

Стратиграфия. Геологическая корреляция, 2021, T. 29, № 1, стр. 59-98
Палеоген Преддонецкой моноклинали и его палинологическая характеристика
А. И. Яковлева 1, *, Г. Н. Александрова 1
1 Геологический институт РАН
Москва, Россия
* E-mail: alina.iakovleva@gmail.com
Поступила в редакцию 27.05.2020
После доработки 11.06.2020
Принята к публикации 30.06.2020
Аннотация
Представлены результаты палинологического изучения палеогеновых и подстилающих их меловых отложений Преддонецкой моноклинали, вскрытых скважиной 1238. Изучение цист динофлагеллат позволило выявить интервалы зон Apectodinium hyperacanthum, Axiodinium augustum, Ochetodinium romanum/Samlandia chlamydophora, а также слоев с Rhombodinium draco–Deflandrea spinulosa в палеогеновой части разреза. Уточнен возраст свит и слоев в регионе: бузиновская свита датируется ранним танетом, вешенская свита – поздним танетом–инициальным ипром; суровикинские и осиновские слои – частью раннего–среднего ипра; венцовские и “полтавские” слои – поздним рюпелем–хаттом. В разрезе скв. 1238 выявлено два крупных стратиграфических перерыва, отвечающих маастрихту–зеландию и верхнему ипру–нижнему рюпелю. Анализ количественных флуктуаций различных групп палиноморф по разрезу позволил реконструировать изменения в обстановках осадконакопления краевой части бассейна Пери-Тетиса в раннем и позднем палеогене: накопление бузиновской, вешенской свит, суровикинских и осиновских слоев происходило в течение длительного трансгрессивного открыто-морского этапа в позднем палеоцене–раннем ипре, тогда как отложения венцовских и “полтавских” слоев формировались в условиях полузамкнутого бассейна в хатте.
ВВЕДЕНИЕ
Принципиально важной задачей современных геологических исследований палеогена является восстановление истории морских бассейнов в течение различных интервалов времени и связанных с этим возможных миграций различных групп фауны и флоры. Очевидно, что наиболее погруженные части палеобассейнов закономерно характеризуются наиболее полными стратиграфической последовательностью и палеонтологической характеристикой, тогда как в краевых частях, как правило, выявляется сильная фациальная изменчивость толщ, зачастую отсутствие или низкая насыщенность отложений палеонтологическими остатками, что достаточно часто приводит к разногласиям в понимании возраста и объемов местных и региональных стратиграфических подразделений.
На юге Русской плиты к числу краевых палеогеновых морских бассейнов относится субрегион восточного обрамления Донбасса (Ахметьев, Беньямовский, 2003). Изначально данная территория относилась к Донецкой синеклизе Днепровско-Донецкой впадины (Леонов, 1961), позднее – к субрегиону Воронежской антеклизы (Семенов, 1965, 1975). В целом Днепровско-Донецкая впадина, Воронежская антеклиза и восточная часть Большого Донбасса представляли собой в палеогене единую морскую акваторию внутри Пери-Тетиса, через которую в течение определенных этапов палеогена осуществлялись субширотные связи с морями на территории современных Белоруссии, Польши, Прибалтики, а также с палеобассейном Северного моря.
В утвержденной в 2000 г. Межведомственным стратиграфическим комитетом России региональной стратиграфической схеме палеогена в субрегионе восточной части Большого Донбасса, с учетом литолого-фациальных и структурных особенностей, были выделены три структурно-фациальные зоны (СФЗ), различающиеся по литологическому строению и мощностям (а иногда и по набору) развитых в них литологических подразделений: (1) Преддонецкая моноклиналь, (2) собственно Восточный Донбасс, (3) южное и восточное обрамление Донбасса (рис. 1) (Постановления…, 2001; Ахметьев, Беньямовский, 2003). Выделенные СФЗ отражают фациальные переходы от более мелководных отложений на севере к более глубоководным на юге, иногда с учетом влияния локальных конседиментационных структур (прогибов и выступов).
Рис. 1.
Географическое расположение изученных разрезов (а) и структурно-фациальное районирование субрегиона восточной части Большого Донбасса (б) (по Ахметьев, Беньямовский, 2003, с изменениями).
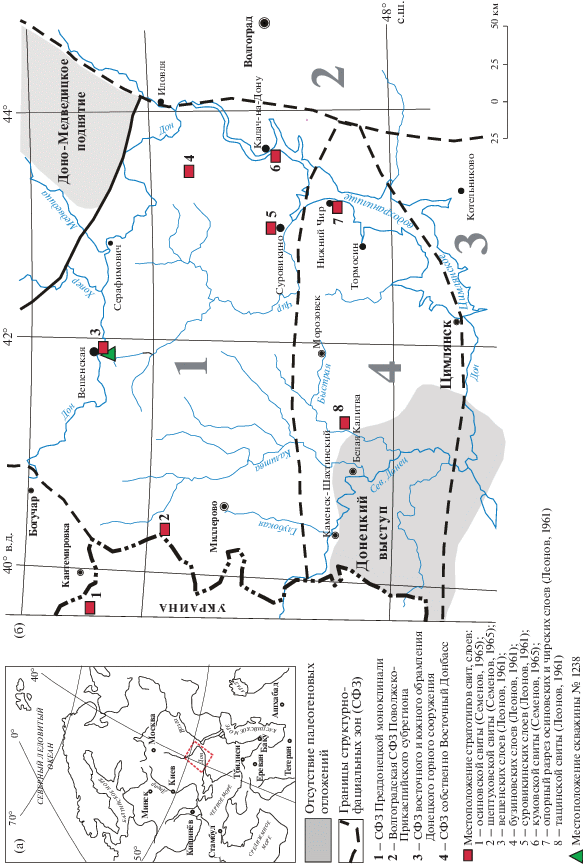
На территории Преддонецкой моноклинали, занимающей самое северное положение в субрегионе и характеризующейся развитием терригенных и терригенно-кремнистых фаций, стратиграфические подразделения были впервые установлены Г.П. Леоновым (1936, 1939, 1961), а позднее В.П. Семеновым (1965) (рис. 2); при этом возраст большинства из них обосновывался данными изучения малакофауны.
Рис. 2.
Стратиграфические схемы расчленения палеогеновых отложений субрегиона восточного обрамления Донбасса.
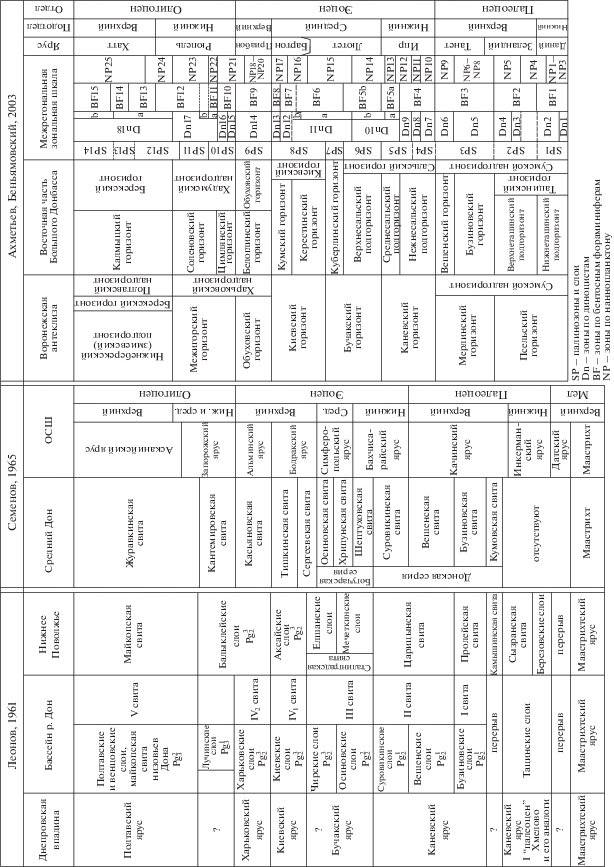
Согласно Ахметьеву и Беньямовскому (2003), развитые на территории Преддонецкой моноклинали горизонты раннего палеогена отвечают одноименным свитам. Так, в стратиграфической схеме 2000 г. для всей территории субрегиона восточной части Большого Донбасса в качестве регионального подразделения палеоцена использован сумской надгоризонт в объеме тацинского, бузиновского и вешенского горизонтов; в эоцене – сальский горизонт (с тремя подгоризонтами), куберлинский, керестинский, кумский, киевский и обуховский горизонты; в олигоцене – хадумский надгоризонт и цимлянский, соленовский, калмыцкий, берекский горизонты (рис. 2) (Постановления…, 2001). Таким образом, в стратиграфической схеме использованы как горизонты Северной Украины (сумской надгоризонт, киевский, обуховский, берекский горизонты), так и подразделения из схемы В.П. Семенова (1975) для Воронежской антеклизы, а также подразделения из схем Нижнего Поволжья, Скифской плиты и Предкавказья. При этом сделано указание (Ахметьев, Беньямовский, 2003, с. 48), что для СФЗ Преддонецкой моноклинали использованы горизонты палеогена Северной Украины. В настоящее время для расчленения палеогеновых отложений региона различными группами специалистов используются местные стратиграфические подразделения, предложенные как для территории Северной Украины, Воронежской антеклизы, так и для Поволжья, Скифской плиты, что приводит к сложности в понимании пространственных и временных взаимоотношений между толщами и установлении их четкого стратиграфического возраста.
Важно отметить, что украинскими и российскими специалистами за последние годы был внесен ряд изменений и уточнений как в стратиграфическую схему северных областей Украины (в частности, был упразднен сумской надгоризонт) (Зосимович и др., 2005, 2013, 2015, 2016, 2017; Зосимович, 2013; Зосимович, Шевченко, 2014, 2015; Орешкина, Яковлева, 2007; Iakovleva, 2015; Мусатов, Рябоконь, 2017), так и в схемы Воронежской антеклизы (Горбаткина, Иосифова, 2004; Шпуль, 2005, 2006, 2007, 2008, 2009, 2010; Бугрова и др., 2016; Мусатов, 2020), Прикаспия и Поволжья (Васильева, 2013, 2019; Мусатов и др., 2014; Мусатов, Богачкин, 2019а, 2019б; Унифицированная…, 2015; Застрожнов и др., 2019).
В последние годы В.Н. Беньямовским (2016, 2017а, 2017б, 2017в) на основании новых и ранее опубликованных данных была предпринята ревизия схемы 2000 г. для субрегиона восточной части Большого Донбасса и субрегиона Воронежской антеклизы, которые были им объединены в единый Воронежско-Придонецкий субрегион. В.Н. Беньямовский предложил собственное структурно-фациальное районирование, а в макете новой региональной схемы использовал все свиты, горизонты и надгоризонты схемы В.П. Семенова. Следует отметить, что в этом варианте региональной стратиграфической схемы субрегиона практически все местные стратиграфические подразделения оказались с перерывами, и ни одно из них не имело согласных границ (Беньямовский, 2017б), однако объяснений этому в публикациях не было дано.
Недостаточная палеонтологическая изученность палеогена юга Русской плиты, использование разными группами специалистов различных местных стратиграфических подразделений (свит, слоев), отсутствие четкого определения стратотипов региоярусов и, в конечном итоге, необходимость актуализации региональной унифицированной схемы палеогена – все это обусловило важность проведения комплексных исследований в регионе Преддонецкой моноклинали и Воронежской антеклизы.
Объектом наших исследований в 2018 г. явилась территория Преддонецкой моноклинали, где мы попытались провести ревизию стратотипов разрезов стратиграфических подразделений палеогена, введенных Г.П. Леоновым в качестве слоев (1936, 1939, 1961) и затем переведенных В.П. Семеновым в ранг свит (1965). При этом лишь некоторые из этих свит имели палеонтологическое обоснование возраста, сделанное по малакофауне. Отдельно следует отметить, что Г.П. Леонов и В.П. Семенов рассматривали датский ярус внутри меловой системы. Поэтому характеристика нижнепалеоценовых пород не включалась в публикации этих авторов.
В ходе проведенных полевых работ нами было установлено, что большинство стратотипических разрезов кумовской, бузиновской, вешенской, суровикинской свит, предложенных Г.П. Леоновым, на сегодняшний день утрачены для непосредственных наблюдений из-за сильной задернованности или залесенности. К сожалению, стратотипы шептуховской и осиновской свит, установленные В.П. Семеновым и расположенные в приграничной с Украиной зоне, в настоящее время не доступны для посещений. Удалось выяснить, что в рассматриваемом регионе палеогеновые отложения обнажаются в изолированных маломощных выходах, в виде “окон” на склонах оврагов и балок, либо вскрываются в неглубоких карьерах, при этом практически нигде не наблюдаются их контакты с ниже- и вышележащими толщами. Исходя из полевых наблюдений и анализа опубликованных материалов, было подтверждено, что палеогеновые отложения в Преддонецкой моноклинали представлены в основном однообразными толщами – песками, песчаниками, алевролитами, опоками с редкими маломощными прослоями и линзами глин, горизонтами галечников и фосфоритов. В результате проведения комплексных литолого-петрографических и микропалеонтологических исследований в более чем 10 естественных выходах палеогена было выявлено отсутствие во всех отобранных образцах фораминифер, диатомовых водорослей и силикофлагеллат, а также цист динофлагеллат, спор и пыльцы высших растений. Палеонтологическая характеристика (в основном по цистам динофлагеллат, диатомовым водорослям) в рассматриваемом регионе получена лишь по нескольким скважинам.
Целью настоящей работы явились детальный комплексный палиностратиграфический анализ палеогеновых отложений разреза скважины № 1238, уточнение возраста местных и региональных стратиграфических подразделений Преддонецкой моноклинали, корреляция этих отложений с последовательностями из смежных регионов, а также восстановление обстановок осадконакопления.
МАТЕРИАЛ И МЕТОДЫ
В рамках настоящего исследования были изучены 63 пробы из разреза скв. 1238, пробуренной в окрестностях станицы Базковской Ростовской области силами АО “Северо-Кавказское ПГО” и ВСЕГЕИ в рамках геолого-съемочных работ на территории листа М-37-XXIV с целью расчленения разреза палеогена (54°89′10′′ с.ш., 77°27′47′′ в.д.) (рис. 1).
Ниже дано полевое литологическое описание (снизу вверх) разреза, вскрытого скв. 1238, и его расчленение в соответствии со схемой Г.П. Леонова (1961) (рис. 3). Литологическое описание выполнено А.С. Застрожновым и С.В. Поповым, расчленение – Г.Н. Александровой.
Рис. 3.
Литологическое строение и стратиграфическое расчленение скв. 1238. Цветовая окраска внутри литологической колонки соответствует цвету пород.
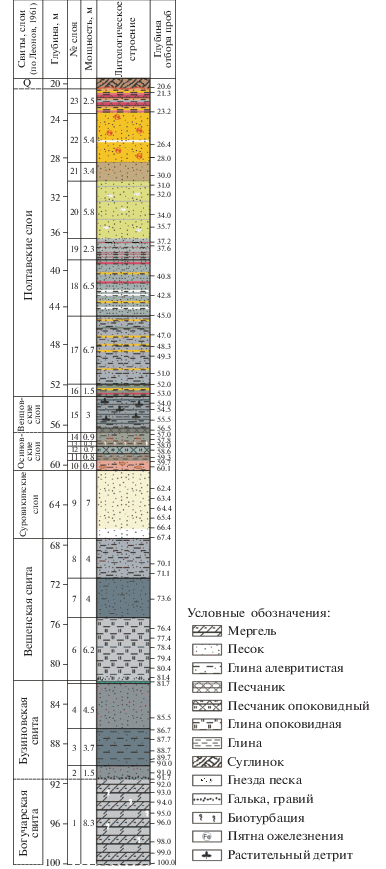
Богучарская свита:
Слой 1, интервал 100–91.7 м. Мергель серый, светло-серый, плотный, массивный, слюдистый, с редкими ходами илоедов, выполненными светлым мергелистым веществом.
Бузиновская свита:
Слой 2, инт. 91.7–90.2 м. С четким контактом залегают пески темно-серые с зеленоватым оттенком, глауконитово-кварцевые, слюдистые, в подошве с мелкой галькой (диаметром 10–15 мм) и крупным гравием фосфоритов, разно- и крупнозернистые, которые вверх по слою постепенно переходят в среднезернистые и глинистые пески.
Слой 3, инт. 90.2–86.5 м. С резким контактом залегают глины темно-серые до черных, плотные, алевритистые, переходящие вверх по слою в глинистые алевриты с зеленоватым оттенком (за счет глауконита), слабокремнистые, слабо биотурбированные, с прослоями (до 0.05 м) песчаников (алевритов) того же состава.
Слой 4, инт. 86.5–82.0 м. Песок темно-серый в основании слоя, средне-мелкозернистый, глинистый с глауконитом, в верхнем метре – средне-крупнозернистый с зернами глауконита.
Слой 5, инт. 82.0–81.7 м. Песчаник крепкий, с характерным изумрудным цветом, окремненный, тонкозернистый до алеврита, глауконитовый.
Вешенская свита:
Слой 6, инт. 81.7–75.5 м. Глина светло-серая, опоковидная, очень легкая, до трепела (5 см), выше – слабокремнистая, слюдистая, с включениями глауконитового вещества. Контакт с нижележащими породами четкий, с базальным горизонтом, сложенным гравийным материалом в виде зерен кварца и фосфоритов (диаметром 1–4 мм).
Слой 7, инт. 75.5–71.5 м. С неясным контактом залегают пески темно-серые с зеленоватым оттенком, глинистые, слабокремнистые, тонко- и мелкозернистые, слюдистые, с участками серого и светло-серого цвета, что придает породе пятнистый облик.
Слой 8, инт. 71.5–67.5 м. Алевриты серые, глинистые, слюдистые, слабокремнистые.
Суровикинские слои:
Слой 9, инт. 67.5–60.5 м. Песок кварцевый, в основании (0.3 м) белый, мелко- и среднезернистый, выше – с желтоватым оттенком, хорошо сортированный. Контакт с подстилающими породами не поднят.
Осиновские слои:
Слой 10, инт. 60.5–59.6 м. Глинисто-алевритовая порода с гнездами песка тонкозернистого, глинистого, с пятнами, прослоями, гнездами oжелезнения; цвет породы – бурый с малиновым, охристым оттенком.
Слой 11, инт. 59.6–58.8 м. Глины коричневые “кофейные”, неизвестковистые, в кровле слоя с гнездами тонкозернистого глауконит-кварцевого серого песка.
Слой 12, инт. 58.8–58.1 м. Песчаник светло-серый, тонкозернистый, опоковидный, легкий, глауконит-кварцевый.
Слой 13, инт. 58.1–57.4 м. Глины коричневые со слабым зеленоватым оттенком, с плитчатой неявной отдельностью, с многочисленными ходами илоедов, выполненными светлым глауконит-кварцевым тонкозернистым песком.
Слой 14, инт. 57.4–56.5 м. Алевролит светло-серый со слабым желтовато-коричневым оттенком, тонкозернистый, слабослюдистый, с пятнами oжелезнения.
Венцовские слои:
Слой 15, инт. 56.5–53.5 м. В подошве песчаник (0.2 м) зеленоватый, среднезернистый и тонкозернистый. Выше – алевролит серый, слюдистый, с линзами и тонкими линзовидными прослойками темно-серых глин, что придает породе слоистый облик, с многочисленными сортированными мелкими обрывками тканей растений. Контакт с нижележащими породами не поднят.
Полтавские слои:
Слой 16, инт. 53.5–52.0. Глина плотная, опесчаненная, пестроцветная: зеленовато-серая с прослоями ожелезнения красного и желтого цвета (интенсивность ожелезнения увеличивается к подошве слоя) и алевролитов светло-серого цвета, что придает породе полосчатый вид. Контакт с нижележащими породами не поднят.
Слой 17, инт. 52.0–45.3 м. Алевролит светло-серый, зеленовато-серый, с прослоями ожелезнения желтоватого цвета, слюдистый. В ожелезненных прослоях встречаются обломки песчаника. В инт. 46.3–46.6 м глина зеленовато-серая.
Слой 18, инт. 45.3–38.8 м. Песок пылеватый, зеленовато-серый, с прослоями ожелезнения красного и желтого цвета. На гл. 44.1–44.2, 42.8, 42.3 м – прослои алевролита светло-серого, мощностью до 5 см. Контакт с нижележащими породами размыт.
Слой 19, инт. 38.8–36.5 м. Песок мелкозернистый, зеленовато-светло-серый, с тонкими прослоями светло-серого алевролита, ожелезнения. Последние придают породе полосчатый облик (9–10 прослоев на 1 м керна). Контакт с нижележащими породами выражен в увеличении количества ожелезненных прослоев.
Слой 20, инт. 36.5–30.7 м. С резким контактом залегает песок мелкозернистый, желто-зеленого цвета, ожелезненный, с включениями песка белого сцементированного, с тонкими прослоями светло-серого глинистого вещества.
Слой 21, инт. 30.7–28.5 м. С резким контактом залегает песок мелкозернистый, светло-коричневый, массивной текстуры.
Слой 22, инт. 28.5–23.1 м. С резким контактом залегает песок желтый, мелкозернистый, слюдистый, слабосцементированный, с неясно слоистой текстурой, с ожелезнением в виде натеков красного цвета. На гл. 26.2–26.1 м – прослой песка белого цвета.
Слой 23, инт. 23.1–20.6 м. С резким контактом залегает пачка чередования прослоев (2–5 см) глин плотных, малиновых, серых, желтых, алевритов охристых и песчаников мелкозернистых, некрепких, красных – кора выветривания по нижележащим отложениям.
Выше разрез перекрывается твердыми бурыми суглинками четвертичного возраста мощностью 20 м.
Обработку палинологических образцов проводили в соответствии со стандартной методикой, принятой в лаборатории палеофлористики ГИН РАН: (1) воздействие на образцы 10%-ной соляной кислотой (HCl) для удаления карбонатов; (2) воздействие горячим раствором пирофосфата натрия (Na4P2O7 ⋅ 10H2O) для дисперсии глинистого материала, а затем отмывка каждые 2 часа с целью удаления глинистых частиц; (3) центрифугирование образцов в тяжелой жидкости (K2CdI4) с плотностью 2.25 с целью отделения органической фракции от более тяжелых минеральных частиц; (4) воздействие 70%-ной плавиковой кислотой (HF) для растворения кремнистых компонентов; (5) воздействие 10%-ной соляной кислотой для удаления флюоро-силикатных гелей; (6) отмывка образца в дистиллированной воде и заливка глицерином. Просеивание мацерата через сита не проводилось.
Таксономия диноцист соответствует индексу DINOFLAJ3 (Williams et al., 2017). Количественное палинологическое изучение материала проводили в 2 этапа: (1) подсчет в каждом продуктивном образце минимум 200–250 морских и наземных палиноморф (цисты динофлагеллат, акритархи, празинофиты, пыльца покрытосеменных и голосеменных растений, внутренние камеры фораминифер); (2) дальнейший подсчет минимум 200 диноцист, после чего препарат просматривался до конца с целью обнаружения редко встречаемых таксонов. Палеоэкологическая интерпретация количественных флуктуаций в палеогеновых палинокомплексах основана на ряде работ, прежде всего (Brinkhuis, 1994; см. табл. 1 в Iakovleva, Heilmann-Clausen, 2020). Для интерпретации возможных палеообстановок цисты динофлагеллат были объединены в 17 групп морфологически близких таксонов: (1) Deflandrea-группа (Deflandrea, Lejeunecysta, Lentinia, Senegalinium); (2) Wetzelielloids; (3) Phthanoperidinium; (4) Fromea fragilis; (5) Lingulodinium machaerophorum; (6) Areoligera-группа (Areoligera, Glaphyrocysta, Emmetrocysta, Adnatosphaeridium, Membranilarnacia); (7) Thalassiphora; (8) Ochetodinium romanum; (9) Elytrocysta brevis; (10) Cribroperidinium; (11) Carpatella rossica; (12) Cordosphaeridium-группа (Cordosphaeridium, Fibrocysta); (13) Operculodinium; (14) Diphyes/Dapsilidinium; (15) Spiniferites-группа (Achomosphaera, Hystrichosphaeropsis, Hystrichostrogylon, Spiniferella, Spiniferites, Rottnestia); (16) Impagidinium; (17) остальные гониаулакоидные.
Хроностратиграфический возраст дается в соответствии с данными, приведенными в работе (Ogg et al., 2016).
Коллекция палинологических образцов хранится в лаборатории палеофлористики Геологического института РАН (г. Москва).
РЕЗУЛЬТАТЫ
Основной задачей настоящих исследований был палинологический анализ палеогеновых отложений, вскрытых скв. 1238, стратиграфические и количественные результаты которого представлены на рис. 4–6. Палинологические данные из подстилающих верхнемеловых отложений в основании пробуренной скважины даны нами в кратком виде. Наиболее характерные таксоны цист динофлагеллат из мел-палеогеновых отложений представлены в табл. I–XI .
Рис. 4.
Стратиграфическое распределение видов диноцист в палеогеновой части разреза скв. 1238.
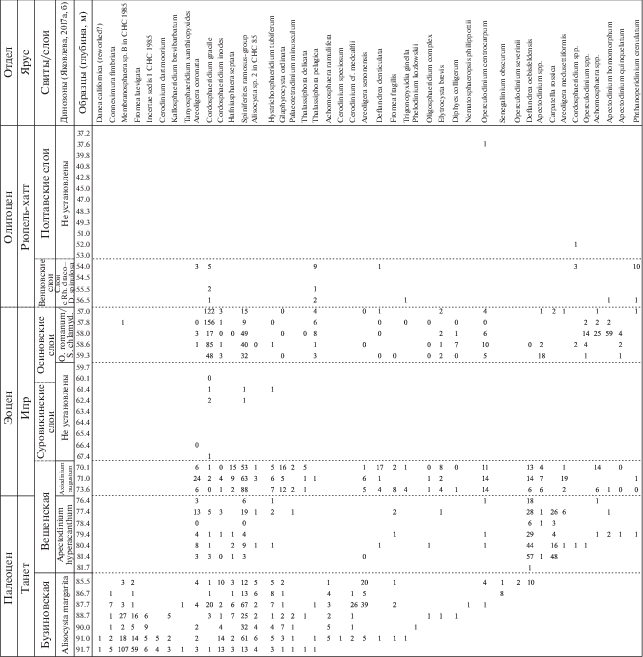
Рис. 4.
Продолжение
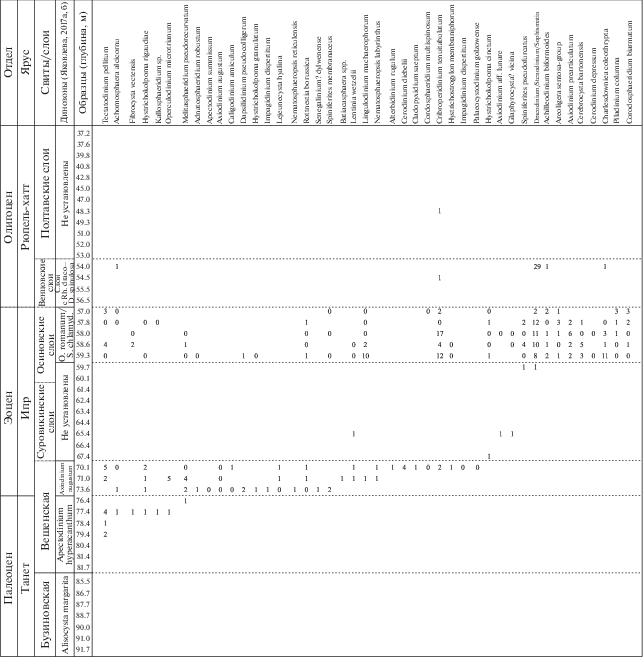
Рис. 4.
Продолжение
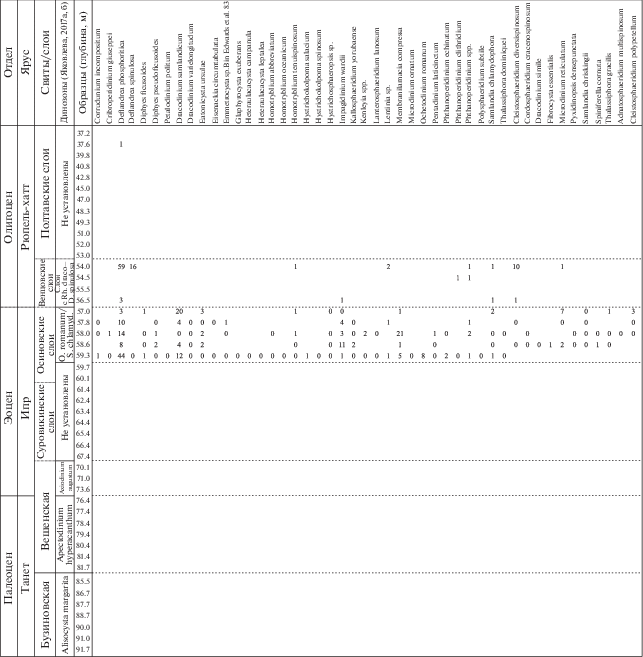
Рис. 4.
Окончание
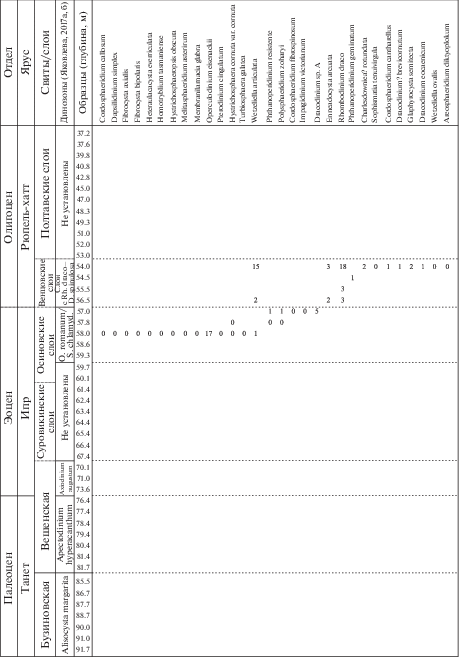
Рис. 5.
Количественное распределение основных групп палиноморф в палеогеновой части разреза скв. 1238.
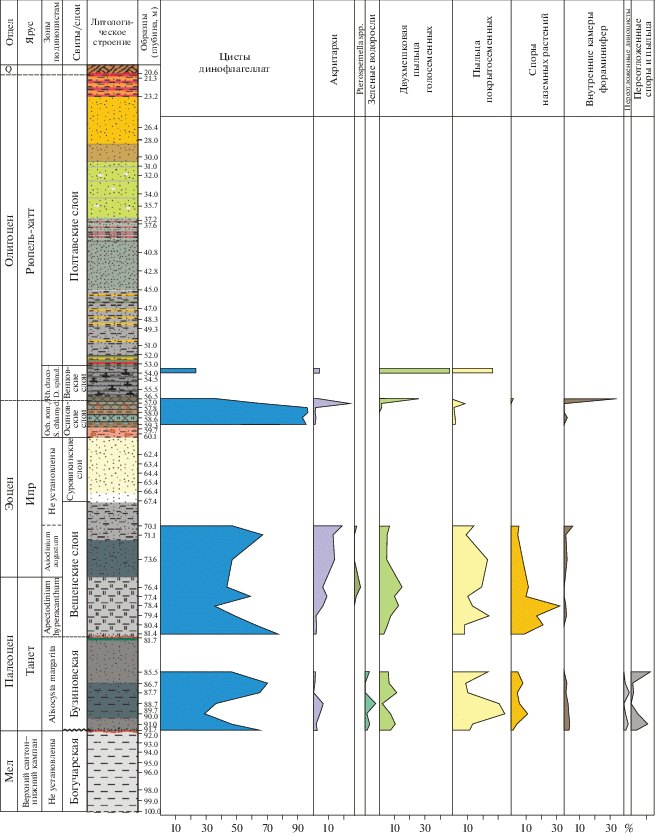
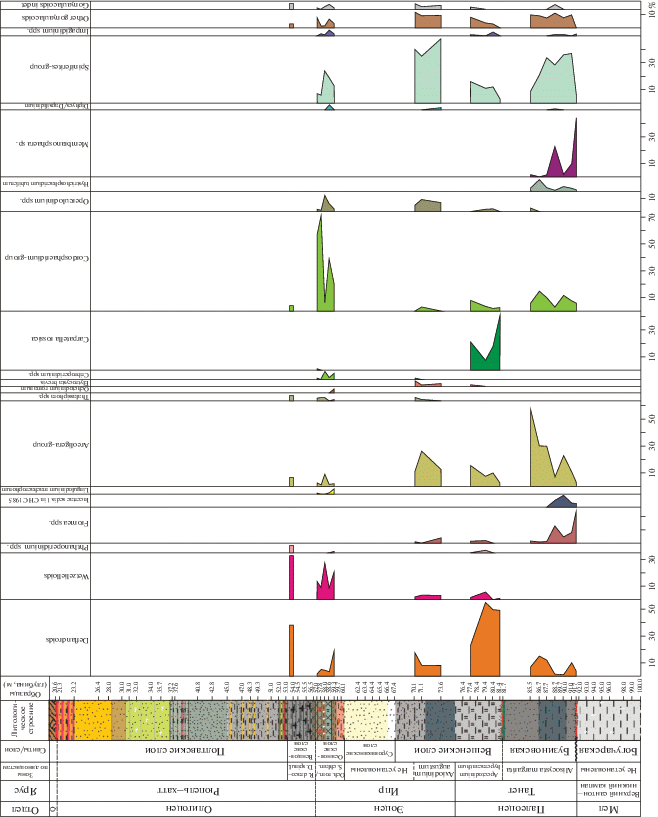
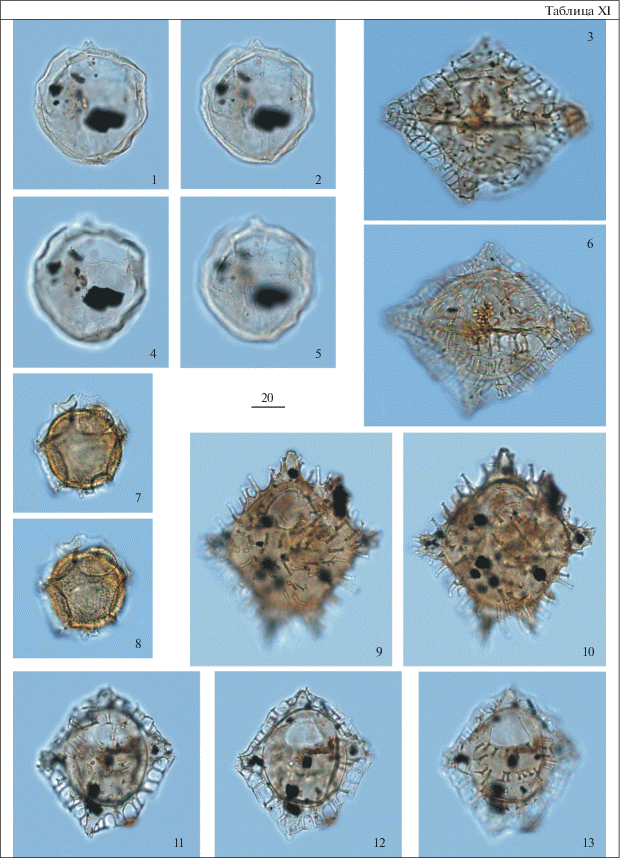
Таблица I . Диноцисты из сантон-кампанских отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1 – Odontochitina costata (Alberti) Clark et Verdier, гл. 92 м; 2 – Manumiella sp., гл. 92 м; 3, 6, 9, 12 – Cribroperidinium wilsonii subsp. trabeculosum Slimani et Louwye, гл. 92 м; 4, 5 – Laciniadinium firmum (Harland) Morgan, гл. 92 м; 7, 10 – Xenascus ceratioides (Deflandre) Lentin et Williams, гл. 92 м; 8, 11 – Isabelidinium microarmum (McIntyre) Lentin et Willams, 8 – гл. 96 м, 11 – гл. 92 м; 13 – Chatangiella vnigrii (Vozzhennikova) Lentin et Willams, гл. 93 м; 14, 15 – Spinidinium echinoideum (Cookson et Eiselanck) Lentin et Willams, гл. 92 м.
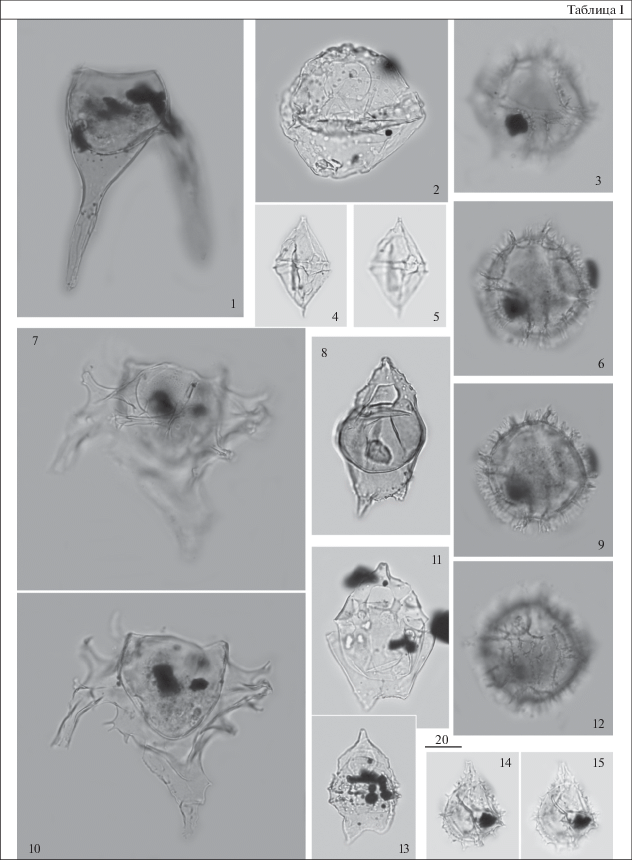
Таблица II. Диноцисты из сантон-кампанских отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 2 – Hystrichosphaeropsis quasicribrata (Wetzel) Gocht, гл. 92 м; 3, 4 – Chatangiella vnigrii (Vozzhennikova) Lentin et Willams, гл. 96 м; 5 – ?Alterbidinium sp., гл. 92 м; 6 – Subtilisphaera perlucida (Alberti) Jain et Millepied, гл. 96 м; 7, 12 ‒ Isabelidinium sp., гл. 92 м; 8 – Isabelidinium pellucidum (Deflandre et Cookson) Lentin et Willams, гл. 92 м; 9, 10 – Apteodinium deflandrei (Clarke et Verdier) Lucas-Clarck, гл. 92 м; 11, 14 – Isabelidinium acuminatum (Cookson et Eisenack) Stover et Evitt, 11 – гл. 92 м, 14 – гл. 93 м; 13 – Isabelidinium microarmum (McIntyre) Lentin et Willams, гл. 93 м; 15, 16 – Alisocysta sp. 2 sensu Heilmann-Clausen, 1985, гл. 90 м; 17, 18 – Conneximura fimbriata (Morgenroth) May, гл. 86.7 м.
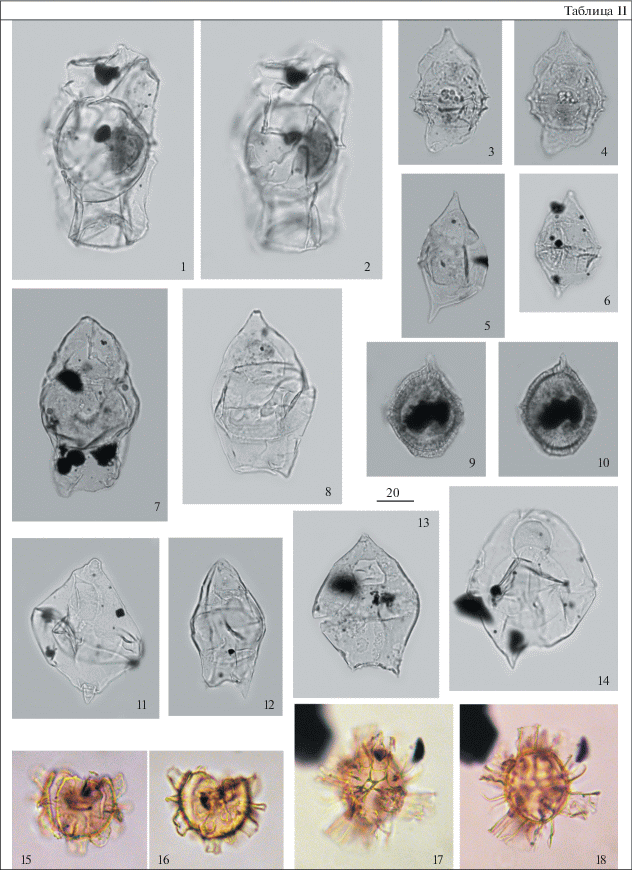
Таблица III. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 5 – Axiodinium augustum (Harland) Williams et al., гл. 70.1 м; 2, 6 – Dracodinium samlandicum (Eisenack) Costa et Downie, гл. 59.3 м; 3, 4 – Samlandia chlamydophora Eisenack, гл. 59.3 м; 7, 8 – Cerebrocysta bartonensis Bujak, гл. 59.3 м; 9, 10, 13 – Impagidinium wardii Iakovleva et Heilmann-Clausen, гл. 58.0 м; 11 – Dracodinium brevicornutum (Heilmann-Clausen) Williams et al., гл. 54.0 м; 12 – Dracodinium sp., археопиль латиепелиформный, гл. 58.0 м; 14, 15 – Phthanoperidinium sp., гл. 58 м; 16, 17 – Membranilarnacia compressa Bujak, гл. 58 м; 18, 19 – Spiniferites ramosus (Ehrenberg) Mantell, гл. 59.3 м; 20 – Ochetodinium romanum Damassa, гл. 59.3 м.
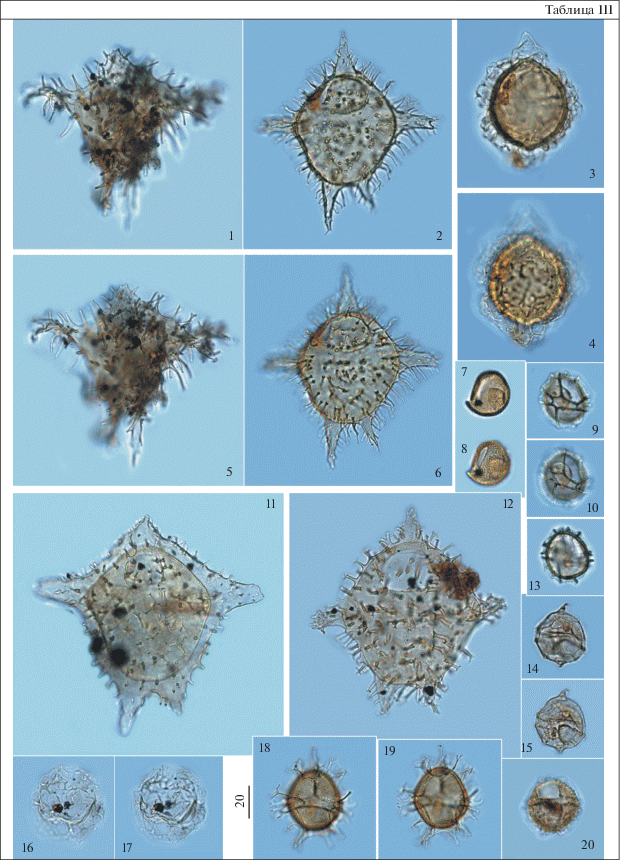
Таблица IV. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 4, 7 – Dracodinium samlandicum (Eisenack) Costa et Downie, гл. 58 м; 2, 3 – Fibrocysta bipolaris (Cookson et Eisenack) Stover et Evitt, гл. 59.3 м; 5 – Fibrocysta essentialis (De Coninck) Brinkhuis et Zachariasse, гл. 59.3 м; 6 – Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, гл. 59.3 м; 8, 9, 11, 12 – Hystrichokolpoma spinosum Wilson, гл. 59.3 м; 10, 13, 14 – Impagidinium wardii Iakovleva et Heilmann-Clausen, гл. 57.8 м; 15 – Heteraulacacysta leptalea Eaton, гл. 59.3 м; 16, 17 – Deflandrea denticulata Alberti, гл. 70.1 м; 18 – Deflandrea cf. denticulata Alberti, гл. 59.3 м.

Таблица V. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 2, 6 – Axiodinium prearticulatum Williams et al., гл. 58.0 м; 3–5 – Impagidinium wardii Iakovleva et Heilmann-Clausen, гл. 58.0 м; 7, 8 – Dracodinium sp. A, археопиль латиепелиформный, гл. 58.0 м; 9, 10 – Membranilarnacia compressa Bujak, гл. 59.3 м; 11, 12 – Phthanoperidinium sp., гл. 58.0 м; 13, 14 – Eatonicysta ursulae (Morgenroth) Stover et Evitt, гл. 58.0 м; 15 – Diphyes pseudoficusoides Bujak, гл. 58.0 м.
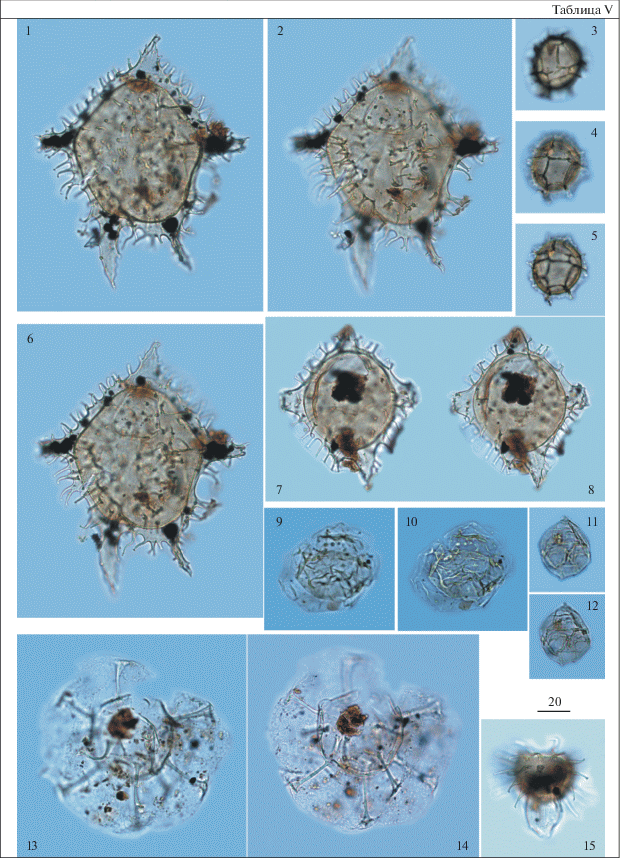
Таблица VI. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 4, 9 – Axiodinium prearticulatum Williams et al., гл. 59.3 м; 2, 3 – Dracodinium sp. A, гл. 57.8 м; 5 – Axiodinium prearticulatum Williams et al., гл. 58.0 м; 6, 7 – Carpatella rossica Iakovleva, гл. 81.4 м; 8 – Dracodinium sp. A, гл. 58.0 м; 9 – Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, гл. 59.3 м; 10, 11 – Diphyes ficusoides Islam, гл. 58.0 м; 12, 13 – Kallosphaeridium yorubaense Jan du Chêne et Adediran, гл. 58.0 м; 14 – Fibrocysta vectensis (Eaton) Stover et Evitt, гл. 58.0 м; 15 – Membranilarnacia compressa Bujak, гл. 58.0 м.

Таблица VII. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1–3 – Cordosphaeridium gracile (Eisenack) Davey et Williams, гл. 57.8 м; 4–7 – Ochetodinium romanum Damassa, гл. 59.3 м; 8 – Axiodinium prearticulatum Williams et al., гл. 58.0 м; 9, 12 – Samlandia chriskingii Iakovleva et Heilmann-Clausen, гл. 59.3 м; 10, 11 – Diphyes pseudoficusoides Bujak, гл. 58.0 м; 13, 14 – Hystrichosphaeropsis sp., гл. 58.0 м.
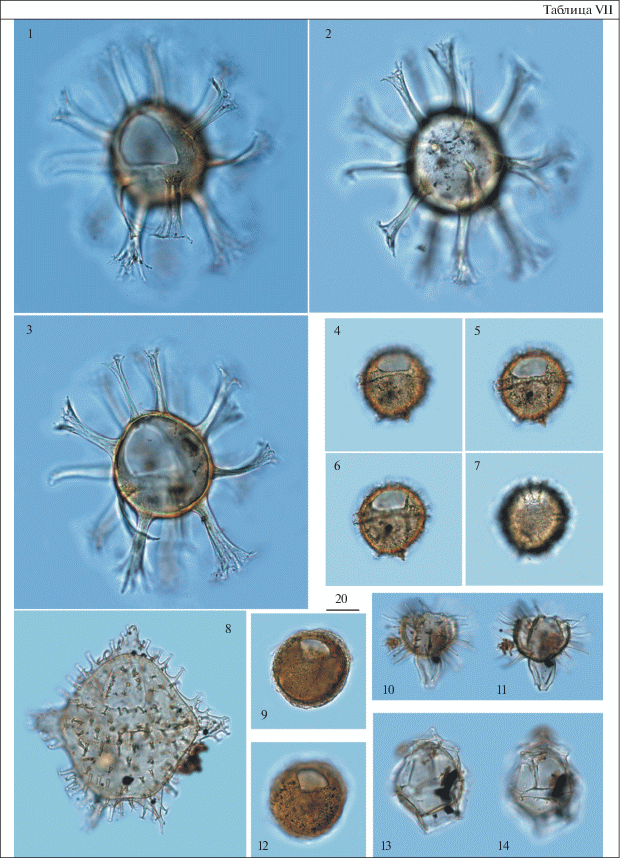
Таблица VIII. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 4 – Thalassiphora dominiquei Iakovleva et Heilmann-Clausen, гл. 59.3 м; 2 – Cribroperidinium tenuitabulatum (Gerlach) Helenes, гл. 59.3 м; 3, 7 – Rhombodinium draco Gocht, гл. 54.0 м; 5, 6 – Eisenackia circumtabulata Drugg, гл. 59.3 м; 8, 11 – Piladinium columna (Michoux) Williams et al., гл. 59.3 м; 9 – Dracodinium eocaenicum (Agelopoulos) Williams et al., гл. 54.0 м; 10 – Dracodinium sp. A, гл. 57.8 м; 12, 13 – Ochetodinium romanum Damassa, гл. 59.3 м; 14 – Rhombodinium draco Gocht, оперкулюм свободен, гл. 54.0 м.
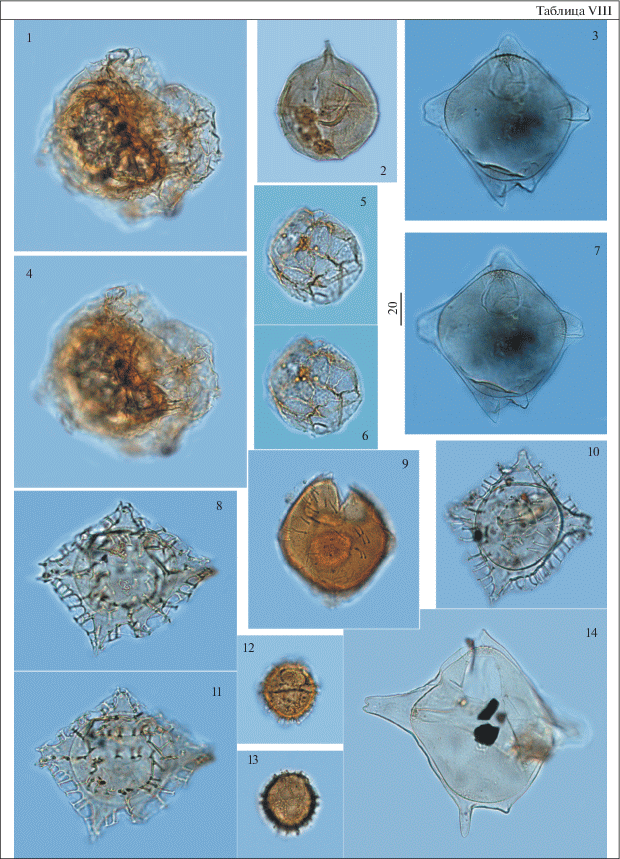
Таблица IX. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 2, 4, 5 – Cordosphaeridium inodes (Klumpp) Eisenack, гл. 57.8 м; 3, 6, 11, 15 – Achilleodinium biformoides (Eisenack) Eaton, гл. 59.3 м; 7–10 – Hystrichokolpoma cinctum Klumpp, гл. 59.3 м; 12–14 – Hystrichokolpoma cf. cinctum Klumpp, гл. 57.8 м.
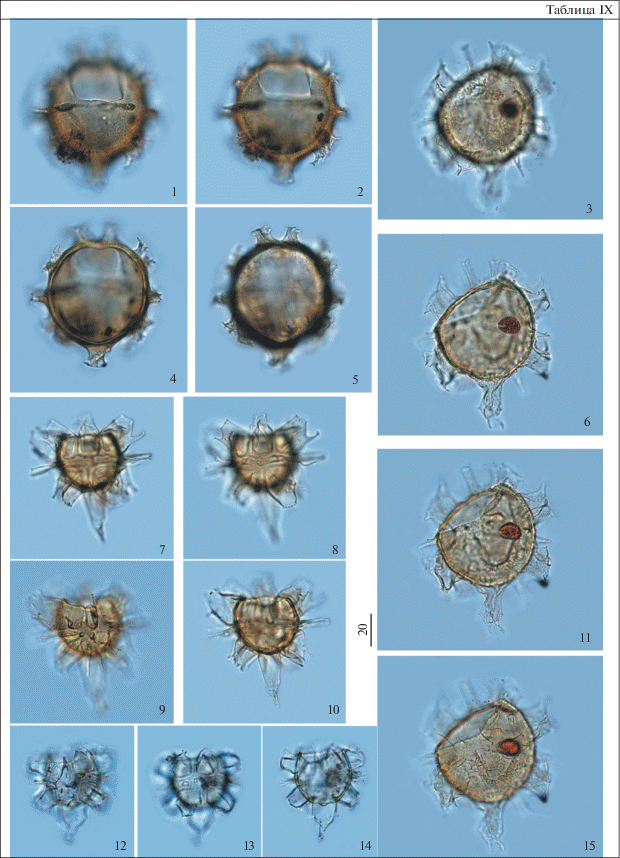
Таблица X. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 5, 9, 13 – Hystrichosphaeropsis sp., гл. 57.8 м; 2, 6, 10 – Piladinium columna (Michoux) Williams et al., гл. 59.3 м; 3, 7, 11 – Hystrichosphaeropsis sp., гл. 57.0 м; 4, 8, 12 – Areoligera sentosa-group, гл. 58 м; 14 – Tectatodinium pellitum Wall, гл. 59.3 м; 15 – Phthanoperidinium sp., гл. 58.0 м; 16 – Piladinium columna (Michoux) Williams et al., гл. 59.3 м; 17, 18 – Deflandrea oebisfeldensis Alberti, гл. 70.1 м.
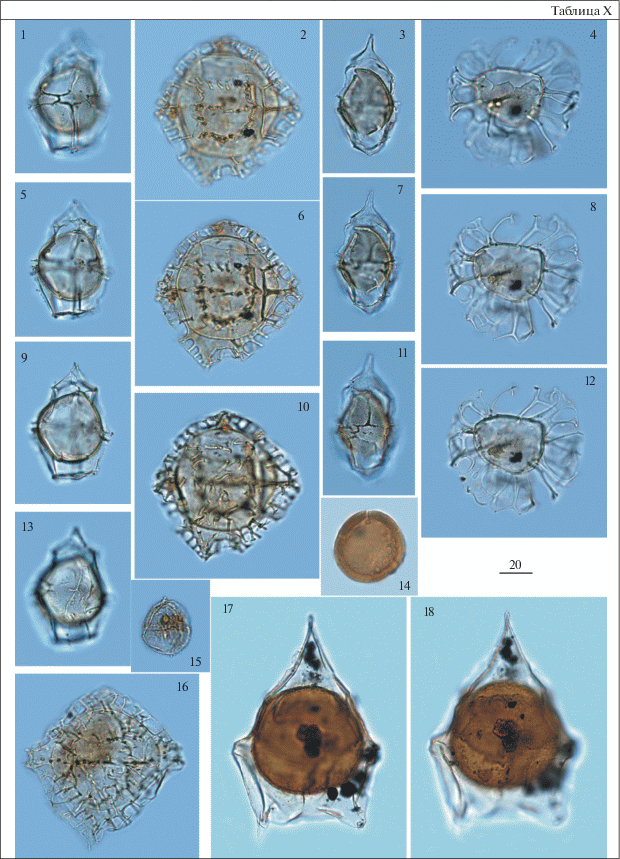
Таблица XI. Диноцисты из палеогеновых отложений разреза скв. 1238. Увеличение показано в микрометрах (мкм). 1, 2, 4, 5 – Cribroperidinium tenuitabulatum (Gerlach) Helenes, гл. 58.0 м; 3, 6 – Charlesdowniea coleothrypta (Williams et Downie) Lentin et Vozzhennikova, гл. 59.3 м; 7, 8 – Pentadinium laticinctum Gerlach, гл. 59.3 м; 9, 10 – Dracodinium samlandicum (Eisenack) Costa et Downie, гл. 58.0 м; 11–13 – Piladinium columna (Michoux) Williams et al., гл. 58.0 м.
В пачке 1, отнесенной к верхнемеловой богучарской свите, во всех образцах установлены количественно представительные ассоциации палиноморф, но содержащие диноцисты преимущественно плохой сохранности, единичные споры папоротников и пыльцу покрытосеменных, а также единичные празинофиты (Palambages morulosa), акритархи (Paralecaniella indentata, Michystridium sp.) и внутренние камеры фораминифер.
К сожалению, из 47 палинологических проб из палеогеновой части разреза лишь в 22 пробах были выявлены количественно репрезентативные палинологические комплексы; остальные же либо содержали небольшое количество палиноморф, либо вообще оказались стерильными. Палинологические комплексы палеоцена–низов эоцена отличаются доминированием цист динофлагеллат (30–90%), существенным (до ~30%) присутствием пыльцы покрытосеменных растений и спор наземных растений и единичным всплеском в содержании внутренних камер фораминифер. Лишь в низах средней части разреза скважины, отнесенной к венцовским слоям олигоцена, комплекс отличается доминированием (~40%) двухмешковой пыльцы хвойных растений.
Комплекс диноцист из богучарской свиты характеризуется доминированием видов Trythirodinium suspectum и Trythirodinium cf. rhomboideum, а также разнообразием и частой встречаемостью родов Chatangiella (Ch. vnigrii, Ch. bondarenkoi, Ch. cassidea, Ch. victoriense), Isabelidinium (I. acuminatum, I. belfastense, I. microarmum, I. magnum) и Spiniferites. В комплексе отмечаются также виды Acantaulax wilsonii, Apteodinium deflandrei, Connexmura fimbriata, Circulodinium distinctum, Alisogymnium euclaense, Dinogymnium acuminatum, D. westralinum, Raetiadinium truncigerum, Exochosphaeridium bifidum, Manumiella cretacea, Odontochitina costata, Laciniadinium rhombiforme, L. arcticum, Lanternosphaeridium lanosum, Spinidinium echinoideum, S. uncinatum, Fromea chytra, Xenascus gochtii, Form 2 sensu Lebedeva. В целом данный комплекс диноцист по таксономическому составу близок к кампанским комплексам Полярного Предуралья, Усть-Енисейского района и Омского прогиба Западной Сибири (Лебедева, 2005, 2007; Lebedeva, 2006; Лебедева и др., 2013). Основываясь на присутствии в комплексе видов Odontochitina costata, Chatangiella vnigrii, Manumiella cretacea, Lanternosphaeridium lanosum и Form 2 sensu Lebedeva, а также отсутствии стратиграфически важного вида Surculosphaeridium? longifurcatum, указанный комплекс может быть предварительно сопоставлен со слоями с Odontochitina costata, установленными в Нижнем Поволжье в верхней части мезинолапшиновской свиты и нижней части пудовкинской свиты и имеющими возраст, близкий к переходу от сантона к кампану (Александрова и др., 2012). Хотя следует отметить, что в Нижнем Поволжье массовая встречаемость рода Trythirodinium была установлена на более низком стратиграфическом уровне – в верхнесантонской части слоев с Surculosphaeridium? longifurcatum–Heterosphaeridium difficile. Исходя из этого, отложения в интервале 100–91.7 м в скв. 1238 предположительно отнесены к верхнему сантону–нижнему кампану.
В количественном отношении комплекс диноцист из богучарской свиты отличается преобладанием перидиниоидных родов Trythirodinium, Chatangiella и Isabelidinium, что может свидетельствовать о накоплении отложений в прибрежных обстановках с существенным привносом питательных веществ.
В низах палеогеновой части разреза, отнесенных к бузиновской свите (глубина 91.7–85.5 м), выявлен диноцистовый комплекс, в котором присутствуют стратиграфически важные виды Deflandrea oebisfeldensis, Deflandrea denticulata и Alisocysta sp. 2 sensu Heilmann-Clausen, а также представлены виды Areoligera coronata, Areoligera senonensis, Palaeotetradinium minusculum, Trigonopyxidia ginella, Tanyosphaeridium xanthiopyxides, Hystrichosphaeridium tubuferum, Cerodinium cf. medcalfii, Cerodinium speciosum и другие. Присутствие вида Deflandrea denticulata позволяет отнести отложения бузиновской свиты к интервалу диноцистовой зоны Alisocysta margarita из шкал Западной Сибири (Яковлева, Александрова, 2013), Западной Европы (Powell, 1992) и к интервалу слоев с Alisocysta sp. 2 Поволжья (Александрова, 2013) и, соответственно, датировать вмещающие отложения ранним танетом (~58.5–56.5 млн лет).
Палинологический комплекс из бузиновской свиты характеризуется доминированием цист динофлагеллат (30–70%); количество акритарх и зеленых водорослей не превышает 5%, пыльца покрытосеменных растений варьирует от 10 до 30%, споры наземных растений не превышают 10%. Комплекс цист динофлагеллат, в свою очередь, отличается безусловным доминированием гониаулакоидных групп, среди которых выделяются группы Areoligera (5–55%), Membranosphaera (45–0%), Spiniferites (10–35%), Fromea (до 20%) и Cordosphaeridium (до 15%); наблюдаются единичные находки Impagidinium; перидиниоидные диноцисты, представленные группой deflandroids, составляют не более 15% от комплекса.
Второй диноцистовый комплекс выявлен в нижней части вешенской свиты (глубина 81.7–76.4 м) по появлению видов Apectodinium homomorphum и Apectodinium quinquelatum и отнесен к интервалу позднетанетской диноцистовой зоны Apectodinium hyperacanthum из зональных шкал Поволжья (Александрова, 2013), Западной Сибири (Яковлева, Александрова, 2013) и Западной Европы (Powell, 1992). В комплексе появляются виды Phthanoperidinium crenulatum, Tectatodinium pellitum, Areoligera medusettiformis, Carpatella rossica, Hystrichokolpoma rigaudiae, Melitasphaeridium pseudorecurvatum и другие. Стратиграфический возраст нижней части вешенской свиты – поздний танет (~56.5–55.8 млн лет).
Палинологический комплекс нижней части вешенской свиты характеризуется существенным преобладанием цист динофлагеллат (35–80%), акритархи составляют до 10%, споры наземных растений достигают 30% от комплекса, пыльца покрытосеменных – 7–20%, двухмешковая пыльца голосеменных – в пределах 12%. Диноцистовый комплекс в этом интервале отличается доминированием перидиниоидной группы deflandroids (до 55%) и пиками в содержании гониаулакоидного вида Carpatella rossica (до 40%); группы Areoligera и Spiniferites не превышают 15% каждая; Impagidinium представлен первыми процентами. Доминирование цист дефляндровых и достаточно заметное количество континентальных палиноморф могут, вероятно, свидетельствовать о накоплении нижней части вешенской свиты в морских мелководных обстановках пониженной солености, с привносом питательных веществ с затопляемой суши. К концу интервала соотношения групп диноцист (относительное уменьшение доли дефляндровых и чуть заметное увеличение количества групп Areoligera, Cordosphaeridium и Spiniferites) свидетельствуют о постепенной нормализации солености воды.
В верхней части вешенской свиты (глубина 73.6–70.1 м), по появлению вида Axiodinium augustum, выявлен третий диноцистовый комплекс, соответствующий зоне Axiodinium augustum восточного Пери-Тетиса (Яковлева, 2017). Ассоциация диноцист в этом интервале достаточно разнообразная: здесь отмечены, впервые для данного разреза, виды Apectodinium summissum, Adnatosphaeridium robustum, Caligodinium amiculum, Dapsilidinium pseudocolligerum, Hystrichokolpoma granulatum, Impagidinium dispertitum и др. Интервал между первым и последним появлением вида Axiodinium augustum характеризует зону Apectodinium (=Axiodinium) augustum Западной Сибири (Яковлева, Александрова, 2013), Южной Англии (Powell et al., 1996), подзону D5A Северо-Западной Европы (Costa, Manum, 1988), зону Viborg-6 в Дании (Heilmann-Clausen, 1988) и подзону P6b в Северном море (Mudge, Bujak, 1994).
Известно, что во многих регионах мира основание зоны Apectodinium (=Axiodinium) augustum совпадает с началом события изотопной аномалии углерода (CIE – carbon isotope excursion), ассоциирующимся с коротким геологическим событием (~200 000 лет) – палеоцен-эоценовым термическим максимумом (ПЭТМ), а верхняя граница зоны оказывается чуть моложе окончания CIE и, следовательно, чуть моложе события ПЭТМ (Frieling et al., 2014; Shcherbinina et al., 2016). Исходя из того, что интервал зоны Ax. augustum большей частью соответствует событию ПЭТМ (~55.8–55.6 млн лет) и связанному с ним глобальному событию CIE, мы предполагаем, что возраст верхней части вешенской свиты – инициальный ипр (~55.8–55.6 млн лет).
Палинологический комплекс внутри этого интервала характеризуется доминированием цист динофлагеллат (50–70%), относительным увеличением доли акритарх (до 25%). Пыльца покрытосеменных составляет от 25 до 10%, споры наземных растений и двухмешковая пыльца хвойных не превышают 5–10% каждая. Внутри комплекса цист динофлагеллат доминируют гониаулакоидные группы Spiniferites (~35–45%), Areoligera (10–25%), Operculodinium (до 10%). Перидиниоидные диноцисты группы дефляндровых составляют не более 15%, а Apectodinium spp. представлены первыми процентами. Подобные соотношения морских и континентальных палиноморф, а также доминирование Spiniferites и Areoligera в ассоциациях диноцист свидетельствуют об открытых нормально-соленых морских условиях и трансгрессивном этапе в развитии бассейна.
Внутри интервала скважины 67.4–59.7 м, отнесенного к суровикинским слоям, палиноморфы практически отсутствуют. Здесь отмечены только единичные находки видов Areoligera coronata, Corodosphaeridium gracile, Spiniferites ramosus и Hystrichosphaeridium tubiferum, имеющих широкий стратиграфический интервал распространения.
Четвертый диноцистовый комплекс выявлен внутри осиновских слоев в интервале глубин 59.3–57.0 м. По присутствию видов Ochetodinium romanum, Piladinium columna и Samlandia chlamydophora этот таксономически разнообразный (более 85 таксонов) и количественно представительный комплекс соответствует диноцистовой зоне Ochetodinium romanum/Samlandia chlamydophora из шкалы восточного Пери-Тетиса (Яковлева, 2017). Внутри интервала также отмечены виды Samlandia chriskingii, Areoligera sentosa-group, Heteralaucacysta everriculata, Homotryblium tasmaniense, Wetzeliella articulata, Hystrichosphaeropsis obscura, Membranilarnacia glabra, Thalassiphora gracilis, Turbiosphaera galatea и другие. Стратиграфический возраст этой части осиновских слоев – средний ипр (~51.3–50.8 млн лет).
Палинологический комплекс в интервале 59.3–57.0 м практически полностью представлен цистами динофлагеллат (более 95%), внутренние камеры фораминифер и пыльца покрытосеменных составляют первые проценты. Только в верхах осиновских слоев на глубине 57.0 м наблюдаются заметные изменения в количестве акритарх (~24%). Диноцистовый комплекс, прежде всего, характеризуется существенными изменениями в содержании группы Cordosphaeridium – от доминирования (20–71%) до незначительного присутствия (4–6%); перидиниоидные диноцисты (deflandroids и wetzelielloids) составляют здесь до 19 и 20% соответственно; содержание группы Spiniferites колеблется от 6 до 19%, и она практически отсутствует в верхах пачки (глубина 57.0 м); Impagidinium (прежде всего Impagidinium wardii) достигает ~5% комплекса. Подобные характеристики палинологического комплекса могут свидетельствовать об очередном, среднеипрском, трансгрессивном цикле в истории бассейна и накоплении осиновских слоев в достаточно мелководных условиях с нормальной соленостью, но, учитывая заметное присутствие перидиниодных групп в диноцистовом комплексе, можно предположить привнос питательных веществ с затопляемой суши.
Пятый диноцистовый комплекс выявлен в венцовских слоях в интервале 56.5–54.0 м и отнесен нами к слоям с Rhombodinium draco и Deflandrea spinulosa. Он характеризуется присутствием видов Rhombodinium draco, Wetzeliella ovalis, Glaphyrocysta semitecta, Areosphaeridium diktyoplokum, Charlesdowniea? rotundata и Phthanoperidinium geminatum, первая совместная встречаемость которых в восточном Пери-Тетисе наблюдается в конце лютета и бартоне на уровне диноцистовой зоны Rhombodinium draco (Яковлева, 2017). Однако сравнение этого комплекса со средне- и верхнеэоценовыми комплексами диноцист, установленными в разрезах Северной Украины, Воронежской антеклизы, Скифской плиты, показывает их существенные различия (Запорожец, 1998, 2001; Орешкина, Яковлева, 2007; Андреева-Григорович, Шевченко, 2007; Андреева-Григорович и др., 2011; Александрова и др., 2011; Васильева, 2013, 2019; Iakovleva, 2015). Так, преобладание Rhombodinium draco, Deflandrea spinulosa, частая встречаемость Thalassiphora pelagica и Wetzeliella articulata позволяют сопоставлять комплекс диноцист из венцовских слоев скв. 1238 с подзоной Rhombodinium draco зоны Chiropteridium partispinatum, установленной ранее в переходном интервале от соленовской к калмыцкой свите в Ергенях в разрезе скв. 45А (инт. 130–94 м; Запорожец, 1998), а также в верхнеморозкинской подсвите–нижней части баталпашинской свиты в разрезе по р. Белая в Предкавказье (Запорожец, Ахметьев, 2017), где вмещающие отложения датируются поздним рюпелем–ранним хаттом (Запорожец, 1998; Запорожец, Ахметьев, 2017; Застрожнов и др., 2019). Следует отметить, что Н.И. Запорожец указывает массовое количество переотложенных диноцист палеогена и даже мезозоя внутри диноцистового комплекса, соответствующего подзоне Rhombodinium draco в различных частях Пери-Тетиса. Вполне возможно, что и в диноцистовом комплексе венцовских слоев часть таксонов может быть переотложена. Выявленный комплекс диноцист из венцовских слоев близок и к комплексу диноцист из пограничного интервала рюпеля и хатта в Датском бассейне (Śliwińska et al., 2012).
В большей части полтавских слоев, представленных песками с подчиненными прослоями глин, выявлены крайне немногочисленные ассоциации палиноморф; лишь палинологическая ассоциация с глубины 54.0 м (слой алевролитов) оказалась количественно представительной: здесь отмечена заметная смена доминирующих групп палиноморф по сравнению с нижележащими породами: преобладание двухмешковой пыльцы хвойных (46%), приблизительно равное содержание пыльцы покрытосеменных (26%) и цист динофлагеллат (23%) и совсем малое (4%) содержание акритарх. Комплекс диноцист при этом характеризуется абсолютным доминированием перидиниоидных групп – deflandroids (38%), wetzelielloids (33%) и Phthanoperidinium (5%); гониаулакоидные диноцисты представлены в малых количествах. Подобные соотношения групп палиноморф позволяют предполагать накопление низов полтавских слоев в условиях прибрежных обстановок пониженной солености с существенным привносом питательных веществ.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Изначально Г.П. Леонов (1936, 1939) расчленил палеогеновые отложения бассейна Среднего Дона на пять свит, имевших цифровое наименование (I–V; рис. 2), а позднее либо заменил эти свиты вспомогательными местными стратиграфическими подразделениями (слоями), либо выделил слои внутри своих свит (Леонов, 1961). Так, свита I стала соответствовать бузиновским слоям, сопоставлявшимся с пролейской свитой Нижнего Поволжья; свита II была подразделена на вешенские и суровикинские слои, соответствовавшие подсвитам царицынской свиты Нижнего Поволжья; в свите III были выделены осиновские и чирские слои, сопоставлявшиеся со сталинградской свитой (мечеткинскими и елшанскими слоями соответственно) Нижнего Поволжья; наконец, свита IV была разделена на киевские и харьковские слои из унифицированной схемы палеогена Северной Украины, которые Г.П. Леонов сопоставлял с аксайскими слоями и нижней частью балыклейских слоев Поволжья. Выше по разрезу Г.П. Леонов выделил лучинские слои, которые опять же сопоставлялись им с верхней частью балыклейских слоев Поволжья. Самая верхняя часть палеогеновой толщи – свита V была отнесена к венцовским слоям, сопоставлявшимся с майкопской свитой Нижнего Поволжья, и так называемым “полтавским” слоям, соответствовавшим, с долей условности, полтавской серии Украины.
Для расчленения палеоцена и нижнего эоцена В.П. Семеновым (1965) было принято большинство вспомогательных местных стратиграфических подразделений, введенных ранее Г.П. Леоновым (1961), таких как бузиновские, вешенские и суровикинские слои, но при этом они были переведены этим автором в ранг свит (рис. 2). В.П. Семенов также ввел две новые свиты – кумовскую (самые низы палеогеновой толщи) и шептуховскую. Кумовская, бузиновская, вешенская и суровикинская свиты были объединены В.П. Семеновым в донскую серию (нижний палеоцен–начало нижнего эоцена). В свою очередь, впервые установленная В.П. Семеновым шептуховская свита (верхняя часть нижнего эоцена), а также среднеэоценовые хрипуновская и осиновская свиты объединились в богучарскую серию. Что касается верхнего эоцена, то здесь В.П. Семенов ввел для Среднего Дона сергеевскую, тишкинскую и касьяновскую свиты, а для олигоцена – кантемировскую и журавкинскую свиты. Возраст указанных свит основывался в основном на сравнении выявленных комплексов малакофауны с комплексами из Приднепровья (Украина). Позднее в схеме 1975 г. для Воронежской антеклизы все указанные свиты были приведены в ранге региональных горизонтов (Семенов, 1975). Следует отметить, что впервые установленные В.П. Семеновым в нижнем и среднем эоцене свиты в значительной мере имеют очень близкий литологический состав и схожие комплексы фауны, а в своем наиболее типичном виде распространены на территории, расположенной западнее Преддонецкой моноклинали, которая, начиная с принятия унифицированной схемы 2000 г., уже отнесена к СФЗ Воронежской антеклизы, тогда как для субрегиона восточной части Большого Донбасса сохранены только бузиновский и вешенский горизонты из схемы Семенова (1975).
В изученном нами разрезе скв. 1238 верхнесантонские–нижнекампанские отложения перекрываются морскими верхнепалеоценовыми (нижний танет) отложениями бузиновской свиты. Исходя из этого, можно сделать вывод о существовании очень крупного (порядка 13 млн лет) перерыва в осадконакоплении, соответствующего маастрихту–зеландию. Отсутствие датско-зеландских отложений в районе бурения скв. 1238 свидетельствует, скорее всего, о достаточно приподнятом положении этой части Русской плиты в раннем палеоцене.
Бузиновская свита представлена в Преддонецкой моноклинали толщей (8–16 м) мелкозернистых зеленовато-серых глауконит-кварцевых песков с непостоянными прослоями кремнистого песчаника; в средней части толщи присутствует горизонт неокатанных желваков фосфорита, ниже которого иногда отмечается второй фосфоритовый горизонт небольшой мощности (Леонов, 1961). В своем типовом виде свита распространена лишь в пределах Преддонецкой моноклинали, а западнее устья р. Тихая и на территории Воронежской антеклизы эта свита не установлена.
Исследование окрестностей села Верхняя Бузиновка, являющегося стратотипическим районом для бузиновских слоев (Леонов, 1961), показало, что здесь имеется только один естественный выход палеогена, в котором наблюдается лишь верхняя часть бузиновской свиты с видимой мощностью 6.2 м, а также ее контакт с вышележащей вешенской свитой (рис. 7а, 7б). Бузиновская свита представлена пачкой песков тонко- и мелкозернистых, кварцево-глауконитовых, с горизонтом фосфоритов (0.5 м) в нижней части. В кровле свиты отмечена биотурбация в виде окремнелых ходов илоедов (диаметром до 1.5 см). Выше по неровной трансгрессивной границе залегает вешенская свита, в базальной части сложенная опокой темно-серой, массивной, плотной, дающей отчетливую бровку в стенке.
Рис. 7.
Фотографии разрезов бузиновской свиты Преддонецкой моноклинали. (а) – обнажение верхней части бузиновской свиты в районе с. Верхняя Бузиновка; (б) – расчистка в верхней части бузиновской свиты в районе с. Верхняя Бузиновка; (в) – обнажение нижней части бузиновской свиты в районе хутора Бирюковский; (г) – фосфоритовый горизонт и контакт бузиновской и вешенской свит в районе хутора Бирюковский.

Западнее, в районе хутора Бирюковский, имеется одно из немногих наиболее полных обнажений нижней части бузиновской свиты (рис. 7в, 7г). В этом разрезе отчетливо видно ритмично-цикличное строение низов бузиновской свиты: она представлена толщей песков кварцево-глауконитовых, зеленовато-серых, разнозернистых, с преобладанием средне- и мелкозернистых, с редкими включениями гравия кварца, трансгрессивно залегающих на писчем меле. Подошва свиты неровная, базальный горизонт (8–10 см) представлен темными глинами с мелкими фосфоритами, окатанной уплощенной галькой, переотложенными белемнитами. Пески заключают в себе прослои песчаников серых, слабослюдистых, крепких. В видимой кровле свиты залегает пачка желваковых фосфоритов с многочисленными включениями (диаметром до 10 мм) кварца и черных алевролитов, к кровле переходящая в плиту пластовых фосфоритов и содержащая обломки раковин моллюсков. Мощность бузиновской свиты составляет 5 м. Исходя из данных Г.П. Леонова о том, что фосфоритовые горизонты наблюдаются в средней части бузиновской свиты, нами предполагается размыв верхней части свиты в разрезе хутора Бирюковский. Бузиновская свита трансгрессивно, со следами размыва в виде многочисленного гравия кварца, гнезд песка, перекрывается вешенской свитой. Близкое строение имеет бузиновская свита и в районе с. Кумовка, где, однако, крайне плохо обнажена.
В схеме 2000 г. бузиновский горизонт содержит комплекс моллюсков с Lyostrea reussi, бентосные фораминиферы зоны BF3, спорово-пыльцевой комплекс SP3 и отвечает нижнему танету (Ахметьев, Беньямовский, 2003), что теперь подтверждается и нашими данными изучения диноцист. Основываясь на новых палинологических данных, свидетельствующих о соответствии бузиновской свиты интервалу раннетанетской диноцистовой зоны Alisocysta margarita, указанная свита может быть сопоставлена с саратовской свитой Ульяновско-Саратовского прогиба и Прикаспийской синеклизы, а также с нижнетанетской новоузенской свитой Прикаспия (Унифицированная…, 2015). По данным палинологического изучения скв. 1238, бузиновская свита накапливалась во время трансгрессии, в условиях внутренней неритической зоны с нормальной соленостью.
Вешенская свита мощностью до 15 м, перекрывающая бузиновскую, сложена плотными песчаными опоковидными или трепеловидными глинами, часто в той или иной степени фациально замещающимися глинистыми, рыхлыми, иногда опоковидными песчаниками или плотными глинистыми песками (Леонов, 1961). В районе южного окончания Доно-Медведицкого вала глины переходят в неоднородные кварцево-глауконитовые пески с прослоями аналогичных песчаников. Свита трансгрессивно залегает на различных горизонтах нижележащих отложений, а ее подошва характеризуется появлением гравийного и галечникового материала. Вешенская свита имеет более широкое распространение, нежели бузиновская свита, – на западе до г. Богучара, на севере – до г. Новохоперска. В схеме 2000 г. вешенский горизонт содержит комплекс моллюсков с Arctica (=Cyprina) morrisi, бентосные фораминиферы зоны BF3, спорово-пыльцевой комплекс SP3 и отвечает верхнему танету (Ахметьев, Беньямовский, 2003).
Выходы вешенской свиты широко распространены в районе Преддонецкой моноклинали и выявлены нами в районе сел Кумовка, Верхняя Бузиновка, Суровикино, хуторов Бирюковский и Калининский, а также в стратотипической местности в окрестностях станицы Базковской. Во всех перечисленных обнажениях присутствуют только отдельные части свиты, полного разреза наблюдать, к сожалению, не удалось (рис. 8а, 8б). В целом наши наблюдения подтверждают данные Г.П. Леонова: формирование вешенской свиты происходило на фоне продолжающейся трансгрессии в открытых нормально-морских условиях.
Рис. 8.
Фотографии разрезов вешенской свиты, суровикинских и “полтавских” слоев Преддонецкой моноклинали. (а) – обнажение верхней части вешенской свиты в районе с. Кумовка; (б) – обнажение нижней части вешенской свиты в районе станицы Базковской; (в) – контакт вешенской свиты и суровикинских слоев в районе с. Суровикино; (г) – горизонт с ходами и норами декапод в суровикинских слоях в районе с. Суровикино; (д) – биотурбация в “полтавских” слоях в районе станицы Базковской; (е) – пологая косая слоистость в “полтавских” слоях в районе станицы Базковской.

По полученным палинологическим данным, в нижней части вешенской свиты скв. 1238 (инт. 81.7–76.4 м) выявлен комплекс диноцист, соответствующий интервалу позднетанетской зоны Apectodinium hyperacanthum. Это действительно доказывает позднетанетский возраст вмещающих отложений, но лишь для нижней части свиты. Исходя из данных изучения цист динофлагеллат, нижняя часть вешенской свиты сопоставляется нами с верхнетанетскими отложениями камышинской свиты Ульяновско-Саратовского прогиба (Александрова, 2013; Унифицированная…, 2015) и нальчикской свиты Северного Кавказа (Shcherbinina et al., 2016). Что касается верхней части свиты, вскрытой скв. 1238, то эти отложения соответствуют интервалу диноцистовой зоны Axiodinium (=Apectodinium) augustum инициального эоцена. Эта часть вешенской свиты может быть сопоставлена в пределах Пери-Тетиса с одновозрастными отложениями нальчикской свиты Северного Кавказа (Shcherbinina et al., 2016), бостандыкской свиты Северного и Центрального Прикаспия (Васильева, Мусатов, 2010а; Vasilyeva, Musatov, 2012), пролейской свиты Волгоградской области (Александрова и др., 2011), песчано-глинистой толщи Восточного Крыма (Александрова, Щербинина, 2011) и калининской свиты Поволжья (балка Дюпа; Aleksandrova, Radionova, 2006). Более того, согласно данным Т.В. Орешкиной и др. (неопубликованные данные), в скважине, пробуренной на крайнем западе Преддонецкой моноклинали (окрестности г. Богучар Воронежской области) (рис. 1), вскрыт более высокий интервал вешенской свиты, соответствующий интервалу диноцистовой зоны Stenodinium meckelfeldense раннеипрского возраста (~54.8–53.4 млн лет). Таким образом, основываясь на данных изучения диноцист, общий возраст вешенской свиты может быть оценен как поздний танет–часть раннего ипра (~56.5–53.4 млн лет).
Суровикинские слои мощностью 8–10 м, перекрывающие в изучаемом регионе вешенскую свиту, представлены толщей кварцевых песков светло-серых и желтоватых, средне-мелкозернистых, мучнистых, слабоглауконитовых в нижних горизонтах, с невыдержанными прослоями и линзами плотных кремнистых песчаников (Леонов, 1961). Начиная с 1950-х годов, суровикинские слои относились к нижнему эоцену и сопоставлялись с нижними частями бахчисарайской свиты Крыма и каневской свиты Украины, а также с верхнецарицынскими слоями Волгоградского Поволжья (Леонов, 1939, 1961; Семенов, 1965, 1975; Геология…, 1969; Стратиграфическая…, 1987).
В ходе недавних полевых работ полные выходы суровикинских слоев не были обнаружены; лишь в разрезе у с. Суровикино удалось зафиксировать их контакт с вешенской свитой (рис. 8в), а у с. Кумовка был выявлен изолированный выход, скорее всего, средней части суровикинских слоев. Следует отметить, что в разрезе у с. Суровикино присутствует горизонт с многочисленными ходами и норами декапод с выраженной бугорчатой скульптурой, весьма близких к ихновиду Ophiomorpha nodosa, описанному ранее из калининской свиты раннего ипра Поволжья (Янин, Беньямовский, 2018) (рис. 8г). Основываясь на литологическом строении суровикинских слоев, можно предположить, что их формирование происходило в мелководных палеообстановках.
К сожалению, в скв. 1238 суровикинские слои оказались охарактеризованы буквально единичными диноцистами, которые не позволяют уточнить возраст вмещающих отложений. В разрезе скважины они занимают промежуточное положение между верхнетанетской–нижнеипрской вешенской свитой и осиновскими слоями с комплексом диноцист зоны Ochetodinium romanum/Samlandia chlamydophora среднего ипра, что в целом подтверждает раннеипрский возраст суровикинских слоев.
Вышележащие осиновские слои в пределах Преддонецкой моноклинали залегают трансгрессивно, что подтверждается и нашими палинологическими данными. В регионе основание этих слоев характеризуется наличием грубозернистых пород и присутствием одного или двух прослоев кремнистого песчаника; выше по разрезу эти породы сменяются пачкой коричневатых глин или тонкозернистых песков (4–5 м), которые, в свою очередь, перекрываются толщей неоднородных зеленовато-серых, уплотненных, тонкозернистых, кварц-глауконитовых песков мощностью до 8–10 м, сменяющихся выше по разрезу среднезернистыми кварцевыми песками (Леонов, 1961). Палеонтологически осиновские слои не охарактеризованы.
Отметим, что в ходе полевых исследований в районе Преддонецкой СФЗ естественные выходы осиновских слоев не обнаружены. Исходя из литологического строения этих слоев, в разрезе скв. 1238 в интервале 60.5–57.0 м вскрыта только нижняя часть слоев, а более высокие горизонты, вероятно, размыты. Полученные нами палинологические данные в разрезе скв. 1238 (глубина 60.5–57.4 м; зона Ochetodinium romanum/Samlandia chlamydophora) свидетельствуют о среднеипрском возрасте осиновских слоев.
Следует напомнить, что, начиная с середины прошлого века (Семенов, 1965; Геология…, 1969; Стратиграфия…, 1975) и до настоящего времени (Государственная…, 2009), принято считать, что осиновские слои (=осиновская свита) с размывом, фиксируемым слоем галечника или конгломерата, залегают на суровикинских слоях, представляют собой нижнюю часть бучакского горизонта и, соответственно, имеют среднеэоценовый (лютетский) возраст. Необходимо подчеркнуть, что за стратотип осиновской свиты В.П. Семеновым был принят разрез у слободы Осиновой на р. Айдар в Луганской области Украины (рис. 1), где, по данным Г.П. Леонова (1961), осиновские слои сокращаются в мощности и приобретают более грубозернистый состав, а вышележащие чирские слои либо отсутствуют из-за размыва, либо плохо обнажены. Как следствие, выделение этих стратонов в бассейне р. Айдар было условным и требовало подтверждения дополнительными данными. В типовом разрезе в овраге Липовском (правый берег р. Дон, в районе устья р. Чир) Г.П. Леонов сопоставлял осиновские слои с нижними частями сталинградской свиты, мечеткинских слоев Поволжья и черкесской свиты Ростовского свода, Предкавказской части Скифской плиты и, с долей условности, с низами бучакского яруса Украины (Леонов, 1961), что и было зафиксировано в Региональной схеме палеогеновых отложений (Геология…, 1969).
Согласно современным данным, черкесская свита в опорном разрезе Северного Кавказа по р. Хеу имеет ипрский (раннеэоценовый) возраст; внутри этого возрастного интервала, помимо прочего, выявлена и диноцистовая зона Ochetodinium romanum/Samlandia chlamydophora среднего ипра (Яковлева, 2017; Shcherbinina et al., 2020). Таким образом, осиновские слои действительно соответствуют части черкесской свиты Северного Кавказа, а также части бостандыкской свиты Прикаспия (Васильева, Мусатов, 2010а, 2010б; Vasilyeva, Musatov, 2012; Васильева, 2013; Мусатов и др., 2014), симферопольской свиты Крыма (King et al., 2018), кульсаринской свиты Северного Устюрта (Iakovleva, Heilmann-Clausen, 2020), части нижнесальского подгоризонта схемы 2000 г. (Ахметьев, Беньямовский, 2003) и верхней части пролейского горизонта Поволжья (Унифицированная…, 2015) (рис. 2).
Многолетние дискуссии вызывают возраст и генезис песчаной толщи, залегающей в верхней части палеогена юга России. Большинство исследователей склонны сопоставлять ее с так называемым полтавским ярусом схемы Н.А. Соколова (1893) и датировать олигоценом–ранним миоценом. В силу отсутствия в этой толще палеонтологических находок, ее олигоцен-раннемиоценовый возраст определялся на основании сопоставлений с подразделениями палеогена в бассейне р. Днепр, Скифской плиты или Нижнего Поволжья (Леонов, 1936, 1939, 1961; Семенов, 1965, 1975; Семенов, Столяров, 1988; Стратиграфическая…, 1987; Воронина и др., 1988; Попов и др., 1993; Горбаткина, Иосифова, 2004 и др.).
Эти отложения мощностью до 30 м в Преддонецкой моноклинали трансгрессивно, с размывом залегают на различных горизонтах мела и палеогена. Достоверные морские олигоценовые отложения, относимые к майкопской серии, распространены на границе структурно-фациальных зон Преддонецкой моноклинали и восточного обрамления Донбасса – в районе Цимлянского гидроузла. Исходя из географического положения и литологического строения, переходные фации от майкопских к полтавским отложениям, установленные в нижнем течении р. Чир, отнесены к венцовским слоям (Леонов, 1961). Они сложены глинами темноокрашенными (серые до черных), гумусированными, с многочисленными тонкими линзовидными прослоями тонкозернистого кварцевого песка, мощностью до 4 м.
Севернее развиты континентальные образования, объединяемые в “полтавскую серию” (Леонов, 1961; Семенов, 1965), которая представлена толщей песков кварцевых, разнозернистых, пестроокрашенных (белых, желтых, красных, малиновых, серых, зеленоватых), сыпучих, иногда слабоглинистых, уплотненных; здесь же отмечаются маломощные прослои и линзы пестрых глин (1–4 м), преимущественно в нижней части, а также стяжения железистых песчаников. Предполагалось, что линией фациального раздела между майкопскими и полтавскими отложениями являлись Донбасс и Доно-Медведицкий вал.
Полученные количественно представительные палинологические данные из венцовских слоев в скв. 1238 (инт. 57.0–54.0 м), где нами установлены слои с Rhombodinium draco–Deflandea spinulosa позднего рюпеля–раннего хатта, наконец вносят некоторую определенность как в вопрос стратиграфического положения нижней границы верхних горизонтов палеогена Преддонецкой моноклинали, так и в вопросы восстановления палеогеографической обстановки формирования этих толщ. Установлено, что венцовские слои в скв. 1238 с размывом залегают на нижнеэоценовых осиновских слоях, что подразумевает наличие крупного перерыва в осадконакоплении, соответствующего верхнему ипру–большей части рюпеля (~18 млн лет). К сожалению, вышележащие “полтавские” слои, в том числе слагающие возвышенные участки Преддонецкой моноклинали, в скв. 1238 не содержат комплексов ископаемых палиноморф, а в ходе полевых изысканий в естественных выходах не были обнаружены и какие-либо другие палеонтологические остатки. Как следствие, пока точно не удается установить и верхнюю возрастную границу “полтавских” слоев.
Г.П. Леонов (1961) рассматривал венцовские слои как прибрежные или даже лагунные отложения. Установление разнообразного комплекса диноцист подтверждает морской генезис венцовских слоев, а их согласное подстилание типичных полтавских фаций свидетельствует о том, что венцовские слои являются базальным горизонтом верхнего осадочного комплекса.
Авторами в рамках экспедиционных работ “полтавские” слои были исследованы и в естественных выходах и карьерах, где они представлены мощными пачками кварцевых песков различной окраски, с пологой косой слоистостью, с горизонтами глин и интенсивной биотурбацией (рис. 8д, 8е), на основании чего сделан вывод об их мелководно-морском генезисе.
С учетом данных Г.И. Горецкого (1982), район Преддонецкой моноклинали был также зоной разгрузки крупной палеореки, названной им “Соленовская река”, чей сток был направлен с севера на юг, а в аллювии преобладали пески, что, возможно, объясняет один из источников поставляемого терригенного материала (рис. 9).
Рис. 9.
Палеогеографическая карта палеобассейнов в хаттское время (по Попов и др., 2009, с изменениями).
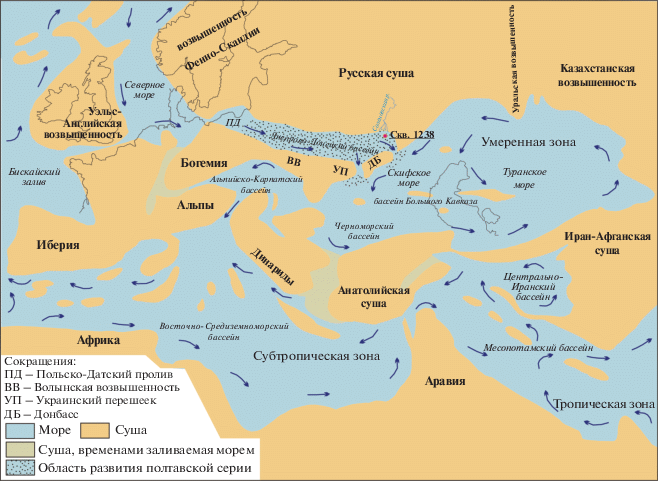
Прибрежно-морские условия формирования полтавской серии, по данным литологических и палеонтологических исследований, также реконструируются и для западных районов юга Русской плиты, вплоть до юга Белоруссии и Северной Украины (Савко и др., 1995, 2012; Зинова, 2001; Скляренко, 2010; Зосимович, Шевченко, 2014, 2015; Бойко, Скляренко, 2014 и др.). Предполагается, что в это время существовал широтный морской пролив, соединявший бассейн Преддонецкой моноклинали (как часть Паратетиса) с бассейнами Западной Европы (рис. 9). С учетом вышесказанного, а также полученных авторами данных о развитии морских фаций вплоть до самых западных районов Преддонецкой моноклинали, очевидно, что морская олигоценовая трансгрессия достигала значительно более северных территорий, чем это предполагалось ранее (Леонов, 1961; Воронина и др., 1988; Попов и др., 1993, 2009; Семенов, Столяров, 1988; Столяров, 1991; Столяров, Ивлева, 2004а, 2004б).
Палинологические данные по динамике изменений ассоциаций диноцист, празинофитов, спор и пыльцы наземных растений в соленовское и калмыцкое время в смежных регионах Нижнего Дона, Нижней Волги, Ергеней и Предкавказья (Заклинская, 1953; Ананова, 1974; Практическая…, 1990; Запорожец, 1998; Ахметьев, Запорожец, 2014; Запорожец, Ахметьев, 2015, 2017; Застрожнов и др., 2019) позволяют реконструировать этапы формирования венцовских и “полтавских” слоев. Так, первый этап характеризуется нарастанием количества пыльцы Taxodiaceae (Taxodium, Glyptostrobus, Sequoia, Metasequoia), пыльцы тепло- и влаголюбивых растений (Juglans, Carpinus, Fagus, Tilia, Ulmus, Quercus, Rhus, Myricaceae, Acer, Fraxinus, Hammamelis, Ilex, Liquidambar, Nyssa, Nelumbo) в течение рюпельского века, что было обусловлено развитием крупной трансгрессии и привело к значительному заболачиванию прилегающих районов суши и потеплению климата, при этом пик потепления приходился на раннеморозкинское время в Предкавказье и раннесоленовское время в Ергенях. Этому интервалу времени отвечает формирование тонких, преимущественно глинистых фаций верхов хадумского надгоризонта Предкавказья. Комплексы морского фитопланктона отражают нестабильные, возможно опресненные, с затрудненным водообменом палеообстановки осадконакопления.
На втором этапе, в позднеморозкинское (или позднесоленовское) время, возникло свободное соединение бассейна Паратетиса с бассейнами севера Европы, при сохраняющемся теплом климате, о чем свидетельствуют первые таксономически разнообразные спектры диноцист, находки моллюсков и состав спорово-пыльцевых комплексов (Застрожнов и др., 2019).
На третьем этапе, в конце позднесоленовского и начале раннекалмыцкого времени, по данным изучения спор и пыльцы растений, происходило похолодание климата при сохранении его высокой влажности, а в комплексах диноцист начинали доминировать перидиниоидные роды Rhombodinium и Deflandrea. Началу третьего этапа отвечает максимальное количество переотложенных палиноморф, а палинологические ассоциации при этом отличаются многочисленной пыльцой сосновых (Pinus, Picea, Tsuga, Abies), преобладанием среди покрытосеменных листопадных Betulaceae (Alnus, Betula, Corylus), Salicaceae, что указывает на гумидизацию климата, усиление процессов денудации и воздымание окружающей суши. Сочетание этих факторов привело к сильному стоку речных вод в бассейн и началу формирования мощных песчаных тел, относимых к калмыцкой свите, по северной периферии Паратетиса (Столяров, 1991; Столяров Ивлева, 2004а, 2004б). Позднекалмыцкое время охарактеризовано комплексом диноцист с доминированием рода Deflandrea, обеднением бентосной фауны моллюсков и фораминифер, что в целом вновь отражает обстановки полузамкнутого морского бассейна.
Мы полагаем, что установленный в венцовских слоях палинокомплекс с доминированием пыльцы Pinaceae, в совокупности с частыми Rhombodinium и Deflandrea, может быть сопоставлен с комплексами позднесоленовского времени, а вышележащие “полтавские” слои могли накапливаться в течение раннекалмыцкого времени. Это позволяет коррелировать венцовские слои с верхнесоленовской подсвитой, а полтавские слои – с калмыцкой свитой Ергеней и Нижнего Поволжья.
Исходя из литологического строения и стратиграфического положения венцовских и полтавских слоев в скв. 1238, эти слои могут быть сопоставлены с берекской свитой Воронежской антеклизы (Горбаткина, Иосифова, 2004), а также со змиевской и сивашской свитами Северной Украины (Стратиграфическая…, 1987; Зосимович, Шевченко, 2014, 2015).
ВЫВОДЫ
По данным изучения диноцист, в низах скв. 1238, пробуренной в Преддонецкой моноклинали, залегают отложения позднесантонского-раннекампанского возраста. В перекрывающих их морских палеогеновых отложениях выявлены интервалы диноцистовых зон из шкал Поволжья и Восточного Пери-Тетиса: Alisocysta margarita, Apectodinium hyperacanthum, Axiodinium augustum, Ochetodinium romanum/Samlandia chlamydophora, а также слои с Rhombodinium draco–Deflandea spinulosa, что позволило уточнить возраст вмещающих отложений. Так, на основании полученных данных подтвержден раннетанетский возраст бузиновского горизонта, возраст вешенского горизонта уточнен и отвечает позднему палеоцену–инициальному эоцену. Вышележащие суровикинские и осиновские слои датированы частью раннего–среднего ипра. Верхняя часть осиновских слоев, а также чирские слои не вскрыты в скважине, что требует поиска более полных непрерывных разрезов. Литологический состав и стратиграфический возраст слоев позволяют сопоставлять их с пролейской свитой Поволжья и частью каневской свиты Северной Украины. Отложения, характеризующие средний и верхний эоцен, а также большую часть нижнего олигоцена, на данной территории не установлены. Венцовские и “полтавские” слои сопоставляются с верхнесоленовской подсвитой и нижней частью калмыцкой свиты Ергеней. В разрезе скв. 1238 выявлены два крупных перерыва в осадконакоплении, первый из которых соответствует верхнему кампану–зеландию, а второй – верхнему ипру–нижнему рюпелю.
Нами продемонстрировано, что для расчленения палеогена Преддонецкой моноклинали вполне применимы местные стратиграфические подразделения, предложенные в 1961 г. Г.П. Леоновым. Установлено, что в пределах Преддонецкой моноклинали формирование отложений в позднем палеоцене–раннем эоцене приурочено к периоду крупной морской трансгрессии, а в конце раннего олигоцена–позднем олигоцене – к регрессии, в условиях полузамкнутого бассейна. Первому этапу отвечают бузиновская и вешенская свиты, а также суровикинские и осиновские слои, второму – венцовские и “полтавские” слои.
Благодарности. Авторы благодарят АО “Северо-Кавказское ПГО” за предоставленный керновый материал, а также признательны коллегам А.С. Застрожнову (ВСЕГЕИ), С.В. Попову (ПИН РАН), С.В. Гришину (ГИН РАН), Н.И. Панченко (г. Волгоград) за помощь в проведении экспедиционных работ. Авторы выражают благодарность Н.К. Лебедевой (ИНГГ СО РАН) и С.В. Попову (ПИН РАН), а также Е.Ю. Барабошкину, Ю.Б. Гладенкову и М.А. Рогову за ценные замечания и комментарии.
Источники финансирования. Работа выполнена в рамках государственного задания ГИН РАН; полевые и аналитические исследования выполнены при финансовой поддержке гранта РФФИ № 18-05-00505а.
Список литературы
Александрова Г.Н. Диноцисты палеоцена Среднего и Нижнего Поволжья: стратиграфия и палеообстановки. Автореф. дисс. … канд. геол.-мин. наук. М.: ГИН РАН, 2013. 24 с.
Александрова Г.Н., Щербинина Е.А. Стратиграфия и палеообстановки переходного палеоцен-эоценового интервала восточного Крыма // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 4. С. 62–88.
Александрова Г.Н., Беньямовский В.Н., Запорожец Н.И., Застрожнов А.С., Застрожнов С.И., Табачникова И.П., Орешкина Т.В., Закревская Е.Ю. Палеоген юго-запада Волгоградской области (скв. 13, Гремячинская площадь). Ст. 1. Биостратиграфия // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 3. С. 70–95.
Александрова Г.Н., Беньямовский В.Н., Вишневская В.С., Застрожнов А.С. Новые данные по биостратиграфии верхнего мела Нижнего Поволжья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 5. С. 25–64.
Ананова Е.Н. Пыльца в неогеновых отложениях юга Русской равнины. Л.: Изд-во ЛГУ, 1974. 196 с.
Андреева-Григорович А.С., Шевченко Т.В. Зональная шкала палеогеновых отложений Украины по диноцистам // Палеонтологiчнi дослiдження в Украïнi: Зб. наук. праць. Кiiв: ИГН НАН Украïнi, 2007. С. 211–214.
Андреева-Григорович А.С., Запорожец Н.И., Шевченко Т.В., Александрова Г.Н., Васильева О.В., Яковлева А.И., Стотланд А.Б., Савицкая Н.А. Атлас диноцист палеогена Украины, России и сопредельных стран. Киев: Наукова думка, 2011. 221 с.
Ахметьев М.А., Беньямовский В.Н. Стратиграфическая схема морского палеогена юга Европейской России // Бюлл. МОИП. Отд. геол. 2003. Т. 78. Вып. 5. С. 40–51.
Ахметьев М.А., Запорожец Н.И. События палеогена в Центральной Евразии, их роль в развитии флоры и растительного покрова, смещении границ фитохорий и изменениях климата // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 3. С. 90–114.
Беньямовский В.Н. Предложения к уточнению стратиграфической схемы палеогена Воронежско-Придонецкого региона // Состояние стратиграфической базы центра и юго-востока Восточно-Европейской платформы. Материалы совещания (Москва, ВНИГНИ, 23 –25 ноября 2015 г.). М.: ВНИГНИ, 2016. С. 122–128. С. 134–139.
Беньямовский В.Н. Предложения по структурно-фациальному районированию и субрегиональным горизонтам палеогена Воронежско-Придонецкого субрегиона // ПАЛЕОСТРАТ-2017. Годичное собрание (научная конференция) секции палеонтологии МОИП и Московского отд. Палеонтологического общества при РАН. Москва, 26–28 января 2017 г. Программа и тезисы докладов. Ред. Алексеев А.С. М.: ПИН РАН, 2017а. С. 15–16.
Беньямовский В.Н. Палеонтологическое и палеогеографическое обоснование региональных горизонтов палеоцена‒эоцена Воронежско-Придонецкого региона юга Европейской России // Материалы LXIII сессии Палеонтологического общества при РАН (3–7 апреля 2017 г., Санкт-Петербург). СПб.: ВСЕГЕИ, 2017б. С. 31–33.
Беньямовский В.Н. Актуализированная стратиграфическая схема нового поколения палеогеновых отложений юга Русской и севера Скифской плит – ключевого бассейна в субширотной коммуникационной системе палеогеновых эпиконтинентальных морей Евразийского Пери-Тетиса // Геология морей и океанов. Материалы XXII Международной научной конференции (школы) по морской геологии. Т. I. М.: ИО РАН, 2017в. С. 45–49.
Бойко Н.И., Скляренко Г.Ю. Титан-циркониевые россыпи полтавской свиты южного склона Воронежской антеклизы // Проблемы минералогии, петрографии и металлогении. Научные чтения памяти П.Н. Чирвинского. 2014. Вып. 17. С. 181–187.
Бугрова Э.М., Беньямовский В.Н., Табачникова И.П., Рябоконь Т.С., Радионова Э.П. Опорный разрез Кантемировка палеогена Воронежского региона // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 6. С. 78–100.
Васильева О.Н. Органикостенный планктон среднего эоцена в парастратотипическом разрезе Кереста-1 (Ергени, Ростовская область) // Ежегодник-2012. Екатеринбург: Изд-во ИГГ УрО РАН, 2013. С. 13–21 (Тр. ИГГ УрО РАН. Вып. 160).
Васильева О.Н. Зональное деление по диноцистам среднеэоценовых отложений Волгоградского Поволжья // Ежегодник-2018. Екатеринбург: Изд-во ИГГ УрО РАН, 2019. С. 3–11.
Васильева О.Н., Мусатов В.А. Биостратиграфия палеогена Северного Прикаспия по диноцистам и наннопланктону (Новоузенская опорная скважина). Статья 1. Обоснование возраста отложений и корреляция // Стратиграфия. Геол. корреляция. 2010а. Т. 18. № 1. С. 88–109.
Васильева О.Н., Мусатов В.А. Биостратиграфия палеогена Северного Прикаспия по диноцистам и наннопланктону (Новоузенская опорная скважина). Статья 2. Биотические события и палеоэкологические обстановки // Стратиграфия. Геол. корреляция. 2010б. Т. 18. № 2. С. 77–98.
Воронина А.А., Кургалимова Г.Г., Попов С.В., Семенов Г.И., Столяров А.С. Биостратиграфия и фациальные особенности майкопских отложений Волго-Донского региона // Изв. АН СССР. Сер. геол. 1988. № 9. С. 39–50.
Геология СССР. Том 46. Ростовская, Волгоградская, Астраханская области и Калмыцкая АССР. Часть 1. Геологическое описание. Ред. Белов Ф.А., Егоров А.И., Погребнов Н.И. М.: Недра, 1969. 666 с.
Горбаткина Т.Е., Иосифова Ю.И. Новые стратиграфические подразделения палеогена Воронежской антеклизы – воробьевская и пасековская свиты // Вестник Воронежского гос. ун-та. Геология. 2004. № 7. С. 28–44.
Горецкий Г.И. Палеопотамологические эскизы Палео-Дона и Пра-Дона. Минск: Наука и техника, 1982. 248 с.
Государственная геологическая карта Российской Федерации. Масштаб 1 : 1 000 000 (третье поколение). Серия Центрально-Европейская. Лист М-38 – Волгоград. Объяснительная записка. Ред. Застрожнов С.И. СПб.: Картфабрика ВСЕГЕИ, 2009. 399 с.
Заклинская Е.Д. Описание некоторых видов пыльцы и спор, выделенных из третичных отложений Пасековского карьера Воронежской обл. // Тр. ГИН АН СССР. Сер. Геология. 1953. № 59. Вып. 142. С. 60–102.
Запорожец Н.И. Новые данные по фитостратиграфии эоцена и олигоцена Северных Ергеней (юг Русской платформы) // Стратиграфия. Геол. корреляция. 1998. Т. 6. № 3. С. 56–73.
Запорожец Н.И. Палинологические комплексы кумского горизонта среднего эоцена Кавказа и его возрастных аналогов в сопредельных регионах // Стратиграфия. Геол. корреляция. 2001. Т. 9. № 6. С. 83–103.
Запорожец Н.И., Ахметьев М.А. Комплексы органикостенного фитопланктона, пыльцы и спор соленовского горизонта (нижний олигоцен) // Стратиграфия. Геол. корреляция. 2015. Т. 23. № 3. С. 96–122.
Запорожец Н.И., Ахметьев М.А. Палеоботаническое изучение разреза майкопской серии олигоцена–нижнего миоцена по р. Белая выше г. Майкопа, Предкавказье // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 6. С. 61–83.
Застрожнов А.С., Попов С.В., Беньямовский В.Н., Мусатов В.А., Ахметьев М.А., Запорожец Н.И., Богачкин А.Б., Столяров А.С. Опорный разрез олигоцена юга Восточно-Европейской платформы // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 4. С. 59–92.
Зинова Р.А. Соленовский горизонт Г.И. Горецкого и его возрастные аналоги // Лiтосфера. 2001. № 14. С. 32–38.
Зосимович В.Ю. Региояруса палеогена Североукраинской палеоседиментационной провинции // Стратиграфия осадочных образований верхнего протерозоя и фанерозоя. Материалы Международной научной конференции (Киев, 23–26 сентября 2013 г.). Киев: LATandK, 2013. С. 66–67.
Зосимович В.Ю., Шевченко Т.В. Етапи розвитку осадових басейнів Північної України в палеогені // Збор. наук. пр. ІГН НАН України. 2014. Т. 7. С. 83–100.
Зосимович В.Ю., Шевченко Т.В. Палеогенові відклади Північноукраїнської палеоседиментаційної провінції // Збор. наук. пр. ІГН НАН України. 2015. Т. 8. С. 68–121.
Зосимович В.Ю., Зернецький Б.Ф., Андреєва-Григорович А.С., Люльєва С.А., Маслун Н.В., Рябоконь Т.С., Шевченко Т.В. Регіояруси палеогену платформної України // Біостратиграфічні критерії розчленування та кореляції відкладів фанерозою України. Зб. наук. пр. Ін-ту НАН України. Киев, 2005. С. 118–132.
Зосимович В.Ю., Шевченко Т.В., Цыба Н.Н. Неостратотип “каневского яруса” // Сб. научн. трудов. Института геол. наук НАН Украины. 2013. Т. 6. Вып. 1. С. 98–110.
Зосимович В.Ю., Зернецкий Б.Ф., Ольштынская А.П., Рябоконь Т.С., Шевченко Т.В. Корреляция региоярусов палеогена Северной и Южной Украины // Стратотипові та опорні розрізи фанерозойських відкладів України: сучасний стан палеонтологічної вивченості та перспективи подальших досліджень. Матер. XXXVI сес. Палеонтол. тов-ва НАН України (Львів, 24–26 вересня 2015 р.). Киев: ІГН НАН, 2015. С. 53–54.
Зосимович В.Ю., Шевченко Т.В., Рябоконь Т.С., Ольштынская А.П. Региональные стратиграфические подразделения палеогена Северной Украины // Состояние стратиграфической базы центра и юго-востока Восточно-Европейской платформы. Материалы совещания (Москва, ВНИГНИ, 23–25 ноября 2015 г.). М.: ВНИГНИ, 2016. С. 140–144.
Зосимович В.Ю., Зернецкий Б.Ф., Ольштынская А.П., Рябоконь Т.С., Шевченко Т.В. Корреляция региоярусов палеогена Северной и Южной Украины // Актуальні проблеми та перспективи розвитку геології: наука й виробництво. Матеріали Міжнародного геологічного форуму (19–24 червня 2017 р., м. Одеса, Україна). Киев: УкрДГРІ, 2017. С. 141–144.
Лебедева Н.К. Биостратиграфия верхнемеловых отложений в бассейне на р. Уса (Полярное Предуралье) по диноцистам // Стратиграфия. Геол. корреляция. 2005. Т. 13. № 3. С. 114–131.
Лебедева Н.К. Биостратиграфия верхнемеловых отложений севера Сибири по диноцистам // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Сб. научн. трудов. Ред. Первушов Е.М. Саратов: Изд-во Сарат. ун-та, 2007. С. 108–122.
Лебедева Н.К., Александрова Г.Н., Шурыгин Б.Н., Овечкина М.Н., Гнибиденко З.Н. Палеонтологическая и магнитостратиграфическая характеристика верхнемеловых отложений, вскрытых скважиной 8 Русско-Полянского района (юг Западной Сибири) // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 1. С. 43–73.
Леонов Г.П. Палеогеновые отложения Сталинградского Поволжья и их соотношения с соответствующими образованиями бассейнов рр. Дона и Днепра // Бюлл. МОИП. 1936. Т. XIV(4). С. 287–321.
Леонов Г.П. Стратиграфия палеогеновых отложений бассейна Среднего Дона // Ученые записки МГУ. 1939. Вып. 26. Геология. С. 3–51.
Леонов Г.П. Основные вопросы региональной стратиграфии палеогена Русской плиты. М.: МГУ, 1961. 552 с.
Мусатов В.А. Лютет или бартон? Возраст сергеевской свиты по наннопланктону в опорном разрезе “Кантемировка” Воронежской антеклизы и корреляция с сопредельными регионами // Недра Поволжья и Прикаспия. 2020. Вып. 101. С. 4–26.
Мусатов В.А., Богачкин А.Б. Бартонский ярус среднего эоцена европейской части России. Стратиграфический объем и критерии определения границ. Статья 1. Пределы точности зональной стратиграфии лютетско-бартонского интервала по наннопланктону и возраст бартонских слоев Англии // Недра Поволжья и Прикаспия. 2019а. Вып. 98. С. 3–24.
Мусатов В.А., Богачкин А.Б. Бартонский ярус среднего эоцена европейской части России. Стратиграфический объем и критерии определения границ. Статья 2. Детальная зональная стратиграфия лютетско-бартонского интервала по наннопланктону и палеомагнитная характеристика разрезов Предкавказья // Недра Поволжья и Прикаспия. 2019б. Вып. 99. С. 3–34.
Мусатов В.А., Рябоконь Т.С. Распределение наннофоссилий и фораминифер в опорном разрезе киевской свиты с. Халепье (Киевское Приднепровье, Украина) // 40 лет Палеонтологическому обществу Украины. Материалы XXXVIII сессии Палеонтологического общества НАН Украины (Канев, 23–26 мая 2017 г.). Киев: ИГН НАНУ, 2017. С. 117–119.
Мусатов В.А., Васильева О.Н., Музылев Н.Г. Результаты исследования наннопланктона и диноцист из неостратотипа керестинской свиты в Южных Ергенях // ПАЛЕОСТРАТ‑2014. Годичное собрание (научн. конф.) секции палеонтологии МОИП и Моск. отд. Палеонтол. общества при РАН (Москва, 27–29 января 2014 г.). Ред. Алексеев А.С. М.: ПИН РАН, 2014. С. 53–54.
Орешкина Т.В., Яковлева А.И. Новые данные по палеонтологической характеристике бучакской, киевской и обуховской свит северного борта Днепровско-Донецкой впадины (скв. 230, пос. Стрелечье) // Палеонтологічні дослідження в Україні: історія, сучасний станта перспективи. Збор. наук. пр. ІГН НАН України. Kyiv, 2007. С. 233–237.
Попов С.В., Ахметьев М.А., Запорожец Н.И., Воронина А.А., Столяров А.С. Эволюция Восточного Паратетиса в позднем эоцене–раннем миоцене // Стратиграфия. Геол. корреляция. 1993. Т. 13. № 6. С. 10–39.
Попов С.В., Ахметьев М.А., Лопатин А.В., Бугрова Э.М., Сычевская Е.К., Щерба И.Г., Андреева-Григорович А.И., Запорожец Н.И., Николаева И.А., Копп М.Л. Палеогеография и биогеография бассейнов Паратетиса. Часть 1. Поздний эоцен–ранний миоцен. М.: Научный мир, 2009. 200 с.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 32. СПб.: ВСЕГЕИ, 2001. С. 18–20.
Практическая палиностратиграфия. Ред. Панова Л.А., Ошуркова М.В., Романовская Г.М. Л.: Недра, 1990. 348 с.
Савко А.Д., Беляев В.И., Иконников Н.Н. Титан-циркониевые россыпи Центрально-Черноземного района. Воронеж: Изд-во Воронеж. гос. ун-та, 1995. 148 с.
Савко А.Д., Звонарев А.Е., Иванов Д.А. Цирконий-титановые россыпи Воронежской антеклизы: типы, эпохи и факторы формирования // Геология рудных месторождений. 2012. Т. 54. № 1. С. 71–94.
Семенов В.П. Палеоген Воронежской антеклизы. Воронеж: Изд-во ВГУ, 1965. 278 с.
Семенов В.П. Воронежская антеклиза // Стратиграфия СССР. Палеогеновая система. Ред. Гроссгейм В.А., Коробков И.А. М.: Недра, 1975. С. 183–191.
Семенов Г.И., Столяров А.С. Вопросы стратиграфии майкопских отложений Волго-Донского региона // Бюлл. МОИП. Отд. геол. 1988. Т. 63. Вып. 2. С. 70–83.
Скляренко Г.Ю. Титан-циркониевые россыпи в отложениях полтавской серии миоцена юго-восточного склона Воронежской антеклизы. Автореф. дис. … канд. геол.-мин. наук. Ростов-на-Дону: ЮФУ, 2010.
Соколов Н.А. Нижнетретичные отложения Южной России // Тр. Геол. комитета. 1893. Т. IX. № 2. 329 с.
Столяров А.С. Палеогеография Предкавказья, Волго-Дона и Южного Магышлака в позднем эоцене и раннем олигоцене // Бюлл. МОИП. Отд. геол. 1991. Т. 66. Вып. 4. С. 64–79.
Столяров А.С., Ивлева Е.И. Верхнеолигоценовые отложения Предкавказья, Волго-Дона и Мангышлака (центральная часть Восточного Паратетиса). Сообщение 1. Основные особенности состава и строения // Литология и полезн. ископаемые. 2004а. № 3. С. 252–270.
Столяров А.С., Ивлева Е.И. Верхнеолигоценовые отложения Предкавказья, Волго-Дона и Мангышлака (центральная часть Восточного Паратетиса). Сообщение 2. Фациально-палеогеографические условия осадконакопления // Литология и полезн. ископаемые. 2004б. № 4. С. 359–368.
Стратиграфия СССР. Палеогеновая система. Ред. Гроссгейм В.А., Коробков И.А. Москва: Недра, 1975. 524 с.
Стратиграфическая схема палеогеновых отложений Украины (унифицированная). Отв. ред. Макаренко Д.Е. Киев: Наукова думка, 1987. 116 с.
Унифицированная стратиграфическая схема палеогеновых отложений Поволжско-Прикаспийского субрегиона. Объяснительная записка. Отв. ред. Ахметьев М.А. М.: ВНИГНИ, 2015. 96 с. + 8 листов.
Шпуль В. Г. Новые данные по фитостратиграфии эоцен-олигоцена юго-восточного склона Воронежской антеклизы // Вестник Воронежского гос. ун-та. Сер. Геология. 2005. № 1. С. 55–70.
Шпуль В.Г. Фитостратиграфия эоцена северо-восточного склона Воронежской антеклизы // Проблеми бiостратиграфii нижнего протерозою и фанерозою Украiни: зб. наук. праць. Iнституту геологiчних наук. Киiв, 2006. С. 208–214.
Шпуль В.Г. Эоцен-олигоценовые флоры Воронежской антеклизы по данным палинологических исследований – Eocene-Oligocenic flora of Voronezh anteclise according to palynological data // Палеонтологiчнi дослiдження в Украiнi: iсторiя, сучасний стан та перспективи: сборник статей. Киев, 2007. С. 255–260.
Шпуль В.Г. Палиностратиграфия палеоцена–нижнего эоцена северо-восточного склона Воронежской антеклизы // Биосферные основы построения стратиграфических схем фанерозоя Украины. Материалы XXXI Сессии Палеонтологического общества НАН Украины, 19–22 мая 2008 г., Симферополь. Киев, 2008. С. 144–149.
Шпуль В.Г. Палеоцен-эоценовые флоры Воронежской антеклизы по данным палинологических исследований // Викопна фауна i флора Украiни: палеоекологiчний та стратиграфiчний аспекти: збiрник наукових праць Iнституту геологiчних наук НАН Украiни. Киiв, 2009. С. 220–225.
Шпуль В. Г. Значение палинологии для обоснования региональных стратиграфических подразделений палеогена Воронежской антеклизы // Эволюция жизни на Земле. Материалы 4-го Международного симпозиума, 10–12 ноября 2010 г. Томск, 2010. С. 426–429.
Яковлева А.И. Детализация эоценовой диноцистовой шкалы для восточного Перитетиса // Бюлл. МОИП. Отд. Геол. 2017. Т. 92. Вып. 2. С. 32–48.
Яковлева А.И., Александрова Г.Н. К вопросу об уточнении зонального деления по диноцистам палеоцен-эоценовых отложений Западной Сибири // Бюлл. МОИП. Отд. Геол. 2013. Т. 88. № 1. С. 59–82.
Янин Б.Т., Беньямовский В.Н. Норы десятиногих ракообразных из палеогеновых отложений Среднего Поволжья и их палеогеографическое значение // Вестник Моск. ун-та. Сер. 4: Геология. 2018. № 3. С. 12–24.
Aleksandrova G.N., Radionova E.P. On the late Paleocene stratigraphy of the Saratov Volga Region: Micropaleontological characteristics of the Kamyshin Formation, Dyupa Gully section // Paleontol. J. 2006. V. 40. № 5. P. 543–557.
Brinkhuis H. Late Eocene to early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994. V. 107. P. 121–163.
Costa L.I., Manum S.B. The distribution of the interregional zonation of the Paleogene (D1-D15) and the Miocene (D16-D20) // The Northwest European Tertiary Basin. Results of the International Geological Correlation Program, Project No. 124. Geol. Jb., Hannover. 1988. A100. P. 321–330.
Frieling J., Iakovleva A.I., Reichart G.-J., Aleksandrova G.N., Gnibidenko Z.N., Schouten S., Sluijs A. Paleocene-Eocene warming and biotic response in the epicontinental West Siberian Sea // Geology. 2014. V. 42. № 9. P. 767–770.
Heilmann-Clausen C. The Danish Subbasin, Paleogene dinoflagellates // The Northwest European Tertiary Basin. Results of the International Geological Correlation Program, Project No. 124 // Geol. Jb., Hannover. 1988. A100. P. 339–343.
Iakovleva A.I. Middle-late Eocene dinoflagellate cysts from NE Ukraine (Borehole No. 230, Dnepr-Donets Depression): stratigraphic and paleoenvironmental approach // Acta Palaeobot. 2015. V. 55. № 1. P. 19–51.
Iakovleva A.I., Heilmann-Clausen C. Early and middle Eocene dinoflagellate cysts from the Aktulagay section, Kazakhstan // Palynology. 2020. https://doi.org/10.1080/01916122.2019.1705933
King Ch., Iakovleva A.I., Heilmann-Clausen C., Steurbaut E. Ypresian (early Eocene) stratigraphy of the Suvlu-Kaya reference section in the Bakhchisaray area (Crimea) // Newslett. Stratigr. 2018. V. 51. № 2. P. 167–208.
Lebedeva N.K. Dinocyst biostratigraphy of the Upper Cretaceous of Northern Siberia // Paleontol. J. 2006. V. 40. Suppl. 5. P. S604–S621.
Mudge D.C., Bujak J.P. Eocene stratigraphy of the North Sea Basin // Mar. Petrol. Geol. 1994. V. 11. P. 166–181.
Ogg J.G., Ogg F.M., Gradstein F.M. A concise Geologic Time Scale 2016. Elsevier, 2016. 229 p.
Powell A.J. A Stratigraphic Index of Dinoflagellate cysts // British Micropaleontol. Soc. Publ. Ser. 1992. 290 p.
Powell A.J., Brinkhuis H., Bujak J.P. Upper Paleocene-lower Eocene dinoflagellate cyst sequence biostratigraphy of southeast England // Correlation of the Early Paleogene in Northwest Europe. Geol. Soc. Spec. Publ. 1996. V. 101. P. 145–183.
Shcherbinina E.A., Gavrilov Yu.O., Iakovleva A.I., Golovanova O.V., Pokrovsky B.G. Aleksandrova G.N. Environmental dynamics during the Paleocene-Eocene thermal maximum (PETM) in the northeastern Peri-Tethys revealed by high-resolution micropaleontological and geochemical studies of the Caucasian key-section // Palaeogeogr. Palaeoecol. Palaeoclimat. 2016. V. 456. P. 60–81.
Shcherbinina E., Iakovleva A., Gavrilov Yu., Golovanova O., Myzylöv N. Lower Eocene sedimentary succession and microfossil biostratigraphy in the central northern Caucasus basin // Geologica Acta. 2020. V. 18. № 1. P. 1–15.
Śliwińska K.K., Abrahamsen N., Beyer C., Brünings-Hansen T., Thomsen E., Ulleberg K., Heilmann-Clausen C. Bio- and magnetostratigraphy of Rupelian–mid Chattian deposits from the Danishland area // Rev. Palaeobot. Palynol. 2012. V. 172. P. 48–69.
Vasilyeva O.N., Musatov V.A. The Paleogene Dinoflagellate Cyst and Nannoplankton Biostratigraphy of the Caspian Depression // Intech: Stratigraphic Analysis of Layered Deposits. 2012. Ch. 7. P. 161–194. ISBN 978-953-51-0578-7.
Williams G.L., Fensome R.A., MacRae R.A. The Lentin and Williams index of fossil dinoflagellates 2017 edition // Am. Assoc. Stratigr. Palynol. Contrib. Ser. 2017. № 48.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция