

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 6, стр. 171-180
Стратиграфия кайнозоя восточной части Опухско-Пекульнейского прогиба, Корякское нагорье, Северо-Восток России
Ю. Б. Гладенков *
Геологический институт РАН
Москва, Россия
* E-mail: gladenkov@ginras.ru
Поступила в редакцию 24.06.2019
После доработки 12.08.2019
Принята к публикации 20.11.2019
Аннотация
Описываются морские отложения (две свиты) кайнозоя, развитые в восточной части Опухско-Пекульнейского прогиба Корякского нагорья. Сведения о них до последнего времени почти отсутствовали. Нижняя свита (ретырвеемская) сложена темными конгломератами и песчаниками с большим количеством карбонатных конкреций и представительными комплексами двустворчатых и брюхоногих моллюсков (изучено более 400 раковин и отпечатков) и бентосных фораминифер раннего эоцена. В ней выделено две подсвиты и шесть пачек. Верхняя свита (моржовская) включает светлые рыхлые гравелиты и песчаники, в которых найдены остатки моллюсков и фораминифер раннего миоцена. Фаунистические комплексы позволили сопоставить изученные свиты со стратиграфическими подразделениями смежных районов и установить перерывы в разрезе, которые охватывают, с одной стороны, большую часть палеоцена, а с другой – верхи эоцена и олигоцен. В ретырвеемской свите отмечены тепловодные комплексы моллюсков, появление которых в северо-бореальных широтах, возможно, связано с глобальным потеплением в раннем эоцене. Впервые приведены изображения остатков характерных моллюсков.
ВВЕДЕНИЕ
Корякское нагорье остается геологически слабо изученной областью. Это касается и кайнозойских толщ, которые встречаются в этом регионе в отдельных прогибах или впадинах (Егиазаров и др., 1965). Один из таких прогибов, который в литературе получил название Опухско-Пекульнейский или Хатырский, расположен на юго-востоке Корякского нагорья и открывается в Берингово море (рис. 1). Он протягивается почти на 300 км в северо-восточном направлении от бухты Опуха до Пекульнейского озера (при ширине наземной части около 30 км). На севере он контактирует обычно по тектоническим нарушениям со сложно дислоцированными образованиями мезозоя, хотя в отдельных случаях отмечается несогласное залегание палеогеновых или неогеновых отложений на породах юры и мела. Особое внимание этот прогиб привлек в 1950-х–1960-х годах, в частности из-за выявленных здесь нефтепроявлений (Белова и др., 1961; Гладенков, 1962). Эти проявления были приурочены в основном к палеогеновым (олигоценовым) толщам. Они первоначально были установлены в западной (нижнехатырской) части прогиба, но позже были выявлены и в других его частях. Автор настоящей работы изучал кайнозойские толщи в 1959–1962 гг. на западной окраине прогиба – в районе р. Хатырка (Богидаева, Гладенков, 1963), когда он проводил в этом районе геологическую съемку, и в 1981 г., когда вместе с В.И. Волобуевой-Богидаевой (Северо-восточный комплексный научно-исследовательский институт – СВКНИИ ДВО РАН, Магадан) исследовал разрезы кайнозоя с целью создания региональной стратиграфической схемы. Данный район посещали в 1960-х–1970х годах и другие геологи (Ю.К. Бурлин, В.В. Донцов, В.Е. Мохов, Н.В. Устинов и др.), но они занимались в основном литологическими и нефтепоисковыми проблемами (Бурлин и др., 1973). В начале 1980-х годов В.И. Волобуева и автор совместно изучили ряд кайнозойских разрезов центральной части Хатырского прогиба: горный массив Хайидин и район Пекульнейского озера, где были пробурены несколько скважин. Эти материалы с использованием данных по двустворчатым и брюхоногим моллюскам и бентосным фораминиферам позволили составить первую корреляционную схему кайнозойских толщ (Гладенков и др., 1985), но самая восточная часть прогиба оставалась практически неизученной. Существовали лишь скудные сведения о кайнозое этого района (Бушуев, 1954). Из работ 1950-х годов следовало, что там развита свита этерет (названная по р. Этеретваям) мощностью 450–600 м. Ее возраст был определен Л.В. Криштофович по редким находкам моллюсков как средне-позднемиоценовый. По данным М.И. Бушуева (1954), в районе небольшого оз. Кайпыльдин свита залегает в тектоническом блоке и не имеет нормальных контактов с более древними отложениями. Она сложена в основном песчаниками с большим количеством конгломератов и карбонатных конкреций, с включением в верхней части прослоев черных углистых сланцев. С 1950-х годов эта свита подробно не изучалась, хотя ее наличие в отдельных блоках отмечалось (Волобуева, Красный, 1969). Поэтому В.И. Волобуева и автор в 1983 г. решили посетить этот геологический объект с целью его более детального исследования. Выяснилось, что кайнозойские толщи по рекам Ретырвеем (ранее называлась Этеретваям), Моржовая, Пика и по морскому побережью Берингова моря сильно дислоцированы, причем в ряде случаев слои стоят вертикально, и местами плохо обнажены, что часто мешает определить стратиграфическую последовательность слоев. Тем не менее нами был составлен разрез кайнозойских толщ с расчленением их на две свиты и несколько пачек. Предварительный анализ собранных здесь органических остатков – моллюсков (определения В.И. Волобуевой и автора) и фораминифер (определения Л.И. Митрофановой) – позволил определить соответственно палеогеновый и неогеновый возраст этих свит. Нижняя свита была названа ретырвеемской (по р. Ретырвеем, в долине которой был описан стратотип свиты) и отнесена к нижнему–среднему (?) эоцену, а верхняя – моржовской (по р. Моржовая) и отнесена к нижнему миоцену. В самом кратком виде эти данные были опубликованы в работе (Гладенков и др., 1985).
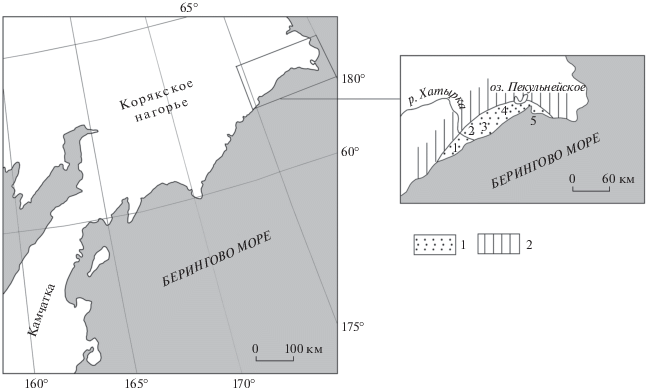
Рис. 1.
Расположение Опухско-Пекульнейского прогиба в Корякском нагорье. 1 – кайнозойские отложения; 2 – палеозойские и мезозойские образования. Цифрами показано положение отдельных разрезов кайнозоя: 1 – правобережье р. Хатырка, 2 – озеро Имыней Гыткин, 3 – горы Хайидин, 4 – Пекульнейское озеро, 5 – р. Ретырвеем.
К сожалению, вскоре В.И. Волобуева ушла из жизни, и собранная нами совместно коллекция моллюсков (более 400 раковин и отпечатков двустворок и гастропод), которую мы определили только предварительно, долгие годы хранилась в СВКНИИ ДВО РАН. По просьбе В.И. Волобуевой коллекция сравнительно недавно была передана автору для дополнительного исследования в ГИН РАН, где она сейчас и хранится под № 5004. Коллекция оказалась интересной и может быть использована, с одной стороны, для подтверждения наличия в восточной части Хатырского прогиба не только миоценовых, но и палеогеновых толщ (что необходимо учитывать при геокартировании и составлении новых стратиграфических схем в этом районе), и с другой стороны – для выявления особенностей природной среды прошлого, в частности температурных характеристик морских вод палеоген-неогенового бассейна.
Выше отмечалось, что определение фораминифер из изученных разрезов проводила в 1980-х годах Л.И. Митрофанова (образцы отбирались с интервалом 10–15 м). Используя эти данные, С.И. Бордунов по моей просьбе уточнил некоторые определения таксонов и возрастной диапазон отмеченных ею комплексов с учетом новых материалов.
Ниже приводятся краткое описание изученного разреза с установленными фаунистическими комплексами, привязанными к впервые выделенным пачкам упомянутых двух свит, а также фототаблицы характерных двустворчатых и брюхоногих моллюсков из этих свит.
ОПИСАНИЕ РАЗРЕЗА И ПАЛЕОНТОЛОГИЧЕСКИЕ КОМПЛЕКСЫ
Обе изучаемые свиты представлены терригенными, часто грубообломочными отложениями значительной мощности, которые имеют определенные литологические и палеонтологические отличия.
Ретырвеемская свита (нижний эоцен)
Свита состоит из двух подсвит (рис. 2, 3).
Рис. 2.
Геологическая схема междуречья Ретырвеем–Пика. 1 – мезозойские образования; 2 – тектонические нарушения; 3, 4 – кайнозойские образования: 3 – угленосные отложения, 4 – маркирующие горизонты песчаников и конгломератов; Р1, Р2 – нижняя и верхняя подсвиты ретырвеемской свиты (эоцен), М – моржовская свита (нижний миоцен), Q – четвертичные отложения, Mz – мезозойские отложения. Тонкими линиями показаны русла рек.
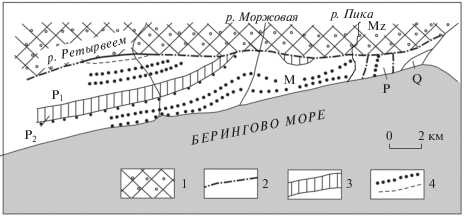
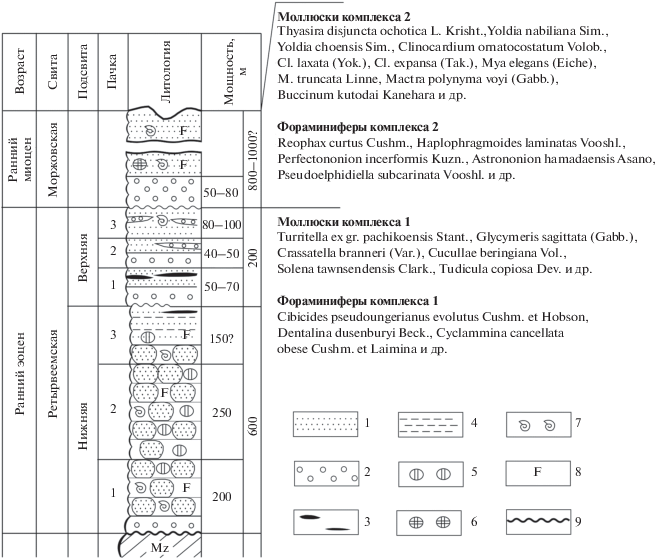
Рис. 3.
Стратиграфическая колонка кайнозойского разреза восточной части Опухско-Пекульнейского прогиба. 1 – песчаники, 2 – конгломераты и гравелиты, 3 – угли и лигниты, 4 – алевролиты и аргиллиты, 5 – карбонатные конкреции, 6 – переотложенные карбонатные конкреции, 7 – находки остатков моллюсков, 8 – находки остатков фораминифер, 9 – размывы.
Нижняя подсвита сложена в основном темными мелкозернистыми песчаниками с карбонатными конкрециями. В ней выделены три пачки.
Пачка 1. Залегает несогласно на породах мелового возраста – черных крепких аргиллитах и песчаниках, часто во флишоидном переслаивании. В основании пачки (3–8 м) присутствуют конгломераты темно-серые, местами ожелезненные, мелкогалечные, с крепким песчанистым цементом. В гальке отмечены обломки черных аргиллитов, песчаников и кремнистых пород. Над конгломератами залегают песчаники обычно темно-серые до черных, полимиктовые, крепкие, мелкозернистые, неяснослоистые, массивные, с крупнокусковыми обломками, видными при выветривании, с характерными карбонатными конкрециями обычно шаровидной формы (преобладают крупные, диаметром до 0.5 м, хотя отмечаются и мелкие, до 0.1 м). Встречаются карбонатные линзообразные стяжения мощностью до 1 м. Мощность пачки около 200 м.
В песчаниках найдены зубы акул и чешуя рыб и встречен представительный комплекс моллюсков 1: Yoldia watasei Kan., Crassatella branneri (Var.), Glycymeris sagittata (Gabb.), Solena aff. townsendensis Clark, Periploma circularis L. Krisht., Arca sp., Cucullaea beringiana Vol., Tudicula aff. copiosa Dev., Turritella aff. pachiесoensis Stant., Polinices nuciformis cowlitzensis (Dickerson) и др. Обращает на себя внимание скопление раковин Turritella, Crassatella и Arca (табл. I ). В этой же пачке отмечены фораминиферы (комплекс фораминифер 1) Cibicides pseudoungerianus evolutus Cushm. et Hobson, Dentalina dusenburyi Beck., D. colei Cushm. et Dusen, D. cf. legumen Reuss, Nodogenerina wegemanni (Cole), Pseudoglandulina cf. ovata d’Orb., Cyclammina cancellata obesa Cushm. et Laiming.
Таблица I . Двустворчатые и брюхоногие моллюски из ретырвеемской свиты (р. Ретырвеем). 1 – Tudicula aff. copiosa Devjatilova, ×1, вид со стороны, противоположной устью, обр. 139; 2–4 – Turritella aff. pachecoensis Stanton, ×1.5, обр. 138, пачка 1: 2, 4 – вид со стороны, противоположной устью, 3 – вид со стороны устья; 5 – Glycymeris sagittata (Gabb.), ×1, левая створка, обр. 56/2, пачка 1; 6 – то же, замок левой створки, ×2, обр. 55/1, пачка 1; 7 – Cucullaea beringiana Volobueva, ×1, левая створка, обр. 56/3, пачка 1; 8 – то же, замок левой створки, ×2, обр. 56/3а, пачка 1; 9 – Periploma circularis L. Krisht., ×1, левая створка, обр. 55/3б, пачка 1; 10 – Solena aff. townsendensis Clark., ×1, правая створка, обр. 55/3, пачка 1; 11 – то же, ×2, обр. 55/3а, пачка 1; 12, 15 – Polinices nuciformis cowlitzensis (Dickerson), ×1, обр. 55, пачка 1: 12 – вид со стороны устья, 15 – вид со стороны, противоположной устью; 13 – Tudicula aff. copiosa Devjatilova, ×1, вид со стороны, противоположной устью, обр. 55/1, пачка 1; 14 – то же, ×1, вид со стороны, противоположной устью, обр. 55/2а, пачка 1.

Таблица II. Двустворчатые и брюхоногие моллюски из моржовской свиты (р. Моржовая). 1 – Bathybembix sachalinensis (Takeda), × 1.5, вид со стороны, противоположной устью, обр. 51/5, нижняя часть свиты; 2 – Ancistrolepis aff. modestoides (Takeda), ×1.5, вид со стороны, противоположной устью, обр. 59/3, нижняя часть свиты; 3, 4 – Buccinum saundersi Martin., ×1.5, обр. 59/5, средняя часть свиты: 3 – вид со стороны, противоположной устью, 4 – вид со стороны устья; 5 – Smekhoviella matchgarensis (Makiyama), ×1, правая створка, обр. 59, средняя часть свиты; 6 – Mactra polynyma voyi (Gabb), ×1, левая створка, обр. 59/6, верхняя часть свиты; 7 – Cyclocardia expansa (Takeda), ×2, правая створка, обр. 14, нижняя часть свиты; 8 – Cyclocardia laxata (Yokoyama), ×2, левая створка, обр. 146, верхняя часть свиты; 9 – Macoma simizuensis L. Krisht., ×2, левая створка, обр. 51/6, средняя часть свиты; 10 – Thracia kavranensis Ilyina, ×1, левая створка, обр. 60/6, верхняя часть свиты; 11 – Mya arenaria Linne, ×1, левая створка, обр. 63/4а, верхняя часть свиты.

Пачка 2. Сложена такими же, как нижележащие, песчаниками темно-серыми, мелкозернистыми, с более редкими карбонатными конкрециями (0.5–0.6 м в диаметре), часто имеющими фиолетовую рубашку. Отмечаются маломощные линзочки гравелитов. Встреченные моллюски по составу близки к комплексу моллюсков пачки 1 и представлены Turritella, Arca, Solen, Polinices и единичными Modiolus sp. Остатки их более редкие, чем в пачке 1, и часто плохой сохранности. Мощность пачки около 250 м.
Пачка 3. Преобладают песчаники темно-серые, мелкозернистые, полимиктовые, с прослоями алевролитов (0.1–0.2 м), разделенными слоями песчаников через 1–3 м. Встречаются редкие карбонатные конкреции (обычно 0.1–0.2 м в диаметре) и отдельные тонкие прослои угля. Среди моллюсков встречены представители тех же Turritella, Arca, Solen, Yoldia, а также Mya sp., Pitaria sp. Мощность пачки около 150 м.
Общая мощность нижней подсвиты около 600 м.
Верхняя подсвита сложена серыми, желтоватыми песчаниками с частыми прослоями конгломератов и гравелитов. В ней выделены три пачки.
Пачка 1. Залегает на подстилающих породах нижней подсвиты согласно, но с размывом, который маркируется линзами гравелитов и мелкогалечных конгломератов. Песчаники серые, местами желтовато-серые, рыхлые, с отдельными сидеритовыми конкрециями (0.2–0.4 м). Отмечены прослои и линзы гравелитов. В верхней части пачки залегают пять прослоев черных лигнитов (каждый мощностью 0.5 м), разделенных слоями песчаников мощностью 1–5 м. Мощность пачки до 50–70 м.
Пачка 2. В нижней части мелкогалечные конгломераты и гравелиты мощностью до 10 м, с крепким песчано-карбонатным цементом. Над ними залегают песчаники серые, желтоватые, местами косослоистые, с линзами гравелитов. Мощность 40–50 м.
Пачка 3. Песчаники желтовато-серые, слоистые, с небольшими прослоями гравелитов. Часто отмечается их ритмическое чередование, мощность ритмов 3–4 м. Встречены редкие раковины моллюсков плохой сохранности: Turritella sp., Mya sp. Мощность пачки до 80–100 м.
Мощность верхней подсвиты 200 м. Общая мощность ретырвеемской свиты до 800 м.
Моржовская свита (нижний миоцен)
Свита залегает с размывом и стратиграфическим перерывом на ретырвеемской свите. В нижней части отмечается пачка конгломератов и песчаников мощностью 50–80 м. Над ней залегают однотипные полимиктовые песчаники светло-серые, желто-серые, рыхлые, средне-крупнозернистые. В них находятся серые песчано-карбонатные конкреции и прослои гальки. В гальке встречаются переотложенные обломки угля и конкреционные стяжения с раковинами Turritella, переотложенными из ретырвеемской свиты. Из-за относительно плохой обнаженности мощность свиты можно оценить только приблизительно до 850–1000 м.
В моржовской свите собран представительный комплекс моллюсков (табл. II). При этом намечается довольно четкая смена группировок с определенными доминантами. В нижней части свиты отмечены скопления Thyasira disjuncta ochotica L. Krisht. и Lucinoma acutilineata (Conrad). В средней части преобладают Yoldia aff. nabiliana Sim. В верхней части свиты характерны Clinocardium ornatocostatum Volobueva. В целом этот комплекс, названный комплексом моллюсков 2, резко отличается от ретырвеемского (комплекса моллюсков 1) и кроме вышеназванных видов включает следующие: Acila (Truncacila) gottschei Bohm., Yoldia chojensis Sim., Glycymeris boraensis Laut., Smechoviella matchgarensis (Mak.), Cyclocardia laxata (Yok.), C. expansa (Tak.), Clinocardium asagaiense (Mak.), C. chikagaganaensis eteretica Vol., Liocyma furtiva (Yok.), Mya elegans (Eiche), M. arenaria Linne, M. truncata Linne, Thracia kavranensis Ilyina, Macoma simizuensis L. Krisht., Periploma aff. besshoense (Yok.), Mactra polynyma voyi (Gabb.), Buccinum saundersi Martin, B. kurodai Kanehara, Bathybembix sachalinensis (Takeda), Ancistrolepis aff. modestoides (Takeda). В этой же свите определены фораминиферы (комплекс фораминифер 2): Reophax curtus Cushm., Haplophragmoides laminatus Vol., Melonis cf. tumiensis Kuzn., Perfectononion incertarformis Kuzn., P. pseudomartoobi Chalil., Astrononion hamadaensis Asano, Cribroelphidium sumitomoi (Asano et Murata), Pseudoelphidiella subcarinata Vol.
ОБСУЖДЕНИЕ И ВЫВОДЫ
Из изложенных материалов следует несколько выводов.
1. В разрезах кайнозоя восточной части Опухско-Пекульнейского прогиба развиты морские терригенные толщи, разделенные на две свиты в диапазоне нижнего эоцена–нижнего миоцена. Нижняя, ретырвеемская, свита представлена в основном черными и темно-серыми мелкозернистыми песчаниками с карбонатными конкрециями. Она охарактеризована комплексом моллюсков 1 и комплексом фораминифер 1. Среди моллюсков встречены в основном сублиторальные формы, отмеченные выше. Их сравнение с комплексами смежных территорий (Японии, Камчатки и Тихоокеанского побережья Северной Америки) показывает, что они являются в общем раннеэоценовыми и соответствуют кыланскому горизонту региональной стратиграфической схемы Корякского нагорья (Weaver et al., 1944; Девятилова, Волобуева, 1981; Решения…, 1998; Гладенков и др., 2013). На это указывает присутствие в комплексе моллюсков 1 Glycymeris sagittata (Gabb.), Solena aff. townsendensis Clark, Periploma circularis Krisht., Polinices nuciformis cowlitzensis (Dickerson), Tudicula aff. copiosa Devyatilova, Turritella aff. pacheoensis Stanton, Yoldia watasei Kan.
Основная часть видов фораминифер комплекса 1, по мнению С.И. Бордунова (с учетом данных В.Н. Беньямовского (Гладенков и др., 2013)), обычно встречается в диапазоне самых верхов палеоцена–нижнего эоцена Северо-Тихоокеанской области, включая Японию, Камчатку и Северную Америку. Особенно характерны Cibicides pseudoungerianus evolutus Cushm. et Hobson, Dentalina dusenburyi Beck., D. colei Cushm. et Dusen, D. cf. legumen Reuss, Nodogenerina wegemanni (Cole), Pseudoglandulina cf. ovate d’Orb., Cyclammina cancellata obese Cushm. et Laiming.
Верхи ретырвеемской свиты почти не содержат органических остатков: здесь отмечены только редкие плохо определимые раковины моллюсков. Поэтому возраст этих слоев определяется достаточно условно. Однако в них отмечены редкие фораминиферы, которые в общем сходны с комплексами более низких слоев.
Залегающая выше со стратиграфическим перерывом моржовская свита представлена светлыми, желтоватыми, часто рыхлыми песчаниками с конгломератами. Она охарактеризована комплексом моллюсков 2 сублиторального типа, заметно отличающимся от комплекса 1 и включающим виды, которые свойственны отложениям нижнего миоцена других районов, в частности Yoldia chojensis Sim., Thyasira disjuncta ochotica L. Krisht., Smekhoviella matchgarensis (Makiyama), Macoma simizuensis L. Krisht., Periploma aff. besshoense (Yok.), Cyclocardia expansa (Takeda), C. laxata (Yok.), Thracia kavranensis Ilyina, Ancistrolepsis aff. modestoides (Takeda), Buccinum saundersi Martin, B. kurodai Kanehara, Bathybembix sachalinensis (Takeda), Polinices ampla Philippi. Многие виды являются обычными для кулувенского и пахачинского горизонтов нижнего миоцена Западной и Восточной Камчатки (Гладенков, Синельникова, 1990), а некоторые из встреченных форм, такие как Mactra polynyma voyi (Gabb.), Mya arenaria Linne, M. truncata Linne, отмечены в неоген-плиоценовых толщах региона и отсутствуют в палеогене. Среди фораминифер встречаются также типичные для нижнего миоцена данного региона Haplophragmoides laminates Vol., Pseudoelphidiella subcarinata (Vol.), Melonis cf. tumiensis Kuzn., Astrononion hamadaense Asano, Perfectononion incertaeformis Kuzn., P. pseudomartobi Chalil, Cribroelphidium sumitomoi (Asano et Murata).
2. Из сказанного следует, что между ретырвеемской и моржовской свитами имеется значительный перерыв от среднего эоцена до олигоцена. Следует заметить, что в более западных частях Хатырского прогиба именно верхнеэоценовые и олигоценовые толщи получили значительное распространение (ионайская свита и ее аналоги). Это свидетельствует о том, что в позднеэоцен-олигоценовое время в разных частях Хатырского прогиба палеогеографические и седиментационные обстановки были неодинаковые. В западной части в целом накапливались грубообломочные отложения большой мощности (мощность малленской свиты превышает 6000 м), представленные песчано-конгломератовыми пачками, формирование которых связывают с активной деятельностью дельтовой системы р. Пра-Хатырки (Гладенков и др., 1985). В общей структуре осадочных комплексов здесь часто образовывались проградации, составляющие громадную линзу в шельфовой зоне. В средней части прогиба обычно одновозрастные слои представлены менее мощными алевролито-песчанистыми отложениями (часть хайидинской свиты), которые формировались в основном, видимо, в пределах сублиторали (Гладенков и др., 1985). Но, как следует из вышеприведенных данных, в самой восточной части прогиба накапливались терригенные образования сначала верхне-нижнесублиторального, а затем относительно мелководного типа. При этом на средне-позднеэоценовое и олигоценовое время пришелся перерыв в осадконакоплении. Неоднородность осадочных разрезов кайнозоя разных частей Опухско-Пекульнейского прогиба обусловлена, видимо, тектоническими процессами с их различным проявлением и активностью в этих частях.
3. Ископаемые моллюски и бентосные фораминиферы ретырвеемской свиты включают виды родов, которые типичны для относительно тепловодных субтропических сообществ. Среди моллюсков это представители Turritella, Crassotella, Tudicula, Glycymeris, Arca и др. Хорошо известно, что ранний эоцен был временем самого большого в кайнозое потепления (Гладенков и др., 2013). Это, видимо, отразилось на распространении теплолюбивых элементов морской биоты в шельфовых зонах Берингова моря. С этим явлением связывают появление термофильных наземных растений (в частности, пальм) на Северной Камчатке и даже на Чукотке (Ахметьев, 2015). Это же привело к появлению в морях бореальных широт карбонатного планктона – теплолюбивых фораминифер (Алабушева, 1996; Беньямовский, 2013). Поэтому наличие тепловодных моллюсков в изученных разрезах можно связать именно с этим потеплением и миграциями теплолюбивой фауны по мелководным зонам от южных морей (Японского) до более северных (Берингова). Обращает на себя внимание, что масштаб таких миграций мог быть достаточно заметным – вероятно, до 2000 км и более.
Полученные данные будут полезны для составления геологических карт нового поколения. При этом необходимо учитывать появившиеся сведения по строению шельфовой зоны Берингового моря на участке Опухско-Пекульнейского прогиба (Харахинов и др., 2014).
Благодарности. Автор приносит искреннюю благодарность С.И. Бордунову за анализ комплекса фораминифер из изученных разрезов. Он благодарит также А.С. Бякова, А.Б. Германа и С.В. Попова за полезные замечания к рукописи и обсуждение поднятых проблем.
Источники финансирования. Работа выполнена по теме государственного задания ГИН РАН № АААА-А18-118021690155-7 и при поддержке гранта РФФИ № 19-05-00361.
Список литературы
Алабушева А.В. Биостратиграфия и фораминиферы палеогеновых отложений Северо-Востока России (Хатырская и Анадырская впадины). Автореф. дисс… канд. геол.-мин. наук. Санкт-Петербург, 1996. 19 с.
Ахметьев М.А. Палеоген высокоширотных регионов Сибири и Северо-Востока России: стратиграфия, флора, климат и угленакопление // Стратиграфия. Геол. корреляция. 2015. Т. 23. № 4. С. 76–90.
Белова М.Б., Васильев В.Г., Власов Г.М. и др. Геологическое строение и перспективы нефтегазоносности Камчатки. М.: Гостоптехиздат, 1961. 344 с.
Беньямовский В.Н. Палеобиогеография бентосных фораминифер в палеоценовых акваториях средних и высоких широт Северной Евразии // Стратиграфия в начале ХХI века – тенденции и новые идеи. М.: Геокарт, ГЕОС, 2013. С. 145–167.
Богидаева В.И., Гладенков Ю.Б. Стратиграфия верхнемеловых и третичных отложений нижнего течения р. Хатырки // Материалы по геологии и полезным ископаемым Северо-Востока СССР. Магадан, 1963. Вып. 16. С. 192–196.
Бурлин Ю.К., Донцов В.В., Новиков Н.К. Особенности геологического строения и перспективы нефтегазоносности Анадырской и Нижнехатырской впадин на Чукотке. М.: ВНИИОЭНГ, 1973. 52 с.
Бушуев М.И. Геология и угленосность северо-восточной части Корякского хребта. М.: Водотранспорт, 1954. 130 с.
Волобуева В.И., Красный Л.Л. Маастрихт-неогеновые отложения восточной части Корякского нагорья. М.: Наука, 1979. 83 с.
Гладенков Ю.Б. Нефтепроявления нижнего течения р. Хатырки (Корякское нагорье) // Изв. АН СССР. Сер. геол. 1962. № 9. С. 101–106.
Гладенков Ю.Б., Синельникова В.Н. Моллюски и климатические оптимумы миоцена Камчатки. М. Наука, 1990. 174 с.
Гладенков Ю.Б., Волобуева В.И., Митрофанова Л.И. Третичные толщи Нижнехатырского прогиба (Корякское нагорье) // Изв. АН СССР. Сер. геол. 1985. № 3. С. 77–93.
Гладенков Ю.Б., Синельникова В.Н., Беньямовский В.Н., Фрегатова Н.А. Стратиграфия морского палеоцена и нижнего эоцена Западной Камчатки. М.: Геокарт–ГЕОС, 2013.160 с.
Девятилова А.Д., Волобуева В.И. Атлас фауны палеогена и неогена Северо-Востока СССР. М.: Недра, 1981. 219 с.
Егиазаров Б.Х., Дундо О.П., Аникеева Л.И., Русаков И.М., Дегтяренко Ю.П. Геология и полезные ископаемые Корякского нагорья. Л.: Недра, 1965. 342 с.
Решения рабочих межведомственных региональных стратиграфических совещаний по палеогену и неогену восточных районов России – Камчатки, Корякского нагорья, Сахалина и Курильских островов. Объяснительная записка к стратиграфическим схемам. М.: ГЕОС, 1998. 147 с.
Харахинов В.В., Шленкин С.И., Вашкевич А.А., Агапитов Д.Д., Обухов А.Н. Нефтегазоносные бассейны Беринговоморского региона (итоги нефтегазопоисковых работ 2000–2009 гг.). М.: Научный мир, 2014. 340 с.
Weaver C.E., Beek S., Bramlette M.N. Correlation of the Marine Cenozoic formations of Western North America // Bull. Geol. Soc. Am. 1944. V. 55. № 5. 268 p.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция