

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 5, стр. 12-26
Состав и возраст тафофлор кривореченской свиты (верхний мел) левобережья реки Анадырь, Северо-Восток Азии
А. Б. Герман 1, *, С. В. Щепетов 2, **
1 Геологический институт РАН
Москва, Россия
2 Ботанический институт РАН
Санкт-Петербург, Россия
* E-mail: alexeiherman@yandex.ru
** E-mail: shchepetov@mail.ru
Поступила в редакцию 29.12.2020
После доработки 01.02.2021
Принята к публикации 08.02.2021
Аннотация
Чинейвеемская тафофлора из кривореченской свиты левобережья р. Анадырь (Северо-Восток Азии) характеризуется невысоким разнообразием входящих в нее растений, представленных гинкговыми, хвойными и покрытосеменными. По их систематическому составу она моложе гребенкинской флоры из кривореченской свиты бассейна р. Анадырь и наиболее близка к пенжинской флоре Северо-Западной Камчатки и п-ова Елистратова, возраст которой надежно определяется как туронский, исключая начало турона. Исходя из этого, возраст чинейвеемского флористического комплекса также туронский, скорее всего позднетуронский. Такая датировка этой тафофлоры отвечает представлениям о позднетуронском или коньякском возрасте морских моллюсков из перекрывающих слоев дуговской свиты на р. Чинейвеем и о согласном стратиграфическом контакте этих слоев с флороносными отложениями. Предполагавшийся ранее тектонический контакт между указанными отложениями в этом районе не находит палеоботанического подтверждения. Последовательность тафофлор из кривореченской свиты в междуречье Убиенка–Чинейвеем позволяет ограничить верхний возрастной предел распространения гребенкинской флоры началом туронского века.
ВВЕДЕНИЕ
Северо-Востоком Азии принято называть часть Евразийского континента к востоку от р. Лена (Самылина, 1974). Здесь широко представлены верхнемезозойские отложения различного генезиса. По характеру осадконакопления в меловом периоде данная территория разделена на три субрегиона (Герман, 1993, 2011) или два региона (Решения…, 2009). Для Анадырско-Корякского субрегиона (рис. 1) характерно чередование в разрезах и/или замещение по простиранию меловых отложений морского и континентального генезиса, причем последние, формировавшиеся преимущественно в условиях приморских низменностей и равнин, содержат многочисленные и разнообразные ископаемые растения. Остатки моллюсков и фораминифер из морских слоев позволяют здесь надежно датировать растительные остатки, содержащиеся в неморских отложениях (Герман, 2011).
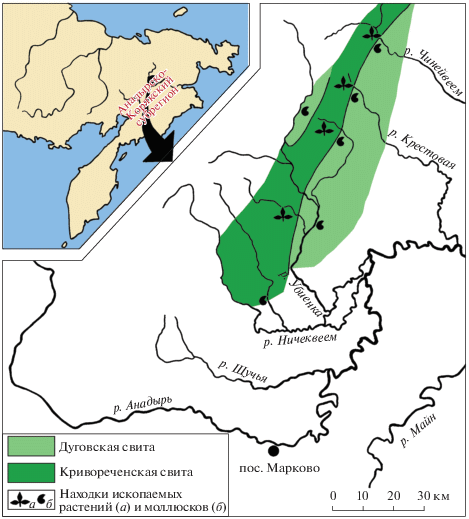
Рис. 1.
Распространение флороносных отложений кривореченской свиты и перекрывающих образований дуговской свиты на левобережье среднего течения р. Анадырь и приуроченные к ним местонахождения ископаемых растений и моллюсков (по Герман, 2011, с изменениями).
Данная особенность делает некоторые разрезы Анадырско-Корякского субрегиона ключевыми для разработки стратиграфических схем неморского альба–верхнего мела Северо-Востока России. При этом в качестве фитостратиграфических реперов рассматриваются ископаемые флоры, характеризующие определенные этапы развития древней флоры на значительной территории; каждая из таких флор включает отдельные тафофлоры, или палеофлористические комплексы, в состав которых входят растения из одного или нескольких территориально и стратиграфически близких местонахождений (Герман, 2011). Для первой половины позднего мела были выделены и описаны гребенкинский (конец альба–начало турона), пенжинский (турон, исключая его начало) и кайваямский (коньяк) этапы развития древней флоры (Самылина, 1974; Герман, Лебедев, 1991; Герман, 2011; Щепетов и др., 1992; Филиппова, Абрамова, 1993; Spicer et al., 2002).
Гребенкинская флора из кривореченской свиты среднего течения р. Анадырь – самая богатая и разнообразная из позднемеловых флор Северо-Востока Азии. Она является одним из наиболее важных фитостратиграфических реперов неморского мела региона. Гребенкинский этап, с которого начинается позднемеловой флорогенез на Северо-Востоке России, был выделен В.А. Самылиной (1974). В течение этого этапа формировались гребенкинская флора среднего течения р. Анадырь и среднегинтеровская флора бухты Угольной (Моисеева, 2010; Герман, 2011). Поскольку в бассейне р. Гребенка тафофлоры гребенкинского этапа датируются не только сеноманом, но и, вероятно, поздним альбом (Щепетов и др., 1992; Spicer et al., 2002; Герман, 2011), начало этого этапа следует датировать в пределах позднего альба. В перекрывающей кривореченскую свиту дуговской свите (рис. 1, 2) были найдены остатки туронских морских моллюсков (Терехова, 1988), что позволило провести границу между гребенкинским и пенжинским этапами развития флоры в начале туронского века или между ранним и поздним туроном. При этом смены тафофлор этих этапов, документированной остатками растений, в едином разрезе известно не было.
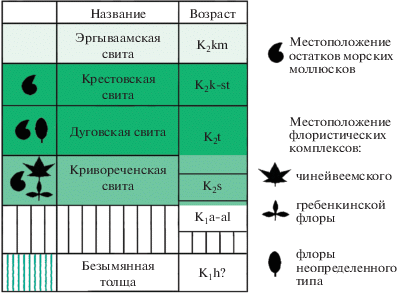
Рис. 2.
Последовательность стратифицированных геологических тел на левобережье р. Чинейвеем в районе руч. Болотный по материалам Б.Д. Трунова (Государственная…, 1977).
Из отложений, относимых к кривореченской свите, в бассейне р. Чинейвеем (левобережье р. Анадырь) геологами научно-производственного объединения “Аэрогеология” были собраны растительные остатки, изученные Е.Л. Лебедевым (Дворянкин и др., 1993). По его мнению, данная тафофлора принципиально отличается по составу от гребенкинской и может быть условно датирована сеноном. Вероятно, именно из-за этого Дворянкин и его соавторы сочли, что остатки растений происходят из тектонического блока пород, не имеющих отношения к кривореченской свите. Позже А.Б. Герман (2011), проанализировав приведенный Лебедевым список растений чинейвеемской тафофлоры и просмотревший коллекцию ее растительных остатков, не согласился со стратиграфическими выводами А.И. Дворянкина с соавторами (1993). Однако сама эта тафофлора так и не была детально изучена, что делает заключения о ее возрасте, сделанные как Лебедевым, так и Германом, недостаточно обоснованными.
В настоящей статье мы восполняем этот пробел и пытаемся ответить на следующие вопросы.
1. Каков систематический состав и возраст чинейвеемского флористического комплекса?
2. Насколько обосновано высказанное ранее мнение о тектоническом контакте флороносных слоев, включающих чинейвеемский комплекс, с дуговской свитой морского генезиса?
3. Каков верхний возрастной предел существования гребенкинской флоры и, соответственно, гребенкинского этапа развития меловой флоры Анадырско-Корякского субрегиона?
СТРАТИГРАФИЯ ВЕРХНЕМЕЛОВЫХ ОТЛОЖЕНИЙ ЛЕВОБЕРЕЖЬЯ р. АНАДЫРЬ (КРИВОРЕЧЕНСКАЯ И ДУГОВСКАЯ СВИТЫ)
На левом берегу р. Анадырь кривореченская свита распространена в бассейнах рек Ничеквеем, Убиенка, Крестовая и Чинейвеем (рис. 1). На р. Кривая (правый приток р. Убиенка) расположен стратотип кривореченской свиты, выделенной в 1963 г. Г.Г. Кайгородцевым.
Кривореченская свита в этом районе, как и на правобережье р. Анадырь (Девятилова и др., 1980; Герман, 2011), состоит из двух подсвит: нижней существенно конгломератовой и верхней, сложенной конгломератами, гравелитами, полимиктовыми и вулканомиктовыми песчаниками и глинистыми алевролитами. В верхней подсвите многочисленны остатки растений, однако они менее разнообразны и худшей сохранности, чем на правобережье р. Анадырь (Филиппова, Абрамова, 1993). Наиболее представительные сборы фитофоссилий из верхнекривореченской подсвиты в бассейнах рек Ничеквеем, Убиенка, Кривая, Дуговая были сделаны А.Д. Девятиловой, Эд.Б. Невретдиновым и Л.А. Анкудиновым, а изучила эти коллекции Г.Г. Филиппова (1979, 1982, 1984; Филиппова, Абрамова, 1993; Девятилова и др., 1980). На р. Левая Ничеквеем (рис. 1) в верхнекривореченской подсвите Л.А. Анкудиновым были найдены остатки Inoceramus cf. nipponicus Nagao et Mat., I. korjakensis Ter., I. ex gr. korjakensis Ter., I. cf. gradilis Perg., свидетельствующие, по мнению Г.П. Тереховой (1988), о принадлежности вмещающих отложений зоне Inoceramus nipponicus средне-позднесеноманского и, возможно, раннетуронского возраста. Мощность кривореченской свиты в этом районе около 1600 м.
По определениям Г.Г. Филипповой (Филиппова, Абрамова, 1993), в кривореченской свите на реках Кривая, Ветвистая и Дуговая (бассейн р. Убиенка) обнаружены (в квадратных скобках приведены более правильные, на наш взгляд, определения): Thallites sp., Osmunda sp., Gleichenites zippei (Corda) Sew., Gleichenites microphylla Philipp., Gleichenites asiatica Philipp., Coniopteris grebencaensis Philipp., Asplenium dicksonianum Heer, Birisia jelisejevii (Krysht.) Philipp., Cladophlebis frigida (Heer) Sew., Cladophlebis ex gr. haiburnensis (L. et H.) Brong., Cladophlebis ubiencaensis Philipp., Arctopteris sp. [Arctopteris penzhinensis E. Lebed.], Pseudocycas hyperborea Krysht. [Cycadites hyperborea (Krysht.) E. Lebed.], Nilssonia serotina Heer, Nilssonia alaskana Hollick, Nilssonia yukonensis Hollick, Ginkgo ex gr. adiantoides (Unger) Heer, Sphenobaiera aff. biloba Pryn. [Sphenobaiera vera Samyl. et Shczep.], Cephalotaxopsis heterophylla Hollick, Cephalotaxopsis intermedia Hollick, Elatocladus gracillimus (Holl.) Sveshn., Araucarites anadyrensis Krysht., Sequoia reichenbachii (Gein.) Heer, Sequoia fastigiata (Sternb.) Heer, Sequoia obovata Knowlton [скорее всего, Elatocladus smittiana (Heer) Seward], Sequoia ambigua Heer [Pagiophyllum triangulare Prynada], Cryptomeria subulata (Heer) Sveshn., Florinia borealis Sveshn. et Budantsev, Podozamites lanceolatus (Lindl. et Hutton) Braun, Pityophyllum nordenskioldii (Heer) Nath. [Pityophyllum ex gr. nordenskioldii (Heer) Nath.], Pityostrobus sp., Pityocladus sp., Magnolia alternans Heer [Magnoliaephyllum alternans (Heer) Seward], Menispermites septentrionalis Hollick, Menispermites marcovoensis Philipp., Ettingshausenia embicola (Vachr.) Herman [Ettingshausenia louravetlanica (Herman et Shczepetov) Herman et Moiseeva], Credneria inordinata Hollick [Arthollia inordinata (Holl.) Herman или, скорее всего, Ettingshausenia louravetlanica], Leguminosites acuminata Philipp. [Myrtophyllum acuminata (Philipp.) Herman], Scheffleraephyllum venustum (Philipp.) Philipp., Grebenkia kryshtofovichii E. Lebed. [Grebenkia anadyrensis (Krysht.) E. Lebed.], Araliaephyllum devjatilovae Philipp., Celastrophyllum latifolium Font.
Проанализировав систематический состав тафофлор кривореченской свиты право- и левобережья р. Анадырь, Г.Г. Филиппова (Филиппова, Абрамова, 1993, с. 19) отмечает, что “все они составляют флористический комплекс, единство которого обусловлено общностью доминирующих родов и видов в составе папоротников (Gleichenites, Coniopteris, Birisia), хвойных (Cephalotaxopsis, Sequoia) и покрытосеменных (Menispermites, Ettingshausenia, Dalembia), присутствием древних мезофитных форм”. Таким образом, Филиппова относит перечисленные ископаемые растения к гребенкинской флоре, наиболее представительные местонахождения растительных остатков которой расположены на правобережье р. Анадырь (Щепетов и др., 1992; Филиппова, Абрамова, 1993; Spicer et al., 2002; Герман, 2011).
Кривореченская свита на левобережье р. Анадырь согласно перекрывается морскими отложениями дуговской свиты (рис. 1, 2), представленной песчаниками, алевролитами, аргиллитами и туффитами различного состава мощностью около 600 м (Верещагин, Терехова, 1960; Паракецов и др., 1974; Решения…, 1982; Терехова, Дундо, 1987; Терехова, 1988). Эти породы содержат остатки морских моллюсков, которые были изучены Г.П. Тереховой (1988): Inoceramus concentricus costatus Nagao et Mat., I. iburiensis Nagao et Mat., I. gibberosus Bodul., I. hobetsensis Nagao et Mat., Scaphites (Otoscaphites) puerculus teshioensis (Yabe), S. (O.) aff. puerculus Jimbo, S. (O.) ex gr. puerculus Jimbo, S. planus (Yabe), Scalarites sp. indet. По мнению Тереховой (1988, с. 110), “комплекс фаунистических остатков из дуговской свиты соответствует зоне Inoceramus iburiensis и одновозрастной ей зоне Jimboiceras planulatiforme Анадырско-Корякского региона”, для которых принят позднетуронский возраст (Решения…, 1982); “самое низкое стратиграфическое положение в дуговской свите занимают слои с Inoceramus concentricus costatus Nagao et Mat., которые выше… сменяются отложениями с Inoceramus iburiensis Nagao et Mat., I. gibberosus Bodul., а еще выше… – породами с… Inoceramus hobetsensis Nagao et Mat. и скафитами”. Исходя из этого, Терехова (1988, с. 111) “считает возраст фаунистического комплекса дуговской свиты позднетуронским, не исключая возможности отнесения самых нижних горизонтов свиты с Inoceramus concentricus costatus к нижнему турону”. При этом она отмечает, что “в бассейне р. Убиенка… континентальные отложения кривореченской свиты с остатками флоры непосредственно перекрываются морскими слоями с Inoceramus concentricus costatus. Севернее, в бассейнах рр. Крестовой и Чинейвеем…, выше континентальных слоев залегают морские отложения с Inoceramus hobetsensis”.
Последнее наблюдение, а также определенные Тереховой (1988) из верхней подсвиты кривореченской свиты моллюски позднеальбского–раннесеноманского возраста на р. Малая Гребенка (правобережье р. Анадырь: Щепетов и др., 1992) и среднесеноманского–(?) раннетуронского возраста на р. Лев. Ничеквеем, позволили ей сделать вывод о том, что возраст содержащей растительные остатки кривореченской свиты “варьирует в зависимости от возраста перекрывающих морских осадков. Верхняя возрастная граница изменяется от позднего альба до позднего турона” (Терехова, 1988, с. 113). С этим выводом о диахронности верхней границы кривореченской свиты и омоложении ее в направлении с юга на север согласен и В.П. Похиалайнен (1994), причем степень ее диахронности он считает еще более существенной, вплоть до конца коньякского века.
Дуговскую свиту согласно перекрывают песчаники и алевролиты крестовской свиты (рис. 2), которые по остаткам содержащихся в них морских моллюсков датируются ранним сеноном (Терехова, 1988) или сантоном (Похиалайнен, 1994).
Принципиально иного воззрения на соотношение кривореченской и дуговской свит придерживаются А.И. Дворянкин, Е.Л. Лебедев и Н.И. Филатова (Дворянкин и др., 1993), считающие границу этих свит (как их понимает Терехова) на всем или почти всем ее протяжении тектонической. Указанные авторы полагают, что “полоса отложений, ранее целиком относившихся к кривореченской свите… включает тектонические блоки пород с сенонской флорой” (Дворянкин и др., 1993, с. 475). Эта тафофлора, названная ими чинейвеемским флористическим комплексом, была обнаружена на правом берегу р. Чинейвеем между ее притоками – руч. Болотный и р. Обильная (рис. 3), в верхней части грубообломочной терригенной толщи, относимой Тереховой (1988) к кривореченской свите. По данным Дворянкина с соавторами, эти флороносные породы образуют “тектонический блок шириной около 4 км, ограниченный дугообразной формы разломами” (Дворянкин и др., 1993, с. 473). Однако геолог научно-производственного объединения “Аэрогеология” В.Л. Самсоненко, принимавший участие в полевых работах и сборах чинейвеемской тафофлоры, утверждает (устное сообщение, 2011 г.), что наблюдать эти тектонические контакты в поле не представляется возможным из-за плохой обнаженности района, закрытого тундровой растительностью. Остается предположить, что эти разломы были установлены Дворянкиным и его коллегами в результате дешифрирования аэрофотоснимков.
Рис. 3.
Схема геологического строения бассейна р. Чинейвеем и местонахождения ископаемых морских моллюсков и растений (по ГГК-200, листы Q-59-XXI, XXII, Б.Д. Трунов, 1972 г.: Государственная…, 1977).
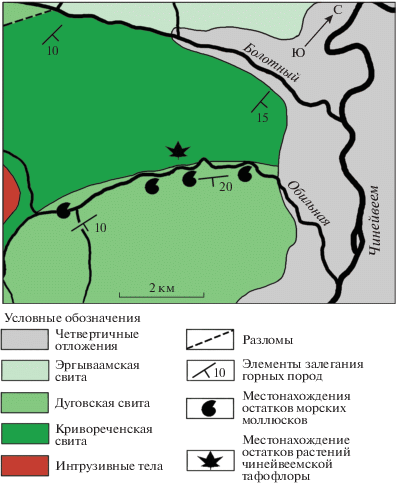
Терригенная толща, содержащая чинейвеемскую тафофлору, с юго-востока по разлому (по данным Дворянкина и соавторов) контактирует с отложениями, относимыми Тереховой (1988) к дуговской свите, в которых во время среднемасштабной геологической съемки Б.Д. Труновым были собраны Inoceramus cf. hobetsensis Nagao et Mat., Inoceramus sp. indet. (возможно, типа I. iburiensis Nagao et Mat.), Pholadomya sp. indet., Parallelodon cf. sachalinensis Schmidt., характерные для позднего турона, однако “тектонический характер контакта между флороносными отложениями и толщей с туронскими иноцерамами не позволяет корректировать возраст толщи, вмещающей чинейвеемский комплекс флоры, по упомянутым палеонтологическим данным” (Дворянкин и др., 1993, с. 474).
Возраст чинейвеемской тафофлоры Е.Л. Лебедев определяет по ее систематическому составу. Он отмечает ее более молодой облик по сравнению с гребенкинской флорой (из-за присутствия многочисленных Metasequoia, Trochodendroides и Arthollia) и сходство с пенжинской тафофлорой Северо-Западной Камчатки. На основании наличия в чинейвеемском комплексе “остатков листьев семейства Fagaceae, относящихся или к Quercus, или к новому роду”, Лебедев считает, что эта флора “несомненно моложе флор пенжинского… типа (поздний турон–коньяк)” (Дворянкин и др., 1993, с. 474–475), и датирует ее сеноном. Стратиграфическим аналогом толщи, содержащей чинейвеемскую тафофлору, цитированные авторы считают сенонскую крестовскую свиту (рис. 2).
МАТЕРИАЛ И МЕТОДЫ
Изученная нами коллекция была собрана Дворянкиным с коллегами при картировании участка правобережья р. Чинейвеем на левобережье р. Анадырь (рис. 3). Она состоит из 81 образца (штуфа) с отпечатками растений и полевой маркировкой “W1306”. При некоторых образцах имеются рукописные этикетки с определениями, сделанными Лебедевым. Материал представлен отпечатками листьев, облиственных побегов и репродуктивных органов (шишек), фитолеймы на них не сохранились. Эта коллекция № 5003 хранится в Геологическом институте (ГИН) РАН, г. Москва.
Ископаемые растения после очистки и препарирования изучались при помощи налобной бинокулярной лупы или под бинокулярным микроскопом Carl Zeiss SM-XX и фотографировались при помощи цифровой камеры Panasonic Lumix DMC-GX9, оснащенной макрообъективом Leica DG Macro-Elmarit 1:2.8/45 мм.
СИСТЕМАТИЧЕСКИЙ СОСТАВ И ВОЗРАСТ ЧИНЕЙВЕЕМСКОЙ ТАФОФЛОРЫ
Чинейвеемский флористический комплекс, получивший название по своему местонахождению в бассейне р. Чинейвеем (рис. 3), представлен 23 видами гинкговых, хвойных и покрытосеменных. В его состав входят следующие растения.
Ginkgoales:
Ginkgo ex gr. adiantoides (Unger) Heer (табл. I , фиг. 1);
Pinales:
Cephalotaxopsis intermedia Hollick (табл. I , фиг. 7, 8 ),
Sequoia rigida Heer (табл. I , фиг. 5, 6 ),
Metasequoia cuneata (Newberry) Chaney (табл. I , фиг. 9, 10 ),
Sequoia sp. (шишка) (табл. I , фиг. 2, 3),
Pityostrobus (?) sp. (табл. I , фиг. 4);
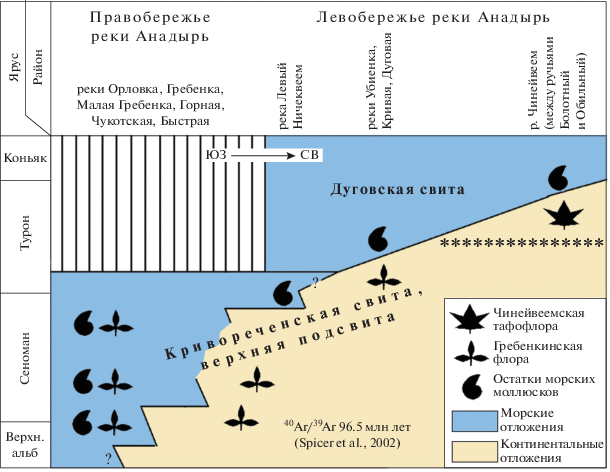
Рис. 4.
Соотношение континентальных флороносных отложений кривореченской свиты с морскими образованиями в среднем течении р. Анадырь; звездочками показана предполагаемая верхняя граница распространения гребенкинской флоры (по Герман, 2011, с изменениями).
Magnoliopsida:
Araliaephyllum cf. armanensis Herman (табл. I , фиг. 11 ),
Magnoliaephyllum ex gr. alternans (Heer) Seward (табл. II, фиг. 3),
Cissites cf. inscissus Herman (табл. II, фиг. 1),
Myrtophyllum (?) sp. (табл. II, фиг. 5 ),
Menispermites orientalis Golovneva (табл. II, фиг. 6 ),
Menispermites sibiricus (Heer) Golovneva (табл. II, фиг. 2),
Trochodendroides ex gr. arctica (Heer) Berry (табл. II, фиг. 9, 10 ),
Arthollia inordinata (Hollick) Herman (табл. III, фиг. 1),
Arthollia pacifica Herman (табл. IV, фиг. 3),
Ettingshausenia louravetlanica (Herman et Shczepetov) Herman et Moiseeva (табл. IV, фиг. 1, 2),
Ettingshausenia primaeva (Lesquereux) Herman (табл. III, фиг. 2, 3),
платаноидные репродуктивные структуры (табл. III, фиг. 4; табл. IV, фиг. 4, 5 ),
Celastrophyllum orientalis Philippova (табл. IV, фиг. 7 ),
Viburniphyllum whymperi (Heer) Herman (табл. IV, фиг. 8 ),
Grebenkia cf. anadyrensis (Kryshtofovich) E. Lebedev (табл. II, фиг. 7, 8 ),
Parvileguminophyllum (?) sp. (табл. II, фиг. 4),
Chachlovia dombeyopsoida (Herman) Herman (табл. IV, фиг. 6 ).
Таблица I . Гинкговые, хвойные и покрытосеменные чинейвеемского комплекса; левобережье р. Анадырь, бассейн р. Чинейвеем; кривореченская свита, верхний мел, турон. Длина масштабной линейки 1 см. 1 – Ginkgo ex gr. adiantoides (Unger) Heer, экз. 5003/65; 2, 3 – Sequoia sp., шишка: 2 – экз. 5003/70, 3 – экз. 5003/1-2; 4 – Pityostrobus (?) sp., экз. 5003/3; 5, 6 – Sequoia rigida Heer: 5 – экз. 5003/51, 6 – экз. 5003/53; 7, 8 – Cephalotaxopsis intermedia Hollick: 7 – экз. 5003/2-2, 8 – экз. 5003/2-1; 9, 10 – Metasequoia cuneata (Newberry) Chaney: 9 – экз. 5003/5, 10 – экз. 5003/25-1; 11 – Araliaephyllum cf. armanensis Herman, листочек сложного листа, экз. 5003/10.

Таблица II. Покрытосеменные чинейвеемского комплекса; левобережье р. Анадырь, бассейн р. Чинейвеем; кривореченская свита, верхний мел, турон. Длина масштабной линейки 1 см. 1 – Cissites cf. inscissus Herman, экз. 5003/61; 2 – Menispermites sibiricus (Heer) Golovneva, экз. 5003/68; 3 – Magnoliaephyllum ex gr. alternans (Heer) Seward, экз. 5003/57-2; 4 – Parvileguminophyllum (?) sp., экз. 5003/22-1; 5 – Myrtophyllum (?) sp., экз. 5003/50-2; 6 – Menispermites orientalis Golovneva, экз. 5003/23; 7, 8 – Grebenkia cf. anadyrensis (Kryshtofovich) E. Lebedev: 7 – экз. 5003/50-1, 8 – экз. 5003/35-1; 9, 10 – Trochodendroides ex gr. arctica (Heer) Berry: 9 – экз. 5003/59, 10 – экз. 5003/77.

Таблица III. Покрытосеменные чинейвеемского комплекса; левобережье р. Анадырь, бассейн р. Чинейвеем; кривореченская свита, верхний мел, турон. Длина масштабной линейки 1 см. 1 – Arthollia inordinata (Hollick) Herman, экз. 5003/18; 2, 3 – Ettingshausenia primaeva (Lesquereux) Herman: 2 – экз. 5003/73, 3 – экз. 5003/8; 4 – платаноидная репродуктивная структура, экз. 5003/26.

Таблица IV. Покрытосеменные чинейвеемского комплекса; левобережье р. Анадырь, бассейн р. Чинейвеем; кривореченская свита, верхний мел, турон. Длина масштабной линейки 1 см. 1, 2 – Ettingshausenia louravetlanica (Herman et Shczepetov) Herman et Moiseeva: 1 – экз. 5003/79, 2 – экз. 5003/27; 3 – Arthollia pacifica Herman, экз. 5003/67; 4, 5 – платаноидная репродуктивная структура, экз. 5003/26; 6 – Chachlovia dombeyopsoida (Herman) Herman, экз. 5003/30; 7 – Celastrophyllum orientalis Philippova, экз. 5003/78; 8 – Viburniphyllum whymperi (Heer) Herman, экз. 5003/43.

В чинейвеемской тафофлоре отсутствуют мохообразные, папоротники, лептостробовые, цикадовые, беннеттитовые и кейтониевые растения. Гинкговые представлены единственным фрагментарным отпечатком Ginkgo ex gr. adiantoides. Хвойные многочисленны, наиболее часто встречаются облиственные побеги Sequoia rigida и Metasequoia cuneata, реже – побеги Cephalotaxopsis intermedia. Генеративные органы хвойных представлены единичными находками ископаемых шишек таксодиевых и, возможно, сосновых. Наиболее разнообразны и многочисленны по количеству отпечатков покрытосеменные растения, включающие 17 видов из 14 родов. Из них часто встречаются Cissites cf. inscissus, Trochodendroides ex gr. arctica, Viburniphyllum whymperi и платанообразные, представленные ископаемыми листьями Arthollia inordinata, A. pacifica и Ettingshausenia louravetlanica, E. primaeva, а также единственной находкой отпечатка платаноидного репродуктивного органа – сидящих на оси головчатых структур. По мнению Н.П. Масловой (устное сообщение, 2020 г.), сохранность их недостаточна для того, чтобы точно определить, являются они соплодиями или мужскими соцветиями. Нельзя исключить, что эти головчатые структуры близки или идентичны сложным соплодиям Anadyricarpa altingiosimila N. Maslova et Herman из кривореченской свиты на р. Гребенка (правобережье р. Анадырь), находки которых ассоциируются с ископаемыми листьями Ettingshausenia louravetlanica (Маслова, Герман, 2004). Остатки остальных покрытосеменных малочисленны или единичны.
Как отмечалось выше, по данным А.Д. Девятиловой и Г.П. Тереховой (Девятилова и др., 1980; Терехова, 1988), чинейвеемская тафофлора происходит из верхов кривореченской свиты, к отложениям которой приурочены также многочисленные находки ископаемых растений богатой и разнообразной гребенкинской флоры (Щепетов и др., 1992; Филиппова, Абрамова, 1993; Spicer et al., 2002; Герман, 2011; Herman, 2013). Чинейвеемский комплекс обнаруживает определенное сходство с этой флорой, в состав которой входит ряд видов, идентичных или близких к чинейвеемским: Ginkgo ex gr. adiantoides, Cephalotaxopsis intermedia, Magnoliaephyllum alternans, Menispermites ex gr. sibiricus, Ettingshausenia louravetlanica, Arthollia cf. pacifica, Trochodendroides ex gr. arctica, Cissites cf. inscissus, Grebenkia anadyrensis. Однако различие этих флор существенно: из доминантов чинейвеемской тафофлоры хвойные Metasequoia в гребенкинской флоре отсутствуют, покрытосеменные Trochodendroides, Ettingshausenia и Arthollia встречаются единично либо сравнительно редко. Перечисленные растения, однако, становятся обычными компонентами и, как правило, многочисленны в более молодых, чем гребенкинская, флорах Северо-Востока Азии. Наоборот, многие характерные растения гребенкинской флоры, являющиеся ее обязательными компонентами, в чинейвеемской отсутствуют. Показательно также, что в последней листья покрытосеменных с цельным краем очень редки, тогда как в гребенкинской флоре они составляют ее заметный компонент. В состав чинейвеемской тафофлоры входят растения, известные из туронских и более молодых флор региона: Metasequoia cuneata, Araliaephyllum cf. armanensis, Arthollia inordinata, Celastrophyllum orientalis, Viburniphyllum whymperi, Chachlovia dombeyopsoida. Обращает внимание доминирование в чинейвеемской тафофлоре крупнолистных платанообразных (четыре вида из двух родов), наибольший расцвет которых приходится на Северо-Востоке Азии на турон (Герман, 2011). Своеобразие чинейвеемской флоре придает отсутствие в ее составе папоротников, цикадовых, беннеттитовых и кейтониевых растений, обычно встречающихся в меловых флорах Северо-Востока Азии.
По нашему мнению, чинейвеемская тафофлора, безусловно, более молодая, чем гребенкинская флора, и наиболее близка к пенжинской тафофлоре Северо-Западной Камчатки и п-ова Елистратова, возраст которой надежно установлен как турон, исключая начало турона (Герман, Лебедев, 1991; Герман, 2011; Herman, 2013). Об одновозрастности пенжинской и чинейвеемской флор свидетельствуют (1) их сходство на видовом (Cephalotaxopsis intermedia, Metasequoia cuneata, Magnoliaephyllum ex gr. alternans, Cissites cf. inscissus, Menispermites sibiricus, M. orientalis, Trochodendroides ex gr. arctica, Arthollia inordinata, A. pacifica, Ettingshausenia primaeva, Viburniphyllum whymperi, Grebenkia cf. anadyrensis) и родовом (Sequoia, Pityostrobus, Araliaephyllum, Myrtophyllum, Celastrophyllum) уровнях; (2) многочисленность в них растений родов Metasequoia, Trochodendroides, Menispermites; (3) значительное разнообразие и количественное доминирование крупнолистных платанообразных покрытосеменных.
Вывод Е.Л. Лебедева о сенонском возрасте чинейвеемского комплекса (Дворянкин и др., 1993) основывается главным образом на присутствии в нем листьев, отнесенных к Fagophyllum sp. (gen. nov.?) современного семейства Fagaceae. Однако такое определение представляется нам ошибочным. Немногочисленные фрагментарные отпечатки этих листьев, обладающие плохой сохранностью, по нашему мнению, близки к гребенкинскому виду Grebenkia anadyrensis, но отличаются от него более крупными размерами листовой пластинки. Следует отметить, что Лебедев, скорее всего, и сам сомневался в определении Fagophyllum sp. (gen. nov.?): на сохранившихся этикетках к этим отпечаткам его рукой написано “Grebenkia (?)”. Сказанное позволяет нам считать чинейвеемскую тафофлору примерно одновозрастной пенжинской и датировать ее туронским веком, скорее всего поздним туроном.
Определенное сходство чинейвеемская тафофлора обнаруживает с флористическими комплексами следующего, кайваямского, этапа развития флоры Анадырско-Корякского субрегиона. Этот этап представлен кайваямским комплексом Северо-Западной Камчатки и п-ова Елистратова, поперечнинским комплексом восточного склона хребта Пекульней и тыльпегыргынайским комплексом западного склона хребта Пекульней (Герман, Лебедев, 1991; Герман, 2011; Herman, 2013; Craggs, 2005; Филиппова, 2010). Отложения, вмещающие растительные остатки этих комплексов, надежно датируются коньякским веком (кайваямский комплекс) или ранним коньяком (поперечнинский и тыльпегыргынайский комплексы) по соотношению с морскими слоями, содержащими остатки моллюсков. Как и для чинейвеемской тафофлоры, для перечисленных флористических комплексов характерно значительное участие хвойных Metasequoia и покрытосеменных Trochodendroides, а также крупнолистных платанообразных. При этом общих с чинейвеемской тафофлорой видов в них меньше, чем в пенжинской флоре: это лишь Ginkgo ex gr. adiantoides, Cephalotaxopsis intermedia, Metasequoia cuneata, Magnoliaephyllum alternans, Menispermites sibiricus, Arthollia pacifica, Cissites inscissus, Viburniphyllum whymperi, Chachlovia dombeyopsoida. Сказанное позволяет считать возраст чинейвеемской тафофлоры более древним, чем коньякский век.
ЗАКЛЮЧЕНИЕ
Отвечая на стоявшие перед авторами вопросы, сформулированные во введении к этой статье, мы можем утверждать следующее.
1. Чинейвеемская тафофлора характеризуется невысоким разнообразием входящих в нее растений, представленных гинкговыми, хвойными и покрытосеменными. Судя по их составу, она моложе гребенкинской флоры и наиболее близка к пенжинской флоре Северо-Западной Камчатки и п-ова Елистратова. Поэтому чинейвеемскую тафофлору мы считаем примерно одновозрастной пенжинской и определяем ее возраст как туронский, скорее всего позднетуронский.
Следует отметить, что В.Ф. Белый (устное сообщение, 1998 г.) обратил внимание авторов на то, что на левобережье р. Чинейвеем, севернее захоронения чинейвеемской тафофлоры (рис. 3), в дуговской свите, содержащей обильные фаунистические остатки позднетуронского возраста, появляются континентальные фации с отпечатками ископаемых растений. По мнению Белого, нельзя в принципе исключить, что и само местонахождение чинейвеемской тафофлоры принадлежит дуговской, а не кривореченской (как полагает Г.П. Терехова) свите.
2. Имеются две взаимоисключающие точки зрения на соотношение кривореченской и дуговской свит левобережья р. Анадырь: а) их контакт постепенный, согласный без следов перерыва (Терехова, 1988; устное сообщение, 1983 г.; рис. 3); б) контакт между свитами в этом районе тектонический (Дворянкин и др., 1993). По результатам изучения коллекции остатков растений чинейвеемской тафофлоры мы поддерживаем точку зрения Г.П. Тереховой. Как и показано на карте Б.Н. Трунова (Государственная…, 1977), местонахождение чинейвеемского комплекса расположено в самой верхней части разреза кривореченской свиты близ подошвы дуговской свиты. Наш вывод о туронском (вероятно, позднетуронском) возрасте чинейвеемской тафофлоры вполне отвечает представлениям о позднетуронском или коньякском возрасте морских моллюсков из перекрывающих слоев дуговской свиты на р. Чинейвеем (Терехова, 1988; Похиалайнен, 1994) и согласном стратиграфическом контакте этих слоев с флороносными отложениями.
Отметим, что наша датировка чинейвеемского комплекса лишает точку зрения А.И. Дворянкина с соавторами ее наиболее серьезного – палеоботанического – доказательства или, может быть, самого основания. Кажется вполне вероятным, что представления о тектоническом контакте двух свит появились ввиду необходимости отделить “более молодые” флороносные отложения от “более древних” морских слоев с остатками моллюсков. Как уже отмечалось, участник полевых работ на р. Чинейвеем В.Л. Самсоненко данный разлом не наблюдал. После выхода статьи Дворянкина с соавторами (1993) С.В. Щепетов проанализировал все опубликованные и фондовые материалы по описанному району, провел дешифрирование аэрофотоснимков и никаких свидетельств тектонических разломов, отделяющих отложения с чинейвеемской тафофлорой от кривореченской и дуговской свит, не обнаружил. Этих разломов не выявил и В.Ф. Белый – признанный мастер дешифрирования аэрофотоснимков (Белый, Белая, 1998, рис. 4).
3. Рассмотренные выше данные о соотношении флороносных слоев кривореченской свиты левобережья р. Анадырь с перекрывающими морскими отложениями и о составе флористических комплексов позволяют, на наш взгляд, считать тафофлоры с рек Убиенка, Кривая и Дуговая аналогом классической гребенкинской флоры правобережья р. Анадырь и датировать их сеноманом и, возможно, ранним туроном, а чинейвеемскую тафофлору самых верхов кривореченской свиты, выделяя ее из состава гребенкинской флоры, считать более молодой (вероятнее всего, позднетуронской). Иными словами, последовательность тафофлор из кривореченской свиты в междуречье Убиенка–Чинейвеем позволяет ограничить верхний возрастной предел распространения гребенкинской флоры началом туронского века. Следовательно, левобережье р. Анадырь интересно прежде всего тем, что здесь удается проследить в едином разрезе смену тафофлор гребенкинского этапа развития флор Анадырско-Корякского субрегиона таковыми пенжинского этапа. Представления авторов о соотношении морских и континентальных меловых отложений среднего течения р. Анадырь, содержащих гребенкинскую и чинейвеемскую флоры, показано на рис. 4.
Благодарности. Авторы искренне признательны Г.П. Тереховой и В.Л. Самсоненко за важные сведения по геологии левобережья р. Анадырь, Н.П. Масловой (Палеонтологический институт РАН, Москва) за ценные советы относительно платаноидного репродуктивного органа, а также Б.Н. Шурыгину (Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, Новосибирск) и Е.В. Бугдаевой (ФНЦ Биоразнообразия ДВО РАН, Владивосток), чьи замечания к рукописи этой статьи позволили нам существенно ее улучшить.
Источники финансирования. Работа выполнена в рамках тем государственного задания Геологического института РАН и Ботанического института РАН и при поддержке Российского фонда фундаментальных исследований, грант № 19-05-00121.
Список литературы
Белый В.Ф., Белая Б.В. Поздняя стадия развития Охотско-Чукотского вулканогенного пояса (верхнее течение р. Энмываам). Магадан: СВКНИИ ДВО РАН, 1998. 108 с.
Верещагин В.Н., Терехова Г.П. Морские верхнемеловые отложения на левобережье р. Анадырь, севернее пос. Марково // Материалы по геологии и полезным ископаемым Северо-Востока СССР. Магадан: Севвостокгеология, 1960. Вып. 14. С. 198–200.
Герман А.Б. Этапность и цикличность развития позднемеловой флоры Анадырско-Корякского субрегиона (Северо-Восток России) и их связь с климатическими изменениями // Стратиграфия. Геол. корреляция. 1993. Т. 1. № 1. С. 87–96.
Герман А.Б. Альбская–палеоценовая флора Северной Пацифики. М.: ГЕОС, 2011. 280 с. (Труды Геол. ин-та РАН. Вып. 592).
Герман А.Б., Лебедев Е.Л. Стратиграфия и флора меловых отложений Северо-Западной Камчатки. М.: Наука, 1991. 189 с. (Труды Геол. ин-та АН СССР. Вып. 468).
Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Анадырская. Лист Q-59-XXI, XXII. Объяснительная записка. Составитель Трунов Б.Д. Магадан, 1977. 69 с.
Дворянкин А.И., Лебедев Е.Л., Филатова Н.И. Чинейвеемский сенонский флористический комплекс на Северо-Востоке России (правобережье р. Анадырь) и его геологическое значение // Докл. АН. 1993. Т. 333. № 4. С. 473–476.
Девятилова А.Д., Невретдинов Э.Б., Филиппова Г.Г. Стратиграфия верхнемеловых отложений бассейна среднего течения р. Анадырь // Геология и геофизика. 1980. № 12. С. 62–70.
Маслова Н.П., Герман А.Б. Новые находки ископаемых Hamamelidales и проблема филогенетических взаимоотношений платановых и гамамелидовых // Палеонтол. журн. 2004. № 5. С. 94–105.
Моисеева М.Г. Новые данные о сеноманской флоре бухты Угольной (Северо-Восток России) // Палеонтол. журн. 2010. № 2. С. 100–110.
Паракецов К.В., Похиалайнен В.П., Терехова Г.П. Биостратиграфическое расчленение меловых отложений Анадырско-Корякского региона // Основные проблемы биостратиграфии и палеогеографии Северо-Востока СССР. Ч. 2. Мезозой. Владивосток: ДВНЦ АН СССР, 1974. С. 196–227. (Тр. СВКНИИ ДВНЦ АН СССР. Вып. 63).
Похиалайнен В.П. Мел Северо-Востока России. Магадан: СВКНИИ ДВО РАН, 1994. 37 с.
Решения 2-го Межведомственного стратиграфического совещания по мелу, палеогену и неогену Корякского нагорья, Камчатки, Командорских островов и Сахалина (Петропавловск-Камчатский, 1974). Петропавловск-Камчатский: ПГО “Камчатгеология”, 1982. 131 с.
Решения Третьего Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России (Санкт-Петербург, 2002). СПб.: Изд-во ВСЕГЕИ, 2009. 268 с.
Самылина В.А. Раннемеловые флоры Северо-Востока СССР (К проблеме становления флор кайнофита) // ХХVII Комаровские чтения. Л.: Наука, 1974. 56 с.
Терехова Г.П. О возрасте кривореченской свиты и гребёнкинского флористического комплекса // Стратиграфия и палеонтология фанерозоя Северо-Востока СССР. Магадан: СВКНИИ ДВО АН СССР, 1988. С. 100–117.
Терехова Г.П., Дундо О.П. Анадырско-Корякская область // Стратиграфия СССР. Меловая система. М.: Недра, 1987. 2-й полутом. С. 137–154.
Филиппова Г.Г. Сеноманская флора р. Гребенка и ее значение для стратиграфии // Дальневосточная палеофлористика. Владивосток: ДВНЦ АН СССР, 1979. С. 91–115. (Тр. Биол-почв. ин-та ДВНЦ АН СССР. Нов. серия. Т. 53 (156)).
Филиппова Г.Г. Новые меловые покрытосеменные из бассейна среднего течения р. Анадырь // Материалы по геологии и полезным ископаемым Северо-Востока СССР. Магадан: Севвостокгеология, 1982. Вып. 26. С. 69–75.
Филиппова Г.Г. Меловые хвойные из бассейна среднего течения р. Анадырь // Стратиграфия и палеонтология палеозойских и мезозойских отложений Северо-Востока СССР. М.: Геол. фонд РСФСР, 1984. С. 154–174.
Филиппова Г.Г. Стратиграфия и флора меловых отложений северной части хребта Пекульней (Чукотка). Магадан: СВКНИИ ДВО РАН, 2010. 203 с.
Филиппова Г.Г., Абрамова Л.Н. Позднемеловая флора Северо-Востока России. М.: Недра, 1993. 348 с.
Щепетов С.В., Герман А.Б., Белая Б.В. Среднемеловая флора правобережья реки Анадырь (стратиграфическое положение, систематический состав, атлас ископаемых растений). Магадан: СВКНИИ ДВО АН СССР, 1992. 166 с.
Craggs H.J. Late Cretaceous climate signal of the Northern Pekulney Range flora of northeastern Russia // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005. V. 217. P. 25–46.
Herman A.B. Albian–Paleocene flora of the North Pacific: systematic composition, palaeofloristics and phytostratigraphy // Stratigr. Geol. Correl. 2013. V. 21. № 7. P. 689–747.
Spicer R.A., Ahlberg A., Herman A.B., Kelley S.P., Raikevich M., Rees P.M. Palaeoenvironment and ecology of the middle Cretaceous Grebenka flora of northeastern Asia // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002. V. 184. № 1–2. P. 65–105.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция