

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 3, стр. 45-71
Силикофоссилии и диноцисты из терригенно-кремнистых отложений нижнего палеогена юга Русской плиты и их значение для обоснования возраста свит
Т. В. Орешкина 1, *, А. И. Яковлева 1, Г. Н. Александрова 1
1 Геологический институт РАН
Москва, Россия
* E-mail: oreshkina@ginras.ru
Поступила в редакцию 16.07.2020
После доработки 16.08.2020
Принята к публикации 08.12.2020
Аннотация
Эоценовые силикофоссилии (диатомовые водоросли и силикофлагеллаты), цисты динофлагеллат и континентальные палиноморфы изучены в двух скважинах Преддонецкой и Павловско-Кантемировской структурно-фациальных зон Донецкой синеклизы. Вёшенская свита, включающая комплекс диноцист зоны Stenodinium meckelfeldense, отнесена к раннему ипру. Тишкинская и касьяновская свиты, охарактеризованные комплексом диатомей зоны Bipalla oamaruensis и первым появлением силикофлагеллаты Corbisema hexacantha, так же как и комплекс диноцист с Enneadocysta pectiniformis, датированы поздним лютетом–ранним приабоном. Комплексы микрофоссилий типичны для мелководных краевых частей эпиконтинентального палеобассейна.
ВВЕДЕНИЕ
На территории Российской Федерации один из блоков широкого развития морских палеогеновых отложений приурочен к северной части Днепровско-Донецкой впадины. Здесь они представлены практически всем разнообразием терригенных и терригенно-кремнистых фаций внутриконтинентального морского бассейна с фрагментарным присутствием в разрезах карбонатных отложений.
Первые работы по диатомеям и силикофлагеллатам в рассматриваемом регионе связаны c именами А.П. Жузе (1949) и Ю.М. Успенской (1950). В 1960–1970 гг. изучение диатомей на территории Днепровско-Донецкой впадины продолжили В.С. Шешукова-Порецкая, З.И. Глезер (Шешукова-Порецкая, Глезер, 1964; Глезер и др., 1965; Глезер, Шешукова-Порецкая, 1968, 1969; Глезер, 1974, 1979) и А.П. Ольштынская (Палатная, 1976; Ольштынская, 1976, 1977, 1978а, 1978б; Ольштинська, 2008). В 1990-е гг. в регионе было начато комплексное микропалеонтологическое изучение палеогеновых отложений (Радионова и др., 1994; Radionova et al., 2003; Радионова, Орешкина, 2016; Khokhlova et al., 1999; Popova et al., 2002; Горбаткина, Иосифова, 2004; Орешкина, Яковлева, 2007; Бугрова и др., 2016). Для расчленения по диатомовым водорослям использовались различные модификации зональных диатомовых шкал З.И. Глезер (1979; Зональная…, 2006) и Н.И. Стрельниковой (1991). В региональную схему палеогеновых отложений юга Русской плиты, утвержденную МСК (Ахметьев, Беньямовский, 2003), вошел синтез зональных шкал З.И. Глезер и Н.И. Стрельниковой, однако детальная характеристика диатомовых зон и стратотипические разрезы, в которых эти подразделения были установлены, в этой публикации, а также и в дальнейшем не рассматривались.
Начало палинологическим исследованиям (изучению спор и пыльцы наземных растений) палеогеновых отложений Воронежской антеклизы было положено еще в 1950-е годы С.Н. Наумовой (Шпуль, 2010), Е.Д. Заклинской (1953), А.А. Чигуряевой (1956). Позднее континентальные палиноморфы здесь изучались В.Г. Шпуль (1972, 1998, 2005, 2006, 2007, 2008, 2009, 2010), А.Б. Стотландом (Крашенинников, Ахметьев, 1996) и Л.А. Пановой (Горбаткина, Иосифова, 2004).
Изучение цист динофлагеллат в Воронежской антеклизе на настоящий момент носит фрагментарный характер. Первые упоминания о диноцистах даны в работе А.А. Чигуряевой (1956). Н.И. Запорожец были изучены комплексы диноцист из аналогов кумской свиты (киевской и дерезовской) в разрезах скважин в окрестностях г. Новый Оскол и сопоставлены с зонами Areosphaeridium diktyoplokum–Wilsodinium intermedium (верхний лютет–нижний бартон) и Rhombodinium porosum–Rhombodinium perforatum (верхний бартон–низы приабона) (Запорожец, 2001; Горбаткина, Иосифова, 2004).
В.Г. Шпуль (2005) в разрезах скважин в районе с. Пасеково установила в верхнекиевской подсвите комплекс диноцист зоны Rhombodinium porosum–Rhombodinium perforatum и в обуховской свите – Charlesdowniea clathrata angulosa (=Talladinium? angulosum) (приабон). По данным А.И. Яковлевой (Iakovleva, 2015; Яковлева, 2017), диноцистовые ассоциации, изученные в скв. 230 (в 30 км севернее г. Харькова), позволяют отнести бучакскую свиту в этой части Северной Украины к среднему–верхнему лютету, киевскую свиту – к бартону–низам приабона, а нижнюю часть обуховской свиты – к приабону.
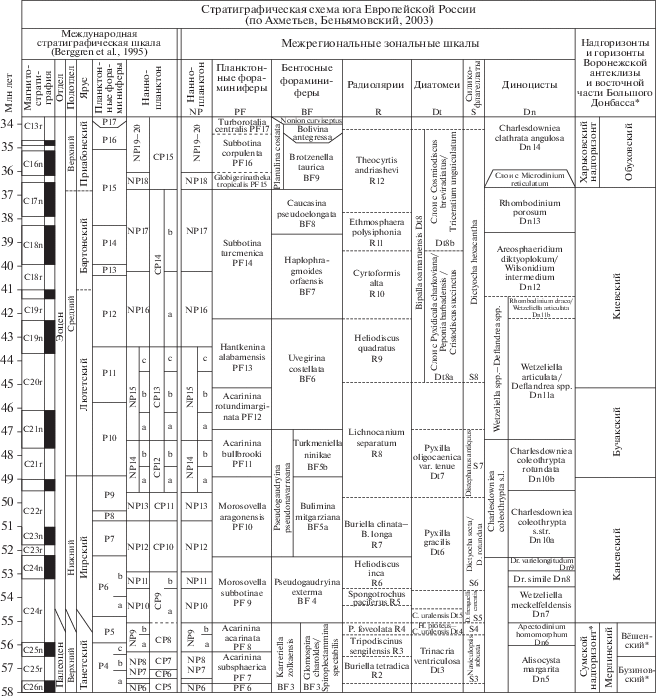
Следует отметить, что в настоящее время региональная стратиграфическая схема (рис. 1), принятая Межведомственным стратиграфическим комитетом (МСК) в 2000 г. (Постановления…, 2001), нуждается в обновлении. В первую очередь это связано с тем, что в схеме использованы названия горизонтов (сумской, каневский, бучакский, харьковский и т.д.), стратотипы которых находятся на территории Украины, тогда как их гипостратотипы в России не утверждены, что не соответствует требованиям Стратиграфического кодекса России (2019). В утвержденной МСК стратиграфической схеме палеогеновых отложений для юга Европейской России (Ахметьев, Беньямовский, 2003) отсутствует свитное наполнение горизонтов, необходимое как для крупномасштабного картирования, так и для более четкого понимания региональных особенностей осадконакопления. Более того, назрела необходимость учесть новые данные по основным группам микропланктона, появившиеся за два последних десятилетия.
Рис. 1.
Региональная стратиграфическая схема палеогена юга Русской плиты (по Ахметьев, Беньямовский, 2003). Звездочкой (*) отмечены региональные стратиграфические подразделения восточной части Большого Донбасса. Сокращения: P. – Petalospyris; C. – Coscinodiscus; H. – Hemiaulus; Dr. – Dracodinium.
По-прежнему актуальной задачей остается и комплексное биостратиграфическое изучение стратотипических разрезов свит палеогена, установленных Г.П. Леоновым (1961) и В.П. Семеновым (1965), а также поиск новых, более полных парастратотипов. Отсутствие на большей части этого региона в палеогеновых отложениях известкового наннопланктона и фораминифер выводит на первый план совместное изучение кремневого и органикостенного планктона в разрезах, которое может обеспечить надежную корреляцию отложений с Международной стратиграфической шкалой (МСШ).
Первая попытка перехода в регионе на российские стратиграфические подразделения была предпринята Т.Е. Горбаткиной и Ю.И. Иосифовой (2004). Ими были предложены воробьевская свита (это название оказалось преоккупированным) в качестве аналога киевского горизонта эоцена и пасековская свита для нижней части межигорского горизонта олигоцена.
Э.М. Бугровой с соавторами (2016) была обоснована замена киевской свиты из стратиграфической схемы Украины на две свиты – сергеевскую и тишкинскую, выделенные В.П. Семеновым (1965). В качестве опорного разреза сергеевской и касьяновской свит и гипостратотипа тишкинской свиты был предложен разрез на окраине г. Кантемировка Воронежской области. В сергеевской свите установлены биостратоны по наннопланктону, планктонным и бентосным фораминиферам: NP16 Discoaster tani nodifer, слои с Acarinina kiewensis и зона Robulus dualis соответственно. Возраст сергеевской свиты определялся как переходный от лютета к бартону. Позднее В.А. Мусатовым (2020) в свите были установлены наннопланктонные зоны CNE12 и CNE13 по шкале C. Agnini et al. (2011, 2014), которые соответствуют верхнему лютету МСШ. Комплекс радиолярий из верхней части сергеевской свиты был отнесен к зоне Heliodiscus quadratus верхнего лютета–нижнего бартона (Козлова, 1999). Диноцисты и континентальные палиноморфы в разрезе не были обнаружены. Граница сергеевской и тишкинской свит проводится по смене карбонатного осадконакопления на терригенно-кремнистое и совпадает с исчезновением карбонатных фоссилий и развитием кремневых групп микроорганизмов, что соответствует рубежу керестинского и кумского горизонтов Северо-Кавказского региона.
В гипостратотипе тишкинской свиты в ее нижней части выделены местные слои с Pseudohastigerina micra по планктонным фораминиферам и слои с Haplophragmoides kiewensis по бентосным фораминиферам, датируемые бартоном (Бугрова и др., 2016). На этом же уровне в свите была выделена небогатая ассоциация кокколитов средней и плохой сохранности (практически все несут следы растворения), которые в еще меньшем количестве встречаются на отдельных уровнях в ее более высоких горизонтах (Мусатов, 2020). На основании присутствия единичных Chiasmolithus solitus В.А. Мусатовым, с долей условности, было высказано предположение о возможной принадлежности свиты к зоне CNE14 пограничного интервала лютета и бартона. Тишкинская и касьяновская свиты в разрезе Кантемировка охарактеризованы диатомовыми водорослями и силикофлагеллатами зоны Bipalla oamaruensis. На основании опосредованных корреляций возраст зоны принят как временной интервал бартона–раннего приабона (Бугрова и др., 2016). По радиоляриям в тишкинской свите установлены зоны Cyrtophormis alta и Ethmosphaera polysiphonia, датируемые бартоном, в нижней части касьяновской свиты – зона Theocyrtis andriashevi нижнего приабона (Козлова, 1999). Г.Э. Козлова (1999) отмечала, что ввиду отсутствия в изученных разрезах Воронежской антеклизы совместных находок радиолярий и ортостратиграфических групп фауны и флоры, возраст радиоляриевых зон и их стратиграфическая позиция определены условно, в соответствии с положением вмещающих толщ: кумской свите и ее аналогам отвечают зоны C. alta и E. polysiphonia, нижней части белоглинской свиты и ее аналогам – зона T. andriashevi.
Следует отметить, что в МСШ пока окончательно не установлены критерии распознавания границы бартонского яруса и еще только принимается нижняя граница приабонского яруса (Agnini et al., 2020). В принятой ОСШ России (Николаева и др., 2006) граница лютета/бартона сопоставляется с границей зон Hantkenina alabamensis и Globigerina turcmenica по планктонным фораминиферам, что отвечает рубежу керестинского и кумского горизонтов, а граница бартона/приабона – с границей зон Globigerina turcmenica и Globigerapsis tropicalis s.l. по планктонным фораминиферам, что отвечает рубежу кумского и белоглинского горизонтов Предкавказья.
Целью настоящей статьи является комплексный микропалеонтологический анализ эоценовых отложений из разрезов двух скважин, пробуренных в стратотипической местности развития региональных подразделений палеогена Преддонецкой и Павловско-Кантемировской структурно-фациальных зон, для получения детальной характеристики зон по диноцистам, диатомовым водорослям и силикофлагеллатам, для уточнения возраста местных стратиграфических подразделений.
МАТЕРИАЛ И МЕТОДЫ
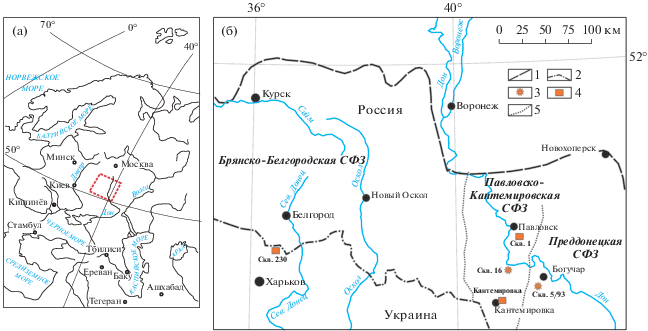
В рамках настоящего исследования были изучены образцы из скважин № 5/93 (Монастырщина) и № 16 (Богучар), пробуренных в Преддонецкой и Павловско-Кантемировской структурно-фациальных зонах (СФЗ) Донецкой синеклизы (Александрова и др., 2020) (рис. 2). В Преддонецкой СФЗ выделяются три крупных этапа осадконакопления. Так, первый, позднепалеоценовый–раннеэоценовый, трансгрессивный этап представлен бузиновской, вёшенской свитами, суровикинскими и осиновскими слоями в понимании Г.П. Леонова (1961). Второй, средне-позднеэоценовый, трансгрессивно-регрессивный этап представлен тишкинской и касьяновской свитами. Третий трансгрессивно-регрессивный этап представлен мощной толщей кварцевых песков, в которых практически невозможно выделить свиты. По особенностям строения и состава нижняя часть толщи соотносится с венцовской свитой Г.П. Леонова, которая, в свою очередь, сопоставляется им с нижней частью майкопской свиты Предкавказья (Леонов, 1961). В Павловско-Кантемировской СФЗ палеоген представлен непрерывной последовательностью свит среднеэоценового–олигоценового возраста. Палеоценовые и раннеэоценовые отложения в данной СФЗ отсутствуют. Это единственный регион в Донецкой синеклизе, где установлен карбонатный тип пород (мергели, карбонатные глины сергеевской свиты).
Рис. 2.
Местоположение изученных скважин и опорных разрезов. 1 – граница распространения палеогеновых отложений; 2 – госграница РФ; 3 – изученные разрезы скважин; 4 – опорные разрезы; 5 – границы структурно-фациальных зон.
Скважина № 5/93 (Монастырщина) пробурена на территории совхоза Радченский, в 16 км к юго-западу от города Богучар Воронежской области (координаты 50°24′ с.ш., 31°40′23′′ в.д.). Образцы керна были отобраны в кернохранилище Павловской геологической партии сотрудниками ГИН РАН Э.П. Радионовой, В.Н. Беньямовским и Т.В. Орешкиной в 1994 г., ими же было составлено описание разреза. Позже этот материал был выборочно (8 образцов из 30) изучен на карбонатные и кремневые группы микропланктона (Khokhlova et al., 1999). В публикации И.Е. Хохловой с соавторами для расчленения отложений скважины была использована схема В.И. Курлаева и Е.Ф Ахлестиной (1988) для нижнего течения Волги и Дона из-за специфического строения разреза с мощной (до 31 м) пачкой песков и песчаников в основании киевского горизонта (рис. 3). Впоследствии Т.Е. Горбаткина и Ю.И. Иосифова (2004) отнесли песчаную толщу и вышележащие отложения опок, опоковидных глин и алевритов разреза скв. 5/93 к нижней части выделенной ими воробьевской свиты в качестве аналога киевской свиты на Украине. Ниже приведено послойное описание керна скв. 5/93 (Монастырщина), при этом предлагаемое в настоящей статье свитное деление отличается от опубликованного ранее в работе (Khokhlova et al., 1999).
Рис. 3.
Биостратиграфия, диапазоны распространения стратиграфически значимых видов силикофоссилий и диноцист в скв. 5/93 (Монастырщина). Условные обозначения: 1 – песчаники, кварциты; 2 – пески; 3 – известняки; 4 – опоки, кремнистые алевролиты; 5 – опоковидные глины; 6 – песчанистые опоки; 7 – мергели; 8 – глаукониты; 9а – ходы илоедов; 9б – фосфориты; 10а – моллюски; 10б – брекчированность пород; 11 – карбонатные глины. Сокращения: Полт. – полтавская свита. Звездочкой (*) отмечены таксоны силикофлагеллат.
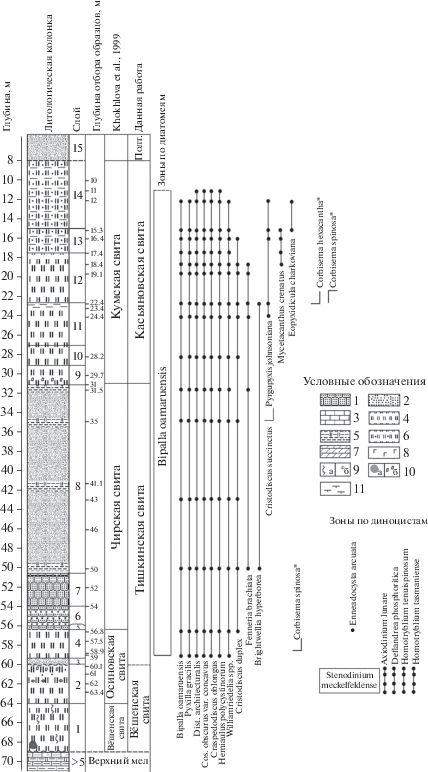
На меловых мергелях с размывом залегают снизу вверх (рис. 3):
Вёшенская свита:
Слой 1. Опока серая, песчанистая, пятнистая, слабокарбонатная, с ходами илоедов и мелкими раковинами двустворок. Интервал 68.8–64.0 м.
Слой 2. Близкая по составу опока, в интервале 61.8–61.0 м существенно обогащенная фосфоритовыми желваками, выше опесчаненная, в верхней части слоя брекчированная. Интервал 64.0–59.4 м.
Тишкинская свита:
Слой 3. Песок мелкозернистый, светлый, с мелкими пятнами ожелезнения. Интервал 59.4–58.6 м.
Слой 4. Опока светло-серая, плотная, легкая, с пятнами ожелезнения в средней части толщи. Интервал 58.6–56.4 м.
Слой 5. Песчаники кварцитовидные, темно-серые, с мелкозернистым цементом. Интервал 56.4–56.0 м.
Слой 6. Песчаник слабосцементированный, опоковидный, с тонкими прослоями опок. Интервал 56.0–54.0 м.
Слой 7. Сливной песчаник. Интервал 54.0–50.4 м.
Слой 8. Песок светлый, мелкозернистый, глауконитовый, с прослоями опоковидных глин. В верхней части отмечается узловатость за счет неравномерного окремнения. Интервал 50.4–31.0 м.
Касьяновская свита:
Слой 9. Опока светло-серая, массивная, со светлыми пятнами ожелезнения, в нижней части брекчиевидная, пестрая, с зеленоватыми и вишневыми пятнами, слабокарбонатная. Интервал 31.0–29.0 м.
Слой 10. Опока розовато-бордовая, неравномерно-слоистая. Интервал 29.0–27.0 м.
Слой 11. Опока более светлая, бордовая, тонкоплитчатая, с неравномерной красноцветностью. В верхней части (23.4–22.6 м) прослой глины. Интервал 27.0–22.6 м.
Слой 12. Опока светлая, почти белая, массивная, на уровне 17.4 м красноцветный прослой. Интервал 22.4–17.4 м.
Слой 13. Опока песчанистая, светлая, слабоожелезненная, с глинистыми прослоями. Интервал 17.4–14.8 м.
Слой 14. Оповидный песчаник светлый, с глинистыми прослоями. Интервал 14.8–8.0 м.
Выше по разрезу толща песков предположительно аналога полтавской свиты мощностью около 4 м.
В рамках настоящего исследования из разреза скважины было изучено 30 образцов.
Скважина 16 (Богучар) пройдена в 1 км восточнее северо-восточной окраины с. Ивановка Россошанского района Воронежской области и в 4.4 км по азимуту 289° от высоты 182.7 м на водораздельной поверхности. Абсолютная отметка устья скважины 204.0 м, глубина 221.5 м, координаты 50°00′26′′ с.ш., 40°01′53′′ в.д. Скважина пробурена в 2000 г. в рамках работы по геологическому доизучению масштаба 1 : 200 000 на площади листа М-37-XVII (Павловск). Описание палеогеновой части разреза скв. 16 дано по материалам отчета по этим работам (Радьков и др., 2002). Образцы керна на микропалеонтологический анализ были получены от Э.М. Бугровой (ВСЕГЕИ). Снизу вверх на меловых отложениях в разрезе вскрываются (рис. 4):
Рис. 4.
Биостратиграфия, диапазоны распространения стратиграфически значимых видов силикофоссилий и диноцист в скв. 16 (Богучар). Условные обозначения см. на рис. 3. Сокращения: Кант. – кантемировская свита. Звездочкой (*) отмечены таксоны силикофлагеллат.
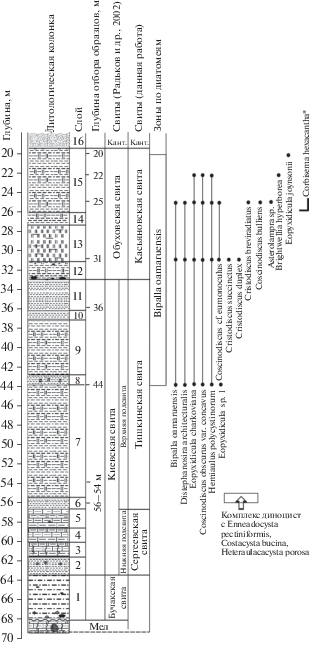
Бучакская свита:
Слой 1. Алевролит ярко-зеленый, крупноалевритовый, слюдисто-глауконитового состава, жирный на ощупь, с неравномерно распределенным гравием черных кремней. Характерны агрегатные скопления глауконита размером 0.3–0.5 мм. В подошве слоя (0.1 м) алеврит с конкрециями коричнево-черных фосфоритов размером до 5 см. Интервал 68.2– 63.8 м.
Сергеевская свита:
Слой 2. Глина темно-зеленая, карбонатная, слюдистая, мелкощебенчатая, при ударе раскалывается на мелкие остроугольные отдельности. По всему слою наблюдаются обохренные песчаные конкреции фосфоритов размером до 2–3 см. Интервал 63.8–61.8 м.
Слой 3. Мергель серый, глинистый, сильноалевритистый, слюдистый. Алеврит в породе представлен тонкими зернами слюды, кварца, иногда размером до 0.1–0.2 мм. Часто наблюдаются нитевидные ходы червей темно-серой окраски. Интервал 61.8–60 м.
Слой 4. Мергель серый с темно-зеленым оттенком, алевритистый, слюдистый, сильноглинистый, во влажном состоянии мягкий до рыхлого. Интервал 60–58.5 м.
Слой 5. Мергель серый, сильноглинистый, слюдистый, слабоалевритистый, с крупной кусковатой отдельностью, с редкими мелкими зернами черного окисленного глауконита. Интервал 58.5–56.6 м.
Тишкинская свита:
Слой 6. Глина темно-зеленая, сильно перемятая, передробленная, с мелкой кусковатой отдельностью, участками превращена в труху. Интервал 56.6–55.6 м.
Слой 7. Глина светло-серая со слабым зеленоватым оттенком, опоковидная, плотная, с раковистым изломом, слабослюдистая, алевритистая. Алеврит тонкий, кварцево-глауконитовый, слабослюдистый. Встречены спикулы губок. Интервал 55.6–43.8 м.
Слой 8. Глина в нижней части слоя темно-зеленая, выше серая с зеленоватым оттенком. Сильно перемята, передроблена, с мелкой кусковатой отдельностью, участками превращена в труху. Интервал 43.8–43.0 м.
Слой 9. Глина опоковидная, бледно-зеленой окраски, слабослюдистая, алевритистая. В сухом состоянии тонкоплитчатая. По плоскостям наслоения часто присыпки светло-серого алеврита. Интервал 43.0–37.6 м.
Слой 10. Глина светло-серая до белесой, с кремоватым оттенком, кремнеземистая, легкая, в сухом состоянии с полураковистым изломом, слабослюдистая. В подошве слоя (0.2 м) глина становится более алевритистой. Алеврит в породе кварцево-слюдистого состава. Зерна кварца размером 0.1–0.2 мм. Интервал 37.6–36.2 м.
Слой 11. Глина, до глубины 35.0 м темно-зеленая, выше – серая с зеленоватым оттенком, слюдистая, алевритистая, запесоченная, с многочисленными ходами илоедов. Интервал 36.2–33.0 м.
Касьяновская свита:
Слой 12. Глауконитово-глинисто-кремнеземистая порода зеленовато-серая с белесыми пятнами, сильноалевритистая. Алеврит в породе кварцево-слюдисто-глауконитового состава. Глауконит в виде крупных зерен размером до 0.1–0.2 мм распределен неравномерно, в подошве слоя его содержание увеличивается до 10%. Интервал 33.0–31.6 м.
Слой 13. Глина светло-серая до белесой, кремнеземистая, опоковидная, более легкая по сравнению с глиной из слоя 12 (видимо, кремнеземистый материал преобладает), слабоалевритистая, с редкими вкраплениями темного глауконита. Интервал 31.6–27.4 м.
Слой 14. Глина серая до белесой, аналогичная глинам слоя 13, но с примесью темных (от зеленых до черных) зерен глауконита до 15–20%. Интервал 27.4–26.0 м.
Слой 15. Глина светло-серая до белесой, в верхней части со слабым зеленоватым оттенком, кремнеземистая, опоковидная, алевритистая, слюдистая, иногда с оранжевыми разводами ожелезнения. Отмечено присутствие спикул губок. Интервал 26.0–19.4 м.
Выше по разрезу залегают пески и алевролиты кантемировской свиты.
В нижней части тишкинской свиты установлен единственный уровень с диноцистами (обр. 54–55 м), присутствие силикофоссилий приурочено к интервалу 44–20 м.
Химическую обработку образцов на диатомовый и палинологический анализ проводили в соответствии со стандартной методикой, используемой в лабораториях палеофлористики и микропалеонтологии ГИН РАН (Александрова и др., 2012).
При таксономической идентификации диатомей были учтены изменения в названиях родов, оказавшихся преоккупированными (Blanco, Wetzel, 2016). Полуколичественную оценку общего содержания диатомей и отдельных видов проводили по балльной системе при увеличении ×400: A (Abundant) = 1 или более экземпляров в каждом поле зрения (ПЗ); C (Common) = 1 экземпляр в 2 ПЗ; F (Few) = 1–2 экземпляра в каждом ряду покровного стекла; R (Rare) = несколько экземпляров в препарате; B (Barren) = полное отсутствие диатомей или их фрагментов; T (Trace) = редкие фрагменты створок. Сохранность диатомей оценивали следующим образом: G (Good) = минимальное количество обломков; M (Moderate) = = примерно равное количество обломков и целых створок; P (Poor) = целые створки встречаются редко. Микросъемку проводили в световом микроскопе Motic BA310 и в сканирующем микроскопе Vega ©Tescan MV-2300.
Морские диатомовые водоросли подразделяются на пелагические (или открыто-морские) и неритические, характерные для шельфовых пространств. Более разнообразная в таксономическом отношении неритическая группа представлена планктонными, бентосными и тихопелагическими видами. Экологические и климатические характеристики ископаемых видов, как правило, определяются распространением ныне живущих родов. Палеоэкологические интерпретации среднеэоценовых таксонов диатомей основаны, в первую очередь, на работе Witkowski et al. (2014).
Для зонального расчленения по диатомеям за основу взята схема З.И. Глезер с изменениями (1979; Зональная..., 2006).
Таксономия диноцист соответствует индексу DINOFLAJ3 (Williams et al., 2017). Количественное палинологическое изучение материала из интервала с продуктивными ассоциациями палиноморф проводилось в 2 этапа: (1) подсчет в каждом образце минимум 200 морских и наземных палиноморф (цисты динофлагеллат, акритархи, пыльца покрытосеменных и хвойных растений, споры наземных растений); (2) дальнейший подсчет минимум 200 диноцист, после чего препарат просматривался до конца с целью обнаружения редко встречаемых таксонов. Для интерпретации возможных палеобстановок цисты динофлагеллат были объединены в 13 групп морфологически близких таксонов: (1) дефляндровые; (2) ветзелиеллоидные, (3) Phthanoperidinium; (4) Homotryblium-группа; (5) Areoligera-группа; (6) Carpatella rossica; (7) Tectatodinium pellitum; (8) Cordosphaeridium-группа, (9) Alisocysta sp. 2; (10) Operculodinium; (11) Spiniferites-группа; (12) Impagidinium; (13) остальные гониаулакоидные. Палеоэкологическая интерпретация количественных флуктуаций в палинокомплексах основана на работах (Brinkhuis, 1994; Powell et al., 1996; Crouch, Brinkhuis, 2005; Sluijs et al., 2005; Iakovleva, 2015; Iakovleva, Heilmann-Clausen, 2020).
РЕЗУЛЬТАТЫ
Скв. 5/93 (Монастырщина). В основании разреза палеогена, в вёшенской свите (интервал 60.1–63.4 м, слой 2), установлен количественно и таксономически представительный комплекс палиноморф (рис. 3, 5, табл. 1). Палинологические ассоциации в этом интервале характеризуются чередованием доминирования диноцист (25–60%) и пыльцы покрытосеменных растений (25–50%); содержание двухмешковой пыльцы хвойных, спор и акритарх не превышает 10% для каждой группы. В свою очередь, в ассоциациях диноцист наибольших количественных значений достигают группы дефляндроидных (~20–40%), Areoligera (10–20%), Carpatella rossica (1–26%) и Spiniferites (~15%). Такой состав комплекса палиноморф свидетельствует о мелководных прибрежных обстановках.
Рис. 5.
Количественное распределение групп палиноморф и экогрупп диноцист в разрезе скв. 5/93 (Монастырщина). Условные обозначения см. на рис. 3.
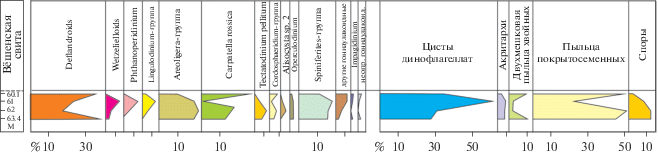
Таблица 1.
Стратиграфическое распределение цист динофлагеллат в скв. 5/93 (Монастырщина)
| Свита | Зоны по диноцистам (Яковлева, 2017) | Глубина отбора образцов, м | Achomosphaera ramulifera | Achomosphaera ramulifera-group | Alisocysta sp. 2 CHC 1985 | Apectodinium homomorphum | Areoligera coronata | Areoligera senonensis | Axiodinium aff. lunare | Carpatella rossica | Cordosphaeridium fibrospinosum | Cordosphaeridium gracile | Dapsilidinium pseudocolligerum | Deflandrea denticulata | Deflandrea oebisfeldensis | Deflandrea phosphoritica | Glaphyrocysta ordinata | Hafniasphaera septata | Homotryblium tasmaniense | Hystrichosphaeridium tubiferum | Hystrichosphaeropsis obscura | Impagidinium victorianum | Lingulodinium machaerophorum | Microdinium ornatum | Operculodinium microtrianum | Phthanoperidinium crenulatum | Polysphaeridium zoharyi | Rottnestia borussica | Spiniferites ramosus-group | Tectatodinium pellitum | Alterbidinium rugulum | Apectodinium quinquelatum-paniculatum | Areoligera medusettiformis | Caligodinium aceras | Cordosphaeridium inodes | Diphyes colligerum | Heteraulacacysta campanula | Hystrichostrogylon membraniphorum | Melitasphaeridium pseudorecurvatum | Membranilarnacia sp. cf. Eatonicysta sensu DC 1996 | Memranosphaera sp. B in CHC 1985 | Oligosphaeridium complex | Operculodinium centrocarpum | Palaeotetradinium minusculum | Thalassiphora delicata | Cerodinium diebelii | Cladopyxidium saeptum | Eocladopyxis peniculata | Fromea fragilis | Glaphyrocysta exuberans | Hystrichokolpoma unispinum | Impagidinium dispertitum | Nematosphaeropsis sp. | Thalassiphora patula | Axiodinium lunare | Cleistosphaeridium diversispinosum | Cordosphaeridium multispinosum | Cribroperidinium tenuitabulatum | Elytrocysta brevis | Homotryblium aculeatum | Homotryblium tenuispinosum | Impagidinium aculeatum | Trigonopyxidia ginella | Gen. Spec. Indet 1 CHC-LC 1989 | Enneadocysta arcuata |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Тишкинская | Не выделены | 41 |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 43 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 50 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 54 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 55 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 56.8 | 1 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 57.5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 58.2 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 58.6 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 59 | 5 | 2 | 1 | 3 | 3 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Вёшенская | Stenodinium meckelfeldense | 60.1 | 1 | 1 | 20 | 60 | 0 | 6 | 1 | 80 | 1 | 1 | 2 | 22 | 0 | 0 | 2 | 1 | 1 | 1 | 3 | 1 | 2 | 0 | 1 | 0 | 0 | 1 | 1 | 0.5 | 1 | 1 | 4 | ||||||||||||||||||||||||||||||||
| 61.8 | 1 | 11 | 3 | 0 | 39 | 4 | 3 | 1 | 0 | 6 | 0 | 47 | 1 | 2 | 4 | 3 | 2 | 2 | 2 | 3 | 15 | 10 | 28 | 9 | 1 | 9 | 0 | 2 | 0 | 1 | 1 | 0 | 2 | 1 | 0 | 1 | 1 | 1 | 4 | 1 | 0 | 0 | 1 | 0 | |||||||||||||||||||||
| 62.1 | 1 | 27 | 7 | 43 | 38 | 0 | 6 | 0 | 0 | 0 | 1 | 4 | 0 | 2 | 0 | 3 | 1 | 1 | 1 | 3 | 1 | 14 | 1 | 3 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 4 | 0 | 1 | ||||||||||||||||||||||||||||
| 63.4 | 2 | 8 | 1 | 0 | 32 | 1 | 1 | 27 | 2 | 0 | 1 | 1 | 80 | ?0 | 3 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0.5 | 1 | 20 | 8 |
В комплексе диноцист (табл. 1) группа стратиграфически значимых видов представлена Axiodinium lunare (Gocht) Williams et al., Deflandrea phosphoritica Eisenack, Homotryblium tenuispinosum Davey et Williams, H. tasmaniense Cookson et Eisenack и Membranilarnacia sp. cf. E-atonicysta sensu De Coninck, 1996. Это позволяет соотнести данный интервал разреза с диноцистовой зоной Stenodinium meckelfeldense шкалы восточного Перитетиса (Яковлева, 2017). Соответственно, отложения в скважине на глубинах 63.4–60.1 м датированы ранним ипром.
Выше по разрезу палиноморфы либо встречены единично, либо вообще отсутствуют. При этом диноцисты представлены таксонами широкого стратиграфического распространения, и лишь на глубине 56.8 м обнаружен единственный экземпляр Enneadocysta arcuata (Eaton) Stover et Williams, указывающий на то, что стратиграфически эта часть разреза не может быть древнее лютета.
Диатомовые водоросли и силикофлагеллаты (рис. 3, табл. 2, табл. I–VI ) присутствуют в разрезе, начиная с подошвы тишкинской свиты (обр. 59.0 м). В интервале 59–35 м комплекс представлен в основном фрагментами створок диатомей и спикулами губок. С основания касьяновской свиты (гл. 31 м) возрастает доля целых створок диатомей, присутствуют силикофлагеллаты, единичные асцидии и эбриидеи.
Таблица 2.
Стратиграфическое распределение силикофоссилий в скв. 5/93 (Монастырщина) и в скв. 16 (Богучар)
| Скв. 5/93 Монастырщина | Скв. 16 Богучар | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| № образцов | 56.8 м | 50 м | 43 м | 35 м | 31.5 м | 28.2 м | 24.4 м | 22.4 м | 19.1 м | 18.4 м | 17.4 м | 16.4 м | 15.3 м | 12 м | 11 м | 45 м | 31 м | 25 м | 22 м | 20 м |
| Сохранность | M | M | M | M | M | M | G | G | G | G | M | G | G | G | G | G | M | M | G | M |
| Диатомовые водоросли | ||||||||||||||||||||
| Actinoptychus intermedius A. Schmidt | R | R | R | R | R | R | F | F | F | F | R | F | F | F | R | R | ||||
| Anuloplicata concentrica (A. Schmidt) Gleser | F | F | F | F | F | F | C | C | C | C | F | C | C | F | R | F | F | F | F | R |
| Anuloplicata ornata (Grunow) Gleser | F | F | F | F | F | F | F | F | F | F | F | F | F | F | R | F | F | F | F | R |
| Aulacodiscus cellulosus Grove et Stuart | R | |||||||||||||||||||
| Asterolampra sp. | R | |||||||||||||||||||
| Arachnoidiscus ehrenbergii Ralf | R | R | R | R | R | R | R | R | ||||||||||||
| Biddulphia rigida Schmidt | F | F | R | F | F | C | R | F | F | F | R | C | ||||||||
| Biddulphia tuomeyi (Bailey) Roper | R | F | F | R | R | |||||||||||||||
| Biddulphia sp. 1 | R | |||||||||||||||||||
| Biddulphia sp. 2 | R | |||||||||||||||||||
| Bipalla oamaruensis (Grove et Stuart) Gleser | R | R | R | R | R | R | R | F | R | R | R | R | R | R | F | C | F | R | R | |
| Brightwellia coronata (Brightwell) Ralfs | R | R | ||||||||||||||||||
| Brightwellia hyperborea Grunow | R | R | R | |||||||||||||||||
| Chaetoceros spp. bristles | F | F | F | F | F | F | C | F | F | C | C | C | C | F | R | F | F | R | R | R |
| Clavicula polymorpha Grunow et Pantocsek | R | |||||||||||||||||||
| Corona retinervis Sheshukova et Gleser | R | |||||||||||||||||||
| Coscinodiscus argus Ehrenberg | F | C | C | R | F | F | C | R | R | |||||||||||
| Coscinodiscus decrescens Grunow | F | C | F | C | F | R | R | R | R | R | ||||||||||
| Coscinodiscus cf. eomonoculus Bukry | F | F | ||||||||||||||||||
| Coscinodiscus heteroporus Ehrenberg | F | |||||||||||||||||||
| “Coscinodiscus marginatus” Ehrenberg | F | F | R | F | R | F | F | F | F | F | F | F | F | F | R | R | F | R | ||
| Coscinodiscus obscurus var. concavus Gleser | R | F | R | F | R | C | C | C | C | C | F | F | F | F | R | F | F | F | F | R |
| Cosmiodiscis breviradiatus Gleser et Olshtynskaya | R | |||||||||||||||||||
| Costopyxis costata (Gleser) Gleser | R | |||||||||||||||||||
| Costopyxis reticulata (Long, Fuge et Smith) Gleser | R | F | F | |||||||||||||||||
| Craspedodiscus coscinodiscus (Ehrenberg) Ehrenberg | F | F | F | |||||||||||||||||
| Craspedodiscus oblongus (Greville) Grunow | F | F | R | R | F | F | R | F | R | R | F | R | R | R | ||||||
| Craspedophorus actinoptychoides Gleser | R | |||||||||||||||||||
| Cristodiscus duplex (Gleser) Gleser et Olshtynskaya | F | R | R | R | R | F | R | R | R | R | R | R | R | |||||||
| Cristodiscus succinctus (Sheshukova et Gleser) Gleser et Olsht. | R | R | R | R | ||||||||||||||||
| Distephanosira architecturalis (Brun) Gleser | C | C | C | C | C | A | A | A | A | C | C | C | C | C | R | F | R | R | ||
| Eunotogramma weissei Ehrenberg | R | R | ||||||||||||||||||
| Eunotogramma productum Grunow | R | F | ||||||||||||||||||
| Eupyxidicula charkoviana (Jousé) Blanco et Wetzel | R | F | F | R | R | F | F | F | F | R | F | |||||||||
| Eupyxidicula crenata (Sheshukova) Oreshkina comb. nov. | F | C | R | R | F | F | F | F | F | F | F | F | R | |||||||
| Eupyxidicula grunowii (Grove et Stuart) Blanco et Wetzel | F | F | F | C | C | C | C | C | C | C | C | F | F | R | F | F | F | F | A | |
| Eupyxidicula joynsonii (A. Schmidt) Blanco et Wetzel | F | F | ||||||||||||||||||
| Eupyxidicula megapora (Grunow) Blanco et Wetzel | R | R | R | R | ||||||||||||||||
| Eupyxidicula spinosa (Jousé) Oreshkina | R | R | ||||||||||||||||||
| Eupyxidicula turris (Greville et Arnott) Blanco et Wetzel | F | F | F | F | F | F | C | C | F | F | F | R | R | R | F | |||||
| Eupyxidicula sp. 1 | F | F | ||||||||||||||||||
| Fenneria brachiata (Brightwell) Witkowski | F | R | R | F | R | R | ||||||||||||||
| Hemiaulus polycystinorum Ehrenberg | F | F | C | C | C | C | C | A | C | A | C | C | C | A | R | F | F | F | ||
| Hemiaulus polymorphus var. charkovianus Jousé | F | F | F | F | F | R | ||||||||||||||
| Hemiaulus sp. | F | F | C | F | ||||||||||||||||
| Hyalodiscus scoticus Ehrenberg | R | R | R | |||||||||||||||||
| Ktenodiscus aculeiferus (Grunow) Blanco et Wetzel | F | R | R | R | ||||||||||||||||
| Liradiscus barbadensis Greville | R | |||||||||||||||||||
| Mastogloia splendida (Gregory) Cleve et Moller | R | |||||||||||||||||||
| Medlinia fenestrata (Witt) P.A. Sims | R | |||||||||||||||||||
| Melosira goretzkii Tscheremisinova | R | |||||||||||||||||||
| Mycetacanthus crenatus (Sheshukova) Strelnikova et Fourtanier | F | C | C | C | F | F | ||||||||||||||
| Odontotropis carinata Grunow | R | R | ||||||||||||||||||
| Paralia clavigera (Grunow) Gleser | F | R | R | |||||||||||||||||
| Paralia crenulata (Grunow) Gleser | C | C | C | F | F | F | F | F | F | C | F | C | F | F | F | F | F | |||
| Paralia fausta (A. Schmidt) Sims et Crawford | R | R | R | R | R | F | F | F | R | R | R | R | R | R | ||||||
| Paralia sulcata (Ehrenberg) Cleve | R | R | R | R | F | C | F | F | F | A | C | F | C | F | R | F | R | |||
| Pseudopodosira westii (W. Smidt) Sheshukova et Gleser | R | R | R | F | F | R | R | C | F | F | F | F | F | R | R | F | F | F | ||
| Pseudopodosira hyalina Jousé | R | F | R | F | R | F | R | R | ||||||||||||
| Pseudopodosira pileiformis Jousé | F | F | F | F | F | C | C | C | C | C | C | F | F | F | R | C | F | F | F | |
| Pseudopyxilla composita Gleser | F | |||||||||||||||||||
| Pseudostictodiscus angulatus Brun | R | R | ||||||||||||||||||
| Pseudostictodiscus kossuthii Pantocsek | R | |||||||||||||||||||
| Pyrgupyxis johnsoniana (Forti) Hendey | R | R | F | F | F | R | ||||||||||||||
| Pyxilla gracilis Tempère et Forti | R | C | F | F | F | F | F | F | F | F | F | F | F | R | R | F | F | |||
| Pyxilla sp. 1 | R | |||||||||||||||||||
| Radioplicata clavigera (Grunow) Gleser | R | F | F | F | ||||||||||||||||
| Radioplicata costulata Gleser | F | F | F | C | C | F | F | F | F | F | F | F | F | F | ||||||
| Sceletonema utriculosum Brun | R | R | ||||||||||||||||||
| Stellarima microtrias (Ehrenberg) Hasle et Sims | R | R | R | |||||||||||||||||
| Strelnikovia? sp. | R | F | R | R | ||||||||||||||||
| Sheshukovia flos (Ehrenberg) Fenner | F | |||||||||||||||||||
| Sheshukovia sp. 1 | R | |||||||||||||||||||
| Trinacria excavata Heiberg | R | R | F | F | F | F | F | F | F | F | F | F | F | F | R | R | ||||
| Trinacria ventricosa Grove et Stuart | R | R | F | F | F | F | F | F | F | R | F | F | F | F | R | F | F | |||
| Vallodiscus sp. 1 | ||||||||||||||||||||
| Vallodiscus sp. 2 | R | R | R | R | R | R | F | F | ||||||||||||
| Vallodiscus sp. 3 | R | F | ||||||||||||||||||
| Williamriedelia claviger (A. Schmidt) Blanco et Wetzel | F | F | R | R | ||||||||||||||||
| Williamriedelia spp. | C | C | C | C | C | F | A | C | A | A | A | C | C | C | ||||||
| Xanthiopyxis globosa Ehrenberg | R | |||||||||||||||||||
| Силикофлагеллаты | ||||||||||||||||||||
| Bachmannocena apiculata apiculata (Schulz) Bukry | F | R | ||||||||||||||||||
| Corbisema apiculata (Lemmermann) Hanna | F | R | ||||||||||||||||||
| Corbisema exilis Bukry | R | R | ||||||||||||||||||
| Corbisema triacantha Ehrenberg | R | R | R | R | ||||||||||||||||
| Dictyocha byronalis Bukry | R | R | R | R | ||||||||||||||||
| Corbisema hexacantha (Schulz) Deflandre | F | R | ||||||||||||||||||
| Corbisema spinosa Deflandre | F | R | ||||||||||||||||||
| Dictyocha sp. 1 | R | |||||||||||||||||||
| Distephanus crux (Ehrenberg) Haeckel | R | R | R | |||||||||||||||||
| Naviculopsis constricta (Schulz) Frenguelli | R | R | R | R | ||||||||||||||||
| Naviculopsis foliacea Deflandre | R | R | F | R | ||||||||||||||||
Таблица I . Диатомеи из разреза скв. 5/93 (Монастырщина). 1 – Eupyxidicula charkoviana (Jousé) Blanco et Wetzel, гл. 24.2 м; 2 – Mycetacanthus crenatus (Sheshukova) Strelnikova et Fourtanier, гл. 22.4 м; 3 – Eupyxidicula grunowii (Grove et Stuart) Blanco et Wetzel, гл. 22.4 м; 4 – Costopyxis reticulata (Long, Fuge et Smith) Gleser, гл. 15.3 м; 5 – Eupyxidicula spinosa (Jousé) Oreshkina comb. nov., гл. 22.4 м; 6 – Eupyxidicula turris (Greville et Arnott) Blanco et Wetzel, гл. 22.4 м; 7 – Hemiaulus sp., гл. 22.4 м; 8 – Hemiaulus polymorphus var. charkovianus Jousé, гл. 15.3 м; 9 – Corona retinervis Sheshukova et Gleser, гл. 22.4 м; 10 – Pseudostictodiscus angulatus Brun, гл. 22.4 м; 11 – Hyalodiscus scoticus Ehrenberg, гл. 22.4 м; 12 – Trinacria excavata Heiberg, гл. 22.4 м.

Таблица II. Диатомеи и силикофлагеллаты из разреза скв. 5/93 (Монастырщина). 1 – Dictyocha spinosa (Deflandre) Gleser, гл. 22.4 м; 2 – Craspedodiscus oblongus (Greville) Grunow, гл. 22.4 м; 3 – Eunotogramma weissei Ehrenberg, гл. 15.3 м; 4 – Biddulphia rigida Schmidt, гл. 22.4 м; 5 – Pseudopodosira westii (W. Smith) Sheshukova et Gleser, гл. 22.4 м; 6 – Pseudopodosira pileiformis Jousé, гл. 22.4 м; 7, 9, 10 – Distephanosira architecturalis (Brun) Gleser, гл. 22.4 м; 8 – Bipalla oamaruensis (Grove et Stuart) Gleser, гл. 15.3 м; 11 – Paralia sulcata (Ehrenberg) Cleve, гл. 15.3 м; 12 – Paralia crenulata (Grunow) Gleser, гл. 22.4 м; 13 – Anuloplicata concentrica (A. Schmidt) Gleser, гл. 22.4 м.

Таблица III. Диатомеи из разреза скв. 5/93 (Монастырщина). 1 – Actinoptychus intermedius A. Schmidt, гл. 22.4 м; 2 – Costopyxis costata (Gleser) Gleser, гл. 22.4 м; 3 – Archeomonodaceae, гл. 22.4 м; 4 – вырост Williamriedelia sp., гл. 22.4 м; 5, 6 – Xanthiopyxis globosa Ehrenberg, гл. 22.4 м; 7 – Clavicula polymorpha Grunow et Pantocsek, гл. 15.3 м; 8 – Vallodiscus sp. 1, гл. 22.4 м; 9 – Biddulphia tuomeyi (Bailey) Roper, гл. 22.4 м; 10 – щетинки Chaetoceros spp., гл. 22.4 м; 11, 13 – Strelnikovia? sp., гл. 22.4 м; 12 – Lyradiscus ellipticus Greville, гл. 22.4 м; 14 – Vallodiscus sp. 3, гл. 18.4 м; 15 – Sceletonema utriculosum Brun, гл. 28.2 м; 16 – Radioplicata costulata Gleser, гл. 56.8 м; 17 – Mastogloia splendida (Gregory) Cleve and Moller, гл. 15.3 м.
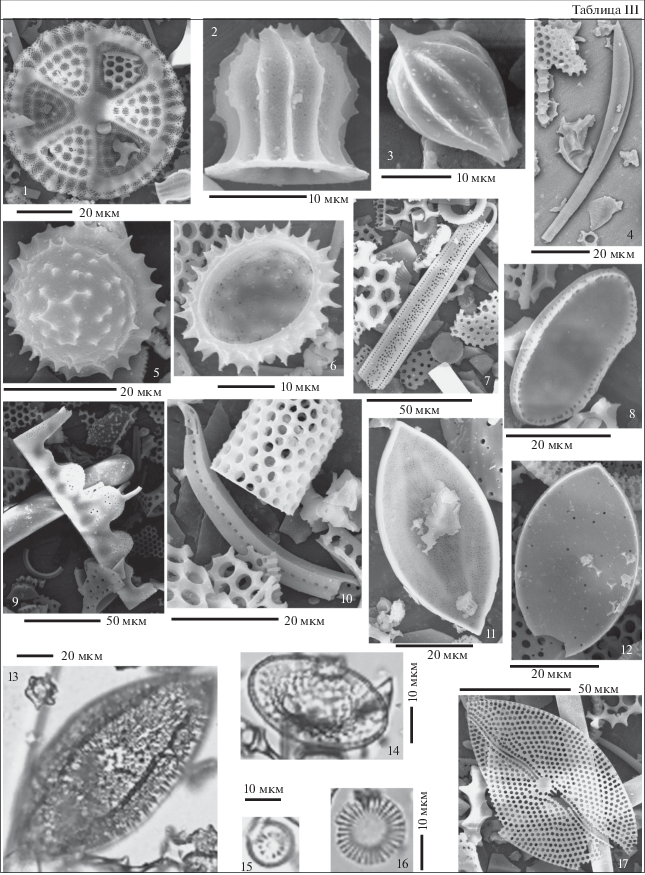
Таблица IV. Силикофоссилии разреза скв. 5/93 (Монастырщина). Длина масштабной линейки 10 мкм. 1 – Coscinodiscus obscurus A. Schmidt var. concavus Gleser, гл. 18.4 м; 2, 4 – Coscinodiscus decrescens Grunow; 2 – гл. 28.4 м, 4 – гл. 22.4 м; 3 – Cristodiscus succinctus (Sheshukova et Gleser) Gleser et Olshtinskaya, гл. 22.4 м; 5 – Brightwellia coronata (Brightwell) Ralfs, гл. 22.4 м; 6 – Coscinodiscus argus Ehrenberg, гл. 59 м; 7 – Craspedodiscus coscinodiscus Ehrenberg, гл. 22.4 м; 8 – Pseudopyxilla composita Gleser, гл. 22.4 м; 9 – Hemiaulus polycystinorum Ehrenberg, гл. 22.4 м; 10 – Cristodiscus duplex (Gleser) Gleser et Olshtinskaya, гл. 22.4 м; 11 – Trinacria ventricosa Grove et Stuart, гл. 18.4 м; 12 – Trinacria excavata Heiberg, гл. 22.4 м; 13, 14, 15 – Fenneria brachiata (Brightwell) Witkowski; 13, 14 – гл. 18.4 м; 15 – гл. 22.4 м; 16 – Anuloplicata concentrica (A. Schmidt) Gleser, гл. 35 м.
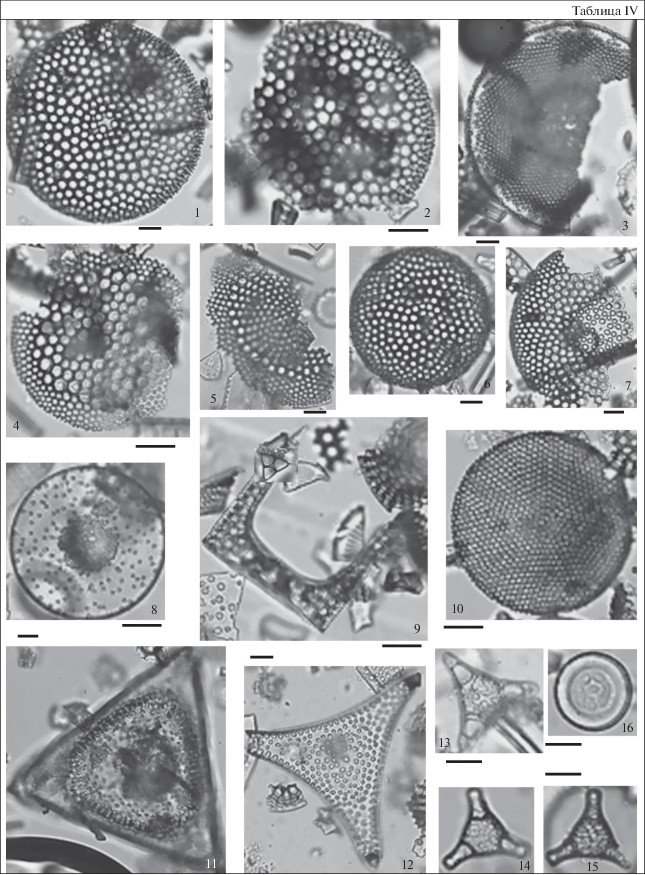
Таблица V. Диатомеи и силикофлагелляты из разреза скв. 5/93 (Монастырщина). Длина масштабной линейки 10 мкм. 1 – Pyxilla gracilis (Tempere et Forti) Hendey, гл. 22.4 м; 2 – Pyrgupyxis johnsoniana (Forti) Hendey, гл. 16.4 м; 3, 4 – Williamriedelia claviger (A. Schmidt) Blanco et Wetzel, 3 – гл. 18.4 м, 4 – гл. 22.4 м; 5 – Ktenodiscus aculeiferus (Grunow) Blanco et Wetzel, гл. 56.8 м; 6 – Distephanus crux (Ehrenberg) Haeckel, гл. 22.4 м; 7, 10 – Corbisema triacantha Ehrenberg, 7 – гл. 35 м, 10 – гл. 59 м; 8 – Odontotropis carinata Grunow, гл. 22.4 м; 9 – Corbisema exilis Bukry, гл. 22.4 м; 11 – Naviculopsis constricta (Schulz) Frenguelli, гл. 59 м; 12, 17 – Dictyocha byronalis Bukry, 12 – гл. 18.4 м, 17 – гл. 28.2 м; 13 – Corbisema hexacantha (Schulz) Deflandre, гл. 22.4 м; 14, 15 – Corbisema spinosa Deflandre, 14 – гл. 22.4 м, 15 – гл. 59 м; 16 – Dictyocha sp., гл. 18.4 м; 18 – Bachmannocena apiculata apiculata (Schulz) Bukry, гл. 28.2 м.
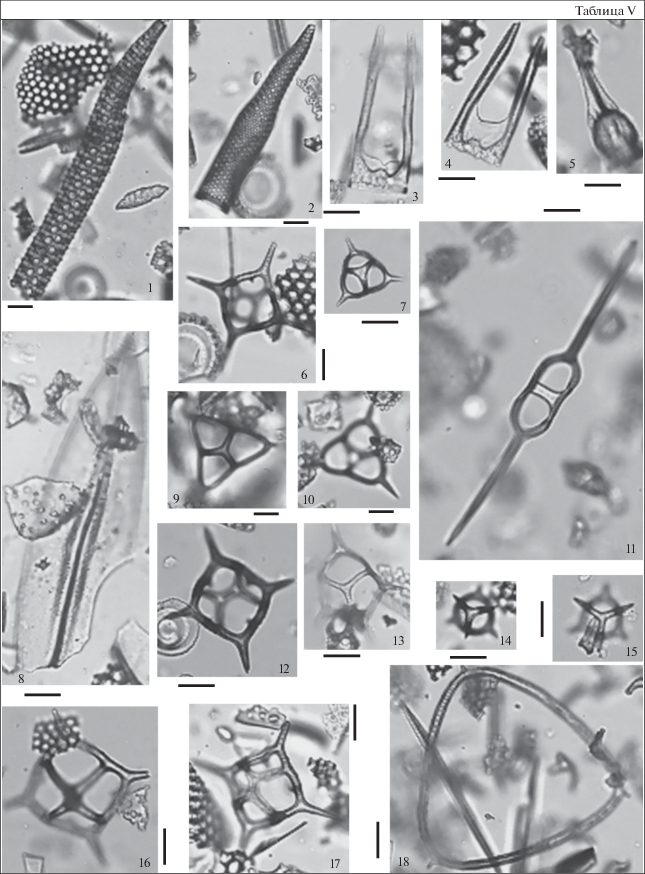
Таблица VI. Диатомеи и силикофлагеллаты из разреза скв. 16 (Богучар). Длина масштабной линейки 10 мкм. 1 – Eopyxidicula sp. 1, гл. 45 м; 2 – Hemiaulus polymorphus var. charkovianus Jousé, гл. 45 м; 3 – Melosira goretzkii Tscheremisinova, гл. 22 м; 4 – Aulacodiscus cellulosus Grove et Stuart, гл. 25 м; 5 – Liradiscus barbadensis Greville, гл. 22 м; 6 – Pseudopodosira hyalina Jousé, гл. 25 м; 7 – Coscinodiscus cf. eomonoculus Bukry, гл. 31 м; 8 – Naviculopsis foliacea Deflandre, гл. 25 м; 9 – Sheshukovia flos (Ehrenberg) Fenner, гл. 45 м; 10 – Sheshukovia sp. 1, гл. 25 м; 11 – Vallodiscus sp. 3, гл. 25 м; 12 – Corbisema apiculata (Lemmermann) Hanna, гл. 31 м; 13 – Eopyxidicula joynsonii (A. Schmidt) Blanco et Wetzel, гл. 20 м; 14 – Biddulphia sp. 1, гл. 25 м; 15 – Bachmannocena apiculata apiculata (Schulz) Bukry, гл. 45 м.

В группе таксонов широкого стратиграфического диапазона преобладают представители родов Pseudopodosira, Paralia, Anuloplicata, Radioplicata, Hyalodiscus, Actinoptychus, характерные для шельфовой области; менее многочисленны Arachnoidiscus, Pseudostictodiscus, Trochosira, Pseudopyxilla, Vallodiscus, Ktenodiscus. Особо следует отметить низкое содержание Paralia sulcata (Ehrenberg) Cleve – обычно массового вида для палеоцена и нижнего эоцена Поволжья и Зауралья. Характерно высокое содержание фрагментов створок диатомей, как правило считающихся выростами Williamriedelia и щетинками Chaetoceros. Постоянными компонентами комплекса являются группа Coscinodiscus (C. argus Ehrenberg, C. decrescens Grunow, C. obscurus var. concavus Gleser), Eupyxidicula (E. turris (Greville et Arnott) Blanco et Wetzel, E. crenata (Sheshukova) Oreshkina, E. grunowii (Grove et Stuart) Blanco et Wetzel, E. megapora (Grunow) Blanco et Wetzel), Trinacria (T. excavata Heiberg, T. ventricosa Grove et Stuart). Также встречены Biddulphia regida Schmidt, Odontotropis carinata Grunow, Stellarima microtrias (Ehrenberg) Hasle et Sims.
Группа стратиграфически значимых таксонов диатомей включает моноспецифичные Bipalla oamaruensis (Grove et Stuart) Gleser и Distephanosira architecturalis (Brun) Gleser, а также Pyxilla gracilis Tempère et Forti, Craspedodiscus oblongus (Greville) Grunow, Fenneria brachiata (Brightwell) Witkowski, Cristodiscus duplex (Gleser) Gleser et Olshtinskaya, Hemiaulus polycystinorum Ehrenberg. Отмечается присутствие силикофлагеллаты Corbisema spinosa Deflandre. В верхней части тишкинской свиты встречен вид Cristodiscus succinctus (Sheshukova et Gleser) Gleser et Olshtinskaya. В целом комплекс типичен для прибрежной зоны морского бассейна.
В касьяновской свите комплекс диатомей пополняется видами Brightwellia coronata (Brightwell) Ralfs, Mycethacanthus crenatus (Sheshukova) Strelnikova et Fourtanier, выше – Pyrgupyxis johnsoniana (Forti) Hendey, Eopyxidicula charkoviana (Jousé) Blanco et Wetzel, Williamriedelia claviger (A. Schmidt) Blanco et Wetzel. Среди силикофлагеллат отмечено появление Corbisema hexacantha Deflandre.
В верхах касьяновской свиты (гл. 11 м) силикофоссилии представлены в основном обломками спикул губок и редкими створками диатомей плохой сохранности, что говорит о начале регрессивного этапа осадконакопления.
Присутствие в тишкинской и касьяновской свитах стратиграфически важных таксонов диатомей Bipalla oamaruensis, Coscinodiscus obscurus var. сoncavus, Distephanosira architecturalis, Cristodiscus succinctus, C. duplex, Hemiaulus polycystinorum Ehrenberg, Fenneria brachiata, Eopyxidicula сharkoviana, Williamriedelia сlaviger позволяет отнести эти свиты к зоне Bipalla oamaruensis зональной шкалы З.И. Глезер (1979). Реперное событие – появление силикофлагеллаты Corbisema hexacantha в средней части касьяновской свиты, что дает возможность установить одноименную зону.
Скв. 16 (Богучар). Палинологический комплекс, выявленный из подошвы тишкинской свиты (гл. 54–55 м), характеризуется абсолютным доминированием цист динофлагеллат (89%); акритархи составляют лишь 8% от общего состава, а споры и пыльца наземных растений представлены единично. В комплексе диноцист определено 48 видов. Очевидных доминантов в ассоциации цист динофлагеллат не наблюдается, в наибольших количествах отмечаются Deflandrea phosphoritica (17%), группа Spiniferites (16%), Corrudinium incompositum (7%), Microdinium reticulatum (7%) и Thalassiphora pelagica (4.5%). Подобные соотношения диноцистовых экогрупп могут свидетельствовать об относительно мелководных обстановках осадконакопления.
В составе комплекса диноцист встречены виды Achilleodinium biformoides (Eisenack) Eaton, Achomosphaera ramulifera (Deflandre) Evitt, Areosphaeridium diktyoplokum (Klump) Eaton, Areosphaeridium michoudii Bujak, Cerebrocysta bartonensis Bujak, C. magna Bujak, Cleistosphaeridium diversispinosum Davey et al., Cordosphaeridium funiculatum Morgenroth, Corrudinium incompositum (Drugg) Stover et Evitt, Costacysta bucina Heilmann-Clausen et Van Simayes, Cribroperdinium giuseppei (Morgenroth) Helenes, Cribroperidinium tenuitabulatum (Gerlach) Helenes, Dapsilidinium pseudocolligerum (Stover) Bujak et al., Deflandrea phosphoritica, Elytrocysta brevis Stover et Hardenbol, Enneadocysta arcuata (Eaton) Stover et Williams, E. pectiniformis (Gerlach) Stover et Williams, Heteraulacacysta campanula Drugg et Loeblich Jr., H. porosa Bujak, Homotryblium tenuispinosum, Hystrichokolpoma cinctum Klumpp, H. aff. granulatum Eaton, Hystrichostrogylon membraniphorum Agelopoulos, Impagidinium dispertitum (Cookson et Eisenack) Stover et Evitt, I. paradoxum (Wall) Stover et Evitt), I. victorianum (Cookson et Eisenack) Stover et Evitt, Lingulodinium machaerophorum (Deflandre et Cookson) Wall, Melitasphaeridium asterium (Eaton) Bujak et al., M. pseudorecurvatum (Morgenroth) Bujak et al., Membranophoridium aspinatum Gerlach, Membranosphaera sp. B sensu CHC (1985), Microdinium reticulatum Vozzhennikova, Operculodinium centrocarpum (Deflandre et Cookson) Wall, O. eisenackii Heilmann-Clausen et Van Simayes, O. microtriainum (Klumpp) Islam, Pentadinium goniferum Edwards, P. laticinctum Gerlach, P. lophophorum (Benedek) Benedek et al., Phthanoperidinium distinctum Bujak, Ph. geminatum Bujak, Ph. stockmansii (De Coninck) Lentin et Williams, Pterodinium cingulatum (Wetzel) Below, Rottnestia borussica (Eisenack) Cookson et Eisenack, Samlandia chlamydophora Eisenack, Spiniferella cornuta (Gerlach) Stover et Hardenbol, Spiniferites pseudofurcatus (Klumpp) Sarjeant, S. ramosus (Ehrenberg) Mantell, Svalbardella clausenii Sliwinska et Head, Tectatodinium pellitum Wall, Wetzeliella articulata Eisenack. Поскольку цисты динофлагеллат выявлены лишь в одном образце из разреза, достаточно сложно четко определить стратиграфический возраст отложений. Присутствие видов Enneadocysta pectiniformis, Heteraulacacysta porosa и Costacysta bucina указывает на возраст не древнее позднего лютета, не ранее интервала диноцистовой зоны Enneadocysta pectiniformis Перитетиса (~44 млн лет и меньше; Яковлева, 2017).
Силикофоссилии, выявленные в верхней части тишкинской свиты и в касьяновской свите, в целом характеризуются преобладанием спикул губок, фрагментов створок диатомовых. В тишкинской свите неритические диатомеи широкого стратиграфического диапазона представлены Pseudopodosira, Anuloplicata, Radioplicata, Paralia sulcatа, отмечены крупные створки Eopyxidicula sp. 1, Biddulphia sp. 2 (рис. 4, табл. 2, табл. I–VI ). Среди стратиграфически важных видов – Bipalla oamaruensis (Grove et Stuart) Gleser, Distephanosira architecturalis (Brun) Gleser, Eopyxidicula сharkoviana (Jousé) Blanco et Wetzel, Coscinodiscus obscurus var. concavus Gleser, Hemiaulus polycystinorum Ehrenberg.
В касьяновской свите комплекс диатомей обогащается за счет появления новых элементов – Cristodiscus succinctus (Sheshukova et Gleser) Gleser et Olshtinskaya, C. duplex (Gleser) Gleser et Olshtinskaya. Выше по разрезу встречены Cristodiscus breviradiatus Gleser et Olshtinskaya, Coscinodiscus heteroporus Ehrenberg, Brightwellia hyperborea Grunow, Eopyxidicula joynsonii (A. Schmidt) Blanco et Wetzel, Hemiaulus tschestnovii Pantochek, H. aff. immanis (Boyer) Hendey et Sims, Asterolampra sp. Спорадически присутствует Coscinodiscus? aff. eomonoculus, описанный Bukry (1987) из интервала наннопланктонной зоны CP12b в скважине 613, пробуренной в рейсе 95 DSDP в Атлантике у берегов штата Нью-Джерси. Рассмотренный комплекс характеризует диатомовую зону Bipalla oamaruensis зональной шкалы З.И. Глезер (1979). Реперным уровнем является появление силикофлагеллаты Corbisema hexacantha.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате проведенного микропалеонтологического анализа палеогеновых отложений скв. 5/93 (Монастырщина) и скв. 16 (Богучар) получены новые данные для уточнения стратиграфического диапазона региональных подразделений эоцена, выделяемых в рассматриваемых структурно-фациальных зонах.
Согласно нашим данным, полученным при изучении палеогеновых отложений Преддонецкой СФЗ (скв. 1238 в Ростовской области; Яковлева, Александрова, 2021), нижняя часть вёшенской свиты соответствует верхнему танету–инициальному ипру (интервал диноцистовых зон Apectodinium hyperacanthum–Axiodinium augustum). С учетом новых данных изучения диноцист в скв. 5/93 (Монастырщина) установлено, что верхняя часть вёшенской свиты включает зону Stenodinium meckelfeldense. Таким образом, общий объем указанной свиты в регионе должен быть расширен и соответствовать верхнему танету–части нижнего ипра. Следует отметить, что в статье (Khokhlova et al., 1999) интервал разреза, рассматриваемый в настоящей работе в качестве вёшенской свиты (гл. 64.3–60.1 м), был отнесен к осиновским слоям; более того, в нем были обнаружены диатомеи и радиолярии среднеэоценового возраста. В ходе проведенных нами исследований в этом интервале в скв. 5/93 не было найдено диатомовых водорослей, а диноцисты четко указали на ипрской возраст пород. Скорее всего, это расхождение в данных связано с технической ошибкой в процессе лабораторной обработки проб при предыдущих исследованиях. Исходя из полученных нами данных, вёшенская свита накапливалась в прибрежных морских обстановках, близких к нормально-соленым.
Из-за редкого отбора проб в скв. 16 (Богучар) данные по палиноморфам и силикофоссилиям носят фрагментарный характер. В частности, не удалось получить данные по палиноморфам бучакской и сергеевской свит. Однако такие данные имеются для скв. 230 Северной Украины (Орешкина, Яковлева, 2007; Iakovleva, 2015; Яковлева, 2017), в которой внутри интервала бучакской и киевской свит выявлены интервалы диноцистовых зон Costacysta bucina, Enneadocysta pectiniformis и Rhombodinium draco лютетского–раннебартонского возраста. Также получены данные (Iakovleva, 2019) по диноцистам из стратотипа бучакской свиты в песчаном карьере Костянец (окрестности г. Канев, Украина): здесь выявлен комплекс диноцистовой зоны Costacysta bucina среднего лютета, что подтверждает общее мнение украинских (Zosimovich, Shevchenko, 2014) и российских стратиграфов о лютетском возрасте бучакской свиты.
В Павловско-Кантемировской СФЗ, помимо разреза у г. Кантемировка, имеются данные по карбонатным группам для скв. 1 (Павловск), где карбонатные глины сергеевской свиты были отнесены к зоне NP16–нижней части зоны NP17 по наннопланктону и слоям с Acarinina rotundimarginata и Acarinina kiewensis по планктонным фораминиферам (Практическое…, 2005; Бугрова и др., 2016). Таким образом, сергеевская свита в Павловско-Кантемировской СФЗ соответствует части верхнего лютета–инициальному бартону ОСШ.
Согласно полученным нами данным, в разрезах скв. 16 и скв. 5/93 тишкинская и касьяновская свиты охарактеризованы диатомеями и силикофлагеллатами зоны Bipalla oamaruensis. При этом в разрезах Павловско-Кантемировской СФЗ (скв. 16 и разрез Кантемировка (Бугрова и др., 2016)) ей отвечает верхняя часть тишкинской свиты, тогда как в Преддонецкой СФЗ (скв. 5/93 (Монастырщина)) этой зоной охарактеризована вся тишкинская свита. В обеих СФЗ внутри касьяновской свиты фиксируется уровень появления силикофлагеллаты Corbisema hexacantha.
В опорном разрезе Кантемировка, содержащем диатомовые комплексы с менее информативным, обедненным составом, верхняя часть тишкинской свиты соответствует нижней части диатомовой зоны Bipalla oamaruensis и радиоляриевым зонам Cyrtophormis alta и Ethmosphaera polysiphonia (Khokhlova et al., 1999; Бугрова и др., 2016). Последние датируются поздним лютетом–бартоном (Козлова, 1999), что согласуется с отнесением свиты к наннопланктонной зоне ?СNE14 шкалы Agnini et al. (2014) (Мусатов, 2020). Однако данные по фораминиферам свидетельствуют уже о бартонском возрасте базальных горизонтов тишкинской свиты (Бугрова и др., 2016).
В изученном нами в Павловско-Кантемировской СФЗ разрезе скв. 16 (Богучар) в основании тишкинской свиты установлен таксономически разнообразный комплекс диноцист, который по присутствию стратиграфически значимых видов Eneadocysta pectiniformis, Heteraulacacysta porosa и Costacysta bucina условно можно отнести к диноцистовой зоне Enneadocysta pectiniformis среднего эоцена Перитетиса (не древнее ~44 млн лет; Яковлева, 2017). Следует отметить, что на крайнем востоке Преддонецкой СФЗ на стыке с Поволжско-Прикаспийским субрегионом в скв. 13 (Котельниково) первое присутствие Eneadocysta pectiniformis зафиксировано в верхней части тишкинской свиты (Александрова и др., 2011), тогда как на севере Украины это событие выявлено стратиграфически ниже – в верхней части бучакской свиты (Iakovleva, 2015; Яковлева, 2017), а в разрезе Актумсук на плато Устюрт – на уровне зоны СР13b (Яковлева и др., 2019). При этом совместная встречаемость в разрезе Актумсук данного таксона, а также Heteraulacacysta porosa, Costacysta bucina, присутствующих в основании тишкинской свиты в скв. 16, приурочена к бартону (терминальная часть зоны СР14а), а в скв. 13 (Котельниково) они имеют разорванный диапазон распространения. На наш взгляд, такие расхождения обусловлены особенностями палеообстановок в различных частях Перитетиса.
На смежной территории Преддонецкой СФЗ, по нашим данным (скв. 5/93 (Монастырщина)), зафиксирован перерыв между вёшенской и тишкинской свитами, объем которого, вероятно, охватывает большую часть ипра и лютета. Согласно предыдущим исследованиям скв. 5/93 (Khokhlova et al., 1999) (рис. 6), в средней части тишкинской свиты установлен бедный комплекс наннопланктона, который может быть отнесен к верхней части зоны CP13 или к зоне CP14а (Okada, Buckry, 1980). Особенностью разреза является отсутствие радиолярий в тишкинской свите. Их присутствие отмечено только начиная с подошвы касьяновской свиты (Khokhlova et al., 1999). Скорее всего, это обусловлено неблагоприятными для их захоронения палеообстановками, с широким развитием нехарактерных для тишкинской свиты песчаных фаций. Последнее, по мнению Горбаткиной и Иосифовой (2004), присуще участкам размыва бучакских и каневских отложений.
Рис. 6.
Корреляция зональных подразделений по разным группам микрофоссилий (показано цветом) в изученных разрезах скв. 5/93 (Монастырщина), скв. 16 (Богучар), в опорном разрезе Кантемировка с региональными зональными шкалами и МСШ по версии Agnini et al. (2014) и предполагаемый возраст региональных литостратонов. Пунктиром показан уровень появления силикофлагеллаты Corbisema heхасаntha. Использованы данные: Khokhlova et al., 1999 (1); Козлова, 1999 (2); Бугрова и др., 2016 (3); Мусатов, 2020 (4). Условные обозначения см. на рис. 3. Сокращения: зоны по диноцистам: Rh. – Rhombodinium, Och.rom./S.chl. – Ochetodinium romanum/Samlandia chlamydophora, E. – Eatonicysta, Ch. – Charlesdownea, Dr. – Dracodinium, D. – Deflandrea, A. – Axioidinium; зоны по радиоляриям: T – Theocyrtis, C – Cyrtophormis; зоны по диатомеям: M. – Moisseevia, H. – Hemiaulus; зоны по фораминиферам: A. – Acarinina; F – планктонные фораминиферы, N – наннопланктон, R – радиолярии, Dt – диатомовые водоросли, Dn – диноцисты; Олиг. – олигоцен, Кан.– кантемировская свита; Пол. – полтавская свита.
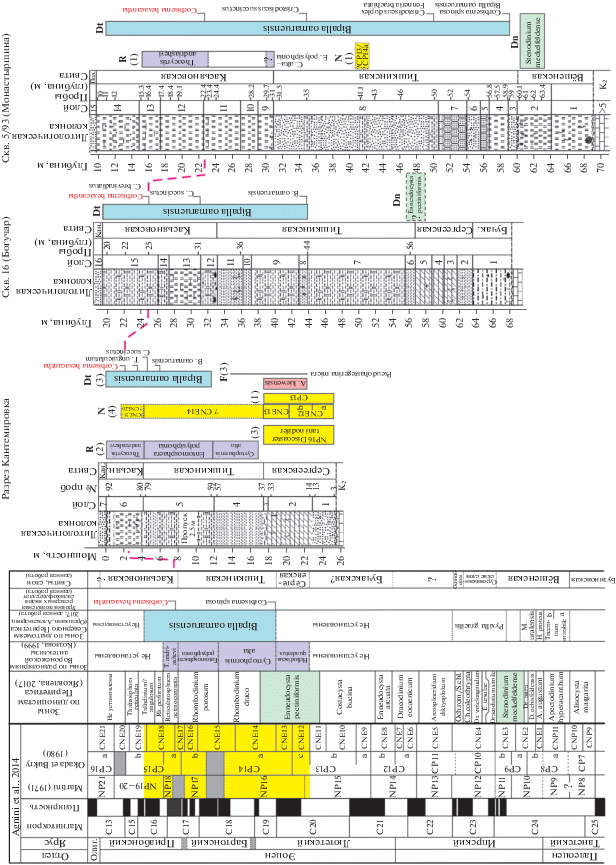
Таким образом, по имеющимся и полученным нами данным, стратиграфический диапазон тишкинской свиты, относящейся к нижней части зоны Bipalla oamaruensis, может быть ограничен терминальной частью лютета–бартоном.
Касьяновская свита относится к верхней части зоны Bipalla oamaruensis с уровнем появления Corbisema hexacantha, в Павловско-Кантемировской и Преддонецкой структурно-фациальных зонах она сопоставляется с радиоляриевой зоной Theocyrtis andriashevi приабона, установленной в верхнеэоценовой белоглинской свите (Козлова, 1999). Обедненный комплекс нерасчлененных зон по радиоляриям Cyrtophormis alta–Ethmosphaera polysiphonia, установленный (Khokhlova et al., 1999) в подошве касьяновской свиты в Преддонецкой СФЗ (скв. 5/93), на наш взгляд, является переотложенным, что подтверждается характером вмещающих пород с признаками перемыва. Приабонский возраст касьяновской (=обуховской) свиты палинологически обоснован и в разрезе Пасеково Павловско-Кантемировской СФЗ (Шпуль, 2005).
Хотя диатомовые ассоциации из тишкинской и касьяновской свит близки по составу, в последней отмечено присутствие новых элементов – Cosmiodiscus breviradiatus, Triceratium unguiculatum, а также силикофлагеллаты Corbisema hexacantha. Стратиграфический диапазон последнего вида, по имеющимся данным для донных осадков океана, оценивается как интервал NP15 (верхи)–NP19/20 (Locker, 1996). Учитывая, что касьяновской свите соответствует радиоляриевая зона Theocyrtis andriashevi, появление вида C. hexacantha в рассматриваемом палеобассейне происходило значительно позже, чем в океанических осадках.
Приабонский возраст верхней части зоны Bipalla oamaruensis подтверждается комплексным изучением обуховской свиты, фациального аналога касьяновской свиты в украинских разрезах (Зосимович и др., 2009), а также данными по разрезу скв. 57 Прикаспия, где верхняя часть зоны Bipalla oamaruensis установлена в интервале диноцистовых зон Reticulatosphaera actinocoronata и Rhombodinium perforatum приабона (Орешкина и др., 2015). На смежной территории Северной Украины в скв. 230 (Стрелечье, Харьковская область), по нашему мнению, представлена только самая верхняя часть зоны B. oamaruensis, охарактеризованная комплексом радиолярий зоны Helterosestrum shabalkini (Радионова и др., 1994) и диноцистовой зоной Talladinium? angulosum приабонского возраста (Яковлева, 2017).
Таким образом, общий стратиграфический диапазон зоны Bipalla oamaruensis в Павловско-Кантемировской и Преддонецкой структурно-фациальных зонах соответствует терминальному лютету–раннему приабону. Такой широкий стратиграфический интервал зоны и невозможность на текущий момент времени уверенного выделения более дробных подразделений внутри зоны Bipalla oamaruensis связаны с нестабильными обстановками в краевых частях эпиконтинентальных бассейнов, где реперные виды диатомей появляются на разных стратиграфических уровнях в зависимости от фациальных обстановок. Экологическая структура диатомовых ассоциаций из тишкинской и касьяновской свит с преобладанием неритических видов характеризует краевые участки палеобассейна. Присутствие практически по всему разрезу эоцена обломков створок диатомей и спикул губок говорит об активном гидрологическом режиме в регионе в течение среднего эоцена и части позднего эоцена. Спецификой рассматриваемого комплекса также является доминирование вида Distephanosira architecturalis, видов рода Pseudopodosira и относительно низкое содержание вида Paralia sulcata. Стабильное присутствие силикофлагеллат отражает условия нормальной морской солености.
Тщательное изучение разрезов скважин показало, что в касьяновской свите отмечается присутствие новых элементов – Cosmiodiscus breviradiatus, Triceratium unguiculatum. Также в качестве относительно стабильного реперного уровня можно рассматривать появление силикофлагеллаты Corbisema hexacantha в касьяновской свите на уровне радиоляриевой зоны Theocyrtis andriashevi, что позволяет фиксировать уровень нижней границы приабона.
Таким образом, исходя из всех имеющихся на данный момент микропалеонтологических данных по региону, можно предположить, что сергеевская свита соответствует верхнему лютету, тишкинская – терминальному лютету–бартону, а касьяновская – низам приабона.
Полученные микропалеонтологические данные позволили выявить различия в строении палеогеновых толщ в Павловско-Кантемировской и Преддонецкой структурно-фациальных зонах (рис. 6). Разрезы Павловско-Кантемировской СФЗ (Кантемировка, скв. 1 (Павловск) и скв. 16 (Богучар)) характеризуются четкой последовательностью литостратонов лютета–бартона–приабона с присутствием карбонатных фаций (сергеевская свита). В Преддонецкой СФЗ происходит наращивание стратиграфического объема палеогена (бузиновская и вёшенская свиты верхнего танета–нижнего ипра), значительное возрастание мощностей тишкинской и касьяновской свит за счет увеличения терригенной составляющей, отсутствуют позднеипрские–большая часть лютетских пород. Возможно, для Преддонецкой СФЗ следует вернуться к предложению Горбаткиной и Иосифовой (2004) о выделении здесь самостоятельного подразделения, соответствующего объему сергеевской и тишкинской свит в соседней Павловско-Кантемировской СФЗ.
ВЫВОДЫ
Впервые в Преддонецкой СФЗ выявлена диноцистовая зона Stenodinium meckelfeldense, что позволило уточнить стратиграфический объем вёшенской свиты, расширив его от танета до нижнего ипра. В основании тишкинской свиты разреза скв. 16 (Богучар) определен таксономически разнообразный комплекс диноцист с Enneadocysta pectiniformis, Heteraulacacysta porosa и Costacysta bucina, что позволяет с некоторой долей условности отнести основание этой свиты к зоне Enneadocysta pectiniformis верхнего лютета.
В тишкинской и касьяновской свитах обеих СФЗ установлена диатомовая зона Bipalla oamaruensis, охватывающая интервал от терминального лютета до раннего приабона. Этот возрастной диапазон примерно соответствует объему этой зоны в региональной схеме (Ахметьев, Беньямовский, 2003). Более дробное деление этого интервала с привязкой к Международной шкале требует дальнейшего подтверждения по другим группам планктона, которые в настоящее время не выявлены.
Выделенные комплексы силикофоссилий и палиноморф характеризуют краевые участки палеобассейна с активным гидрологическим режимом. Присутствие силикофлагеллат и состав комплексов цист динофлагеллат указывают на нормальную соленость.
Проведенная биостратиграфическая корреляция разрезов наглядно демонстрирует сложную ситуацию в решении вопроса соотношения карбонатных, кремневых и органикостенной групп микрофоссилий и различия в строении палеогеновых толщ. Актуальной задачей по-прежнему остаются поиск и комплексное изучение более насыщенных микропалеонтологическими остатками разрезов палеогена в южной части Русской плиты.
Благодарности. Авторы признательны Э.М. Бугровой за образцы керна скв. 16 (Богучар), Э.П. Радионовой за материалы по скв. 5/93 (Монастырщина), С.В. Гришину за помощь в лабораторной обработке проб, Н.В. Горьковой за съемку в сканирующем электронном микроскопе. Важные для нас рекомендации и замечания Е.Б. Пещевицкой и С.В. Наугольных дали возможность значительно улучшить содержание статьи.
Источники финансирования. Исследования проведены в соответствии с планами научно-исследовательской работы ГИН РАН и гранта РФФИ № 18-05-00505.
Список литературы
Александрова Г.Н., Беньямовский В.Н., Запорожец Н.И., Застрожнов А.С., Застрожнов С.И., Табачникова И.П., Орешкина Т.В., Закревская Е.Ю. Палеоген юго-запада Волгоградской области (скв. 13, Гремячинская площадь). Статья 1. Биостратиграфия // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 3. С. 70–95. https://doi.org/10.1134/S0869593811030014
Александрова Г.Н., Орешкина Т.В., Яковлева А.И., Радионова Э.П. Диатомеи и диноцисты в интервале позднего палеоцена–раннего эоцена в биокремнистых фациях Среднего Зауралья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 4. С. 68–94.
Александрова Г.Н., Орешкина Т.В., Яковлева А.И. Актуализация региональной стратиграфической схемы палеогена юга Русской плиты: проблемы и пути решения // Проблемы региональной геологии Северной Евразии. Материалы конференции “XII Научные чтения, посвященные памяти проф. М.В. Муратова”. М.: МГРИ-РГГРУ, 2020. С. 3–5.
Ахметьев М.А., Беньямовский В.Н. Стратиграфическая схема морского палеогена юга Европейской России // Бюлл. МОИП. Отд. геол. 2003. Вып. 5. С. 40–51.
Бугрова Э.М., Беньямовский В.Н., Табачникова И.П., Рябоконь Т.С., Радионова Э.П. Опорный разрез Кантемировка палеогена Воронежского региона // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 6. С. 78–100.
Глезер З.И. Диатомовые водоросли палеогена // Диатомовые водоросли СССР. Ископаемые и современные. Л.: Наука, 1974. Т. 1. С. 109–146.
Глезер З.И. Зональное расчленение палеогеновых отложений по диатомовым водорослям // Сов. геол. 1979. № 11. С. 19–30.
Глезер З.И., Шешукова-Порецкая В.С. Позднеэоценовые диатомовые, золотистые и перидиниевые водоросли и эбриидеи северо-восточной окраины Украинского кристаллического массива // Палеонтол. сборник. 1968. Вып. 1. № 5. С. 117–123.
Глезер З.И., Шешукова-Порецкая В.С. К истории формирования позднеэоценовой морской диатомовой флоры Украины // Вестник Ленинградского ун-та. 1969. № 9. С. 60–73.
Глезер З.И., Зосимович В.Ю., Клюшников М.Н. Диатомовые водоросли палеогеновых отложений бассейна Северный Донец и их стратиграфическое положение // Палеонтол. сборник. 1965. Вып. 2. № 2. С. 73–83.
Горбаткина Т.Е., Иосифова Ю.И. Новые стратиграфические подразделения палеогена Воронежской антеклизы – воробьевская и пасековская свиты // Вестник Воронежского ун-та. Геология. 2004. № 2. С. 28–44.
Жузе А.П. Диатомовые третичных отложений // Диатомовый анализ. М.–Л.: Госгеолиздат, 1949. Т. 1. С. 114–144.
Заклинская Е.Д. Описание пыльцы и спор некоторых видов растений полярной тундры. Описание некоторых видов пыльцы и спор, выделенных из третичных отложений Пасековского карьера Воронежской области // Труды ГИН АН СССР. Сер. геол. 1953. Вып. 142. № 59. 119 с.
Запорожец Н.И. Палинологические комплексы кумского горизонта среднего эоцена Кавказа и его возрастных аналогов в сопредельных регионах // Стратиграфия. Геол. корреляция. 2001. Т. 9. № 6. С. 83‒103.
Зональная стратиграфия фанерозоя России. Ред. Корень Т.Н. СПб.: Изд-во ВСЕГЕИ, 2006. 256 с.
Зосимович В.Ю., Ольштинська О.П., Рябоконь Т.С. и др. Особливості складу і датування середньо-верхньоеоценових відкладів зони зчленування Дніпровсько-Донецької западини та окраїн Донбасу // Ископаемая фауна и флора Украины: палеоэкологический и стратиграфический аспекты. Киев: ИГН НАН Украины, 2009. С. 262–276.
Козлова Г.Э. Практическое руководство по микрофауне России. Том 9. Радиолярии палеогена бореальной области России. СПб.: ФГУП ВНИГРИ, 1999. 312 с.
Крашенинников В.А., Ахметьев М.А. Геологические и биотические события позднего эоцена–раннего олигоцена на территории бывшего СССР. Часть I. М.: ГЕОС, 1996. 314 с.
Курлаев В.И., Ахлестина Е.Ф. Палеоген Среднего и Нижнего Поволжья. Деп. в ВИНИТИ, № 885б В88. 1988. 204 с.
Леонов Г.П. Основные вопросы региональной стратиграфии палеогеновых отложений Русской плиты. М.: Изд-во МГУ, 1961. 552 с.
Мусатов В.А. Лютет или бартон? Возраст сергеевской свиты по наннопланктону в опорном разрезе “Кантемировка” Воронежской антеклизы и корреляция с сопредельными регионами // Недра Поволжья и Прикаспия. 2020. Вып. 98. С. 4–26.
Николаева И.А., Бугрова Э.М., Глезер З.И., Табачникова И.П., Александрова Г.Н., Яковлева А.И., Закревская Е.Ю. Палеогеновая система // Зональная стратиграфия фанерозоя России. СПб.: Изд-во ВСЕГЕИ, 2006. С. 172-192.
Ольштынская А.П. Диатомовые водоросли стратотипического разреза тишкинской свиты // Геол. журн. 1976. Т. 36. Вып. 4. С. 148–151.
Ольштынская А.П. Позднеэоценовые диатомовые водоросли и силикофлагеллаты северо-востока Украины // Геол. журн. 1977. Т. 37. Вып. 3. С. 46–52.
Ольштынская А.П. Диатомовые водоросли верхнего эоцена северо-восточной Украины и их стратиграфическое значение. Автореф. дисс. … канд. геол.-мин. наук. Киев, 1978а. 21 с.
Ольштынская А.П. Новые диатомовые водоросли позднего эоцена Украины // Палеонтол. сборник. 1978б. № 15. С. 75–79.
Ольштинська О.П. Сучасний стан вивченості кайнозойських діатомових водоростей в Україні // Біостратиграфічні основи побудови стратиграфічних схем фанерозою України. Kyiv: ІГН НАН України, 2008. С. 351–359.
Орешкина Т.В., Александрова Г.Н. Палеонтологическая характеристика палеоцена–нижнего эоцена Ульяновско-Сызранской структурно-фациальной зоны Поволжско-Прикаспийского субрегиона // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 3. С. 73–98.
Орешкина Т.В., Яковлева А.И. Новые данные по палеонтологической характеристике отложений киевской и обуховской свит северного борта Днепровско-Донецкой впадины (скв. 230, пос. Стрелечье) // Палентологiчнi дослiдження в Украiнi. Вiдп. ред. Гожик П.Ф. Kyiv: Нора-прiнт, 2007. С. 233–237.
Орешкина Т.В., Яковлева А.И., Щербинина Е.А. Комплексный микропалеонтологический анализ эоценовых отложений восточного Прикаспия (скв. 57, шубарсайская мульда, Казахстан) // Бюлл. МОИП. Отд. геол. 2015. Т. 90. Вып. 1. С. 42–80.
Палатная А.П. Новые данные о диатомовых водорослях стратотипического разреза касьяновской свиты // Геол. журн. 1976. Т. 36. Вып. 2. С. 144–147.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 32. СПб.: Изд-во ВСЕГЕИ, 2001. С. 18–20.
Практическое руководство по микрофауне. Т. 8. Фораминиферы кайнозоя. Ред. Бугрова Э.М. СПб.: Изд-во ВСЕГЕИ, 2005. 324 с.
Радионова Э.П., Орешкина Т.В. Проблема границы среднего–верхнего эоцена в разрезах северо-восточного борта Днепровско-Донецкой впадины. Проблемы обоснования региональных стратонов фанерозоя Украины // Материалы XXXVII сессии Палеонтологического общества НАН Украины. Киев, 2016. С. 84–85.
Радионова Э.П., Орешкина Т.В., Хохлова И.Е., Беньямовский В.Н. Эоценовые отложения северо-восточного борта Днепровско-Донецкой впадины (зональная стратиграфия и циклический анализ) // Стратиграфия. Геол. корреляция. 1994. Т. 2. № 6. С. 85–102.
Радьков В.М., Паничев В.В., Богданов В.М., Шевченко Л.Н., Мишакова Н.Т., Молотков Л.С., Черкашин Д.Н. и др. Отчет о результатах геологического доизучения масштаба 1 : 200 000 на площади листов М-37-XVII, XI (Павловск, Бобров) за 1999–2001 г.г. (Воронежская область), в 5 книгах и 2 папках. Воронеж, 2002.
Семенов В.П. Палеоген Воронежской антеклизы. Воронеж: Изд-во Воронежского ун-та, 1965. 279 с.
Стратиграфический кодекс России. Ред. Жамойда А.И. СПб.: Изд-во ВСЕГЕИ, 2019. 96 с.
Стрельникова Н.И. Палеогеновые диатомовые водоросли. СПб.: Изд-во Санкт-Петербургского ун-та, 1991. 312 с.
Успенская Ю.М. Современное состояние изучения микрофлоры и микрофауны харьковского яруса Днепрово-Донецкой впадины // Ученые записки Харьковского ун-та. 1950. Т. 10. С. 71–74.
Чигуряева А.А. Атлас микроспор из третичных отложений СССР. Харьков: Изд-во Харьковского гос. ун-та, 1956. 261 с.
Шешукова-Порецкая В.С., Глезер З.И. Новые виды морских палеогеновых диатомовых водорослей УССР // Новости систематики низших растений. М.–Л.: Наука, 1964. С. 78–92.
Шпуль В.Г. Споры и пыльца кантемировской и журавкинской свит верхнего палеогена Среднего Дона // Сб. студенческих научн. работ Воронежского гос. ун-та (естественные науки). Воронеж: Изд-во Воронежского ун-та, 1972. С. 70–73.
Шпуль В.Г. Палинологическая характеристика стратотипического разреза эоцен-олигоценовых отложений в окрестностях с. Сергеевки бассейна р. Россошь // Современные проблемы геологии. Материалы юбилейной научн. сессии геол. ф-та Воронежского гос. ун-та. Воронеж, 1998. С. 19–20.
Шпуль В.Г. Новые данные по фитостратиграфии эоцен-олигоцена юго-восточного склона Воронежской антеклизы // Вестник Воронежского гос. ун-та. Сер. Геология. 2005. № 1. С. 55–70.
Шпуль В.Г. Фитостратиграфия эоцена северо-восточного склона Воронежской антеклизы // Проблеми бiостратиграфii нижнего протерозою и фанерозою Украiни. Збiрник наукових праць. Iнституту геологiчних наук. Киiв: Ін-т геологічних наук НАН України, 2006. С. 208–214.
Шпуль В.Г. Эоцен-олигоценовые флоры Воронежской антеклизы по данным палинологических исследований // Палеонтологiчнi дослiдження в Украiнi: iсторiя, сучасний стан та перспективи. Киев: Ін-т геологічних наук НАН України, 2007. С. 255–260.
Шпуль В.Г. Палиностратиграфия палеоцена–нижнего эоцена северо-восточного склона Воронежской антеклизы // Биосферные основы построения стратиграфических схем фанерозоя Украины. Материалы XXXI сессии палеонтологического общества НАН Украины, 19–22 мая 2008 г., Симферополь. Киев: Ін-т геологічних наук НАН України, 2008. С. 144–149.
Шпуль В.Г. Палеоцен-эоценовые флоры Воронежской антеклизы по данным палинологических исследований // Викопна фауна i флора Украiни: палеоекологiчний та стратиграфiчний аспекти. Збiрник наукових праць Iнституту геологiчних наук НАН Украiни. Киiв: Ін-т геологічних наук НАН України, 2009. С. 220–225.
Шпуль В.Г. Значение палинологии для обоснования региональных стратиграфических подразделений палеогена Воронежской антеклизы // Эволюция жизни на Земле. Материалы 4-го Международного симпозиума. Томск, 2010. С. 426–429.
Яковлева А.И. Детализация эоценовой диноцистовой шкалы для восточного Перитетиса // Бюлл. МОИП. Отд. геол. 2017. Т. 92. Вып. 2. С. 32–48.
Яковлева А.И., Щербинина Е.А., Музылев Н.Г., Александрова Г.Н. Диноцисты среднего–верхнего эоцена разреза Актумсук, Устюрт, Узбекистан: биостратиграфия и палеообстановки // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 6. С. 78–102.
Яковлева А.И., Александрова Г.Н. Палеоген Преддонецкой моноклинали и его палинологическая характеристика // Стратиграфия. Геол. корреляция. 2021. Т. 29. № 1. С. 59–98.
Agnini C., Backman J., Boscolo-Galazzo F., Condon D.J., Fornaciari E., Galeotti S., Giusberti L., Grandesso P., Lanci L., Luciani V., Monechi S., Muttoni G., Pälike H., Pampaloni M.L., Papazzoni C.A., Pearson P.N., Pignatti J., Premoli Silva I., Raffi I., Rio D., Rook L., Sahy D., Spofforth D.J.A., Stefani C., Wade B.S. Proposal for the Global Boundary Stratotype Section and Point (GSSP) for the Priabonian Stage (Eocene) at the Alano section (Italy) // Episodes. 2020. https://doi.org/10.18814/epiiugs/2020/020074
Agnini C., Fornaciari E., Raffi I., Catanzariti R., Pälike H., Backman J., Rio D. Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes // Newslett. Stratigr. 2014. V. 47. P. 131–181.
Berggren W.A., Kent D.V., Swisher C.C. III, Aubry M.-P. A revised Cenozoic geochronology and chronostratigraphy. Geochronology, Time Scales, and Global Stratigraphic Correlation. Eds. Berggren W.A., Kent D.V., Aubry M.-P., Hardenbol J.A. SEPM Spec. Publ. 1995. V. 54. P. 129–212.
Blanco S., Wetzel C.E. Replacement names for botanical taxa involving algal genera // Phytotaxa. 2016. V. 266. № 3. P. 195–205. https://doi.org/10.11646/phytotaxa.266.3.3
Brinkhuis H. Late Eocene to early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994. V. 107. № 1–2. P. 121–163.
Bukry D. Eocene siliceous and calcareous phytoplankton, Deep Sea Drilling Project Leg 95 // Init. Repts. DSDP. 1987. V. 95. P. 395–415.
Crouch E.M., Brinkhuis H. Environmental change across the Paleocene–Eocene transition from eastern New Zealand: a marine palynological approach // Mar. Micropaleontol. 2005. V. 56. P. 138–160.
Iakovleva A.I. Middle-Late dinoflagellate cysts from NE Ukraine (Borehole No 230, Dnepr-Donets Depression): stratigraphic and palaeoenvironmental approach // Acta Palaeobot. 2015. V. 55. № 1. P. 19–51.
Iakovleva A.I. Carpatella rossica sp. nov., a new Late Paleocene-Eocene dinoflagellate species from European Russia and Ukraine // Acta Palaeobot. 2019. V. 59. № 2. P. 277–288.
Iakovleva A.I., Heilmann-Clausen C. Early and middle Eocene dinoflagellate cysts from the Aktulagay section, Kazakhstan // Palynology. 2020. https://doi.org/10.1080/01916122.2019.1705933
Khokhlova I.E., Radionova E.P., Beniamovskii V.N., Shcherbinina E.K. Eocene stratigraphy of key sections of the Dnieper-Donets Depression based on calcareous and siliceous microplankton // Geodiversitas. 1999. V. 21. № 3. P. 453–476.
Locker S. Cenozoic siliceous flagellates from the Frame strait and the east Greenland margin: biostratigraphic and paleoceanographic results // Proc. Ocean Drilling Program, Scientific Results. Eds. Thiede J. et al. 1996. V. 151. P. 101–124.
Martini E. Standard Tertiary and Quaternary calcareous nannoplankton zonation // Proc. II Planktononic Conference. Roma, 1970. P. 739–785.
Okada H., Bukry D. Supplementary modification and introduction of code numbers to the low-coccolith biostratigraphic zonation (Bukry, 1973; 1975) // Mar. Micropaleontol. V. 5. № 3. P. 321–325.
Popova I.M., Baumgartner P.O., Guex J., Tochilina S.V., Glezer Z.I. Radiolarian biostratigraphy of Paleogene deposits of the Russian Platform (Voronesh Anticline) // Geodiversitas. 2002. V. 24. № 1. P. 7–59.
Powell A.J., Brinkhuis H., Bujak J.P. Upper Paleocene-lower Eocene dinoflagellate cyst sequence biostratigraphy of southeast England // Correlation of the Early Paleogene in Northwest Europe. Geol. Soc. Spec. Publ. 1996. V. 101. P. 145–183.
Radionova E.P., Beniamovski V.N., Iakovleva A.I., Muzylöv N.G., Oreshkina T.V., Shcherbinina E.A., Kozlova G.E. Early Paleogene transgressions: stratigraphical and sedimentological evidence from the northern Peri-Tethys // Geol. Soc. Am. Spec. Pap. 2003. V. 369. P. 239–261. https://doi.org/10.1130/0-8137-2369-8.239
Sluijs A., Pross J., Brinkhuis H. From greenhouse to icehouse: organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene // Earth-Sci. Rev. 2005. V. 68. P. 281–315.
Williams G.L., Fensome R.A., MacRae R.A. The Lentin and Williams index of fossil dinoflagellates 2017 edition // Am. Assoc. Stratigraphic Palynologists Contrib. Ser. 2017. № 48.
Witkowski J., Bohaty S.M., Edgar K.M., Harwood D.M. Rapid fluctuations in mid-latitude siliceous plankton production during the Middle Eocene Climatic Optimum (ODP Site 1051, western North Atlantic) // Mar. Micropaleontol. 2014. № 106. P. 110–129.
Zosimovich V.Yu., Shevchenko T.V. Development stages of Paleogene sedimentary basins of Northern Ukraina // Сб. научных трудов Института геологических наук Украины. 2014. Т. 7. С. 83–100.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция