата, определенным как Greencephalites sp. nov. (Митта, Альсен, 2013, табл. I , фиг. 2), G. freboldi (Spath) (Митта, 2009, табл. II , фиг. 2) и G. belli (Poulton) (Mitta et al., 2015, рис. 7 ). Следовательно, нет оснований для сведения названия Greencephalites в синонимию Arcticoceras или Arctocephalites.
C оговорками о том, что последовательность хроновариаций Arcticoceras ishmae “может быть намечена только проспективно”, Д.Н. Киселев (2020, c. 99, рис. 7 ) предлагает свой вариант инфразонального деления зоны Ishmae Бореально-Арктической области. Однако здесь как “интервалы высокого обоснования” для Восточной Гренландии по-прежнему показаны фаунистические горизонты Дж. Калломона (Callomon, 1993), включая горизонт A. harlandi, хотя этот вид сводится Д.Н. Киселевым в синонимию A. ishmae. Эти горизонты полностью соответствуют биогоризонтам, общим для всей Бореально-Арктической области, но названия подразделений в нижней части зоны Ishmae заменены Д.Н. Киселевым на A. ishmae subsp. A, A. ishmae subsp. B, A. ishmae ishmae.
Завершая статью, Д.Н. Киселев (2020, с. 100) пишет, что “зона Harlandi является младшим синонимом зоны Ishmae и должна быть упразднена”. Однако в ранге зоны или подзоны интервал с A. harlandi принимался лишь сибирскими (Меледина, 1994) и канадскими (Poulton, 1987) исследователями (в последнем случае это объясняется самобытными правилами североамериканских стратиграфов, не признающими инфразональных подразделений). К настоящему времени и сибирские исследователи пришли к пониманию этого интервала как эквивалента базальной части зоны Ishmae (например, Меледина и др., 2011), поэтому упразднение зоны Harlandi немного запоздало.
ЗАКЛЮЧЕНИЕ
Данные по аммонитам, изложенные в статьях (Захаров и др., 2020; Киселев, Ипполитов, 2020), полностью подтверждают наши полевые наблюдения и окончательные результаты их обработки (Mitta et al., 2015) и дополняют их. Особенно важны новые находки Arctocephalites arcticus (Newton), подтверждающие правильность отнесения первой находки аммонита этого вида к пачке I (Митта, 2006, 2009). Важны также находки ретроцерамов для дальнейшей калибровки биостратиграфических подразделений по этой космополитной группе двустворчатых моллюсков.
Однако нельзя согласиться с предложенным (Ипполитов и др., 2019; Захаров и др., 2020; Киселев, Ипполитов, 2020) вариантом корреляции разрезов по р. Дрещанка, в результате которой в два раза завышается видимая мощность сысольской свиты. Как нельзя согласиться и с датировкой слоев с Mezounio zverkovi ранним байосом, даже как условной – эти слои, по моему мнению, относятся к верхнему байосу, исходя из их непрерывной связи с вышележащими отложениями и обоснованного еще Дж. Калломоном (Callomon, 1985 и др.) сопоставления нижних зон “бореального бата” с верхним байосом. Соответственно, в настоящее время нет никаких оснований датировать начало среднеюрской бореальной трансгрессии ранним байосом (Захаров и др., 2020, с. 91). Все имеющиеся данные указывают на близость этого события к границе байоса и бата.
Благодарности. Седиментолог В.В. Костылева (ГИН РАН), специалист по микрофауне Л.А. Глинских (ИНГГ СО РАН) и геолог-съемщик А.В. Ступаченко (Москва) принимали активное участие в изучении среднеюрских разрезов бассейна р. Ижма. Любители палеонтологии из Германии О. Нагель (O. Nagel, Radeberg) и Ш. Гребенштайн (S. Gräbenstein, Bodelshausen) оказали дружескую помощь при проведении полевых работ. Специалисты по разным группам фауны В.А. Захаров (ГИН РАН), О.С. Дзюба, Б.Л. Никитенко, Б.Н. Шурыгин (ИНГГ СО РАН) приняли участие в обработке собранных коллекций. ПИН РАН оказывал содействие в организации Юрского полевого отряда. Я благодарен всем лицам и организациям, способствовавшим изучению Печорской юры. Приношу также свою искреннюю признательность рецензентам Д.Б. Гуляеву (Ярославль) и М.А. Рогову (ГИН РАН), чьи критические замечания позволили улучшить рукопись.
Список литературы
Захаров В.А., Ипполитов А.П., Зверьков Н.Г., Безносов П.А., Киселев Д.Н. Ретроцерамы и униониды из байоса и нижнего бата (средняя юра) бассейна р. Ижма, север Европейской России // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 4. С. 73–95.
Ипполитов А.П., Киселев Д.Н., Зверьков Н.Г. О положении границы байоса и бата в опорных разрезах р. Ижмы (Тимано-Печорская область) // Палеострат-2019. Годичное собрание (научная конференция) секции палеонтологии МОИП и Московского отделе- ния Палеонтологического общества при РАН. Москва, 28–30 января 2019 г. Программа и тезисы докладов. Ред. Алексеев А.С., Назарова В.М. М.: Палеонтологический ин-т им. А.А. Борисяка РАН, 2019. С. 32–33.
Киселев Д.Н. Род Аrcticoceras Spath, 1924 (Аmmonoidea) и инфразональная биостратиграфия нижнего бата Бореальной области // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 2. С. 77–102.
Киселев Д.Н., Ипполитов А.П. О последовательности арктоцефалитин (Ammonoidea) в верхнем байосе и нижнем бате Тимано-Печорского региона // Юрская система России: проблемы стратиграфии и палеогеографии. Материалы VIII Всероссийского совещания с международным участием. Онлайн-конференция, 7–10 сентября 2020 г. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 107–112.
Кравец В.С., Месежников М.С., Яковлева С.П. Важнейшие разрезы юрской системы на Севере СССР. Русская равнина. Реки Ижма и Пижма // Стратиграфия юрской системы Севера СССР. М.: Наука, 1976. С. 14–24.
Меледина С.В. Аммониты и зональная стратиграфия келловея суббореальных районов СССР. М.: Наука, 1987. 182 с.
Меледина С.В. Бореальная средняя юра России // Тр. ИГиГ СО РАН. 1994. Вып. 819. 184 с.
Меледина С.В. О корреляции зон байоса и бата Сибири в свете новых палеонтологических данных // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 6. С. 45–56.
Меледина С.В., Никитенко Б.Л., Шурыгин Б.Н., Дзюба О.С., Князев В.Г. Современная комплексная зональная шкала юры Сибири и бореальный стандарт // Новости палеонтологии и стратиграфии. 2011. Вып. 16–17. С. 17–40.
Митта В.В. Первая находка Arctocephalites (Cardioceratidae, Ammonoidea) в средней юре бассейна Печоры // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция и биостратиграфия. Материалы Всероссийского совещания, Москва, 8–10 ноября 2006 г. Ред. Барсков И.С., Леонова Т.Б. М.: ПИН РАН, 2006. С. 82–84.
Митта В.В. Верхний байос и нижний бат бассейна Печоры и бореально-тетическая корреляция // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 1. С. 77–87.
Митта В.В., Альсен П. Аммониты и зональная шкала батского яруса Гренландии // Юрская система России: проблемы стратиграфии и палеогеографии. Тюмень: ЗапСибНИИГГ; Екатеринбург: ИздатНаукаСервис, 2013. С. 149–151.
Митта В.В., Захаров В.А., Барсков И.С., Сельцер В.Б., Иванов А.И. Верхний байос и нижний бат окрестностей Саратова: малакологическая характеристика и биостратиграфия // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 5. С. 32–45.
Митта В.В., Костылева В.В., Глинских Л.А. О байосе и бате (средняя юра) бассейна р. Ижма // Проблемы геологии Европейской России. Сборник научных трудов Всероссийской научной конференции, посвященной 130-летию со дня рождения профессора Бориса Александровича Можаровского, Саратов, 11–14 сентября 2013 г. Саратов: СГТУ, 2013. С. 93–103.
Митта В.В., Глинских Л.А., Костылева В.В. Предварительный отчет по полевым работам 2013 г. в бассейнах рек Сысола и Ижма // Геология и минеральные ресурсы Европейского Северо-Востока России. Материалы XVI Геологического съезда Республики Коми. Т. 2. Сыктывкар: Геопринт, 2014. С. 188–190.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г., Меледина С.В., Дзюба О.С., Лебедева Н.К., Пещевицкая Е.Б., Глинских Л.А., Горячева А.А., Хафаева С.Н. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный зональный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082.
Унифицированная стратиграфическая схема юрских отложений Восточно-Европейской платформы. Объяснительная записка. Ред. Митта В.В., Алексеев А.С., Шик М.С. М.: ПИН РАН–ВНИГНИ, 2012. 14 л. + 64 с.
Холодов В.Н., Недумов Р.И., Голубовская Е.В. Фациальные типы осадочных железорудных месторождений и их геохимические особенности. Сообщение 1. Фациальные группы осадочных руд, их литология и генезис // Литология и полезн. ископаемые. 2012. № 6. С. 503–531.
Шурыгин Б.Н., Никитенко Б.Л., Меледина С.В., Дзюба О.С., Князев В.Г. Комплексные зональные шкалы юры Сибири и их значение для циркумарктических корреляций // Геология и геофизика. 2011. Т. 52. № 8. С. 1051–1074.
Callomon J.H. A review of the biostratigraphy of the post-Lower Bajocian Jurassic ammonites of the western and northern North America // Geol. Assoc. Can. Spec. Pap. 1984. V. 27. P. 143–174.
Callomon J.H. The evolution of the Jurassic ammonite family Cardioceratidae // Spec. Pap. Paleont. 1985. № 33. P. 49–90.
Callomon J.H. The ammonite succession in the Middle Jurassic of East Greenland // Bull. Geol. Soc. Denmark. 1993. V. 40. P. 83–113.
Callomon J.H., Alsen P., Surlyk F. The ammonites of the Middle Jurassic Cranocephalites beds of East Greenland // Geol. Surv. Denmark Greenland Bull. 2015. V. 34. P. 1–86.
Howarth M.K. Systematic descriptions of the Stephanoceratoidea and Spiroceratoidea // Treatise Online. № 84. Pt L, revised. V. 3B. Chap. 6. Paleontol. Inst. Univ. Kansas, 2017. 101 p.
Kelly S.R.A., Gregory F.J., Braham W., Strogen D.P., Whitham A.G. Towards an integrated Jurassic biostratigraphy for eastern Greenland // Volumina Jurassica. 2015. V. XIII. № 1. P. 43–64.
Mitta V., Kostyleva V., Dzyuba O., Glinskikh L., Shurygin B., Seltzer V., Ivanov A., Urman O. Biostratigraphy and sedimentary settings of the Upper Bajocian–Lower Bathonian in the vicinity of Saratov (Central Russia) // N. Jb. Geol. Palaont. Abh. 2014. V. 271. № 1. S. 95–121.
Mitta V., Glinskikh L., Kostyleva V., Dzyuba O., Shurygin B., Nikitenko B. Biostratigraphy and sedimentary settings of the Bajocian–Bathonian beds in the Ishma River basin (European North of Russia) // N. Jb. Geol. Palaont. Abh. 2015. V. 277. № 3. S. 307–335.
Morton N., Mitta V.V., Underhill J.R. Ammonite occurrences in North Sea cores: implications for Jurassic Arctic-Mediterranean marine seaway connectivity // Scottish J. Geol. 2020. V. 56. № 2. P. 175–195.
Poulton T.P. Zonation and correlation of Middle Boreal Bathonian to Lower Callovian (Jurassic) ammonites, Salmon Cache Canyon, Porcupine river, Northern Yukon // Bull. Geol. Surv. Can. 1987. № 358. P. 1–155.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция