

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 5, стр. 146-156
Моллюски в стратиграфическом расчленении опорного разреза кайнозоя бухты Квачина, Западная Камчатка
Ю. Б. Гладенков *
Геологический институт РАН
Москва, Россия
* E-mail: gladenkov@ginras.ru
Поступила в редакцию 22.01.2018
После доработки 20.09.2019
Принята к публикации 20.11.2019
Аннотация
Дана характеристика комплексов моллюсков одного из опорных разрезов кайнозоя Западной Камчатки – бухты Квачина. В осадочном разрезе мощностью около 450 м выделены 16 стратиграфически сменяющих друг друга слоев с моллюсками, которые группируются в 5 горизонтов. Проведена корреляция биостратонов с региональными горизонтами и определен их возраст в интервале средний эоцен–средний миоцен. Выявлен ряд особенностей палеогеографических и климатических обстановок.
ВВЕДЕНИЕ
Стратиграфическое расчленение геосинклинальных толщ всегда сталкивалось с большими трудностями. Это относится, в частности, к шельфовым формациям кайнозоя переходного региона от океана к континенту Сахалино-Камчатской области. В целом она входит в состав альпийской зоны складчатости, что обуславливает широкое развитие складчатых и разрывных дислокаций, затрудняющих изучение последовательностей древних толщ. Именно здесь развиты осадочные образования большой мощности (следствие лавинной седиментации), отличающиеся значительной фациальной пестротой (с наличием клиноформ разного масштаба); в осадочных сериях часто выявляются перерывы, размывы, а также угловые несогласия (как следствие тектонических и эвстатических событий); вместе с осадочными толщами отмечаются комплексы вулканических образований.
В последние 20 лет стратиграфическое изучение кайнозоя Камчатско-Сахалинской области в широком масштабе, к сожалению, не проводилось. В результате стратиграфические схемы палеогена и неогена этой области, которые были утверждены Межведомственным стратиграфическим комитетом России около 20 лет назад (Решения…, 1998), фактически не изменялись. Вот почему все новые сведения по актуализации стратиграфических схем региона с ориентацией на их возможную детализацию вызывают повышенный интерес. Особенно если они, с одной стороны, базируются на детальном изучении опорных разрезов шельфовых толщ, а с другой – получены с использованием разнообразных методов: биостратиграфического (по разным палеонтологическим группам), палеомагнитного, циклического, изотопного, палеоклиматического и др.
В настоящей работе излагаются новые данные о моллюсках одного из типовых разрезов кайнозоя Западно-Камчатской структурно-формационной зоны – бухты Квачина, относящейся к Тигильскому району, где находится и Точилинский разрез со стратотипами горизонтов региональной стратиграфической схемы (рис. 1) (крупномасштабная геологическая схема района бухты Квачина опубликована в работе (Гладенков, 2016)). Разрез бухты Квачина имеет стопроцентную обнаженность в береговых обрывах морского побережья Охотского моря, что позволяет изучать осадочные толщи и биотические комплексы в стратиграфической последовательности. Здесь на сильно дислоцированных породах (крепкие неразмокающие черные алевролиты и песчаники) нижнего эоцена с резким угловым несогласием и размывом залегают среднеэоценовые–среднемиоценовые отложения.
Рис. 1.
(а) Местонахождение изученного разреза кайнозоя бухты Квачина Западной Камчатки (показано стрелкой) и (б) расположение разреза в бухте Квачина (черная линия).
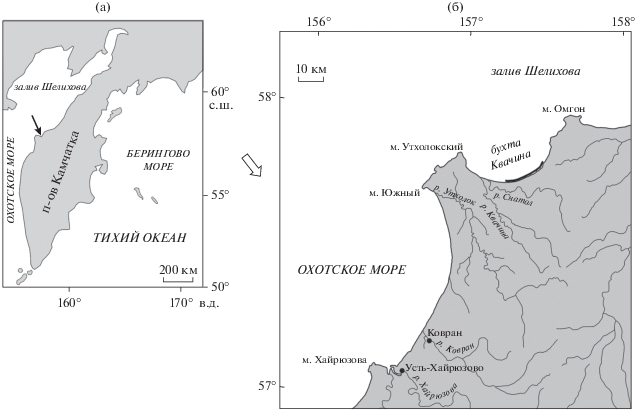
Этот разрез, начиная с 1930-х годов, посещали многие геологи. Среди них сотрудники ВНИГРИ Б.Ф. Дьяков, И.Б. Плешаков (Плешаков, 1939), Ю.Н. Григоренко (Григоренко, 2011) и др., геологи Камчатского геологического управления С.И. Сингаевский, Е.П. Кленов (Геология…, 1964), специалисты из академических институтов (Серова, 2001). В 1984 г. его исследовали сотрудники ГИН РАН Ю.Б. Гладенков, В.Н. Синельникова, А.Е. Шанцер, А.И. Челебаева и Г.М. Братцева. Однако описание этих толщ было сделано недостаточно полно, а определение фауны и флоры было дано в основном для нижней части разреза (Гладенков и др., 1991, 1997). Моллюски тогда были предварительно определены В.Н. Синельниковой и Ю.Б. Гладенковым, флора – А.И. Челебаевой и Г.М. Братцевой, диатомовые (только в верхней части разреза) – Т.В. Орешкиной (2009), а фораминиферы по ряду причин остались неизученными. Недавно в отдельных образцах коллекции 1984 г. были определены диноцисты (Запорожец и др., 2015).
В 2014 г. Ю.Б. Гладенков, С.И. Бордунов и А.Ю. Гладенков дополнительно изучили этот разрез и послойно собрали новые образцы для комплексного исследования палеосообществ моллюсков, фораминифер, диатомовых и диноцист. Частично эти данные уже нашли отражение в публикации (Гладенков, 2016), в которой была приведена детальная разбивка разреза на пачки с привязкой к ним палеонтологических комплексов.
ОБЩИЕ ЗАМЕЧАНИЯ О РАЗРЕЗЕ КАЙНОЗОЯ БУХТЫ КВАЧИНА
Поскольку, как было сказано, этот разрез недавно описан автором (Гладенков, 2016), его подробная характеристика здесь не приводится. Однако представляется полезным отметить некоторые его важные литологические особенности. Разрез включает осадочные толщи (мощностью около 450 м) палеогенового и неогенового возраста. Стратиграфически снизу вверх автором обособлены 37 литологических пачек, относящихся к двум “толщам”, выделенным ранее (Плешаков, 1939): “тигильской” (пачки 1–13) и “белесоватой” (пачки 14–35). Самые верхние пачки 36 и 37 относятся к более молодой “кавранской” серии. Все найденные моллюски привязаны к этим пачкам.
При описании кайнозойских отложений бухты Квачина выявились некоторые особенности литологического строения разреза. Прежде всего, изученные образования (пачки 1–35) составляют крупный седиментационный цикл первого порядка (400 м), который соответствует возрастному интервалу от среднего эоцена до раннего миоцена включительно. В этом цикле выделяются три части: нижняя (пачки 1–13, тигильская толща), средняя (пачки 14–29, низы белесоватой толщи) и верхняя (пачки 30–35, верхи белесоватой толщи). Нижняя часть (средний–верхний эоцен) представлена в основном мелководными песчаниками, конгломератами и гравелитами. Для них характерны косая слоистость, небольшие размывы, углистые линзы, остатки листовой флоры, куски деревьев, шишки и раковины мелководных моллюсков. В терминальной части тигильской толщи (пачки 12–13) появляются туфогенные породы, которые выделены И.Б. Плешаковым в “туфогенный горизонт”.
Средняя часть осадочного цикла (олигоцен) состоит из тонких слоев (0.1–0.3 м) кремнистых туфоалевролитов и диатомитов с прослоями туфов (мощностью от нескольких сантиметров до 0.1–0.2 м). В ее низах отмечаются отдельные глыбы магматических пород и экзотическая галька, которые связываются с ледовым разносом. Иногда присутствуют глендониты. Встречаются редкие маломощные глауконитовые прослои. Характерны остатки относительно глубоководных моллюсков. Терминальная часть седиментационного цикла (нижний миоцен) характеризуется песчаниками и гравелитами с прослоями алевролитов и комплексами относительно мелководных моллюсков.
В разрезе отмечаются два крупных размыва – в основании тигильской толщи и в кровле белесоватой толщи. Нижний из них связан с угловым несогласием между дислоцированными образованиями верхнего эоцена и отложениями среднего эоцена. Верхний приурочен к контакту между белесоватой толщей и кавранской серией миоцена.
Отметим еще три особенности разреза. Во-первых, в породах часто присутствует рассеянная галька (особенно ее много в тигильской толще и в верхней части белесоватой толщи), а в низах белесоватых слоев отмечаются крупные (до нескольких метров в поперечнике) экзотические глыбы. Обычно они сложены вулканическими породами (базальты, андезиты и диориты). Во-вторых, для описанного разреза характерно большое количество карбонатных и песчано-карбонатных конкреций разного размера. Они имеют шаровидную и караваеобразную форму и рассеяны в породе. В тигильской толще отмечены сидеритовые конкреции с коричневатой “рубашкой”. В белесоватой толще крупные конкреции собраны в шесть маркирующих горизонтов (1.0–1.5 м). Местами конкреции приурочены к прослоям туфов. Наконец, практически весь разрез вмещает тонкие прослои туфогенного материала, большая часть которых содержится в белесоватой толще.
СЛОИ С МОЛЛЮСКАМИ И ГОРИЗОНТЫ
Первые обобщения об ископаемых остатках моллюсков из кайнозоя бухты Квачина осуществила в середине прошлого века Л.В. Криштофович (1947). Хотя ее коллекция моллюсков была не очень представительной, но именно на ее основе тогда было намечено сопоставление данного разреза с разрезами соседних районов. В обосновании западнокамчатских горизонтов, выделявшихся в региональных стратиграфических схемах 1950–1970 -х гг., моллюски играли важнейшую роль. Проведенное в последующие годы изучение этого разреза (прежде всего в 1984 и 2014 гг.) позволило собрать более богатые, чем раньше, палеонтологические коллекции с точной привязкой комплексов к литологическим пачкам. Новые материалы стали основой для внесения в стратиграфическое расчленение разреза существенных дополнений. Изученная коллекция состоит из более 600 раковин, ядер и отпечатков моллюсков. Их сохранность в породах не всегда хорошая (лучшая – в верхних слоях), а частота их встречаемости по отдельным пачкам заметно варьирует от скоплений до единичных находок. В пачке 1 (3.7 м) моллюски не найдены. Фотоизображения ряда характерных моллюсков приведены в работах (Гладенков и др., 1991, 2018).
В разрезе бухты Квачина автором выделены 16 последовательно сменяющих друг друга “слоев с моллюсками”. Они объединены в пять “групп слоев” или фаунистических горизонтов, которые для удобства обозначены как “М-горизонты” (моллюсковые горизонты), охарактеризованные определенными биосообществами. В ряде случаев приходится считаться с некоторой размытостью их границ. Отметим, что выделение слоев с моллюсками в горизонтах ранее было осуществлено при изучении Точилинского опорного разреза Тигильского региона Западной Камчатки (Гладенков и др., 2005). Этот опыт был использован при выделении слоев в данной работе.
Ниже дается характеристика слоев в стратиграфическом порядке (рис. 2, 3). Возраст М-горизонтов определялся путем их корреляции с ранее датированными региоярусами (горизонтами) палеогена и неогена стратотипического Точилинского разреза, который расположен в 80–100 км севернее данного разреза. Кроме того, были учтены данные последних лет, полученные по планктонным группам (см. ниже). Ниже при описании слоев подчеркиванием выделены количественно преобладающие формы.
Рис. 2.
Стратиграфический разрез среднего эоцена–среднего миоцена бухты Квачина Западной Камчатки и положение в нем слоев с моллюсками (слои 1–16). 1 – конгломераты, гравелиты; 2 – песчаники, в том числе косослоистые; 3 – аргиллиты и алевролиты; 4 – опоковидные алевролиты; 5 – туфогенные породы; 6 – карбонатные конкреции (мелкие и крупные); 7 – экзотические валуны и глыбы; 8 – размывы и несогласия; 9 – остатки фауны и флоры. Цифры в кружках – номера горизонтов крупных карбонатных конкреций. Х-Н – хулгунско-напанские толщи палеоцена–нижнего эоцена (?), П – палеоцен.
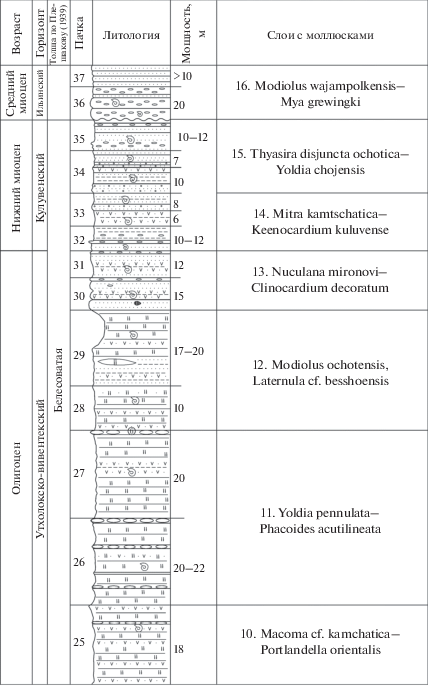
Рис. 2.
Окончание
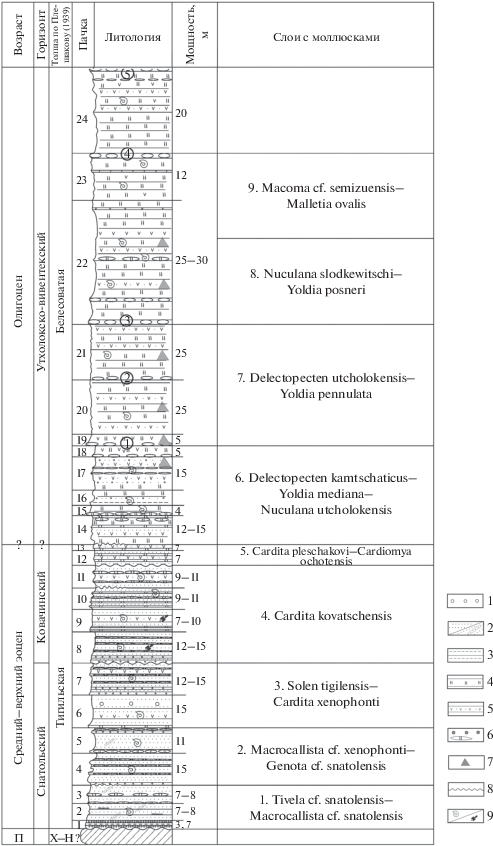
Рис. 3.
Корреляция слоев с моллюсками с горизонтами региональной шкалы кайнозоя Западной Камчатки. Условные обозначения см. рис. 2.

М-горизонт I
Слои 1 с Tivela snatolana–Macrocallista snatolensis (пачки 2 и 3, песчаники, около 20 м). Здесь, помимо указанных форм (Tivela snatolana Slod. и Macrocallista snatolensis L. Krisht.), встречены плохо сохранившиеся мелководные Ostrea sp., Mya sp., Pitar sp., Spisula sp. и др.
Слои 2 с Macrocallista xenophonti–Genota snatolensis (пачки 4 и 5, песчаники, 26 м). Слои характеризуются присутствием Spisula snatolensis L. Krisht., Thyasira snatolensis L. Krisht., Nemocardium cf. marysvillensis Dick., Diplodonta utcholokensis Ilyina, Mya tigilensis L. Krisht., Solen tigilensis Slod., Macoma tigilensis L. Krisht., Venericardia aff. hornii (Gabb.), Gari snatolensis Sin., Macrocallista pleshakov Sin., M. cf. xenophonti Slod., Pholadomya utcholokensis Sin., Genota cf. snatolensis Ilyina. Отмечены также обломки раковин Corbicula sp., Cardita sp., Mytilus sp., Thracia sp., Acila sp., Botula sp., Musculus sp., Ostrea sp., Polinices sp., Dentalium sp.. Этот комплекс в целом является тоже относительно мелководным.
Слои 3 с Solen tigilensis–Cardita xenophonti (пачки 6 и 7, песчаники, около 30 м). Комплекс этих слоев включает Thracia kamtschatica L. Krisht., Mya kovatchensis L. Krisht., Cardita tigilensis L. Krisht., Mytilus cf. littoralis Slod., Macoma tigilensis L. Krisht., Ostrea cf. tigiliana Slod., Limatula kovatchensis L. Krisht., Solen tigilensis Slod., Cardita xenophonti L. Krisht., Crassatella sp., Yoldia sp., Nemocardium sp., Musachia sp. и др. Раковины Mya, Mytilus, Ostrea и Cardita встречаются наиболее часто.
Ранее в интервале слоев 1–3 Л.В. Криштофович (1947) отмечала, помимо указанных видов, следующие формы: Macrocallista snatolensis L. Krisht., Nuculana snatolensis Slod., Nemocardium kovatschensis L. Krisht., Melania cf. snatolensis L. Krisht., Pitar sp., Turritella sp.
Комплекс моллюсков слоев 1–3 в целом весьма сходен с таковым снатольского горизонта (средний эоцен), который широко развит на Западной Камчатке и хорошо распознается по ряду характерных форм – Mactocallista snatolensis, Genota snatolensis, Solen tigilensis и другим видам (хотя в данном разрезе не отмечен Mytilus yakoyama Slod., обычный для этого уровня многих разрезов).
М-горизонт II
Слои 4 с Cardita kovatschensis (пачки 8–11, песчаники, 45 м). В комплекс этих слоев входят Сardita kovatschensis L. Krisht., C. tigilensis, C. xenophonti, Ostrea kovatschensis Slod., Mya kovatschensis, Nemocardium kovatschensis, Sanguilaria snatolensis Sin., Pitar tigilensis L. Krisht., Limatula kovatschensis L. Krisht., Delectopecten cf. kamtschaticus L. Krisht., Nuculana snatolensis L. Krisht., Thyasira snatolensis, Solen sp., Yoldia sp. Cuspidaria sp., Limopsis sp., Natica sp. В сообществе постепенно увеличивается роль тонкораковинных форм средних горизонтов сублиторали (Delectopecten, Nuculana, Thyasira и др.).
Слои 5 с Cardita pleschakovi–Cardiomya ochotensis (пачки 12, 13, песчаники, туфоалевролиты, около 14 м). Здесь отмечены Yoldia cf. longissima Slod., Y. cf. cerussata Slod., Y. watasei Kan., Y. uvutschensis Glad., Anomya iljinensis Sin., Huxleyia snatolana Sin., Cardita pleschakovi L. Krisht., Cardiomya ochotensis L. Krisht., Papyridea utcholokensis Slod., Miltha snatolensis Sin., Acantocardia snatolensis L. Krisht., Limopsis sp., Cuspidaria sp., Delectopecten sp., Ostrea sp., Solen sp., Turritella sp., Dentalium sp. Этот комплекс выглядит несколько обедненным. В нем заметную роль играют представители рода Yoldia.
Ранее Л.В. Криштофович (1947) на этом уровне были отмечены также Cuspidaria napanica L. Krisht., Limatula kovatschensis, Mytilus tigilensis Ilyina, M. littoralis, Nuculana cf. snatolensis, Laternula sp., Ostrea sp., Panope sp. По данным Л.В. Криштофович (1947), в возможных аналогах слоев 4 у мыса Омгон (на северо-восточном крыле синклинали г. Увуч) содержатся Modiolus restorationensis Van Wincle, Yoldia cerussata, Thyasira pervulgata L. Krisht., Nuculana napanica Slod., Turritella sp. Автором там же собраны, помимо указанных видов, остатки Cardita sp., Turcicula sp. и Macoma sp.
Комплекс слоев 4 может сопоставляться с комплексом ковачинского горизонта стратотипического разреза (верхний эоцен). Что касается слоев 5, то они охарактеризованы моллюсками недостаточно представительно, из-за чего их корреляция остается достаточно провизорной. Несколько условным выглядит и их объединение со слоями 4 в один горизонт – М-горизонт II. Присутствие в слоях 5 типичных для аманинской свиты стратотипического разреза видов Yoldia longissimi, Y. cerussata и др. может указывать на наличие в разрезе бухты Квачина пород аманинско-гакхинского уровня (верхний эоцен–олигоцен).
M-горизонт III
Слои 6 с Delectopecten kamtschaticus–Yoldia mediana–Nuculana utcholokensis (пачки 14–18, туфоалевролиты, 50 м). В этих слоях фауна моллюсков заметно меняется. Здесь и выше (слои 7–10) преобладают раковины мелких размеров с тонкостенным строением. В этих слоях встречены скопления остатков Delectopecten kamtschaticus. Помимо них отмечаются Nuculana utcholokensis Ilyina, Yoldia mediana Gladikova, Y. cf. uranoi Kan., Y. cerrusata, Limatula kovachensis L. Krisht., Cardita sp. Указанные формы (Delectopecten, Nuculana и Yoldia) типичны для сообществ нижней сублиторали.
Cлои 7 с Delectopecten utcholokensis–Yoldia pennulata (пачки 19–21, опоковидные аргиллиты, 55 м). Здесь встречены Yoldia mira Gladikova, Y. pennulata Slod., Delectopecten utcholokensis L. Krisht., Nuculana sp., Macoma sp., Cardita sp., Liocyma sp., Malletia sp., Turritella sp., Dentalium sp. Они характерны для относительно больших глубин (нижняя сублитораль или даже батиаль). Раковины обычно мелкие, тонкие и хрупкие.
Cлои 8 c Nuculana slodkevitschi–Yoldia posneri (нижняя часть пачки 22, аргиллиты, 15 м). Комплекс моллюсков этих слоев несколько обедненный. Наиболее часто встречаются два вида-индекса (Nuculana slodkevitschi Kogan и Yoldia posneri L. Krisht.). Они сопровождаются единичными представителями родов Macoma, Laternula, Lima и Cuspidaria.
Слои 9 с Macoma simizuensis–Malettia ovalis (пачки 22 и 23, туфоаргиллиты, около 30 м). Раковины вида Macoma simizuensis L. Krisht. образуют скопления. В этих слоях отмечены также обычно мелкие раковины Yoldia cf. mira Gladikova, Malettia cf. inermis (Yok.), M. ovalis L. Krisht., Portlandella sp., Acila sp., Laternula sp., Nuculana sp., Delectopecten sp. Такое сообщество типично для нижней сублиторали, то есть является относительно глубоководным в шельфовой зоне.
Слои 10 с Macoma kamchatica–Portlandella orientalis (пачки 24 и 25, туфоаргиллиты, 38 м). Комплекс моллюсков, характеризующий эти слои, по набору родов близок к комплексам слоев 8–9. Но здесь появляются новые виды, среди которых преобладают Yoldia aff. chojensis Sim. и Macoma kamchatica Slod. Их сопровождают Y. pennulata Slod., Macoma cf. semizuensis L. Krisht., Portlandella cf. orientalis L. Krisht., Laternula sp., Nuculana sp., Modiolus sp., Delectopecten sp., Dentalium sp.
Слои 11 с Yoldia pennulata–Phacoides acutilineata (пачки 26 и 27, туфоаргиллиты, около 40 м). Здесь отмечено скопление раковин Yoldia pennulata Slod., что делает данный комплекс легко узнаваемым в разрезе. Кроме этого вида встречаются Y. cf. posneri L. Krisht., Phacoides acutilineata (Conrad), Nuculana slodkewichi, Modiolus cf. tichonovichi Mak., Laternula sp., Macoma sp., Musculus sp., Malletia sp., Thyasira cf. disjuncta Gabb., Nuculana sp., Laternula sp., Epitonium sp. С уровня этих слоев появляются относительно крупнораковинные формы моллюсков (модиолусы, тиазиры и др.), обычные для верхнесублиторальной зоны.
Слои 12 с Modiolus ochotensis–Laternula besshoensis (пачки 28 и 29, туфоаргиллиты, около 30 м). В этих слоях руководящими видами являются Modiolus ochotensis Slod. и Laternula cf. besshoensis Yok. И хотя в комплексе еще присутствуют отдельные сравнительно глубоководные Delectopecten aff. pedroanus (Traski), Thyasira sp., Portlandella sp., Macoma sp., постепенно увеличивается число более мелководных форм – Musculus sp., Chlamys sp. и др.
Cлои 13 с Nuculana mironovi–Clinocardium decoratum (пачки 30 и 31, алевролиты и песчаники, 27 м). Ядро сообщества составляют многочисленные представители Nuculana mironovi L. Krisht., Yoldia chojensis., Mya truncata Linney, Serripes groenlandicus (Brug.), Clinocardium decoratum Kogan, Thyasira disjuncta Gabb. Отмечаются отдельные раковины Macoma sp., Modiolus sp., Papyridea sp., Glycymeris sp.
Группа слоев 6–13 по составу комплексов моллюсков может сопоставляться с утхолокско-вивентекским горизонтом Западной Камчатки, который в стратиграфических схемах относится к олигоцену. Он характеризуется типичными для относительно глубоководных ассоциаций моллюсков видами: Delectopecten utcholokensis, Nuculana utcholokensis Ilyina, N. majamraphensis (Khom.), Yoldia pennulata, Y. posneri Slod., Laevicardium puchlense Ilyina, Clinocardium decoratum Kogan, Laternula aff. besshoensis, Turritella gackhensis L. Krisht. и др. Эти формы составляют основу и в слоях 6–13 этого разреза. В нижней части горизонта М-горизонта III отмечены находки Yoldia cerrusata, типичной для аманинско-гакхинского уровня. Маркирующим в верхней части М-горизонта III является сообщество с характерной Nuculana mironovi. Этот уровень хорошо прослеживается во всем Западно-Камчатском регионе.
М-горизонт IV
Cлои 14 с Mitra kamchatica–Keenocardium kuluvense (пачки 32 и 33, песчаники, около 55 м). Комплекс этих слоев заметно отличается от комплексов более древних слоев и в значительной мере представлен сравнительно мелководными верхнесублиторальными видами: Mytilus ochotensis Slod., Mytilus cf. tichanovici Mak., Mitra kamchatica Sin., Keenocardium kuluvense Kafanov, Crepidula cf. ungana Dall, Serripes groenlandicus. Здесь же отмечены Yoldia orientalis L. Krisht., Y. chojensis., Y. nabiliana (Sim.), Y. thraciaeformis (Stor.), Thyasira disjuncta, Laternula sachalinensis Slod., Macoma osacoensisа L. Krisht., a также Glycymeris sp., Panomya sp., Chlamys sp., Mya sp., Spisula sp., Modiolus sp., Musachia sp., Turritella sp. Раковины многих видов часто отличаются большими размерами.
Слои 15 с Thyasira disjuncta ochotica–Yoldia chojensis (пачки 34 и 35, песчаники с конкрециями, около 30 м). Комплекс моллюсков этих слоев в общем близок к таковому слоев 14, но отличается скоплением раковин Yoldia thraciaeformis и Thyasira disjuncta ochotica Gabb. Им сопутствуют Laternula sachalinensis, Serripes groenlandicus, Crepidula cf. ungana, Modiolus sp., Mya sp., Spisula sp. и др. Раковины часто разбиты и деформированы.
В целом комплексы слоев 14 и 15 идентичны сообществу кулувенского горизонта западнокамчатской схемы. Он датируется ранним миоценом и содержит следующие характерные формы: Yoldia chojensis, Y. nabiliana, Laternula sakhalinensis, Crepidula ungana, Thyasira disjuncta ochotica и другие виды. Подошва кулувенского горизонта иногда проводится на литологической основе – по первому появлению в разрезе песчаников (в основании пачек 30–31).
М-горизонт V
Слои 16 с Modiolus wajampolkensis–Mya grewingki (пачки 36 и 37, конгломераты, песчаники, более 30 м). Слои вмещают богатый комплекс моллюсков сублиторального и литорального типов. Они характеризуются скоплением обычно крупных раковин моллюсков, среди которых типичными являются Modiolus wajampolkensis Slod., Mya grewingki Mak., M. cf. cuneiformis (Buhm.), Chlamys cf. kaneharai (Yok.), Arca cf. watanabe (Kanno), Papyridea securiformis Khom., Serripes groenlandicus, Thracia kavranensis Ilyina, Macoma cf. osacaensis L. Krisht., Crassacardia pvella (Yok.), Pododesmus cf. macroshisma (Desh.), Crepidula ungata Dall, Colus aff. rekinnensis Devjat., Ostrea sp., Acmaea sp. и др.
Слои 16 по характерному комплексу моллюсков (Modiolus wajampolkensis, Chlamys cf. kaneharai, Papyridea securiformis и др.) четко коррелируются с ильинским горизонтом региональной схемы, возраст которого определен как начало среднего миоцена. Слои прослеживаются на сотни километров по всей Западной Камчатке.
КОРРЕЛЯЦИЯ БИОСТРАТОНОВ И ПАЛЕОГЕОГРАФИЧЕСКИЕ ОБСТАНОВКИ
В изученном разрезе, в котором ранее выделялись два стратиграфических подразделения – тигильская и белесоватая “толщи”, по комплексам моллюсков можно выделить ряд кайнозойских горизонтов Западной Камчатки (рис. 3): снатольский (средний эоцен), ковачинский (средний–верхний эоцен), утхолокско-вивентекский (олигоцен), кулувенский (нижний миоцен) и ильинский (средний миоцен) (Решения…, 1998). Однако в разрезе с трудом распознается аманинско-гакхинский горизонт верхнего эоцена–нижнего олигоцена. Для него в Точилинском опорном разрезе (мощность до 1000 м) характерны в большом количестве моллюски Yoldia cerussata, Y. nitida, Y. longissima, Modiolus solea, Papyridea harrimani, Turritella tokunagai и иные виды, которые встречаются и в других районах Тигильского региона. Однако в тигильской толще бухты Квачина они практически отсутствуют, хотя в слоях 5 и, возможно, в слоях 6 все же отмечаются редкие Y. cerussata и Yoldia cf. longissima, обычные для аманинского уровня. Именно это обстоятельство позволяет считать, что мощность аманинско-гакхинского горизонта здесь сокращена или в разрезе имеется размыв.
При общем анализе палеокомплексов моллюсков учитывалось, что они приурочены к определенным трофическим группировкам сублиторали и батиали шельфовой зоны (Кузнецов, 1964). На основании данных по 20 разрезам разных районов Западной Камчатки в снатольском горизонте выявлено порядка 135 видов моллюсков, а в ковачинском – около 45. В комплексах этих горизонтов фиксируется большой процент эндемиков (до 40–60%). Но вместе с тем в одновозрастных образованиях Японии, Сахалина и Северной Америки отмечено много общих видов. Исходя из состава комплексов моллюсков снатольского и ковачинского горизонтов и формаций смежных районов, можно говорить о достаточно тесных связях бентосных сообществ шельфовых зон Северо-Тихоокеанской области. По-видимому, эти коммуникации особенно заметно проявились во время значительной трансгрессии и потепления в среднем–позднем эоцене при относительно широком распространении тепловодных форм (субтропических и тропических) (Гладенков и др., 1991).
Устойчивые связи между Северояпономорским, Охотоморским (Присахалинским) и Прикамчатским бассейнами сохранялись и в олигоцене. Сообщества моллюсков этого возраста с характерными видами родов Yoldia, Nuculana, Delectopecten и других, которые относятся в целом к сравнительно холодноводному типу, широко развиты в указанных районах и прослеживаются в Беринговоморский бассейн, к Корякскому нагорью, Аляске и Североамериканскому побережью (Гладенков и др., 2005). Об относительном похолодании в олигоценовое время может свидетельствовать появление в этой части разреза гальки и глыб, которое связывают с ледовым разносом.
Сообщества моллюсков раннего миоцена (кулувенский горизонт) в общем наследуют олигоценовые биоты, но в них отмечается увеличение доли относительно теплолюбивых форм (до 25% и более) и появляется несколько современных форм (4–6), число которых заметно увеличивается выше по разрезу.
Комплексы моллюсков ильинского горизонта начала среднего миоцена (время климатического оптимума) с типичными тепловодными Arca, Chlamys, Crepidula и пр. тоже четко прослеживаются в Северо-Тихоокеанской области – от Японии до Берингова пролива (Гладенков, Синельникова, 1990). Это свидетельствует о тесных связях шельфовых биот того времени на громадных пространствах этих акваторий и о значительных миграциях донных относительно теплолюбивых комплексов от южных (японских) широт к северным (на 12–15 градусов по широте).
В настоящее время уже изучены палеонтологические остатки других групп (фораминиферы, диатомовые, споры и пыльца с диноцистами) из разреза бухты Квачина, что позволяет проводить его корреляции по разным группам фоссилий (Гладенков и др., 2018). По комплексам фораминифер (данные С.И. Бордунова, Т.В. Дмитриевой и Н.А. Фрегатовой), как и по моллюскам, выделяются практически все горизонты Западной Камчатки от среднего эоцена до среднего миоцена: снатольский, ковачинский, утхолокско-вивентекский, кулувенский, ильинский. Аманинско-гакхинский горизонт верхнего эоцена–нижнего олигоцена выделен в сокращенной мощности. С другой стороны, наличие планктонных групп позволило на основе сопоставления с комплексами зон стратиграфических шкал по кайнозою Мирового океана более точно, чем раньше, определить возраст местных стратонов и положение их границ. Фактически впервые (данные Г.Н. Александровой по диноцистам и А.Ю. Гладенкова по диатомовым) достаточно уверенно намечено положение границ отделов и ярусов: эоцена и олигоцена (в самой верхней части тигильской толщи), нижнего и верхнего олигоцена (рюпеля и хатта), олигоцена и миоцена. Решена важная геологическая проблема – впервые точно установлен возраст базальных пачек этого разреза (как средний эоцен) и подстилающих деформированных толщ (как ранний эоцен, а не мел, как предполагалось раньше). Это сделало возможным определить время и продолжительность проявления крупной фазы тектогенеза на Камчатке, после которой наступила широкая морская трансгрессия (снатольско-ковачинская), захватившая практически всю Камчатку. Данные по бухте Квачина представляются чрезвычайно важными, так как в Точилинском стратотипическом разрезе нижняя часть снатольской свиты не обнажена и проблема возраста ее нижних слоев не была решена. Кроме того, впервые на Камчатке было проведено обоснованное выделение ярусов олигоцена (с их границей внутри белесоватой толщи).
К сказанному следует добавить, что, помимо биостратиграфических, в бухте Квачина были выполнены также изотопные и палеомагнитные исследования, которые существенно дополнили данные по стратиграфическим корреляциям и палеогеографическим обстановкам. Так, изотопное изучение диагенетических карбонатных конкреций, как и раковин морских моллюсков, показало, что в конкрециях в средней части разреза (олигоцен) отмечается относительно повышенное содержание метаногенной углекислоты (Покровский, Гладенков, 2017). Это свидетельствует о сравнительно глубоководных обстановках формирования вмещающих конкреции пород (что подтверждает данные, полученные по моллюскам) и переработке органического вещества, сносимого в морской бассейн. Палеомагнитный анализ (данные В.Ю. Водовозова и В.М. Трубихина) позволил провести корреляцию разреза с палеомагнитными хронами Мировой палеомагнитной шкалы (наилучшие результаты получены для олигоценового интервала, в котором намечены хроны С13–С9). Предложенный вариант корреляции в целом согласуется с биостратиграфическими построениями. Все новые материалы (в частности, по разным палеонтологическим группам разреза) подробно отражены в монографии (Гладенков и др., 2018).
ЗАКЛЮЧЕНИЕ
Изучение кайнозойского разреза бухты Квачина Западной Камчатки показало, что при расшифровке особенностей развития морских шельфовых систем Северной Пацифики одно из важных мест, как и раньше, принадлежит бентосным группам, в частности моллюскам. Установление стратиграфической последовательности комплексов (палеосообществ) моллюсков позволяет использовать ее в качестве основы как при расчленении древних толщ отдельных районов на биостратоны, так и при стратиграфической корреляции регионального масштаба в пределах Японско-Сахалинской и Камчатской областей. Это сейчас особенно востребовано при геологическом картировании и бурении в шельфовых зонах.
Но, конечно, сопоставление выявленных планктонных групп с комплексами шкал по кайнозою Северной Пацифики и Мирового океана позволяет более точно, чем раньше, определять возраст местных стратонов и положение их границ. Появление новых и детальных стратиграфических сведений по опорным разрезам кайнозоя данного региона дает возможность лучше понять особенности эволюции морских экосистем региона и использовать эти данные для расшифровки строения осадочных формаций геосинклинального типа.
Источники финансирования. Работа выполнена по теме госзадания ГИН РАН № АААА-А18-118021690155-7, при поддержке проекта РФФИ № 19-05-00361 и проекта Программы фундаментальных исследований Президиума РАН № 17.
Список литературы
Геология СССР. Т. XXXI. Камчатка, Курильские и Командорские острова. Ч. 1. Геологическое описание. М.: Недра, 1964. 734 с.
Гладенков Ю.Б. Расчленение опорного разреза кайнозоя бухты Квачина Западной Камчатки и проблемы его корреляции // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 6. С. 101–113.
Гладенков Ю.Б., Синельникова В.Н. Моллюски и климатические оптимумы миоцена Камчатки. М.: Наука, 1990. 174 с.
Гладенков Ю.Б., Синельникова В.Н., Шанцер А.Е. и др. Эоцен Западной Камчатки. М.: Наука, 1991. 184 с.
Гладенков Ю.Б., Шанцер А.Е., Челебаева А.И. и др. Нижний палеоген Западной Камчатки (стратиграфия, палеогеография, геологические события). М.: ГЕОС, 1997. 367 с.
Гладенков Ю.Б., Синельникова В.Н., Челебаева А.И., Шанцер А.Е. Биосфера–экосистема–биота в прошлом Земли. Экосистемы кайнозоя Северной Пацифики: эоцен–олигоцен Западной Камчатки и сопредельных районов. М.: ГЕОС, 2005. 480 с. (Труды ГИН РАН. Вып. 540).
Гладенков Ю.Б., Гладенков А.Ю., Бордунов С.И. и др. Опорный разрез кайнозоя Западно-Камчатской структурно-фациальной зоны. М.: Геокарт–ГЕОС, 2018. 202 с.
Григоренко Ю.Н. Палеоцен-эоценовый граувакковый комплекс тыловых прогибов Притихоокеанской окраины (строение и формирование). СПб.: ВНИГРИ, 2011. 322 с.
Запорожец Н.И., Ахметьев М.А., Витухин Д.И. и др. Шкала морского палеогена Западной Камчатки (Тигильский район) по диноцистам // Бюлл. МОИП. Отд. геол. 2015. Т. 90. Вып. 1. С. 18–41.
Криштофович Л.В. Стратиграфия и фауна тигильской толщи западного побережья Камчатки. Л.: Ленгостоптехиздат, 1947. 151 с. (Труды ВНИГРИ. Новая сер. Вып. 23).
Кузнецов А.П. Распространение донной фауны западной части Берингова моря по трофическим зонам и некоторые общие вопросы трофической зональности // Труды Института океанологии. 1964. Т. 69. 180 с.
Орешкина Т.В. Новые данные по диатомеям из морских палеогеновых отложений Западной Камчатки // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 3. С. 104–119.
Плешаков И.Б. Третичные отложения Утхолокского района на западном побережье Камчатки. Л.–М.: ГОНТИ НКТП СССР, 1939. 38 с. (Труды ИГРИ. Сер. А. Вып. 123).
Покровский Б.Г., Гладенков Ю.Б. Условия диагенетического карбонатообразования в кайнозойских отложениях Западной Камчатки по изотопным (С и О) данным // Вестник КРАУНЦ. Науки о Земле. 2017. № 4. Вып. 36. С. 5–12.
Решения рабочих межведомственных региональных стратиграфических совещаний по палеогену и неогену восточных районов России – Камчатки, Корякского нагорья, Сахалина и Курильских островов. Объяснительная записка к стратиграфическим схемам. М.: ГЕОС, 1998. 147 с.
Серова М.Я. Фораминиферы и биостратиграфия верхнего палеогена Северной Пацифики. М.: Наука, 2001. 215 с.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция