

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 5, стр. 59-84
Опыт глубоководного бурения в Мировом океане: методическое и практическое значение для стратиграфических исследований
А. Ю. Гладенков 1, *, Ю. Б. Гладенков 1, **
1 Геологический институт РАН
Москва, Россия
* E-mail: agladenkov@ilran.ru
** E-mail: gladenkov@ginras.ru
Поступила в редакцию 07.12.2020
После доработки 27.12.2020
Принята к публикации 25.01.2021
Аннотация
Приводятся результаты стратиграфических, прежде всего биостратиграфических, исследований, проведенных в процессе осуществления Международного проекта по глубоководному бурению за последние 50 лет. Они вносят весомый вклад в разработку и совершенствование методики детальных стратиграфических исследований и определения возраста морских комплексов осадочных пород, а также в реконструкции океанологических и климатических событий прошлого. Полученные результаты имеют важное методическое значение для проведения стратиграфических работ по всему фанерозою. Показано, что выделенные зональные подразделения кайнозоя могут быть реально прослежены на громадной территории – по всей тропическо-субтропической области. Отмечено, что в этих работах значительную роль сыграло изучение комплексов планктонных микроорганизмов (карбонатных и кремнистых), которые являлись неотъемлемой частью мезозойских и кайнозойских морских экосистем. Развитие этих ассоциаций во времени проходило на фоне смены режимов океанической циркуляции и осадконакопления, изменения степени продуктивности глубинных и поверхностных вод, их температур и других характеристик. Направленность и тенденции эволюции биотических сообществ в целом отражают этапы развития океанических экосистем прошлого и происходивших в них перестроек. Все это позволяет выявить реальную картину последовательности не только биотических, но и абиотических (климатических, океанографических, палеогеографических, эвстатических) событий в Мировом океане для последних 70–75 млн лет.
ВВЕДЕНИЕ
В истории стратиграфических исследований имеются, пожалуй, два международных проекта, которые сыграли особенно важную роль в развитии стратиграфии, как одного из значительных разделов геологии. Первый проект был реализован в конце XIX века. Тогда была поставлена задача – создать геологическую карту Европы. Чтобы приступить к ее практическому решению, геологи разных стран в 1878 г. собрали 1-й Международный геологический конгресс (МГК) и образовали специальную комиссию, поручив ей разработать Международную стратиграфическую шкалу (МСШ). Комиссия сосредоточила свое внимание на создании универсальной стратиграфической классификации и номенклатуры, опираясь на накопленный к тому времени опыт стратиграфических работ европейских стран (Леонов, 1973; Меннер, 1991). Эта работа заняла более 20 лет, и в 1900 г. на 8-й сессии МГК произошло утверждение МСШ, которая, таким образом, в 2020 г. отмечает свой 120-летний юбилей. Именно на основе этой шкалы и была создана геологическая карта Европы, а позднее – карты отдельных регионов других континентов и мира. В упомянутой шкале была предложена определенная иерархия основных стратиграфических подразделений (эратемы, системы и отделы), которая просуществовала до середины 70-х годов XX века. И только в 1970-е годы в нее были добавлены ярусы (ранее они относились к региональным категориям), что сделало шкалу значительно детальнее. Трудно переоценить значение МСШ как геологического документа мирового масштаба. Прежде всего, это замечательное геологическое обобщение, отразившее определенную этапность развития Земли и ее биосферы, которое продолжалось свыше 4 млрд лет. С другой стороны, она определила методическую базу расчленения древних толщ разных областей и их корреляций. В ее основу лег историко-геологический подход к выделению стратиграфических подразделений с важной ролью палеонтологического метода в их обосновании. Наконец, МСШ оказалась незаменимым средством профессионального общения геологов разных стран мира. Прошло около 120 лет после ее создания, а МСШ, которая к настоящему времени усовершенствована под эгидой Международной комиссии по стратиграфии, продолжает играть важнейшую роль во всех геологических дисциплинах, которые связаны с расшифровкой истории нашей планеты.
Вторым важным международным геологическим проектом прошлого столетия, который также оказал значительное влияние на стратиграфические исследования, явился Проект глубоководного бурения в Мировом океане, который, как известно, занимает более 70% поверхности планеты, но который оставался к середине XX столетия геологически крайне слабо изученным. Проект был начат в 1968 г., и в 2018 г. международным научным сообществом было отмечено 50-летие его старта. Этот гигантский геологический эксперимент по изучению строения морского дна дал возможность получить большой объем материалов о вещественном составе и возрасте отложений осадочного чехла, а также истории геологического развития океанов в целом. К настоящему времени в рамках международного проекта Deep Sea Drilling Project (1968–1983 гг.), а также международных программ Ocean Drilling Program (1985–2003 гг.), Integrated Ocean Drilling Program (2004–2012 гг.) и International Ocean Discovery Program (2013 г.–настоящее время) на морском дне пробурено более 3000 глубоководных скважин (рис. 1). За прошедшие годы произошло значительное усовершенствование технологий бурения и извлечения керна, а также модернизация и увеличение количества судовых буровых платформ. Это позволяет сейчас проводить бурение при высоком проценте выхода керна практически во всех районах Мирового океана и в породах различной степени плотности. Современные технологии дали возможность достигать глубин бурения до 1500 м и проводить его при глубине моря до 4000 м. Более того, судно последнего поколения “Chikyu” (“Чикю”), построенное в Японии, в целом способно бурить скважины до глубины 10 000 м ниже уровня моря и более 2000 м ниже дна моря. В 2012 г. им уже осуществлено бурение скважин при глубине воды более 6960 м, а также на глубину 2111 м ниже морского дна. Краткие обзоры, в которых представлены полезные сведения по истории, основным результатам и перспективам бурения в океанах, даны в разные годы в статьях И.А. Басова (2001), Н.К. Рубаник (2008) и А.Г. Матуля (2010). Широкий спектр современных научных и технических достижений глубоководного бурения регулярно освещается в специальном журнале “Scientific Drilling”, который издается с 2005 г.
Рис. 1.
Карта точек бурения в Мировом океане (https://iodp.tamu.edu/scienceops/maps.html). 1–4 – глубоководные скважины: 1 – проекта DSDP (рейсы 1–96); 2 – программы ODP (рейсы 100–210); 3 – программы Integrated Ocean Drilling Program (экспедиции 301–348), 4 – программы International Ocean Discovery Program (экспедиции 349–371).
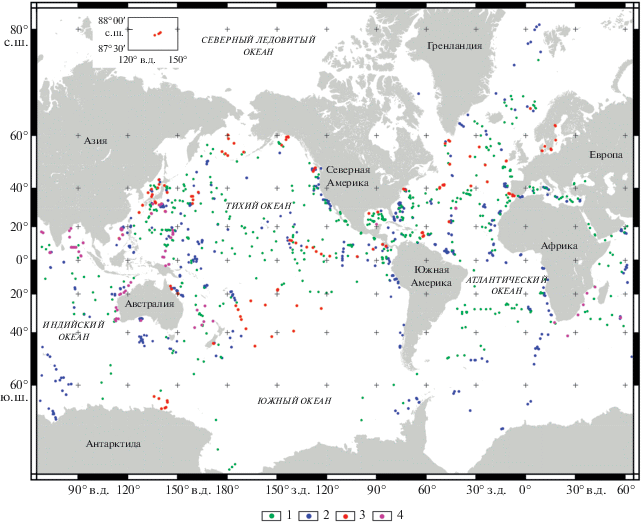
Необходимо подчеркнуть, что проводимые работы имеют международный характер: в их реализации участвовали ученые многих стран мира, а полученные материалы доступны всем заинтересованным специалистам. Эти исследования следует рассматривать как огромное научное достижение не только в отношении изучения строения морского дна и геологической истории океанов, но и для разработки и совершенствования методики стратиграфических исследований. Кроме того, они предоставили богатый материал для реконструкций геологических событий и изменений природных обстановок прошлых эпох. Выбор актуальных направлений исследований и их реализация привели в итоге к появлению как комплексных научных материалов каждой экспедиции и многочисленных региональных сводок, так и крупных обобщений (например, Океанология…, 1980; Kennett, 1982; Seibold, Berger, 1982; The Miocene…, 1985; Plankton…, 1985; Литвин, 1987; Удинцев, 1987; Productivity…, 1989; Geological…, 1990 и др.).
Отметим, что советские и российские специалисты (в том числе в области стратиграфии) до 1993 г. принимали достаточно активное участие как непосредственно в научных рейсах буровых судов, так и в обработке полученных в них материалов. Среди стратиграфов это В.А. Крашенинников, И.А. Басов, А.П. Жузе, Н.И. Стрельникова, М.Г. Петрушевская, Е.Д. Заклинская, Н.Г. Музылев, В.В. Шилов, А.Ю. Гладенков и другие. К сожалению, затем в силу разных причин такое участие прекратилось, и Россия в отличие от СССР не входит в число стран (ныне 26), участвующих в международной программе глубоководного бурения.
Следует специально заметить, что керновые материалы всех пробуренных скважин хранятся в нескольких специальных депозитариях, расположенных в США, Германии и Японии. Они доступны для изучения ученым различных стран.
В настоящей статье авторы хотели бы с учетом полученных при глубоководном бурении стратиграфических данных остановиться на основных методических и практических подходах, использованных при стратиграфических исследованиях палеогена и неогена, которые могут оказаться полезными для решения стратиграфических вопросов не только кайнозоя, но и других частей фанерозойской шкалы – мезозоя и палеозоя.
МЕТОДИКА БИОСТРАТИГРАФИЧЕСКОГО РАСЧЛЕНЕНИЯ, ОПРЕДЕЛЕНИЕ ВОЗРАСТА И КОРРЕЛЯЦИЯ МОРСКИХ ОСАДОЧНЫХ ТОЛЩ РАЗРЕЗОВ ОКЕАНИЧЕСКОГО ДНА
До начала глубоководного бурения знания ученых о вещественном составе и строении океанического дна были достаточно ограниченными и основывались, прежде всего, на результатах исследования образцов, отобранных из самого верхнего, молодого, слоя осадков (мощностью до нескольких метров) с помощью донных трубок или в результате драгирования отдельных участков дна.
Развернувшееся с конца 1960-х годов регулярное бурение глубоководных скважин дало возможность получать огромный объем материалов по разрезам океанических донных толщ мощностью в сотни метров, вскрытых в различных климатических зонах и районах Мирового океана. Полученные данные показали, что осадочные отложения океанического дна имеют в основном кайнозойский возраст. Реже отмечены осадки мела и в некоторых случаях – юры; при этом самые древние, среднеюрские, толщи с возрастом более 150 млн лет вскрыты в окраинных районах Тихого и Атлантического океанов. Осадочные отложения залегают на океаническом фундаменте, обычно сложенном мезозойскими вулканогенными породами (базальтами). Возраст древнейших (батско-келловейских) осадочных отложений установлен по радиоляриям в разрезе, вскрытом в котловине Пигафетта, Западная Пацифика (Matsuoka, 1992). Основным предметом нашего анализа служили прежде всего кайнозойские толщи.
Значение микропалеонтологических данных
Проводимое бурение позволило получить данные по ненарушенным колонкам образцов в разрезах морских толщ (в первую очередь кайнозоя), сложенных разнофациальными отложениями различной мощности. Как показала практика, попытки коррелировать эти толщи в масштабе океанов на литостратиграфической основе оказались несостоятельными и самым эффективным методом для этого оказался биостратиграфический, основанный на изучении смены в разрезах ископаемых микроорганизмов. При изучении разрезов глубоководных скважин было выявлено, что остатки морских микроорганизмов в донных отложениях находятся практически повсеместно. Они представили целый “мир” древней биоты, ранее недостаточно изученный. Микрофоссилии начали использоваться в стратиграфических целях еще в первой половине XX века. Однако наземные разрезы морских осадков, которые изучались в то время, не всегда были полными и непрерывными, а также содержали преимущественно бентосные мелководные организмы. Поэтому прослеживание в них смены типично морских комплексов микробиоты (с выделением маркирующих форм и определением интервалов их стратиграфического распространения), а также проведение их корреляций сталкивались с большими трудностями. В отличие от толщ, сформировавшихся в окраинно-океанических и приконтинентальных районах, отложения открытого океана в целом представлены достаточно выдержанными по латерали фациями, сложенными в основном биогенными осадками и глубоководными глинами, имеющими сравнительно небольшие мощности. В ходе обработки палеонтологических материалов из них было установлено, что использование прежде всего микрофоссилий дает наиболее эффективные результаты по расчленению осадочных толщ кайнозоя и мезозоя. Изучение пробуренных осадочных толщ, с одной стороны, позволило проследить в относительно полных разрезах пелагических фаций в различных регионах последовательную стратиграфическую смену комплексов микроорганизмов. С другой стороны, были проведены оценки интервалов стратиграфического распространения и ареалов многих ископаемых форм (в том числе ранее неизвестных в наземных разрезах), что дало возможность определить их важность для биостратиграфического расчленения и корреляции. При этом наибольший интерес представляли данные по микропланктонным организмам, так как они быстро эволюционируют и имеют широкое географическое распространение. Важно, что частая встречаемость микрофоссилий в породах и их распределение в разрезах без крупных перерывов обеспечивает послойную характеристику исследуемых отложений. Вот почему изучение микроостатков в итоге позволяет осуществлять дробное расчленение и широкие корреляции древних толщ. При этом результаты были получены на основе исследований микропланктона как карбонатного (в первую очередь планктонных фораминифер и наннопланктона), особенно характерного для тропических и субтропических широт, так и кремнистого (диатомей, радиолярий), типичного для бореального и натального поясов (хотя кремнистые микроорганизмы развиты и в экваториальном поясе, где в ряде случаев могут даже доминировать в планктоне). Специально следует отметить, что большой вклад в эти исследования внесло совершенствование оборудования для идентификации микроостатков, в частности широкое внедрение в практику с 1970-х годов электронной микроскопии. Это позволило добиться существенного прогресса в изучении ультраструктуры и морфологических особенностей строения скелетов, раковин и панцирей, что привело не только к выделению новых родов и видов, но и к ревизии классификации и таксономии различных палеонтологических групп.
Зональные подразделения и методика их выделения
Следует подчеркнуть, что во многом именно на материалах глубоководного бурения была отработана методика выделения таких стратиграфических подразделений, как биостратиграфические зоны, с последующим широким их внедрением в геологическую практику. Хотя, как известно, первые зональные схемы юры по аммонитам были созданы еще в середине XIX в. А. Оппелем, многие методические вопросы их составления долгие годы оставались не освещенными (Степанов, Месежников, 1979; Гладенков, 2004, 2010). Изучение в разрезах глубоководных скважин разных океанов смены ископаемых комплексов различных групп микроорганизмов кайнозоя предоставило реальную основу для разработки детальных океанических схем и шкал, представляющих собой последовательности смыкающихся зональных подразделений. В результате были разработаны шкалы, состоящие из зон с продолжительностью в среднем от 1–2 до 0.1–0.2 млн лет, которые выделяются с учетом этапности эволюции ископаемых микроорганизмов. При этом необходимо отметить, что при расчленении океанических разрезов была отработана методика выделения зон различного типа. В Международном стратиграфическом руководстве (International…, 1999) и Стратиграфическом кодексе России (2019) этим типам дана развернутая характеристика. Как показала практика, важнейшее значение для расчленения разрезов имеют две-три разновидности биостратиграфических зон. В первую очередь надо назвать комплексную зону и интервал-зону (рис. 2). Комплексная зона – это совокупность слоев, охарактеризованная комплексом из трех или более ископаемых таксонов, который отличен от комплексов подстилающих и перекрывающих слоев. Интервал-зона – это совокупность слоев, которые заключены между двумя установленными биогоризонтами (уровнями первого появления или последнего нахождения каких-либо характерных таксонов). Именно они чаще всего используются при широких корреляциях. Однако в практике используются и другие разновидности зон (зоны совместного распространения, зоны распространения таксона и др.) в зависимости от геологической ситуации и палеонтологического материала.
Рис. 2.
Комплексная зона (а) и интервал-зоны (б) (по International…, 1999). 1 – стратиграфические разрезы, 2 – временная поверхность, 3 – границы интервал-зон, 4 – самое высокое положение таксона в конкретном разрезе, 5 – самое низкое положение таксона в конкретном разрезе, 6 – таксоны.
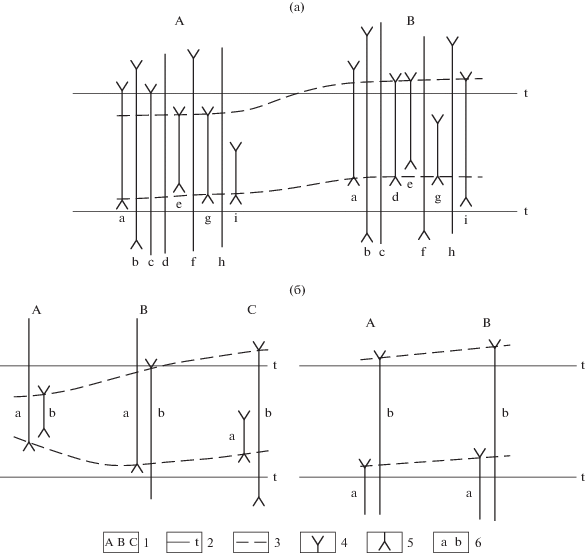
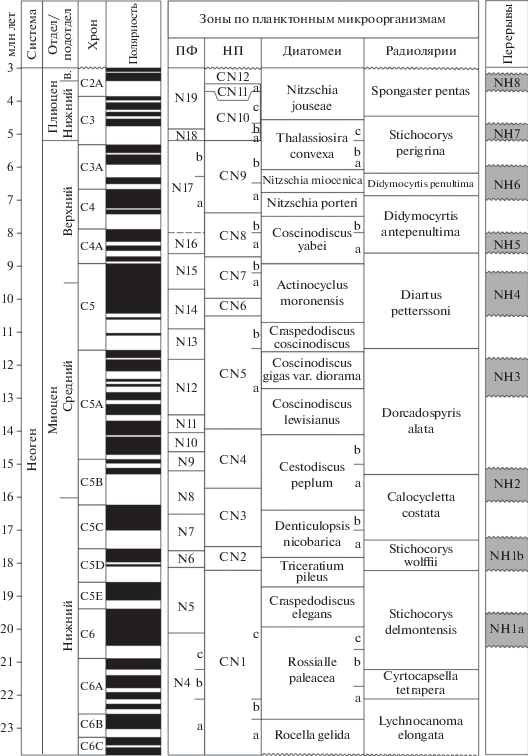
Специально отметим, что границы зон проводятся по датированным уровням – датум-плейнам, прежде всего по уровням появления и исчезновения маркирующих планктонных форм, или с их учетом. Как показал накопленный опыт, наиболее эффективные результаты по выделению зон достигаются именно через анализ стратиграфического распространения отдельных видов, реже – родов. Подобный прием выделения границ предполагает, что зоны часто не отражают значительные этапы развития той или иной группы организмов, а зональные комплексы не всегда обнаруживают преемственность в разрезе. Однако применение такого подхода, при “комбинировании” разного типа зон, позволяет устанавливать дробные и смыкающиеся биостратиграфические подразделения с относительно изохронными границами. Использование датированных уровней представляет удобный практический инструмент, который, по существу, в первую очередь направлен именно на выделение дробных биостратиграфических подразделений и маркирующих горизонтов. Например, на такой основе для олигоценовых–четвертичных отложений Северной Пацифики при участии одного из авторов статьи разработана океаническая зональная шкала по диатомеям, которая сопоставлена с МСШ и включает более 20 зон (рис. 3).
Рис. 3.
Зональная шкала по диатомеям для расчленения олигоцена–квартера Северной Пацифики (по Barron, Gladenkov, 1995; Гладенков, 2007), скоррелированная с геохронологической и палеомагнитными шкалами по (Ogg et al., 2016). УП – уровень появления, УИ – уровень исчезновения, УМП – уровень массового появления, УСЧ – уровень резкого снижения численности, а–с – подзоны, Э. – эоцен, в. – верхний.

В качестве индекс-форм, которые используются для характеристики зональных границ в океанических шкалах, стали выбираться таксоны, отвечающие определенным требованиям. К ним, прежде всего, относятся постоянная встречаемость и широкое площадное распространение остатков, четкость и устойчивость пределов их стратиграфического распространения. При отсутствии каких-либо из основных критериев датированные уровни в ряде случаев могут использоваться в качестве характеристик границ подзон или локальных маркеров, которые не столь устойчивы по сравнению с зональными.
В заключение раздела сделаем два замечания. Во-первых, следует помнить, что при расчленении какого-либо разреза (скважины) может быть выделено достаточно много биостратиграфических зон (“параллельных” – созданных по разным палеонтологическим группам) и часто с несовпадающими границами (рис. 4). И, следовательно, для практического их использования приходится выбирать одну (две) из зональных шкал в качестве опорной или “стандартной”. Во-вторых, надо иметь в виду, что в Стратиграфическом кодексе России (2019) перечисленные зоны относятся к разряду “специальных” (биостратиграфических) подразделений. К основным же единицам Общей стратиграфической шкалы, которые являются подразделениями комплексного обоснования и единицами дробнее яруса, в кодексе выделены “хронозоны” (некоторые исследователи называют их “оппельзонами”). Хотя хронозоны устанавливаются по биостратиграфическим данным, к ним могут относиться отложения с комплексом остатков, отличным от зонального, или без него, если доказана одновозрастность сравниваемых отложений. Однако отметим, что в последнем Международном стратиграфическом руководстве (справочнике) (International…, 1999) хронозона рассматривается как подразделение неопределенного ранга, не входящее в иерархию общепринятых хроностратиграфических единиц.
Рис. 4.
Соотношение зон по разным группам микропланктона нижнего миоцена–плиоцена низких широт (скоррелированных со шкалой магнитной полярности) и перерывы, выявленные в океанических отложениях неогена (по Keller, Barron, 1983, 1987, с упрощением). ПФ – планктонные фораминиферы, НП – наннопланктон, а–с – подзоны.
На практике встречаются случаи, когда неполнота разрезов приводит к трудностям выделения полного объема биостратиграфических зон, и приходится обращаться к тейльзонам – слоям, отвечающим реальному распространению какого-либо таксона (или их группы) в конкретном разрезе конкретного района.
Возраст зон и интеграция данных, полученных разными методами
При выделении зон океанических шкал встал ряд вопросов практического характера: как определить их возраст и как установить возрастное положение датируемых уровней. В этом отношении помогает, конечно, корреляция зон, выявленных в океанических осадках, с зонами, которые установлены в стратотипах ярусов наземных разрезов, особенно если для этих стратотипов имеются определенные данные по их возрастным характеристикам – различным геохронологическим уровням (например, по магнитостратиграфическим или радиологическим определениям). Проблема здесь заключается в том, чтобы корректно провести соответствующие корреляции.
Что касается возраста датированных уровней, то он в океанических разрезах определяется, прежде всего, с помощью данных по палеомагнетизму и радиологическому датированию. Во многих случаях в разрезах глубоководных скважин проведена прямая корреляция границ зон с магнитостратиграфической шкалой. Это дало возможность не только датировать границы зональных подразделений и определять “продолжительность” зон, но и напрямую сопоставлять их с МСШ (Berggren et al., 1995; A Geologic…, 2004; The Geologic…, 2012; Ogg et al., 2016; Geologic…, 2020). Выделение подразделений в современных шкалах геомагнитной полярности кайнозоя (палеомагнитные хроны, субхроны, эпизоды прямой и обратной полярности) и их датирование основаны прежде всего на анализе магнитных профилей при изучении магнитных аномалий в океанических спрединговых зонах, в первую очередь в Южной Атлантике (Cande, Kent, 1992, 1995; The Geologic…, 2012; Geologic…, 2020). На рис. 5 и 6 приведены наглядные примеры выявленного соотношения биостратиграфических зон (по диатомеям) с магнитостратиграфической шкалой в интервале олигоцена–квартера, с одной стороны, тропическо-субтропической области, а с другой – южных высоких широт (эти материалы были получены при бурении в этих регионах).
Рис. 5.
Океаническая зональная шкала по диатомеям для расчленения неогена–квартера низких широт, сопоставленная с палеомагнитной шкалой Cande, Kent (1992, 1995) (по Barron, 2003, 2005, с упрощением). УП – уровень появления, УИ – уровень исчезновения, a–c – подзоны, П. – палеоген, О. – олигоцен, нижн. – нижний, верхн. – верхний, C. – Coscinodiscus, Fr. – Fragilariopsis.
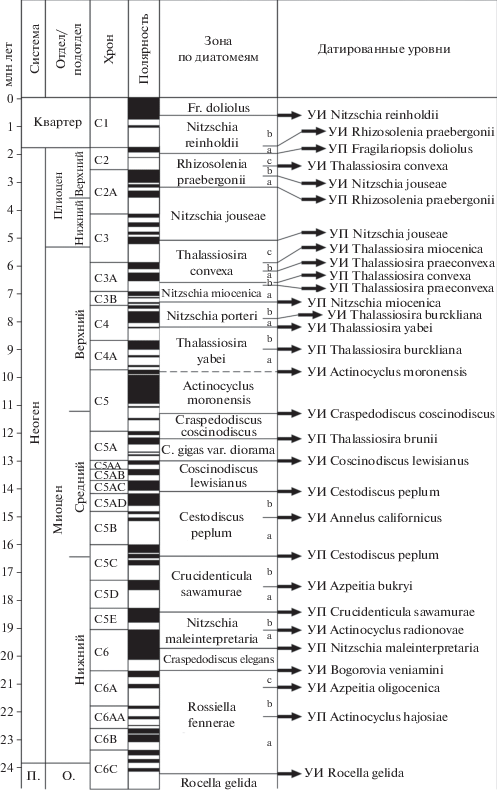
Рис. 6.
Океаническая зональная шкала по диатомеям для расчленения неогена–квартера высоких южных широт, сопоставленная с палеомагнитной шкалой Cande, Kent (1992, 1995) (по Harwood, Maruyama, 1992; Barron, 2003, с упрощением). Fr. – Fragilariopsis, N. – Nitzschia, D. – Denticulopsis, A. – Actinocyclus; остальные аббревиатуры см. на рис. 5.

Вместе с тем стоит обратить внимание на полученные оценки продолжительности интервалов между датум-плейнами. Они часто достигают впечатляющей для фанерозоя дробности (до сотен тысяч лет и даже меньше). В качестве примера в табл. 1 приведены датировки биостратиграфических уровней (биогоризонтов) по диатомеям, полученные по материалам бурения в Северной Пацифике. Такой дробности расчленения фанерозойских разрезов геологическая практика раньше, видимо, не знала (к подобной точности приближаются, наверно, лишь мелкие юрские биостратоны, выделенные в отдельных районах Европы по аммонитам (Page, 2003), но эти подразделения практически не обеспечены палеомагнитными и радиологическими характеристиками в той мере, как кайнозойские зоны океанических шкал).
Таблица 1.
Возраст стратиграфически важных неоген-четвертичных датированных уровней по диатомеям, определенный в результате прямой привязки к палеомагнитной шкале в разрезах глубоководных скважин в субарктической Пацифике (по Barron, Gladenkov, 1995, с дополнениями)
| Датированные уровни | Возраст, млн лет |
|---|---|
| УП Crucidenticula sawamurae | 18.2 |
| УП Crucidenticula kanayae | 16.9 |
| УП Denticulopsis praelauta | 16.3 |
| УП Denticulopsis lauta | 15.9 |
| УП Actinocycluus ingens var. nodus | 15.8 |
| УП Denticulopsis hyalina | 14.9 |
| УП Denticulopsis hustedtii s. ampl. | 14.6 |
| УМП Denticulopsis hustedtii s. ampl. | 13.1 |
| УП Denticulopsis praedimorpha | 13.0 |
| УП Proboscia barboi | 12.6 |
| УCЧ Denticulopsis praedimorpha | 11.6 |
| УИ Mediaria splendida | 11.0 |
| УП Denticulopsis dimorpha | 10.0 |
| УП Thalassionema schraderi | 9.6 |
| УCЧ Denticulopsis katayamae | 9.4 |
| УИ Denticulopsis dimorpha | 9.3 |
| УCЧ Denticulopsis hustedtii s. ampl. | 8.7 |
| УП Thalassiosira gravida | 8.2 |
| УП Pyxidicula zabelinae | 8.0 |
| УСЧ Thalassionema schraderi | 7.7 |
| УП Neodenticula kamtschatica | 7.5 |
| УИ Cavitatus jouseanus | 6.7 |
| УП Thalassiosira praeoestrupii | 6.2 |
| УИ Rouxia californica | 6.0 |
| УИ Ikebea tenuis | 5.5 |
| УП Thalassiosira oestrupii | 5.5 |
| УП Thalassiosira jacksonii (пликатная) | 5.2 |
| УП Thalassiosira latimarginata | 5.1 |
| УИ Cosmiodiscus insignis | 4.8 |
| УИ Thalassiosira jacksonii (пликатная) | 4.8 |
| УП Neodenticula koizumii | 4.0 |
| УСЧ Neodenticula kamtschatica | 2.6 |
| УИ Pyxidicula zabelinae | 2.0 |
| УИ Neodenticula koizumii | 2.0 |
| УСЧ Actinocyclus oculatus | 1.0 |
| УИ Proboscia curviristris | 0.3 |
Необходимо также отметить, что отсутствие полного “набора” установленных зон в ряде разрезов донных отложений позволяет выявлять или размывы отдельных слоев, или перерывы в осадконакоплении с определением их длительности. Одним из впечатляющих примеров исследований по датированию и площадному распространению выявленных перерывов в океанических отложениях неогена являются работы, проведенные на основе данных по планктонным микроорганизмам (Keller, Barron, 1983, 1987). При этом анализ распространения восьми выделенных перерывов (рис. 4) позволил авторам подойти к выявлению основных палеоокеанографических и палеоклиматических перестроек в неогене с этапами изменений в характере осадконакопления, биогеографическом распространении и продуктивности планктонных комплексов.
Особенности и возможности применения биостратиграфических зон в глобальном и региональном масштабе
Результаты, полученные при изучении материалов глубоководного бурения, впервые позволили показать возможность прослеживания зон по различным группам микропланктона в пределах обширных регионов Мирового океана. В первую очередь ярким примером служат кайнозойские зоны по планктонным фораминиферам (Berggren et al., 1995; Berggren, Pearson, 2005; Wade et al., 2011) и наннопланктону (Bukry, 1973, 1975; Okada, Bukry, 1980; Agnini et al., 2014; Raffi et al., 2016), выделенные при изучении скважин, пробуренных в низких широтах. При этом надо отметить, что при разработке зональных шкал учитывались биостратиграфические данные, полученные ранее по различным группам микрофоссилий, прежде всего по планктонным фораминиферам и наннопланктону, из наиболее представительных и палеонтологически хорошо охарактеризованных разрезов на суше (Bramlette, Riedel, 1954; Морозова, 1959; Субботина, 1960; Bandy, 1964; Bolli, 1966; Hay et al., 1967; Blow, 1969; Berggren, 1969; Крашенинников, 1969; Шуцкая, 1970; Martini, 1971 и др.).
В то же время полученные данные свидетельствуют о том, что глобальных зон по микропланктону для всего кайнозоя в строгом смысле не существует. В целом к субглобальным биостратонам могут быть отнесены, например, зоны по планктонным фораминиферам палеоцена–эоцена (рис. 7) (время, когда отмечалось потепление и ареалы тепловодного карбонатного планктона расширялись от тропических широт к аркто-бореальным и натальным). Для характеризующих зоны этого возраста ассоциаций зоо- и фитопланктона типичны космополитные формы. Однако начиная с олигоцена, когда наступило глобальное похолодание и оформилась близкая к современной широтная климатическая зональность, микропалеонтологические ассоциации низких, средних и высоких широт стали заметно различаться. Поэтому в их пределах используются в той или иной степени различные зональные шкалы с разным количеством зон, границы которых часто устанавливаются по разным видам (рис. 8). Зональные комплексы разных широт могут иметь заметные отличия в таксономическом составе, а в качестве датированных уровней во многих случаях выбираются различные формы. Ну а в связи с тем, что в осадочных толщах высоких широт с возрастом моложе эоцена остатки карбонатного планктона редки или отсутствуют, в качестве основных инструментов при биостратиграфическом расчленении и определении возраста здесь используются кремнистые микроорганизмы.
Рис. 7.
Зональная шкала палеоцена–эоцена по планктонным фораминиферам, скоррелированная с палеомагнитной шкалой A Geologic Time Scale 2004 (по Wade et al., 2011, с упрощением). а–с – подзоны. Аббревиатуры см. на рис. 5.
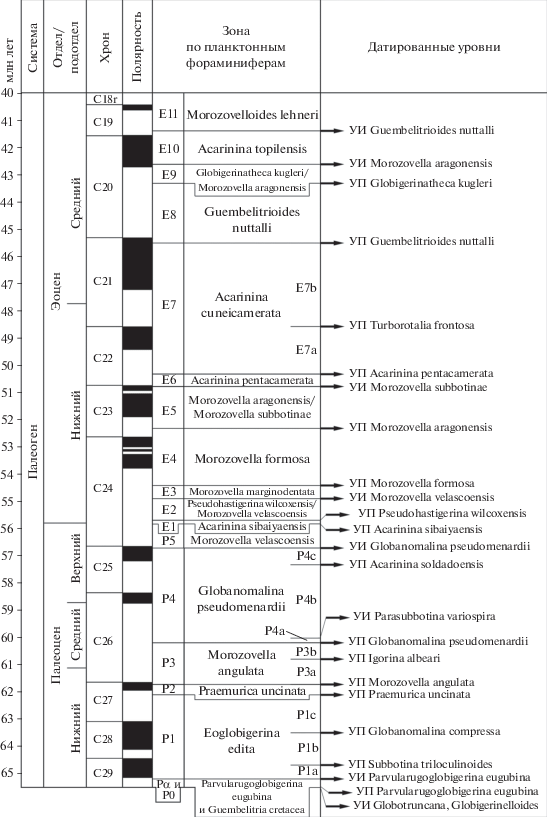
Рис. 8.
Зональные шкалы нижнего–среднего миоцена по планктонным фораминиферам различных климатических широтных зон (по Berggren et al., 1995, с упрощениями). Ол. – олигоцен, верх. – верхний, сер. – серравалий, a–b – подзоны.
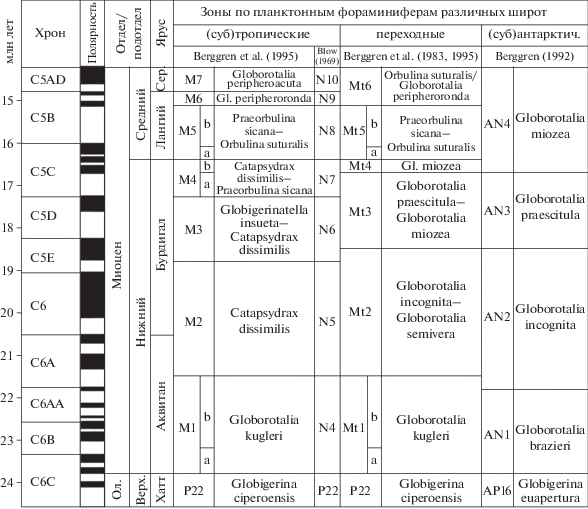
Отдельно следует подчеркнуть, что датум-плейны одних и тех же важных в стратиграфическом отношении видов при переходе из одной широтной области в другую могут оказаться диахронными, что мешает осуществлять корректные сопоставления разрезов разных климатических поясов. На это приходится обращать внимание, так как в ряде случаев под влиянием “гипноза” изучения зон в одном разрезе исследователи “подравнивают” зональные границы в других разрезах, не задумываясь об объеме допуска такого “подравнивания”. В связи с тем, что при проведении границ зон (в частности, по планктону) следует учитывать полный интервал стратиграфического распространения маркирующих форм, при оценке возраста появления таксонов необходимо использовать уровень с наиболее древней из имеющихся датировок (и наоборот, при определении возраста исчезновения таксонов следует использовать уровень с наиболее молодой датировкой). При этом “разброс” возрастных датировок какого-либо из уровней, выбранного в качестве характеристики зональных границы, не должен превышать пределов необходимой точности. Поэтому в конкретных ситуациях надо ориентироваться на то, чтобы имеющийся допуск, или “люфт”, составлял самую минимальную часть зонального интервала. Вот почему при проведении границ большое значение имеет как изучение ископаемых комплексов в широкой серии разрезов, так и использование для контроля других палеонтологических групп, а также анализ различных маркирующих реперов (палеомагнитных, изотопных, литологических и др.).
Однако, несмотря на определенные осложнения, создание субглобальных океанических зональных шкал кайнозоя, а также шкал для обширных сегментов океанов (например, для северной части Тихого океана или высоких южных широт) на основе данных, полученных в результате осуществления Международного проекта глубоководного бурения, представляется одним из важнейших достижений геологии последних десятилетий. Сейчас считается общепринятым фактом, что биостратиграфические зоны тропического пояса прослеживаются во всех трех океанах – Тихом, Атлантическом и Индийском, то есть действительно в субглобальном масштабе. И именно на их основе вносятся коррективы в определение объемов ярусов в стратотипах кайнозоя МСШ, установленных в наземных разрезах. В настоящее время практически все границы ярусов палеогена и неогена в стратотипических разрезах Западной Европы датируются во многом с помощью изученных в них комплексов карбонатного планктона, сопоставленных с ассоциациями океанических зональных шкал.
В то же время из материалов глубоководного бурения следует еще одно обстоятельство. Оказалось, что зоны кайнозоя, отмеченные в одном океане (например, в Тихом), не всегда точно совпадают с зонами другого океана (Атлантического или Индийского). Другими словами, палеонтологическая характеристика и стратиграфическое распространение ряда форм отдельных зон кайнозоя, используемых в пределах одного океана (например, низких широт Тихого океана), не всегда точно совпадают с таковыми зон тех же широт в другом океане (низких широт Атлантического океана) (например, Berggren et al., 1995; Wade et al., 2011; Agnini et al., 2014). Это свидетельствует об определенных различиях водных масс таких крупных экосистем, как океаны. Такой факт четко устанавливается при детальном анализе ископаемых биокомплексов и свидетельствует о том, что океанские воды не являлись во всем однородными в различных областях и частях океанов. При этом приходится учитывать различную роль многих биотических и абиотических факторов, влияющих на формирование палеокомплексов (эволюционные преобразования, конкурентные взаимоотношения таксонов в биотических сообществах, деятельность течений, химический состав вод, процессы, влияющие на сохранность остатков микроорганизмов при опускании на морское дно, а также при их захоронении в осадках и фоссилизации в осадочных породах и др.). Здесь же уместно заметить и о случаях определенного своеобразия биотических комплексов, отмечаемых в различных районах океанов (в частности, на их окраинах и в бассейнах эпиконтинентального типа, где помимо широтной проявляется циркумконтинентальная зональность) и заключающихся в появлении отдельных эндемиков, подвидов и морфотипов и исчезновении некоторых форм, типичных для других акваторий. Практически это свидетельствует о проявлении провинциализма, что, видимо, было свойственно также палеозойским и мезозойским морским бассейнам.
Как показала практика, разработанные зональные шкалы по различным группам планктона могут использоваться также успешно при датировании, расчленении и корреляции толщ и наземных разрезов. Изучение в таких разрезах микропалеонтологических комплексов и сопоставление их с ассоциациями зональных шкал во многих случаях позволило провести дробное расчленение кайнозойских толщ, а также пересмотреть или уточнить возраст различных формаций, свит и региональных ярусов. При этом в шельфовых разрезах часто достаточно трудно проследить зоны, выделенные в океанических толщах, в их полных объемах и определить их границы. В таких разрезах океанические фации, содержащие “эталонные” зональные палеонтологические комплексы, представлены только в идеальных случаях. Это может приводить к несмыкаемости зон в относительно мелководных фациях. К тому же уровни появления и исчезновения стратиграфически важных таксонов здесь могут не совпадать с таковыми их реального возникновения и вымирания. Кроме того, при переходе от океанических к более мелководным отложениям число типично океанических видов в составе комплексов уменьшается (вплоть до их отсутствия), и доминировать начинают формы, характерные для прибрежных вод. Поэтому нередко выделяются местные или локальные биостратиграфические подразделения (охарактеризованные в основном относительно мелководными ассоциациями), которые затем коррелируются с океаническими зонами.
ИСПОЛЬЗОВАНИЕ ДАННЫХ ПО ГЛУБОКОВОДНОМУ БУРЕНИЮ В РЕКОНСТРУКЦИЯХ УСЛОВИЙ И ПАРАМЕТРОВ ОКРУЖАЮЩЕЙ СРЕДЫ ПРОШЛОГО
Обработка материалов глубоководного бурения не только внесла важный вклад в отработку методик и приемов зонального биостратиграфического расчленения морских толщ кайнозоя и проведение их обоснованного датирования, но и позволила получить огромный массив новых данных, представляющих значительную ценность для реконструкций параметров окружающей среды прошлого, прослеживания изменений режимов океанической циркуляции, климата и характера седиментации, а также выявления особенностей развития древней морской биоты.
Палеоокеанология и палеоклимат кайнозоя по данным анализа изотопного состава раковин ископаемых фораминифер
Большую ценность для проведения палеоклиматических и палеоокеанологических реконструкций представляют результаты изучения количественного изменения изотопного состава кислорода (δ18O) и углерода (δ13C) в раковинах глубоководных бентосных фораминифер из кайнозойских разрезов скважин, пробуренных в Мировом океане. Данные по δ18O представляют основу для оценок скорости и масштаба изменений во времени температур глубинных океанических вод, а также объема континентальных льдов. Так как формирование глубинных вод в основном связано с охлаждением и опусканием морских вод в приполярных регионах, их температуры одновременно отражают и температуры поверхностных вод в высоких широтах (Kennett, 1982; Zachos et al., 2001 и др.). Метод определения соотношения тяжелых и легких изотопов кислорода в состоящих из карбоната кальция раковинах бентосных фораминифер является основным инструментом для восстановления абсолютных значений палеотемператур глубинных вод (выявлено, что чем больше значения δ18O, тем ниже температуры, так как при потеплениях раковины фораминифер обогащены легким изотопом кислорода, а при похолоданиях – тяжелым изотопом). Одновременно он дает возможность получить информацию о температурных градиентах палеосреды и изменениях состава морских вод. С другой стороны, анализ изотопно-углеродного состава (δ13C) раковин бентосных фораминифер дает ценную информацию о потоке питательных веществ и крупных изменениях в круговороте этих веществ и углекислого газа в океанских глубинах. Метод основан на выявлении соотношения редких стабильных изотопов и обычных изотопов углерода в раковинах и, как правило, проводится параллельно с изотопно-кислородным анализом.
Как уже говорилось, в глубоководных скважинах, пробуренных в районах различных климатических поясов, были вскрыты серии разрезов, в которых представлены практически непрерывные последовательности осадочных толщ, накопившихся за те или иные интервалы кайнозоя. Использование современных методик дало возможность проводить детальный анализ летописей изменений δ18O и δ13C высокой разрешаемой способности. Впервые был накоплен большой массив данных по изотопии, который ранее было трудно представить, и это позволило выявить интервалы изменений климата, определить скорость и масштаб климатических перестроек, а также сделать выводы об их влиянии на режим океанической циркуляции и условия окружающей среды прошлого (Miller et al., 1987, 1991; Zachos et al., 2001, 2008; Cramer et al., 2009 и др.). Ниже приводится более подробная характеристика полученных результатов, так как подобные данные, основанные на исследованиях тысяч образцов из океанических толщ, до начала глубоководного бурения отсутствовали. Именно они дали возможность по-новому расшифровать многие особенности палеоклиматических и палеоокеанологических обстановок.
Согласно реконструкциям, выполненным на основе использования изотопного метода, в начале кайнозоя, в первой половине палеоцена, на Земле в целом господствовал одинаково теплый и ровный климат с небольшими температурными градиентами между экватором и полюсами. Термическая структура океана была относительно однородной, с температурами донных вод около +8°С. Резкие меридиональные термические градиенты отсутствовали, а тропические зоны были более широкими по сравнению с современными. Поэтому океан характеризовался широтной тепловодной циркуляцией. Циркуляция и температура донных вод были обусловлены в целом не термической, а соленостной стратификацией. В середина палеоцена (около 59 млн лет назад) отмечается начало отчетливого тренда глобального потепления продолжительностью ~10 млн лет. В конце палеоцена, около 56 млн лет назад, фиксируется повышение температур донных вод более чем на 5°C. Этот кратковременный интервал (продолжительностью ~170 тыс. лет) назван палеоцен-эоценовым термальным максимумом (Paleocene–Eocene Thermal Maximum = PETM). Потепление продолжилось в раннем эоцене, достигнув максимума около 50 млн лет назад (раннеэоценовый климатический оптимум – Early Eocene Climatic Optimum = EECO). Согласно полученным данным, климат раннего эоцена (в интервале 53–50 млн лет назад) был самым теплым во всем кайнозое, с температурами донных вод +10°С и более; около 50 млн лет назад температуры глубинных вод достигали +14°С. Это потепление обусловило дальнейшее снижение температурных градиентов между приполярными и приэкваториальными областями.
После температурного пика, в самом конце раннего эоцена (около 49 млн лет назад), в океанах по соотношению изотопов кислорода фиксируется начало общего тренда глобального кайнозойского похолодания, характеризующего историю развития Земли за последние 50 млн лет. Этот климатический тренд, однако, имел сложный и неоднородный характер, при котором эпизоды падения температур водных масс чередовались с интервалами стабильных условий и относительных потеплений. Например, наиболее отчетливое потепление на фоне тренда глобального похолодания в среднем эоцене фиксируется в интервале ~40.6–40.0 млн лет назад (среднеэоценовый климатический оптимум – Middle Eocene Climatic Optimum = MECO), с пиком около 40 млн лет назад, когда температуры донных вод достигали +8°С. Однако на протяжении среднего и позднего эоцена фиксируется усиление контраста между температурами вод низких и высоких широт по причине падения температур последних в результате похолодания. Понижение температуры вод приполярных областей вело к формированию более холодных донных вод. Придонные же воды отличаются от поверхностных тем, что после формирования и растекания в горизонтальном направлении они мало меняются на огромных океанических пространствах. Поэтому их проникновение по направлению к экватору могло вызывать усиление температурного контраста между поверхностными и донными водами в низких и средних широтах, а также приводить к усилению широтного термического градиента. Увеличение вертикального температурного градиента вело к появлению термоклина, который обусловливал “разъединение” поверхностных и глубинных вод, что препятствовало подъему последних к поверхности. Интересно отметить, что согласно данным, полученным при обработке материалов глубоководного бурения у восточного побережья Антарктиды (Barron et al., 1991), оледенение на участках побережья Восточной Антарктиды и образование морских льдов на ее шельфе, вероятно, могли иметь место уже в конце среднего эоцена (около 39 млн лет назад).
Однако, как свидетельствуют результаты исследований по соотношению изотопов кислорода, наиболее знаменательное событие произошло вблизи границы между эоценом и олигоценом (~34 млн лет назад). В это время фиксируется значительное падение температур донных вод в Мировом океане, а также поверхностных вод в южной приполярной области (олигоценовое кислородно-изотопное событие Oi-1). Многочисленные данные указывают на то, что к этому времени относится начало интенсивного формирования оледенения Антарктиды и морских льдов на ее шельфовой окраине. За счет охлаждения и опускания ко дну поверхностных вод с нормальной соленостью в прилегающих к Антарктиде мелководных районах началось образование низкотемпературных плотных антарктических придонных вод, растекавшихся к северу. Этот отток стал компенсироваться подъемом менее плотных циркумполярных глубинных вод, что формировало зону антарктической дивергенции повышенной биологической продуктивности. Поэтому анализ изотопного состава раковин бентосных фораминифер этих районов дает информацию не только об объеме ледников, составе и температурах донных вод, но и о температурах приполярных поверхностных вод. Таким образом, процесс формирования холодных донных вод во время пика похолодания в самом начале раннего олигоцена привел к появлению в высоких широтах Южного полушария криосферы, а в океанических глубинах – психросферы, то есть к изменению всего температурного режима планеты (так называемый “оранжерейный” режим – “green house” сменился “ледниковым” режимом – “ice house”). Значительно возросшее влияние полярных областей на формирование глобальной океанической циркуляции и климатического режима в целом к началу олигоцена привело к смене режимов общей океанической циркуляции и климата. Появление более резких температурных градиентов между высокими и низкими широтами вследствие высокоширотного похолодания вызвало усиление вертикальной и поверхностной океанической циркуляции. Возникновение в водной толще выраженных температурных градиентов обусловило изменение комплекса характеристик и циркуляции поверхностных вод, а также интенсификацию поверхностных течений. Преимущественно широтная тепловодная циркуляция на всех глубинах, характерная в целом для вод Мирового океана в раннем палеогене, сменилась преимущественно меридиональной термогалинной холодноводной циркуляцией. Появление более резких температурных градиентов между высокими и низкими широтами вследствие высокоширотного похолодания вызвало усиление вертикальной и поверхностной океанической циркуляции. Это, в свою очередь, приводило к усилению деятельности прибрежных ветров и пассатов, вызывавшей активизацию прибрежных и экваториальных апвеллингов. Изменения в океанической циркуляции особенно ярко выразились в высоких южных широтах, где в начале олигоцена появился циркумполярный пояс кремненакопления, который отражает обширную зону подъема глубинных вод, богатых питательными веществами. Термические барьеры в высоких южных широтах в виде зон антарктической и субтропической конвергенции стали основными биогеографическими преградами, влияющими на распространение планктонных организмов.
Итак, общий тренд глобального кайнозойского похолодания имел на протяжении последних 50 млн лет сложный и неоднородный характер. Хотя климатические и океанографические изменения, произошедшие в конце среднего эоцена, привели к некоторому усилению широтного температурного градиента и контраста между глубинными и поверхностными водами, коренная перестройка океанической циркуляции и климата имела место вблизи границы между эоценом и олигоценом. Именно в это время был нарушен общий циркуляционный и температурный баланс, существовавший в раннем палеогене, что явилось толчком к формированию психросферы и криосферы и началу перехода планеты от общего “оранжерейного” температурного режима к “ледниковому” режиму. Однако происходившие изменения не сразу привели к современной широтной зональности с самыми низкими температурами в высоких широтах, по сравнению с предыдущими эпохами. Начиная с олигоцена, эпизоды скачкообразного падения температур чередовались с интервалами стабильных условий и периодами относительных потеплений. Наиболее масштабные потепления с повышением температур глубинных вод отмечены в конце олигоцена и в середине миоцена (миоценовый климатический оптимум вблизи границы между ранним и средним миоценом). В целом результаты анализа изотопов кислорода свидетельствуют, что температуры вод приполярных областей до конца среднего миоцена (время начала постепенного похолодания) оставались на несколько градусов выше по сравнению с современными, а постоянных континентальных льдов не существовало. Постоянное оледенение Восточной Антарктиды формируется только начиная с конца среднего миоцена, а Западной Антарктиды – с начала плиоцена. Начало же континентального оледенения Северного полушария относится к среднему плиоцену. Таким образом, в начале раннего олигоцена тропические и субтропические зоны были шире современных. Климат на Земле был мягче по сравнению с современным, оледенения высоких южных широт не были постоянными, а значения температур вод в этих районах не были такими низкими, как сейчас. Однако несмотря на это, формирование психросферы и образование термической стратификации вод имели необратимый характер. Это, наряду с изменением глобальной океанической циркуляции, имело решающее значение для изменения закономерностей развития и распространения как планктонных организмов, так и океанической биоты в целом. В целом палеоклиматическая эволюция в кайнозое имеет три главные особенности: (1) общее понижение температуры начиная с конца раннего эоцена; (2) более резкое похолодание в высоких широтах и более слабое в тропических широтах; (3) не постепенный, а скачкообразный характер похолодания (Zachos et al., 2001) (рис. 9). А к настоящему времени выполнены обобщения, основанные на еще более обширном массиве данных по изотопии, которые позволили значительно дополнить и усовершенствовать кривые изменений δ18O и δ13C, а также уточнить возраст происходивших перестроек (Cramer et al., 2009).
Рис. 9.
Кривая глобальных изменений δ18O в глубоководных отложениях кайнозоя и “календарь” важных климатических событий палеоцена–квартера (по Zachos et al., 2001). 1, 2 – ледовые покровы: 1 – частичные или кратковременные, 2 – полномасштабные или постоянные; PETM – палеоцен-эоценовый термальный максимум, EECO – раннеэоценовый климатический оптимум. пл. – плиоцен, ч. – четвертичный период.
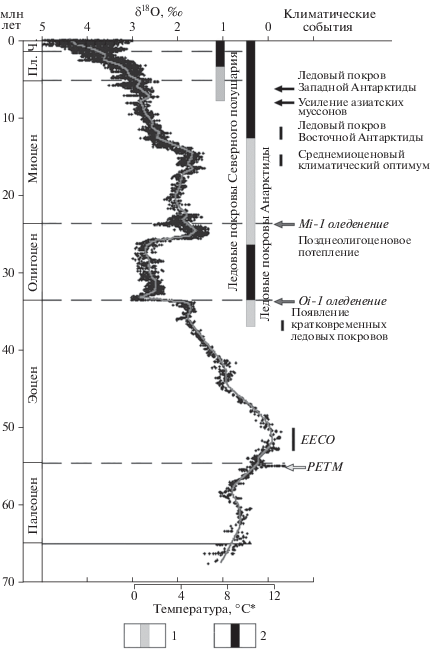
Отдельно следует сказать, что результаты изучения количественного изменения изотопного состава кислорода в разрезах океанических отложений внесли огромный вклад в выделение “морских изотопных стадий” (Marine Isotope Stages = MIS) неогена–квартера. Разработанная к настоящему времени детальная шкала этих стадий, отражающих последовательность относительно кратковременных периодов потеплений и похолоданий, представляет основу дробного климатостратиграфического расчленения и датирования четвертичных морских отложений и их корреляции с континентальными толщами.
К сказанному выше следует добавить, что при исследовании керна из глубоководных скважин, помимо биостратиграфического и магнитостратиграфического методов, а также изучения изменения изотопного состава кислорода (δ18O) и углерода (δ13C), для геологических и палеоклиматических реконструкций, как и определения возраста пород, сейчас в той или иной мере используются и другие методы и подходы (стронциевая изотопная, осмиевая изотопная, серная изотопная стратиграфия, радиоизотопная геохронология, астрохронология, ТЕХ86 и проч.). Возможно, часть из них пока не имеет определяющего и масштабного значения, но их роль постоянно растет. В частности, все больше получает развитие стронциевая изотопная хемостратиграфия (основанная на изучении вариаций 87Sr/86Sr), которая в ряде случаев помогает совершенствовать стратиграфические схемы, их корреляции и палеогеографические модели. Так, изучение образцов кайнозойских карбонатных отложений, прежде всего неогеновых, из разных частей Мирового океана выявило одинаковое отношение 87Sr/86Sr в одновозрастных осадках, что позволяет использовать Sr-изотопную характеристику как средство корреляции (DePaolo, Ingram, 1985; Koepnick et al., 1985; Hodell et al., 1989; Farrell et al., 1995 и др.). А поскольку отношение 87Sr/86Sr в кайнозое неуклонно повышалось, высокий градиент этого отношения позволяет различать возраст морских отложений с точностью до яруса и даже дробнее (Кузнецов и др., 2018). Но подробная характеристика разных методов не входит в нашу задачу, и мы отсылаем читателя к соответствующим сводкам последних лет (например, см. McArthur et al., 2001, 2020).
Некоторые особенности развития и распространения микропалеонтологических комплексов в кайнозое
Большой вклад в реконструкции изменений морской среды, океанической циркуляции и климатов прошлого внес детальный анализ данных по таксономическому составу и характеру пространственного распределения разновозрастных комплексов микропланктона, полученных в ходе изучения глубоководных скважин. Это объясняется тем, что, во-первых, микроорганизмы являются достаточно чуткими индикаторами изменений температур морских вод, а во-вторых, характер площадного распространения и скорости накопления биогенных осадков отражают особенности режимов океанической циркуляции и условий осадконакопления.
В современном Мировом океане таксономический состав планктонных ассоциаций во многом определяется широтной климатической зональностью: в различных географических зонах доминируют группы карбонатных или кремнистых организмов определенного видового состава. От положения климатических зон и деятельности течений в этих поясах зависит температурный режим поверхностных водных масс. Они разделены переходными – фронтальными – зонами (фронтами), которые простираются субширотно в соответствии с границами климатических зон. Во фронтальных зонах развиты сообщества смешанного состава, то есть переходные между комплексами соседних водных масс. Фронты служат термическими барьерами для миграции многих представителей планктона, поэтому распространение этих видов ограничено определенной поверхностной водной массой и является для нее наиболее показательным. Иными словами, различные планктонные ассоциации приурочены к поясам, которые в целом протягиваются параллельно экватору. Подобные пояса прослеживаются и в донных осадках океанов, куда остатки микроорганизмов попадают в процессе седиментации (Лисицын, 1978). С другой стороны, для теплых водных масс низких-средних широт в целом характерно доминирование карбонатного планктона, который является основным источником биогенного карбонатонакопления в донных осадках. В холодных водах высоких широт карбонатный планктон развит намного реже или практически отсутствует. Здесь доминирует кремнистый микропланктон, прежде всего диатомовые водоросли, что обуславливает процесс преимущественно биогенного кремненакопления. Исключением, пожалуй, являются Северная Атлантика и примыкающие к ней моря, где глубинные воды отчасти образуются за счет поступления “молодых” вод северного происхождения, которые формируются на границе с Арктическим бассейном и недостаточно обогащены растворенным кремнеземом. Поэтому панцири и скелеты кремнистых микроорганизмов на дне подвергаются растворению за счет взаимодействия с глубинными водами, что объясняет здесь незначительное накопление биогенных кремнистых осадков. Отметим также, что ввиду недонасыщенности морских вод кремниевой кислотой активное развитие кремнистого фитопланктона (основного продуцента биогенного кремнезема и поставщика его в осадки) возможно только в областях, где происходит вертикальное перемешивание вод, в результате которого к поверхности поднимаются глубинные воды, богатые минеральными веществами, в том числе кремнеземом (например, в зонах дивергенций и апвеллингов). Таким образом, характер развития диатомовых водорослей в планктоне (в том числе их продуктивность) и их участие в биогенной седиментации в разных частях Мирового океана неоднородны и, в конечном счете, зависят от режима глобальной океанической циркуляции. В соответствии с этим в Мировом океане выделяются три основных пояса современного осадочного кремненакопления (Лисицын, 1978). Положение этих поясов в целом совпадает с таковым регионов, поверхностные воды которых характеризуются наибольшим развитием диатомовой флоры. Следовательно, по смене характера распространения и интенсивности накопления кремнистых толщ можно судить об изменениях степени продуктивности диатомовой флоры в поверхностных водах того или иного региона в соответствующий интервал геологического времени.
Изучение материалов глубоководного бурения показало, что широтная зональность была характерна и для распространения древнего микропланктона. Исследования таксономического состава разновозрастных ископаемых ассоциаций в разрезах скважин различных регионов океана дали возможность прослеживать смещение во времени поверхностных масс по отношению к экватору в результате климатических флуктуаций, отражающих периоды потеплений и похолоданий. С другой стороны, анализ пространственного распространения ископаемых комплексов карбонатных и кремнистых микроорганизмов в разрезах позволил сделать выводы о смещении во времени “теплых” и “холодных” водных масс, степени контраста широтных температурных градиентов в океанах, а также об изменениях океанической циркуляции. В частности, анализ особенностей распространения морских кайнозойских биогенных кремнистых толщ (Baldauf, Barron, 1990; Barron et al., 2015) дал возможность выявить различия в развитии кремнистого фитопланктона в океанах в разные интервалы палеогена и неогена, а также связать их с перестройками климата и океанической циркуляции.
Кроме того, полученные в ходе обработки материалов глубоководного бурения данные позволили установить особенности палеобиотических сообществ кайнозоя открытых бассейнов и шельфовых зон, определить специфику развития комплексов открытоокеанических, полуизолированных и окраинно-океанических экосистем, с анализом эволюции ассоциаций микроорганизмов отдельных провинций.
Общие особенности эволюции океанических экосистем
Суммирование изложенных выше данных служит основой расшифровок ряда общих особенностей развития морских палеоэкосистем и трендов изменения палеогеографических обстановок. Этому посвящено множество работ (Kennett, 1982 и др.), поэтому ниже они излагаются в кратком виде.
– Получены данные по этапности развития различных групп морской фауны и флоры (с фиксацией периодов вспышек формообразования и таксономического разнообразия), с рассмотрением развития как отдельных видов, так и сообществ. Именно с учетом этих данных был проведен анализ этапности геологического развития крупных морских экосистем (Атлантического, Тихоокеанского, Антарктического и Арктического бассейнов) и их частей.
– Установлены различия между биокомплексами в зависимости от их широтной приуроченности, то есть их принадлежности к разным климатическим поясам (тропическому, бореальному, натальному и арктическому).
– Изучение микропалеонтологических комплексов в разрезах океанических экотонных зон дало возможность определить особенности биосообществ этих переходных зон и их смещение во времени в зависимости от климатических флуктуаций.
– При изучении и восстановлении условий развития и формирования древних ассоциаций микроорганизмов было определено влияние на распространение биоты морских течений и апвеллингов. В этом же отношении важными оказались многие морские проливы (пролив Дрейка в Южном полушарии, Гибралтарский пролив в Северном полушарии и проч.).
– Обращают на себя внимание случаи определенной диахронности уровней появления и исчезновения тех или иных таксонов микропланктонных организмов в разрезах различных морских бассейнов. В одних случаях это, видимо, связано с миграциями биоты, обусловленными климатическим фактором (при потеплениях, например, тропические формы мигрируют к северу, часто отставая во времени появления в высоких широтах на десятки тысяч и даже, возможно, на первые миллионы лет). В других случаях это связывается с раскрытием морских проливов и миграциями палеосообществ из одного бассейна в другой, в результате чего появление одних и тех же таксонов в разных бассейнах может быть неодновременным. Однако точно определить масштаб таких расхождений часто достаточно сложно. Вместе с тем установлены различия палеобиотических сообществ открытых бассейнов и шельфовых зон, в которых упомянутые выше “уровни появления и исчезновения” отдельных таксонов тоже обнаруживают диахронность. Наконец, мы вынуждены считаться со спецификой развития биоты эпиконтинентальных, полуизолированных и окраинно-океанических экосистем с их провинциями, в которых тоже могут отмечаться случаи диахронности уровней.
– Выявлены случаи биполярности в распространении некоторых древних биотических комплексов планктона. Например, недавно определены биполярные ареалы для ряда видов планктонных диатомей начала раннего олигоцена и предложены возможные механизмы их формирования (Gladenkov, 2014).
– На стратиграфической основе построены графики палеоклиматических флуктуаций и осуществлены реконструкции палеогеографических обстановок по различным временным срезам. На материалах по микроорганизмам были получены наиболее детальные данные по изменениям климатических обстановок прошлого, в частности по смене “оранжерейного” типа климата на “ледниковый” вблизи границы между эоценом и олигоценом (Zachos et al., 2001; Cramer et al., 2009; настоящая работа, рис. 9).
– Накопленный опыт при изучении палеонтологических комплексов в разрезах глубоководных скважин внес существенный вклад в оценку разнообразия различных групп морских микроорганизмов в прошлые эпохи. В частности, это позволило выявить, что биоразнообразие на родовом и видовом уровнях карбонатного микрофитопланктона (кокколитофорид) и динофлагеллат достигало максимума в позднем мелу, а с эоцена и до настоящего времени прослеживается тренд его уменьшения. И, наоборот, биоразнообразие кремнистого микрофитопланктона (диатомей) на протяжении кайнозоя возрастало с отчетливой активизацией вблизи границы между эоценом и олигоценом, а затем в интервале среднего–позднего миоцена. Таким образом, кремнистый фитопланктон и карбонатный фитопланктон имеют различные, не совпадающие по времени тренды увеличения разнообразия. Вероятно, такой контраст отражает специфику развития фитопланктона в различных экологических условиях, которая проявляется в приспособлении к изменениям условий окружающей среды (Falkowski et al., 2004). В частности, диатомеи могут существовать в условиях, когда поставка необходимых для жизнедеятельности веществ в зону фотосинтеза носит не постоянный, а пульсационный характер. При благоприятных условиях водоросли активно используют появляющуюся “подпитку”, и происходит интенсивное и продуктивное развитие диатомовой флоры. Таким образом, для существования диатомей благоприятны условия, когда периоды высокой гидродинамической активности чередуются с периодами относительной стабильности в водной толще. И, напротив, наннопланктон и динофлагеллаты имеют тенденцию к доминированию и продуктивному развитию в относительно спокойных и стабильных условиях. Подобные оптимальные условия для карбонатного планктона существовали в мезозое и раннем палеогене, когда на Земле господствовал “оранжерейный” тип климата с небольшими широтными температурными градиентами между экваториальной и полярными областями и отсутствием сильных ветров. Такой режим предопределял медленную стабильную циркуляцию вод в относительно теплых океанах при отсутствии психросферы и резких термоклинных барьеров в водной толще. На этот период приходится расцвет карбонатного фитопланктона. Переход к “ледниковому” типу климата и появление глобальной термогалинной циркуляции в начале раннего олигоцена явились причинами формирования выраженного термоклина и усиления циркуляции поверхностных вод. Изменения условий окружающей среды создали предпосылки для расцвета и увеличения разнообразия диатомей и, напротив, для регресса и снижения разнообразия карбонатного фитопланктона.
– Отдельно надо отметить, что материалы глубоководного бурения позволили по-новому осветить сложную проблему эвстатики и относительного колебания уровня Мирового океана (подробно см., например, Vail, Hardenbol, 1979; Haq et al., 1987; Hardenbol et al., 1998; Miller et al., 2020). В связи с этим надо вспомнить о широком использовании сейсмостратиграфического метода, который сейчас сопровождает практически все океанологические исследования по выявлению строения дна океанов и их окраин.
– Внесены коррективы в определение ярусных границ в стратотипах верхнего мела, палеогена и неогена, установленных в разрезах на материковых блоках, с уточнением границ стратиграфических подразделений разного ранга и возрастного положения маркеров.
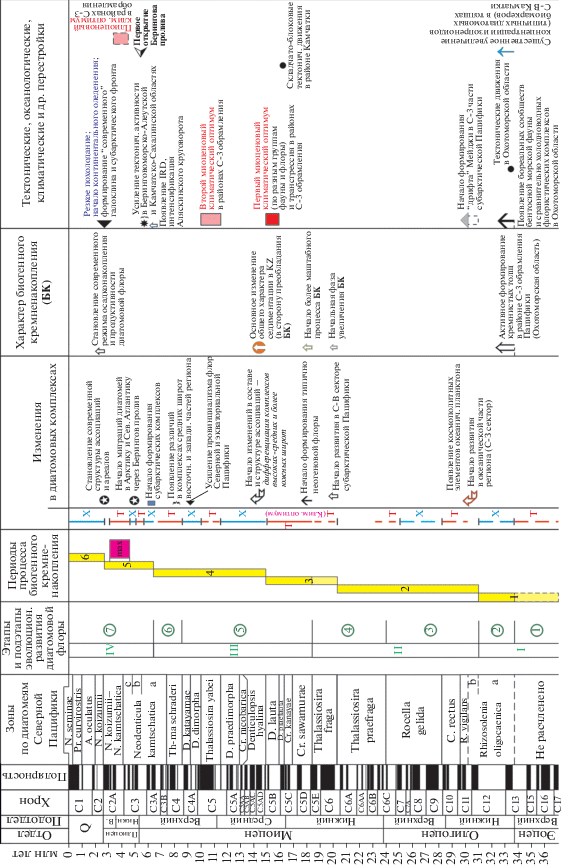
– Выявлена общая картина последовательности и связи ряда биотических и абиотических событий в Мировом океане для последних 70–75 млн лет. Это позволяет составлять календари различных геологических событий, которые наглядно отражают определенные этапы природных процессов прошлого и их масштаб. Такие календари могут быть как субглобальными, так и региональными. В качестве иллюстрации субглобального календаря можно привести известную схему событий кайнозоя из работы (Zachos et al., 2001). К календарям регионального масштаба могут быть отнесены построения одного из авторов статьи для позднего кайнозоя Северо-Тихоокеанского региона (Гладенков, 2007) (рис. 10).
Рис. 10.
Изменения комплексов диатомей и геологические события олигоцена–неогена в Северо-Тихоокеанском регионе (по Гладенков, 2007). Т – тепловодные диатомовые ассоциации, Х – холодноводные диатомовые ассоциации.
По-видимому, этот перечень общих и частных особенностей развития морских экосистем может быть продолжен. Но при этом важно одно – все эти фактические и методические сведения, полученные при обработке материалов глубоководного бурения, уже сейчас могут быть использованы при построении региональных и межрегиональных стратиграфических схем не только мела и кайнозоя, но и всего фанерозоя в целом.
Изложенные выше выводы представляют собой современную оценку стратиграфических результатов глубоководного бурения. Но мы, конечно, должны помнить и о мнении по разбираемой проблеме тех специалистов, которые сами в прошлые годы принимали активное участие в этих работах и которые еще раньше оценили многие их результаты с позиций не только региональных построений, но и стратиграфических проблем общего плана. В этом отношении уместно вспомнить, например, о некоторых выводах В.А. Крашенинникова – одного из самых известных стратиграфов и специалистов в области изучения ископаемых фораминифер, который оставил несколько крупных монографий о результатах глубоководного бурения, отличающихся громадным объемом привлеченного материала и широтой его анализа. Напомним лишь некоторые из этих выводов, не боясь определенных повторов и учитывая, что они были сделаны много лет назад (Крашенинников, 1969, 1973; Крашенинников, Басов, 2007). В.А. Крашенинников один из первых отметил, что бурение в океанических бассейнах приносит обширнейшие, ранее неизвестные сведения об их геологическом строении и истории развития на протяжении мезозойского и кайнозойского времени. Из опыта бурения, по его мнению, следует, что биостратиграфический метод является основным при изучении осадочного чехла планеты, несмотря на интенсивное развитие в последние десятилетия различных физических и химических методов исследования осадочных толщ. Он же заметил, что с течением времени изменилась палеонтологическая база стратиграфии мезозоя и – прежде всего – кайнозоя. Основой субглобальных стратиграфических шкал стали планктонные микроорганизмы (фораминиферы, наннопланктон, диатомеи, радиолярии). Бентосные организмы (нуммулитиды, мелкие бентосные фораминиферы, моллюски, морские ежи) сохраняют свое важное значение для разработки региональных стратиграфических шкал, но, как правило, не являются носителями субглобальной стратиграфической информации. Среди планктонных микроорганизмов ведущее положение в кайнозое занимают фораминиферы, которые, с одной стороны, исторически являются наиболее изученной традиционной группой микрофауны, а с другой – отличаются всесветным распространением и быстрым темпом эволюции. Максимальное развитие планктонные фораминиферы получили в тропической области. Здесь они послужили основой для разработки зональной (Карибской) шкалы, предложенной в 1957 г. Г. Болли (Bolli, 1957а, 1957b, 1957c) и модифицированной в 1969 г. У. Блоу (Blow, 1969), а глубоководное бурение подтвердило ее высокий стратиграфический и корреляционный потенциал. В.А. Крашенинникову давно было ясно, что при переходе к субтропическим отложениям палеогена и неогена дробность стратиграфического расчленения по планктонным фораминиферам несколько снижается, а зональные границы могут располагаться на других уровнях. Хотя при переходе к более высокоширотным областям детальность расчленения отложений по планктонным фораминиферам уменьшается, однако основы стратиграфической шкалы сохраняются. По мнению В.А. Крашенинникова, в методическом плане следует рекомендовать расчленение разрезов с помощью комплексов видов, а не отдельных таксонов (с этим кто-то может и не согласиться). Последние следует рассматривать в качестве дополнительного аргумента при обосновании границ подразделений. Параллельное использование различных групп планктонных организмов (например, планктонных фораминифер и наннопланктона) повышает степень достоверности стратиграфических построений. Безусловно, эти и многие другие соображения В.А. Крашенинникова свидетельствуют о его широком понимании результатов глубоководного бурения в отношении разработки стратиграфии осадочных отложений нашей планеты и остаются весьма полезными до сих пор.
ЗАКЛЮЧЕНИЕ
Таким образом, результаты стратиграфических исследований, проведенных при глубоководном бурении за 50 лет, во многом имеют “прорывной характер”. Они вносят весомый вклад в разработку и совершенствование методики детальных стратиграфических исследований и определения возраста морских комплексов осадочных пород, а также в реконструкции океанологических и климатических событий прошлого. Полученные результаты имеют важное методическое значение для проведения стратиграфических работ по всему фанерозою. Не случайно полученные в процессе глубоководного бурения материалы постоянно обсуждаются на международных конгрессах по стратиграфии и других научных форумах. Многие из этих материалов были использованы, в частности, в недавно опубликованной обширной сводке по различным стратиграфическим проблемам (Geologic…, 2020).
Впервые показано, что выделенные зональные подразделения кайнозоя могут быть реально прослежены на громадной территории – во всем тропическом поясе трех океанов (Тихого, Индийского и Атлантического). В настоящее время подобные дробные биостратоны стали уже обязательным элементом стратиграфических схем, которые разрабатываются для древних морских толщ, вскрытых на суше.
При этом в проведенных работах значительную роль сыграло изучение комплексов микроорганизмов, которые являлись неотъемлемой частью мезозойских и кайнозойских морских экосистем. Развитие этих ассоциаций во времени проходило на фоне смены режимов океанической циркуляции и осадконакопления, изменения степени продуктивности глубинных и поверхностных вод, их температур и других характеристик. Направленность и тенденции эволюции биотических сообществ в целом отражают этапы развития океанических экосистем прошлого и происходивших в них перестроек. Поэтому исследование этапности развития микроорганизмов дало возможность подойти к установлению последовательности и синхронности разномасштабных природных процессов, пониманию их взаимосвязи и влияния на формирование биоты. Все это позволило выявить общую картину последовательности не только биотических, но и абиотических (климатических, океанографических, палеогеографических, эвстатических) событий в Мировом океане для последних 70–75 млн лет. Весь комплекс работ по особенностям зональной стратиграфии кайнозоя, выполненный при глубоководном бурении за последние 50 с небольшим лет, позволяет дать ему самую высокую оценку. Опыт стратиграфических исследований, полученный в процессе осуществления Международного проекта по глубоководному бурению, достоин дальнейшего изучения и осмысления.
Благодарности. Авторы признательны А.Г. Матулю, В.С. Вишневской и Л.Ф. Копаевич за конструктивные советы, которые были учтены при доработке статьи.
Источники финансирования. Работа выполнена по теме государственного задания ГИН РАН и при поддержке гранта РФФИ № 19-05-00361.
Список литературы
Басов И.А. Глубоководное бурение в океанах // Соросовский образовательный журн. 2001. Т. 7. № 10. С. 59–66.
Гладенков А.Ю. Детальная стратиграфия и морские экосистемы позднего кайнозоя севера Тихоокеанского региона (по диатомеям). М.: ГЕОС, 2007. 296 с. (Труды ГИН РАН. Вып. 571).
Гладенков Ю.Б. Биосферная стратиграфия (проблемы стратиграфии начала XXI века). М.: ГЕОС, 2004. 120 с. (Труды ГИН РАН. Вып. 551).
Гладенков Ю.Б. Зональная биостратиграфия в решении фундаментальных и прикладных задач геологии // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 6. С. 104–117.
Крашенинников В.А. Географическое и стратиграфическое распределение планктонных фораминифер в отложениях палеогена тропической и субтропической областей. М.: Наука, 1969. 187 с. (Труды ГИН АН СССР. Вып. 202).
Крашенинников В.А. Стратиграфия миоценовых отложений области Атлантического, Индийского и Тихого океанов по фораминиферам. М.: Наука, 1973. 224 с. (Труды ГИН АН СССР. Вып. 233).
Крашенинников В.А., Басов И.А. Стратиграфия палеогеновых отложений Мирового океана и корреляция с разрезами на континентах. М.: Научный мир, 2007. 316 с. (Труды ГИН РАН. Вып. 583).
Кузнецов А.Б., Семихатов М.А., Горохов И.М. Стронциевая изотопная хемостратиграфия: основы метода и его современное состояние // Стратиграфия. Геол. корреляция. 2018. Т. 26. № 4. С. 3–23.
Леонов Г.П. Основы стратиграфии. М.: Изд-во МГУ, 1973. Т. 1. 530 с.
Лисицын А.П. Процессы океанской седиментации: литология и геохимия. М.: Наука, 1978. 366 с.
Литвин В.М. Морфоструктура дна океанов. Л.: Недра, 1987. 275 с.
Матуль А.Г. Глубоководное научное бурение в океане: структура, итоги, планы // Природа. 2010. № 7. С. 24–33.
Меннер В.В. Избранные труды. Общие вопросы стратиграфии. М.: Наука, 1991. 288 с.
Морозова В.Г. Зональная стратиграфия палеогена на основе развития микроскопических фораминифер // Труды совещания по разработке унифицированной стратиграфической шкалы третичных отложений Крымско-Кавказской области. М.: Изд-во АН СССР, 1959. С. 277–293.
Океанология. Геология океана. Геологическая история океана. Отв. ред. Монин А.С., Лисицын А.П. М.: Наука, 1980. 464 с.
Рубаник Н.К. Глубоководное бурение в океанах: история и перспективы. К 40-летию Международной программы глубоководного бурения // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 6. С. 104–109.
Степанов Д.Л., Месежников М.С. Общая стратиграфия (методы стратиграфических исследований). Л.: Недра, 1979. 423 с.
Стратиграфический кодекс России. Издание третье, исправленное и дополненное. СПб.: Изд-во ВСЕГЕИ, Межведомственный стратиграфический комитет, 2019. 96 с.
Субботина Н.Н. Пелагические фораминиферы палеогеновых отложений СССР // Палеогеновые отложения юга европейской части СССР. М.: Изд-во АН СССР, 1960. С. 24–38.
Удинцев Г.Б. Рельеф и строение дна океанов. М.: Недра, 1987. 239 с
Шуцкая Е.К. Стратиграфия, фораминиферы и палеогеография нижнего палеогена Крыма, Предкавказья и западной части Средней Азии. М.: Недра, 1970. 256 с. (Труды ВНИГРИ. Вып. 70).
A Geologic Time Scale 2004. Eds. Gradstein F.M., Ogg J.G., Smith A.G. Cambridge: Cambridge Univ. Press, 2004. 589 p.
Agnini C., Fornaciari E., Raffi I., Catanzariti R., Pälike H., Backman J., Rio D. Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes // Newslett. Stratigr. 2014. V. 47. P. 131–181.
Baldauf J.G., Barron J.A. Evolution of biosiliceous sedimentation patterns – Eocene through Quaternary: paleoceanographic response to polar cooling // Geological History of the Polar Oceans: Arctic versus Antarctic. Eds. Bleil U., Thiede J. Kluwer Academic Publ., 1990. P. 575–607.
Bandy O.L. Cenozoic planktonic foraminiferal zonation // Micropaleontology. 1964. V. 10. P. 1–17.
Barron J.A. Planktonic marine diatom record of the past 18 m.y.: appearances and extinctions in the Pacific and Southern Oceans // Diatom Res. 2003. V. 18. P. 203–224.
Barron J.A. Diatom biochronology for the Early Miocene of the Equatorial Pacific // Stratigraphy. 2005. V. 2. P. 281–309.
Barron J.A., Gladenkov A.Y. Early Miocene to Pleistocene diatom stratigraphy of Leg 145 // Proc. ODP. Sci. Res. College Station. TX (Ocean Drilling Program). 1995. V. 145. P. 3–19.
Barron J.A., Larsen B., Baldauf J.G. Evidence for late Eocene to early Oligocene Antarctic glaciation and observations on late Neogene glacial history of Antarctica: results from Leg 119 // Proc. ODP. Sci. Res. College Station. TX (Ocean Drilling Program). 1991. V. 119. P. 869–891.
Barron J.A., Stickley C.E., Bukry D. Paleoceanographic and paleoclimatic constraints on the global Eocene diatom and silicoflagellate record // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015. V. 422. P. 85–100.
Berggren W.A. Rates of evolution of some Cenozoic planktonic foraminifera // Micropaleontology. 1969. V. 15. P. 351–365.
Berggren W.A., Pearson P. Tropical to subtropical planktonic foraminiferal zonation of the Eocene and Oligocene // J. Foraminiferal Res. 2005. V. 35. P. 279–298.
Berggren W.A., Kent D.V., Swisher C.C.III, Aubry M.-P. A revised Cenozoic Geochronology and Chronostratigraphy // Geochronology, time scales and global stratigraphic correlation. Eds. Berggren W.A., Kent D.V., Aubry M.-P., Handerbol J. SEPM Spec. Publ. 1995. № 54. P. 129–212.
Blow W.H. Late middle Eocene to Recent planktonic foraminiferal biostratigraphy // Proc. First Int. Conf. on Planktonic microfossils, Geneva, 1967. Eds. Bronnimann P.R., Renz H.H. Leiden: E.J. Brill, 1969. V. 1. P. 199–421.
Bolli H.M. Planktonic foraminifers from the Oligocene-Miocene Cipero and Lengua formations of Trinidad, B.W.I. // U.S. Nat. Mus. Bull. 1957a. № 215. P. 97–123.
Bolli H.M. Planktonic foraminifers from the Eocene Navet and San Fernando formations of Trinidad, B.W.I. // U.S. Nat. Mus. Bull. 1957b. № 215. P. 155–172.
Bolli H.M. The genera Globigerina and Globorotalia in the Paleocene-lower Eocene Lizard Formation of Trinidad, B.W.I. // U.S. Nat. Mus. Bull. 1957c. № 215. P. 61–81.
Bolli H.M. Zonation of Cretaceous to Pliocene marine sediments based on planktonic foraminifera // Boletin Informativo Asociacion Venezolana de Geologia, Mineraria y Petroleo. 1966. V. 9. P. 3–32.
Bramlette M.N., Riedel W.R. Stratigraphic value of discoasters and some other microfossils related to recent coccolithophores // J. Paleontol. 1954. V. 28. P. 385–403.
Bukry D. Low-latitude coccolith biostratigraphic zonation // Init. Repts. DSDP. Washington: U.S. Government Printing Office, 1973. V. 15. P. 685–703.
Bukry D. Coccolith and silicoflagellate stratigraphy, northwestern Pacific Ocean, Deep Sea Drilling Project Leg 32 // Init. Repts. DSDP. Washington: U.S. Government Printing Office, 1975. V. 32. P. 677–701.
Cande S.C., Kent D.V. A new geomagnetic polarity time scale for the Late Cretaceous and Cenozoic // J. Geophys. Res. 1992. V. 97. P. 13917–13951.
Cande S.C., Kent D.V. Revised calibration of the geomagnetic polarity time scale for the Late Cretaceous and Cenozoic // J. Geophys. Res. 1995. V. 100. P. 6093–6095.
Cramer B.S., Toggweiler J.R., Wright J.D., Katz M.E., Miller K.G. Ocean overturning since the Late Cretaceous: inferences from a new benthic foraminiferal isotope compilation // Paleoceanography. 2009. V. 24. PA4216.
DePaolo D.J., Ingram B. High-resolution stratigraphy with strontium isotopes // Science. 1985. V. 227. P. 938–941.
Falkowski P.G., Katz M.E., Knoll A.H., Quigg A., Raven J.A., Schofield O., Taylor F.J.R. The evolution of modern eukaryotic phytoplankton // Science. 2004. V. 305. P. 354–360.
Farrell J.W., Clemens S.C., Gromet L.P. Improved chronostratigraphic reference curve of late Neogene seawater 87Sr/86Sr // Geology. 1995. V. 23. P. 403–406.
Geologic Time Scale 2020. Eds. Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Elsevier BV, 2020. 1357 p.
Geological History of the Polar Oceans: Arctic versus Antarctic. Eds. Bleil U., Thiede J. Kluwer Academic Publ., 1990. 834 p.
Gladenkov A.Yu. Bipolar distribution of some earliest Oligocene marine diatoms // Nova Hedwigia. 2014. Beih. 143. P. 337–368.
Haq B.U., Hardenbol J., Vail P.R. Chronology of fluctuating sea levels since the Triassic // Science. 1987. V. 235. P. 1156–1167.
Hardenbol J., Thierry J., Farley M.B., Jacquin T., de Graciansky P.-C., Vail P.R. Mesozoic and Cenozoic sequence chronostratigraphic framework of European basins // SEPM Spec. Publ. 1998. № 60. P. 3–13, 763–781.
Harwood D.M., Maruyama T. Middle Eocene to Pleistocene diatom biostratigraphy of Southern Ocean sediments from the Kerguelen Plateau, Leg 120 // Proc. ODP. Sci. Results. College Station. TX (Ocean Drilling Program). 1992. V. 120. P. 683–733.
Hay W.W., Mohler H., Roth P.H., Schmidt R.R., Boudreau J.E. Calcareous nannoplankton zonation of the Cenozoic of the Gulf Coast and Caribbean–Antillean area, and transoceanic correlation // Transactions of the Gulf Coast Association of Geological Societies. 1967. V. 17. P. 428–480.
Hodell D.A., Mueller P.A., McKenzie J.A., Mead G.A. Strontium isotope stratigraphy and geochemistry of the late Neogene ocean // Earth Planet. Sci. Lett. 1989. V. 92. P. 165–178.
International Stratigraphic Guide – An abridged version. Eds. Murphy M.A., Salvador A. // Episodes. 1999. V. 22. P. 255–271.
Keller G., Barron J.A. Paleoceanographic implications of Miocene deep-sea hiatuses // Bull. Geol. Soc. Am. 1983. V. 94. P. 590–613.
Keller G., Barron J.A. Paleodepth distribution of Neogene deep-sea hiatuses // Paleoceanography. 1987. V. 2. P. 697–713.
Kennett J.P. Marine Geology. Prentice-Hall: Englewood Cliffs, 1982. 813 p.
Koepnick R.B., Burke W.H., Denison R.E., Hetherington E.A., Nelson H.F., Otto J.B., Waite L.E. Construction of the seawater 87Sr/86Sr curve for the Cenozoic and Cretaceous: supporting data // Chem. Geol. 1985. V. 58. P. 55–81.
Martini E. Standard Tertiary and Quaternary calcareous nannoplankton zonation // Proc. Second Planktonic Conf., Roma, 1970. Ed. Farinacci A. Roma: Edizioni Tecnoscienza, 1971. V. 2. P. 739–785.
Matsuoka A. Jurassic and Early Cretaceous radiolarians from Leg 129, Sites 800 and 801, Western Pacific Ocean // Proc. ODP. Sci. Results. College Station. TX (Ocean Drilling Program). 1992. V. 129. P. 203–220.
McArthur J.M., Howarth R.J., Bailey T.R. Strontium isotope stratigraphy: LOWESS Version 3: Best fit to the marine Sr-Isotope curve for 0–509 Ma and accompanying look-up table for deriving numerical age // J. Geology. 2001. V. 109. P. 155–170.
McArthur J.M., Howarth R.J., Shields G.A., Zhou Y. Strontium Isotope Stratigraphy // Geologic Time Scale 2020. Eds. Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Elsevier BV, 2020. P. 211–238.
Miller K.G., Fairbanks R.G., Mountain G.S. Tertiary isotope synthesis, sea level history, and continental margin erosian // Paleoceanography. 1987. V. 2. P. 1–19.
Miller K.G., Wright J.D., Fairbanks R.G. Unlocking the Ice House: Oligocene–Miocene oxygen isotopes, eustasy, and margin erosion // J. Geophys. Res. 1991. V. 96 (B4). P. 6829–6848.
Miller K.G., Browning J.V., Schmelz W.J., Kopp R.E., Mountain G.S., Wright J.D. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records // Science Advances. 2020. V. 6 (20), eaaz1346.
Ogg J.G., Ogg G.M., Gradstein F.M. Concise Geologic Time Scale 2016. Elsevier, 2016. 229 p.
Okada H., Bukry D. Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975) // Marine Micropaleontology. 1980. V. 5. P. 321–325.
Page K.N. The Lower Jurassic of Europe: its subdivision and correlation // GEUS Bull. 2003. V. 1. P. 23–59.
Plankton Stratigraphy. Eds. Bolli H.M., Saunders J.B., Perch-Nielsen K. Cambridge: Cambridge Univ. Press, 1985. 1032 p.
Productivity of the Oceans: Present and Past. Eds. Berger W.H., Smetacek V.S., Wefer G. New York: Wiley-Interscience, 1989. 471 p.
Raffi I., Agnini C., Backman J., Catanzariti R., Pälike H. A Cenozoic calcareous nannofossil biozonation from low and middle latitudes: a synthesis // J. Nannoplankton Res. 2016. V. 36. P. 121–132.
Seibold E., Berger W.H. The sea floor. An Introduction to Marine Geology. Berlin, New York: Springer-Verlag, 1982. 288 p.
The Geologic Time Scale 2012. Eds. Gradstein F.M., Ogg J.M., Schmitz M.D., Ogg G.M. Elsevier, 2012. 1144 p.
The Miocene Ocean: Paleoceanography and Biogeography. Ed. Kennett J.P. // Mem. Geol. Soc. Am. 1985. V. 163. 343 p.
Vail P.R., Hardenbol J. Sea-level changes during the Tertiary // Oceanus. 1979. V. 22. P. 71–79.
Wade B.S., Pearson P.N., Berggren W.A., Pälike H. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale // Earth-Sci. Rev. 2011. V. 104. P. 111–142.
Zachos J., Pagain M., Sloan L., Thomas E., Billups K. Trends, rhythms, and aberrations in global climate 65 Ma to present // Science. 2001. V. 292. P. 686–691.
Zachos J.C., Dickens G.R., Zeebe R.E. An Early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics // Nature. 2008. V. 451. P. 279–283.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция