

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 5, стр. 78-100
Аммониты семейства Cardioceratidae из верхнего келловея севера Сибири
А. С. Алифиров 1, *, В. Г. Князев 2
1 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
2 Институт геологии алмаза и благородных металлов ЯНЦ СО РАН
Якутск, Россия
* E-mail: alifirovas@ipgg.sbras.ru
Поступила в редакцию 11.04.2019
После доработки 16.07.2019
Принята к публикации 20.11.2019
Аннотация
Представлены результаты изучения келловейских аммонитов рода Longaeviceras и близких к нему таксонов c восточного берега Анабарской губы (обн. 109, север Сибири). Установлены ранее неизвестные здесь роды и виды кардиоцератид: Rondiceras milaschevici, Pseudolongaeviceras densicostatum gen. et sp. nov., Platylongoceras holtedahli, Pseudocadoceras laminatum. Не подтвердилось присутствие в изученном разрезе зоны Eboraciceras subordinarium. Приведены аргументы, доказывающие неправомерность разделения рода Longaeviceras на два рода – Longoceras и Longaeviceras, а также включения в последний подрода Soaniceras. По находке Cardioceras maltonense уточнен возраст оксфордской части разреза.
ВВЕДЕНИЕ
Аммониты семейства Cardioceratidae Siemiradzki, 1891, которое существовало с байоса по кимеридж, имеют первостепенное значение для стратиграфии средне-верхнеюрских отложений Панбореальной надобласти. В настоящей статье представлены новые данные по систематике верхнекелловейского рода кардиоцератид Longaeviceras и близких к нему таксонов, а также по ранее неизвестным находкам аммонитов келловея и оксфорда в разрезе юры на восточном берегу Анабарской губы.
Род Longaeviceras имел широкое географическое распространение в бореальных и суббореальных акваториях позднего келловея. Он является хорошим показателем главным образом нижней зоны верхнего келловея Европы – Peltoceras athleta или ее бореального аналога – Keyserlingi. Типовая коллекция кардиоцератид, по которой был выделен этот род, происходит из верхнего келловея Англии (Buckman, 1918 in Buckman 1909–1930; Callomon, Wright, 1989), при этом все изображенные в публикациях до настоящего времени топотипы представлены внутренними оборотами макроконхов. Характеристика внешних оборотов, включая жилую камеру, для представителей данного рода была дана на экземплярах Д.Н. Соколова (1912) из бассейна р. Печора (Arkell et al., 1957; Меледина, 1977), описанных под другими видовыми названиями. Таким образом, основу видового состава рода Longaeviceras образовали виды L. longaevum (Leckenby), L. placenta (Leckenby), L. nikitini (Sokolov), L. keyserlingi (Sokolov). В дальнейшем к рассматриваемому роду были отнесены такие виды, как “L.” staffinense Sykes (нижний оксфорд), “L.” novosemelicum Bodylevsky (верхний келловей) и “L.” polonicum Callomon et Wright. Некоторые специалисты отметили несоответствие морфологических признаков этих видов и родового диагноза Longaeviceras и стали относить их к новым таксонам: Percacosticeras Kiselev, Platylongoceras Repin (Киселев, 1996; Репин, 2002). Дж. Калломон и Дж. Райт (Callomon, Wright, 1989) пришли к выводу, что дальнейшее систематическое исследование аммонитов переходных от Cadoceras к Quenstedtoceras (т.е. Longaeviceras) должно быть основано на новом материале из Арктики. Позднее ревизию систематики аммонитов, относящихся к рассматриваемой группе, провел Ю.С. Репин, который установил новые роды, а также предложил выделять новые зональные подразделения, основанные на данных аммонитах (Репин, 2002, 2008).
Материалом для настоящего исследования послужила коллекция кардиоцератид, собранная авторами в разные годы на восточном берегу Анабарской губы и насчитывающая несколько десятков экземпляров различной степени сохранности (рис. 1). Часть изученной коллекции уже публиковалась ранее (Меледина, 1977, 1994), однако новые сборы указали на необходимость ревизии прежних данных. В ранних работах все аммониты из обнажения 109 (пачки 46–47б; по Меледина, 1994) относились главным образом к роду Longaeviceras: L. keyserlingi (Sok.), L. aff. nikitini (Sok.), L. stenolobum (Sok.), L. bodylevskyi (Meled.). Также указывались редкие находки Pavloviceras omphaloides (Sowerby) и Eboraciceras subordinarium Buckman (Столярова, Меледина, 2004). Исследование обновленной коллекции аммонитов с восточного берега Анабарской губы позволяет установить новый верхнекелловейский род Pseudolongaeviceras, а также выявить ранее неизвестные в данном местонахождении виды.
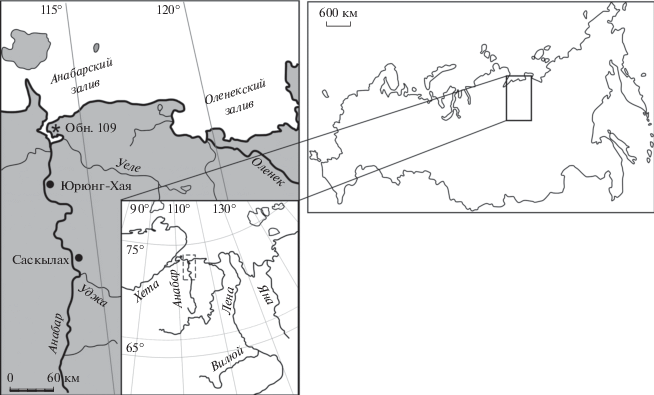
СТРАТИГРАФИЧЕСКИЙ ОЧЕРК
Описание разреза келловея восточного берега Анабарской губы неоднократно приводилось в литературе (Басов и др., 1967; Стратиграфия…, 1976; Меледина, 1977, 1994; Захаров, Шурыгин, 1978; Каплан и др., 1979; Князев и др., 2010).
Верхний келловей на восточном берегу Анабарской губы подстилается слоем алевролитов крупнозернистых, крупнооскольчатых, темно-серых, глауконитовых, с линзами ракушников, пиритовых конкреций в нижней части слоя (мощность 2.2 м; рис. 2). Возраст этого слоя определялся как ранний келловей по находкам Cadochamoussetia tschernyschewi (D. Sokolov) (Князев и др., 2010). Поскольку этот слой накапливался в мелководных условиях при активной гидродинамике, он может быть конденсированным, а находка в нем Rondiceras milaschevici (Orbigny) может свидетельствовать о ранне-среднекелловейском возрасте пород (табл. VI, фиг. 4 ). Также в литературе высказывалось мнение о том, что этот слой частично имеет позднекелловейский возраст (Басов и др., 1967; Захаров, Шурыгин, 1978).
Рис. 2.
Разрез келловея на восточном берегу Анабарской губы (обн. 109). Показанная на рисунке зона Longaeviceras keyserlingi переименована в настоящей работе в L. nikitini. Условные обозначения: 1 – алевролиты известковистые, 2 – алевролиты, 3 – глины алевритовые, 4а – глауконит, 4б – пирит, 5 – поверхность несогласия, 6 – конкреции известковистые. Зоны бореального стандарта приведены по (Никитенко и др., 2013; Князев и др., 2020), название свиты – по (Галабала, 1991), номера пачек – по работам (Меледина, 1994, в колонке “Пачка”) и (Басов и др., 1967, в колонке “Пачка*”).
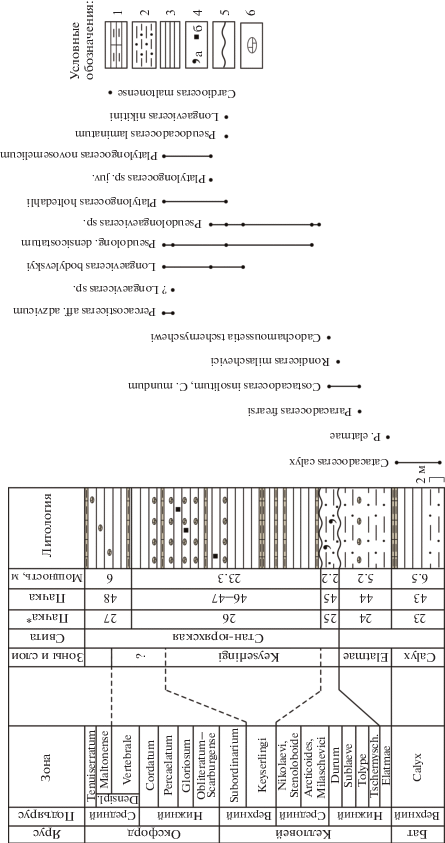
Следы размыва или перерыва в осадконакоплении между пачками 45 и 46 в явном виде не выражены. Верхнекелловейские породы представлены аргиллитами темно-серыми, оскольчатыми, ожелезненными, с многочисленными горизонтами конкреций, в которых последние либо образуют сплошные пласты, либо встречаются разрозненно. Аммониты присутствуют на разных уровнях в нескольких конкреционных горизонтах. Данные о мощности верхнего келловея разнятся: 23.3 м (по А.С. Алифирову), 26 м (по В.Г. Князеву и В.А. Басову) и 30 м (по С.В. Мелединой).
После выхода в свет работ (Басов и др., 1967; Стратиграфия…, 1976; Меледина, 1977; Захаров, Шурыгин, 1978) данная толща относилась к нижней зоне верхнего келловея Longaeviceras keyserlingi. Однако впоследствии здесь была установлена еще одна зона – Eboraciceras subordinarium (Меледина, 1994; Шурыгин и др., 2000), которая была обособлена в пачку 47б с поверхностью размыва в основании, отмеченной галькой и кусками окремнелой древесины. В конкрециях были обнаружены аммониты диаметром до 150 мм, определенные С.В. Мелединой (1994) как Eboraciceras subordinarium Buckman (коллекция № 489), но не изображенные. Отметим, что таксономический состав аммонитов в пачках 45 и 46 по данным разных исследователей отличается друг от друга. В пачке 45 (= 25 по: Басов и др., 1967) В.А. Басовым упоминаются Longaeviceras spp., а в пачке 46 (= 26) – Cadoceras aff. stenolobum Keyserling, C. sp. (cf. innocentii Bodyl.), Longaeviceras cf. novosemelicum Bodyl., L. cf. nikitini (Sokolov).
Верхний келловей перекрывается глинами черными, рыхлыми, мелкооскольчатыми, с караваеобразными и округлыми септариевыми конкрециями глинистого известняка. Видимая мощность этой пачки (= 48 по: Меледина, 1994 или = 27 по: Басов и др., 1967) составляет 6 м. Возраст пачки определен как поздний оксфорд по находке Amoeboceras cf. kostromense (Nikitin). Далее разрез прерывается распадком, за которым выходят нижнемеловые отложения.
Новые полевые наблюдения и палеонтологические сборы, проведенные В.Г. Князевым и А.С. Алифировым, позволяют предложить иное стратиграфическое расчленение верхнекелловейской и оксфордской частей разреза восточного берега Анабарской губы (обн. 109) и дополнить его палеонтологическую характеристику по аммонитам.
Верхний келловей в обн. 109 представляет собой литологически однородную толщу без видимых следов размыва в основании и со стратиграфическим несогласием в кровле. Присутствие здесь зоны Subordinarium, выделенной С.В. Мелединой (1994), не подтвердилось. Пачка 47б (мощность 2.5 м) нами не идентифицирована.
По всей пачке 46–47 распространен новый род кардиоцератид Pseudolongaeviceras gen. nov. Определимые до вида представители рода Longaeviceras встречаются в нижней части пачки до уровня 11.6 м (L. nikitini (Sok.)). На этом уровне обнаружены первые Platylongoceras holtedahli (Salfeld) и один экземпляр Pseudocadoceras laminatum (Buckman), а выше (в 13.5 м и в 19.3 м выше подошвы) встречены также P. novosemelicum (Bodyl.). В верхней части пачки на уровне 18.2 и 19.3 м обнаружены два экземпляра Percacosticeras aff. adzvicum Repin (= Pavloviceras omphaloides (Sowerby) в Столярова, Меледина, 2004, табл., фиг. 3 ). Для рода Pavloviceras характерны только двойные ребра, тогда на экземпляре, изображенном Л.Р. Столяровой, присутствуют вставные ребра. На одном уровне с ними найдены Pseudolongaeviceras bodylevskyi (Meled.) и Longaeviceras/Pseudolongaeviceras sp. (экз. 489-69; Меледина, 1977, табл. 26 , фиг. 3 а, 3б).
В осыпи пачки В.Г. Князевым обнаружены по одному экземпляру Percacosticeras aff. adzvicum Repin и ? Dolganites sp. (табл. VI, фиг. 1).
По-видимому, за Eboraciceras subordinarium (Buckman) ранее были приняты крупные Longaeviceras nikitini (Sok.) (см. рубрику “Замечание” в описании вида).
Таким образом, стратиграфическое распространение определенных нами видов аммонитов из пачки 46–47 разреза на восточном берегу Анабарской губы свидетельствует в пользу отнесения этой толщи к зоне Longaeviceras keyserlingi.
ОПИСАНИЕ АММОНИТОВ
При описании аммонитов используются следующие сокращения замеров параметров раковин: Д – диаметр (мм), Ш – ширина (толщина) (мм), В – высота (мм), У – диаметр умбиликуса (мм), РВ – ребра вентральные (мм), РУ – ребра умбиликальные (первичные) (мм), РО – реберное отношение РВ/РУ. Все отношения параметров приводятся в %, кроме РО. Пропуски измерений связаны с плохой сохранностью экземпляров.
НАДСЕМЕЙСТВО STEPHANOCERATOIDEA NEUMAYR, 1875
СЕМЕЙСТВО CARDIOCERATIDAE SIEMIRADZKI, 1891
ПОДСЕМЕЙСТВО CADOCERATINAE HYATT, 1900
Род Longaeviceras Buckman, 1918 emend. Arkell, 1957
Longaeviceras: Buckman, 1918 in Buckman, 1909–1930, p. 504; Reeside, 1919, p. 14; Arkell et al., 1957, p. 302; Бодылевский, 1960, с. 79; Меледина, 1977, с. 134; Århus et al., 1989, p. 42, figs. 3b–3c ; Callomon, Wright, 1989, p. 823; Меледина, 1994, с. 20; Репин и др., 2006, с. 62; Howarth, 2017, p. 73; Kiselev, Rogov, 2018, p. 94.
Longoceras: Репин, 2002, с. 33; Репин, 2008, с. 1.
Типовой вид: Ammonites longaevum Leckenby (Bean MS), 1859. Лектотип типового вида изображен в (Leckenby, 1859, pl. 2, fig. 1), переизображен С. Бакмэном в 1918 г. (Buckman, 1909–1930, pl. CXLVIII); келловей, Йоркшир, Англия.
Диагноз. Конечные размеры взрослых особей достигают 150 мм (возможно, и более) в диаметре. Обороты до диаметра 50–80 мм оксиконические, с большей или меньшей степенью килеватости, с узким умбиликусом, дугообразными двойными, одиночными и вставными ребрами. На этой стадии появляется привентральное ветвление ребер или ребра с высокой точкой вставления. С ростом раковины, по мере формирования кадикона, скульптура трансформируется в толстые редкие ребра; в нижней части боковых сторон появляются буллы, начинающиеся от умбиликального перегиба, а ребра имеют слабый наклон вперед; вентральный выгиб ребер к устью ослабевает или исчезает вовсе. Отношение первичных и вторичных ребер меняется в широких пределах (от 2 до 3.5) и увеличивается по мере роста раковины. Последние 1.5–2 оборота лишены скульптуры, они имеют кадиконическую форму раковины.
Сравнение. Род Longaeviceras отличается от Rondiceras уже при диаметре 10 мм. Для Rondiceras, близких к виду R. milaschevici, характерно раннее сглаживание ребер, начиная с боковых сторон от умбиликального шва, и большее реберное отношение. Другие виды Rondiceras имеют более длинную скульптурированную стадию, однако при диаметре 50–60 мм скульптура у них также сглаживается, начиная с нижней части боковых сторон. Род Stenocadoceras отличается сохранением уплощенной, но не килеватой формы раковины до конца ее роста и характером развития скульптуры в онтогенезе – отсутствием высокого вставления и ветвления, дифференциации первичных и вторичных ребер, булл и т.д. Для рода Quenstedtoceras характерно раннее развитие дифференциации первичных и вторичных ребер с высоким реберным отношением. У типичных Quenstedtoceras, таких как Q. lamberti, существенно более широкий (в 1.5–2 раза) умбиликус, а у многих видов этого рода отсутствует кадиконическая стадия в онтогенезе раковины.
Видовой состав. L. longaevum (Leckenby), L. placenta (Leckenby), L. nikitini (Sokolov), L. lahuseni (Bodylevsky), ? L. principale (Sasonov), L. bodylevskyi (Meledina).
Замечание. Род Longaeviceras выделен С. Бакмэном в 1918 г. (Buckman, 1909–1930) с типовым видом L. longaevum (Leckenby, 1859). В работе Дж. Лекенби был упомянут и другой вид, очень близкий к типовому – Longaeviceras placenta (Leckenby, 1859). Лектотипы L. longaevum и L. placenta представлены ядрами фрагмоконов средних размеров (50 и 58 мм). Важное дополнение в диагноз рода внес В. Аркелл (Arkell et al., 1957), указав на наличие у Longaeviceras стадии с гладкой кадиконовой формой раковины, которая была описана на экземплярах из Печорского бассейна (Соколов, 1912). Этот уточненный диагноз был принят отечественными специалистами по аммонитам (Бодылевский, 1960; Меледина, 1977, 1994). Со времени выделения рода и до публикации работ Ю.С. Репина (2002, 2008; Репин и др., 2006) менялся в основном лишь его видовой состав. Подродовая классификация Longaeviceras использовалась в работе С.В. Мелединой (1998), где, кроме номинального подрода, в его состав был включен подрод L. (Percacosticeras) с такими видами, как L. novosemelicum, L. polonicum, L. holtedahli. Д.Н. Киселев отнес к роду Longaeviceras несколько среднекелловейских видов: L. praestenolobum Kiselev et Meledina, L. stenolobum (Keyserling), L. alpha Kiselev, родовая принадлежность которых трактуется по-разному (Киселев, 2001, 2006; Киселев, Меледина, 2004; Репин и др., 2006; Князев и др., 2015, 2020; Митта, 2016).
Ю.С. Репин предложил для аммонитов, ранее относившихся к роду Longaeviceras, систему, существенно отличающуюся от традиционной. Из рода Longaeviceras были исключены все печорские и сибирские виды, для которых было предложено название Longoceras Repin, 2002. Сам род Longaeviceras разделен Ю.С. Репиным на два подрода: L. (Longaeviceras), включающий только типовой вид L. longaevum, и установленный ранее С.В. Мелединой (1977) L. (Soaniceras) c типовым видом Quenstedtoceras (Soaniceras) angustatum Meledina. Ряд видов (L. polonicum, L. novosemelicum, L. staffinense, L. hotedahli) включены им в состав родов Percacosticeras Kiselev, 1996 и Platylongoceras Repin, 2002.
Судя по имеющимся в литературе изображениям, а также по описанию, приведенному в работе Дж. Калломона и Дж. Райта (Callomon, Wright, 1989), лектотип типового вида рода Longaeviceras (L. longaevum) имеет следующие особенности: это фрагмокон макроконха с раковиной близкой к оксикону, но плохой сохранности и небольшого размера (диаметром около 50 мм, с 24 первичными и 65 вторичными ребрами). Эти же авторы предполагают, что различия между L. longaevum и другим английским видом L. placenta могут быть связаны с изменчивостью. Для вида L. placenta характерна сглаженная кадиконическая жилая камера (Callomon, Wright, 1989, p. 824), которая, как справедливо отмечает Ю.С. Репин, до сих пор не показана ни на одном экземпляре из Англии.
По мнению Ю.С. Репина (Репин и др., 2006), для диагностики родов Longoceras и Longaeviceras исследование их кадиконовых стадий не представляется необходимым, а основные родовые признаки проявляются при диаметре от 30 до 70 мм (Репин и др., 2006). Определенные Ю.С. Репиным особенности формы раковины и скульптуры, служащие для отличия обсуждаемых таксонов, приведены в табл. 1.
Таблица I . Здесь и в табл. II–VI все изображенные экземпляры собраны на восточном берегу Анабарской губы, обнажение 109. Коллекция хранится в Центре геологических коллекций “ГЕОХРОН” ИНГГ СО РАН под номером 2060. Образцы, кроме особо отмеченных, происходят из пачки 46–47 (или ее осыпи), зона Longaeviceras keyserlingi. Все экземпляры изображены в натуральную величину. Длина масштабной линейки 1 см. 1–2 – Longaeviceras nikitini (Sokolov, 1912): 1а–1в – экз. 2060/179 (109/31), 1а, 1б –внешний оборот, 1а – вид сбоку, 1б – вид с вентральной стороны, 1в – вид со стороны устья, по С.В. Мелединой образец происходит из пачки 31 (47б в Меледина, 1994); 1г–1е – экз. № 2060/152, без внешнего оборота, 1г – вид сбоку, 1д – поперечное сечение, 1е – вид с вентральной стороны; 11.6 м от подошвы, осыпь; 2 – экз. 2060/151, вид сбоку, 11.6 м от подошвы; 3 – Cardioceras (Maltoniceras) maltonense (Yong et Bird, 1822), экз. 2060/170: 3а – вид сбоку, 3б – вид со стороны устья, 3в – вид с вентральной стороны; пачка 48, 2 м от подошвы, средний оксфорд, зона Cardioceras densiplicatum.
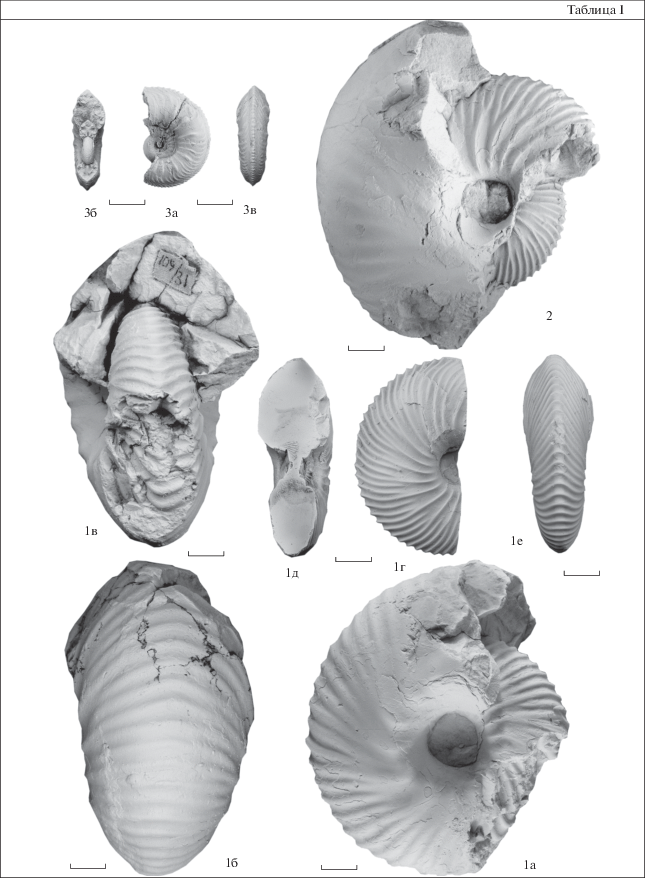
Таблица 1.
Признаки родов Longaeviceras и Longoceras по Ю.С. Репину (Репин и др., 2006)
| Стадия | Longaeviceras | Longoceras |
|---|---|---|
| 30–70 мм | Приостренная вентральная сторона, без килеватости, ребра от слабо изогнутых до серповидных, точка ветвления и вставления ребер находится в пределах от середины боков до их верхней привентральной части. | Приостренная, слабокилевидная вентральная сторона, резко дугообразная форма ребер, точка ветвления смещается в верхнюю четверть боков. |
| 60–70 мм | Слабо дуговидные и серповидные ребра с устойчивой точкой ветвления на середине боков. | Килеватость исчезает, вентральная сторона округляется. Высокая привентральная точка ветвления ребер. |
Отнесение английских и печорских лонгаевицерасов к разным таксономическим группам не лишено логики. Отличительные признаки Longoceras заметно проявляются на фоне малочисленности английских форм L. placenta и L. longaevum и отсутствия достоверных данных о морфологии внешних оборотов этих видов.
Стратиграфическое распространение лонгаевицерасов ограничивается главным образом нижней зоной верхнего келловея Athleta (арктический аналог – Keyserlingi). Указания на находки лонгаевицерасов в терминальной зоне келловея Lamberti связано со спорной родовой трактовкой таких видов, как “L.” novosemelicum, “L.” holtedahli, “L.” filarum Meledina. В.В. Митта изобразил экземпляр аммонита под названием Longaeviceras sp. nov. из зоны Lamberti Ульяновского Поволжья (урочище Тархановская Пристань; Митта и др., 2014, табл. III, фиг. 1, 2), но, по мнению авторов настоящей работы, этот образец не относится к роду Longaeviceras.
М. Ховарт (Howarth, 2017) в род Longaeviceras включает макро- и микроконхи. К последним, в качестве синонимов Longaeviceras, М. Ховарт отнес роды Longoceras Repin, Platylongoceras Repin, Percacosticeras Kiselev.
Районы распространения аммонитов Longaeviceras включают территории севера Сибири, севера бассейна р. Печора, Русской платформы, Северо-Западной Европы (Англия), Гренландии (Земля Милна), ? Дальнего Востока и Северо-Востока России, Аляски. Следует отметить, что экземпляры из Гренландии в литературе не изображены, однако имеются в коллекциях Дж. Калломона и Т. Биркелунд (Callomon, Birkelund, 1980), а аляскинские экземпляры относятся к видам “L.” pomeroyense и “L.” stenolobum (Imlay, 1953, 1976, а также синонимика в Callomon, 1984), родовая принадлежность которых дискуссионна.
Предковым для Longaeviceras нами рассматривается род Protolongaeviceras Knyazev, Meledina, Alifirov (Князев и др., 2019).
Географическое распространение. Север Сибири, Северо-Западная Европа, Русская платформа, Новая Земля, Земля Франца-Иосифа, Восточная Гренландия, архипелаг Шпицберген, ? Норвежское море.
Стратиграфическое распространение. Верхний келловей, зона Athleta (частично) Западной Европы и Центральной России, зона Keyserlingi севера бассейна р. Печора, Западной и Восточной Сибири, Земли Франца-Иосифа, Шпицбергена и Восточной Гренландии.
Longaeviceras nikitini (Sokolov, 1912)
Табл. I , фиг. 1, 2; табл. II, фиг. 1, 2; табл. III, фиг. 1
Cadoceras (Quenstedtoceras) keyserlingi: Соколов, 1912, с. 25, табл. II, фиг. 2; Меледина, 1977, табл. 32 , фиг. 2.
Cadoceras nikitini: Соколов, 1912, с. 24, табл. I , фиг. 1–3 ; Howarth, 2017, p. 72, figs. 56c–56e .
Longaeviceras stenolobum: Меледина, 1977, с. 140, табл. 26 , фиг. 3 а, 3б.
Longaeviceras aff. stenolobum: Меледина, 1977, с. 143, табл. 40 , фиг. 1, табл. 41 , фиг. 1.
non Longaeviceras nikitini: Callomon, 1985, text-fig. 8 (iv), figs. P , p.
Longoceras nikitini: Ершова, 1983, табл. 3 , фиг. 1–4 ; Репин, 2002, табл. 28 , фиг. 2, 7 , табл. 27 , фиг. 4 .
Лектотип: Cadoceras nikitini Sokolov, 1912, изображен в работе Д.Н. Соколова (1912, табл. 1, фиг. 1–3 ); Россия, р. Вишера, верхний келловей, зона Nikitini; назначен в настоящей работе.
Таблица II. 1–2 – Longaeviceras nikitini (Sokolov, 1912): 1 – экз. 2060/151, 1а – вид со стороны устья, 1б – вид с вентральной стороны, 1в–1д – без внешнего оборота, 1в – вид сбоку, 1г – вид с вентральной стороны, 1д – поперечное сечение; 11.6 м от подошвы; 2 – экз. № 2060/150, 2а – вид сбоку, 2б – вид со стороны устья, 2в – вид с вентральной стороны; 11.6 м от подошвы; 3 – Pseudolongaeviceras densicostatum sp. nov., экз. № 2060/166, вид сбоку; пачка 46–47, 11.6 м от подошвы; 4 – Platylongoceras novosemelicum (Bodylevsky, 1960), экз. № 2060/171, 4а – вид сбоку, 4б – поперечное сечение, 4в – вид с вентральной стороны; пачка 46–47, 13.5 м от подошвы.

Таблица III. 1 – Longaeviceras nikitini (Sokolov, 1912), экз. № 2060/172, поперечный скол оборотов раковины, осыпь; 2, 4 – Pseudolongaeviceras densicostatum sp. nov.: 2 – экз. № 2060/158, 2а – вид сбоку, 2б – вид со стороны устья, 2в – вид с вентральной стороны, 13.5 м от подошвы; 4 – экз. 2060/157, 4а – вид сбоку, 4б – вид со стороны устья, 4в – вид с вентральной стороны, 11.6 м от подошвы; 3 – Longaeviceras bodylevskyi (Meledina, 1977), экз. 489-77 (голотип, переизображен из (Меледина, 1977)), 3а – вид сбоку, 3б – вид со стороны устья, 3в – вид с вентральной стороны, 19.3 м от подошвы (= уровень 24е по Меледина, 1977).

Описание. Макроконхи диаметром до 100 мм и более. Жилая камера начинается обычно при диаметре около 80 мм, хотя есть и отклонения как в большую (при Д около 115–120 мм), так и в меньшую сторону (при Д около 60 мм). Эта часть раковины имеет форму кадикона и у наших экземпляров, как правило, сильно деформирована или вовсе не сохраняется. Ее длина составляет не менее полуоборота. Раковина на средних оборотах (Д = 50–70 мм) уплощенная или умеренно вздутая (Ш/Д = 35–44%), с закругленной вентральной стороной. Форма сечения на этой стадии – вытянутый в высоту овал (Ш/В около 0.83). Начиная с Д = 55–60 мм раковина быстро увеличивается в ширину, приобретая пахиконовую или кадиконовую форму. Умбиликус умеренно узкий (У/Д = 15–23%), постепенно расширяющийся по мере роста раковины. Скульптура до Д = 50 мм не наблюдалась. На фрагмоконе до начала формирования кадиконовых оборотов скульптура представлена двойными ребрами, между которыми наблюдается одно вставное ребро. Ветвление происходит на середине боков или чуть выше. Одиночные ребра редки. Чередование двойных и вставных ребер иногда нарушается. На этой стадии проявляется повторное ветвление ребер в привентральной части, здесь же могут появляться вставные ребра. РО превышает 2 и может достигать 3. При Д около 70 мм РО достигает 3.5. По мере роста раковины расстояние между первичными ребрами возрастает. Сами ребра становятся более толстыми. На умбиликальном перегибе они могут трансформироваться в буллы, вентральный изгиб вперед ослабевает. Жилая камера гладкая.
Измерения и отношения:
| № экземпляра | Д | Ш | В | У | Ш/Д | В/Д | У/Д | Ш/В | РВ/РУ | РО |
| 2060/150 | 57.9 | 19.2 | 30.1 | 7.8 | 33.2 | 52.0 | 13.5 | 63.8 | 60/24 | 2.5 |
| 2060/151 | 92.1 | 49.5 | 42.6 | 18.5 | 53.7 | 46.3 | 20.1 | 116.2 | – | – |
| 2060/151 | 59 | 24.3 | 28.1 | 11 | 41.2 | 47.6 | 18.6 | 86.5 | 60/21 | 2.8 |
| 2060/152 | 89.5 | 47.4 | 37.7 | 17.2 | 53.0 | 42.1 | 19.2 | 125.7 | – | – |
| 2060/152 | 56 | 20.9 | 26.4 | 9.8 | 37.3 | 47.1 | 17.5 | 79.2 | 32/12 | 2.6 |
| 2060/179 | 87.9 | 55.5 | 47.6 | 15.5 | 63.1 | 54.2 | 17.6 | 116.5 | – | – |
| 2060/179 | 50.4 | 21.3 | 25.2 | 10.1 | 42.2 | 50 | 20 | 84.5 | 30/13 | 2.3 |
| 2060/153 | 85.1 | 48.7 | 33.2 | 19.5 | 57.2 | 39.0 | 22.9 | 146.7 | – | – |
| 489-89 | 50.3 | 22.2 | 24 | 9 | 44.1 | 47.7 | 17.9 | 92.5 | 49/21 | 2.3 |
Сравнение. Внутренние обороты вида L. nikitini (Sok.) идентичны таковым у L. keyserlingi (Sok.), как указал сам автор вида, до диаметра 60 мм. Поскольку лектотип L. keyserlingi является фрагмоконом, можно с большой долей вероятности предположить, что этот экземпляр Д.Н. Соколова представляет средние обороты L. nikitini. Сравнение с видами L. placenta (Leck.) и L. longaevum (Leck.) затруднено ввиду малого количества данных об этих таксонах в литературе. Высокое реберное отношение (до 3.5), характерное для L. placenta и L. longaevum на оборотах, не достигших кадиконовых стадий (Kiselev, Rogov, 2018, pl. 7, figs. 1, 2, pl. 9, fig. 6 ), наблюдается и у некоторых экземпляров описываемого вида.
Замечание. В работе С.В. Мелединой (1994) крупные аммониты с маркировкой 109/31 (= 2060/179) (т.е. из самых верхних конкреционных горизонтов аргиллитовой толщи обн. 109, пачка 47б) не изображены, но определялись как E. subordinarium. Нами эти экземпляры отнесены к описываемому виду (табл. I , фиг. 1а–1в). В любом случае они явно отличаются онтогенезом формы раковины и скульптуры от типичных представителей Eboraciceras, изображенных, например, в работе Дж. Калломона (Callomon, 1985).
Материал. 8 неполных ядер (сборы В.Г. Князева и С.В. Мелединой, А.С. Алифирова), обн. 109, пачка 46.
Распространение. Земля Франца-Иосифа, архипелаг Шпицберген, бассейн р. Печора, ? Восточная Гренландия, север Восточной Сибири, верхний келловей, зона Keyserlingi.
Longaeviceras bodylevskyi (Meledina, 1977)
Табл. III, фиг. 3 , табл. IV, фиг. 1
Longaeviceras bodylevskyi: Князев и др., 1973, с. 655; Меледина, 1977, с. 145, табл. 25 , фиг. 3 , табл. 30 , фиг. 2 (голотип).
non Longaeviceras bodylevskyi: Меледина, 1977, табл. 44 , фиг. 1, табл. 45 , фиг. 2.
Longaeviceras keyserlingi (pars): Меледина, 1977, с. 136, табл. 32 , фиг. 3 , табл. 34 , фиг. 3 , табл. 35 , фиг. 2.
Голотип: экз. № 489-77, коллекция С.В. Мелединой. Хранится в Центральном Сибирском геологическом музее г. Новосибирска; восточный берег Анабарской губы, пачка 46–47, уровень 109/24е; верхний келловей, зона Keyserlingi.
Таблица IV. 1 – Longaeviceras bodylevskyi (Meledina, 1977), экз. № 2060/154, 1а – вид сбоку, 1б – вид со стороны устья, 1в – вид с вентральной стороны, 13.5 м от подошвы; 2, 3 – Pseudolongaeviceras densicostatum sp. nov. (Repin, 2002): 2 – экз. № 2060/176, вид сбоку, 19.3 м от подошвы; 3 – экз. № 2060/177, вид сбоку, 16–18 м от подошвы; 4–6 – Platylongoceras holtedahli (Salfeld et Frebold, 1924): 4 – экз. № 2060/175, 5 – экз. № 2060/174, 6 – экз. № 2060/173; все вид сбоку; 19.3 м от подошвы.

Описание. Макроконх с фрагмоконом, достигающим диаметра 82 мм. Конечная жилая камера начинается при Д около 75 мм, однако у самых крупных экземпляров вида в изученной коллекции она неизвестна, поэтому верхний предел конечных размеров этого вида пока не установлен. Внутренние обороты с округлым сечением (Ш/В = 1.1) и умеренно широким умбиликусом (У/Д = 37%). При Д = 20 мм раковина становится оксиконической, c превышением высоты над шириной в среднем в 1.5 раза. На фрагмоконе (до Д = 70 мм) происходит постепенное увеличение ширины оборотов. Сохраняется килеватость вентральной стороны (сужение от боков к вершине вентральной стороны). Некоторые экземпляры (например, экз. № 489-77) на этой стадии роста имеют параллельные боковые стороны, хотя и узкие, но без килеватости. Умбиликус узкий, слабо расширяющийся по мере роста раковины. Умбиликальные стенки с плавным умбиликальным перегибом. Истинная форма жилой камеры неизвестна. Деформированные фрагменты позволяют предположить, что она была ближе к платикону. Видовые особенности скульптуры проявляются на последнем обороте или полуобороте перед жилой камерой. Ребра становятся примерно в 1.5 раза более редкими и утолщенными. Точка ветвления часто наблюдается выше середины боковой стороны, еще выше появляются вставные ребра. В привентральной части раковины может происходить повторное ветвление ребер. Отношение количества первичных и вторичных ребер на средних оборотах (Д = 40–70 мм) в среднем составляет около 2.5. РО повышается с увеличением диаметра раковины от 2.1 до 3.4. При этом первичные ребра становятся более редкими, слегка приподнятыми вблизи умбиликального перегиба и не заходят на умбиликальную стенку раковины.
Измерения и отношения:
| № экземпляра | Д | Ш | В | У | Ш/Д | В/Д | У/Д | Ш/В | РВ/РУ | РО |
| 2060/154 | 75.7 | 29 | 35.5 | 11.8 | 38.3 | 46.9 | 15.6 | 81.7 | 72/21 | 3.4 |
| 2060/155 | 63.7 | 23 | 31.2 | 10.2 | 36.1 | 49.0 | 16.0 | 73.7 | 56/22 | 2.5 |
| 2060/156 | 85.7 | 42.9 | 43.8 | 11.9 | 50.1 | 51.1 | 13.9 | 97.9 | – | – |
| 2060/156 | 71.8 | 28.2 | 33.4 | 12.4 | 39.3 | 46.5 | 17.3 | 84.4 | 32/9 | 3.5 |
| 2060/156 | 64 | 24.8 | 30 | 10.5 | 38.8 | 46.9 | 16.4 | 82.7 | – | – |
| 489-77 | 79.4 | 23.4 | 36.8 | 13.3 | 29.5 | 46.3 | 16.8 | 63.6 | – | – |
Сравнение. От близкого вида L. nikitini (Repin) данный вид отличается более редкой и грубой ребристостью на последнем обороте/полуобороте фрагмокона перед жилой камерой. Вероятно, эти виды различаются формой поперечного сечения конечной жилой камеры, которая у вида L. bodylevskyi неизвестна.
Материал. Сборы В.Г. Князева: 1 экземпляр с деформированной жилой камерой и два неполных крупных ядра (2060/154, 2060/155). Сборы С.В. Мелединой: экз. № 489-77 – неполное ядро, обн. 109, пачка 46–47.
Распространение. Север Сибири, верхний келловей, зона Keyserlingi.
Род Pseudolongaeviceras gen. nov.
Longaeviceras: Меледина, 1977, с. 134 (частично).
Longoceras: Репин, 2002 с. (частично); Репин, 2008, с. 8.
Этимология. Название рода происходит от морфологически и стратиграфически близкого рода Longaeviceras Buckman и лат. pseudo – ложный, фальшивый.
Типовой вид. Pseudolongaeviceras densicostatum sp. nov., экз. № 2060/157, Центр геологических коллекций “ГЕОХРОН”, ИНГГ СО РАН; Анабарский залив (море Лаптевых), западный берег, обн. 109, пачка 46–47, зона Keyserlingi; изображен здесь, табл. III, фиг. 4 .
Диагноз. Конечные размеры фрагмокона у особей данного рода достигают диаметра 60–80 мм. Форма раковины оксиконическая или платиконическая на стадии конечной жилой камеры. Умбиликус узкий, очень слабо расширяется в онтогенезе. Для рода характерны два типа скульптуры на раковине. Первый тип: тонкая ребристость с сильным дугообразным выгибом назад на боках и вперед на вентральной стороне с одинаковой рельефностью по всей длине ребра. Второй тип: спрямленная более грубая ребристость с наклоном вперед на боках и слабым изгибом в сторону устья на вентральной стороне. Вставные ребра достигают середины боковых сторон. Характерно привентральное ветвление. Реберное отношение, как правило, более 2 и может достигать 3. Конечная жилая камера несет модифицированную скульптуру вплоть до полного сглаживания в конце нее.
Сравнение. Внутренние обороты по всем признакам близки к Longaeviceras Buckman emend. Arkell и Platylongoceras Repin. От Longaeviceras новый род отличается отсутствием кадиконической стадии раковины и меньшими конечными размерами, а от Platylongoceras – наличием ребристости первого типа (см. диагноз) и более узким умбиликусом последних оборотов в более крупных конечных размерах.
Видовой состав. Pseudolongaeviceras raricostatum (Repin), Pseudolongaeviceras densicostatum sp. nov.
Замечание. К настоящему времени в коллекции авторов крупные представители рода либо не имеют жилой камеры, либо она сильно деформирована. Вследствие этого для данного рода не установлен максимальный размер особей с конечной жилой камерой.
Географическое распространение. Север Сибири, бассейн р. Печора.
Стратиграфическое распространение. Верхний келловей, зона Keyserlingi севера бассейна р. Печора; зона Keyserlingi севера Восточной Сибири.
Pseudolongaeviceras densicostatum sp. nov.
Табл. II, фиг. 3 ; табл. III, фиг. 2, 4 ; табл. IV, фиг. 2, 3 ; табл. V, фиг. 1, 2; табл. VI, фиг. 3
Этимология. Название вида происходит от лат. densus – частый и costa – ребро.
Голотип: Pseudolongaeviceras densicostatum sp. nov., табл. III, фиг. 4 , экз. № 2060/157, Центр геологических коллекций “ГЕОХРОН”, ИНГГ СО РАН; Анабарский залив (море Лаптевых), западный берег, обн. 109, пачка 46–47, зона Keyserlingi.
Таблица V. 1, 2 – Pseudolongaeviceras densicostatum sp. nov. (Repin, 2002): 1 – экз. № 2060/159, 1а – вид сбоку без части внешнего оборота, 1в – вид сбоку с жилой камерой, 1г – вид с вентральной стороны, часть внутреннего оборота; пачка 46–47, 13.5 м от подошвы; 2 – экз. № 2060/160, 2а – вид сбоку, 2б – вид со стороны устья; пачка 46–47, 13.5 м от подошвы; 3 – Pseudocadoceras laminatum (Buckman, 1918), экз. № 2060/170, 3а – вид сбоку, 3б – вид со стороны устья, 3в – вид с вентральной стороны; пачка 46–47, 11.6 м от подошвы; 4–6 – ? Platylongoceras sp. juv.: 4 – экз. № 2060/178, 5 – экз. № 2060/177, 6 – экз. № 2060/176; все вид сбоку; 13.5 м от подошвы.
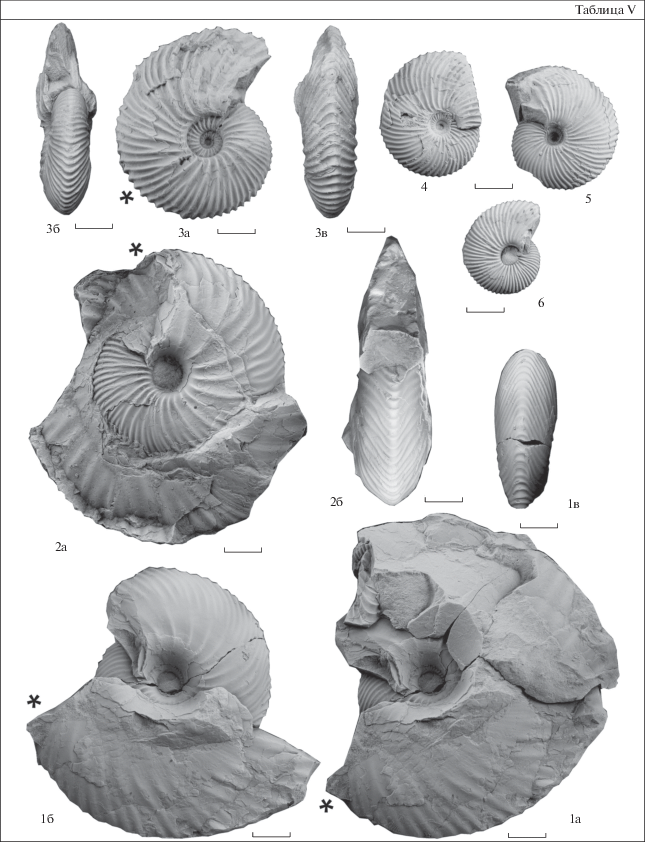
Таблица VI. 1 – ? Dolganites sp., экз. № 2060/169, 1а – вид сбоку, 1б – вид с вентральной стороны, 1в – вид с вентральной стороны последнего полуоборота; осыпь; 2 – Platylongoceras holtedahli (Salfeld et Frebold, 1924), экз. № 489-68, 2а – вид сбоку, 2б – вид с вентральной стороны; 19.3 м от подошвы; 3 – Pseudolongaeviceras densicostatum sp. nov. (Repin, 2002), экз. № 489-53, 3а – вид сбоку, 3б – вид с вентральной стороны, 3в – вид со стороны устья; 19.3 м от подошвы; 4 – Rondiceras milaschevici (Nikitin, 1881), экз. № 2060/168, 4а – вид сбоку, 4б – вид сбоку без внешнего полуоборота, 4в – поперечное сечение, 4г – вид с вентральной стороны; пачка 45, подошва слоя.

Описание. Макроконхи достигают 86 мм в диаметре. Конечная жилая камера начинается при диаметре около 55 мм. При диаметре 25–30 мм раковина уплощенная, со слабо приостренной вентральной стороной. Умбо узкий на всех оборотах. По мере роста толщина раковины медленно нарастает, но килеватость сохраняется вплоть до начала жилой камеры. Жилая камера занимает около половины оборота и сохраняет форму, близкую к оксикону. Скульптура характеризуется одиночными, двойными и вставными ребрами. Точка ветвления и появления вставных ребер повышается по мере роста раковины. В целом для ребер характерен наклон вперед. Дугообразный изгиб ребер на боках и их изгиб вперед усиливаются до последнего полуоборота, но проявляются не у каждого ребра. На жилой камере скульптура сохраняется, хотя становится более грубой. Имеется предустьевой пережим.
Измерения и отношения:
| № экземпляра | Д | Ш | В | У | Ш/Д | В/Д | У/Д | Ш/В | РВ/РУ | РО |
| 2060/157 | 71.1 | 23 | 30.9 | 11.4 | 32.3 | 43.5 | 16.0 | 74.4 | 33/14 | 2.3 |
| 2060/158 | 45.8 | 14.4 | 22.2 | 7 | 31.4 | 48.5 | 15.3 | 64.9 | – | – |
| 2060/159 | 82.5 | – | – | – | – | – | – | – | – | – |
| 2060/160 | 86.5 | – | – | – | – | – | – | – | – | – |
| 2060/160 | 61 | 21.9 | 28.6 | 9.7 | 35.9 | 46.9 | 15.9 | 76.6 | /19 | – |
| 2060/161 | 76.7 | – | – | – | – | – | – | – | – | – |
| 489-53 | 67.8 | 16.8 | 31.5 | 12.2 | 24.8 | 46.5 | 18.0 | 53.3 | 69/29 | 2.3 |
| 2060/162 | 53.4 | 14.3 | 25.8 | 8.7 | 26.8 | 48.3 | 16.3 | 55.4 | 36/14 | 2.57 |
| 2060/163 | 55.7 | – | 27.8 | 9.5 | – | 49.9 | 17.1 | 1 | 55/26 | 2.1 |
Сравнение. От вида Pseudolongaeviceras raricostatum (Repin) отличается более узким умбиликусом и более частой и тонкой скульптурой на последнем обороте.
Материал. 9 экземпляров из сборов А.С. Алифирова, В.Г. Князева, С.В. Мелединой; восточный берег Анабарской губы, обн. 109, пачка 46–47.
Распространение. Верхний келловей, бассейн р. Печора, зона Keyserlingi, cевер Сибири, зона Keyserlingi.
Род Platylongoceras Repin, 2002
Platylongoceras holtedahli (Salfeld et Frebold, 1924)
Табл. IV, фиг. 4–6 ; табл. VI, фиг. 2
Quenstedticeras holtedahli: Salfeld, Frebold, 1924, pl. 1, figs. 3, 3a, 4 .
Longaeviceras holtedahli: Бодылевский, 1960, табл. X , фиг. 3 ; Callomon, Wright, 1989, p. 826.
Platylongoceras holtedahli: Репин и др., 2006, с. 107, табл. 33 , фиг. 1, 2; Репин, 2008, с. 8, табл. I , фиг. 3 .
Лектотип. Quenstedtoceras holtedahli Salfeld et Frebold, 1924 (pl. 1, figs. 3, 3a ), выделен в работе Дж. Калломона и Дж. Райта (Callomon, Wright, 1989); Новая Земля, из валунов.
Описание. Раковины диаметром до 55–60 мм. Конечная жилая камера занимает до ¾ оборота и начинается при диаметре фрагмокона от 36 до 50 мм. Раковины оксиконические до конца жилой камеры. Умбиликус узкий или умеренно узкий (У/Д = 13.5–26%), заметно расширяющийся на последнем обороте. Умбональные стенки низкие. Скульптура представлена одиночными, двойными и вставными ребрами без закономерного чередования, с преобладанием двойных ребер. На фрагмоконе ребра появляются на умбиликальной стенке, проявляясь более отчетливо на умбиликальном перегибе с уклоном в сторону от устья. Через боковые стороны ребра проходят по дуге. Их наклон вперед несколько усиливается на вентральной стороне. Высокое привентральное ветвление встречается редко. На жилой камере межреберные расстояния заметно увеличиваются, умбональная стенка сглаживается. Отношение первичных и вторичных ребер составляет, как правило, 2–2.5.
Сравнение. По форме раковины и некоторым особенностям скульптуры представители вида сходны с Platylongoceras novosemelicum (Bodylevsky), который в отличие P. holtedahli характеризуется существенно более рельефными и широко расставленными ребрами.
Измерения и отношения:
| № экземпляра | Д | Ш | В | У | Ш/Д | В/Д | У/Д | Ш/В | РВ/РУ | РО |
| 2060/164 | 60 | 11.2 | 28.8 | 12.9 | 18.7 | 48.0 | 21.5 | 38.9 | 28/14 | 2 |
| 2060/165 | 49.2 | 11.2 | 22.6 | 11.5 | 22.8 | 45.9 | 23.4 | 49.6 | 36/15 | 2.4 |
| 2060/166 | 58.7 | 17.6 | 27.8 | 10.2 | 30.0 | 47.4 | 17.4 | 63.3 | 63/27 | 2.3 |
| 489-68 | 54.7 | 16.2 | 23.2 | 14.2 | 29.6 | 42.4 | 26.0 | 69.8 | 25/14 | 1.8 |
| 2060/167 | 23.8 | 8 | 12.3 | 4.2 | 33.6 | 51.7 | 17.6 | 65.0 | 31/14 | – |
Замечания. Экземпляр описываемого вида из работы Ю.С. Репина (Репин и др., 2006, табл. 33 , фиг. 1) имеет более крупные размеры фрагмокона (Д около 70 мм), чем его сибирские представители. Вид P. holtedahli отличается от лектотипа L. longaevum (Leck.) большими значениями У/Д (на 6–10%) и меньшими значениями РО (у L. longaevum РО = 2.7). Внутренние обороты L. placenta (Leck.) при сопоставимых с описываемым видом диаметрах (Д = 40–50 мм) имеют более толстую (в 1.5–2 раза) раковину и грубую скульптуру. В коллекции С.В. Мелединой (№ 489) имеется экземпляр Longaeviceras cf. stenolobum (Sok.) (экз. № 489-68, табл. VI, фиг. 2), который нами отнесен к описываемому виду.
Материал. Всего 9 экземпляров: 4 из коллекции С.В. Мелединой (489-68, 109/30), 2 из сборов А.С. Алифирова, 3 из сборов В.Г. Князева. Все экземпляры имеют полностью или частично сохранившуюся жилую камеру. Также в коллекции имеются около 10 обломков ядер в основном из сборов В.Г. Князева; восточный берег Анабарской губы, пачка 46–47, верхний келловей, зона Keyserlingi.
Распространение. Верхний келловей, зона Keyserlingi cевера Русской платформы (бассейн р. Печора), зона Keyserlingi севера Сибири (восточный берег Анабарской губы); верхний келловей, зона Lamberti Новой Земли, ? Шотландии (о-в Скай, залив Штаффин).
ОБСУЖДЕНИЕ
Longaeviceras в среднем келловее
Дискуссионным является вопрос о присутствии рода Longaeviceras в среднем келловее. Дж. Калломон и Дж. Райт предположили, что вид L. stenolobum (Keyserling, 1846) из зоны Coronatum является переходным между типичными формами Longaeviceras и группой Cadoceras (Rondiceras) milaschevici (Callomon, Wright, 1989). Д.Н. Киселев установил в среднем келловее Русской платформы, кроме L. stenolobum (Keyserling, 1846 emend. Nikitin, 1881), еще два вида: L. praestenolobum Kisel. и L. alpha Kisel. Однако не все исследователи согласились с тем, что все эти виды принадлежат роду Longaeviceras (Репин и др., 2006; Митта, 2016; Алифиров и др., 2017).
Longaeviceras praestenolobum Kiselev et Meledina, 2004. Признаки внутренних и внешних оборотов, показанных Д.Н. Киселевым для этого вида, формально отвечают диагнозу рода Longaeviceras, данному В. Аркеллом (Arkell et al., 1957). Однако лонгаевицерасы верхнего келловея (L. nikitini и др.) отличаются от L. praestenolobum более высоким В/Ш (1.25 против 1.1) и килеватым поперечным сечением, а реберное отношение первых может превышать 3, как у L. placenta. Тем не менее у верхнекелловейских Longaeviceras наблюдается преемственность признаков скульптуры и формы раковины от описываемого вида. Поэтому помещение рассматриваемого вида в начало лонгаевицерасовой филолинии, как предложено Д.Н. Киселевым, в целом не лишено оснований (Киселев Меледина, 2004; Киселев, 2006). С другой стороны, Д.Н. Киселев показал, что потомком вида L. praestenolobum является вид L. stenolobum, родовая принадлежность которого дискуссионна (Митта, 2000; Репин и др., 2006). Среднекелловейский возраст L. praestenolobum обосновывается совместным нахождением с Kosmoceras obductum (Buckman) в разрезах на р. Сысола (с. Вотча, север Европейской России) и Михайловцемент (Рязанская область; Kiselev, Rogov, 2018).
Rondiceras stenolobum (Keyserling, 1846). Как правило, представление о виде stenolobum основывается на экземплярах С.Н. Никитина (1881, табл. 12 , фиг. 28–30 ), что ясно из работ Дж. Калломона (Callomon, Wright, 1989), Д.Н. Киселева (2006) и В.В. Митта (2000). После публикации Д.Н. Киселевым фотографии лектотипа вида (Киселев, 2006, табл. IV, фиг. 1–6 ) авторы согласились с его отнесением к роду Longaeviceras (Князев и др., 2015). Кроме того, при изучении онтогенезов представителей рода Rondiceras (виды R. tschefkini (d’Orbigny), R. milaschevici (Nikitin), R. nikolaevi (Bodylevsky)) выяснилось, что характерной особенностью рода является исчезновение скульптуры, начиная от умбиликального шва. При этом продолжительность скульптурированной стадии увеличивается в ряду видов R. milaschevici–R. nikolaevi. Данный признак на лектотипе L. stenolobum не наблюдается (возможно, из-за плохой сохранности последнего). С другой стороны, предложенный В.И. Бодылевским и опубликованный Д.Н. Киселевым лектотип имеет округлую вентральную сторону, ребра проходят через нее без характерного для Longaeviceras изгиба. То же самое наблюдается и у других экземпляров данного вида (Репин и др., 2006, табл. 26 , фиг. 1, 5 ; табл. 28 , фиг. 1).
В процессе исследования возник вопрос о сходстве видов R. nikolaevi (Bodylevsky) и R. stenolobum (Keyserling, 1846 emend. Nikitin, 1881). Различия этих видов В.И. Бодылевский (1960) выявил в степени расчлененности лопастных линий и местоположении третьей боковой лопасти. При этом он указал на сходство этих линий у молодых экземпляров сравниваемых видов, а также на сходство лопастной линии последнего оборота голотипа C. nikolaevi Bodyl. с таковой у Cadoceras stenolobum (Keyserling). Апелляция к особенностям строения лопастей у кардиоцератид на уровне видов, на наш взгляд, не убедительна. Тем более что зарисовка лопастной линии у вида C. nikolaevi в работе В.И. Бодылевского отсутствует. Остается одно отличие, также приведенное В.И. Бодылевским, – это отсутствие ребер на умбиликальных стенках и умбиликальном перегибе внутренних оборотов у R. nikolaevi. В.В. Митта (2000) указывает на то, что ребра у R. stenolobum могут опускаться до верхней трети умбиликальной стенки. По данным Ю.С. Репина (Репин и др., 2006), у Rondiceras stenolobum stenolobum ребра начинаются от умбиликального перегиба. Лектотип вида stenolobum (слепок умбиликального столбика) также имеет ребристые умбиликальные стенки (Keyserling, 1846, табл. 20 , фиг. 7 ). Из приведенных данных можно заключить, что обсуждаемый признак может быть сильно изменчивым. На ранних стадиях роста ребра есть и в умбиликусе у R. nikolaevi. Сходство признаков раковины у экземпляров С.Н. Никитина (1881, табл. 12 , фиг. 29 , р. Ока), лектотипа “L.” stenolobum (Киселев, 2006, табл. IV, фиг. 1–2) и голотипа R. nikolaevi (Бодылевский, 1960, табл. IX , фиг. 1–2) может быть дополнительным основанием для отнесения “L.” stenolobum к роду Rondiceras.
L. alpha Kiselev. Этот вид относится в отечественной литературе к разным родам. Ю.С. Репин рассматривает его в составе рода Rondiceras, а В.В. Митта (2016) считает предковым для рода Platychamoussetia Repin, в который помещены виды Funiferites allae Kiselev, F. patruus (Eichwald), F. funiferus (Phillips). Следует также обратить внимание на сходство формы раковины L. alpha с таковой у Stenocadoceras, отмеченное автором вида Д.Н. Киселевым. Достаточно долго сохраняющаяся ребристость у голотипа L. alpha (до Д = 80 мм; Киселев, 2001, табл. 10 , фиг. 1, 2) и характер ее исчезновения (последними исчезают вентральные ребра) позволяют рассматривать этот вид в качестве потомка R. stenolobum. Точка ветвления не имеет регулярного положения, что не характерно для типичных верхнекелловейских Longaeviceras.
Замечания к системе родов Longaeviceras и Longoceras (Репин, 2002; Репин и др., 2006)
Отдельно следует обсудить состав родов Longaeviceras и Longoceras, предложенный Ю.С. Репиным. По Ю.С. Репину (Репин, 2006, 2008) род Longaeviceras состоит из номинального подрода с одним типовым видом L. longaevum и подрода Soaniceras Meledina с видами L. (S.) placenta, L. (S.) angustatum, L. (S.) sazonovi Repin, L. (S.) cupressum (Sazonov), L. (S.) principale (Sazonov). Подрод Soaniceras первоначально был выделен С.В. Мелединой (1977) в составе рода Quenstedtoceras Hyatt. Д.Н. Киселев (1999) рассматривал его как подрод в составе роде Pseudocadoceras. М. Ховарт считал Soaniceras самостоятельным родом в подсемействе Cardioceratinae (Howarth, 2017). Синонимом Soaniceras, по мнению М. Ховарта, является род Anabariceras Stolyarova (Столярова, 2007).
Голотип типового вида Q. (Soaniceras) angustatum представляет собой фрагмокон (Д = 55 мм) с сильно затертым внешним оборотом (экз. № 489-111, о-в Б. Бегичев, р. Иннокентьевка). Отличительной особенностью данного экземпляра является высокое отношение вторичных и первичных ребер уже при диаметре 35 мм (КВ = 3.3), что является характерным признаком Quenstedtoceras s.l. (Quenstedtoceras, Eboraciceras и других кардиоцератин). Основная выборка вида Q. (S.) angustatum представлена микроконховыми формами диаметром менее 40 мм с жилыми камерами (Восточный Таймыр, р. Чернохребетная), что затрудняет ее сравнение как с типовым экземпляром вида Q. (S.) angustatum с о-ва Большой Бегичев, так и с L. placenta. Таким образом, мнение Ю.С. Репина о систематическом положении Soaniceras нами не принимается. Кроме того, голотип и типовая выборка рассматриваемого вида Q. (S.) angustatum не могут быть объединены под одним видовым и родовым названием. Голотип типового вида Soaniceras может оказаться представителем рода Anabariceras, как указал М. Ховарт (в таком случае последний является его младшим синонимом), или рода Dolganites. Для остальных аммонитов, описанных как Q. (S.) angustatum (из коллекции С.В. Мелединой), следует установить новое родовое название или отнести эту группу к какому-либо известному микроконховому роду кардиоцератид, такому как Pseudocadoceras.
В род Longoceras Ю.С. Репин поместил совершенно разные по своим признакам виды. C одной стороны, L. nikitini и L. keyserlingi (макроконхи), которые легли в основу диагноза рода Longaeviceras по В. Аркеллу (Arkell et al., 1957), и, с другой стороны, L. parvulum (Meled.) – микроконхи из Quenstedtoceras (Soaniceras). Выделение печорских и северосибирских видов в отдельный род кажется логичным лишь на фоне недостатка данных о морфологии типовых для Longaeviceras s.str. видов – L. longaevum и L. placenta. Виды “L.” bodylevskyi (Meled.) и “L”. raricostatum Repin, как показано выше, помещены здесь в новый род Pseudolongaeviceras.
Вид Longaeviceras filarum Meledina (Меледина, 1977, табл. 21 , фиг. 3 , табл. 22 , фиг. 2) описан главным образом с р. Иннокентьевка (о-в Б. Бегичев). Типовая серия включает в себя экземпляры с частично сохранившейся жилой камерой при диаметре 38–45 мм, а также фрагмоконы диаметром 46–51 мм (и менее), к которым относится и голотип вида (экз. № 489-90). Ю.С. Репин отнес этот вид к роду Rondiceras (Репин и др., 2006). Исследование голотипа и близких к нему по размеру фрагмоконов из коллекции С.В. Мелединой показало, что они имеют сходство с Longaeviceras по форме раковины (приостренное сечение) и скульптуре (дугообразные двойные и вставные ребра) до диаметра 30 мм. В дальнейшем вентральная сторона L. filarum округляется, изогнутость ребер ослабевает, при диаметре 45–50 мм намечается тенденция к сглаживанию, как у Rondiceras. По-видимому, часть экземпляров, описанных как L. filarum Meled., включая голотип, относится к виду R. nikolaevi (Bodylevsky).
Стратиграфические выводы
В зональной схеме верхнего келловея в бассейне р. Печора Ю.С. Репиным (2002, 2007) зона Longaeviceras keyserlingi заменена двумя зонами – Longoceras nikitini и Dolganites adzviensis. Последняя также охватывает часть терминальной зоны келловея Европейской шкалы – Quenstedtoceras lamberti. Эквивалентом печорской зоны D. adzviensis является зона Dolganites sibiricus (Репин и др., 2006).
Выделение зоны Sibiricus проблематично, поскольку экземпляры вида-индекса (в том числе и голотип) входят в состав комплекса разновозрастных аммонитов, собранного С.В. Мелединой на реках Иннокентьевка и Чернохребетная и ранее считавшегося показателем зоны Eboraciceras subordinarium. Пересмотр сборов С.В. Мелединой привел нас к заключению, что в слое 2, обн. 5 на р. Чернохребетная и в слое 2, обн. 3 на р. Иннокентьевка (Меледина, 1977, 1994), трактуемых ранее как терминальный келловей (зона Subordinarium), присутствуют среднекелловейские Rondiceras и нижнеоксфордские Scarburgiceras (или Vertumniceras).
В основании верхнего келловея Сибири традиционно выделялась зона Longaeviceras keyserlingi, которая целиком сопоставлялась с европейской зоной Peltoceras athleta. Представление об объеме зоны и таксономическом составе ее аммонитового комплекса для Сибири основывалось главным образом на данных по разрезу на восточном берегу Анабарской губы. Проведенный авторами анализ вертикального распространения аммонитов с указанием их точных привязок позволил сделать следующие выводы.
1. Пачка 46–47 верхнего келловея (зона Keyserlingi) на восточном берегу Анабарской губы залегает на конденсированных отложениях ранне-среднекелловейского возраста.
2. Как уже отмечалось выше, присутствие зоны E. subordinarium в разрезе келловея на восточном берегу Анабарской губы не подтвердилось в связи с переопределением нами аммонитов, относимых ранее к роду Eboraciceras (из коллекции С.В. Мелединой), в Longaeviceras nikitini. В.Г. Князевым в осыпи обн. 109 был найден аммонит ? Dolganites sp. (табл. VI, фиг. 1), который может свидетельствовать о возможном присутствии в данном разрезе аналогов зоны Lamberti.
В других районах Панбореальной надобласти, таких как Гренландия, Западная Европа, Арктическая Канада, лонгаевицерасы не являются основой для построения зональных шкал, отмечаются лишь фаунистические горизонты в зоне Athleta верхнего келловея, содержащие аммониты этого рода.
Географическое распространение Longaeviceras
Западная Европа. Хотя род Longaeviceras был установлен С. Бакмэном на материале из английских разрезов (Йоркшир), представительность европейских лонгаевицерасов весьма скудна. Единичные экземпляры приводятся в работах Дж. Калломона (Callomon, Wright, 1989) и К. Пейджа (Page, 1991). Присутствие Longaeviceras в разрезах келловея Франции установлено Ф. Курвилем (Courville et al., 1996; Courville, 2011). Однако экземпляр Longaeviceras sp., изображенный в (Courville, 2011, pl. 2, fig. 4 , Zone Lamberti), по-нашему мнению, относится к роду Eboraciceras. Аммониты, приведенные в (Courville et al., 1996, pl. 2, figs. 1–2), свидетельствуют о присутствии Longaeviceras в зоне Athleta этой части Европы.
Русская платформа и Печорская плита. В европейской части России присутствие Longaeviceras предполагалось В.И. Бодылевским (1960) на основании переопределенных им экземпляров из коллекции Н.Т. Сазонова (1957, табл. 11 , фиг. 3 , табл. 2 , фиг. 4 ) и Е.А. Троицкой (Камышева-Елпатьевская и др., 1956, табл. 19 , фиг. 47 ). В дальнейшем мнение Бодылевского не было поддержано, однако появились новые, хотя и дискуссионные, данные о присутствии Longaeviceras в среднем келловее данного региона (Киселев, Меледина, 2004; Киселев, 2006). Верхнекелловейские Longaeviceras keyserlingi упоминались из разреза на р. Сысола (с. Вотча; Киселев, 2006) и обнажения на р. Сухая Песчанка (Киселев, Меледина, 2004), а несколько экземпляров L. placenta недавно были изображены из келловея окрестностей г. Михайлова (Рязанская область; Kiselev, Rogov, 2018, pl. 9, fig. 6 , pl. 7, figs. 1, 2). Из района Печорской плиты известны многочисленные находки Longaeviceras и близких к нему родов. Именно данные по аммонитам из бассейна р. Печора послужили основанием для уточнения диагноза рода (Arkell et al., 1957) и выделения ряда новых родов и видов (Репин и др., 2006; Репин, 2008).
Западная Сибирь. Верхнекелловейские аммониты в Западной Сибири известны по находкам из керна скважин и являются наиболее многочисленными по сравнению с находками аммонитов из других подъярусов келловея (Атлас…, 1990). Учитывая вышеизложенные данные по систематике верхнекелловейских Longaeviceras s.l., следует с особой осторожностью подходить к определению этих аммонитов в керне скважин из-за существенной потери признаков при литификации осадка. Западносибирские лонгаевицерасы представлены отпечатками ядер диаметром до 50–60 мм (Атлас…, 1990, табл. 44 , фиг. 11, 12 ; Меледина, 1998; Алифиров, Меледина, 2010), что серьезно ограничивает достоверность родовой идентификации. Многие определения Longaeviceras из керна скважин Западной Сибири следует пересмотреть на предмет их отнесения к более молодым родам, таким как Quenstedtoceras или Scarburgiceras (Атлас…, 1990, табл. 44 , фиг. 13 , табл. 45 , фиг. 2).
Восточная Гренландия. Верхнекелловейские отложения в Восточной Гренландии развиты не повсеместно. Редкие находки верхнекелловейских Kosmoceras и Longaeviceras известны с Земли Милна и Земли Джеймсона (Callomon, Birkelund, 1980; Engkilde, Surlyk, 2003; Larsen et al., 2003; Vosgerau et al., 2004) из пачки Kosmocerasdal Member формации Kap Leslie (фауна M2) и из формации Olympen (фауна J2) соответственно. Лонгаевицерасы обнаружены в выветрелых известковистых септариевых конкрециях (Longaeviceras nikitini (Sok.), экз. № 3816, 3817, коллекция Дж. Калломона, MGUH, Геологический музей Копенгагена, Дания, определение авторов).
Аляска. По данным Р. Имлея, зона Athleta может быть представлена верхними 150–200 футами формации Чинитна, однако фоссилии, доказывающие это предположение, здесь отсутствуют (Imlay, 1975). Тем не менее в указанной работе (Imlay, 1975, fig. 7 ) показано распространение Cadoceras (Stenocadoceras) в зоне Athleta верхнего келловея. Последующая ревизия аляскинской и североканадской аммонитовой фауны, проведенная Дж. Калломоном (Callomon, 1984), позволила ему выделить ряд фаунистических уровней, в том числе и в верхнем келловее: B10 – Longaeviceras pomeroyense, D6 – L. cf. stenolobum. По нашему мнению, экземпляр Longaeviceras pomeroyense (Imlay, 1953, pl. 43, fig. 1–3 ) относится к среднекелловейскому роду Protolongaeviceras (Князев и др., 2020). Среди экземпляров, которые Дж. Калломон определил как L. cf. stenolobum (см. синонимику в Callomon, 1984, p. 167; Imlay, 1976, pl. 3, figs. 1–4 , 7, 8, 11–16 ), мы не исключаем присутствия представителей Longaeviceras в фаунистическом горизонте D6.
ЗАКЛЮЧЕНИЕ
Новый ископаемый материал из разрезов на севере Сибири и переизучение прежних сборов позволили выделить среди аммонитов, традиционно определяемых как Longaeviceras, несколько родов: Longaeviceras s.s., Pseudolongaeviceras gen. nov., Platylongoceras Repin, Percacosticeras Kiselev. По мнению авторов, разделение Longaeviceras s.l. на Longaeviceras s.s. и Longoceras, а также включение в Longaeviceras s.s. подрода Quenstedtoceras (Soaniceras) не имеет весомых палеонтологических оснований.
Новый род Pseudolongaeviceras gen. nov. стратиграфически, вероятнее всего, распространен лишь в зоне Nikitini. Более точные данные по этому вопросу могут быть получены при изучении непрерывных разрезов верхнего келловея.
Аммониты верхнего келловея на восточном берегу Анабарской губы представлены следующими таксонами: Longaeviceras nikitini (Sok.), Longaeviceras bodylevskyi (Meled.), Pseudolongaeviceras densicostatum sp. nov., Platylongoceras holtedahli (Salfeld), Pseudocadocers laminatum (Buckman), Percacosticeras aff. adzvicum Repin, ? Dolganites sp. Присутствие аммонитов рода Eboraciceras в данном разрезе не подтвердилось. Последнее говорит об отсутствии зоны Eboraciceras subordinarium в толще аргиллитов верхнего келловея (пачка 46–47) обнажения 109. Данный вывод ставит вопрос об аналогах европейской зоны Lamberti в келловее Сибири, однако это не является предметом исследования настоящей работы.
Возраст оксфордской части разреза (пачка 48) уточнен благодаря находке Cardioceras (Maltoniceras) maltonense Spath, сделанной В.Г. Князевым (табл. I , фиг. 3 ). Ранее эта пачка относилась к верхнему оксфорду по обнаруженному в ней Amoeboceras kostromense (Nikitin), однако проверить это определение невозможно, так как экземпляр, вероятнее всего, потерян, место его хранения неизвестно.
Ставятся под сомнение данные о присутствии рода Longaeviceras в среднем келловее. Среднекелловейские виды, относимые к Longaeviceras (stenolobum, alpha), не имеют признаков классических лонгаевицерас: приострение вентральной стороны раковины, изгиб ребер на вентральной стороне, высокое повторное ветвление ребер.
Географическое расселение аммонитов Longaeviceras s.l. имеет следующие особенности. Эти аммониты были наиболее характерны для Северо-Сибирской, Печорской и Восточно-Гренландской палеобиогеографических провинций, в которых они многочисленны и разнообразны. В суббореальной палеобиогеографической области эти аммониты более редки.
Благодарности. Авторы выражают свою глубокую признательность М.А. Рогову (ГИН РАН, Москва) и Д.Н. Киселеву (ЯГПУ, Ярославль) за тщательное рецензирование рукописи, которое привело к ее существенному улучшению.
Источники финансирования. Работа выполнена при финансовой поддержке РФФИ (проект № 19-05-00130), а также является вкладом в проект ФНИ № 0331-2019-0004 “Палеонтология, стратиграфия, биогеография бореальных и смежных с ними палеобассейнов и комплексное обоснование усовершенствования региональных стратиграфических схем мезозоя и кайнозоя Сибири”.
Список литературы
Алифиров А.С., Меледина С.В. Келловейские аммониты Западной Сибири: хорология и хронология // Новости палеонтологии и стратиграфии. Приложение к журн. “Геология и геофизика”. 2010. Т. 51. Вып. 14. С. 61–84.
Алифиров А.С., Князев В.Г., Меледина С.В. К систематике и филогении позднебатских–среднекелловейских Cardioceratidae (Ammonoidea) // Интегративная палеонтология: перспективы развития для геологических целей. Материалы LXIII сессии Палеонт. общества при РАН (3–7 апреля, 2017 г., Санкт-Петербург). Санкт-Петербург, 2017. С. 12–15.
Атлас моллюсков и фораминифер морских отложений верхней юры и неокома Западно-Сибирской нефтегазоносной области. М.: Недра, 1990. Т. 1. 285 с.
Басов В.А., Великжанина Л.С., Джиноридзе Н.М, Меледина С.В., Нальняева Т.И. Новые данные по стратиграфии юры Лено-Анабарского района // Проблемы палеонтологического обоснования детальной стратиграфии мезозоя Сибири и Дальнего Востока. Л.: Наука, 1967. С. 74–94.
Бодылевский В.И. Келловейские аммониты Северной Сибири // Записки ЛГУ. 1960. Т. 37. Вып. 2. С. 49–82.
Галабала Р.О. К стратиграфии юрских отложений Лено-Анабарского прогиба // Региональная геология и полезные ископаемые Якутии. Сборник научных трудов. Якутск: Изд-во Якутского госуниверситета, 1991. С. 155–158.
Ершова Е.С. Объяснительная записка к биостратиграфической схеме юрских и нижнемеловых отложений архипелага Шпицберген. Л.: ПГО “Севморгеология”, 1983. 88 с.
Захаров В.А., Шурыгин Б.Н. Биогеография, фации и стратиграфия средней юры Советской Арктики (по двустворчатым моллюскам). Новосибирск: Наука, 1978. 206 с.
Камышева-Елпатьевская В.Г., Николаева В.П., Троицкая Е.А. Определитель юрских аммонитов саратовского Поволжья. М.: Госгеолтехиздат, 1956. 61 с.
Каплан М.Е., Меледина С.В., Шурыгин Б.Н. Келловейские моря Северной Сибири (условия осадконакопления и существования фауны). Новосибирск: Наука, 1979. 79 с.
Киселев Д.Н. Морфогенез и систематика рода Pseudocadoceras (Ammonoidea) // Палеонтол. журн. 1996. № 3. С. 15–27.
Киселев Д.Н. Зональные и подзональные комплексы среднего келловея Центральной России // Проблемы стратиграфии и палеонтологии мезозоя. Научные чтения, посвященные М.С. Месежникову. Научн. ред. Козлова Г.Э., Прозоровский В.А. СПб.: ВНИГРИ, 1999. С. 87–116.
Киселев Д.Н. Зоны, подзоны и биогоризонты среднего келловея Центральной России // Спец. выпуск трудов естественно-географического факультета Ярославского гос. пед. ун-та. 2001. № 1. 38 с.
Киселев Д.Н. Аммониты и биостратиграфия келловейских отложений (река Сысола у села Вотча, Русская платформа) // Новости палеонтологии и стратиграфии. Приложение к журн. “Геология и геофизика”. 2006. Т. 47. Вып. 9. С. 47–69.
Киселев Д.Н., Меледина С.В. Аммонитовые комплексы и биогоризонты подзоны Kosmoceras Jason (средний келловей) на Русской платформе // Новости палеонтологии и стратиграфии. Приложение к журн. “Геология и геофизика”. 2004. Т. 45. Вып. 6–7. С. 157–175.
Князев В.Г., Кутыгин Р.В., Меледина С.В. Новая аммонитовая зональная шкала нижнего келловея севера Сибири // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 4. С. 45–64.
Князев В.Г., Меледина С.В., Алифиров А.С., Кутыгин Р.В. Среднекелловейский этап эволюции сибирских кардиоцератид // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция, экология и биостратиграфия. Материалы совещания (Москва, 2–4 апреля 2015 г.). Ред. Леонова Т.Б., Барсков И.С., Митта В.В. М.: ПИН РАН, 2015. С. 40–45.
Князев В.Г., Меледина С.В., Алифиров А.С. Protolongaeviceras (Cardioceratidae, Ammonoidea) – индикатор среднего келловея на севере Cибири // Геология и минерально-сырьевые ресурсы Сибири. 2019. № 2. С. 16–23.
Князев В.Г., Меледина С.В., Алифиров А.С. Средний келловей Сибири: аммониты и зональное деление // Стратиграфия. Геол. корреляция. 2020. Т. 28. № 3. С. 63–81.
Меледина С.В. Аммониты и зональная стратиграфия келловея Сибири. М.: Наука, 1977. 289 с. (Труды ИГиГ СО АН СССР. Вып. 356).
Меледина С.В. Бореальная средняя юра России. Новосибирск: Наука, 1994. 184 с. (Труды ИГиГ СО АН СССР. Вып. 819).
Меледина С.В. Новые находки келловейских и оксфордских кардиоцератид (Ammonoidea) Западной Сибири // Геология и геофизика. 1998. Т. 39. № 8. С. 1032–1038.
Митта В.В. Аммониты и биостратиграфия нижнего келловея Русской платформы // Бюлл. коллекционного фонда ВНИГНИ. 2000. № 3. 144 с.
Митта В.В. О филогении ранних Cardioceratidae (Ammonoidea) и среднерусских представителях Cadoceratinae на рубеже бата и келловея // Палеонтол. журн. 2016. № 4. С. 42–51.
Митта В.В., Костылева В.В., Глинских Л.А., Шурыгин Б.Н., Стародубцева И.А. Стратиграфия средней юры юго-запада Республики Татарстан // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 1. С. 31–46.
Никитин С.Н. Юрские образования между Рыбинском, Мологой и Мышкиным // Материалы для геологии России. 1881. Т. 1. № 2. 131 с.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г., Меледина С.В., Дзюба О.С., Лебедева Н.К., Пещевицкая Е.Б., Глинских Л.А., Горячева А.А., Хафаева С.Н. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082.
Репин Ю.С. Новые аммониты из Печерского верхнего келловея // Палеонт. журн. 2002. № 5. С. 33–38.
Репин Ю.С. Аммонитовая шкала Печорской юры // Бюллетень МОИП. Отд. геол.. 2007. Т. 82. Вып. 2. С. 24–31.
Репин Ю.С. Новости аммонитологии Печорской юры // Нефтегазовая геология. Теория и практика. 2008. Т. 3. № 3. С. 1–15.
Репин Ю.С., Захаров В.А., Меледина С.В., Нальняева Т.И. Атлас моллюсков Печорской юры // Бюлл. ВНИГРИ. 2006. № 3. 262 с.
Сазонов Н.Т. Юрские отложения центральных областей Русской платформы. Л.: Гостоптехиздат, 1957. 157 с.
Соколов Д.Н. К аммонитовой фауне Печорской юры // Тр. Геол. ком. Нов. сер. 1912. Вып. 76. 65 с.
Столярова Л.Р. Новый род кардиоцератид (Ammonoidea) из верхнего келловея–нижнего оксфорда Севера Сибири // Палеонт. журн. 2007. № 3. С. 10–15.
Столярова Л.Р., Меледина С.В. Первые находки рода Pavloviceras (Cardioceratidae, аммониты) в верхнем келловее Северной Сибири // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2004. Т. 45. Вып. 6–7. С. 177–181.
Стратиграфия юрской системы севера СССР. Ред. Сакс В.Н. М.: Наука, 1976. 436 с.
Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П., Ильина В.И., Меледина С.В., Гайдебурова Е.А., Дзюба О.С., Казаков А.М., Могучева Н.К. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск: Изд-во СО РАН, филиал “Гео”. 2000. 476 с.
Arkell W.J., Kummel B., Wright C.W. Systematic description // Treatise on Invertebrate Paleontology. Pt 4. Mollusca 4. Ed. Moor R.C. Lawrence, Kansas: University of Kansas Press, 1957. P. L129–L437.
Århus N., Birkelund T., Smelror M. Biostratigraphy of some Callovian and Oxfordian cores off Vega, Helgeland, Norway // Norsk Geologisk Tidsskrift. 1989. V. 69. P. 3956.
Buckman S.S. Yorkshire Type Ammonites. London: Wesley and Son, 1909–1930. V. 1–7.
Callomon J.H. A review of the biostratigraphy of the post-lower Bajocian Jurassic ammonites of western and nothern North America // Geol. Assoc. Can. Spec. Pap. 1984. № 27. P. 143–174.
Callomon J.H. The evolution of the Jurassic ammonite family Cardioceratidae. // Spec. Pap. in Paleontologie. 1985. № 33. P. 49–90.
Callomon J.H., Birkelund T. The Jurassic transgression and the mid-late Jurassic succession in Milne Land, central East Greenland // Geol. Mag. 1980. V. 117. № 3. P. 211–310.
Callomon J.H., Wright J.K. Cardioceratid and kosmoceratid ammonites from the Callovian of Yorkshire // Palaeontology. 1989. V. 32. Pt. 4. P. 799–836.
Courville P. Baptiser les Cardioceratinae: des noms et de prénoms “buckmano-arkelliens”. Systématique des genres et sous-genres // Fossiles. Hors série II. 2011. P. 27–38.
Courville P., Marchand D., Bonnot A., Raffray M., Double S. Cadoceratinae (ammonites d’origine boreale) dans le Callovien moyen et supérieur de la Haute-Marne et de la Côte-d’Or // Bulletin science Bourgogne. 1996. V. 48. P. 53–68.
Engkilde M., Surlyk F. Shallow marine syn-rift sedimentation: Middle Jurassic Pelion Formation, Jameson Land, East Greenland // Geol. Surv. Denmark and Greenland Bull. 2003. № 1. P. 813–863.
Howarth M.K. Systematic descriptions of the Stephanoceratoidea and Spiroceratoidea // Treatise Online 2017. Pt L, Revised. Vol. 3B, Ch. 6. P. 84–101.
Imlay R.W. Callovian (Jurassic) ammonites from the United States and Alasca. Pt. 2. Alasca Peninsula and Cook Inlet regions // US Geol. Surv. Prof. Pap. № 249-B. 1953. P. 41–108.
Imlay R.W. Stratigraphic distribution and zonation of Jurassic (Callovian) ammonites in Southern Alaska // US Geol. Surv. Prof. Pap. 1975. № 836. 28 p.
Imlay R.W. Middle Jurassic Bajocian and Bathonian Ammonites from Northern Alaska // US Geol. Surv. Prof. Pap. 1976. № 854. 22 p.
Keyserling A. Geognostische Beobachtungen. Wissen-schaftliche Beobachtungen auf einer Reise in das Petschora-Land im Jahre 1843. St. Petersburg, 1846. S. 149–406.
Kiselev D.N., Rogov M.A. Detailed biostratigraphy of the Middle Callovian–lowest Oxfordian in the Mikhaylov reference section (Ryazan region, European part of Russia) by ammonites // Volumina Jurassica. 2018. V. XVI. P. 73–186.
Larsen M., Piasecki S., Surlyk F. Stratigraphy and sedimentology of a basement-onlapping shallow marine sandstone succession, the Charcot Bugt Formation, Middle–Upper Jurassic, East Greenland // Geol. Surv. Denmark and Greenland Bull. 2003. № 1. P. 893–930.
Leckenby J. On the Kalloway rock of the Yorkshire Coast // The Quarterly J. Geol. Soc. London. 1859. V. 15. № 1. P. 4–15.
Page K.N. Ammonites // Fossils of the Oxford Clay. Eds. Martill D.M., Hudson J.D. Paleontol. Assoc. Field Guides to Fossils. London. 1991. V. 4. P. 86–143.
Reeside J.B. Some American Jurassic ammonites of the genera Quenstedtoceras, Cardioceras and Amoeboceras, family Cardioceratidae // U.S. Geol. Surv. Prof. Pap. 1919. V. 118. 64 p.
Salfeld H., Frebold H. Jura- und Kreidofossilen von Nowaya Zemlya // Rep. scient. Result Norw. Exped. Novaya Zemlya. 1924. V. 23. P. 1–12.
Vosgerau H., Larsen M., Piaseki S., Therkelsen J. A new Middle–Upper Jurassic succession on Hold with Hope, North-East Greenland // Geol. Surv. Denmark and Greenland Bull. 2004. № 5. P. 51–71.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция