

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 3, стр. 72-92
Морской палеоген Корякского нагорья, Северо-Восточная Азия: стратиграфия, моллюски, корреляция и геологические события
Ю. Б. Гладенков *
Геологический институт РАН
Москва, Россия
* E-mail: gladenkov@ginras.ru
Поступила в редакцию 06.10.2020
После доработки 16.11.2020
Принята к публикации 06.12.2020
Аннотация
Рассматриваются морские отложения среднего эоцена–олигоцена, а также подстилающие и перекрывающие их толщи, которые приурочены к четырем кайнозойским прогибам и двум впадинам, расположенным в периферийных зонах мезозоид Корякского нагорья Северо-Востока Азии. Эти отложения были сформированы во время значительной морской трансгрессии, охватившей также смежные регионы Камчатки, Сахалина и Японии. Для расчленения формаций древних шельфовых зон использованы комплексы моллюсков, которые позволили обосновать корреляцию стратиграфических горизонтов верхней части палеогена во всей Северо-Тихоокеанской области. На основе анализа этих комплексов выявлены палеоклиматические флуктуации (потепления в эоцене и похолодание в позднем эоцене–олигоцене), которые вызывали миграции морской палеобиоты.
ВВЕДЕНИЕ
Корякское нагорье (или Корякская горная страна) расположено в северо-восточной части Азии. С юго-востока и востока оно омывается Беринговым морем, а на юго-западе – водами Пенжинского залива Охотского моря. На северо-западе его ограничивает Пенжинская депрессия, на севере – р. Анадырь, а на крайнем северо-востоке – Анадырский залив. Нагорье состоит из небольших хребтов, гряд и кряжей высотой от 600 до 1800 м. Горная система ориентирована в целом с северо-востока на юго-запад, простираясь на 880–1200 км при ширине от 80 до 270 км. Общая площадь структуры более 500 000 км2. Основная часть горной системы имеет альпинотипный рельеф. Хотя Корякское нагорье в целом относится к зоне мезозойской складчатости, в его пределах, в основном по его периферии (побережье Пенжинской губы Охотского моря, Ильпинско-Олюторский, Хатырский и Анадырский районы Беринговоморского побережья), достаточно широко развиты морские толщи палеогена и неогена, приуроченные к нескольким протяженным прогибам и впадинам. Несмотря на проведенное во многих районах мелко- и среднемасштабное геологическое картирование, морские кайнозойские толщи изучены здесь не всегда полно и с разной детальностью. Достаточно сказать, что некоторые участки с третичными образованиями геологами совсем не посещались или сведения о них опираются на данные, полученные в отдельных маршрутах далеких лет. Изучению кайнозойских толщ мешает также не всегда хорошая их обнаженность и отсутствие во многих районах материалов бурения. Наконец, нельзя забывать, что палеоген-неогеновые отложения часто сильно дислоцированы.
КРАТКИЕ СВЕДЕНИЯ О СТРАТИГРАФИЧЕСКОЙ ИЗУЧЕННОСТИ РЕГИОНА И МАТЕРИАЛ К ХАРАКТЕРИСТИКЕ МОРСКИХ ОТЛОЖЕНИЙ ПАЛЕОГЕНА
В течение последних десятилетий стратиграфия морских третичных толщ тематически изучалась в основном геологами Северо-Восточного геологического управления и СВКНИИ ДВО РАН г. Магадана (прежде всего В.И. Волобуевой и А.М. Девятиловой), Чукотприродресурса г. Анадыря (Д.И. Агапитовым) и Геологического института РАН г. Москвы (Ю.Б. Гладенковым). Ими был опубликован ряд работ по этой тематике, включая статьи и монографии (Гладенков, 1972, 2020; Волобуева, Красный, 1979; Волобуева и др., 1988; Девятилова, Волобуева, 1981; Агапитов, Митрофанова, 1998 и др.). В этих публикациях приводились оригинальные данные по стратиграфии палеогена и неогена отдельных районов данного региона с изложением новых материалов по литологической и палеонтологической характеристике изученных образований. Автор данной статьи долгие годы сотрудничал с В.И. Волобуевой, вместе с которой были организованы несколько тематических экспедиций в регионе, прежде всего в районы Олюторского (Ильпинский п-ов) и Хатырского прогибов. Совместные изыскания, сопровождавшиеся анализом палеонтологических остатков (прежде всего морских моллюсков), позволили прийти к ряду стратиграфических обобщений по кайнозою Корякского нагорья, отраженных, в частности, в региональных стратиграфических схемах, которые широко используются при геолого-съемочных и буровых работах (Решения…, 1998), а также в ряде сводок (Волобуева и др., 1988, 1994). К сожалению, Вилена Исаевна Волобуева, плодотворно работавшая в данном регионе около 50 лет, в 1990-х годах ушла из жизни, не закончив изучение части коллекции моллюсков, собранных нами совместно. Впоследствии эта коллекция была передана автору данной статьи, который к настоящему времени изучил ее лишь частично – ревизована фауна верхней части палеогена Ильпинского п-ова и нижнего палеогена и нижнего миоцена восточного фланга Хатырского прогиба (А. Гладенков, Ю. Гладенков, 2007; Гладенков, 2020). Только недавно представилась возможность снова обратиться к этой коллекции, чтобы с учетом новых материалов внести определенные коррективы в характеристику комплексов моллюсков древних толщ разных районов, уточнить сопоставление палеогеновых толщ различных структур региона и на этой основе попытаться реконструировать отдельные геологические события в регионе. Следует напомнить, что объектом изучения служили осадочные образования геосинклинального типа, отличающиеся большой мощностью, фациальной невыдержанностью, часто сильной дислоцированностью.
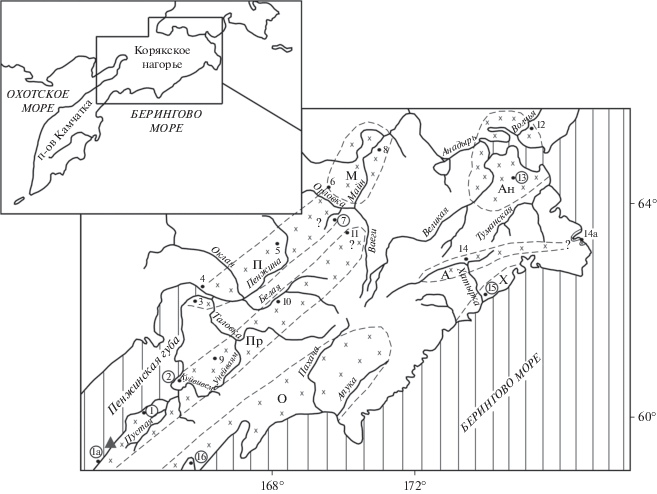
На рис. 1 показаны местонахождения 16 основных разрезов морского палеогена и неогена, которые были изучены в прошлые годы. Эти разрезы находятся в разных районах Корякского нагорья, которые, как говорилось, приурочены к разным прогибам и впадинам. Отдельные разрезы отличаются относительной полнотой и могут считаться опорными: это разрезы Западной Камчатки и Ильпинского п-ова. Именно они дают наиболее полное представление о литологическом характере морских образований и их палеонтологических комплексах в определенных структурных зонах. Предметом дальнейшего анализа будут палеогеновые (прежде всего среднеэоценовые–олигоценовые) отложения упомянутых 16 разрезов. Их описание дается по прогибам и впадинам, к которым они приурочены. В связи с тем, что отдельные разрезы уже были охарактеризованы в ряде публикаций, ниже они описаны в кратком виде и с разной полнотой. При этом особое внимание уделяется выявлению роли ископаемых моллюсков разных свит и горизонтов в корреляционных построениях. Это относится в первую очередь к осадочным образованиям и комплексам среднего–верхнего эоцена и олигоцена, которые изучались с целью сопоставления древних толщ разных районов и реконструкции геологических событий именно этого в определенном отношении переломного этапа в геологической истории данного региона. Вместе с тем, помимо материалов по горизонтам среднего эоцена–олигоцена, приводятся данные по подстилающим и перекрывающим толщам, чтобы нагляднее представлять особенности кайнозойских формаций и их границы.
Рис. 1.
Кайнозойские прогибы и впадины Корякского нагорья и местонахождение разрезов палеогена. Прогибы: Пр – Парапольский, П – Пенжинский, О – Олюторский, Х – Хатырский (или Опухско-Пекульнейский), А – Алькатваамский (или Иомраутский); впадины: М – Марковская, Ан – Анадырская; 1–16 – номера разрезов, номера в кружках – разрезы, в которых автор изучал моллюски. Треугольником показано положение обнажения, приведенного на рис. 3.
При необходимости для определения возраста отдельных стратонов привлекались данные и по другим палеонтологическим группам, прежде всего по фораминиферам (Беньямовский и др., 1992; Серова, 2001 и др.). Работы М.Я. Серовой по фораминиферам кайнозоя Камчатской области широко известны, но в последние 25–30 лет в изучение этой группы много нового внес В.Н. Беньямовский. Он являлся автором соответствующих разделов во многих монографиях по палеогену Западной Камчатки и юга Корякского нагорья (Волобуева и др., 1994; Гладенков и др., 1997, 2013 и др.). В дальнейшем при описании разрезов используются сведения по фораминиферам прежде всего упомянутых специалистов. С.И. Бордунов по просьбе автора уточнил определения некоторых приводимых ниже таксонов фораминифер с учетом их стратиграфического положения в современных региональных схемах.
ПРОГИБЫ И ВПАДИНЫ КАЙНОЗОЯ КОРЯКСКОГО НАГОРЬЯ И ОСОБЕННОСТИ МОРСКИХ РАЗРЕЗОВ
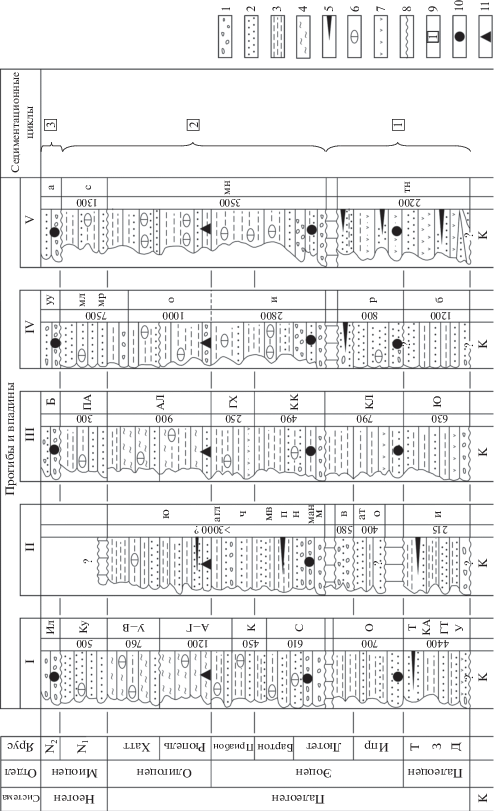
Выше говорилось, что в пределах Корякского нагорья выделяются несколько прогибов и впадин, выполненных третичными образованиями. Наиболее значительные из них показаны на рис. 1. Впервые они были намечены в 1960–1980-е гг. (Егиазаров и др., 1965; Волобуева и др., 1988). Всего было выявлено семь крупных таких структур, которые приурочены в основном к периферии нагорья и сложены терригенными образованиями различной мощности. Среди крупных кайнозойских прогибов можно выделить Парапольский и Пенжинский в западной части нагорья; Олюторский, Хатырский (или Опухско-Пекульнейский) и Алькатваамский (или Иомраутский) в южной и юго-восточной частях. Кроме них отмечаются Марковская и Анадырская впадины, приуроченные к северной и восточной частям нагорья. Эти наложенные и частично унаследованные прогибы и впадины не всегда имеют четкие границы (из-за недостаточной обнаженности и тектонических дислокаций), но все же достаточно уверенно распознаются в современной структуре, что позволяет, хотя и с определенными допущениями, судить о составе и мощности выполняющих их толщ и восстанавливать палеогеографические обстановки. Как уже говорилось, наше внимание будет сосредоточено на характеристике отложений морского происхождения, прежде всего, среднего эоцена–олигоцена, с целью выявления особенностей формирования древних толщ в разных седиментационных бассейнах. Приводимый ниже стратиграфический материал дается в обобщенном виде и включает не только ранее опубликованные сведения, но и ряд новых и пересмотренных данных (рис. 2). При этом по возможности сохраняются названия ранее выделенных свит. На рис. 2 изображены западнокамчатский разрез Парапольского прогиба (I) и ильпинский разрез Олюторского прогиба (III), которые являются опорными соответственно для северо-западной и юго-восточной частей Корякского нагорья, а также сводные разрезы для Пенжинского прогиба и Марковской впадины (II), для Хатырского прогиба (IV) и для Анадырской впадины (V).
Рис. 2.
Схема корреляции эоценовых и олигоценовых отложений разных прогибов и впадин Корякского нагорья. 1 – конгломераты и гравелиты; 2 – песчаники; 3 – алевролиты, аргиллиты; 4 – кремнистые породы; 5 – угли; 6 – карбонатные конкреции, 7 – вулканические породы, 8 – размывы и несогласия; 9 – седиментационные циклы (серии); 10, 11 – комплексы моллюсков: 10 – тепловодные, 11 – холодноводные. Цифры справа от литологических колонок обозначают мощность стратонов (м). Система, подотделы и ярусы: К – меловая система, Д – даний, З – зеландий, Т – танет, N1 – нижний миоцен, N2 – средний миоцен. I – Парапольский прогиб, II – Пенжинский прогиб, III – Олюторский прогиб, IV – Хатырский прогиб, V – Анадырская впадина. Сокращения стратонов: (I) У – улэвенейский горизонт, ГТ – геткилнинский горизонт, КА – камчикский горизонт, Т – ткаправаямский горизонт, О – оммайский горизонт, С – снатольский горизонт, К – ковачинский горизонт, АГ – аманинско-гакхинский горизонт, УВ – утхолокско-вивентекский горизонт, Ку – кулувенский горизонт, Ил – ильинский горизонт; (II) и – ичигемская толща, о – оммайская свита, ат – атольская толща, в – вильгильвэемская свита, м – марковская свита, ман – манильская толща, н – никлекуюльская толща, п – пальматкинская свита, мв – мавринская толща, ч – чокмавээмская свита, агл – агликичская толща, ю – южнинская свита; (III) Ю – южно-ильпинский горизонт, КЛ – кыланский горизонт, КК – килакирнунский горизонт, ГХ –гаилхавиланский горизонт, АЛ – алугинский горизонт, ПА – пахачинский горизонт, Е – ежовый горизонт; (IV) б – быстрореченская толща, р – ретырвеемская свита, и – ионайская свита, о – ольховская свита, мр – моржовская свита, мл – маллэнская свита, уу – ундал-уменская свита; (V) тн – танюрерский горизонт, мн – майницкий горизонт, с – собольковская свита, а – автоткульская свита.
Парапольский прогиб
Палеогеновые толщи Парапольского прогиба в пределах его южного фланга в последние десятилетия весьма полно изучены сотрудниками ГИН РАН. Прогиб расположен в западной части Корякского нагорья между мезозойскими структурами и простирается от побережья Пенжинской губы Западной Камчатки на северо-восток, в бассейн р. Белая. Его длина достигает 700–800 км при максимальной ширине 45–50 км (рис. 1, разрезы 1, 1а, 2, 9, 10, 11). Самыми изученными в этом прогибе являются западнокамчатские разрезы Чемурнаутской и Маметченской бухт (разрезы 1 и 2 соответственно на рис. 1). Опорным разрезом Западной Камчатки всегда считался Точилинский (рис. 1, разрез 1а), который расположен в Тигильском районе, несколько южнее разрезов 1 и 2 (рис. 1). Этому разрезу была посвящена монография с большим блоком палеонтологических описаний (Атлас…, 1984), но, к сожалению, палеоген в ней был охарактеризован лишь частично (дополнением служили публикации по палеогену для Тигильского района Л.В. Криштофович (1947, 1961)). Позднее палеогеновые толщи районов, примыкающих к местонахождению Точилинского разреза с севера и юга (Чемурнаутская бухта, Паланский район, Майначская бухта, бухта Квачина, Каврано-Утхолокская бухта), были детально охарактеризованы Ю.Б. Гладенковым и др. (1991, 1997, 2005, 2013, 2018). Моллюски из них были изучены В.Н. Синельниковой и Ю.Б. Гладенковым. В итоге в данном регионе в стратиграфической последовательности были выделены несколько палеогеновых горизонтов (или региоярусов) региональной схемы (Решения…, 1998), которые широко используются в геологической практике.
Улэвенейский горизонт (верхи маастрихта (?)–датский ярус). Предполагается, что его нижний контакт несогласный. Горизонт включает толщу вулканитов (базальты, андезиты и риолиты, мощность 700 м), свиту Мыса Зубчатого (ритмично чередующиеся тонкослоистые песчаники и алевролиты с флорой нижнего палеоцена, мощность 750 м), а также сосопханскую и хулгунскую свиты Тигильского района (песчаники, конгломераты, алевролиты, мощность более 1000 м; разрез 1а на рис. 1). К этому горизонту относится и напанская толща, сложенная угленосными образованиями с флорой. В отдельных слоях горизонта отмечены относительно редкие раковины морских моллюсков палеоцена: Macrocallista kovatschensis L. Krisht., Modiolus ornatus Gabb, Batissa cf. magna Dev., Eucrasatella lincolnensis (Weaver), Aturia angusta (Conrad) и др. (Криштофович, 1947).
Геткилнинский горизонт (датский–танетский ярусы). Описан в Чемурнаутской бухте (рис. 1, разрез 1), сложен преимущественно алевролитами с прослоями песчаников, мощность до 1400 м. В нем выявлены несколько слоев с моллюсками и бентосными фораминиферами. Среди моллюсков А.Д. Девятиловой (Девятилова, Волобуева, 1981) и Ю.Б. Гладенковым отмечены Acila decisa (Conrad), Nuculana gabbii (Conrad), Thyasira baca Dev., Periploma eodiscus Dev., Pitar murphy Zinsm., Molopophorus atquicostatus Vokes. Из фораминифер выявлены палеоценовые Silicosigmoilina californica Cushman et Church, Kolchidina danica Morozova, Lituotuba lituiformis (Brady) и др. (здесь и далее определения В.Н. Беньямовского).
Камчикский горизонт (танетский ярус). Залегает согласно, состоит из песчаников и алевролитов с углистыми прослоями, мощность до 1000 м. Среди палеонтологических остатков выявлены моллюски и бентосные фораминиферы. Из моллюсков отмечены Yoldia kilanskensis Glad., Miltha parsoni Waring, Acanthocardia brewerii (Gabb), Callista conradiana (Gabb), Nemocardium linteum (Conr.), Acrilla durhami Zinsm. Из фораминифер, согласно В.Н. Беньямовскому, типичны Spiroplectammina paleocenica Cushman и Rabdammina robusta Grzyb. Морским толщам в Паланском районе соответствует анадыркская континентальная свита (конгломераты, песчаники, мощность 380 м) с флорой.
Ткаправаямский горизонт (верхняя часть палеоцена–нижний эоцен). Нижний контакт согласный. Типичными для горизонта являются толщи морского и континентального генезиса, которые включают ритмично чередующиеся конгломераты, песчаники и алевролиты, местами с углями, мощность до 1000 м и более. В морских отложениях Чемурнаутского района отмечены остатки тепловодных моллюсков Barbatia landesi (Weaver et Palm.), Venericardia venturensis Waring, Callista stantoni (Nelson), Pitar sinedepressa (Nelson), Gari hornii (Gabb), Eocernina hannibali Dick., Modiolus speciosus Dev., Amaurupsis meierensis Zinsm. Фораминиферы представлены комплексом верхнего палеоцена с Nonionella insecta (Schwger).
В Маметченской бухте (рис. 1, разрез 2) вскрываются толщи палеоцена (зеландия–танета), ипра и нижнего лютета. К ним относятся две свиты: унэльская и оммайская. Они входят в состав камчикского и ткаправаямского горизонтов, отличаясь от их стратотипов некоторыми фациальными особенностями. Унэльская свита (зеландий–часть ипра) с размывом залегает на меловых образованиях. В ней выделяются две части. Нижняя (мощность 500–600 м) сложена песчаниками с линзами гравелитов и конгломератов. Моллюски, по В.Н. Синельниковой и Ю.Б. Гладенкову (Гладенков и др., 2013), представлены Nuculana snatolensis Slod., Crassatella stillwaterensis Weav., Venericardia mandaica (Yok.), Macoma tigilensis L. Krisht., Solen tigilensis Slod., Thracia kovatschensis L. Krisht. Верхняя часть свиты представлена песчаниками и алевролитами (мощность 700–800 м). Здесь отмечен представительный комплекс моллюсков: Yoldia chehalisensis Arn., Eucrassatella washingtoniana (Weav.), Crassatella lincolnensis (Weav.), Cr. stillwaterensis Weav., Cr. yessoensis Minato et Kimano, Pitar tigilensis L. Krisht., Venericardia mandaica (Yok.), V. mulleri Verastegui, Acesta perrini (Waring), Modiolus napanensis Sin., Thyasira baca Dev., Dacridium penjicus Sin., Glycymeris rosecanyonensis Hanna, Periploma efimovae Dev., Nemocardium linteum (Conrad). В нижней части свиты встречаются бентосные фораминиферы палеоцена–нижнего эоцена Alabamina wilcoxensis (Toulmin), Rhizammina indivisa Brady, Dentalina colei Cushman et Dusenbury, Cyclamina cf. pacifica Beck, Plectochammina poronaiensis (Asano), Bulimina debilis Martin и др. (определения В.Н. Беньямовского).
Оммайский горизонт (нижний эоцен–низы среднего эоцена). Его стратотип (оммайская свита) находится в Маметченской бухте. Горизонт надстраивает палеогеновый разрез и сложен чередующимися песчаниками и алевролитами мощностью до 700 м. Среди моллюсков отмечены Acila parva Dev., Yoldia kilanskiensis Glad., Nuculana semenova Dev., Turcicula praesakhalinensis (Dev. et L. Krisht.), Variamussium devjatilovae L. Krisht., Eucrassatella washingtonensis (Weav.). В.Н. Беньямовским определены также планктонные и бентосные фораминиферы ипра и нижнего лютета: Pseudohastigerina wilcoxensis (Cushman et Ponton), Morozovella wilcoxensis (Cushman), Subbotina patagonica Todd et Kniker, Acarinina coalingensis (Cushman et Hanna), Plectofrondicularia kerni Cook, Dentalina catenula Reuss и др.
Стратиграфически выше оммайского горизонта залегают особенно интересующие нас горизонты среднего эоцена–олигоцена (рис. 1, разрез 1а). Отметим, что стратотипами для них служат одноименные свиты.
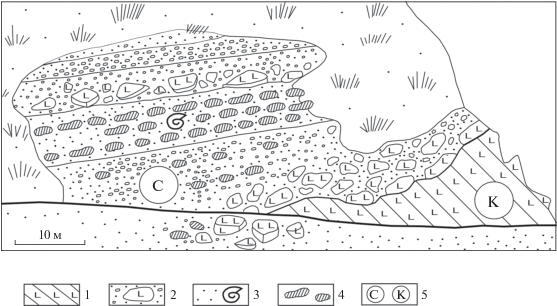
Снатольский горизонт (верхняя часть среднего эоцена). Его стратотип (снатольская свита) находится в Тигильском районе Западной Камчатки. Горизонт начинает новый седиментационный цикл в регионе. Это важный стратиграфический рубеж. Нижняя граница горизонта резко несогласная. Она прослеживается на сотни километров вдоль всего побережья Западной Камчатки. Горизонт залегает с размывом и конгломератами в основании на разновозрастных породах мезозоя и низов палеогена. На рис. 3 приведена зарисовка углового несогласия в подошве данного горизонта на охотоморском побережье в районе Кахтанинского мыса, расположенного в 17 км южнее устья р. Палана (его местонахождение отмечено на рис. 1 треугольником). Горизонт сложен в основном зеленовато-серыми полимиктовыми песчаниками с прослоями конгломератов в нижней части и алевролитов в верхней. Характерны карбонатные мелкие и крупные конкреции, редко отмечаются мелкие глендониты. Местами породы угленосные. Общая мощность до 450–670 м. В горизонте отмечены слои с многочисленными моллюсками, среди которых типичны Macrocallista snatolensis L. Krisht., M. xenophonti Slod., Mytilus yokoyamai Slod., Mytilus littoralis Slod., Modiolus restorationensis Van Winkle, Solen tigilensis Slod., Pitar micrum Sin., Plicacesta someshomai (Oyama et Mizuno), Nuculana kovatschensis Sin., Yoldia watasei Kaneh., Y. watasei ogasawarai Uozumi, Nemocardium kovatschensis L. Krisht., N. morysvillensis Dick., Venericardia aff. hornii (Gabb), Miltha kamtschatica Pronina, Acantocardia snatolensis L. Krisht., Papyridea utcholokensis Slod., Neverita washingtonensis (Weav.), Trominina dispar Takeda, Colus asagaiensis Mak., Turcicula nagaoi Noda, Genota snatolensis Ilyina. Этот сравнительно представительный комплекс моллюсков среднего эоцена хорошо трассируется по всему региону. Среди бентосных фораминифер В.Н. Беньямовский идентифицировал Rhizammina indivisa Brady, Bathysiphon nodosariaformis Subbotina, Cyclammina ezoensis Asano и др. В Паланском районе в усть-анадыркской свите, относимой к нижней части снатольского горизонта, встречены редкие планктонные фораминиферы среднего эоцена: Globigerina boweri Bolli, Pseudohastigerina wilcoxensis (Cushman et Ponton) (Серова, 2001).
Рис. 3.
Зарисовка несогласного залегания снатольской свиты среднего эоцена на вулканитах толщи мыса Кахтанинского палеоцена–нижнего эоцена в Паланском районе на побережье Охотского моря Западной Камчатки (севернее Точилинского разреза) (по Гладенков и др., 1991). 1 – вулканиты; 2 – глыбы, конгломераты и гравелиты; 3 – песчаники с моллюсками; 4 – песчано-карбонатные конкреции; С – снатольская свита, К – толща мыса Кахтанинского.
Ковачинский горизонт (верхи среднего эоцена–верхний эоцен). Залегает согласно, со следами незначительного размыва в основании. Он сложен в основном коричневато-бурыми и черными тонкослоистыми алевролитами и аргиллитами, в верхней части голубовато-серого цвета, с прослоями песчаников и туфов. Отмечаются небольшие карбонатные конкреции и мелкие глендониты. Мощность до 450 м. Выделен комплекс моллюсков с Yoldia takaradaiensis acuta L. Krisht., Y. watasei Kaneh., Y. kovatschensis (Slod.), Malletia poronaica (Yok.), Cardita kovatschensis L. Krisht., Modiolus restorationensis Van Wincle, Laternula korniana L. Krisht., Delectopecten kamtschaticus L. Krisht., Hataiella aff. poronaitnsis (Takeda), H. hellenica L. Krisht., Pitar kryshtofovichii Sin., Turcicula praesakhalinensis (Dev. et L. Krisht.), Turritella tokunagai Yok. Многие моллюски этого сообщества встречаются на этом уровне и в других районах прогиба. В сообществе фораминифер cреднего и верхнего эоцена отмечены: Ammodiscus concinnus Kuznetsova, Haplophragmoides subimpressus Voloshinova, Plectofrondicularia packardi Cushman et Schenck, Globulina landesi (Hanna et Hanna), Gyroidina condonei Cushman et Schenck, Melonis shimokinensis (Asano), Caucasina cf. schwageri Yok. (Серова, 2001; Бордунов и др., 2020).
Аманинско-гакхинский горизонт (верхний э-оцен–олигоцен). Его нижний контакт согласный, горизонт сложен двумя свитами. Нижняя, аманинская, включает темно-серые кремнистые и глинистые туфоаргиллиты с прослоями глауконитовых песчаников и туфов. Вторая, гакхинская, состоит из светло-серых кремнистых туфоаргиллитов, туфоалевролитов, перекристаллизованных опок, туфопесчаников и туфов. Типичны карбонатные конкреции, встречаются глендониты. Общая мощность до 1200 м. Комплекс моллюсков включает часто встречающиеся виды: Yoldia longissima Slod., Y. nitida Slod., Y. watasei Kaneh., Y. deformis Slod., “Yoldia” multidentata Khom., Acila praedivaricata Nagao et Husioka, Malletia inermis Yok., Modiolus solea Slod., Nemocardium iwakiense (Mak.), Chlamys matschgarensis Mak., Ch. gackhensis L. Krisht., Papyridea harrimani Dall, Periploma besshoensis (Yok.). Представители этого характерного комплекса отмечаются во многих разрезах сопредельных районов – от Японии на юге до Чукотки на севере. Бентосные фораминиферы, по данным М.Я. Серовой (2001) и С.И. Бордунова (Бордунов и др., 2020), представлены Melonis tumiensis Kuznetsova, Gavelinella glabrata (Cushman), Glandulina laevigata d’Orbigny, Cyclammina placenta (Reuss), Guttulina problema d’Orbigny. В верхней части горизонта отмечены диатомеи олигоценового возраста Pyxilla aff. reticulatа Grove et Sturt, Pseudotriceratium radiosoreticulatum Gruniw, Kisseleviella carina Sheshukova, Actinostephanus rudus Chursevich et Tsoy, Odontella aff. aurita (Lyngbye) Agardh. (Атлас…, 1984; Гладенков, 2007).
Утхолокско-вивентекский горизонт (олигоцен). Залегает согласно. Нижняя его часть представлена темно-серыми и зеленовато-серыми кремнистыми туфогенными алевролитами с частыми маломощными прослоями туфов, верхняя – тонкослоистыми кремнистыми туфоаргиллитами с опоками. В основании этих частей местами отмечается галечно-валунный материал. Для горизонта в Точилинском разрезе характерны мелкие и крупные (до 0.5 м) глендониты разнообразной формы, а также карбонатные конкреции. Местами встречаются скопления галек. Мощность до 750–760 м. Комплексы моллюсков включают в основном относительно глубоководные мелкораковинные виды: Nuculana slodkewitschi Kogan, N. utcholokensis Ilyina, N. majamraphensis (Khom.), N. mironovi L. Krisht., Yoldia mediana Gladikova, Y. pennulata Slod., Y. mira Gladikova, Delectopecten utcholokensis L. Krisht., Macoma kamtschatica Slod., Periploma besshoensis (Yok.), Papyridea aff. harrimani Dall. Присутствуют фораминиферы, среди которых, по данным М.Я. Серовой и С.И. Бордунова, наиболее типичны олигоценовые виды Grabratella isabeliana d’Orbigny, Astrononion hamadaensis Asano, Ammodiscus macilentus Chapman, A. concinnus Kuznetsova (Бордунов и др., 2020). В горизонте отмечены также находки диатомовых водорослей олигоцена (Pyxilla aff. reticulatа Grove et Sturt, Kisseleviella carina Sheshukova, Stephanopyxis grunowii Grove et Sturt, St. spinosissima Grunow, Odontella aff. aurita (Lyngbye) Agardh, Cosmodiscus asteromphalus Ehrenberg) (Атлас…, 1984). Недавно А.Ю. Гладенковым (2018; A. Gladenkov, 2019) были получены новые данные по зональному расчленению олигоценовых отложениий горизонта бухты Квачина (юго-западная часть Тигильского района), которые впервые позволили достоверно выделить на Камчатке олигоценовые ярусы международной шкалы (рюпель и хатт).
Кулувенский горизонт (нижний миоцен). Залегает без видимого несогласия и сложен преимущественно туфопесчаниками c карбонатными конкрециями, вверху с прослоями конгломератов (общая мощность до 300–500 м). Он характеризуется моллюсками Yoldia chojensis Sim., Y. thraciaeformis (Stor.), Keenocardium kuluvense Kafanov, Mytilus ochotensis Slod., Serripes groenlandicus (Brug.), Thiasira disjuncta ochotica Gabb и др. Этот комплекс является маркирующим для нижнего миоцена всего Камчатско-Сахалинского региона (Решения…, 1998).
Ильинский горизонт (нижняя часть среднего миоцена). Залегает трансгрессивно, с размывом в основании, начинает новый седиментационный цикл кайнозойского разреза в регионе и поэтому является хорошим маркером. Его слагают в основном конгломераты и песчаники мощностью до 400 м. Горизонт охарактеризован богатым относительно тепловодным комплексом моллюсков (до 150 видов) с Panomya elongate Kanno, Modiolus wajampolkensis Slod., Chlamys kaneharai (Yok.), Arca watanabai Kanno, Crepidula princeps Conrad и др. Комплекс фауны этого уровня распространен по всей Северо-Тихоокеанской провинции.
В других районах Парапольского прогиба палеогеновые образования представлены хуже. Из-за фрагментарной обнаженности и фациальной изменчивости отложений, а также из-за их неполной палеонтологической характеристики детально определить последовательность слоев и оценить их возраст часто затруднительно. Тем не менее здесь помимо континентальных образований (вулканитов и угленосных толщ) выделяются морские толщи с фауной. В северо-восточной части Парапольского прогиба в разрезах 9 и 10 (междуречье Уюл–Унэнваям и бассейн рек Ванетат и Учхичхиле) палеоцен-эоценовые морские толщи практически не представлены (хотя имеются небольшие выходы алевролитов неопределенного возраста). На олигоценовом уровне появляются вулканиты (липариты, андезиты, базальты, мощность до 1500 м), в нижней части которых выявлены прослои алевролитов и углистых сланцев. Зато северо-восточнее, в разрезе 11 (междуречье Майн–Ваега и верховье р. Пальматкина) отмечаются отложения с морской фауной. На майнской толще, видимо, нижнего палеогена, которая представлена континентальными песчаниками с прослоями алевролитов и бурого угля, с размывом залегает пальматкинская толща (песчаники, местами с прослоями карбонатизированных алевролитов и аргиллитов, мощностью до 450 м). В ней отмечены редкие остатки раковин мелководных моллюсков: Ostrea palmatkensis L. Krisht., Lima oskvillensis Clark., Acesta cf. sameshima (Oyama et Mizumo). По-видимому, эта толща относится к среднему–верхнему эоцену. Стратиграфически выше залегают песчаники и алевролиты с Variamussium pillarense Slod., Venericardium mandsi Yok., Periploma kariboensis L. Krisht., которые обычны для олигоцена.
Таким образом, в Парапольском прогибе в том или ином виде выявляются следы морской трансгрессии конца среднего эоцена–олигоцена, которая наиболее ярко проявилась в пределах Западной Камчатки, но затронула и другие районы прогиба. Она маркируется литологическими особенностями формаций (сменой грубообломочных образований сравнительно глубоководными алевролитовыми и часто кремнистыми отложениями) с замещением вверх по разрезу соответствующих биотических сообществ. В 1994 г. при обсуждении стратиграфических схем палеогена–миоцена Западной Камчатки было предложено выделять три серии: тигильскую (палеоцен–низы среднего зоцена), воямпольскую (верхи среднего эоцена–нижний миоцен), кавранскую (средний миоцен–верхний миоцен), которые соответствуют определенным седиментационным циклам (Решения…, 1998). Эти серии были намечены раньше (Геология…, 1964), но их объемы и возраст были несколько изменены с учетом новых данных. Приведенные выше материалы прямо соответствуют этим построениям. В изученных разрезах четко выделяются названные серии, в частности уверенно обособляется вторая серия, служившая предметом нашего повышенного внимания.
Пенжинский прогиб
Этот прогиб расположен между двумя антиклинорными структурами и вытянут в северо-восточном направлении на 600 км с максимальной шириной 45–50 км. Северо-западный борт прогиба соприкасается с краевой зоной Охотско-Чукотского вулканического пояса. В этом прогибе, расположенном севернее Парапольского прогиба, можно отметить разрезы, в которых были найдены морские отложения палеогена (рис. 1, разрезы 3, 4, 5, 7). Но из-за относительно слабой изученности района и недостаточной обнаженности сопоставить многие толщи отдельных участков крайне затруднительно (возможно, из-за фациального разнообразия пород разобщенных участков геологи выделили здесь слишком много свит). Сводный разрез здесь можно составить только по отдельным блокам часто с условными границами. В таловской структурно-фациальной зоне (на западе прогиба) наиболее древней из палеогеновых отложений считается ичигемская толща, относимая к палеоцену и сложенная на правобережье р. Пенжина конгломератами, песчаниками и туфоалевролитами с флорой (мощность 55–215 м). Она с перерывом и размывом перекрывается оммайской свитой морского генезиса, состоящей из песчаников и гравелитов (мощность 85–100 м) с моллюсками Yoldia kilanskiensis Glad., Nuculana semenova Dev., Turcicula praesakhalinensis (Dev. et L. Krisht.), Variamussium devjatilovae L. Krisht. Она относится к нижнему и частично среднему эоцену. Выше несогласно залегает манильская толща верхнего эоцена–олигоцена, которая представлена континентальными конгломератами и песчаниками (с флорой) мощностью свыше 850 м.
На восточном побережье Пенжинской губы, от мыса Астрономического до р. Таловка (рис. 1, разрез 3), палеоген представлен свитой Мыса Астрономического среднего эоцена. Она залегает несогласно на меловых образованиях и включает песчаники, алевролиты и аргиллиты с карбонатными стяжениями (мощность 300 м). Среди моллюсков отмечены: Nuculana napanica L. Krisht., Yoldia watasei Kaneh., Y. aff. nitida Slod., Spirotropis kincaidi (Weav.), встречается большое количество раковин Nautilus. Эта свита перекрывается южнинской свитой олигоцена, представленной алевролитами и песчаниками с прослоями конгломератов и линзами лигнитов. Мощность до 1000–1300 м. Среди моллюсков определены Ostrea kirikovi Slod., Modiolus utcholokensis Slod., Nemocardium kovatschensis L. Krisht., Spisula callistaeformis Dall.
На северном побережье Пенжинской губы (рис. 1, разрез 4) к нижнему палеогену отнесена атольская толща песчаников (до 300 м) с пластами конгломератов, реже алевролитов с растительными остатками и редкими моллюсками Solen tigilensis Slod., Macrocallista tigilensis L. Krisht., Corbicula kamtschatica L. Krisht. В верховьях р. Пенжина и в бассейнах рек Черная, Березовая, Осиновая (рис. 1, разрез 5) выделена вильгильвэемская свита (мощность 500–580 м) среднего эоцена. Она представлена крупногалечными конгломератами и песчаниками, в которых отмечены мелководные моллюски: Modiolus esutornensis L. Krisht., Lucina hannai Clark, Ostrea rekinikensis Slod., O. idriaensis Gabb, O. agnevoensis Laut., Anomia maganiglensis Hanna, Periploma circularis L. Krisht. Эта свита перекрывается чокмавээмской свитой верхнего эоцена, сложенной мелкогалечными конгломератами и туфогенными аргиллитами мощностью до 420–480 м. В них присутствуют моллюски Acila shumardi (Dall), Yoldia packardi (Clark), Malletia kovachensis L. Krisht., Delectopecten kornensis L. Krisht., Dentalium circularis Weav. Северо-восточнее несогласно залегают грубозернистые песчаники с линзами аргиллитов и ракушняков мощностью до 300–430 м, выделенные в пальматкинскую и никлекуюльскую толщи среднего–верхнего эоцена. В нижней части преобладают конгломераты и песчаники (70–180 м), в которых отмечены моллюски Nuculana snatolensis Slod., Yoldia takaradaiensis L. Krisht., Y. chehalinensis (Arn.), Glycymeris tenuimbricata Clark, Thyasira snatolensis L. Krisht., Spisula packardii Dick., Taras kovatschensis L. Krisht., Pitar tigilensis L. Krisht. и др. Выше они сменяются алевролитами, аргиллитами с прослоями песчаников (около 250 м). Среди моллюсков встречаются Yoldia chehalinensis (Weav.), Malletia kovachensis L. Krisht., Lima twinensis Slod., Variamussium pillarense Slod., Modiolus restorationensis Van Winkle, Turritella washingtonensis (Weav.).
Северный фланг Пенжинского прогиба к северо-востоку смыкается с Марковской впадиной (рис. 1, разрезы 6, 8). Впадина характеризуется морскими отложениями значительной мощности. В бассейне рек Гребенка, Майн и др. (разрез 6) выделяется марковская свита среднего эоцена общей мощностью свыше 700 м. Она залегает несогласно на меловых образованиях и сложена песчаниками, конгломератами и алевролитами с углистыми прослоями в нижней части. В них отмечаются моллюски: Yoldia olympiana Clark, Y. takaradaiensis L. Krisht., Modiolus trinominata (Hanna), M. porterensis Clark, Mytilus snochomisensis Weav., M. yokoyamai Slod., Tellina lincolnensis Weav., Solen tigilensis Slod., Acila packardi Clark, Cardita ex gr. ksenophonti L. Krisht., Glycymeris andersoni Dall, Gl. chehalisensis Arn., Gl. branneri Arn., Crassatella lincolnensis Weav., Ostrea ex gr. esutoriensis L. Krisht. Морские отложения палеогена встречены также в бассейне р. Майн (разрез 8). Здесь развита мавринская толща, относимая к среднему эоцену. В основании отмечаются крупногалечные конгломераты. Для толщи типичны конгломераты и песчаники с прослоями алевролитов и аргиллитов и с линзами лигнитов. Общая мощность толщи от 300 до 500 м. Среди моллюсков определены: Glycymeris banner Arn., Crassatella lincolnensis Weav., Turritella tembloroensis Wiedy. На левобережье р. Майн (разрез 8) на мавринской свите, видимо, с перерывом залегает агликичская толща. Она сложена песчаниками, пластами конгломератов, в средней части с линзами угля. Мощность 600–850 м. В ней найдены моллюски: Nuculana calkinsi Moore, Yoldia nairoensis Evs., Y. ex gr. watasei Kaneh., Y. excavata Dall, Papyridea ex gr. matschigarica Khom., Nemocardium cf. yokoyamai Tak., Cardita ex gr. tokudai Tak., Anomia schmidti L. Krisht., Acila conradi (Meek.), Crassatella amaninensis L. Krisht., Neptunea vinjukovi L. Krisht. По-видимому, агликичская толща имеет позднеэоцен-олигоценовый возраст (Афанасьева и др., 1974).
Из представленного материала можно сделать вывод, что в Пенжинском прогибе и Марковской впадине обнаруживаются явные следы среднеэоценовой–олигоценовой трансгрессии, которая особенно ярко проявилась в Западно-Камчатском регионе, в Парапольском прогибе. Вместе с тем создается впечатление, что морские толщи более древнего возраста здесь распространены относительно мало, а в чередовании с морскими отмечаются континентальные образования. Тем не менее мощность морского среднего эоцена–олигоцена здесь солидная – до нескольких тысяч метров (Решения…, 1998).
Если обратиться к южным и восточным окраинам Корякского нагорья (Олюторский, Хатырский, Алькатваамский прогибы и Анадырская впадина), то там тоже обнаруживаются следы палеогеновой трансгрессии, хотя в ряде случаев она проявилась по-разному.
Олюторский прогиб
Этот прогиб является, возможно, продолжением Восточно-Камчатского кайнозойского прогиба. Он вытянут вдоль побережья Берингова моря на 450 км при ширине 80–100 км. Здесь на Ильпинском п-ове (рис. 1, разрез 16) расположен один из лучших кайнозойских разрезов всей Северо-Тихоокеанской области, охарактеризованный богатыми планктонными комплексами фораминифер (данные М.Я. Серовой (2001) и В.Н. Беньямовского (Волобуева и др., 1994)). В данном разделе использованы материалы по зональной схеме В.Н. Беньямовского. В отличие от вышеописанных, данный разрез представлен относительно глубоководными фациями. Он расчленен на несколько горизонтов (региоярусов), которые для прогиба и прилегающих территорий являются опорными. Они подробно описаны в монографии (Волобуева и др., 1994). Ниже дана их краткая характеристика.
Южно-ильпинский горизонт (палеоцен). Залегает с размывом и, видимо, с некоторым несогласием на меловых толщах. Он представлен туфопесчаниками (в нижней части) и туфоалевролитами мощностью до 630 м. Для горизонта характерны бентосные фораминиферы палеоцена Rzehakina epigone Rzehak., Rz. minima Cushman et Renz, Glomospira corona Cushman et Jurvis, Silicosigmoilina californica Curch. В верхней части горизонта встречены планктонные фораминиферы танетского яруса Globigerina nana Bolli, Subbotina velascoensis (Cushman), Acarinina nitida (Martin), Morozovella aequa (Cushman et Renz) (здесь и далее определения В.Н. Беньямовского). Моллюски здесь редкие и плохой сохранности, в нижней части отмечены, в частности, Ostrea sp.
Кыланский горизонт (нижний эоцен–низы среднего эоцена). Он представлен флишоидным чередованием тефроидных аргиллитов и алевролитов (с включением прослоев туфопесчаников) мощностью 750–790 м. Здесь отмечен комплекс относительно глубоководных моллюсков: Nuculana semenovi Pron., Nucula suworovi Pron., Yoldia kilanskensis Glad., Y. breviscapha distincta Pron., Acila decisa ilpinensis Pron., Malletia ilyina Pron., Variamussium indigenum Pron., Orectospira wadana Yok. В породах В.Н. Беньямовским обнаружены также планктонные и бентосные фораминиферы ипра и лютета: Globigerina pileata Kchalilov, Globorotalia subbotinae Morozova, G. wilcoxensis Cushman, Acarinina clara Chal., Globigerina linaperta Finl., Gl. pseudoeocenica Subbotina, Gl. boveri Bolli, Silicosigmoilina sogabei Fuk. и др.
Килакирнунский горизонт (верхи среднего э-оцена–низы верхнего эоцена). Хотя нижний контакт является согласным, в основании горизонта отмечается характерная пачка (мощностью до 20 м) чередующихся гравелитов, туфопесчаников и туфоалевролитов. Комплекс моллюсков представлен Periploma altarata L. Krisht., Neilo semenovi Pron., Variamussium indigenus Pron., Orectospira wadana (Yok.). В этом горизонте отмечены планктонные фораминиферы: Globigerina boweri Bolli, G. praebulloides Blow, G. pseudoeocaena compacta Subbotina, Globigerapsis index (Finl.) (лютетский и бартонский ярусы), Globigerapsis tropicalis (Blow et Banner) (приабонский ярус). Бентосные фораминиферы представлены Chilostomella ovoidea Reuss, Bulimina corrugate Cushman et Schenck, Globobulimina pacifica Cushman, Gyroidina octocamerata Cushman et Hanna, Praebulimina reussi (Morrow), Plectofrondicularia packardi Cushman et Schenck, P. trilineata Cushman, Caucasina eocaenica (Yok.), C. kamtschatica Serova.
Гаилхавиланский горизонт (верхний эоцен). Залегает согласно, в его основании находится лапареламский слой кислых туфов (20 м). Горизонт сложен в основном тефроидными аргиллитами с карбонатными конкрециями, общей мощностью около 250 м. В комплексе моллюсков определены Variamussium pillarense Slod., Cyclocardia kovatschensis (Slod.), Lima twinensis Slod., Turritella poronaiensis Tak., Acesta kovatschensis Sin., Malletia poronaica Tak. Много морских ежей. Из бентосных фораминифер отмечены Caucasina schwageri (Yok.), Poronaia poronaiensis (Asano), Globobulimina pacifica Cushman, Plectofrondicularia sersi Cushman, Haplophragmoides laminates Voloshinova.
Алугинский горизонт (олигоцен). Надстраивает разрез с согласным нижним контактом. Он сложен тефроидными аргиллитами с карбонатными конкрециями, с прослоями кремнистых опоковидных пород (диатомитов). В нижней части залегает мулатханский горизонт глауконитовых песчаников. Мощность алугинского региояруса 900 м. Среди моллюсков встречаются Acila praedivaricata Nag. et Hug, Yoldia watasei Kaneh., Cyclocardia ilpinensis Pron., Papyridea harrimani Dall, Periploma besshoensis (Yok.), Modiolus solea Slod., Trominina bicordata (Hatai et Koike), Neptunea ezoana Tak. Этот в целом бореальный комплекс является весьма типичным и для олигоцена сопредельных районов. Фораминиферы представлены олигоценовыми Ammudiscus ariacensis Mur., Asanospira carinata Cushm., Melonis shimokinensis (Asano), Cribrelphilium sumitomoi Asano et Mur. и др. Здесь же А.Ю. Гладенковым отмечены олигоценовые комплексы диатомовых водорослей: Cavitatus cf. jouseanus Williams, Stephanopyxis grunowii Grove et Sturt, St. superba Grunow, Odontella sawamurae Akiba, Ikebea tenuis (Brun) Akiba и др. (А. Гладенков, Ю. Гладенков, 2007; A. Gladenkov, 2009).
Пахачинский горизонт (нижний миоцен). Залегает согласно, сложен песчаниками и алевролитами с моллюсками Yoldia chojensis Sim., Thracia schenckii Clark, Thyasira ochotica Gabb, Clinocardium asagaiensis (Mak.), Mya truncata L. и др.
Ежовый горизонт (средний миоцен). Залегает трансгрессивно, с размывом в основании. Он характеризуется тепловодным комплексом моллюсков Dosinia kaneharai Yok., Mytilus tichanovichi Mak., Clinocardium pseudofastatum (Nomura), Macoma optiva (Yok.), Mya cuneiformis (Bohm) и др.
Из рассмотренных данных следует, что в разрезе палеогена Олюторского прогиба, где развиты в основном относительно глубоководные отложения (алевролиты и аргиллиты), можно выделить толщи верхов среднего эоцена–олигоцена, в нижней части которых отмечаются конгломератовые пачки, а в верхней части (олигоценовой) – алевролиты с прослоями опок и диатомитов, с фауной бореального типа.
Хатырский (Опухско-Пекульнейский) прогиб
Прогиб протягивается вдоль побережья Берингова моря от нижнего течения р. Опуха до мыса Наварина на 285 км при ширине до 30–40 км наземной части. Положение разреза палеогеновых морских отложений (точнее, серии разрезов) показано на рис. 1 (разрез 15). Надо отметить, что в пределах северной приграничной зоны прогиба развиты многочисленные тектонические нарушения, в частности надвигового типа, что затрудняет изучение контактов мезозойских и кайнозойских толщ. В прогибе выделяются три части: западная, центральная и восточная, каждая из которых отличается некоторыми фациальными особенностями и мощностью развитых в них третичных толщ (Гладенков и др., 1985). Важно, что в прогибе, кроме наземных обнажений, для характеристики палеогеновых образований были использованы около 15 скважин и сейсмические материалы (Агапитов, Митрофанова, 1998).
В центральной части структуры в одной из скважин (Ямракоимская 33) на глубине 721 м вскрыты датские слои с планктонными фораминиферами Globorotalia trinidadensis Bolli, G. pseudobulloides (Plum.) и др. (Алабушева, 1996). По всей видимости, эти слои входят в состав основания Хатырского бассейна. На сейсмических разрезах центральной части прогиба отчетливо видно, что отложения, относимые к среднему эоцену, залегают с угловым несогласием на верхнем мелу и, видимо, нижнем палеогене, представляя собой нижний элемент нового седиментационного цикла. Он начинается ионайской свитой среднего–верхнего эоцена–олигоцена. Свита представлена внизу конгломератами, переходящими в темно-серые алевролиты и аргиллиты с прослоями песчаников и мелкогалечных конгломератов. Мощность до 800 м. В комплексе моллюсков определены Acila praedivaricata Nagao et Huzioka, Yoldia longissima Slod., Y. watasei Kaneh., Modiolus solea Slod., Turritella tokunagai Yok. Среди фораминифер отмечены Plectofrondicularia packardi multilineata Cushman, Haplophragmoides gratus Ter-Grig., Cibicides elmaensis Rau, Cyclamina pacifica Beck, Cribroelphidium minimum Cushman и др. (Серова, 2001). К олигоцену здесь относится и хайидинская свита, которая внизу представлена конгломератами и песчаниками, в средней части – флишоидным чередованием алевролитов и песчаников, а вверху – алевролитами и аргиллитами (Волобуева и др., 1988). Общая мощность свиты до 2150–2500 м. Среди моллюсков встречаются Malletia inermis (Yok.), Variamussium kronokiensis Pron., Cyclocardia yakatagensis (Clark.), Neptunea ombetsuensis Matsui.
В то же время в восточной части прогиба на меловых толщах с угловым несогласием залегает ретырвеемская свита нижнего–среднего (?) эоцена (Гладенков, 2020). Лежащие в основании конгломераты (3–8 м) сменяются пачками черных крепких аргиллитов и массивных песчаников часто во флишоидном переслаивании. В породах отмечаются карбонатные конкреции – крупные шаровидной формы (в диаметре до 0.5 м) и мелкие конкреции (до 0.1 м). Общая мощность около 800 м. В породах найдены моллюски: Yoldia watasei Kaneh., Crassatella branneri (Var.), Glycymeris sagitata (Gabb), Solena aff. townsendensis Clark, Periploma circularis L. Krisht., Cucullaea beringiana Vol., Tudicula aff. copiosa Dev., Turritella aff. pachikoensis Stant., Polinices nuciformis cowlitzensis (Dickerson) и др. В этой толще Л.И. Митрофановой (Гладенков и др., 1985) отмечены фораминиферы Cibicides pseudoungerianus evolutus Cushman et Hobson, Dentalina dusenburyi Beck, D. colei Cuchman et Dusen, D. cf. legumen Reuss, Nodogenerina wegemanni (Cole), Pseudoglandulina cf. ovate d’Orbigny, Cyclammina cancellata obese Cuchman et Laiming. Перечисленные толщи соответствуют южно-ильпинскому и кыланскому горизонтам Ильпинского разреза, составляющим седиментационный цикл палеоцена и нижнего эоцена, который наращивается выше среднеэоценовым–нижнемиоценовым циклом. Выше ретырвеемской свиты со значительным стратиграфическим перерывом трансгрессивно залегает моржовская свита нижнего миоцена. В нижней части отмечается пачка конгломератов и песчаников (50–80 м). Над ней залегают песчаники светло-серые, желто-серые, рыхловатые, средне-крупнозернистые, мощностью более 850 м. В них находятся серые песчано-карбонатные конкреции и прослои гальки. В гальке встречаются переотложенные обломки угля и конкреционных стяжений (с раковинами Turritella, перемытыми из рытервеемской свиты). Фауна включает комплекс моллюсков с Yoldia chojensis Sim, Glycymeris boraensis Laut., Thyasira disjuncta ochotica L. Krisht., Lucinoma acutilineata (Conrad), Clinocardium ornatocostatum Volobueva, Mya arenaria Linne, M. truncata Linne, Thracia kavranensis Ilyina, Mactra polynyma voyi (Gabb), Ancistrolepis aff. modestoides (Takeda). В этой же свите Л.И. Митрофановой определены фораминиферы: Reophax curtus Cushman, Haplophragmoides laminatus Voloshinova, Melonis cf. tumiensis Kuznetsova, Perfectononion incertarformis Kuznetsova. Свита соответствует пахачинскому горизонту нижнего миоцена Олюторского прогиба (возрастному аналогу кулувенского горизонта Западной Камчатки).
В западной части Хатырского прогиба вскрыт разрез эоцена–среднего миоцена. В нижней части залегает быстрореченская толща (видимо, нижнего палеогена), контакты которой остаются неизученными (Волобуева, Красный, 1979). Она сложена в основном песчаниками мощностью до 1000–1200 м. Стратиграфически выше находится ольховская свита (или толща), нижний контакт которой не выявлен, но по фауне она относится к среднему (?)–верхнему эоцену–олигоцену (Гладенков и др., 1985). Свита сложена темно-серыми алевролитами с прослоями песчаников. Местами, прежде всего внизу, отмечаются конгломераты (в частности, глинистые) и плавающая галька. Мощность свиты около 1000 м. Встречаются моллюски: Yoldia watasei Kaneh., Nemocardium iwakiense Mak., Papyridea matschigarica Khom., P. cf. harrimani Dall, Thyasira inflata Yabe et Nomura, Ostrea sp., Crassatella sp., Trominina japonica (Tak.). Комплекс фораминифер включает Rhabdammina aspera Voloshinova, Bathysiphon cf. eocenicus Cushman et Hanna, Cyclammina ezoensis Asano и др. (определения Л.И. Митрофановой). Стратиграфически выше согласно залегает маллэнская свита олигоцена–нижнего миоцена, включающая три подсвиты значительной мощности. Нижняя подсвита олигоцена сложена крупными пачками (около 50–70 м) глинисто-песчанистых пород с Yoldia laudabilis Yok., Y. watasei Kaneh., Neptunea ezoana Takeda. Мощность до 2100 м. Средняя подсвита, тоже относящаяся к олигоцену, представлена песчанистыми пачками, в которых отмечены прослои конгломератов, алевролитов и песчано-карбонатных конкреций. Ее мощность около 3000 м. Верхняя подсвита (2400 м) нижнего миоцена включает массивные и мусорные комковатые песчаники, в верхней части с песчано-карбонатными конкрециями. В средней подсвите отмечены моллюски Yoldia caudata Khom., Y. matschigarica L. Krisht., Clinocardium asagaiense Mak., C. schinjiense (Yok.), Neptunea onnaica Takeda, Turritella tocunagai Yok. В верхней подсвите идентифицированы Yoldia chojensis Sim., Mya arenaria Linne, Macoma simizuensis L. Krisht., Clinocardium californiense (Desh.). Суммарная мощность маллэнской свиты около 7500 м. Стратиграфически выше с трансгрессивным контактом залегает ундал-уменская свита (средний миоцен). Ее нижняя часть представлена конгломератами и песчаниками, верхняя – песчаниками и алевролитами. В породах найдены остатки Mya karaginskiensis Khark., Laevicardium taracaicum (Yok.), Taras gouldi sertunaiensis (Kogan) и др. Отмечены остатки фагусовых. Суммарная мощность до 980 м.
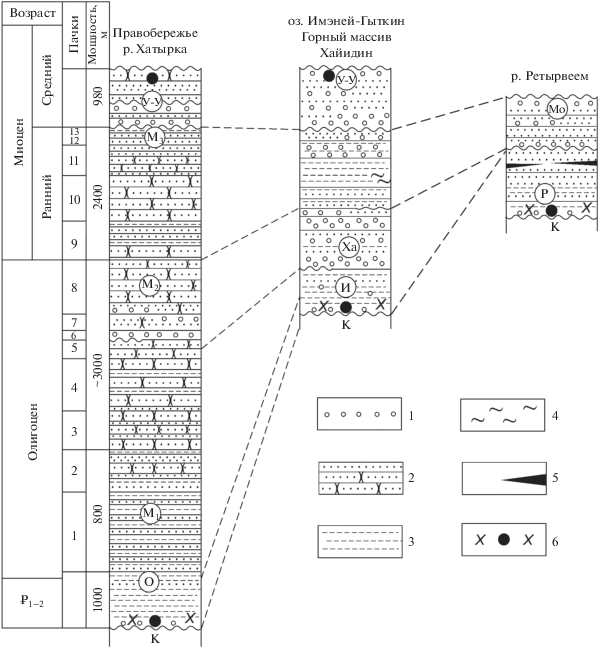
Приведенные материалы свидетельствуют о том, что в средне-позднеэоценовое и олигоценовое время в разных частях Хатырского прогиба палеогеографические и седиментационные обстановки были неодинаковые (Гладенков, 2020) (рис. 4). Так, в западной части (правобережье р. Хатырка) в целом накапливались грубообломочные песчано-конгломератовые отложения большой мощности (в частности, мощность одной маллэнской свиты превосходит 7000 м), формирование которых связывается с гипераккумуляционной деятельностью дельтовой системы р. Пра-Хатырка. Здесь часто отмечаются проградации в структуре осадочных комплексов, приведшие к образованию громадной линзы в шельфовой зоне осадконакопления (Гладенков и др., 1985). В средней части прогиба (оз. Имэней-Гыткин) одновозрастные слои представлены менее мощными (около 2000–2500 м) алевролито-песчанистыми отложениями (часть хайидинской свиты), которые формировались в сублиторальных условиях без активного участия речных систем. В самой восточной части прогиба (р. Ретырвеем) накапливались терригенные образования сначала верхне-нижнесублиторального, а затем относительно мелководного типа. При этом на средне-позднеэоценовое и олигоценовое время здесь, возможно, пришелся перерыв в осадконакоплении. Отмеченная неоднородность разрезов кайнозоя разных частей Хатырского прогиба обусловлена, видимо, различным проявлением и активностью тектонических процессов в этих частях, что приводило, видимо, к разным условиям осадконакопления и различной морфологии дна отдельных участков древнего бассейна.
Рис. 4.
Схема сопоставления разрезов кайнозоя западной, центральной и восточной частей Хатырского прогиба. 1 – конгломераты; 2 – слоистые песчаники; 3 – массивные песчаники; 4 – кремнистые породы; 5 – угли; 6 – комплексы тепловодных моллюсков. Сокращения: О – ольховская свита, М1 – нижнемаллэнская подсвита, М2 – среднемаллэнская подсвита, М3 – верхнемаллэнская подсвита, У-У – ундал-уменская свита, И – ионайская свита, Ха – хайидинская свита, Р – ретырвеемская свита, Мо – моржовская свита, К – мезозойские отложения, 1–2 – палеоцен–эоцен.
Алькатваамский прогиб
Прогиб расположен между двумя палеозойскими и мезозойскими выступами, вытянут на 350 км, ширина от 6 до 60 км. Протягивается от среднего течения р. Хатырка на северо-восток к бухте Угольной (рис. 1, разрез 14). Этот прогиб (его называют в литературе также Иомраутским) состоит, видимо, из двух частей: западной иомраутской и восточной краснореченской (известной как Краснореченский грабен) (Волобуева, Красный, 1979; Волобуева и др., 1988). Разрез здесь изучен недостаточно детально. Его низы в восточной и центральной частях прогиба представлены глухореченской толщей палеоцен-эоценового возраста мощностью 1000–1200 м. В ней отмечаются темно-серые алевролиты, в нижней части которых выявлены планктонные фораминиферы Globigerina triloculinoides Plumm., Globorotalia angulata (White), G. velanscoensis (Cushman). Из бентосных видов найдены Rhabdammina eocenica Cushm., Ammodiscus pennyi Cushman et Yary, Haplophragmoides coalingensis Cushman et Hanna (определения М.Я. Серовой). В западной части прогиба выделена мольская толща (до 800 м), в основании которой залегают песчаники, переходящие в алевролиты и аргиллиты со сходным комплексом микрофауны. Она перекрывается вычхынейской свитой среднего–верхнего? эоцена. Свита включает полимиктовые желтовато-серые разнозернистые песчаники мощностью 2000–2500 м. В них В.И. Волобуевой определены моллюски Crassatella utcholokensis L. Krisht., Acila decisa (Conr.), Solen sugenensis Conr., Venericardia horni Gabb и др. Выше по разрезу данную свиту сменяет змейковская свита верхнего эоцена–олигоцена. Она представлена темно-серыми алевролитами мощностью до 900 м. В верхней части отмечены пласты песчаников и конгломератов с характерными для олигоцена морскими моллюсками Acila praedivaricata Nag. et Hus., Naculana napanica L. Krisht., Yoldia watasei Kaneh., Callista furtiva (Yok.), Thracia condoni Dall, Mya grevingki elongata Nag. et Inoue. В центральной части прогиба распространена белогорская свита, которая тоже относится к олигоцену. Ее слагают полимиктовые песчаники с рассеянной галькой (до 1000 м), с комплексом моллюсков Acila praedivaricata Nag. et Hus., Papyridea harrimani Dall, Chlamys branneri Arnold, Clinocardium cf. okushirense Uozumi et Fujie.
В краснореченской части прогиба (в Краснореченском грабене) к среднему и верхнему эоцену относится краснореченская свита. В ее нижней части преобладают конгломераты и полимиктовые песчаники (600 м) с моллюсками Tivela aragoensis Turn., Epitonion turneri Durham, Turritella uvasana clequahensis Weav. et Palm. В верхней части типичными являются алевролиты, аргиллиты, глинистые песчаники (800 м). В этой свите В.И. Волобуевой определены моллюски Lima oakvillensis Clark., Crassatella washingtoniana Weav. и др.
В свете приведенных данных можно утверждать, что морская трансгрессия палеогена в Алькатваамской структуре оставила достаточно явные следы.
Восточнее данного прогиба находится небольшой Беринговский прогиб (бухта Угольная и лагуна Амаам), где обнажаются палеогеновые образования (соответственно чукотская и амаамская свиты; рис. 1, разрез 14a) (Бушуев, 1954; Волобуева, Красный, 1979). В основании разреза залегают нижнечукотская (бухта Угольная) и нижнеамаамская (лагуна Амаам) подсвиты, относимые к палеоцену. Первая из них мощностью 330 м состоит из полимиктовых песчаников с глинистой пачкой (25 м) в средней части и угольным пластом (“Речным”) в верхней. В подсвите отмечены бентосные фораминиферы Haplophragmoides formosus Takayanagi, Rzehakina cf. epigona (Rzehak), Ammodiscoides cf. conica Cushman (определения Н.А. Волошиновой). Среднечукотская подсвита (250 м) верхнего эоцена сложена песчаниками и глинистыми породами с тремя угольными пластами (“Мощный”, “Двойной”, “Карьерный”) с остатками растений (в частности, пальм). Верхнечукотская подсвита среднего эоцена (500 м) включает вулканомиктовые песчаники с прослоями конгломератов и тремя угольными пластами. В подсвите отмечены типичные для среднего эоцена морские моллюски Mytilus littoralis Slod., M. yokoyamai Slod., Spisula rushi aragoensis Turn., S. acutirostrata Packard.
Нижнеамаамская подсвита палеоцена (200–330 м) сложена гравелитами, песчаниками и в верхней части алевролитами. В основании находятся вулканомиктовые конгломераты (3 м), которые трансгрессивно, хотя без резкого несогласия перекрывают кампан-маастрихтскую корякскую серию. В подсвите В.И. Волобуевой отмечены моллюски Glycymeris amaaensis Volob., Crassatella uniodes Stanton, Turritella aff. pachecoensis Stanton. Комплекс фораминифер, по М.Я. Серовой, включает Bathysiphon kushiroensis (Yosh.), Haplophragmoides kushiroensis Yosh., Alabamina cf. hokkaidoensis (Yosh.). Среднеамаамская подсвита нижнего эоцена (580–600 м) состоит из песчаников и алевролито-аргиллитовых пород с четырьмя пластами каменного угля (1–1.5 м). В верхней части подсвиты встречены моллюски Сrassatella cf. uvasana semidentata (Coop.), Glycymeris aff. branneri Arn, Corbicula triangular Volob. Верхнеамаамская подсвита среднего эоцена сложена песчаниками и алевролитами в верхней части (400–450 м). Ее характеризуют моллюски Nuculana washingtoniana (Weaver), Crassatella stillwaterensis emimica Volob., Solen coosensis Turn., Pitar campi Vokes. Скорее всего, в прошлом существовала связь между Алькатваамским и Беринговским прогибами. Наличие в разрезах, наряду с морскими толщами, угленосных образований свидетельствует о неустойчивом режиме осадконакопления вблизи прибрежных зон.
Анадырская впадина
Впадина расположена в низовьях рек Анадырь и Великая (рис. 1, разрезы 12, 13). В этом районе пробурено большое число скважин (более 60), что обеспечивает достаточно хорошую корреляцию отложений разных частей впадины. В ее пределах выделяются три структурно-фациальных зоны: южная, центральная и северная, которые отличаются по литологическому характеру и мощности отложений (Агапитов, 1991; Агапитов, Митрофанова, 1998). В целом разрез палеогена представляется следующим образом.
В основании залегает танюрерский горизонт (палеоцен–нижний эоцен). Он представлен угленосными прибрежно-осадочными (песчаники и аргиллиты) и вулканогенными образованиями. В его пределах выделяют танюрерскую и ягельную толщи мощностью до 2000–2200 м. В ягельной толще А.В. Алабушевой (1996) отмечены планктонные фораминиферы (Globigerina trivialis Subbotina, G. triloculinoides Plummer) и бентосные комплексы палеоцена (Bathysiphon eocenicus Cushman et Hanna, Glomospira corona Cushman et Jarvis, Haplophragmoides exacavata Cushman et Waters, Cyclammina akkeshiensis (Yoshida)). На танюрерском горизонте с несогласием, перерывом и размывом залегает майницкий горизонт, включающий усть-черынайскую и майницкую свиты. По данным бурения, проводившегося в прибрежном районе Берингова моря, в разрезе под этим горизонтом отмечается кора выветривания (Маргулис и др., 2011). Майницкий горизонт сопоставляется со средним–верхним эоценом и олигоценом соседних районов и сложен преимущественно морскими терригенными толщами (песчаники и алевролиты, в верхней части с редкими прослоями диатомитов) большой мощности (от 1300 до 3500 м). В породах этого горизонта В.И. Волобуевой и Ю.Б. Гладенковым отмечены моллюски: Yoldia kovatschensis Slod., Y. cf. longissima Slod., Cardita striata L. Krisht., Laevicardium esutoruensis L. Krisht., Papyridea cf. utcholokensis Slod., Chlamys aff. gackhensis L. Krisht., Clinocardium cf. asagaiensis (Mak.). Здесь же выявлены фораминиферы Bathysiphon eocenicus Cushman et Hanna, Rhabdammina eocenica Cushman et Hanna, Cyclammina pacifica Beck, Plectofrondicularia packardi (Cushman et Schenck), Pl. multiliniata Cushman et Simonson (определения Л.И. Митрофановой).
Выше залегает собольковский горизонт нижнего миоцена. К нему относятся две свиты: собольковская и гагаринская, сложенные песчаниками и алевролитами с углями. Среди моллюсков отмечены Yoldia chojensis Sim., Hiatella pleshakovi (Sim.), Delectopecten cf. pedroanus (Tr.). Мощность горизонта до 950–1300 м. Упомянутые свиты сменяются вверх по разрезу автоткульской свитой среднего миоцена, залегающей трансгрессивно и представленной в основном песчаниками (200–690 м) с типичными среднемиоценовыми моллюсками Yoldia nabiliana (Sim.), Y. kuluntunensis Slod., Spisula voyi (Gabb), Macoma nasuta (Conrad) и др.
Майницкий горизонт в данной впадине маркирует значительную морскую трансгрессию среднего эоцена–олигоцена, сопровождавшуюся опусканием дна бассейна и накоплением терригенных толщ большой мощности.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ И ВЫВОДЫ
Синтез приведенных выше материалов по стратиграфии палеогена разных районов Корякского нагорья позволяет прийти к нескольким общим выводам.
1. В рассмотренных разрезах различных прогибов и впадин Корякского региона Северо-Востока Азии отмечена серия морских осадков, которая связана с крупной морской трансгрессией конца среднего эоцена–олигоцена. Строение этого седиментационного цикла можно представить в следующем виде (рис. 2, цикл 2). В основании серии (верхи среднего эоцена) залегают обычно мелководные конгломератовые и песчаниковые толщи, которые содержат гальку с обломками мезозойских и нижнепалеогеновых пород. Они надстраиваются относительно глубоководными глинистыми толщами (алевролиты, аргиллиты, местами кремнистые породы – опоки и диатомиты; верхний эоцен и олигоцен), которые отвечают максимуму трансгрессии. Выше эти отложения постепенно сменяются регрессивными песчано-конгломератовыми образованиями нижнего миоцена. Эта серия (на Камчатке она называется “воямпольской”) практически повсеместно залегает на подстилающих породах мела и палеоцена–нижнего эоцена с угловым несогласием или с размывом, а перекрывается трансгрессивно залегающими среднемиоценовыми толщами, относящимися к другому, средне-верхнемиоценовому, седиментационному циклу (“кавранская серия”).
В Парапольском и Пенжинском прогибах (западные и северо-западные районы Корякского нагорья) среднеэоценовое угловое несогласие выражено резко и прослеживается практически во всех описанных разрезах. В третичных структурах юго-восточной части нагорья (в Анадырской впадине и Хатырском прогибе) оно отмечается тоже достаточно четко. В Олюторском прогибе (юг нагорья), в котором развиты сравнительно глубоководные образования, начало трансгрессивной серии отмечается в виде размыва и появления в разрезе пачек гравелитов, залегающих без видимого углового несогласия. Следы этой значительной трансгрессии фиксируются и на Восточной Камчатке, где во многих районах также отмечается угловое несогласное залегание верхов среднего эоцена на подстилающих отложениях, в частности в Валагинском хребте (р. Корниловская), где автору пришлось его описывать раньше (Гладенков, 2016). И в пределах Сахалина морские толщи среднего эоцена (краснопольевский горизонт) тоже с размывом перекрывают континентальные угленосные образования нижнего палеогена, знаменуя и здесь начало “снатольской” трансгрессии (Гладенков и др., 2002). Во всех случаях возрастные аналоги сравнительно мелководной снатольской свиты (горизонта) перекрываются относительно глубоководными алевролитами и песчаниками “ковачинского–утхолокско-вивентекского” интервала (на Сахалине это такарадайские и аракайско-холмские толщи), который соответствует пику этой трансгрессии. Из этого следует, что отмеченное выше геологическое событие было достаточно широкомасштабным, ибо оно проявилось на значительной площади северо-восточной окраины Азии.
В данной статье не разбираются причины возникновения среднеэоценовой трансгрессии и появления в разрезах углового несогласия и размывов в основании трансгрессивно залегающих толщ среднего эоцена (обычно его середины). Одна из попыток объяснения этого явления была сделана несколько лет назад (Гладенков, 2013), когда было высказано предположение, что отмечающиеся в кайнозойских разрезах Камчатско-Корякского региона несогласия и трансгрессивные серии связаны с проявлением в этом регионе нескольких (до 5–6) фаз тектогенеза, приведших к структурным перестройкам и прогибанию отдельных структурных зон, в том числе и в пределах Корякского нагорья. Одна из таких фаз приходилась как раз на ранний эоцен–начало среднего эоцена, что и вызвало появление несогласий и размывов в середине среднего эоцена. С другой стороны, следует учитывать, что в литературе имеются указания на возможное относительное поднятие уровня Мирового океана (более 100 м) в конце эоцена и олигоцене, связаннoе с одним из эвстатических циклов второго порядка (Vail, Hardenbol, 1979; Haq et al., 1987). Но в активных тектонических зонах оценить роль этого явления очень трудно, хотя и списывать его нельзя.
2. В северо-западной части Корякского нагорья охарактеризованные выше прогибы имеют явно линейное простирание, что, возможно, отражает расположение древних рифтовых и депрессионных зон, а на востоке и юге наложенные впадины образовывались, видимо, вследствие опускания в палеогене по периферии мезозоид отдельных блоков, связанного с тектоническими дислокациями. Конечно, эти вопросы нуждаются еще в дополнительном освещении со стороны специалистов. Но в нашем случае уже сейчас ясно, что одна из тектонических активизаций в данном регионе, произошедшая в раннем эоцене–начале среднего эоцена, в значительной мере отразилась на седиментационных процессах в структурах Корякского нагорья. Несмотря на то, что она проявилась в разных районах различным образом, общая тенденция их развития все же улавливается. При анализе кайнозоя этого региона обращает на себя внимание относительно большая мощность (до нескольких тысяч метров) терригенных отложений в отмеченных выше прогибах и впадинах, что указывает на лавинную седиментацию. Видимо, это является следствием контрастных тектонических движений и интенсивного сноса осадочного материала при формировании структур. При определенной условности приведенных выше оценок мощности свит разных структур, все же очевидно, что мощность палеогена в них сравнительно большая – до 5000–7000 м (лишь в разрезе Ильпинского п-ова около 3360 м). Что касается интервала верхов среднего эоцена–олигоцена (осадки трансгрессивной части осадочного цикла), то его мощность оценивается в 3000–5000 м, за исключением Ильпинского разреза, где она около 1650 м. Эти показатели наглядно характеризуют одну из особенностей развития активных зон перехода от океана к континенту, где широко образуются формации геосинклинального типа большой мощности (Григоренко, 2011).
Вопросы общей тектоники и геодинамической эволюции мезозоид и альпид Корякского региона здесь не рассматриваются, и мы отсылаем читателя к специальной литературе и моделям по этой тематике (Егиазаров и др., 1965; Александров, 1978; Руженцев и др., 1982; Бондаренко, 2004; Чехович и др., 2009; Харахинов и др., 2014 и др.). Отметим только, что в современной структуре границы перечисленных выше прогибов и впадин Корякского нагорья часто осложнены молодыми, постмиоценовыми и постплиоценовыми, тектоническими нарушениями. Например, почти по всей северной части Хатырского прогиба отмечены следы надвигания с севера мезозойских образований на кайнозойские толщи. Тектонические разломы, в частности надвиги, фиксируются по периферии Анадырской впадины и Алькатваамского прогиба. Разломы в большом количестве отмечаются и в приграничных зонах западных прогибов. При этом разрывные нарушения имеются также в пределах самих кайнозойских прогибов и впадин, третичные толщи в которых обычно смяты в складки. Все это затрудняет изучение этих структур.
3. Снатольский и надстраивающие его палеогеновые горизонты вмещают характерные комплексы бентосной фауны (моллюски и фораминиферы), а в последнее время для датировки этих отложений стали использовать планктонные остатки (фораминиферы, частично диатомовые, наннопланктон и диноцисты). Установленные в разных районах комплексы моллюсков, которым было уделено специальное внимание, несмотря на некоторые различия палеогеновых разрезов в разных структурах Корякского нагорья (различия в фациях осадочных толщ, их мощности и пр.), достаточно уверенно сопоставляются друг с другом и с комплексами моллюсков региоярусов (или горизонтов) палеогена Камчатки и Корякского нагорья (Решения…, 1998). Сравнительное изучение палеосообществ моллюсков других районов послужило основой составления межрегиональной корреляционной схемы морского палеогена всей северо-западной части Азии (от Японии и Сахалина на юге до Аляски на севере). Она сопоставлена со схемой, составленной для Северотихоокеанского побережья Америки (штаты Вашингтон и Орегон) еще в прошлом веке и опубликованной в 1970-х годах (Гладенков, 1978). Не повторяя подробно уже опубликованные палеонтологические данные по трем горизонтам среднего эоцена–олигоцена Камчатско-Корякского региона, все же еще раз напомним, что каждый из них охарактеризован определенным комплексом моллюсков.
Для снатольского горизонта и его возрастных аналогов (верхи среднего эоцена) наиболее типичными являются Macrocallista snatolensis L. Krisht., Mytilus yokoyamai Slod., Mytilus littoralis Slod., Solen tigilensis Slod., Pitar micrum Sin., Plicacesta someshomai (Oyama et Mizuno), Yoldia watasei ogasawarai Uozumi, Nemocardium morysvillensis Dick., Venericardia hornii (Gabb), Miltha kamtschatica Pronina, Acantocardia snatolensis L. Krisht., Papyridea utcholokensis Slod., Neverita washingtonensis (Weav.), Trominina dispar Takeda, Colus asagaiensis Mak., Turcicula nagaoi Noda, Genota snatolensis Ilyina. Для следующего, ковачинского, региояруса и его возрастных эквивалентов (верхний эоцен) обычны Yoldia takaradaiensis acuta L. Krisht., Malletia poronaica (Yok.), Cardita kovatschensis L. Krisht., Modiolus restorationensis Van Wincle, Laternula korniana L. Krisht., Hataiella aff. poronaiensis (Takeda), Pitar kryshtofovichii Sin., Turcicula praesakhalinensis (Dev. et L. Krisht.), Turritella tokunagai Yok. Что касается аманинско-гакхинского горизонта (верхний эоцен–олигоцен), то он характеризуется широко распространенным в регионе характерным сообществом с Yoldia longissima Slod., Y. nitida Slod., Y. watasei Kaneh.,“Yoldia” multidentata Khom., Acila praedivaricata Nagao et Husioka, Malletia inermis Yok., Modiolus solea Slod., Nemocardium iwakiense (Mak.), Chlamys matschgarensis Mak., Papyridea harrimani Dall, Periploma besshoensis (Yok.).
Конечно, сопоставление комплексов моллюсков из разных разрезов такой значительной территории – дело непростое. И одна из причин этого – пока еще недостаточное их сравнительное изучение. Базовым источником сведений о палеогеновых моллюсках Японии до сих пор является одна большая сводка японских специалистов начала 1960-х годов (Oyama et al., 1960). Трудно переоценить и крупное обобщение по моллюскам из разрезов штатов Вашингтон и Орегон США Ч. Вивера (Weaver, 1942). Эти обобщения по объему палеонтологического материала являются уникальными и пока непревзойденными в Японии и Северной Америке. Автору в какой-то мере повезло, так как ему удалось познакомиться со многими палеогеновыми разрезами и коллекциями, находящимися в Японии и США. В Северной Японии ему удалось изучить формации Исикари, Поронай (эоцен) и Мамидзияма (олигоцен) вместе со свойственными им комплексами моллюсков. В Северной Америке особенно важными в этом же отношении оказались формации Ковлитц, Линкольн (эоцен), Твин-Ривер (верхи эоцена–олигоцен) и Блекли (олигоцен). Это изучение привело к выводу, что многие формы, выделенные в этих странах и в России под разными именами, являются либо одними и теми же видами, либо подвидами. Например, в американских коллекциях обнаружились виды, которые сходны с камчатскими и японскими и с которыми они образуют “пары”: Yoldia reagani = Y. orientalis, Y. scissurata = Y. ochotensis, Conchlodesma bainbridgensis = Laternula besshoensis, Variamussium wailandi = V. pillarense, “Cardium” hamiltonensis = Papyridea matchigarica и др. (Гладенков, 1979). Конечно, для стратиграфических корреляций приходится использовать многие виды японских и американских авторов. И это хорошо иллюстрируется приведенными выше списками моллюсков, отмеченных в горизонтах палеогена Корякского нагорья: часть использованных видов описана американскими авторами, вторая часть – японскими, а третья часть – российскими специалистами. При этом имеется достаточно много общих видов, которые находятся в разрезах шельфовых отложений всего Северо-Тихоокеанского региона или его азиатской части. В одной из монографий (Гладенков и др., 2005) приведены 55 биогеографических схем для эоцен-олигоценового интервала данной области, которые наглядно иллюстрируют эту мысль (были использованы данные по ареалам 75 видов моллюсков). В целом можно сказать, что многие книги и публикации российских геологов по стратиграфии палеогена Камчатки и Корякского нагорья (Криштофович, 1947, 1961; Гладенков, 1972, 2020; Гладенков и др., 1991, 1997, 2005, 2013, 2018; Волобуева, Красный, 1979; Волобуева и др., 1988; Девятилова, Волобуева, 1981 и др.) сопровождаются значительным блоком палеонтологического материала и в этом отношении не уступают публикациям наших коллег из соседних стран. Это свидетельствует о том, что уже сейчас заложена солидная база как для проведения широких стратиграфических и палеогеографических корреляций, так и для продолжения будущих более детальных исследований по стратиграфии восточных районов России. Изученные в последнее время планктонные комплексы (фораминиферы, диатомовые, наннопланктон, диноцисты), найденные в одних и тех же слоях вместе с бентосными остатками, позволяют не только контролировать корреляции регионального масштаба, но и привязывать региональные схемы к глобальной шкале с достаточно большой точностью.
4. Помимо значительной роли, которую играют ископаемые комплексы моллюсков в построении местных и региональных стратиграфических схем, эти сообщества помогают реконструировать некоторые особенности природных условий прошлого, в том числе палеогеографические обстановки и характер биособытий. Моллюски обычно обитают в шельфовых условиях, поэтому в ископаемом состоянии они особенно важны для определения обстановок формирования древних формаций шельфовых зон. Так, видовой и родовой состав комплексов определенных литологических фаций может использоваться для определения относительных глубин и гидрорежима их формирования (Кузнецов, 1963). Наличие в палеогеновых осадках тех или иных районов представителей неподвижных сестонофагов (Mytilus, Modiolus, Musculus, Ostrea) свидетельствует о мелководных условиях осадконакопления (глубина обычно 0–50 м) в прибрежной части древнего бассейна. Преобладание подвижных и малоподвижных фильтраторов (Cardium, Serripes и пр.) говорит об обстановке верхней сублиторали с глубиной до 100 м. Более глубокая зона маркируется доминированием в сообществах собирающих детритофагов (Macoma, Acila, Yoldia, Tellina), которые являются показателем спокойного гидрорежима и относительно больших глубин (до 300 м и более), свойственных нижнесублиторальным и верхнебатиальным обстановкам. Следующая по глубине зона грунтоедов развита в условиях малоподвижных вод батиали, нередко с дефицитом кислорода. Здесь моллюсков мало (отдельные виды Nucula, Malletia). Исходя из синтеза данных по моллюскам палеогена Корякского нагорья, а также сведений по литологическому составу изученных формаций, можно в целом сказать, что рассмотренные ранее среднеэоценовые толщи формировались в мелководных условиях в пределах верхней сублиторали (где развиты конгломераты, гравелиты и песчаники), а затем, в позднем эоцене и олигоцене, морские отложения накапливались в обстановках преимущественно верхней–нижней сублиторали (алевролиты и аргиллиты). Относительно глубоководные условия осадконакопления были характерны прежде всего для олигоценового времени, когда накапливались не только алевролито-аргиллитовые осадки, но и опоки и диатомиты. В ряде случаев палеонтологические находки достоверно маркируют изменения в положении береговой линии морских бассейнов.
С другой стороны, смена сообществ ископаемых моллюсков в разрезах северотихоокеанской зоны показывает, что в течение палеогена происходили эволюционные процессы, вызывавшие изменения родового и видового состава комплексов (в частности, изменение состава руководящих и характерных видов, доли впервые появившихся и исчезнувших видов, процента соотношения вымерших и ныне живущих форм, относительного количества эндемиков и пр.). Эти процессы отражались в определенной этапности развития палеокомплексов, что и учитывалось при выделении перечисленных выше горизонтов разных провинций (подробно см. Гладенков, 2018). Фактически каждый горизонт маркирует определенный этап развития морской экосистемы и свойственной ей палеобиоты. Среди наиболее важных границ в “палеосукцессиях” интервала среднего эоцена–нижнего миоцена следует считать рубеж эоцена и олигоцена, когда относительно тепловодные сообщества в регионе стали замещаться бореальными комплексами на фоне глобальной смены “оранжерейного” типа климата “ледниковым” типом (Гладенков, 2018).
5. Анализ сообществ моллюсков разных стратиграфических уровней позволяет предполагать, что в течение палеогена происходило масштабное изменение климата в сторону общего похолодания, отражавшееся, в частности, в изменении температуры морских водных масс. В ряде работ (Гладенков и др., 2005, 2013, 2018) уже было показано, как в связи с этим менялась термотропная характеристика комплексов моллюсков по разрезу: если эоценовые сообщества региона являлись в целом тепловодными и включали представителей Crassatella, Glycymeris, Arca, Turritella, Tudicula и других родов, то в позднем эоцене и особенно олигоцене появились относительно холодноводные комплексы с Yoldia, Nuculana, Acila, Масоmа, Chlamys, Periploma, Nemocardium, Variamussium, Neptunea и др. С олигоцена (а возможно, с позднего эоцена) в Северо-Тихоокеанской области начал формироваться бореальный тип биоты, который получил развитие в неогене и далее в квартере. На этом фоне происходили более мелкие климатические флуктуации, которые при потеплении вызывали кратковременные миграции относительно тепловодных сообществ с юга на север (от японских широт до чукотских), а при похолодании, наоборот, смещение бореальных представителей с севера на юг. Это объясняет, в частности, нахождение в палеоценовых и эоценовых разрезах Корякского нагорья (особенно в верхнем палеоцене–нижнем эоцене и среднем эоцене) субтропических элементов биоты, в частности моллюсков (представители родов Crassatella, Glycymeris, Cucullaea др.). Масштаб этого явления – “климатических миграций” – мог быть, видимо, весьма значительным. Пространственно представители субтропической биоты перемещались вдоль шельфовой зоны Северо-Восточной Азии от южных широт к северным на сотни (и даже, возможно, на тысячи) километров (Гладенков и др., 2005), фактически до широт Анадырской впадины. Недавно появилось сообщение о находке планктонных фораминифер эоцена севернее Анадырской впадины, к северу от залива Креста Восточной Чукотки – до 66° с.ш. (Бугрова, Гульпа, 2019). Это, пожалуй, самая северная находка карбонатного планктона палеогена на азиатском побережье.
Следует отметить одну особенность олигоценового этапа развития шельфовых биотических сообществ. Как указывалось выше, в это время в морских бассейнах Камчатско-Чукотского региона началось формирование бореального типа фауны, который получил позже развитие в неогене. При этом замещение тепловодных ценозов моллюсков, свойственных в целом палеоцен-среднеэоценовым бассейнам, бореальными сообществами шло, видимо, не резко, а достаточно постепенно, было растянуто во времени, о чем свидетельствует отмечаемое в ряде случаев сонахождение в олигоценовых толщах как относительно тепловодных (Glycymeris, Ostrea, Papyridea, Turritella), так и сравнительно холодноводных элементов биоты (в частности, Yoldia, Масоmа, Chlamys, Periploma, Variamussium, Neptunea и др.). Тем не менее с периодом общего относительного похолодания и появления психросферы связано отступление ареалов сравнительно теплолюбивых бентосных и планктонных сообществ от чукотско-камчатских широт в южном направлении (до Японии), что сопровождалось появлением в камчатско-корякских бассейнах богатых комплексов диатомовых водорослей бореального типа, получивших развитие практически во всей Северо-Тихоокеанской области. Именно с этим связывают накопление кремнистых толщ как на Камчатке и Корякском нагорье (свиты гакхинская, утхолокская, вивентекская, алугинская), так и в районах американского побережья (формация Монтерей). Надо добавить, что имеющиеся данные по палеогеновой флоре этого региона (Ахметьев, 2015) в принципе соответствуют выводам о палеоклиматических флуктуациях, полученным по морским отложениям.
В заключение уместно обратить внимание еще на одно обстоятельство. Кайнозойские морские отложения, развитые в пределах Корякского нагорья, обнаруживают в ряде структур признаки нефтегазоносности, хотя изученность с этой точки зрения данной территории далека от полной. Имеется не очень много сводных работ, посвященных данной проблеме, и среди них выделяется важная сводка В.В. Харахинова и его соавторов (Харахинов и др., 2014) о нефтегазоносных бассейнах Беринговоморского региона (с детальной характеристикой коллекторских свойств пород, флюидоупоров и пр.). В ней, в частности, разбираются особенности строения и формации Анадырской впадины и Хатырского прогиба (Агапитов, 1991). Использование материалов бурения, проведенного на суше и в прилегающей к ней акватории, как и данных сейсмостратиграфии, позволило, например, определить, что в этих достаточно больших и тектонически сложно построенных структурах (с широким развитием разрывных нарушений) имеются более мелкие части с их специфическим набором фаций. В Анадырском бассейне (впадине) намечены два суббассейна: Нижне-Анадырский и Восточно-Анадырский с учетом акваториальной части, в которых нефтегазоносность уже доказана. При этом предполагается, что нефтегазогенерационный потенциал шельфовой части бассейна в два раза превышает таковой наземного его сектора. Считается, что в создании общего углеводородного потенциала бассейна значительная роль принадлежит, в частности, палеогеновым толщам. В Хатырском прогибе выделены три структуры – западный и восточный субпрогибы и разделяющая их центральная часть. В них палеогеновые образования также обнаружили признаки присутствия углеводородов. В частности, автор статьи по результатам геологической съемки в западной части Хатырского прогиба давно обращал внимание на проявление нефтегазоносности в его пределах (Гладенков, 1962). Надо учитывать, что материковая часть Хатырского бассейна является его северной окраиной, занимающей не более 10–15% общей площади бассейна. В основной акваториальной части бассейна, простирающейся под водами Берингова моря, структурный план осадочного выполнения становится более спокойным, что, видимо, может свидетельствовать о благоприятных условиях для нефтегазонакопления.
ЗАКЛЮЧЕНИЕ
Выше в развернутом виде приведены основные результаты проведенного исследования. Кратко их можно изложить следующим образом. Изучение морских отложений среднего эоцена–олигоцена Корякского нагорья Северо-Востока Азии выявило их приуроченность к четырем кайнозойским прогибам и двум впадинам, расположенным в периферийных зонах мезозоид региона. Эти образования сформировались во время морской трансгрессии, охватившей смежные регионы не только Камчатки, но и Сахалина, Японии и Северной Америки. Большую роль при расчленении древних формаций играли комплексы моллюсков, которые позволили осуществить корреляцию стратиграфических горизонтов верхней части палеогена практически во всей Северо-Тихоокеанской области. Анализ этих комплексов выявил их различную термотропность, отразившую палеоклиматические флуктуации (прежде всего масштабные потепления в эоцене и похолодание в конце эоцена–олигоцене), которые вызывали заметные миграции морской палеобиоты в пределах субтропических и бореальных широт Северо-Тихоокеанской области.
В заключение хочется еще раз напомнить о большом вкладе в изучение стратиграфических проблем кайнозоя Корякского нагорья во второй половине прошлого столетия трех замечательных женщин – специалистов-малакологов Л.В. Криштофович, А.Д. Девятиловой и В.И. Волобуевой.
Проведенное обобщение по морскому палеогену различных прогибов и впадин Корякского нагорья с суммированием данных по моллюскам изученных разрезов может оказаться полезным для корректировки региональных стратиграфических схем и составления геологических карт нового поколения.
Благодарности. Автор выражает благодарность С.И. Бордунову за помощь в оценке возраста комплексов фораминифер в разрезах палеогена изученного региона. Отдельно хотелось бы поблагодарить за конструктивные советы рецензентов А.С. Бякова, Ю.Д. Захарова и С.В. Попова.
Источники финансирования. Работа выполнена по теме государственного задания ГИН РАН и при поддержке проекта РФФИ № 19-05-00361.
Список литературы
Агапитов Д.И. Новые данные о стратиграфии неогеновых и палеогеновых отложений Анадырской впадины // Геология и стратиграфия кайнозойских отложений Северо-Западной Пацифики. Владивосток: ДВО АН CCCH, 1991. С. 70–95.
Агапитов Д.И., Митрофанова Л.И. Стратиграфия палеогеновых отложений Хатырской впадины (Чукотка) // Тихоокеанская геология. 1998. Т. 17. № 6. С. 53–60.
Алабушева А.В. Биостратиграфия и фораминиферы палеогеновых отложений Северо-Востока России (Хатырская и Анадырская впадины). Автореф. дисс. … канд. геол.-мин. наук. СПб.: СПбГУ, 1996. 16 с.
Александров А.А. Покровные и чешуйчатые структуры в Корякском нагорье. М.: Наука, 1978. 122 с.
Атлас фауны и флоры палеогеновых отложений Дальнего Востока. Точилинский опорный разрез. Отв. ред. Меннер В.В. М.: Наука, 1984. 335 с. (Труды ГИН АН СССР. Вып. 385).
Афанасьева Н.С., Гладенков Ю.Б., Серова М.Я. Новые данные о расчленении и возрасте третичных отложений центральной части Пенжинского прогиба (Корякское нагорье) // Советская геология. 1974. № 1. С. 53–67.
Ахметьев М.А. Палеоген высокоширотных регионов Сибири и Северо-Востока России: стратиграфия, флора, климат и угленакопление // Стратиграфия. Геол. корреляция. 2015. Т. 23. № 4. С. 76–90.
Беньямовский В.Н., Фрегатова Н.А., Спирина Л.В., Бояринова М.Е., Волобуева В.И., Гладенков Ю.Б., Таривердиева Т.Н. Зоны планктонных и бентосных фораминифер в палеогене Восточной Камчатки // Изв. РАН. Сер. геол. 1992. № 1. С. 110–123.
Бондаренко Г.Е. Тектоника и геодинамическая эволюция мезозоид северного обрамления Тихого океана. Автореф. дисс. … докт. геол.-мин. наук. М.: МГУ им. М.В. Ломоносова, 2004. 46 с.
Бордунов С.И., Дмитриева Т.В., Фрегатова Н.А. Вопросы стратиграфии кайнозоя Западной Камчатки и этапы развития палеосообществ фораминифер // Вестник Моск. ун-та. Сер. 4. Геология. 2020. № 5. С. 17–24.
Бугрова Э.М., Гульпа И.В. Новые сведения по стратиграфии верхнего мела и находка морского палеогена на востоке Чукотского полуострова (по данным изучения фораминифер) // Региональная геология и металлогения. 2019. № 78. С. 70–75.
Бушуев М.И. Геология и угленосность северо-восточной части Корякского хребта. М.: Водотранспорт, 1954. 130 с.
Волобуева В.И., Красный Л.Л. Маастрихт-неогеновые отложения восточной части Корякского нагорья. М.: Наука, 1979. 83 с.
Волобуева В.И., Гладенков Ю.Б., Белая Б.В. Палеоген Северо-Востока СССР // Стратиграфия и палеонтология фанерозоя Северо-Востока СССР. Магадан: СВКНИИ ДВО АН СССР, 1988. С. 118–156.
Волобуева В.И., Гладенков Ю.Б., Беньямовский В.Н., Витухин Д.И., Минюк П.С., Музылев Н.Г., Олейник А.Э., Синельникова В.Н., Соколова З.Ш., Титова Л.В., Фрегатова Н.А., Ширая О.А. Опорный разрез морского палеогена севера Дальнего Востока (п-ов Ильпинский). Ч. 1. Стратиграфия. Магадан: СВКНИИ ДВО РАН, 1994. 64 с.
Геология СССР. Том 31. Камчатская область, Курильские и Командорские острова. Ч. 1. М.: Недра, 1964. 729 с.
Гладенков А.Ю. Детальная стратиграфия и морские экосистемы позднего кайнозоя севера Тихоокеанского региона (по диатомеям). М.: ГЕОС, 2007. 296 с. (Труды ГИН РАН. Вып. 571).
Гладенков А.Ю. Ярусы олигоцена и граница между палеогеном и неогеном на Западной Камчатке – новые данные по диатомеям // Неоген и квартер России: стратиграфия, события и палеогеография. М.: ГЕОС, 2018. С. 23–29.
Гладенков А.Ю., Гладенков Ю.Б. Новые данные по палеонтологической характеристике олигоцена в разрезе п-ова Ильпинский (Северо-Восточная Камчатка) // Стратиграфия. Геол. корреляция. 2007. Т. 15. № 2. С. 119–123.
Гладенков Ю.Б. Нефтепроявления нижнего течения реки Хатырки (Корякское нагорье) // Изв. АН СССР. Сер. геол. 1962. № 9. С. 101–106.
Гладенков Ю.Б. Неоген Камчатки (вопросы биостратиграфии и палеоэкологии). М.: Наука, 1972. 251 с. (Труды ГИН АН СССР. Вып. 214).
Гладенков Ю.Б. Морской верхний кайнозой северных районов. М.: Наука, 1978. 194 с. (Труды ГИН АН СССР. Вып. 313).
Гладенков Ю.Б. Верхнекайнозойские толщи тихоокеанского побережья Северной Америки и корреляция с ними дальневосточных разрезов // Изв. АН СССР. Сер. геол. 1979. № 8. С. 74–84.
Гладенков Ю.Б. Проявление фаз тектогенеза в кайнозое северо-западного обрамления Пацифики // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 4. С. 117–123.
Гладенков Ю.Б. Опорные разрезы палеогена и неогена Восточной Камчатки // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 1. С. 64–80.
Гладенков Ю.Б. Стратиграфические горизонты и проблемы эволюции биотических сообществ морских экосистем в рамках геомериды и биосферы // Тихоокеанская геология. 2018. Т. 37. № 5. С. 16–30.
Гладенков Ю.Б. Стратиграфия кайнозоя восточной части Опухско-Пекульнейского прогиба, Корякское нагорье, Северо-Восток России // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 6. С. 171–180.
Гладенков Ю.Б., Волобуева В.И., Митрофанова Л.И. Третичные толщи Нижнехатырского прогиба (Корякское нагорье) // Изв. АН СССР. Сер. геол. 1985. № 3. С. 77–93.
Гладенков Ю.Б., Синельникова В.Н., Шанцер А.Е., Челебаева А.И., Олейник А.Э., Титова Л.В., Братцева Г.М., Зырянов Е.В., Фрегатова Н.А., Казаков К.Г. Эоцен Западной Камчатки. М.: Наука, 1991. 184 с. (Труды ГИН АН СССР. Вып. 467).
Гладенков Ю.Б., Шанцер А.Е., Челебаева А.И., Синельникова В.Н., Антипов М.П., Беньямовский В.Н., Братцева Г.М., Полянский Б.В., Ступин С.И., Федоров П.И. Нижний палеоген Западной Камчатки (стратиграфия, палеогеография, геологические события). М.: ГЕОС, 1997. 336 с. (Труды ГИН РАН. Вып. 488).
Гладенков Ю.Б., Баженова О.К., Гречин В.И., Маргулис Л.С., Сальников Б.А. Кайнозой Сахалина и его нефтегазоносность. М.: ГЕОС, 2002. 225 с.
Гладенков Ю.Б., Синельникова В.Н., Челебаева А.И., Шанцер А.Е. Биосфера–экосистема–биота в прошлом Земли. Экосистемы кайнозоя Северной Пацифики: эоцен–олигоцен Западной Камчатки и сопредельных районов (К 100-летию со дня рождения академика В.В. Меннера). М.: ГЕОС, 2005. 480 с. (Труды ГИН РАН. Вып. 540).
Гладенков Ю.Б., Синельникова В.Н., Беньямовский В.Н., Фрегатова Н.А. Стратиграфия морского палеоцена и нижнего эоцена Западной Камчатки (палеонтологическая характеристика, палеогеографические обстановки). М.: ГЕОКАРТ, ГЕОС, 2013. 160 с.
Гладенков Ю.Б., Гладенков А.Ю., Бордунов С.И., Чехович В.Д., Александрова Г.Н., Водовозов В.Ю., Гептнер А.Р., Дмитриева Т.В., Покровский Б.Г., Трубихин В.М., Фрегатова Н.А. Опорный разрез кайнозоя Западно-Камчатской структурно-формационной зоны. М.: ГЕОКАРТ, ГЕОС, 2018. 202 с.
Григоренко Ю.Н. Палеоцен-эоценовый граувакковый комплекс тыловых прогибов Притихоокеанской окраины (строение и формирование). СПб.: ВНИГРИ, 2011. 322 с.
Девятилова А.Д., Волобуева В.И. Атлас палеогена и неогена Северо-Востока СССР. М.: Недра, 1981. 219 с.
Егиазаров Б.Х., Дундо О.П., Аникеева Л.И., Русаков И.М., Дегтяренко Ю.П. Геология и полезные ископаемые Корякского нагорья. Л.: Недра, 1965. 343 с.
Криштофович Л.В. Стратиграфия и фауна тигильской толщи западного побережья Камчатки. Л.: Ленгостоптехиздат, 1947. 151 с. (Труды ВНИГРИ. Новая сер. Вып. 23).
Криштофович Л.В. Сопоставление третичных отложений северной части Тихоокеанского кольца кайнозойской складчатости // Материалы совещания по разработке унифицированных стратиграфических схем Сахалина, Камчатки и Командорских островов. М.: Гостоптехиздат, 1961. С. 83–90.
Кузнецов А.П. Фауна донных беспозвоночных прикамчатских вод Тихого океана и северных Курильских островов. М.: Изд-во АН СССР, 1963. 271 с.
Маргулис Л.С., Агапитов Д.Д., Маргулис Л.А., Пылина Л.М., Гладенков А.Ю., Жидкова Л.С., Дмитриева Т.В., Федорова В.А., Фрегатова Н.А. Первые данные о кайнозойском разрезе Чукотского шельфа Берингова моря // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 2. С. 103–114.
Решения рабочих межведомственных региональных стратиграфических совещаний по палеогену и неогену восточных районов России – Камчатки, Корякского нагорья, Сахалина и Курильских островов. Объяснительная записка к стратиграфическим схемам. М.: ГЕОС, 1998. 147 с.
Руженцев С.В., Бялобжеский С.Г., Григорьев В.Н., Казимиров А.Д., Пейве А.А., Соколов С.Д. Тектоника Корякского хребта // Очерки тектоники Корякского нагорья. М.: Наука, 1982. С. 136–189.
Серова М.Я. Фораминиферы и биостратиграфия верхнего палеогена Северной Пацифики. М.: Наука, 2001. 215 с. (Труды ПИН РАН. Т. 279).
Харахинов В.В., Шленкин С.И., Вашкевич А.А., Агапитов Д.Д., Обухов А.Н. Нефтеносные бассейны Беринговоморского региона (итоги нефтегазопоисковых работ 2000–2009 гг.). М.: Научный мир, 2014. 340 с.
Чехович В.Д., Сухов А.Н, Кононов М.В., Паланджян С.А. Геодинамика северо-западного сектора Тихоокеанского подвижного пояса в позднемеловое–палеогеновое время // Геотектоника. 2009. № 4. С. 37–62.
Gladenkov A.Yu. Fossil diatom flora from the marine Paleogene stratigraphic key section of northeast Kamchatka, Russia // Acta Botanica Croatica. 2009. V. 68(2). P. 199–209.
Gladenkov A.Yu. New data on diatoms from the marine Cenozoic section of West Kamchatka at the Kvachina Bay // Paleontol. J. 2019. V. 53. № 8. P. 799–802.
Haq B.U., Hardenbol J., Vail P.R. Chronology of fluctuating sea levels since the Triassic // Science. 1987. V. 235. P. 1156–1167.
Oyama K., Mizuno A., Sakamoyo T. Illustrated handbook of Japanese Paleogene mollusks. Geol. Surv. Japan, 1960. 244 p.
Vail P.R., Hardenbol J. Sea-level changes during the Tertiary // Oceanus. 1979. V. 22. P. 71–79.
Weaver C.E. Paleontology of the marine Tertiary formations of Oregon and Washington. Part III // Univ. Washington Publications in Geology. 1942. V. 5. P. 563–789.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция