

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 5, стр. 116-145
Кампан-маастрихтские радиолярии террейнов Корякского нагорья, Северо-Восток России
Т. Н. Палечек *
Геологический институт РАН
Москва, Россия
* E-mail: tpalechek@yandex.ru
Поступила в редакцию 05.09.2019
После доработки 25.10.2019
Принята к публикации 20.11.2019
Аннотация
Анализ выделенных позднемеловых радиоляриевых ассоциаций из различных структурно-формационных зон Олюторского и Алганского террейнов Корякского нагорья показал существенную разницу в их таксономическом составе, количественном содержании радиолярий, их морфологических особенностях. Олюторский террейн, занимающий южную часть Корякского нагорья, практически на 100% состоит из калифорнийских видов. Алганский террейн, расположенный в северо-западной части Корякского нагорья, на 50% и более состоит из видов, описанных из разрезов Русской плиты и Западной Сибири, и не более чем на 50% из калифорнийских видов. По этим данным отчетливо прослеживается связь Алганского палеобассейна с Палеоарктикой и Западно-Сибирским морем, вероятно, при одновременном влиянии тихоокеанских течений, с помощью которых могли проникнуть калифорнийские виды; а для Олюторского террейна устанавливается тесная связь с Палеопацификой в позднемеловое время. При этом бóльшая часть таксонов, наиболее характерных для Олюторской зоны, распространены также в верхнем мелу/палеоцене Новой Зеландии, включая как космополиты, так и, вероятно, биполярные виды, к которым, скорее всего, относятся Stichomitra livermorensis, Theocampe altamontensis, T. vanderhoofi, Bathropyramis sanjoaquinensis, Praestylosphaera pusilla, P. hastata. В Алганской зоне (северо-западная часть Корякского нагорья) было впервые выделено биостратиграфическое подразделение в ранге слоев с фауной – слой с Prunobrachium articulatum. В настоящее время уровень с Prunobrachium articulatum является субглобальным и трассируется от Западной Сибири, где он был впервые установлен, как на запад на Русскую плиту, так и на восток до Тихоокеанской окраины включительно. В Олюторской зоне (южная часть Корякского нагорья) намечено несколько уровней: на границе сантона и кампана – первое появление Praestylosphaera pusilla (F.O.) и Phaseliforma meganosensis (F.O.) и последнее присутствие Orbiculiforma quadrata (L.O.); в нижнем кампане – первое появление и обилие Dictyomitra densicostata; в верхнем кампане – первое появление Phaseliforma carinata, Stichomitra livermorensis, представителей рода Theocampe (T. altamontensis, T. yaoi, T. vanderhoofi); в верхах верхнего кампана/маастрихте – обилие Phaseliforma carinata, Stichomitra livermorensis, Archaeodictyomitra regina, первое появление Clathrocyclas hyronia и Lithomespilus mendosa. Проведена корреляция выделенных в Корякском нагорье биостратонов с биостратонами смежных территорий в Тихоокеанском регионе.
ВВЕДЕНИЕ
Складчатые комплексы Корякского нагорья образованы в результате аккреционных процессов, развивавшихся вдоль края Азиатского континента и Северо-Запада Пацифики. Возраст формирования структур определен как среднеальбский для Западно-Корякской складчатой системы и как позднесенонский для Корякской (Соколов, 1992, 2010; Парфенов, 1993; Геодинамика…, 2006). Меловое время характеризуется рядом крупных тектонических перестроек, связанных с отмиранием одних и заложением других активных окраин. Реконструкция позднемезозойских палеогеографических и палеотектонических обстановок осадконакопления окраины Азиатского континента возможна при изучении меловых вулканогенно-кремнисто-терригенных отложений Корякского нагорья, широко развитых на исследуемой территории. В решении вопроса о возрасте существенно кремнистых толщ первостепенное значение имеет изучение микрофауны радиолярий, которые часто являются породообразующими.
В пределах Корякского нагорья отложения коньяк-кампанского возраста представлены прибрежно-морскими и континентальными (на северо-западе), морскими (в центральной части) и глубоководными (на юго-востоке) фациями (Зинкевич, 1981; Соколов, 1992). Для изучения были выбраны две площади, включающие несколько ключевых участков: Олюторская зона (или Олюторский террейн, занимающий южную часть Корякского нагорья) и Алганская зона (или Алганский террейн, расположенный в северо-западной части Корякского нагорья) (рис. 1). Основной задачей настоящей работы являлось сравнение микрофауны радиолярий из кампанских отложений различных структурно-формационных зон Олюторского и Алганского террейнов, установление характера распределения таксонов, их морфологических особенностей, связей с различными палеобассейнами, возможных путей их миграции.
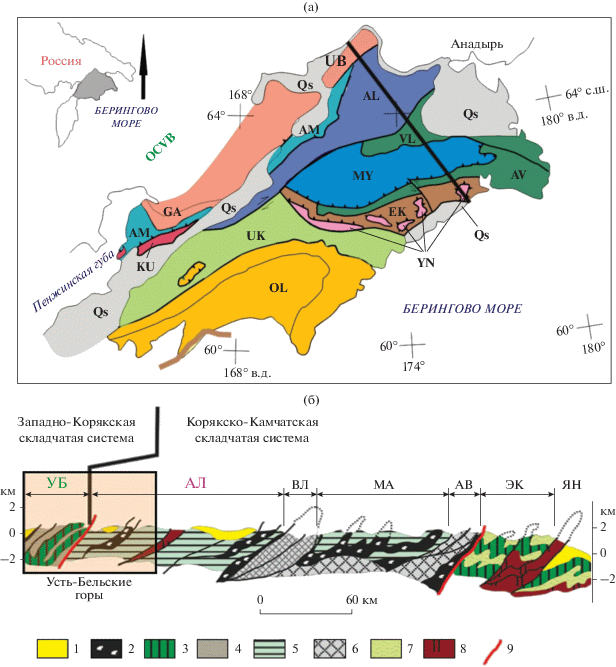
Рис. 1.
(а) Тектоническая схема Корякского нагорья (по Sokolov et al., 2003) и (б) геологический профиль через северную часть Корякского нагорья (по Соколов, 2003, с изменениями А.В. Моисеева (2015)). Рис. 1а: Qs – палеоген-четвертичные отложения; террейны: UB – Усть-Бельский, GA – Ганычаланский; КU – Куюльский, АМ – Айнынско-Майнский, АL – Алганский, VL – Великореченский; МY – Майницкий; АV – Алькатваамский; EK – Эконайский; YN – Янранайский; UK – Укэлаятский; OL – Олюторский; ОCVB – Охотско-Чукотский вулканогенный пояс. Рис. 1б: 1 – постаккреционные позднемеловые–палеогеновые образования; 2 – серпентинитовые меланжи; 3 – офиолиты; 4 – терригенные отложения и известняки (палеозой–нижний мезозой); 5–7 – позднеюрские–раннемеловые ассоциации: 5 – вулканогенно-кремнисто-терригенная; 6 – туфотерригенная (мелкая клетка – для Великореченского террейна); 7 – терригенная; 8 – базальт-яшмовая ассоциация; 9 – тектонические границы. Буквами обозначены террейны: УБ – Усть-Бельский; АЛ – Алганский; ВЛ – Великореченский; МА – Майницкий; АВ – Алькатваамский; ЭК – Эконайский; ЯН – Янранайский.
Материал был собран в ходе полевых сезонов 1993–1998, 2007, 2008, 2011, 2012, 2016 гг. на территории Корякского нагорья. Проведено комплексное исследование различных разрезов. На основании изучения литологических особенностей отложений и выяснения характера взаимоотношений различных типов пород были выделены тектоностратиграфические комплексы. Из кремнистых разностей большинства комплексов были получены объемные формы радиолярий. Для радиоляриевого анализа использовались методики, предложенные Э. Пессаньо (Pessagno, Newport, 1972), Б.Б. Назаровым и Д.И. Витухиным (Назаров, Витухин, 1981). Наиболее эффективным оказалось сочетание двух методов: изучения радиолярий в шлифах и отпрепарированных форм. Предварительно был проведен просмотр всех шлифов кремнисто-терригенных пород и подобрана методика травления для каждого образца (кислота/чередование кислот, концентрация, время травления). Детально исследованы различные срезы пруноидных форм в шлифах в проходящем свете под поляризационным микроскопом Meijitechno MT-9300 в ГИН РАН. Съемка радиолярий велась в сканирующем электронном микроскопе Vega Tescan MV-2300 в ГИН РАН Н.В. Горьковой и в ПИН РАН Е.А. Жегалло.
ОЛЮТОРСКИЙ ТЕРРЕЙН
Олюторская зона занимает восточную часть юга Корякского нагорья, расположенную севернее 58° с.ш. (рис. 1). Фронтальная часть Олюторской зоны протягивается вдоль ее северо-западной границы, характеризуется покровно-чешуйчатым строением и широким распространением меловых отложений (Богданов и др., 1982; Чехович, 1993; Соловьев, 2008). Структура фронтальной части определяется региональным Ватыно-Вывенским надвигом (Митрофанов, 1977) протяженностью более 500 км. Простирание структур конформно сместителю Ватыно-Вывенского надвига, по которому образования Олюторской зоны шарьированы на отложения Укэлаятского прогиба.
В районе Олюторского хребта структуры имеют субмеридиональное простирание и почти ортогонально соотносятся со структурами фронтальной части, здесь широко развиты осадочно-вулканогенные отложения мел-палеогенового возраста.
Фронтальную часть Олюторской зоны слагают преимущественно меловые вулканогенно-кремнисто-терригенные комплексы. К концу 1980-х годов сложилось представление о двух главных региональных стратиграфических единицах – ватынской серии и ачайваямской свите (Липман, 1959; Жамойда, 1972; Казинцова, 1979; Алексеев, 1979). Изучение вулканогенных пород позволило выделить два различных генетических типа меловых отложений: субокеанические и островодужные (Богданов и др., 1982; Геология…, 1987; Чехович, 1993). Отложения ватынской серии считались близкими к океаническим, а образования ачайваямской свиты – к островодужным (Алексеев, 1979).
К концу 1980-х годов возраст субокеанических образований ватынской серии определялся как альб-кампанский, а возраст островодужных отложений ачайваямской свиты как маастрихт-палеоценовый (Вишневская, 1985; Геология…, 1987). Позже появились данные о присутствии в ватынской серии, наряду с альб-кампанскими, и более молодых отложений кампан-маастрихтского возраста (Палечек, 1997; Соловьев и др., 1998, 2000), а также о том, что формирование ачайваямской свиты происходило, по-видимому, уже в кампанское время (Палечек, 1997). Это позволило предположить одновременное образование океанических (окраинно-морских) и островодужных отложений, по крайней мере, в кампане–маастрихте и формирование ими латерального ряда (Палечек, 1997; Соловьев и др., 2000). Ранее предполагались вертикальные соотношения между этими стратонами и считалось, что более молодые отложения ачайваямской свиты без видимого несогласия перекрывают более древние образования ватынской серии (Астраханцев и др., 1987; Казимиров и др., 1987; Шапиро, 1995). При анализе литературных данных о возрасте меловых отложений Олюторской зоны во внимание принимались датировки, полученные из кремнистых отложений, ассоциирующих с вулканическими породами с известными петрохимическими характеристиками, что позволяет предполагать ту или иную геодинамическую обстановку формирования отложений.
Фронтальная часть Олюторской зоны. Изучена по трем ключевым участкам: район бухты Анастасии (северный участок), верховья рек Ильпи и Матыскен (северо-западный участок) и бассейн р. Тапельваям (юго-западный участок) (рис. 2). В географическом отношении районы исследований расположены на западном побережье Алеутской впадины Берингова моря. Их тектоническая позиция определяется областью сочленения Олюторской и Укэлаятской зон. Детально рассмотрены вулканогенно-кремнистые и кремнисто-терригенные образования, из которых были получены радиолярии.
Рис. 2.
Схема расположения изученных участков Олюторского и Алганского террейнов в современных координатах. Олюторская зона (I): 1–3 – фронтальная часть: 1 – район бухты Анастасии; 2 – район верховий рек Ильпи и Матыскен, 3 – бассейн р. Тапельваям; 4–6 – восточная часть (комплексы Олюторского хребта): 4 – район мыса Витгенштейна, 5 – район лимана Мачевна, 6 – Олюторский полуостров; Алганская зона (II): 7 – район г. Кымъылннай; 8 – отроги г. Пик.
Северный участок. В районе бухты Анастасии были выделены два комплекса: вулканогенно-кремнистый и осадочно-вулканогенный (Палечек, 1997; Соловьев и др., 1998). Нижней структурной границей вулканогенно-кремнистого комплекса является Ватыно-Вывенский надвиг, по которому данные образования обдуцированы на флишоидные отложения Укэлаятской зоны. Гипсометрически выше вулканогенно-кремнистого комплекса залегают породы осадочно-вулканогенного комплекса (рис. 3). На территории изученного района контакт между этими комплексами тектонический, в одних местах это надвиги с пологим субгоризонтальным сместителем, в других разрывы взбросового типа с крутопадающим сместителем. Вулканогенно-кремнистый комплекс представлен пиллоу-базальтами, гиалокластитами и долеритами, а также кремнистыми породами и алевропелитами. Для разных тектонических пластин характерны различные парагенетические ассоциации пород: а) пиллоу-базальты, гиалокластиты и сургучные яшмы; б) черные алевропелиты, зеленые и серые силициты. Характерная черта первой ассоциации – сургучные яшмы с призматическими слоями иноцерамов, которые являются хорошим реперным горизонтом (рис. 4а–4в). Вторая ассоциация слагает чешуи, ограниченные поверхностями сместителей как сверху, так и снизу. В черных алевропелитах заключены линзы и прослои зеленых и серых кремнистых пород. Эти фрагменты наиболее деформированы.
Рис. 3.
Тектоностратиграфические колонки изученных комплексов Олюторского и Алганского террейнов. Состав комплексов показан схематично. Слева от колонок приведен возраст, установленный по радиоляриям. Справа от колонок цифрами показаны прослеженные биостратоны по радиоляриям: 1 – Orbiculiforma quadrata, 2 – Dictyomitra densicostata, 3 – Phaseliforma carinata, 4 – Clathrocyclas hyronia, 5 – Prunobrachium articulatum (см. табл. 2 ). Сокращения: ВК – вулканогенно-кремнистый комплекс; ОВ – осадочно-вулканогенный; ФО – флишоидно-олистостромовый; К – кремнистый.
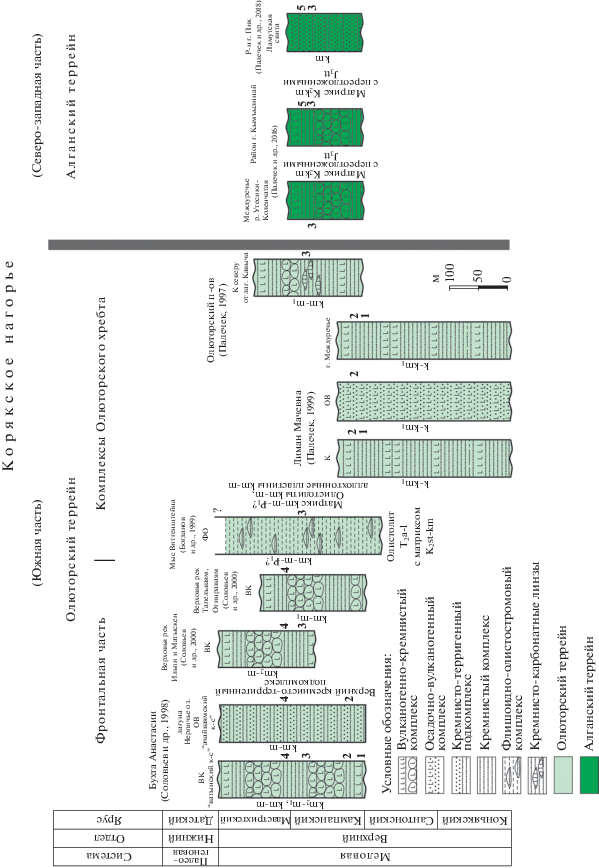
Рис. 4.
Характерные выходы кремнистых пород. а–в – Олюторская зона: выходы сургучных яшм (а, в) и яшм с призматическими слоями иноцерамов (а, б, в) на побережье бухты Анастасии; г–и – Алганская зона: г – отроги г. Кымъылннай, общий вид; д – выходы сургучных яшм в районе г. Кымъылннай; е – контакт яшм и базальтов в районе г. Кымъылннай; ж, з, и – серо-зеленые, черные и красные кремни в береговых обнажениях по р. Кымылнайвеем.

Пробы на микрофауну отбирали из кремнистых и терригенных пород различных тектоностратиграфических единиц. Из пород вулканогенно-кремнистого комплекса изучено около 100 образцов, однако только из 9 образцов, представленных сургучными яшмами, удалось экстрагировать и определить радиолярии, свидетельствующие о кампан-маастрихтском возрасте вмещающих отложений. В отдельных случаях выделены среднекампанские–раннемаастрихтские радиолярии из сургучных яшм, обожженных перекрывающими их пиллоу-базальтами. Сообщества радиолярий характеризуются невысоким таксономическим разнообразием и численностью (табл. 1). Количество видов радиолярий составляет 10–13 в наиболее представительных пробах, а максимальное число экземпляров оценивается первыми десятками. Наиболее типичными являются Praestylosphaera pusilla (Campbell et Clark), Amphipyndax stocki (Campbell et Clark), Stichomitra livermorensis (Campbell et Clark), Dictyomitra densicostata Pessagno, D. multicostata Zittel, Clathrocyclas tintinnaeformis Campbell et Clark, различные виды рода Phaseliforma (P. ex gr. carinata, P. cf. subcarinata). Кроме этого, встречены единичные экземпляры Cornutella californica Campbell et Clark, Stichomitra cf. shirshovica Vishnevskaya, Xitus cf. asymbatos (Foreman), Phaseliforma cf. meganosensis Pessagno (табл. I ).
Таблица 1.
Таксономический состав изученных радиоляриевых ассоциаций в разрезах Олюторского и Алганского террейнов Корякского нагорья
| Виды | Олюторский террейн | Алганский террейн | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| фронтальная часть | комплексы Олюторского хребта | |||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| ВК | ОВ | ВК | ВК | ФО, ол. | ал. пл. | К | ОВ | |||||
| Spumellaria | ||||||||||||
| Actinomma douglasi Pessagno | ||||||||||||
| Actinomma sp. | ||||||||||||
| Alievium sp. | ||||||||||||
| Archaeospongoprunum sp. | ||||||||||||
| Cromyodruppa concentrica Lipman | cf | |||||||||||
| Cromyosphaera vivenkensis Lipman | cf | |||||||||||
| Cromyosphaera sp. | ||||||||||||
| Crucella aster (Lipman) | ||||||||||||
| Crucella lata (Lipman) | ||||||||||||
| Crucella membranifera (Lipman) | ||||||||||||
| Haliomma minor Campbell et Clark | ||||||||||||
| Kreuzstella vierkantiga Empson-Morin | ||||||||||||
| Lithomespilus mendosa (Krasheninnikov) | ||||||||||||
| Orbiculiforma quadrata Pessagno | cf | |||||||||||
| Orbiculiforma rennilaeformis (Camp. et Cl.) | ||||||||||||
| Orbiculiforma sempiterna Pessagno | cf | |||||||||||
| Orbiculiforma vacaensis Pessagno | cf | |||||||||||
| Orbiculiforma sp. | ||||||||||||
| Patulibrachium sp. | ||||||||||||
| Phaseliforma carinata Pessagno | ||||||||||||
| Phaseliforma subcarinata Pessagno | ||||||||||||
| Phaseliforma meganosensis Pessagno | ||||||||||||
| Phaseliforma laxa Pessagno | ||||||||||||
| Phaseliforma sp. | ||||||||||||
| Porodiscus cretaceous Campbell et Clark | ||||||||||||
| Porodiscus vulgaris Lipman | ||||||||||||
| Praestylosphaera hastata (Campbell et Cl.) | ||||||||||||
| Praestylosphaera pusilla Campbell et Clark | ||||||||||||
| Prunobrachium articulatum (Lipman) | cf | cf | ||||||||||
| Prunobrachium crassum (Lipman) | cf | |||||||||||
| Prunobrachium incisum Kozlova | ||||||||||||
| Prunobrachium sp. | ||||||||||||
| Pseudobrachium ornatum (Lipman) | cf | |||||||||||
| Pseudobrachium mucronatum (Lipman) | ||||||||||||
| Pseudoaulophacus lenticulatus (White) | ||||||||||||
| Pseudoaulophacus sp. | ||||||||||||
| Rhopalastrum trigonale Lipman | ||||||||||||
| Rhopalastrum sp. | ||||||||||||
| Spongosaturnalis spiniferus Campbell et Cl. | ||||||||||||
| Saturnalis sp. | ||||||||||||
| Spongodiscus impressus Lipman | ||||||||||||
| Spongodiscus volgensis Lipman | ex gr | |||||||||||
| Spongodiscus sp. | ||||||||||||
| Spongurus concentricum (Lipman) | ||||||||||||
| Spongurus spongiosus (Lipman) | ||||||||||||
| Spongurus quadratus Campbell et Clark | ||||||||||||
| Spongurus sp. | ||||||||||||
| Stylotrocus nativus Lipman | ||||||||||||
| Stylotrocus paciferum Lipman | ||||||||||||
| Stylotrocus sp. | ||||||||||||
| Nassellaria | ||||||||||||
| Amphipyndax stocki Campbell et Clark | ||||||||||||
| Amphipyndax stocki var A Vishnevskaya | ||||||||||||
| Amphipyndax stocki var B Vishnevskaya | ||||||||||||
| Amphipyndax streckta Empson-Morin | ||||||||||||
| Archaeodictyomitra regina (Camp. et Cl.) | ||||||||||||
| Archaeodictyomitra simplex Pessagno | cf | |||||||||||
| Archaeodictyomitra sp. | ||||||||||||
| Bathropyramis sp. | ||||||||||||
| Clathrocyclas aff. diceros Foreman | ||||||||||||
| Clathrocyclas cf. gravis Vishnevskaya | ||||||||||||
| Clathrocyclas hyronia Foreman | ||||||||||||
| Clathrocyclas tintinnaeformis Camp. et Cl. | ex gr | |||||||||||
| Clathrocyclas sp. | ||||||||||||
| Cornutella californica Campbell et Clark | ||||||||||||
| Dictyomitra densicostata Pessagno | ||||||||||||
| Dictyomitra formosa Squinabol | ||||||||||||
| Dictyomitra multicostata Zittel | ||||||||||||
| Dictyomitra napaensis Pessagno | cf | |||||||||||
| Dictyomitra sp. | ||||||||||||
| Lithostrobus rostovzevi Lipman | ||||||||||||
| Stichomitra cf. campi Foreman | ||||||||||||
| Stichomitra aff. compsa Foreman | ||||||||||||
| Stichomitra livermorensis (Camp. et Clark) | ||||||||||||
| Stichomitra shirshovica Vishnevskaya | ||||||||||||
| Stichomitra sp. | ||||||||||||
| Theocampe altamontensis (Camp. et Clark) | ||||||||||||
| Theocampe cf. vanderhoofi (Camp. et Cl.) | ||||||||||||
| Theocampe yaoi Taketani | ||||||||||||
| Theocampe sp. | ||||||||||||
| Wildeus punctulatus (Pessagno) | ||||||||||||
| Xitus asymbatos (Foreman) | ||||||||||||
| Xitus sp. | ||||||||||||
| Xitus sp. B Iwata et Tajika | ||||||||||||
| Xitus sp. С Gorka | ||||||||||||
Примечание. 1, 2 – бухта Анастасии (Палечек, 1997; Соловьев и др., 1998); 3 – район верховий рек Ильпи и Матыскен (Палечек, 1997; Соловьев и др., 2000); 4 – бассейн р. Тапельваям, Огинраваям (Палечек, 1997); 5, 6 – мыс Витгенштейна (Палечек, 1997; Богданов и др., 1999); 7, 8 – Лиман Мачевна (Палечек, 1997, 1999); 9 – Олюторский п-в (Палечек, 1997); 10 – междуречье Утесики–Коленчатая (Палечек и др., 2016); 11 – район г. Кымъылннай (Палечек и др., 2016, с дополнениями после доизучения); 12 – отроги г. Пик (Палечек и др., 2018). ВК – вулканогенно-кремнистый комплекс; ОВ – осадочно-вулканогенный комплекс; ФО – флишоидно-олистостромовый комплекс, ол. – олистолиты, ал. п. – аллохтонные пластины; К – кремнистый комплекс
Таблица I . Кампан-маастрихтские радиолярии Олюторской зоны (южная часть Корякского нагорья). Длина масштабной линейки равна 100 мкм для всех изображений. Обр. 4, 29, А12, 136/ж происходят из бухты Анастасии; обр. Г26 – из района верховий рек Ильпи и Матыскен; обр. 27/88 – с Олюторского п-ова. 1 – Phaseliforma carinata Pessagno, обр. 27/88; 2, 3 – Lithomespilus mendosa (Krasheninnikov), обр. Г26; 4 – Haliomma sp., обр. 27/88; 5, 6 – Praestylosphaera pusilla (Campbell et Clark), обр. Г26; 7 – Cromyosphaera sp., обр. 27/88; 8, 9 – Actinomma sp., обр. 27/88; 10, 11 – Haliomma aff. minor Campbell et Clark, обр. 27/88; 12 – Spongosaturnalis spiniferus Campbell et Clark, обр. 27/88; 13 – Quinquecapsularia sp., обр. 27/88; 14 – Pseudoaulophacus cf. lenticulatus (White), обр. 27/88; 15 – Stylotrocus paciferum Lipman, обр. 27/88; 16 – Peritiviator ? sp., обр. 27/88; 17, 18 – Amphipyndax stocki (Campbell et Clark), обр. 29, Г26; 19, 20 – Stichomitra livermorensis (Campbell et Clark), обр. Г26; 21, 22 – Theocampe cf. yaoi Taketani, обр. 136/ж; 23 – Theocampe cf. altamontensis (Campbell et Clark), обр. 136/ж; 24, 25 – Clathrocyclas hyronia Foreman, обр. 27/88, Г26; 26 – Clathrocyclas tintinnaeformis Campbell et Clark, обр. 27/88; 27, 28 – Cornutella californica Campbell et Clark: 27 – обр. А12, 28 – обр. 4; 29 – Bathropyramis sp., обр. 4; 30, 31 – Archaeodictyomitra regina (Campbell et Clark), обр. Г26; 32 – Dictyomitra densicostata Pessagno, обр. 29; 33, 34 – Dictyomitra multicostata Zittel, обр. 29, Г26; 35 – Lithostrobus rostovzevi Lipman, обр. 12; 36 – Xitus asymbatos Foreman, обр. 27/88; 37 – Xitus sp. B Iwata et Tajika, обр. 27/88.
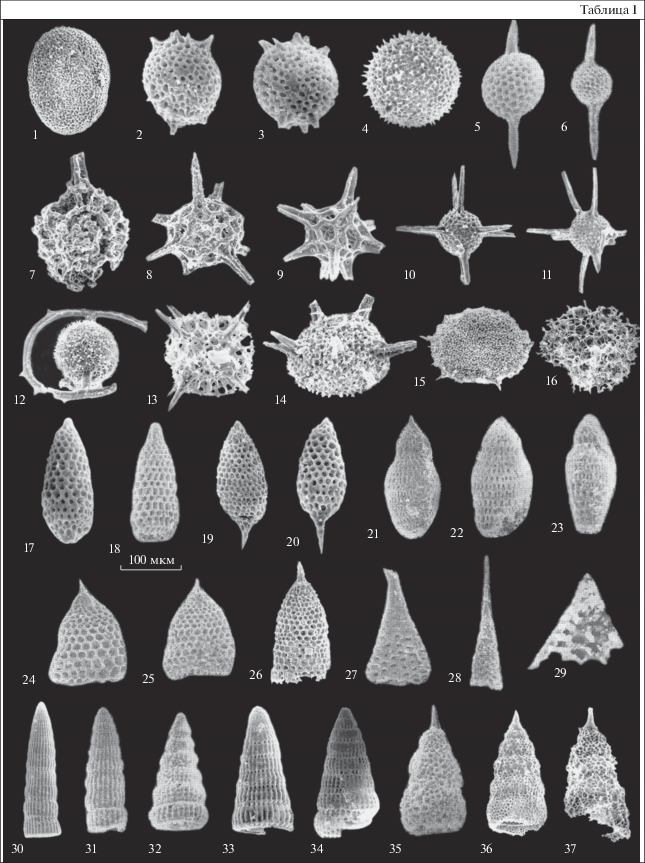
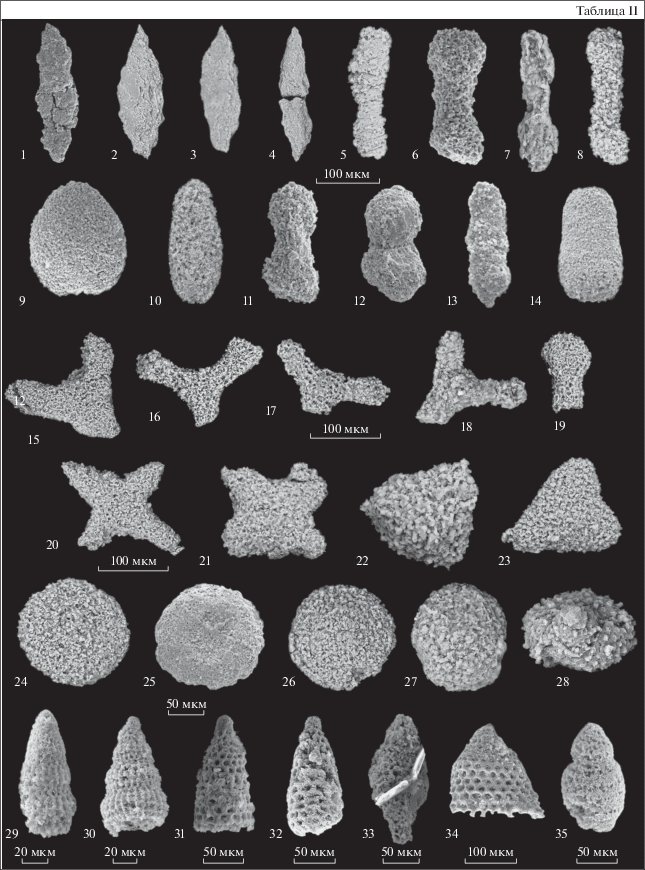
Таблица II. Кампанские радиолярии Алганской зоны (северо-западная часть Корякского нагорья). Длина масштабной линейки равна 100 мкм для фиг. 1–23 ; 50 мкм для фиг. 24–28, 20 мкм для фиг. 29, 30 ; 50 мкм для фиг. 31–33, 35 ; 100 мкм для фиг. 34 . Обр. 491.03 происходит из г. Кымъылннай (Усть-Бельские горы); обр. 176tp16, 184tp16, 187tp16 – из отрогов г. Пик (Алганские горы). 1, 2 – Prunobrachium cf. articulatum (Lipman): 1 – обр. 491.03; 2 – обр. 184tp16; 3, 4 – Prunobrachium sp.: 3 – обр. 176tp16, 4 – обр. 187tp16; 5 – Pseudobrachium cf. ornatum (Lipman), обр. 184 tp16; 6 – Pseudobrachium ornatum (Lipman), обр. 491.03; 7 – Prunobrachium cf. incizum Kozlova, обр. 491.03; 8 – Spongurus spongiosus (Lipman), обр. 491.03; 9 – Phaseliforma carinata Pessagno, обр. 491.03; 10 – Phaseliforma meganosensis Pessagno, обр. 176tp16; 11, 12 – Pseudobrachium mucronatum (Lipman), обр. 491.03; 13 – Prunobrachium sp., обр. 491.03; 14 – Spongurus quadratus Campbell et Clark, обр. 491.03; 15–19 – Rhopalastrum trigonale Lipman, обр. 491.03; 20 – Crucella aster (Lipman), обр. 491.03; 21 – Crucella membranifera (Lipman), обр. 491.03; 22 – Alievium sp., обр. 491.03; 23 – Patulibrachium sp., обр. 491.03; 24, 25 – Orbiculiforma vacaensis Pessagno, обр. 491.03; 26 – Porodiscus vulgaris Lipman, обр. 491.03; 27 – Porodiscus cf. cretaceous Campbell et Clark, обр. 491.03; 28 – Pseudoaulophacus lenticulatus (White), обр. 491.03; 29, 30 – Dictyomitra densicostata Pessagno, обр. 491.03; 31, 32 – Amphipyndax stocki (Campbell et Clark), обр. 491.03; 33 – Stichomitra cf. livermorensis (Campbell et Clark), обр. 491.03; 34 – Clathrocyclas cf. hyronia Foreman, обр. 491.03; 35 – Theocampe vanderhoofi (Campbell et Clark), обр. 491.03.
Осадочно-вулканогенный комплекс занимает большую часть площади полуострова между бухтой Анастасии и лагуной Нерпичье озеро. Фрагменты разрезов данного комплекса были изучены в ряде районов полуострова (Соловьев и др., 1998). В результате проведенных исследований осадочно-вулканогенный комплекс был подразделен на три подкомплекса с некоторой долей условности: 1) собственно вулканогенный (нижний); 2) вулканогенно-терригенный (средний или переходный); 3) кремнисто-терригенный (верхний).
Радиолярии были получены из кремнисто-терригенного (верхнего) подкомплекса осадочно-вулканогенного комплекса. Было изучено более 50 образцов, из трех образцов были выделены комплексы радиолярий, но только один из них (обр. 136/ж) характеризуется удовлетворительной сохранностью и представительностью (табл. 1, табл. I ). В этом комплексе также присутствуют кампан-маастрихтские формы: Praestylosphaera pusilla (Campbell et Clark), Cornutella californica Campbell et Clark, Clathrocyclas cf. tintinnaeformis Campbell et Clark, C. cf. hyronia Foreman, Theocampe cf. altamontensis (Campbell et Clark), Lithostrobus cf. rostovzevi Lipman, Phaseliforma sp., Orbiculiforma sp., Stichomitra sp., Dictyomitra sp., доминирует Theocampe yaoi Taketani.
Северо-западный участок. В районе верховий рек Ильпи и Матыскен (рис. 2) радиолярии были также изучены из вулканогенно-кремнистого комплекса (Палечек, 1997). На этой площади были выделены следующие структурные элементы (снизу вверх): автохтон: 1) флишоидные отложения Укэлаятского прогиба, 2) зона тектонического меланжа; 3) Ватыно-Вывенский надвиг; аллохтон: 4) вулканогенно-кремнистые отложения Олюторской зоны (Соловьев и др., 2000). Отложения вулканогенно-кремнистого комплекса слагают водоразделы в районе верховий рек Ильпи и Матыскен и обнажаются в скальных выходах около гривок хребтов и на вершинах гор. В долинах рек обнажаются флишоидные отложения Укэлаятской зоны. Образования вулканогенно-кремнистого комплекса, относимые к фронтальной части Олюторской зоны, шарьированы на флиш Укэлаятского прогиба по Ватыно-Вывенскому надвигу.
Вулканогенно-кремнистый комплекс в данном районе занимает наиболее высокое гипсометрическое положение, часто слагает изолированные тектонические пластины на вершинах гор (рис. 3) и представлен пиллоу-базальтами, лавобрекчиями и гиалокластитами базальтового состава и кремнистыми породами. В долинах рек Матыскен и Ильпи залегают различные горизонты вулканогенно-кремнистого комплекса. В северном и южном бортах долины р. Матыскен комплекс представлен в основном вулканогенными толщами, в бассейне р. Ильпи – кремнистыми разрезами. Кремнистые породы в изученном районе достаточно разнообразны: сургучные яшмы с прослоями, обогащенными битыми раковинами иноцерамов, массивные черные, зеленые и серые кремни. Образования вулканогенно-кремнистого комплекса сильно деформированы и дезинтегрированы, что не позволяет восстановить единый разрез по маломощным разрозненным фрагментам.
Из кремнистых пород вулканогенно-кремнистого комплекса было изучено 20 образцов, из 7 были экстрагированы радиолярии различной сохранности. Комплексы радиолярий характеризуются невысоким таксономическим разнообразием и численностью (табл. 1). В наиболее представительной пробе (Г26) определено 7 видов радиолярий, максимальное число экземпляров оценивается первыми десятками. В комплексе присутствуют: Phaseliforma carinata Pessagno, Lithomespilus mendosa (Krasheninnikov), Stichomitra livermorensis (Campbell et Clark), Archaeodictyomitra regina (Campbell et Clark), Dictyomitra densicostata Pessagno, D. multicostata Zittel, Clathrocyclas cf. hyronia Foreman (табл. I ). Выделенная ассоциация радиолярий позволяет говорить о том, что вулканогенно-кремнистый комплекс района верховий рек Ильпи и Матыскен включает толщи позднекампанского–маастрихтского возраста. По таксономическому составу она близка, хотя и несколько беднее, одновозрастной ассоциации района бухты Анастасии (обр. 29) (табл. 1). В вышеуказанной ассоциации радиолярий (Г26), полученной непосредственно из зоны надвига, следует отметить ярко выраженную деформированность некоторых скелетов радиолярий. Особенно отчетливо это проявляется у фазелиформид (Phaseliforma carinata Pessagno), несколько слабее у насселлярий (Archaeodictyomitra regina (Campbell et Clark)).
Юго-западный участок. Этот участок расположен в бассейне р. Тапельваям, которая протекает по северным отрогам Корякского хребта и является притоком реки Ветвей (рис. 2). Район работ располагался в пределах крупного тектонического останца Ватынского аллохтона (Митрофанов, 1977; Алексеев, 1979). Автохтоном являются флишоидные отложения Укэлаятской зоны позднемелового–палеогенового возраста (Митрофанов, Шелудченко, 1981). Параавтохтон сложен “олистостромовой” толщей маастрихтского возраста (Митрофанов, 1977). Тектонический останец представляет собой сложный пакет аллохтонных пластин, сложенных различными образованиями (Астраханцев и др., 1987; Kravchenko-Berezhnoy et al., 1993). Нижняя пластина состоит из вулканогенно-кремнистых пород ватынской серии сантон-кампанского (Астраханцев и др., 1987) и кампан-маастрихтского возраста (Kravchenko-Berezhnoy et al., 1993). Средняя пластина представлена туфово-кремнистыми толщами ачайваямской свиты позднемелового (Kravchenko-Berezhnoy et al., 1993) или маастрихт-палеоценового возраста (Астраханцев и др., 1987). Верхнюю пластину образуют гипербазиты и габброиды Сейнавского, Гальмоэнанского и Имланского массивов.
А.В. Соловьевым совместно с американскими геологами летом 1996 г. были изучены территории верховий реки Огинраваям (восточный склон массива Имлан) и западный склон массива Гальмоэнан. Т.Н. Палечек были обработаны образцы кремнистых пород (из коллекции А.В. Соловьева) и получены комплексы радиолярий (Палечек, 1997). В указанных местах на Укэлаятский флиш шарьированы аллохтонные пластины, сложенные вулканогенно-кремнистым комплексом и гипербазит-базитовыми породами (Соловьев, 1997, 2008).
Вулканогенно-кремнистый комплекс представлен пиллоу-базальтами, гиалокластитами и лавобрекчиями основного состава, кремнистыми породами и кремнистыми алевропелитами. Кремнистые породы достаточно разнообразны: зеленые и серые, массивные черные кремни, красные яшмы с прослоями, обогащенными битыми раковинами иноцерамов. В районе Гальмоэнана широко развиты белесые, серые грубослоистые кремни. Алевропелиты темно-красного цвета часто заполняют межподушечное пространство в базальтовых потоках. Фрагменты разрезов вулканогенно-кремнистого комплекса описаны в верховьях р. Огинраваям (Палечек, 1997) (рис. 3).
Из кремнистых пород вулканогенно-кремнистого комплекса было изучено 10 образцов, из трех были экстрагированы радиолярии различной сохранности. Были определены: Stichomitra cf. livermorensis (Campbell et Clark), Cornutella cf. californica Campbell et Clark, Amphipyndax stocki (Campbell et Clark), Pseudoaulophacus cf. lenticulatus (White), Phaseliforma ? sp., Orbiculiforma sp., Prunobrachium sp., Archaeospongoprunum sp., Theocampe ? sp., Dictyomitra sp., свидетельствующие о кампан-раннемаастрихтском возрасте вмещающих отложений (табл. 1). Комплексы радиолярий характеризуются невысоким таксономическим разнообразием и плохой сохранностью. Их можно сопоставить с поздневатынским и раннеинетываямским комплексами.
Близкиe по таксономическому составу радиоляриевыe комплексы позднекампан-маастрихтского возраста, которые включают Phaseliforma laxa Pessagno, Xitus asymbatos Foreman, Stichomitra ex gr. livermorensis (Campbell et Clark), Dictyomitra rhadina Foreman, а также комплексы сантон-кампанского возраста, где определены Pseudoaulophacus floresensis Pessagno, Ampipyndax stocki var. A Vishnevskaya, A. stocki var. B Vishnevskaya, Dictyomitra densicostata Pessagno, D. multicostata Zittel, были ранее установлены В.С. Вишневской из кремнистых пород района горы Сейнав (Kravchenko-Berezhnoy et al., 1993).
Олюторский хребет. Район исследований расположен в восточной части Олюторской зоны, где структуры имеют субмеридиональное простирание и почти ортогонально соотносятся со структурами фронтальной части. Здесь широко развиты осадочно-вулканогенные отложения мел-палеогенового возраста.
В качестве ключевых участков были выбраны: район мыса Витгенштейна, где был выделен флишоидно-олистостромовый комплекс, и район лимана Мачевна, где были выделены два комплекса: кремнистый и осадочно-вулканогенный, между которыми наблюдается постепенный переход (Палечек, 1999) (рис. 2, 3).
В районе мыса Витгенштейна флишоидно-олистостромовый комплекс занимает наиболее низкое структурное положение в системе покровов Олюторского хребта, обнажаясь вдоль побережья мыса Витгенштейна и между бухтами Глубокая и Амаян по долинам крупных ручьев. Этот комплекс перекрыт изолированными аллохтонными пластинами кремнистых, вулканогенных и вулканогенно-кремнистых пород и разделен на два подкомплекса. Первый подкомплекс, названный типично олистостромовым, представлен терригенной матрицей, в которую включены олистолиты и олистоплаки базальтов, кремнистых и терригенных пород. Второй подкомплекс сложен исключительно терригенными породами флишоидного облика. Соотношение подкомплексов окончательно не выяснено; предполагается, что типично олистостромовые толщи надвинуты на флишоидные. Падение поверхности надвига, вероятно, юго-западное на западном побережье мыса и северо-западное на восточном побережье (Богданов и др., 1999).
Возраст олистолитов кремнистых пород из флишоидно-олистостромового комплекса определен как кампан-маастрихтский на основе радиолярий: Phaseliforma laxa Pessagno, Clathrocyclas cf. gravis Vishnevskaya, Amphipyndax stocki (Campbell et Clark), Archaeodictyomitra regina (Campbell et Clark), Dictyomitra densicostata Pessagno, Xitus asymbatos (Foreman), Stichomitra aff. compsa Foreman, Theocampe sp. Кроме того, на западном побережье бухты Амаян был обнаружен олистолит сургучных яшм с кластогенно-турбидитной структурой, которая видна только на микроскопическом уровне. Важно отметить, что матрикс этих яшм содержит холодноводный комплекс радиолярий сантон-кампанского возраста: Amphipyndax ex gr. stocki (Campbell et Clark), Dictyomitra cf. lamellicostata Foreman, D. cf. densicostata Pessagno, D. ex gr. multicostata Zittel. Возраст включений соответствует среднему триасу (анизий–ладиний) и определен по комплексу радиолярий: Pseudostylosphaera cf. tenuis Nakaseko et Nishimura, P. aff. goestlingensis Kozur et Mostler, Triassocampe scalaris Dumitrica, Kozur et Mostler. Эти яшмы, несомненно, являются чужеродными по отношению к образованиям Олюторской зоны, так как среди последних неизвестны породы древнее второй половины раннего мела. Ранее, по данным Н.Ю. Брагина (Астраханцев и др., 1987), позднетриасовые (поздний норий–рэт) радиолярии Canoptum triassicum Yao, Kojurastrum quadriradiatus (Kozur et Mostler), Dreyericyrtium sp., Haeckelicyrtium sp. были встречены в маастрихтской матрице олистостромовой толщи вблизи Ватыно-Вывенского надвига в среднем течении р. Вывенка.
Глинисто-алевритовые породы матрикса олистостромы весьма бедны микрофауной. Комплекс наннопланктона (Micula staurophora, Watznaueria barnesae, Prediscosphaera cretacea, Rucinolithus sp., Ceratolithus cf. camptneri, Glaucolithus compactus, Cribrosphaera ehrenbergi, Zeugzhabdotus pseudoanthophorus) с небольшим таксономическим разнообразием, по заключению Е.А. Щербининой (Щербинина, Коваленко, 1996), указывает на кампан-маастрихтский возраст этих толщ. Вместе с тем единичные находки радиолярий кайнозойского облика из матрикса олистостромы, вероятно, свидетельствуют о возможном присутствии более молодых частей флишоидно-олистостромового комплекса (Палечек, 1997; Богданов и др., 1999).
Кремнистые породы аллохтонных пластин в районе мыса Витгенштейна, залегающие на флишоидно-олистостромовом комплексе, по мнению В.Д. Чеховича (1993), имеют островодужную природу и содержат до 10–15% радиолярий и спикул губок. Изредка встречаются бурые до кирпично-красных яшмы, содержащие битые раковины иноцерамов. Возраст пластин по комплексам радиолярий (Spongodiscus ex gr. volgensis Lipman, Phaseliforma ex gr. carinata Pessagno, Stichomitra cf. livermorensis (Campbell et Clark), S. cf. campi Foreman, Clathrocyclas ex gr. tintinnaeformis Campbell et Clark, C. aff. diceros Foreman, Xitus ex gr. asymbatos (Foreman)) соответствует кампану–маастрихту, т.е. близок к возрасту кремнистых пород из олистолитов и возрасту части разреза терригенной матрицы флишоидно-олистостромового комплекса.
Низкое таксономическое разнообразие радиоляриевых комплексов из олистолитов и пластин, а также морфологические особенности самих радиолярий (присутствие массивных губчатых дискоидей, отсутствие иглистых форм, редкие формы, снабженные короткими иглами или армированные шипами) указывают на холодноводность комплекса.
В районе лимана Мачевна были выделены два комплекса: кремнистый и осадочно-вулканогенный, между которыми описан постепенный переход (рис. 2, 3). В качестве переходного был выделен кремнисто-вулканогенный подкомплекс, который включен в осадочно-вулканогенный комплекс по преобладанию вулканогенного материала. Осадочно-вулканогенный комплекс подразделен на три подкомплекса: кремнисто-вулканогенный (переходный), собственно вулканогенный и кремнисто-терригенный (Палечек, 1999).
Отложения кремнистого комплекса на полуострове занимают наиболее низкое гипсометрическое положение и распространены только в его южной части. В районе лимана Мачевна кремнистый комплекс представлен двумя типами пород: а) cургучными яшмами, яшмами с призматическими слоями иноцерамов; б) зеленовато-серыми и черными силицитами. Базальты не встречены.
Из пород кремнистого комплекса радиолярии удалось экстрагировать только из сургучных яшм. Комплекс радиолярий, как и большинство вышеописанных, не отличается высоким таксономическим разнообразием и численностью (табл. 1). Здесь присутствуют Orbiculiforma cf. vacaensis Pessagno, Orbiculiforma (?) cf. sempiterna Pessagno, Cromyosphaera cf. vivenkensis Lipman, Amphipyndax stocki (Campbell et Clark), Amphipyndax stocki var. A Vishnevskaya, Dictyomitra cf. densicostata Pessagno, Dictyomitra cf. formosa Squinabol, Archaeodictyomitra sp. (обр. 102), которые указывают на то, что в состав кремнистого комплекса района лимана Мачевна входят отложения коньяк-раннекампанского возраста. По-видимому, это более низкие, чем во фронтальной части Олюторской зоны (район бухты Анастасии, верховья рек Ильпи и Матыскен, Тапельваям), горизонты, относящиеся к ватынской серии. Выделенные комплексы радиолярий могут быть сопоставлены с ранне- и средневатынскими комплексами Беринговоморского региона (Вишневская, 1985).
Гипсометрически выше кремнистого комплекса залегает осадочно-вулканогенный комплекс. В изученном районе контакт между этими комплексами стратиграфический. Основная площадь полуострова между лиманом Мачевна и лиманом Таман сложена образованиями осадочно-вулканогенного комплекса. В результате проведенных исследований осадочно-вулканогенный комплекс с некоторой степенью условности удалось подразделить на три подкомплекса: 1) кремнисто-вулканогенный (переходный), 2) собственно вулканогенный и 3) кремнисто-терригенный.
Из сургучных яшм кремнисто-вулканогенного подкомплекса были определены радиолярии коньяк-раннекампанского возраста (обр. 91): Archaeodictyomitra cf. simplex Pessagno, Dictyomitra cf. densicostata Pessagno, Dictyomitra multicostata Zittel, Dictyomitra cf. napaensis Pessagno, Amphipyndax cf. stocki (Campbell et Clark), Amphipyndax stocki var. B Vishnevskaya. По таксономическому составу этот радиоляриевый комплекс несколько отличается от одновозрастной ассоциации радиолярий из пород кремнистого комплекса. В первом присутствуют амфипиндациды и многосегментные циртиды, во втором многочисленные орбикулиформиды наряду с циртидами. Выделенные сообщества радиолярий могут быть сопоставлены с ранневатымским и средневатынским комплексами Беринговоморского региона (Вишневская, 1985).
Из пород кремнисто-терригенного подкомплекса было проанализировано около 50 образцов. К сожалению, из-за плохой сохранности не было обнаружено представительных комплексов радиолярий. В обр. 1 были встречены Orbiculiforma sp., Cromyosphaera ? sp., Spongurus ? sp., не позволяющие установить возраст вмещающих отложений.
Дополнительно на Олюторском полуострове были изучены радиолярии из кремнисто-карбонатных пород (серия образцов 27/88 из коллекции В.Д. Чеховича, отобранных непосредственно к северу от лагуны Кавача Олюторского полуострова из маломощных кремнисто-карбонатных линз среди кремней ватынского облика; рис. 2, 3; Палечек, 1997). Следует отметить, что описываемые карбонатные породы не характерны для ватынской и ачайваямской свит. Их уникальность заключается в том, что они содержат микрофауну и микрофлору. Радиолярийсодержащие породы представлены светло-серыми микритовыми известняками со стяжениями темно-серых до черных кремней. Вместе с радиоляриями были отмыты многочисленные диатомеи, относящиеся к классу Centrophyceae. На единицу объема отмытой породы приходится 15–30% радиолярий, до 5% спикул губок и 5–10% диатомей.
Из радиолярий определены: Spongosaturnalis spiniferus Campbell et Clark, который известен из сенонских отложений Калифорнии (Campbell, Clark, 1944) и кампана формации Тоясито (Toyosato Formation) Японии (Iwata, Tajika, 1986); Actinomma douglasi Pessagno, впервые описанная из верхней части формации Форбес (Forbes Formation) верхнего кампана Северной Калифорнии (Pessagno, 1976); Haliomma minor Campbell et Clark, характерная для сенона Калифорнии (Campbell, Clark, 1944); Cromyosphaera vivenkensis Lipman, распространенная в верхнемеловых отложениях Беринговоморского региона, р. Ватына (Геология…, 1987); Kreuzstella vierkantiga Empson-Morin, известная из комплекса кампанских радиолярий из скв. 313 DSDP, Центрально-Тихоокеанские горы (Empson-Morin, 1981); Phaseliforma carinata Pessagno, являющаяся индекс-видом подзоны низов верхнего кампана Северной Калифорнии (Pessagno, 1976); Stylotrochus paciferum Lipman и S. nativus Lipman, описанные из верхнего эоцена Западно-Сибирской низменности (Липман, 1962; Козлова, Горбовец, 1966); Spongodiscus impressus Lipman и S. volgensis Lipman, также характерные виды позднего мела Западно-Сибирской низменности (Липман, 1962; Козлова, Горбовец, 1966); Clathrocyclas ex gr. tintinnaeformis Campbell et Clark, описанный из сенонских отложений Калифорнии (Campbell, Clark, 1944); Xitus asymbatos (Foreman), впервые описанный из маастрихтских отложений Калифорнии (Foreman, 1968), позднее встреченный в верхнемеловых отложениях многих районов; Xitus sp. B Iwata et Tajika, ранее описанный из кампана формации Онари (Onari Formation) (Iwata, Tajika, 1986); Xitus sp. C Gorka, впервые встреченный в кампане Польши (Gorka, 1989); Stichomitra livermorensis (Campbell et Clark), известная из сенонских отложений Калифорнии (Campbell, Clark, 1944) (табл. I ).
Выделенный комплекс радиолярий был датирован кампаном–ранним маастрихтом. Он имеет общие черты с поздневатынской радиоляриевой ассоциацией и описанным в бассейне р. Мачевна раннеинетываямским комплексом (Геология…, 1987).
Южнее, в районе горы Междуречье (рис. 2), из кремнистых пород (коллекция А.В. Ландера) был выделен комплекс коньяк-раннекампанских радиолярий (табл. 1). Комплекс характеризуется невысоким таксономическим разнообразием и включает Orbiculiforma cf. vacaensis Pessagno, O. cf. quadrata Pessagno, Pseudoaulophacus sp., Archaeodictyomitra cf. squinaboli Pessagno, Dictyomitra cf. densicostata Pessagno, D. cf. multicostata Zittel, Ampipyndax cf. stocki (Campbell et Clark). Изученную ассоциацию радиолярий можно сопоставить с ранневатымским и средневатынским комплексами Беринговоморского региона (Геология…, 1987).
Таким образом, проведен радиоляриевый анализ трех структурно-формационных комплексов Олюторского террейна: вулканогенно-кремнистого, осадочно-вулканогенного и флишоидно-олистостромового.
Кампан-маастрихтские радиоляриевые ассоциации из вулканогенно-кремнистых отложений различных участков фронтальной части Олюторской зоны в целом схожи по своему таксономическому составу, хорошо коррелируются между собой (табл. 1) и сопоставляются с поздневатынским и раннеинетываямским комплексами Беринговоморского региона (Вишневская, 1985).
Вулканогенно-кремнистые отложения на всех изученных участках фронтальной части Олюторской зоны близки по составу, структурному положению и содержат, наряду с более древними, горизонты кампан-маастрихтского возраста, что позволяет коррелировать их между собой на этом временнóм интервале. По вещественному составу они сопоставляются с океаническими и/или окраинно-морскими образованиями ватынской серии (Богданов и др., 1982; Геология…, 1987; Чехович, 1993), формирование которой, на основании вышесказанного, могло продолжаться до конца мела. Осадочно-вулканогенные отложения, изученные лишь на северном участке (район бухты Анастасии), также содержат горизонты кампан-маастрихтского возраста (Палечек, 1997; Соловьев и др., 1998), а по вещественному составу они сопоставляются с островодужными образованиями ачайваямской свиты (Чехович, 1993), формирование которой, таким образом, могло начаться уже в кампане.
На основании полученных датировок и литературных данных составлена схема возрастных соотношений меловых океанических (окраинно-морских) и островодужных отложений Олюторской зоны (Палечек, 1997; Соловьев и др., 2000). Очевидно, что островодужные отложения кампан-маастрихсткого возраста могут перекрывать, возможно и без видимого несогласия, океанические и/или окраинно-морские докампанские комплексы, что отмечалось в ряде публикаций. Такая ситуация, по-видимому, возможна при зарождении островной дуги на океанической коре (Астраханцев и др., 1987; Казимиров и др., 1987). С другой стороны, одновременное формирование островодужных и океанических (окраинноморских) отложений в кампане–маастрихте позволяет считать их первичные взаимоотношения латеральными, а наблюдаемые в современной структуре тектонические контакты между ними могут быть результатом более позднего тектонического скучивания (Соловьев и др., 2000).
Проведенные в районе мыса Витгенштейна исследования позволили выделить во флишоидно-олистостромовом комплексе два подкомплекса: типично-олистостромовый и безолистолитовый. Изучение комплексов радиолярий и наннопланктона показало, что формирование флишоидного матрикса происходило в кампане–маастрихте и, возможно, продолжалось в более позднее время, кремнистые породы олистолитов и олистоплак, а также аллохтонных пластин образовались в кампане–маастрихте. Радиоляриевые комплексы из флишоидно-олистостромового комплекса сопоставимы с одновозрастными радиоляриевыми комплексами фронтальной части Олюторской зоны.
Находка тепловодного триасового (анизий–ладиний) комплекса радиолярий, самого древнего из известных ранее в Олюторской зоне, заключенного в ватынский матрикс, содержащий холодноводный сантон-кампанский комплекс радиолярий, является еще одним подтверждением того, что триасовые образования, сформировавшиеся в низких широтах, были транспортированы в более высокие широты до формирования ватынской серии. Образования самой ватынской серии имеют гораздо более сложное строение, чем предполагалось ранее, и требуют новой интерпретации.
Наблюдается сходство изученных ассоциаций радиолярий, выделенных из комплексов тектоностратиграфических разрезов, с ассоциациями радиолярий позднего мела Калифорнии (Campbell, Clark, 1944; Foreman, 1968; Pessagno, 1976) и Японии (Taketani, 1982), а также с сообществами, описанными из скв. 275 DSDP (Pessagno, 1975) и Новой Зеландии (Hollis, 1997).
АЛГАНСКИЙ ТЕРРЕЙН
В пределах северо-западной части Корякского нагорья, так же как и в его южной части, широко распространены вулканогенно-кремнисто-терригенные образования. Они входят в состав покровно-складчатых структур, сформированных в результате аккреционных процессов, проявленных вдоль Азиатского континента. В северной части нагорья подобные образования объединены в алганскую (ранее пекульнейвеемскую) свиту и чирынайскую серию (Кайгородцев, 1961; Кальянов, Беляцкая, 1961; Зинкевич, 1981; Терехова, 1987; Шмакин, 1988). Область их распространения соответствует Алганской и Майницкой структурно-формационным зонам (Зинкевич, 1981; Чехов, 1982; Ставский и др., 1989; Соколов, 1992), позже выделенным в одноименные террейны (Парфенов и др., 1993; Соколов, Бялобжеский, 1996). Большинством исследователей возраст алганской свиты и чирынайской серии рассматривался как позднеюрско-раннемеловой. В ходе разномасштабного картирования в Анадырско-Корякском регионе среди пород, отнесенных к указанным свите и серии, была обнаружена фауна триаса, ранней и средней юры (Невретдинов, Лебедев, 1987; Терехова, 1987; Грецкий, 1989; Вяткин, 1990), а также кампана (Гульпа, 2014; Палечек и др., 2016). Детальное изучение отдельных участков позволило выделить несколько различных структурно-вещественных комплексов и продемонстрировать участие мезозойских комплексов в многочисленных покровно-надвиговых дислокациях (Очерки…, 1982; Соколов, 1992; Филатова, Вишневская, 1992; Вишневская, Филатова, 1996; Палечек и др., 2013, 2016; Моисеев, 2015).
Был изучен ряд ключевых участков в Усть-Бельских и Алганских горах, расположенных в северо-западной части Корякского нагорья (Палечек и др., 2013, 2016, 2018; Моисеев, 2015) (рис. 1, 2). В географическом отношении Усть-Бельские горы расположены на правобережье крупной излучины среднего течения р. Анадырь. Тектоническая позиция района исследования определяется областью сочленения разновозрастных складчатых областей (Соколов, 1992, 2010; Парфенов и др., 1993; Соколов, Бялобжеский, 1996; Геодинамика…, 2006). В западной части распространены породы Усть-Бельского террейна Западно-Корякской складчатой области, которые включены в состав аллохтона (Соколов, Бялобжеский, 1996). Восточнее бассейна р. Левая Маврина широко развиты вулканогенно-кремнисто-терригенные породы. Допозднемеловые вулканогенно-кремнисто-терригенные породы Алганского террейна включены в алганскую (ранее пекульнейвеемскую) свиту (Кайгородцев, 1961; Кальянов, Беляцкая, 1961; Захаров, 1974; Зинкевич, 1981; Терехова, 1987; Шмакин, 1988; Легенда…, 1999).
В пределах Алганского террейна к неоавтохтону относятся перекатнинская и ламутская свиты, представленные флишоидными отложениями позднеальб-туронского и сенонского возрастов соответственно (Зинкевич, 1981; Соколов, Бялобжеский, 1996). Отложения датированы по единичным находкам фауны, и на большей части территории их возраст неясен, что не исключает возможного присутствия в их составе разновозрастных отложений. Особенно это актуально для вулканогенно-кремнистых образований, которые описаны в единичных разрезах (Зинкевич, 1981; Очерки…, 1982; Вяткин, 1990). В структурном отношении образования этих свит рассматривают как слабодеформированный чехол (Соколов, Бялобжеский, 1996).
Образования вулканогенно-кремнистого комплекса изучены в правом борту р. Анадырь, на р. Перевальная, в нижнем течении р. Правый Коначан, в междуречье Утесики–Коленчатая, районе г. Кымъылннай (Моисеев, 2015; Палечек и др., 2016). Вулканиты представлены темно-коричневыми и зеленоватыми трещиноватыми базальтами с реликтами подушечной отдельности, с редкими прослоями гиалокластитов и кремнистых туфоаргиллитов. В подчиненном количестве встречаются жильные тела долеритов. Базальты содержат изометричные будины вулканитов кислого состава (дациты, риолиты) до 6 м в длину. Кремнистые породы представлены метаморфизованными бордовыми яшмами и радиоляритами (рис. 4г–4и). Внутри комплекса взаимоотношения вулканитов с кремнистыми породами различны. Чаще всего породы данного комплекса оказываются сильно тектонизированы, кремнистые породы образуют вытянутые будины и фрагменты “слоев” в базальтовом матриксе. В других случаях наблюдаются четкие стратиграфические контакты тонкослоистых кремней с базальтами. Возможное основание вскрыто в виде габбро-долеритовых тел в районе г. Кымъылннай (Палечек и др., 2016).
Кампанские радиолярии были изучены из вулканогенно-кремнистого комплекса в междуречье Утесики–Коленчатая, в районе г. Кымъылннай (Усть-Бельские горы) (Гульпа, 2014; Палечек и др., 2016), а также из ламутской свиты в отрогах г. Пик (Алганские горы) (Палечек и др., 2018).
В районе г. Кымъылннай (Усть-Бельские горы) из прослоев кремней (обр. 491.03) среди базальтов, в непосредственной близости от коренных выходов габбро-долеритов была выделена кампанская ассоциация радиолярий: Prunobrachium cf. articulatum (Lipman), P. incizum Kozlova, Spongurus spongiosus (Lipman), S. quadratus Campbell et Clark, S. concentricum (Lipman), Pseudobrachium ornatum (Lipman), P. mucronatum (Lipman), Cromyosphaera vivenkensis Lipman, Cromyodruppa concentrica Lipman, Phaseliforma cf. carinata Pessagno, P. meganosensis Pessagno, Porodiscus vulgaris Lipman, P. cretaceous Campbell et Clark, Pseudoaulophacus lenticulatus (White), Orbiculiforma vacaensis Pessagno, Crucella aster (Lipman), Rhopalastrum trigonale Lipman, Crucella membranifera (Lipman), Patulibrachium sp., Alievium sp. (табл. 1, табл. II). Дополнительное изучение радиолярий из прослоев кремней позволило уточнить и дополнить таксономический состав изученной ассоциации. Здесь определены: Amphipyndax stocki Campbell et Clark, Stichomitra cf. livermorensis (Camp. et Clark), Dictyomitra densicostata Pessagno, Clathrocyclas cf. hyronia Foreman, Theocampe vanderhoofi (Campbell et Clark). Следует отметить, что такие таксоны, как Clathrocyclas cf. hyronia Foreman, Theocampe vanderhoofi (Campbell et Clark), Stichomitra cf. livermorensis (Camp. et Clark), характерны для позднего кампана–маастрихта. Кроме того, в этом же образце были встречены радиолярии, характерные для титона: Zhamoidellum ventricosum Dumitrica, Z. ovum Dumitrica, Williriedellum carpaticum Dumitrica, Parvicingula ex gr. boesii (Parona), Loopus sp., Triversus sp. Совместное нахождение кампанских и титонских радиолярий было подтверждено еще в нескольких образцах (обр. 492.02, 11-10, 11-14), отобранных в непосредственной близости.
В отрогах г. Пик (Алганские горы) из кремнисто-терригенных отложений ламутской свиты были выделены радиолярии кампанского возраста. Изученная ассоциация радиолярий достаточно разнообразна (табл. 1, табл. II); спумеллярии в ней составляют 75%, насселлярии 25%. Здесь насчитывается 12 родов, в каждом из которых присутствует от одного до трех видов, при наибольшем разнообразии пруноидных форм, таких как Prunobrachium articulatum, P. cf. incisum, P. cf. crassum, Pseudobrachium cf. mucronatum, P. cf. ornatum. Из насселлярий присутствуют Amphipyndax sp., Stichomitra cf. livermorensis Campbell et Clark, S. cf. campi Foreman, Theocampe cf. vanderhoofi (Campbell et Clark). Скелеты радиолярий частично или нацело замещены органическим веществом, что затрудняло проведение химического препарирования микрофоссилий. Стоит также отметить маленький размер раковин радиолярий (менее 100 мкм). Сходная картина отмечена при изучении радиолярий района Чаунской губы на Чукотке (Палечек, 2018).
Из определенных в ламутской свите 17 видов радиолярий, относящихся к 12 родам, 9 видов (53% в изученной ассоциации) впервые описаны из кампанских отложений Западной Сибири и Тургайского прогиба (Козлова, Горбовец, 1966; Липман, 1962), остальные 8 видов (47%) – из позднесенонских отложений Калифорнии (Campbell, Clark, 1944; Pessagno, 1976) (рис. 5, 6). P. articulatum встречен во всех 14 образцах, отобранных из ламутской свиты. Похожая картина наблюдается и в кампанской ассоциации, изученной в районе г. Кымъылннай: здесь насчитывается 18 родов, из них 14 родов Spumellaria (78%) и всего 4 рода Nassellaria (22%), в каждом из которых присутствует от одного до трех видов (рис. 6). Так же, как и в случае с ламутской свитой, больше половины присутствующих здесь видов (52% изученной ассоциации) впервые описаны из кампанских отложений Западной Сибири и Тургайского прогиба (Липман, 1962; Козлова, Горбовец, 1966), остальные описаны из позднесенонских отложений Калифорнии (Campbell, Clark, 1944; Pessagno, 1976) и Камчатки (Липман, 1967) (рис. 6).
Рис. 5.
Видовое разнообразие радиолярий в Олюторской (а) и Алганской (б) зонах. По вертикальной оси – количество видов. 1 – Spumellaria, 2 – Nassellaria.
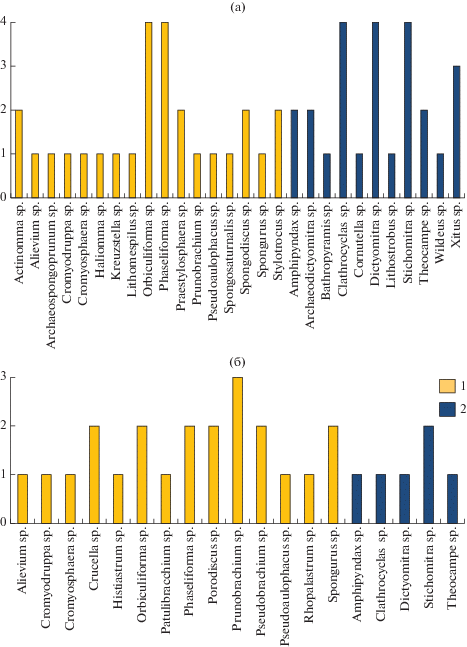
Рис. 6.
Диаграммы распределения таксонов радиолярий в Олюторском и Алганском террейнах. Под “сибирскими” понимаются виды, описанные Р.Х. Липман (1952, 1962) из кампанских отложений Русской платформы и Западно-Сибирской низменности.
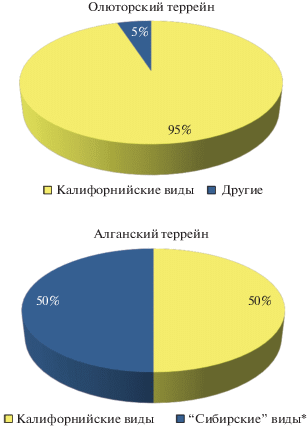
В настоящее время выделено биостратиграфическое подразделение в ранге слоев с Prunobrachium articulatum в разрезах Корякского нагорья, п‑ова Камчатка и о-ва Шикотан в Тихоокеанском регионе. Эти слои прослежены в Западной Сибири, на Урале и Русской платформе и являются прекрасным репером для кампанского интервала в Бореальной области (Палечек, 2018). В большинстве образцов, отобранных из ламутской свиты, совместно с P. articulatum встречена Phaseliforma carinata Pessagno, являющаяся индекс-видом подзоны низов верхнего кампана Северной Калифорнии (Pessagno, 1976). Представители семейства Prunobrachidae распространены симметрично и биполярно относительно экватора и заселяли моря Бореального и Нотального климатических поясов (Петрушевская, 1986; Амон, 2003). Все новые местонахождения прунобрахид на Северо-Востоке России, рассмотренные ранее (Палечек, 2018), протягиваются в меридиональном направлении и большей частью расположены вдоль береговой линии, что подтверждает точку зрения Э.О. Амона (2000) о характере обитания прунобрахид, а также о связи Палеосибири с Палеоарктикой и Палеопацификой и наличии меридиональных течений, с помощью которых представители этого семейства заселили данные территории в кампанское время (Палечек, 2018).
Таким образом, в пределах Алганского террейна Усть-Бельских гор были выделены несколько тектоностратиграфических комплексов: вулканогенно-кремнистый, вулканогенно-туфотерригенный и туфотерригенный. Изучение особенностей их состава и строения позволило реконструировать ряд палеоструктур, сформированных во фронте Удско-Мургальской островодужной системы. В результате исследований был определен возраст выделенных тектоностратиграфических комплексов и установлено, что кремнистые горизонты различных стратиграфических уровней были образованы в разных палеоклиматических зонах (Палечек и др., 2016).
Описаны мощные выходы кремнисто-базальтовых пород в районе г. Кымъылннай, которые датированы кампанскими формами радиолярий, найденными в регионе впервые (Палечек и др., 2016).
Из ламутской свиты впервые выделены радиолярии кампанского возраста. Во всех образцах, отобранных из ламутской свиты, встречен P. articulatum, являющийся репером для кампанского интервала в Бореальной области. Данные U–Pb датирования обломочных зерен цирконов указывают на накопление осадочного материала в постконьякское время и не противоречат микропалеонтологическим данным (Палечек и др., 2018).
Проведенный анализ показал, что в кампанском веке на территории северо-западной части Корякского нагорья в радиоляриевой фауне доминировали пруноидные и дискоидные формы, характерные для умеренно-холодноводных бассейнов, при присутствии единичных насселлярий. Более половины (52–53%) видов в изученных ассоциациях впервые описаны из кампанских отложений Западной Сибири и Тургайского прогиба (Липман, 1962; Козлова, Горбовец, 1966), остальные – из позднесенонских отложений Калифорнии (Campbell, Clark, 1944; Pessagno, 1976).
Для северо-западной части Корякского нагорья было впервые выделено биостратиграфическое подразделение в ранге слоев с фауной – слои с Prunobrachium articulatum (Палечек, 2018). В настоящее время уровень с Prunobrachium articulatum прослежен на Русской плите, Урале, в Западной Сибири и на Тихоокеанской окраине.
СРАВНИТЕЛЬНЫЙ АНАЛИЗ РАДИОЛЯРИЕВЫХ КОМПЛЕКСОВ ТЕРРЕЙНОВ КОРЯКСКОГО НАГОРЬЯ
Изученные кампан-маастрихтские радиоляриевые ассоциации из вулканогенно-кремнистых отложений различных участков фронтальной части Олюторской зоны в целом схожи по своему таксономическому составу и хорошо коррелируются между собой (табл. 1). В районе мыса Витгенштейна (комплексы Олюторского хребта) выделены менее богатые, но схожие с вышеописанными по таксономическому составу радиоляриевые ассоциации. На Олюторском полуострове из кремнисто-карбонатных пород выделена более богатая радиоляриевая ассоциация, что определяется в первую очередь условиями захоронения. Но у всех изученных ассоциаций есть одна характерная черта – они на 90–95% состоят из видов, первоначально описанных из верхнего мела Калифорнии (Campbell, Clark, 1944; Pessagno, 1976; Foreman, 1968), скв. 275 DSDP (Pessagno, 1975) (рис. 5, 6). Исключением являются два вида, встречаемые в разрезах Олюторского террейна: Cromyosphaera vivienkensis Lipman, описанная Р.Х. Липман (1967) из верхнего мела Северной Камчатки, и Lithomespilus mendosa (Krasheninnikov), отмеченный в разрезах фронтальной части Олюторской зоны, прежде описанный из раннего эоцена Западного Предкавказья (Крашенинников, 1960) и распространенный в позднем палеоцене–раннем эоцене Мексиканского залива (Sanfilippo, Riedel, 1973). В позднемеловых (кампан-маастрихтских) разрезах Олюторского террейна наиболее часто встречаются следующие таксоны: Phaseliforma carinata Pessagno, Lithomespilus mendosa (Krasheninnikov), Amphipyndax stocki (Campbell et Clark), Stichomitra livermorensis (Campbell et Clark), Archaeodictyomitra regina (Campbell et Clark), Dictyomitra densicostata Pessagno, D. multicostata Zittel, Cornutella californica Campbell et Clark, Clathrocyclas hyronia Foreman, Xitus asymbatos (Foreman) (табл. 1, табл. I ). Для фронтальной части Олюторской зоны также характерно присутствие Praestylosphaera hastata (Campbell et Clark) и P. pusilla Campbell et Clark, реже встречается Theocampe altamontensis. В позднемеловых олюторских разрезах отмечено присутствие 17 родов/18 видов Spumellaria и 11 родов/22 вида Nassellaria. При этом практически все вышеперечисленные таксоны, наиболее характерные для Олюторской зоны, распространены также в позднем мелу/палеоцене Новой Зеландии (Hollis, 1997). Так, если Ampipyndax stocki, Dictyomitra multicostata являются космополитами, то такие таксоны, как Stichomitra livermorensis, Theocampe altamontensis, T. vanderhoofi, Bathropyramis sanjoaquinensis, Praestylosphaera pusilla, P. hastata, вероятно, являются биполярными.
Из кремнисто-терригенных пород различных тектоностратиграфических комплексов Алганского террейна получены представительные кампанские ассоциации радиолярий (табл. 1, табл. II). Изученные ассоциации по своему составу заметно отличаются от олюторских. В Алганском террейне повсеместно распространены дискоидные и пруноидные формы, часто составляющие от 85 до 95% изученных ассоциаций и встречаемые в массовом количестве. Здесь присутствуют: Phaseliforma carinata Pessagno, P. meganosensis Pessagno, Prunobrachium articulatum (Lipman), P. crassum (Lipman), P. incizum Kozlova, Pseudobrachium ornatum (Lipman), P. mucronatum (Lipman) Spongurus spongiosus (Lipman), S. quadratus Campbell et Clark, S. concentricus (Lipman), Cromyosphaera vivenkensis Lipman, Cromyodruppa concentrica Lipman, Porodiscus vulgaris Lipman, P. cretaceous Campbell et Clark, Pseudoaulophacus lenticulatus (White), Orbiculiforma vacaensis Pessagno, Crucella aster (Lipman), Rhopalastrum trigonale Lipman, Crucella membranifera (Lipman). Из насселлярий встречены единичные экземпляры Amphipyndax stocki (Campbell et Cl.), Stichomitra livermorensis Camp. et Cl., Dictyomitra densicostata Pessagno, Clathrocyclas cf. hyronia Foreman, Theocampe vanderhoofi (Camp. et Cl.). В общем в изученных ассоциациаях Алганского террейна отмечены 14 родов/20 видов Spumellaria и 5 родов/6 видов Nassellaria (рис. 5). При этом 50% этих ассоциаций составляют виды, первоначально описанные из кампанских отложений Русской платформы и Западно-Сибирской низменности (Липман, 1952, 1962; Козлова, Горбовец, 1966), где эти виды зачастую нацело слагают выделенные ассоциации (Палечек и др., 2018). Калифорнийские виды составляют не более 50% изученных ассоциаций (рис. 6). Важно также отметить существенно меньший размер раковин радиолярий из разрезов Алганского террейна по сравнению с таковыми из Олюторского террейна, что потребовало применения для изучения микрофоссилий оптики с большей разрешающей способностью. По мнению автора настоящей работы, тенденция к существенному уменьшению размера раковин радиолярий наблюдается в ряде меловых разрезов, приуроченных к акваториям Арктического палеобассейна, на что, скорее всего, повлияли условия обитания и геодинамическй режим палеобассейна. Анализ выделенных позднемеловых радиоляриевых ассоциаций различных террейнов Корякского нагорья показал их существенную разницу в таксономическом составе, количественном содержании радиолярий, их морфологических особенностях.
В разрезах Олюторского террейна, занимающего южную часть Корякского нагорья, ассоциации радиолярий практически на 100% состоят из “калифорнийских” видов. В Алганском террейне, расположенном в северо-западной части Корякского нагорья, ассоциации радиолярий на 50% и более состоят из видов, описанных из разрезов Русской плиты и Западной Сибири, и не более чем на 50% из “калифорнийских” видов. Это указывает на отчетливую связь Алганского палеобассейна с Палеоарктикой и Западно-Сибирским морем при одновременном влиянии тихоокеанских течений, с помощью которых могли проникнуть “калифорнийские” виды; а для Олюторского террейна устанавливается тесная связь с Палеопацификой в позднемеловое время (рис. 7).
Рис. 7.
(а) Схема возможной миграции таксонов в кампанское время и (б) гипотетическая реконструкция основных морских течений (показаны стрелками) в Северном полушарии в позднем мелу. Рис. 7а: приведена палеореконструкция для 75 млн лет (PALEOMAP PaleoAtlas для GPlates (http://www.odsn. de/odsn/services/paleomap/paleomap.html)). Римскими цифрами обозначены: I – Олюторская зона, II – Алганская зона. 1 – миграция “сибирских” видов, описанных Р.Х. Липман (1952, 1962) из Русской платформы и Западно-Сибирской низменности; 2 – миграция “калифорнийских” видов (Campbell, Clark, 1944; Foreman, 1968; Pessagno, 1976); 3 – суша; 4 – морские бассейны; 5 – современное положение береговой линии. Рис. 7б: длина стрелок приблизительно соответствует относительной силе течения; палеогеография приведена для раннего маастрихта (по Zeigler, Rowley, 1997, c изменениями А.Б. Германа (2004)). Буквами обозначены: АО – Арктический океан, HS – Гудзонов пролив, NPC – Северо-Пассатное течение, RS – Русский пролив, TS – пролив Тейхерта, TuS – Тургайский пролив, WIS – Западный внутренний пролив, WSB – Западно-Сибирский бассейн. 1 – суша, 2 – мелководные морские бассейны, 3 – глубоководные морские бассейны.
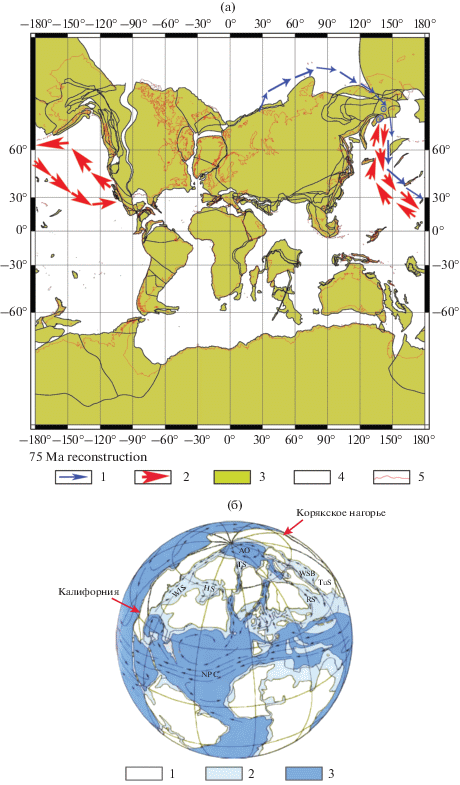
В Алганском палеобассейне обитали виды, как отмечено выше, более чем на 50% состоящие из мигрантов из Сибирского палеобассейна, которые большей частью были представлены пруноидными формами. Географическое распространение позднемелового рода Prunobrachium было детально проанализировано Э.О. Амоном (2003), который пришел к выводу, что этот род возник в сантонское время в Западной Сибири и Зауралье, а затем проник в Поволжье. Из Сибири через Палеоарктику род достиг Северной Пацифики и Калифорнии, а с помощью меридиональных течений в позднем кампане проник в приантарктические акватории, где был образован его нотальный ареал расселения.
По-видимому, в это же время за счет меридиональных течений могло произойти некоторое смешение фаун, когда некоторые калифорнийские виды достигли арктических акваторий. Большей частью это относится к дискоидным пористо-губчатым формам, таким как Phaseliforma carinata Pessagno, P. meganosensis Pessagno, Spongurus quadratus Camp. et Cl., описанных из разрезов Калифорнии (Campbell, Clark, 1944; Foreman, 1968; Pessagno, 1976), в меньшей степени – к насселляриям. Как уже отмечалось выше, находки Nassellaria здесь единичны и представлены 5 родами – Amphipyndax (A. stocki (Camp. et Cl.)), Stichomitra (S. livermorensis (Camp. et Cl.)), Dictyomitra (D. densicostata Pess.), Clathrocyclas (Cl. hyronia Foreman), Thocampe (T. vanderhoofi (Camp. et Cl.)). Но более вероятным представляется сценарий, согласно которому эти таксоны (в первую очередь дискоидные формы) уже существовали в меловое время в изученных палеобассейнах, а право их первоописания принадлежит нашим американским коллегам, так как в нашей стране радиоляриевый анализ начал развиваться позднее. Хорошо известно, что дискоидные формы широко распространены как в древних, так и в современных морях Мирового океана и чаще всего являются космополитными (Кругликова, 1979; Петрушевская, 1986).
Всего несколько таксонов было совместно встречено в Олюторском и Алганском террейнах: из Spumellaria – Phaseliforma carinata Pess., P. meganosensis Pess. и в единичных случаях Cromyosphaera vivenkensis Lipman; из Nassellaria – Amphipyndax stocki (Camp. et Cl.), Stichomitra livermorensis (Camp. et Cl.), Dictyomitra densicostata Pessagno, Clathrocyclas hyronia Foreman, а также единичные представители рода Theocampe (в Олюторском – T. altamontensis, T. yaoi; в Алганском – T. vanderhoofi).
РАДИОЛЯРИЕВЫЕ БИОСТРАТОНЫ
Детальное изучение кампан-маастрихтских радиоляриевых ассоциаций из различных тектоностратиграфических комплексов Олюторской и Алганской зон позволило выделить несколько биостратонов. Так, для Алганской зоны в позднем кампане было выделено биостратиграфическое подразделение в ранге слоев с фауной – слои с Prunobrachium articulatum, которые были впервые прослежены в разрезах северо-западной части Корякского нагорья, п-ова Камчатка и о-ва Шикотан (Палечек, 2018). Эти слои установлены в различных местонахождениях на Северо-Востоке России: в Усть-Бельских и Алганских горах (Палечек и др., 2016, 2018), в районе Чаунской губы (Палечек, 2018), в бассейне р. Ватына на Олюторском п-ове (Вишневская, Басов, 2007) и на п‑ове Камчатка: на Камчатском перешейке (Цуканов и др., 2017), на Западной Камчатке (междуречье Анадырка–Палана) (Сухов, Кузьмичев, 2005), п-ове Камчатский Мыс (Цуканов и др., 2008), Шипунском п-ове и в Леховских горах (Палечек, 2014; Цуканов и др., 2014), а также на о. Шикотан (Курильские о-ва) (Палечек и др., 2008). Совместно с P. articulatum практически во всех изученных нами разрезах присутствует Phaseliforma carinata Pessagno, являющаяся индекс-видом подзоны начала позднего кампана Северной Калифорнии (Pessagno, 1976). Кроме того, на острове Шикотан совместно с радиолярими в малокурильской свите обнаружены единичные остатки иноцерамов, а также проведено радиологическое датирование магматических пород, входящих в состав свиты или располагающихся на контакте с ней (Палечек и др., 2008).
В настоящее время уровень с Prunobrachium articulatum является субглобальным и трассируется от Западной Сибири, где он был впервые установлен, как на запад на Русскую плиту, так и на огромное расстояние на восток.
В Олюторской зоне можно наметить несколько уровней: на границе сантона–кампана – последнее присутствие Orbiculiforma quadrata; в нижнем кампане – присутствие/обилие Dictyomitra densicostata; в верхнем кампане – появление Phaseliforma carinata, Stichomitra livermorensisа; в верхнем кампане/маастрихте – обилие Phaseliforma carinata, Stichomitra livermorensis, Archaeodictyomitra regina, появление Clathrocyclas hyronia. Amphipyndax tylotus в изученных разрезах Олюторской зоны не встречен. Выделяемые биостратоны в ранге слоев с фауной прослежены в разрезах на побережье бухты Анастасии, лагуны Нерпичье озеро (Палечек, 1997; Соловьев и др., 1998), в верховьях рек Ильпи и Матыскен (Палечек, 1997; Соловьев и др., 2000), на мысе Витгенштейна (Палечек, 1997; Богданов и др., 1999), в районе лимана Мачевна (Палечек, 1997, 1999), на Олюторском полуострове (Палечек, 1997) (табл. 2 ).
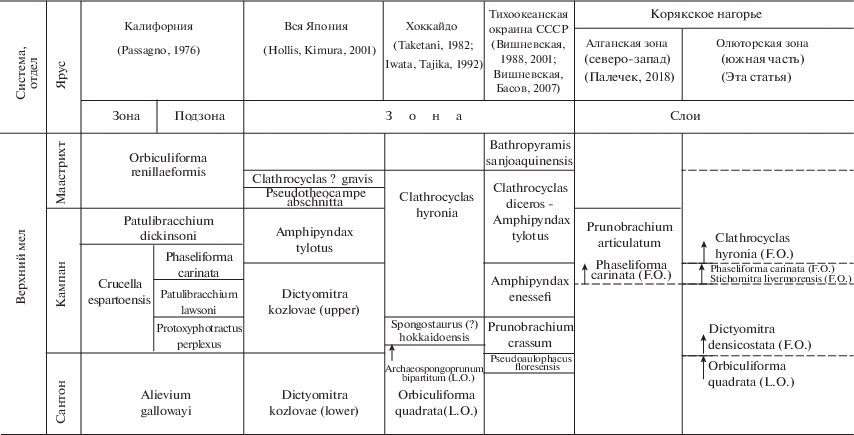
Граница сантона–кампана фиксируется по последнему присутствию Orbiculiforma quadrata, первому появлению представителей рода Praestylosphaera (P. pusilla) и Phaseliforma meganosensis в разрезах вулканогенно-кремнистого комплекса, описанного по побережью бухты Анастасии и в безымянных ручьях, впадающих в бухту (Палечек, 1997; Соловьев и др., 1998), а также в районе лимана Мачевна (Палечек, 1997, 1999).
В нижнем кампане наблюдается первое появление и обилие Dictyomitra densicostata в разрезах вулканогенно-кремнистого комплекса в районе бухты Анастасии, в разрезах осадочно-вулканогенного комплекса в районе руч. Ущельный, лагуны Нерпичье озеро (Палечек, 1997; Соловьев и др., 1998) и в верховьях рек Ильпи и Матыскен (Палечек, 1997; Соловьев и др., 2000), в разрезах флишоидно-олистостромового комплекса мыса Витгенштейна (Палечек, 1997; Богданов и др., 1999), а также кремнистого и осадочно-вулканогенного комплексов в районе лимана Мачевна (Палечек, 1997, 1999). В радиоляриевой ассоциации также присутствуют Praestylosphaera pusilla, P. hastata, Phaseliforma meganosensis, Ampipyndax stocki var. B Vishnevskaya.
В верхнем кампане первое появление Phaseliforma carinata, Stichomitra livermorensis, представителей рода Theocampe (T. altamontensis, T. yaoi, T. vanderhoofi) отмечено в разрезах вулканогенно-кремнистого комплекса в районе бухты Анастасии, верховий рек Ильпи и Матыскен, в разрезах осадочно-вулканогенного комплекса в районе лагуны Нерпичье озеро, мыса Витгенштейна (Палечек, 1997).
В верхах верхнего кампана/маастрихте обилие Phaseliforma carinata, Stichomitra livermorensis, Archaeodictyom-itra regina, первое появление Lithomespilus mendosa, Clathrocyclas hyronia, а также редкие экземпляры Clathrocyclas diceros установлены в разрезах вулканогенно-кремнистого комплекса в районе бухты Анастасии, в верховьях рек Ильпи и Матыскен и бассейне р. Тапельваям (Палечек, 1997; Соловьев и др., 2000).
Ранее В.С. Вишневской (1988, 2001; Вишневская, Басов, 2007) для Тихоокеанского региона для этого интервала были предложены следующие зональные комплексы: с Pseudoaulophacus floresensis (верхний сантон), с Prunobrachium crassum (нижний кампан), с Amphipyndax enesseffi (средний кампан), с Clathrocyclas diceros–Amphipyndax tylotus (верхний кампан–нижний маастрихт), с Bathropyramis sanjoaquinensis (верхний маастрихт–нижний палеоцен?).
По данным В.С. Вишневской (Вишневская, Басов, 2007), в нижнекампанской зоне появляются 6 видов рода Prunobrachium (P. angustum (Lipman), P. crassum (Lipman), P. mucronatum (Lipman), P. sibiricum (Lipman), P. incisum (Kozlova), P. longum (Pessagno)), что согласуется с нашими новыми данными по северо-западной части Корякского нагорья. Нижнекампанская радиоляриевая зона Prunobrachium crassum коррелируется с иноцерамовой зоной Pennatoceramus orientalis, индекс-вид которой определила Е.А. Языкова в типовом местонахождении радиолярий № 64 (61°40′ с.ш., 172°50′ в.д.).
Л.И. Казинцовой (Практическое…, 1999) для Корякского нагорья в маастрихте по шлифам предложены слои с Haliomma (?) intracanthacea–Clathtocyclas hyronia, которые прослежены на Камчатке и Восточном Сахалине.
Таким образом, для северо-западной и южной частей Корякского нагорья для верхнего кампана–маастрихта устанавливаются различные биостратоны по радиоляриям. Выполненное стратиграфическое расчленение базируется на эволюционных изменениях радиоляриевых сообществ. Возрастные интервалы выделены по рубежам одновременного появления и исчезновения комплексов видов или видов-индексов. Границы фиксируются по смене комплексов радиолярий или по первому и, соответственно, последнему достаточно частому присутствию в комплексе видов-индексов, т.е. по резкому увеличению или сокращению обилия экземпляров вида-индекса. По возможности, для сложно построенных тектоностратиграфических разрезов на Северо-Востоке России параллельно с микропалеонтологическим анализом при выделении биостратонов были проведены колибровки и другими возможными методами, такими как радиологическое датирование магматических пород, трековое датирование цирконов. В редких случаях, например на острове Шикотан, в изучаемых отложениях описаны совместные находки макро- и микрофауны (Палечек и др., 2008).
КОРРЕЛЯЦИЯ С РАДИОЛЯРИЕВЫМИ СХЕМАМИ СЕВЕРНОЙ АМЕРИКИ И ЯПОНИИ
Результаты корреляции выделенных подразделений различных районов Тихокеанского региона показаны в табл. 2 . В схеме использованы данные по сопредельным территориям Северо-Востока России: зональные схемы по радиоляриям, предложенные Э. Пессаньо для Калифорнийского побережья (Pessagno, 1976), У. Такетани, Ивата и др. для острова Хоккайдо (Taketani, 1982; Iwata, Tajika, 1992) и всей Японии (Hollis, Kimura, 2001), В.С. Вишневской для Тихоокеанского обрамления бывшего СССР (1988, 2001; Вишневская, Басов, 2007).
ВЫВОДЫ
Таким образом, анализ радиоляриевой микрофауны из различных структурно-формационных зон двух разных террейнов, расположенных в южной и северо-западной частях Корякского нагорья, показал существенную разницу в распределении таксонов, морфологии раковин и связи с различными палеобассейнами. В южной части Корякского нагорья (Олюторский террейн) позднемеловая радиоляриевая фауна практически на 100% состоит из калифорнийских видов. На территории северо-западной части Корякского нагорья (Алганский террейн) в кампанском веке в радиоляриевой фауне доминировали пруноидные и дискоидные формы, характерные для умеренно-холодноводных бассейнов, при присутствии единичных насселлярий. В изученных ассоциациях более половины (52–53%) видов впервые описаны из кампанских отложений Западной Сибири и Тургайского прогиба (Липман, 1962; Козлова, Горбовец, 1966), остальные известны из позднесенонских отложений Калифорнии (Campbell, Clark, 1944; Pessagno, 1976). Размер раковин радиолярий, изученных из позднемеловых разрезов приарктических акваторий, существенно меньше по сравнению с радиоляриями из других разрезов Северо-Востока России. По этим данным отчетливо прослеживается связь Алганского палеобассейна с Палеоарктикой и Западно-Сибирским морем, вероятно, при одновременном влиянии тихоокеанских течений, с помощью которых могли проникнуть калифорнийские виды; а для Олюторского террейна устанавливается тесная связь с Палеопацификой в позднемеловое время. Для северо-западной части Корякского нагорья было впервые выделено биостратиграфическое подразделение в ранге слоев с фауной – слои с Prunobrachium articulatum (Палечек, 2018). В настоящее время уровень с Prunobrachium articulatum является субглобальным и прослежен на всей территории России: на Русской плите, Урале, в Западной Сибири и на Тихоокеанской окраине. Для Олюторской зоны намечено несколько уровней: на границе сантона–кампана – последнее присутствие Orbiculiforma quadrata (L.O.) и первое появление Praestylosphaera pusilla (F.O.) и Phaseliforma meganosensis (F.O.); в нижнем кампане – первое появление и обилие Dictyomitra densicostata; в верхнем кампане – первое появление Phaseliforma carinata, Stichomitra livermorensis, представителей рода Theocampe (T. altamontensis, T. yaoi, T. vanderhoofi); в верхах верхнего кампана/маастрихте – обилие Phaseliforma carinata, Stichomitra livermorensis, Archaeodictyomitra regina, первое появление Clathrocyclas hyronia, а также Lithomespilus mendosa. Рассмотрена корреляция выделенных биостратонов в Корякском нагорье со смежными территориями в Тихоокеанском регионе.
Благодарности. Автор глубоко признателен идейным вдохновителям работ на Северо-Востоке России – ${\text{Н}}{\text{.А}}{\text{.}}\,\,{\text{Богданову}}$, С.Д. Соколову; своим друзьям и коллегам А.В. Соловьеву, Г.В. Ледневой, А.В. Ландеру, А.В. Моисееву за совместные работы в экспедициях на Корякском нагорье; Н.В. Горьковой и Е.А. Жегалло за фотосъемку радиолярий под СЭМ; В.С. Вишневской, Ю.Д. Захарову, А.Б. Герману за советы и замечания при обсуждении статьи.
Источники финансирования. Работа выполнена по теме госзадания ГИН РАН (№ гос. регистрации АААА-А18-118021690155-7) и частично при поддержке РФФИ (грант мол_а_вед № 18-35-20037).
Список литературы
Алексеев Э.С. Основные черты развития и структуры южной части Корякского нагорья // Геотектоника. 1979. № 1. С. 85–95.
Амон Э.О. Верхнемеловые радиолярии Урала // Материалы по стратиграфии и палеонтологии Урала. Вып. 5. Екатеринбург: ИГГУрО РАН, 2000. 209 с.
Амон Э.О. Некоторые закономерности глобального географического распространения позднемеловых радиолярий рода Prunobrachium // Литосфера. 2003. № 4. С. 78–83.
Астраханцев О.В., Казимиров А.Д., Хейфeц А.М. Тектоника северной части Олюторской зоны // Очерки по геологии Северо-Западного сектора Тихоокеанского тектонического пояса. М.: Наука, 1987. С. 161–187.
Богданов Н.А., Чехович В.Д., Сухов А.Н., Вишневская В.С. Тектоника Олюторской зоны // Очерки тектоники Корякского нагорья. М.: Наука, 1982. С. 189–217.
Богданов Н.А., Гарвер Дж.И., Чехович В.Д., Палечек Т.Н., Леднева Г.В., Соловьев А.В., Коваленко Д.В. Обстановки формирования флишоидно-олистостромового комплекса западного побережья Алеутской глубоководной впадины // Геотектоника. 1999. № 5. С. 52–66.
Вишневская В.С. Биостратиграфия вулканогенно-кремнистых образований позднего мела Беринговоморского региона СССР по радиоляриям // Тихоокеанская геология. 1985. № 4. С. 84–93.
Вишневская В.С. О возможностях расчленения юрско-палеогеновых вулканогенно-кремнистых формаций северо-западного обрамления Пацифики (в пределах СССР) // Очерки по геологии Камчатки и Корякского нагорья. М.: Наука, 1988. С. 8–17.
Вишневская В.С. Радиоляриевая биостратиграфия юры и мела России. Москва: ГЕОС, 2001. 376 c.
Вишневская В.С., Басов И.А. Новые данные о биотических событиях на границе сантона и кампана (на примере микропланктона Тихоокеанской окраины России) // Докл. АН. 2007. Т. 417. № 3. С. 364–368.
Вишневская В.С., Филатова Н.И. Радиоляриевая биостратиграфия мезозоя Северо-Востока России // Тихоокеанская геология. 1996. Т. 15. № 1. С. 16–43.
Вяткин Б.В. Государственная геологическая карта СССР м-ба 1 : 200 000. Серия Корякская. Лист Q-59-XXXV, XXXVI. М., 1990.
Геодинамика, магматизм и металлогения Востока России. Ред. Ханчук А.И. Владивосток: Дальнаука, 2006. Кн. 1. 572 с.
Геология юга Корякского нагорья. М.: Наука, 1987. 167 с.
Герман А.Б. Позднемеловой климат Евразии и Аляски: по палеоботаническим данным. Отв. ред. Ахметьев М.А. М.: Наука, 2004. 157 с.
Грецкий В.А. Государственная геологическая карта СССР м-ба 1 : 200 000. Серия Корякская. Лист P-59-IX. М., 1989.
Гульпа И.В. Объяснительная записка к Геологической карте Российской Федерации масштаба 1 : 200 000. Корякская серия. Лист Q_59_XXIX, XXX (Отрожненская площадь). СПб.: ВСЕГЕИ, 2014.
Жамойда А.И. Биостратиграфия мезозойских кремнистых толщ Востока СССР. Л.: Недра, 1972. 243 с.
Захаров В.А. Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Анадырская. Лист Q-59-ХХIХ. Ред. Кайгородцев Г.Г. М.: Аэрогеология, 1974.
Зинкевич В.П. Формации и этапы тектонического развития севера Корякского нагорья. М.: Наука, 1981. 107 с.
Казимиров А.Д., Крылов К.А., Федоров П.И. Тектоническая эволюция окраинных морей на примере юга Корякского нагорья // Очерки по геологии Северо-Западного сектора Тихоокеанского тектонического пояса. М.: Наука, 1987. С. 200–225.
Казинцова Л.И. Меловые радиолярии Корякского нагорья // Советская геология. 1979. № 4. С. 81–85.
Кайгородцев Г.Г. Офиолитовые формации хребта Пекульней // Материалы по геологии и полезным ископаемым Северо-Востока СССР. 1961. Вып. 15. С. 93–104.
Кальянов В.Г., Беляцкая Б.П. Отчет о работе Верхне-Коначанской геологической партии м-ба 1 : 200 000 за 1961 г. Анадырь, 1961.
Козлова Г.Э., Горбовец А.Н. Радиолярии верхнемеловых и верхнеэоценовых отложений Западно-Сибирской низменности. Л.: Недра, 1966. 158 с.
Крашенинников В.А. Некоторые радиолярии нижнего и среднего эоцена Западного Предкавказья // Палеонтологический сборник. Труды Всесоюзного научно-исслед. геологоразвед. нефт. ин-та. 1960. Вып. 16. С. 271–307.
Кругликова С.Б. Характерные черты распределения радиолярий (высоких таксонов) в отложениях Тихого океана и окраинных морей // Ископаемые и современные радиолярии. Л.: Наука, 1979. С. 42–52.
Легенда Корякской серии Государственной геологической карты Российской Федерации масштаба 1 : 200 000 (Чукотская часть). Анадырь, 1999.
Липман Р.Х. Материалы к монографическому изучению радиолярий верхнемеловых отложений Русской платформы // Палеонтология и стратиграфия. Л.: ВСЕГЕИ, 1952. С. 24–51.
Липман Р.Х. Значение радиолярий для стратиграфического расчленения осадочных пород // Бюлл. МОИП. Отд. геол. 1959. Т. 34. Вып. 6. С. 67–88.
Липман Р.Х. Позднемеловые радиолярии Западно-Сибирской низменности и Тургайского прогиба // Материалы по стратиграфии мезо-кайнозоя Тургайского прогиба, Северного Приаралья и Западно-Сибирской низменности. Труды ВСЕГЕИ. Нов. сер. 1962. Т. 77. С. 271–323.
Липман Р.Х. Новые виды радиолярий Северной Камчатки // Биостратиграфический сборник. Труды ВСЕГЕИ. 1967. Т. 129. Вып. 3. С. 89–103.
Митрофанов Н.П. Ватынский тектонический покров в Центрально-Корякской складчатой зоне // Геология и геофизика. 1977. № 4. С. 144–149.
Митрофанов Н.П., Шелудченко С.Д. О возрасте терригенных отложений на юго-западе Центрально-Корякской складчатой зоны // Геология и геофизика. 1981. № 4. С. 128–131.
Моисеев А.В. Структура и история тектонического развития Усть-Бельского сегмента Западно-Корякской складчатой системы (СВ России, Корякия). Автореф. дисс. … канд. геол.-мин. наук. М., 2015. 30 с.
Назаров Б.Б., Витухин Д.И. Методы выделения ископаемых радиолярий // Изв. АН СССР. Сер. геол. 1981. № 10. С. 95–101.
Невретдинов Э.Б., Лебедев В.В. Государственная геологическая карта СССР масштаба 1: 200 000. Серия Анадырская. Лист Q-60-XXXI, XXXII. СПб.: ВСЕГЕИ, 1987.
Очерки тектоники Корякского нагорья. Ред. Пущаровский Ю.М., Тильман С.М. М.: Наука, 1982. 220 с.
Палечек Т.Н. Строение и условия формирования верхнемеловых вулканогенно-кремнистых отложений Олюторского района (на основе радиоляриевого анализа). Автореф. дисс. … канд. геол.-мин. наук. М.: Ин-т литосферы РАН, 1997. 25 с.
Палечек Т.Н. Радиоляриевый анализ осадочно-вулканогенных отложений в восточной части Олюторской зоны (Корякское нагорье) // Геология и разведка. 1999. № 5. С. 34–39.
Палечек Т.Н. Меловые радиолярии Шипунского п-ова (Восточная Камчатка) // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Сборник научных трудов. Ред. Барабошкин Е.Ю., Маркевич В.С., Бугдаева Е.В. и др. Владивосток: Дальнаука, 2014. С. 232–234.
Палечек Т.Н. Распространение и стратиграфический потенциал мезозойских радиолярий семейства Prunobrachidae // Стратиграфия. Геол. корреляция. 2018. Т. 26. № 4. С. 116–129.
Палечек Т.Н., Терехов Е.П., Можеровский А.В. Кампан-маастрихтские радиолярии из малокурильской свиты острова Шикотан // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 5. С. 76–89.
Палечек Т.Н., Моисеев А.В., Соколов С.Д. Новые данные о строении и возрасте юрско-нижнемеловых отложений Алганского террейна (район р. Перевальная, Корякское нагорье, Чукотка) // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 2. С. 43–60.
Палечек Т.Н., Моисеев А.В., Гульпа И.В. Тектоностратиграфия северо-западной части Корякского нагорья, район Усть-Бельских гор // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 4. С. 55–81.
Палечек Т.Н., Моисеев А.В., Гущина М.Ю. Новые данные о возрасте ламутской свиты (Алганские горы, северо-западная часть Корякского нагорья) // Вестник КРАУНЦ. Науки о Земле. 2018. № 4. Вып. 40. С. 105–119.
Парфенов Л.М., Натапов Л.М., Соколов С.Д., Цуканов Н.В. Террейны и аккреционная тектоника Северо-Востока Азии // Геотектоника. 1993. № 1. С. 68–78.
Петрушевская М.Г. Радиоляриевый анализ // Методы зоологических исследований – практике. Л.: Наука, 1986. 200 с.
Практическое руководство по микрофауне. Т. 6. Радиолярии мезозоя. СПб.: ВСЕГЕИ, 1999. 272 с.
Соколов С.Д. Аккреционная тектоника Корякско-Чукоткого сегмента Тихоокеанского пояса. М.: Наука, 1992. 182 с.
Соколов С.Д. Аккреционная тектоника: современное состояние проблемы // Геотектоника. 2003. № 1. С. 13–18.
Соколов С.Д. Очерк тектоники Северо-Востока Азии // Геотектоника. 2010. № 6. С. 60–78.
Соколов С.Д., Бялобжеский С.Г. Террейны Корякского нагорья // Геотектоника. 1996. № 6. С. 68–80.
Соловьев А.В. Геологическое строение и кинематика Ватыно-Вывенского надвига (Корякское нагорье): Автореф. дисс. … канд.-геол. мин. наук. Москва, 1997. 24 с.
Соловьев А.В. Изучение тектонических процессов в областях конвергенции литосферных плит методами трекового датирования и структурного анализа. М.: Наука, 2008. 319 с. (Тр. ГИН РАН. Вып. 577).
Соловьев А.В., Палечек Т.Н., Палечек Р.М. Тектоностратиграфия северной части Олюторской зоны (Корякское нагорье, район бухты Анастасии) // Стратиграфия. Геол. корреляция. 1998. Т. 6. № 4. С. 92–105.
Соловьев А.В., Палечек Т.Н., Леднева Г.В. Кампан-маастрихтские отложения фронтальной части Олюторской зоны (юг Корякского нагорья) // Стратиграфия. Геол. корреляция. 2000. Т. 8. № 2. С. 88–96.
Ставский А.П., Березнер О.С., Сафонов В.Г., Злобин С.К. Тектоника Майницкой зоны Корякского нагорья // Тихоокеанская геология. 1989. № 3. С. 72–80.
Сухов А.Н., Кузьмичев А.Б. Верхнемеловые отложения Западной Камчатки // Западная Камчатка: геологическое развитие в мезозое. Отв. ред. Гладенков Ю.Б., Паланджян С.А. М.: Научный мир, 2005. С. 121–162.
Терехова Г.П. Отчет по теме “Биостратиграфия сеноман-туронских отложений северо-восточной части Корякского нагорья для целей крупномасштабного картирования” за 1985–1987 гг. Магадан, 1987.
Филатова Н.И., Вишневская В.С. Аллохтонные формации среднего мезозоя северо-западного континентального обрамления Тихого океана // Докл. АН. 1992. Т. 323. № 4. С. 734–740.
Цуканов Н.В., Сколотнев С.Г., Палечек Т.Н. Новые данные о составе и строении аккреционной призмы п-ова Камчатский Мыс (Восточная Камчатка) // Вестник КРАУНЦ. Сер. Науки о Земле. 2008. № 12. С. 42–50.
Цуканов Н.В., Палечек Т.Н., Соловьев А.В., Савельев Д.П. Тектоно-стратиграфические комплексы южного сегмента Кроноцкой палеодуги (Восточная Камчатка): строение, возраст и состав // Тихоокеанская геология. 2014. Т. 33. № 4. С. 3–17.
Цуканов Н.В., Палечек Т.Н., Федорчук А.В. Меловые осадочно-вулканогенные комплексы Камчатского перешейка: строение, состав и геодинамические условия формирования // Тихоокеанская геология. 2017. Т. 36. № 2. С. 3–16.
Чехов А.Д. Тектоника Таловско-Пекульнейской зоны // Очерки тектоники Корякского нагорья. М.: Наука, 1982. С. 70–106.
Чехович В.Д. Тектоника и геодинамика складчатого обрамления малых океанических бассейнов. М.: Наука, 1993. 272 с.
Шапиро М.Н. Позднемеловая Ачайваям-Валагинская вулканическая дуга (Камчатка) и кинематика плит Северной Пацифики // Геотектоника. 1995. № 1. С. 58–70.
Шмакин В.Б. Литология верхнемезозойских кремнево-терригенных отложений Севера Корякского нагорья. Автореф. … дисс. канд. геол.-мин. наук. Москва, 1988. 16 с.
Щербинина Е.А., Коваленко Д.В. О возрасте кайнозойских флишево-олистостромовых толщ Олюторского хребта (юг Корякского нагорья) // Стратиграфия. Геол. корреляция. 1996. Т. 4. № 2. С. 110–112.
Campbell A., Clark B. Radiolaria from Upper Cretaceous of Middle California // Geol. Soc. Am. Spec. Pap. 1944. Iss. 57. P. 1–61.
Empson-Morin K. Campanian Radiolaria from DSDP Site 313, Mid-Pacific Mountains // Micropaleontology. 1981. V. 27. № 3. P. 249–292.
Foreman H. Upper Maastrichtian Radiolaria of California // Spec. Pap. Paleontol. 1968. № 3. P. 1–82.
Gorka H. Les Radiolarires du Campanien inferieur de Cracovie (Pologne) // Palaeontologica. Warszawa. 1989. V. 34. № 4. P. 327–354.
Hollis C.J. Cretaceous–Paleocene Radiolaria from Eastern Marlborough, New Zealand // Institute of Geological & Nuclear Sciences Monograph. 1997. № 17. P. 1–152.
Hollis C.J., Kimura K. A unified radiolarian zonation for the Late Cretaceous and Paleocene of Japan // Micropaleontology. 2001. V. 47. № 1. P. 235–255.
Iwata K., Tajika J. Late Cretaceous radiolarians of the Yubetsu group, Tokoro Belt, Northeast Hokkaido // J. Fac. Sci. Hokkaido Univ. Ser. IV. 1986. V. 21. № 4. P. 619–644.
Iwata K., Tajika J. Early Paleogene radiolarians from green and red mudstones in the Yubetsu Group and reconsideration of the age of their sedimentation // Report of the Geological Society of Hokkaido. 1992. V. 63. P. 23–31.
Kravchenko-Berezhnoy I.R., Ledneva G.V., Ivanova E.A., Vishnevskaya V.S. Allochthonous lithotectonic units of the NW Olutor terrane (NE Kamchatka) // Ofioliti. 1993. V. 18 (2). P. 177–180.
Pessagno E. Upper Cretaceous Radiolaria from DSDP Site 275 // Init. Rep. DSDP. 1975. V. 29. P. 1011–1029.
Pessagno E. Radiolarian zonation and stratigraphy of the Upper Cretaceous portion of the Great Valley Sequence, California Coast Ranges // Micropaleontol. Spec. Publ. 1976. V. 2. P. 1–95.
Pessagno E., Newport R. A technique for extracting Radiolaria from radiolarian cherts // Micropaleontology. 1972. V. 18. P. 231–234.
Sanfilippo A., Riedel W. Cenozoic Radiolaria (exclusive of Theoperids, Artostrobiids and Amphipyndacids) from the Gulf of Mexico, DSDP-leg.X. // Init. Rep. DSDP. 1973. V. X. P. 475–612.
Sokolov S.D., Luchitskaya M.V., Silantyev S.A., Morozov O.L., Ganelin A.V., Bazylev B.A., Osipenko A.B., Palanzhyan S.A., Kravchenko-Berezhnoy I.R. Ophiolites in accretionary complexes along the Early Cretaceous margin of NE Asia: age, composition, and geodynamic diversity // Ophiolites in Earth History. London Spec. Publ. 2003. V. 218. № 1. P. 619–664.
Taketani Y. Cretaceous radiolarian biostratigraphy of the Urakawa and Obira areas, Hokkaido // Tohoku Univ. Sci. Repts. 2nd ser. (Geol.). 1982. V. 52. № 1–2. P. 1–76.
Zeigler A.M., Rowley D.B. The vanishing record of epeiric seas, with emphasis on the Late Cretaceous “Hudson Seaway”. Eds. Growley T.J., Burke K. Oxford: Oxford Univ. Press, 1997. P. 147–166.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция