

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 5, стр. 101-115
Чинганджинская палеофлора и возраст меловых вулканитов междуречья Балыгычан–Сугой и прилегающей части Охотско-Колымского водораздела
С. В. Щепетов 1, *, Д. А. Юдова 1
1 Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
* E-mail: shchepetov@mail.ru
Поступила в редакцию 15.05.2018
После доработки 17.07.2019
Принята к публикации 20.11.2019
Аннотация
Большинство ископаемых флор Охотско-Чукотского вулканогенного пояса существенно отличается по систематическому составу от фаунистически датированных флор приморских низменностей соседнего Анадырско-Корякского субрегиона. Это сильно затрудняет их сопоставление и, соответственно, осложняет определение возраста вулканических накоплений. Исключение составляет лишь чинганджинская флора из одноименной свиты междуречья Вилига–Туманы (Омсукчанский район Магаданской области). Эта флора уверенно сопоставляется с приморскими флорами и ранее датировалась туроном–коньяком. Новые данные о систематическом составе чинганджинской флоры позволяют уточнить ее возраст. Она наиболее сходна с кайваямской флорой Анадырско-Корякского субрегиона и, следовательно, должна датироваться коньякским веком. Стратиграфическое положение местонахождений остатков растений чинганджинской флоры позволяет датировать коньякским веком три нижние в разрезе толщи вулканитов внешней зоны Охотско-Чукотского пояса. В ходе среднемасштабного геологического картирования в ряде случаев к ним ошибочно были отнесены и более молодые образования. Проникнуть далеко вглубь континента, сохранив свой приморский облик, чинганджинская флора могла по низменности, возможно возникшей в линейной зоне тектонических нарушений.
ВВЕДЕНИЕ
На Северо-Востоке России широко распространены континентальные отложения мелового возраста. Они заполняют несколько осадочных бассейнов и слагают огромную геологическую структуру – Охотско-Чукотский вулканогенный пояс (ОЧВП) (рис. 1). Планомерное изучение меловых вулканогенных образований началось в середине XX в., и сразу же встал вопрос об обосновании их возраста. Остатков морских моллюсков и фораминифер континентальные образования не содержат, споры и пыльца растений в вулканогенных толщах, как правило, не сохраняются, а изотопные методы тогда были мало пригодны для детальных стратиграфических корреляций. Однако в терригенных отложениях и туфовых прослоях вулканогенных толщ нечасто, но регулярно встречаются остатки ископаемых растений. К началу 1970-х годов был накоплен значительный палеофлористический материал, который впервые обобщила В.А. Самылина (1974), что дало начало бурному развитию меловой фитостратиграфии Северо-Востока Азии.
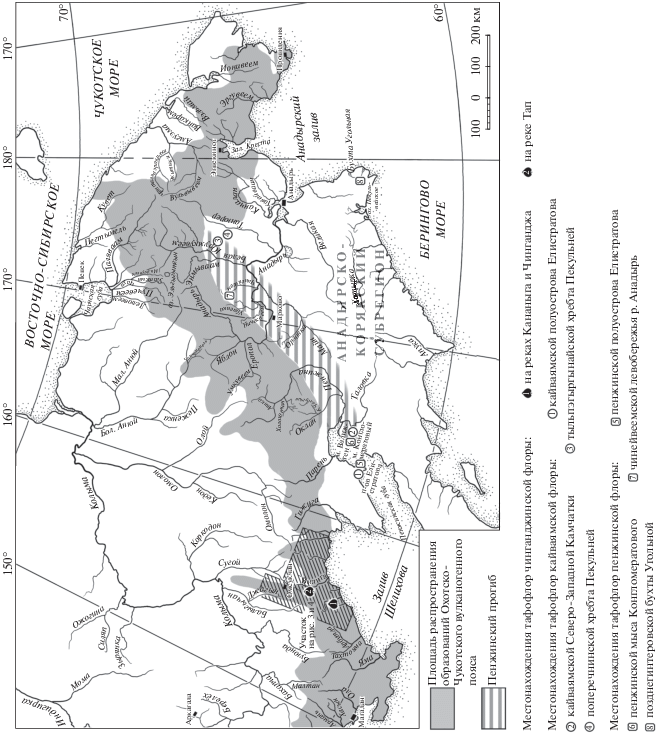
Рис. 1.
Местоположение захоронений остатков растений чинганджинской, пенжинской и кайваямской палеофлор на Северо-Востоке России.
В данной отрасли стратиграфии используются термины “палеофлористический комплекс” или “тафофлора” – это элементарные палеофлористические единицы, представляющие собой совокупность ископаемых растений из одного или нескольких территориально и стратиграфически близких местонахождений. Тафофлоры или палеофлористические комплексы, обладающие существенно сходными чертами, т.е. характерным сочетанием таксонов, качественным и количественным соотношением групп растений и т.д., мы, вслед за А.Б. Германом (2011), рассматриваем как ископаемую флору или палеофлору, характеризующую этап развития флоры значительной территории (например, Северного Приохотья).
О возрасте палеофлористических комплексов ОЧВП и, соответственно, о возрасте вмещающих вулканитов споры ведутся вот уже несколько десятилетий. Дело в том, что фаунистически датированные палеофлоры соседнего Анадырско-Корякского субрегиона существовали на приморских равнинах (Герман, 2011), а флоры ОЧВП – вдали от моря в условиях контрастного рельефа и активного вулканизма. Для палеофлор вулканической области характерно отсутствие или малое количество видов, встреченных в потенциально одновозрастных флорах приморских низменностей, а также значительное количество раннемеловых реликтов. Различия в систематическом составе сильно затрудняют сопоставление флор соседних регионов и часто делают фитостратиграфические корреляции условными. Исключение составляет лишь чинганджинская палеофлора из одноименной свиты междуречья Вилига–Туманы (Самылина, 1988; Головнева и др., 2011), которая по составу мало чем отличается от пенжинской и кайваямской палеофлор приморских низменностей (Герман, 1999, 2011). Это делает чинганджинскую флору одним из важнейших фитостратиграфических реперов неморского мела Северо-Востока России.
Первые сведения о систематическом составе комплекса одного из местонахождений чинганджинской флоры опубликовала В.А. Самылина (1974, 1976). Первоначально она сопоставляла комплекс р. Тап с гребенкинской флорой (Самылина, 1974, 1976), которая ныне датируется концом альба, сеноманом и началом турона (Герман, 2011). Позже В.А. Самылина (1984а, 1984б) и А.Б. Герман (1991) стали считать чинганджинскую флору туронской, то есть одновозрастной пенжинской флоре. Затем В.А. Самылина (1988) включила комплексы с рек Тап, Кананыга и Чинганджа в состав валижгенской стратофлоры, в которую, кроме прочего, вошли приморские пенжинская (турон, исключая начало) и кайваямская (коньяк) флоры. Понятие “валижгенская стратофлора” в дальнейшем не получило распространения в научной литературе, однако ныне большинство исследователей (Герман, 2011; Головнева и др., 2011; Щепетов, Герман, 2014, 2019; Herman et al., 2016 и др.) сравнивают чинганджинскую флору ОЧВП с пенжинской и кайваямской флорами Анадырско-Корякского субрегиона без разделения и датируют ее туроном–коньяком.
В 2002 г. в Санкт-Петербурге прошло Третье Межведомственное стратиграфическое совещание по докембрию, палеозою и мезозою Северо-Востока России. На нем было признано, что кайнофитная чинганджинская флора, в которой доминируют покрытосеменные и нет раннемеловых реликтов, одновозрастна арманской, в которой обильно представлены мезофитные формы растений (Решения…, 2009). Арманская палеофлора включает две локальных флоры – собственно арманскую из Примагаданья и зоринскую, известную из междуречий Балыгычан–Сугой и Вилига–Туманы (Herman et al., 2016). В предыдущей работе (Щепетов, 2019) подробно рассмотрено строение частных (в том числе флороносных) разрезов междуречья Вилига–Туманы и показано, что зоринская и чинганджинская флоры являются одновозрастными лишь в геологическом масштабе времени (период–эпоха–век). Их разделяет одно или два локальных вулканических события, которые могли длиться многие тысячи лет.
Монографическое описание чинганджинской флоры еще не завершено, однако авторы полагают, что новых данных о ее составе уже достаточно, чтобы провести более точное сравнение с одной из фаунистически датированных флор приморских низменностей. Анализ опубликованных в настоящее время картографических материалов позволяет, пусть иногда и условно, провести корреляцию и установить возраст вулканогенных толщ Охотско-Колымского водораздела между реками Аслачан и Наяхан.
СИСТЕМАТИЧЕСКИЙ СОСТАВ ЧИНГАНДЖИНСКОЙ ФЛОРЫ И ЕЕ СРАВНЕНИЕ С ИСКОПАЕМЫМИ ФЛОРАМИ БЛИЗКОГО ВОЗРАСТА
Чинганджинская флора включает растительные остатки из трех местонахождений (рис. 1). На р. Тап сборы проводили С.И. Филатов в 1959 г., Филатов и В.А. Самылина в 1965 г., В.Ф. Белый при участии В.В. Акинина и С.В. Щепетова в 1982 г. На р. Кананыга остатки растений собирали Белый при участии Акинина и Щепетова в 1982 г., Щепетов в 1984 г., а также Л.Б. Головнева, Щепетов и П.И. Алексеев в 2010 г. На р. Чинганджа сборы проводили Щепетов в 1984 г., Головнева, Щепетов и Алексеев в 2010 г. Собранные коллекции (№№ 519, 524, 538, 547, 548, 1563) хранятся в Ботаническом институте РАН (Санкт-Петербург).
Первый сводный список растений чинганджинской флоры по определениям Самылиной составил Щепетов (1988). После проведения дополнительных сборов в местонахождениях на реках Кананыга и Чинганджа в 2010 г. Головнева, Щепетов и Алексеев провели ревизию систематического состава чинганджинской флоры и опубликовали новый список ископаемых растений (Головнева и др., 2011). В этой же работе приводится детальное описание флороносных разрезов и местонахождений чинганджинской флоры. В настоящее время в составе данной флоры установлено 36 видов. Ниже отмечены изменения в списке Головневой и ее соавторов (2011), сделанные в результате более поздних работ (Юдова, Головнева, 2014, 2015; Юдова и др., 2016; Головнева и др., 2017; Shczepetov, Golovneva, 2009):
Marchantiophyta:
Thallites sp.;
Lycopodiophyta:
Selaginellites sp.;
Equisetophyta:
Equisetum sp.;
Polypodiophyta:
Asplenium dicksonianum Heer, A. rigidum Vassilevsk., Anemia ochotica Samyl., Arctopteris sp., Coniopteris opposita Samyl., C. tschuktschorum (Krysht.) Samyl., Cladophlebis inaequipinnulata Samyl., Osmunda tapensis Samyl., Lobifolia sp. (установлен впервые; Грабовский А.А., устное сообщение);
Pinophyta;
Caytoniales:
Sagenopteris sp.;
Ginkgoopsida:
Ginkgo ex gr. adiantoides (Ung.) Heer, G. ex gr. sibirica Heer;
Pinopsida:
Sequoia ochotica Yudova et Golovn. (новый вид, ранее – Sequoia sp. (Юдова, Головнева, 2015)), Metasequoia sp., Taxodium sp., Pityophyllum sp., Elatocladus sp., Ditaxocladus planiphyllus Guo et Sun, Cupressinocladus sp., Protophyllocladus subintegrifolius (Lesq.) Berry;
Magnoliophyta:
Trochodendroides tumanensis Yudova (новый вид, ранее – Trochodendroides sp. 2 (Юдова, Головнева, 2014)), T. deminii Yudova et Golovn. (новый вид, ранее – Trochodendroides sp. 1(Юдова, Головнева, 2014)), Trochodendroides sp., Dalembia argentea Yudova et Golovn. (новый вид, ранее – D. vachrameevii E. Lebedev et Herman (Юдова и др., 2016)), D. vachrameevii E. Lebedev et Herman, Menispermites sibirica (Heer) Golovn., Menispermites sp., Pseudoprotophyllum cordatum Samyl., Celastrophyllum kryshtofovichii Samyl., Cissites sp., Araliaephyllum speciosum Philipp. (ранее Araliaephyllum sp. 1), A. arenaria (Philipp.) Philipp. (ранее Araliaephyllum sp. 2), Ievlevia dorofeevii Samyl., Quereuxia angulata (Newb.) Krysht. et Baik.
Как видно из списка, в состав чинганджинской флоры входят печеночники, плауны, хвощи, папоротники (табл. I ), кейтониевые, гинкговые, хвойные (табл. II) и покрытосеменные (табл. III, рис. 2). Последние особенно разнообразны, им принадлежит чуть менее половины видового состава – 13 видов. Папоротники и хвойные менее разнообразны (по 8 видов). Остальные группы представлены 1–2 видами.
Рис. 2.
Отпечатки листьев Pseudoprotophyllum cordatum Samyl. чинганджинской флоры. а – экз. 1565/256; б – экз. 1565/305; в – экз. 1565/257; г – экз. 1565/310; д – экз. 1565/258; е – экз. 1565/261; ж – экз. 1565/317; з – экз. 1565/312; и – экз. 1565/285; к – экз. 1565/262; л – экз. 1565/291; м – экз. 1565/286; н – экз. 1565/409; о – экз. 1565/394; п – экз. 1565/410; р – экз. 1565/389; с – экз. 1565/272; т – экз. 1565/265; у – экз. 1565/269. Длина масштабной линейки 1 см.

Таблица I . Ископаемые печеночники и папоротники чинганджинской флоры. Длина масштабной линейки 1 см. 1 – Thallites sp., экз. 1565/227; 2 – Asplenium dicksonianum Heer, экз. 1565/202; 3 – Asplenium rigidum Vassilevsk., экз. 1565/205; 4, 5 – Anemia ochotica Samyl.: 4 – голотип, экз. 524/1; 5 – то же, фрагмент; 6, 7 – Coniopteris opposita Samyl.: 6 – голотип, экз. 538/174, 7 – экз. 1565/504; 8, 9 – Coniopteris tschuktschorum (Krysht.) Samyl.: 8 – экз. 1565/571, 9 – экз. 519/10; 10, 11 – Cladophlebis inaequipinnulata Samyl.: 10 – голотип, экз. 519/12, 11 – экз. 519/13; 12 – Osmunda tapensis Samyl., голотип, экз. 519/1; 13, 14 – Lobifolia sp.: 13 – экз. 1565/500, 14 – то же, фрагмент.

Таблица II. Ископаемые голосеменные чинганджинской флоры. Длина масштабной линейки 1 см. 1 – Sagenopteris sp., экз. 1565/585; 2 – Ginkgo ex gr. adiantoides (Ung.) Heer, экз. 1565/357; 3, 4 – Ginkgo ex gr. sibirica Heer: 3 – экз. 538/160, 4 – экз. 1565/502; 5–7 – Sequoia ochotica Yudova et Golovn.: 5 – голотип, экз. 1565/451, 6 – то же, фрагмент, 7 – экз. 1565/67; 8 – Sequoia sp., шишка, экз. 1565/169; 9–11 – Metasequoia sp.: 9 – экз. 1565/454а, 10 – экз. 1565/567, 11 – экз. 1565/454; 12–14 – Taxodium sp.: 12 – экз. 1565/473а, 13 – экз. 1565/473b, 14 – экз. 1565/107; 15–17 – Cupressinocladus sp.: 15 – экз. 538/137, 16 – то же, фрагмент, 17 – экз. 1565/67.

Таблица III. Ископаемые покрытосеменные чинганджинской флоры. Длина масштабной линейки 1 см. 1 – Trochodendroides tumanensis Yudova, голотип, экз.1565/27; 2–3 – Trochodendroides deminii Yudova et Golovn.: 2 – голотип, экз. 1565/233, 3 – экз.1565/234; 4 – Trochodendroides sp., экз. 1565/10; 5–7 – Dalembia argentea Yudova et Golovn.: 5 – апикальный и латеральный листочки, голотип, экз. 1565/355, 6 – латеральный листочек, экз. 1565/351, 7 – латеральный листочек, экз. 1565/357; 8 – Dalembia vachrameevii E. Lebedev et Herman, экз. 1565/352; 9 – Menispermites sp., экз. 1565/513; 10 – Celastrophyllum kryshtofovichii Samyl., голотип, экз. 519/63; 11 – Cissites sp., экз. 1565/358; 12 – Araliaephyllum speciosum Phillip., экз. 1565/373; 13 – Araliaephyllum arenaria (Phillip.) Phillip., экз. 1565/495; 14 – Quereuxia angulata (Newb.) Krysht. et Baik., фрагменты листочков, экз. 1565/321.

Как уже отмечалось, чинганджинская флора наиболее сходна с палеофлорами приморских низменностей Анадырско-Корякского субрегиона пенжинского и кайваямского этапов развития. К первому из них относятся пенжинская тафофлора Северо-Западной Камчатки, пенжинская тафофлора полуострова Елистратова, чинейвеемская тафофлора левобережья среднего течения р. Анадырь и позднегинтеровская тафофлора Беринговского полуострова (Герман, 2011) (рис. 1). Покрытосеменные в этих палеофлорах составляют чуть больше половины таксономического состава. Среди них преобладают крупнолистные платанообразные – Ettingshausenia, Arthollia, Paraprotophyllum, возможно, Pseudoprotophyllum и Credneria. Обычными компонентами пенжинских тафофлор являются представители родов Menispermites, Celastrophyllum, Trochodendroides. Хвойные многочисленны, чаще всего встречаются Cephalotaxopsis и Sequoia. Появляется Metasequoia.
У чинганджинской и пенжинской флор общими являются роды Arctopteris, Asplenium (A. dicksonianum Heer), Cladophlebis, Ginkgo (G. ex gr. adiantoides (Ung.) Heer), Pityophyllum, Sequoia (S. ochotica Yudova et Golovn. = S. tenuifolia (Schmalh.) Sveshn. et Budants.), Metasequoia, Elatocladus, Protophyllocladus, Pseudoprotophyllum, Menispermites, Celastrophyllum, Cissites, Trochodendroides, Dalembia. В пенжинских тафофлорах большим разнообразием отличаются представители сем. Platanaceae (16 видов), но род Pseudoprotophyllum не занимает доминирующего положения. В чинганджинской флоре также заметную роль играют платанообразные, но листья Pseudoprotophyllum cordatum в некоторых прослоях образуют листовые кровли. В пенжинской флоре встречены четыре вида рода Menispermites, а род Araliaephyllum отсутствует. В чинганджинской флоре установлено только два вида Menispermites, но выделяется два вида Araliaephyllum – A. speciosum и A. arenaria.
Палеофлора кайваямского этапа развития включает одноименную тафофлору Северо-Западной Камчатки, кайваямскую тафофлору полуострова Елистратова, поперечнинскую тафофлору восточного склона хребта Пекульней и тыльпэгыргынайскую тафофлору западного склона того же хребта (Герман, 1999, 2011) (рис. 1). В перечисленных палеофлористических комплексах покрытосеменные составляют не менее половины списочного состава. Платанообразные доминируют, как и в пенжинской флоре, но в основном это виды родов Paraprotophyllum и Pseudoprotophyllum. Заметна роль Trochodendroides и Araliaephyllum, видовое разнообразие которых очень велико, а вот представители родов Menispermites и Celastrophyllum, по сравнению с пенжинской флорой, не так разнообразны. Цикадофиты – Nilssonia yukonensis и Ctenis sp. – установлены только в составе тафофлор хребта Пекульней. Среди хвойных многочисленны находки Cephalotaxopsis и Sequoia в сочетании с Metasequoia, Elatocladus, Cupressinocladus, Glyptostrobus.
Общими для чинганджинской флоры и тафофлор кайваямского этапа являются роды Equisetum, Osmunda, Arctopteris, Asplenium, Cladophlebis, Ginkgo, Elatocladus, Sequoia, Metasequoia, Cupressinocladus, Pityophyllum, Menispermites, Araliaephyllum, Celastrophyllum, Cissites, Trochodendroides, Dalembia, Quereuxia. В кайваямской флоре виды Araliaephyllum speciosum и A. arenaria являются общими с чинганджинской флорой (табл. III, фиг. 12, 13 ). В последней установлено три вида Trochodendroides, два из них недавно описаны как новые – T. tumanensis (табл. III, фиг. 1) и T. deminii (табл. III, фиг. 2–3) (Юдова, Головнева, 2014). Именно они, по нашему мнению, представляют род Trochodendroides в тыльпэгыргынайской тафофлоре хребта Пекульней. В отличие от пенжинских тафофлор, в кайваямских присутствует водное растение Quereuxia angulata, которое обильно представлено в чинганджинской флоре (табл. III, фиг. 14 ).
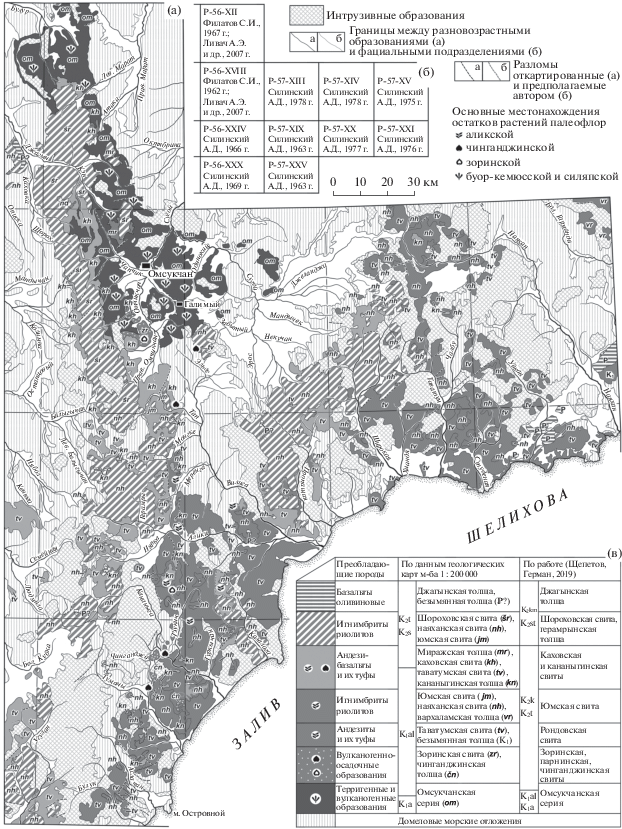
Рис. 3.
Схема распространения меловых отложений в междуречье Балыгычан–Сугой и прилегающей части Охотско-Колымского водораздела (а), разграфка использованных листов геологических карт масштаба 1 : 200 000 (б) и стратиграфическая последовательность меловых отложений района (в).
Проведенное сравнение говорит в пользу того, что чинганджинская флора более сходна с тафофлорами кайваямского этапа развития, чем пенжинского, и датировать ее следует коньякским веком.
Как показано выше, в междуречье Вилига–Туманы отложения с остатками растений чинганджинской флоры залегают на одном стратиграфическом уровне со слоями, включающими зоринскую палеофлору. Общими для зоринской и чинганджинской флор являются Thallites sp. (табл. I , фиг. 1), Equisetum sp., Osmunda tapensis (табл. I , фиг. 12 ), Asplenium dicksonianum (табл. I , фиг. 2), Arctopteris sp., Cladophlebis inaequipinnulata (табл. I , фиг. 10, 11 ), Sagenopteris sp. (табл. II, фиг. 1), Ginkgo ex gr. adiantoides (табл. II, фиг. 2), Ginkgo ex gr. sibirica (табл. II, фиг. 3, 4), Sequoia ochotica (табл. II, фиг. 5–7 ), Taxodium sp. (табл. II, фиг. 12–14 ), Pityophyllum sp., Elatocladus sp., Trochodendroides tumanensis (табл. III, фиг. 1). Кроме того, их объединяет наличие в систематическом составе представителей родов Menispermites, Cissites, Pseudoprotophyllum, Dalembia. Однако в зоринской флоре разнообразие покрытосеменных значительно меньше – только восемь видов. Причем в захоронениях они чрезвычайно редки и могут быть обнаружены лишь в результате целенаправленных поисков. По количеству отпечатков в зоринских захоронениях иногда резко доминируют раннемеловые реликты Sphenobaiera, Вaiera, Phoenicopsis, Podozamites, отсутствующие в составе чинганджинской флоры.
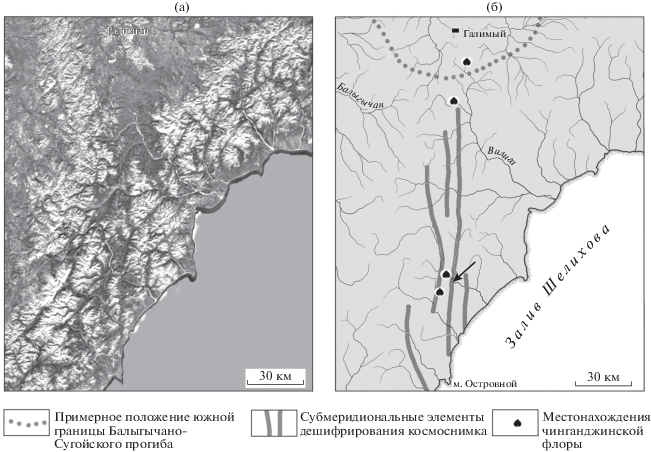
Рис. 4.
(а) Вид из космоса южной части междуречья Балыгычан–Сугой и прилегающего участка Охотско-Колымского водораздела, а также (б) положение субмеридиональных элементов дешифрирования космоснимка и местонахождений чинганджинской флоры (стрелкой показано местоположение современной сейсмодислокации).
Слои с чинганджинской и зоринской флорами в междуречье Вилига–Туманы перекрываются туфами и лавами амфиболовых и амфибол-пироксеновых андезитов, выше залегают вулканиты кислого состава, а венчают разрез района туфы и лавы двупироксеновых андезибазальтов. Из этих трех толщ происходят остатки растений аликской флоры (рис. 3в). В ней мало цветковых растений и велико участие реликтовых элементов: Hausmannia, Lobifolia, Heilungia, Phoenicopsis, Sphenobaiera, Ginkgo ex gr. sibirica (Щепетов, Головнева, 2010). Общими для аликской и чинганджинской флор являются Coniopteris tschuktschorum, Asplenium dicksonianum, Ginkgo ex gr. adiantoides, Dalembia argentea, роды Lobifolia, Sequoia, Metasequoia, Menispermites и Trochodendroides.
КОРРЕЛЯЦИЯ ВУЛКАНОГЕННЫХ ТОЛЩ МЕЖДУРЕЧЬЯ БАЛЫГЫЧАН–СУГОЙ И ПРИЛЕГАЮЩЕЙ ЧАСТИ ОХОТСКО-КОЛЫМСКОГО ВОДОРАЗДЕЛА
На большей части внешней зоны ОЧВП наблюдается гомодромная последовательность вулканизма, включающая пять этапов (Устиев, 1949, 1959; Белый, 1977; Акинин, Миллер, 2011 и др.). Для продуктов извержений первого этапа в целом характерен относительно разнообразный состав – лавы и туфы основного и среднего состава при доминировании последних. На втором этапе преимущественно извергались лавы и туфы кислого состава. В толщах, сформированных на третьем этапе вулканизма, преобладают лавы и туфы двупироксеновых андезибазальтов. Затем следует толща игнимбритов, туфов и лав кислого состава. Для субщелочных базальтов и андезибазальтов завершающей стадии развития вулканизма ОЧВП характерно присутствие оливинов в качестве темноцветных вкрапленников. Кислые вулканиты второго и четвертого этапов вулканизма мало различаются по геохимическому и петрографическому составу. Три нижние толщи гомодромной последовательности представляют ранний этап развития ОЧВП (Белый, 1977; Белый, Белая, 1998). По современным представлениям они датируются туроном–коньяком (Щепетов, Герман, 2019). Две верхние толщи представляют поздний этап развития вулканизма пояса (Белый, 1977; Белый, Белая, 1998) и датируются сантоном–кампаном (Щепетов, Герман, 2019).
В пределах междуречья Балыгычан–Сугой образования ОЧВП подстилаются накоплениями омсукчанской серии, заполняющей субмеридиональный Балыгычано-Сугойский прогиб (Матвеенко, 1957) (рис. 3). Серия включает (снизу вверх) аскольдинскую свиту кислых вулканитов, базальтовую арылахскую толщу, а также терригенные, частично угленосные уликскую, галимовскую, айгурскую и топтанскую свиты (Ливач и др., 2009). В арылахской толще остатков растений не обнаружено. Аскольдинская и уликская свиты содержат растительные остатки силяпской флоры аптского возраста, остальные свиты включают многочисленные остатки буор-кемюсской флоры (Самылина, 1976; Головнева и др., 2017), существовавшей в альбе и, возможно, в сеномане и туроне (Щепетов, Герман, 2019).
На терригенных отложениях омсукчанской серии залегает локально распространенная зоринская свита, включающая остатки растений одноименной флоры. Последняя входит в состав региональной арманской флоры (Щепетов, Головнева, 2014). Стратиграфические аналоги зоринской свиты – парнинская и чинганджинская свиты – также имеют локальное распространение и залегают непосредственно на эродированной поверхности юрских отложений. Стратиграфически выше располагаются вулканогенные толщи ОЧВП.
Тафофлоры чинганджинской флоры с рек Кананыга и Чинганджа происходят из отложений одноименной свиты в основании вулканогенного разреза, а местонахождение на р. Тап приурочено, скорее всего, к базальным слоям третьей в разрезе вулканогенной толщи (Щепетов, 1995, 2019). Аналогичное положение занимают и остатки растений с левобережья р. Эльге. Там удалось обнаружить только обрывки довольно крупных листьев покрытосеменных. Этот сильно обедненный комплекс можно отнести к чинганджинской флоре лишь условно – просто потому, что в местонахождениях зоринской флоры, в отличие от чинганджинской, покрытосеменные встречаются редко. Таким образом, есть основания полагать, что возраст трех первых толщ гомодромной последовательности вулканизма ОЧВП в рассматриваемом районе соответствует коньякскому веку. В этих же толщах в маломощных прослоях туффитов и тефроидов содержатся остатки растений еще одной палеофлоры – аликской, которая не имеет аналогов за пределами ОЧВП и датирована коньякским веком по косвенным данным. Она обладает определенными чертами сходства не только с чинганджинской флорой, но и с палеофлорой из чаунской серии Чукотки, коньякский возраст которой определен изотопными методами (Kelley et al., 1999; Щепетов, Головнева, 2010).
На рассматриваемой территории распространены все пять толщ гомодромной последовательности вулканизма ОЧВП (рис. 3в). Однако в строении вулканических полей обычно участвуют только две, реже три толщи пород различного состава (Щепетов, 1991, 1995). При этом любая из толщ общей последовательности в конкретном районе может выпадать из разреза. Кроме того, среднемасштабное геологическое картирование данной части Охотско-Колымского водораздела велось в 60-х и 70-х гг. ХХ в., когда считалось, что здесь представлены только три меловых вулканогенных толщи: таватумская свита вулканитов среднего состава, наяханская свита риолитов и локально распространенная кананыгинская толща вулканитов среднего состава, то есть в разрезе присутствует только одна кислая толща. Позже чинганджинская и кананыгинская толщи были описаны как свиты (Щепетов, 1988), однако, анализируя старые карты, здесь мы используем прежние названия.
С высокой долей уверенности можно предположить, что в тех случаях, когда из разреза выпадает первая кислая толща, все вулканиты среднего состава показаны на картах как таватумская свита, поскольку состав пород сходен. Если же в разрезе отсутствуют две нижние толщи, то третья толща андезибазальтов и андезитов показана как первая – все та же таватумская свита. По мнению авторов, первоначально именно такой была ситуация в междуречье Балыгычан–Сугой (Геологическая…, 1965), где выходы пород ОЧВП образуют субмеридиональный апофиз вглубь материка. В новой редакции геологических карт P-56-XII и P-56-XVIII (Ливач и др., 2009) выделены все толщи гомодромной последовательности вулканизма ОЧВП, за исключением финальных базальтов, которые на данной территории просто отсутствуют. Большая часть вулканитов “наяханской свиты” отнесена к шороховской свите (четвертая толща в общем разрезе, она же вторая кислая), а “таватумской свиты” – к каховской (первая толща общего разреза). С.В. Щепетову (1995, 2019) кажется более вероятным, что основной объем вулканитов среднего состава представляет здесь третью толщу гомодромной последовательности, поскольку во всех изученных им частных разрезах преобладают лавы и туфы с вкрапленниками и кристаллокластами клино- и ортопироксенов.
На рассматриваемой территории Охотско-Колымского водораздела в ряде случаев “лишние” толщи на картах зафиксированы как фациальные разности в составе таватумской или наяханской свит. Так, на западной границе листа P-57-XXV – на правобережье нижнего течения р. Кананыга – показаны (снизу вверх): андезиты (таватумская свита), риолиты, вновь андезиты (то и другое – наяханская свита). Территория распространения верхних андезитов примыкает к полю распространения таватумской свиты на листе P-56-XXX, которая перекрывает чинганджинскую толщу. Это дает основание (пусть и слабое!) предположить, что на флороносных вулканогенно-осадочных отложениях залегает здесь не первая в разрезе толща вулканитов среднего состава, как в местонахождениях на реках Чинганджа и Кананыга, а вторая толща, как, вероятно, на р. Тап (Щепетов, 2019).
По данным геологических карт масштаба 1 : : 200 000 (рис. 3а), базальты самой верхней толщи гомодромной последовательности на территории встречаются редко. Обычно они слагают небольшие по площади останцы на вершинах водоразделов. К юго-западу от рассматриваемой территории, в Примагаданье, такие образования (мыгдыкитская свита) подстилаются слоями с остатками растений ольской флоры (Белый, Самылина, 1987), датируемой сантоном–кампаном (Герман, 2011; Щепетов, Герман, 2019). По наблюдениям Щепетова (1991, 1995), оливиновые базальты на левобережье р. Джагын ничем не отличаются от пород мыгдыкитской свиты Примагаданья, но обнаружить остатки растений там не удалось. По данным А.Д. Силинского (Государственная…, 1977), в бассейне р. Уйкан покровы двупироксеновых и оливиновых базальтов подстилаются терригенными отложениями с остатками растений, которые А.Ф. Ефимова датировала палеогеном. Мы не имеем возможности ознакомиться с этой коллекцией, поэтому не можем исключить, что в этом районе был еще один – более поздний – этап базальтового вулканизма.
Суммируя изложенное выше, можно утверждать, что на рассматриваемой территории коньякские вулканогенные накопления закартированы как таватумская, наяханская, каховская и юмская свиты, кананыгинская, миражская и вархаламская толщи. Сантон-кампанские вулканиты представлены шороховской свитой, а в ряде случаев картировались как наяханская и юмская свиты. Такой же возраст имеет по крайней мере часть венчающих разрез оливиновых базальтов. В легенде для крупномасштабных геологических карт междуречья Балыгычан–Сугой они получили название “джагынская толща”.
КАК СМОГЛА ПРИМОРСКАЯ ФЛОРА ПОПАСТЬ ВГЛУБЬ КОНТИНЕНТА?
Тафофлоры зоринской флоры и, по крайней мере, две тафофлоры чинганджинской флоры располагаются на одном стратиграфическом уровне, однако сами эти палеофлоры существовали не одновременно. В бассейнах рек Сугой, Вилига, Кананыга и Туманы вулканогенно-осадочные слои с остатками зоринской и чинганджинской палеофлор непосредственно подстилают разрез вулканогенных образований внешней зоны Охотско-Чукотского пояса. Однако стратиграфические и петрографические данные свидетельствуют в пользу того, что в междуречье Вилига–Туманы после формирования слоев с остатками растений зоринской флоры имела место локальная вспышка вулканизма умеренно-кислого состава, затем на поверхность стали поступать амфиболовые лавы и туфы среднего состава. Лишь после этого началось формирование слоев с остатками растений чинганджинской флоры (Щепетов, 2019).
Появление полностью лишенной реликтовых элементов чинганджинской флоры на территории, где уже существовала зоринская флора, в которой, наряду с покрытосеменными, обильно представлены раннемеловые формы растений, требует рационального объяснения. Исходя из реконструкций В.Ф. Белого (1994), можно предположить, что под водой внешняя (относительно материка) граница ОЧВП проходит в нескольких десятках километров от современного берега примерно параллельно северо-западному побережью залива Шелихова. Не будет большой натяжкой допустить, что к юго-востоку от этой границы располагалось продолжение Пенжинского прогиба с его приморскими низменностями, заселенными кайнофитными сообществами растений. С этих низменностей покрытосеменные и сопутствующие им группы растений в туроне–коньяке проникали вглубь континента, смешиваясь с растениями буор-кемюсской флоры и постепенно вытесняя их. Данный эпизод флорогенеза представлен в геологической летописи тафофлорами парнинской и зоринской свит, которые относятся к арманскому этапу развития флоры. На космоснимках и мелкомасштабных топографических картах хорошо видно, что в полосе шириной около 60 км вдоль современного морского побережья речные долины и водоразделы ориентированы преимущественно с юго-запада на северо-восток и с северо-запада на юго-восток. Однако на линии пос. Галимый–мыс Островной появляются элементы ландшафта с субмеридиональной ориентировкой (рис. 4). На среднемасштабных геологических картах (рис. 3) эти “линеаменты” почти никак не выражены (если не считать таковым выражением присутствие в разрезах отложений с остатками растений чинганджинской флоры), однако на местности здесь наблюдаются относительно недавно перехваченные (резко изменившие направление) русла рек и палеодолины.
Можно предположить, что в коньякском веке произошло некое тектоническое событие. Возможно, это была серия сейсмодислокаций, подобных той, что относительно недавно резко повернула русло р. Кананыга на юго-восток близ устья руч. Ронд (рис. 4б). За прошедшие несколько сотен (или тысяч?) лет в районе этой дислокации на площади в несколько квадратных километров современная растительность так и не смогла полностью восстановиться. В начале формирования ОЧВП сейсмоактивность, конечно, была гораздо выше современной. Серия подобных сейсмодислокаций вдоль субмеридиональных тектонических нарушений вполне могла создать протяженное субмеридиональное понижение рельефа или полосу разрушенного ландшафта, в которой местная растительность была уничтожена или сильно пострадала. Вот по этой полосе с приморских низменностей кайваямская флора и проникла вглубь континента, не встречая на своем пути неповрежденных, инвазионно устойчивых сообществ местных растений.
Примерно в это же время начала свое существование и аликская флора. Она включает шесть тафофлор из вулканогенных толщ (Щепетов, Головнева, 2010). Местонахождение самой представительной из них – тэукичского флористического комплекса – расположено примерно в 15 км к северо-востоку от местонахождения чинганджинской флоры на р. Кананыга (рис. 3). Остатки растений происходят из слоев тефроидов амфиболовых андезитов, которые залегают непосредственно на дислоцированных отложениях юры (Щепетов, 2019, рис. 1). По-видимому, аликская флора включает растения, которые наиболее активно осваивали ювенильные поверхности формирующихся вулканических полей. Там, где появлялись такие поля, чинганджинская флора прекращала свое существование – комплексы ее растений переставали попадать в захоронения. По представлениям Щепетова (1995, 2019), в северо-западной части рассматриваемого района изверженный материал стал поступать на дневную поверхность лишь на третьем этапе вулканизма, и чинганджинская флора существовала здесь более длительное время, чем на юге.
ЗАКЛЮЧЕНИЕ
По систематическому составу чинганджинская флора ОЧВП ранее сопоставлялась с фаунистически датированными пенжинской и кайваямской флорами Анадырско-Корякского субрегиона и считалась турон-коньякской. За последние годы в ее составе описано несколько новых видов, пересмотрены некоторые прежние определения. С учетом этих данных авторы полагают, что чинганджинская флора обладает наибольшими чертами сходства именно с кайваямской флорой и, соответственно, должна датироваться коньякским веком.
Стратиграфическое положение остатков растений чинганджинской флоры позволяет датировать коньякским веком первые три толщи гомодромной последовательности вулканизма ОЧВП. В ходе среднемасштабного геологического картирования междуречья Балыгычан–Сугой и примыкающей части Охотско-Колымского водораздела в ряде случаев к ним были отнесены и более молодые образования, которые следует датировать сантоном–кампаном. Исправить это, вероятно, можно лишь в ходе новых полевых исследований.
Остается неясным, каким образом приморская по составу чинганджинская флора смогла оказаться далеко в глубине материка, не приняв при этом в свой состав ни одного местного реликтового вида. Следы “залеченных” субмеридиональных тектонических нарушений, дешифрируемые на космоснимках, позволяют предположить, что в коньякском веке здесь возникло протяженное понижение рельефа. В нем могли сформироваться речные долины, по которым покрытосеменные и сопутствующие им группы растений стремительно продвинулись на север, не ассимилируя в пути местные растительные сообщества.
Благодарности. За помощь при подготовке статьи авторы выражают искреннюю признательность А.Б. Герману и М.Г. Моисеевой (Геологический институт РАН, Москва).
Источники финансирования. Работа выполнена в рамках государственного задания Ботанического института им. В.Л. Комарова РАН “Ископаемые растения России и сопредельных территорий: систематика, филогения, палеофлористика и палеофитогеография” (АААА-А19-119021190031-8), а также при поддержке РФФИ, грант № 19-05-00121.
Список литературы
Акинин В.В., Миллер Э.Л. Эволюция известково-щелочных магм Охотско-Чукотского вулканогенного пояса // Петрология. 2011. Т. 19. № 3. С. 249–290.
Белый В.Ф. Стратиграфия и структуры Охотско-Чукотского вулканогенного пояса. М.: Наука, 1977. 171 с.
Белый В.Ф. Геология Охотско-Чукотского вулканогенного пояса. Магадан: СВКНИИ ДВО РАН, 1994. 76 с.
Белый В.Ф., Белая Б.В. Поздняя стадия развития Охотско-Чукотского вулканогенного пояса (верхнее течение р. Энмываам). Магадан: СВКНИИ ДВО РАН, 1998. 108 с.
Белый В.Ф., Самылина В.А. О заключительном этапе развития Охотско-Чукотского вулканогенного пояса // Тихоокеанская геология. 1987. № 5. С. 76–85.
Геологическая карта СССР. Серия Верхнеколымская. Лист P-56-XVIII. Объяснительная записка. М.: Недра, 1965. 65 с.
Герман А.Б. Меловая флора Анадырско-Корякского субрегиона (Северо-Восток России): систематический состав, возраст, стратиграфическое и флористическое значение. М.: ГЕОС, 1999. 122 с.
Герман А.Б. Альбская–палеоценовая флора Северной Пацифики. М.: ГЕОС, 2011. 280 с. (Тр. ГИН РАН. Вып. 592).
Головнева Л.Б., Щепетов С.В., Алексеев П.И. Чинганджинская флора (поздний мел, Северо-Восток России): систематический состав, палеоэкологические особенности и стратиграфическое значение // Чтения памяти А.Н. Криштофовича. 2011. Вып. 7. С. 37–61.
Головнева Л.Б., Алексеев П.И., Гниловская А.А., Юдова Д.А. Род Trochodendroides (Cercidiphyllaceae) в позднемеловых флорах Северо-Востока России // Палеоботаника. 2017. Т. 8. С. 122–179.
Государственная геологическая карта СССР. Серия Магаданская. Лист P-57-XXI. Объяснительная записка. Москва, 1977. 55 с.
Ливач А.Э., Третьякова Н.И., Чуравцов А.П. Государственная карта РФ масштаба 1 : 200 000 (издание второе), серия Сугойская. P-56-XVIII (Омсукчан). Объяснительная записка. М., 2009. 88 с.
Матвеенко В.Т. Петрология и общие черты металлогении Омсукчанского рудного узла (Северо-Восток СССР) // Тр. ВНИИ-1. Геол. 1957. Вып. 31. 73 с.
Решения 3-го Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России (Санкт-Петербург, 2002). СПб.: ВСЕГЕИ, 2009. 266 с.
Самылина В.А. Раннемеловые флоры Северо-Востока СССР (к проблеме становления флор кайнофита) // ХХVII Комаровские чтения. Л.: Наука, 1974. 56 с.
Самылина В.А. Меловая флора Омсукчана. Л.: Наука, 1976. 207 с.
Самылина В.А. О меловой флоре с р. Кананыга (Северное Приохотье) // Стратиграфия и палеонтология палеозойских и мезозойских отложений Северо-Востока СССР. М.: Геол. фонд РСФСР, 1984а. С. 178–190.
Самылина В.А. Позднемеловая флора р. Тап (Северное Приохотье) // Ежегодник Всесоюзного палеонтол. об-ва. Т. 27. М.: ВПО, 1984б. С. 236–247.
Самылина В.А. Аркагалинская стратофлора Северо-Востока Азии. Л.: Наука, 1988. 131 с.
Устиев Е.К. Мезозойский и кайнозойский магматизм Северо-Востока СССР // Материалы по геологии и полезным ископаемым Северо-Востока СССР. Магадан. 1949. Вып. 4. С. 30–51.
Устиев Е.К. Охотский тектоно-магматический пояс и некоторые связанные с ним проблемы // Сов. геол. 1959. № 3. С. 2–26.
Щепетов С.В. Стратиграфия меловых континентальных накоплений Омсукчанского района (Северное Приохотье). Магадан: СВКНИИ ДВО АН СССР, 1988. 60 с.
Щепетов С.В. Стратиграфия континентального мела юго-западного фланга Колымского нагорья. Магадан: СВКНИИ ДВО РАН, 1991. 160 с.
Щепетов С.В. Стратиграфия континентального мела Северо-Востока России. Магадан: СВКНИИ ДВО РАН, 1995. 122 с.
Щепетов С.В. Об относительном возрасте зоринской и чинганджинской флор позднего мела Северо-Востока России // Тихоокеанская геология. 2019. Т. 38. № 4. С. 35–44.
Щепетов С.В., Герман А.Б. Проблемы биостратиграфии континентального мела Северо-Востока Азии // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Сборник научных трудов. Ред. Барабошкин Е.Ю., Маркевич В.С., Бугдаева Е.В., Афонин М.А., Черепанова М.В. Владивосток: Дальнаука, 2014. С. 331–335.
Щепетов С.В., Герман А.Б. К вопросу о стратиграфии и флорах неморского мела Северо-Востока России // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 3. С. 40–52.
Щепетов С.В., Головнева Л.Б. Позднемеловая флора из вулканогенных образований Северного Приохотья (Охотско-Чукотский вулканогенный пояс) // Палеоботаника. 2010. Т. 1. С. 45–95.
Щепетов С.В., Головнева Л.Б. Зоринская флора Северного Приохотья (поздний мел) и проблема фитостратиграфии нижней части разреза Охотско-Чукотского вулканогенного пояса // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 4. С. 52–66.
Юдова Д.А., Головнева Л.Б. Род Trochodendroides Berry в чинганджинской флоре Северо-Востока России // Палеоботаника. 2014. Т. 5. С. 60–72.
Юдова Д.А., Головнева Л.Б. Новый вид рода Sequoia Endli (Cupressaceae) из позднемеловых отложений Северо-Востока России // Палеоботаника. 2015. Т. 6. С. 80–95.
Юдова Д.А., Головнева Л.Б., Алексеев П.И. Распространение рода Dalembia E. Lebedev et Herman (Magnoliophyta) в позднемеловых флорах Северной Пацифики // Палеоботаника. 2016. Т. 7. С. 66–79.
Herman A.B., Golovneva L.B., Shczepetov S.V., Grabovsky A.A. The Late Cretaceous Arman Flora of Magadan Oblast, Northeastern Russia // Stratigraphy and Geological Correlation. 2016. V. 24. № 7. P. 651–760. https://doi.org/10.1134/S0869593816070029
Kelley S.P., Spicer R.A., Herman A.B. New 40Ar/39Ar dates for Cretaceous Chauna Group tephra, North-Eastern Russia, and their implications for the geologic history and floral evolution of the North Pacific region // Cretaceous Res. 1999. V. 20. № 1. P. 97–106.
Shczepetov S.V., Golovneva L.B. Morphological diversity of leaves of Pseudoprotophyllum (Platanaceae) from the Turonian–Coniacian of the northern coast of the sea of Okhotsk // Paleontol. J. 2009. V. 43. № 10. P. 1330–1338.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция