

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 4, стр. 73-95
Ретроцерамы и униониды из байоса и нижнего бата (средняя юра) бассейна р. Ижма, север Европейской России
В. А. Захаров 1, *, А. П. Ипполитов 1, Н. Г. Зверьков 1, П. А. Безносов 2, Д. Н. Киселев 3
1 Геологический институт РАН
Москва, Россия
2 Институт геологии Коми НЦ УрО РАН
Сыктывкар, Россия
3 Ярославский государственный педагогический университет им. К.Д. Ушинского
Ярославль, Россия
* E-mail: mzarctic@gmail.com
Поступила в редакцию 15.10.2019
После доработки 15.11.2019
Принята к публикации 20.11.2019
Аннотация
Из байос-батских отложений, выходящих на поверхность по р. Ижма и вдоль ее правого притока р. Дрещанка (Тимано-Печорский регион), впервые собрана и описана представительная коллекция двустворок, принадлежащих к роду Retroceramus. Установлено присутствие 7 таксонов, из которых 4 определены до вида (из них 2 новых), а еще 3 – в открытой номенклатуре. Наиболее древний комплекс обнаружен стратиграфически ниже уровней, охарактеризованных находками позднебайосских аммонитов Arctocephalites arcticus, и допускает возможность датировки вмещающих отложений ранним байосом. Это предположение основано на определениях R. (R.) elongatus Kosh., 1960 и R. (R.) cf. lungershauseni Kosh., 1962 – видов, ранее упоминавшихся из нижнебайосских отложений азиатской части России. В этом же комплексе встречается R. (R.) alliensis Zakharov, sp. nov., а также двустворки сем. Unionidae, иногда образующие массовые скопления и относящиеся к новому роду и виду Mezounio zverkovi Zakharov, gen. et sp. nov. Вышележащие отложения верхнебайосской аммонитовой зоны Arctocephalites arcticus охарактеризованы единичными находками R. (R.) startensis Polub., 1992, R. (R.) elongatus Kosh., 1960 и R. (?) inopinatus Zakharov, sp. nov. Более высокая часть последовательности, не охарактеризованная аммонитами и относящаяся к пограничному байос-батскому интервалу, содержит R. (R.) cf. eximius (von Eichw.), а в нижнебатских отложениях зоны Arcticoceras ishmae найден единственный экземпляр, определенный как R. (R.) sp. indet.
ВВЕДЕНИЕ
Выходы докелловейских (байос-батских) отложений в бассейне р. Ижма ниже г. Сосногорска, в районе устья р. Дрещанка (рис. 1), являются ключевыми для расшифровки геологической летописи севера Европейской России в среднеюрскую эпоху. Они были впервые обнаружены в 1843 г. экспедицией А.А. Кейзерлинга, который описал из нижней части последовательности новый вид аммонитов Ammonites ishmae (Keyserling, 1846) [=Arcticoceras ishmae, согласно современной классификации]. Впоследствии этот вид был найден практически по всей Арктике и стал важным межрегиональным зональным репером, маркирующим начало обширной морской трансгрессии во многих регионах. На протяжении почти всего XX в. в отечественной литературе слои с/зона A. ishmae рассматривались как относящиеся к низам келловея (Соколов, 1912; Бодылевский, 1949; Сазонов, 1957; Меледина и др., 1987), однако в соответствии с данными зарубежных авторов (Callomon, 1985; Poulton, 1987) эта точка зрения была пересмотрена, и слои с/зона A. ishmae были перемещены в бат – сначала в верхний (Меледина и др., 1991; Сей, Калачева, 1992), затем в средний (Меледина, 1994); а в настоящее время хорошо обоснован их раннебатский возраст (Митта, Сельцер, 2002; Mitta et al., 2014).
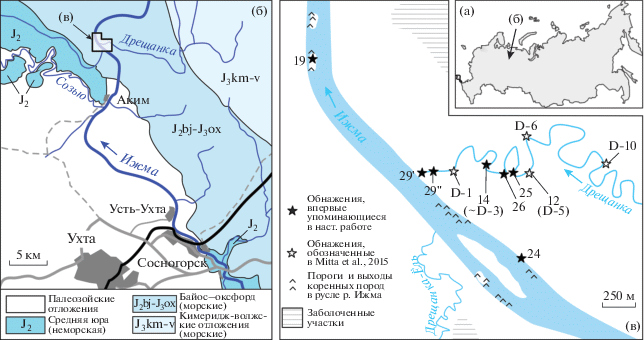
Рис. 1.
Географическое (а–в) и геологическое (б) положение разрезов, материал из которых описан в настоящей работе. Геологическая основа приведена по Государственной геологической карте масштаба 1 : 200 000 (Леонова, 1964), с незначительными изменениями.
В последние годы обширный фактический материал по разрезам средней юры р. Ижма и ее притока р. Дрещанка был опубликован В.В. Митта с соавторами (Митта, 2006, 2009; Митта и др., 2014a; Mitta et al., 2015). Важным открытием этих лет является установление (Митта, 2006) в разрезе еще более древних морских слоев, чем нижнебатские слои с A. ishmae, по находке в осыпи аммонита Arctocephalites arcticus – индекса одноименной аммонитовой зоны Бореального стандарта (Захаров и др., 1997; Меледина и др., 2011). Эта зона, согласно современным представлениям, соответствует верхнему байосу в полном его объеме (Меледина, 2014). Поэтому начало бореальной среднеюрской трансгрессии на территорию Европейской России сейчас по умолчанию датируется именно поздним байосом (Митта и др., 2014б).
Необходимо отметить, что изучение юрских разрезов по рекам Ижма и Дрещанка осложнено фрагментарностью выходов коренных пород и их в целом плохой обнаженностью, что не позволяет наблюдать единую и непрерывную последовательность. В естественных выходах не описано ни видимое основание морских отложений, которые, судя по смежным районам и данным скважин, залегают поверх континентальных и переходных фаций юры (см. Кравец и др., 1976; Чирва, Яковлева, 1982; Чирва, Куликова, 1986), ни общая подошва среднеюрского комплекса, залегающего на терригенно-карбонатных отложениях карбона (Леонова, 1964). А учитывая то, что выходы наиболее древних морских отложений, датированные ранее зоной A. arcticus, расположены в 3 км севернее предполагаемой общей подошвы среднеюрской толщи (рис. 1б), при падении слоев на СВ под углом ~0.5°, можно предполагать ~30–40 м скрытой мощности. Одно это обстоятельство автоматически допускает присутствие в разрезе еще более древних морских отложений, чем верхнебайосские.
В июле–августе 2018 г. авторами настоящей статьи проводились полевые исследования в бассейне р. Ижма. Переизучение коренных выходов средней юры по рекам Ижма и Дрещанка в районе устья последней позволило сопоставить многочисленные изолированные выходы и построить более-менее полную сводную последовательность, а также уточнить стратиграфические построения предшественников (Ипполитов и др., 2019). В интервале, соответствующем верхнебайосской зоне A. arcticus, и в более древних слоях были найдены остатки ретроцерамов – двустворчатых моллюсков, лежащих в основе зонального расчленения среднеюрских отложений по двустворкам на севере Сибири и в Северо-Восточной Азии. Дополнительно в 2019 г. А.П. Ипполитовым было найдено несколько экземпляров в более высокой части последовательности, относящейся к пограничному байос-батскому интервалу и нижнему бату. В настоящей статье представлены результаты таксономического и биостратиграфического изучения этих находок.
Вклад авторов. Определения и палеонтологические описания двустворок выполнены В.А. Захаровым, им же сформулированы основные положения разделов “Обсуждение” и “Заключение”, в том числе вывод о возрасте слоев. Часть экземпляров собрана П.А. Безносовым в 2014 г., а полноценное изучение разрезов и сбор дополнительного материала проводились А.П. Ипполитовым, Н.Г. Зверьковым и Д.Н. Киселевым в 2018 и 2019 годах. Н.Г. Зверькову принадлежит основная заслуга в части препарирования и подготовки к публикации описанного в настоящей статье фактического материала. А.И. Ипполитовым даны описания разрезов, составлены стратиграфические колонки и написаны разделы “Введение”, “Материал” и, частично, “Обсуждение”.
МАТЕРИАЛ
Последовательность среднеюрских отложений в бассейне р. Ижма близ устьевой части р. Дрещанка представлена терригенной толщей мощностью несколько десятков метров, сложенной тонким переслаиванием алевро-глинистых и алевро-песчаных разностей. На многих уровнях хорошо заметна пологая однонаправленная косая слоистость, а также присутствуют многочисленные обугленные растительные остатки. На отдельных уровнях располагаются прослои плотных песчаников (и алевролитов) на карбонатном цементе. Два прослоя песчаников, по-видимому, латерально выдержанны по всему району исследований и могут являться надежными литостратиграфическими реперами.
Верхний из них – это “арктикоцерасовый песчаник” (= пласт С на рис. 2), из которого происходят описанные ранее в литературе многочисленные находки раннебатской макро- и микрофауны зоны A. ishmae, в том числе довольно разнообразный комплекс двустворок (Кравец и др., 1976; Репин и др., 2006; Митта и др., 2011; Mitta et al., 2015). Именно к этому пласту, по-видимому, относятся упоминания находок ретроцерамов в старых работах (Бодылевский, 1949: “Inoceramus ex. gr. retrosus”; Бодылевский, 1963: “Inoceramus sp.”) и совершенно определенно – в недавних публикациях (Митта и др., 2013: “Retroceramus sp.”)11.
Рис. 2.
Распределение описанных двустворок и аммонитов в изученных разрезах по рекам Ижма и Дрещанка. Сокращения: R. – Retroceramus, M. – Mezounio.
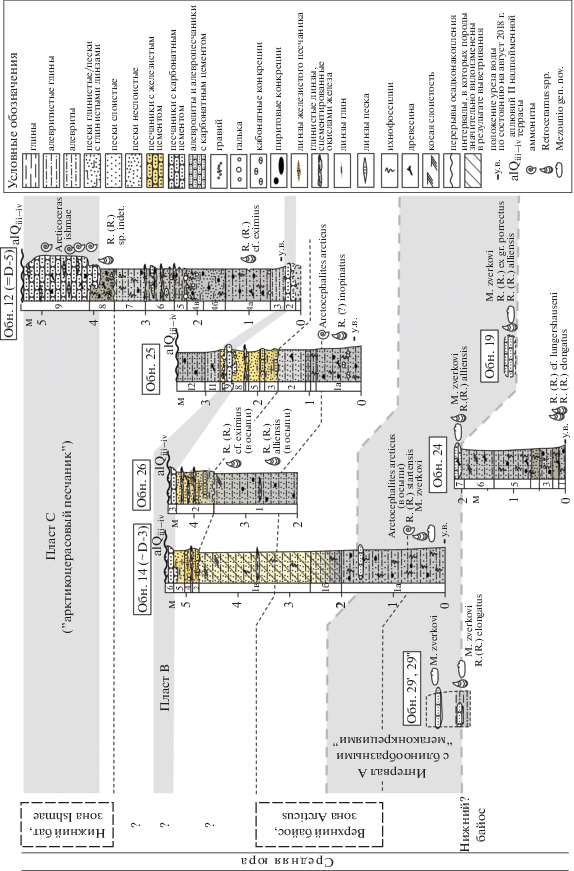
Второй репер – это прослой песчаника, залегающий на ~4 м ниже “арктикоцерасового песчаника” и не охарактеризованный макрофоссилиями (= пласт B на рис. 2). Ранее он был ошибочно интерпретирован В.В. Митта с соавторами как фациальный аналог арктикоцерасового песчаника в разрезах D-3 и D-1 (Митта, 2009; Mitta et al., 2015).
Описанный в настоящей работе материал происходит из нескольких обнажений (№№ 12, 19, 24, 25, 26, 29', 29''; см. рис. 1в), представленных как изолированными выходами песчаников, так и полноценными разрезами, примерно половина которых ранее не фигурировала в литературе. Они кратко охарактеризованы ниже, в порядке от более молодых к более древним горизонтам.
Обнажение 12 расположено в левом берегу р. Дрещанка и соответствует разрезу D-5 sensu Mitta et al., 2015. В основании на урезе воды в виде порога вскрывается пласт песчаника В, выше которого в крутом берегу обнажается рыхлая терригенная толща сложного строения, мощностью 3.3 м, а еще выше вскрыт арктикоцерасовый песчаник (= пласт С) видимой мощностью 1.5 м. Последний охарактеризован многочисленными находками единственного вида аммонитов Arcticoceras ishmae (Киселев, 2020a, 2020б) и относится к нижнему бату (Mitta et al., 2015). Ретроцерамы в обнажении 12 были собраны in situ в рыхлых разностях: один экземпляр – немного выше пласта В, второй – сразу под пластом С, в слое зеленоватого песка. Этот песок, за исключением его рыхлого сложения, литологически совершенно идентичен арктикоцерасовому песчанику и уже в пределах изученного обнажения замещается последним по латерали.
Обнажение 25 представляет собой расчистку в левом берегу р. Дрещанка между разрезами 12 (D-5) и D-3 sensu Mitta et al., 2015. Разрез вскрывает пласт B (= пачка 0 в разрезе D-5), а также подстилающие и перекрывающие его алевро-песчаные отложения. Ретроцерамы были обнаружены в глинистых алевритах в 1.75–2 м ниже подошвы пласта B, и примерно на этом же уровне in situ был найден аммонит Arctocephalites arcticus – зональный индекс бореального верхнего байоса.
Обнажение 14 представляет собой разрез в правом крутом берегу р. Дрещанка протяженностью около 30 м. По-видимому, оно соответствует разрезу D-3 sensu Mitta et al., 2015. Указание положения последнего на карте в упомянутой публикации является некорректным, соответствуя низкой I надпойменной террасе р. Дрещанка. В кровле обнажения 14 вскрыта нижняя часть горизонта B, а ниже по разрезу ~5 метров терригенной толщи. Ретроцерамы были собраны примерно в 4.5 м ниже горизонта B. На этом же уровне, но в осыпи был подобран пиритизированный фрагмокон аммонита A. arcticus, аналогичный по сохранности более ранней находке, описанной В.В. Митта из разреза D-3 (Митта, 2006). Поскольку пиритовые конкреции в верхней части обнажения имеют сильно выветрелый облик, с большой долей вероятности можно предполагать происхождение аммонитов примерно с тех же уровней, где были найдены ретроцерамы.
Обнажение 26 по латерали смыкается с близко расположенным обнажением 14 и в целом повторяет его верхнюю половину. Однако оно характеризуется значительно меньшей выветрелостью коренных пород. Один отпечаток крупного ретроцерама был найден в осыпи верхней части разреза, а на бечевнике было подобрано несколько пиритизированных ювенильных раковин с сочлененными створками хорошей сохранности.
Обнажения 29' и 29'' – это небольшие изолированные выходы пластов карбонатного алевропесчаника, залегающие на 3.5–4.5 м ниже пласта B и расположенные в устьевой части русла р. Дрещанка (0–100 м от устья). Они примерно соответствуют “пачке 0” в разрезе D-1 (sensu Mitta et al., 2015), а в верхней части – уровню находки A. arcticus в разрезе 14 (~D-3), однако их точная корреляция с последним затруднительна в связи с тем, что слои алевропесчаника не выдерживаются по латерали. По сути, они представляют собой огромные блинообразные тела конкреционной природы диаметром от нескольких до десятков метров при мощности не более 0.5 м. Резко закругленные латеральные замыкания таких “мегаконкреций” можно наблюдать в береговых обнажениях р. Дрещанка вблизи ее устья. Эти конкреции, судя по всему, в массе приурочены к узкому интервалу разреза мощностью не более 1.5–2 м (интервал А на рис. 2). Ретроцерамы были обнаружены на отдельных плоскостях напластования в конкрециях алевропесчаников; именно отсюда происходят первые образцы, собранные в 2014 г. П.А. Безносовым (обнажение 29', рис. 3). Также на одном из изолированных блоков, лежащих непосредственно в русле р. Дрещанка (обнажение 29''), было обнаружено небольшое скопление раковин сем. Unionidae, описанных ниже как Mezounio zverkovi Zakharov, gen. et sp. nov.
Рис. 3.
Скопление раковин ретроцерамов в алевропесчанике, обнажение 29'. Длина молотка 28 см. Находка П.А. Безносова, 2014 г.

Обнажение 19 находится в левой половине русла р. Ижма в 1 км ниже устья р. Дрещанка и представляет собой изолированный площадной выход блинообразной конкреции алевропесчаника, располагающейся примерно на том же уровне последовательности, что и алевропесчаники в точках наблюдения 29' и 29''. В этой конкреции на одной из плоскостей напластования было обнаружено массовое (>150 экз.) скопление M. zverkovi Zakharov, gen. et sp. nov. (рис. 4), а также единичные изолированные створки ретроцерамов.
Рис. 4.
Массовое скопление раковин Mezounio zverkovi Zakharov, gen. et sp. nov. в алевропесчанике из обнажения 19. а – общий вид места находки; б – плита со скоплением унионид.

Обнажение 24 расположено в 1 км выше устья р. Дрещанка в правом берегу р. Ижма, под рыбацкой избушкой. Над урезом воды здесь вскрывается примерно 2 м алевро-глинистой толщи, венчающейся конкрециеобразным прослоем карбонатного алевропесчаника. Относительное положение этого обнажения в общей последовательности определяется следующим образом. Пласт тонкослоистого карбонатного алевропесчаника в верхах разреза литологически идентичен блинообразным конкрециям обнажений 19 и 29' (и также содержит остатки Mezounio zverkovi Zakharov, gen. et. sp. nov.), но при этом отличается от залегающего выше пласта B. Соответственно, на рис. 2 конкреции, наблюдавшиеся в точках 19, 29' и 24, условно помещены на единый стратиграфический уровень, а более низкая часть разреза 24, не содержащая конкреций/прослоев песчаника, интерпретируется нами как наиболее древняя часть изученной последовательности, непосредственно подстилающая интервал с блинообразными мегаконкрециями. Ретроцерамы были обнаружены на двух уровнях обнажения 24 – в верхнем прослое алевропесчаника, где был найден также неопределимый фрагмент альвеолярной части белемнита шириной около 12 мм, и в алевритистых глинах в нижней части разреза.
Таким образом, распределение находок ретроцерамов в последовательности является неравномерным: они приурочены к отдельным уровням, иногда образуя автохтонные скопления из нескольких (обычно 2–4) экземпляров. Часть находок происходит из карбонатных песчаников, где ретроцерамы представлены внутренними ядрами, а часть – из глинисто-алевритовой толщи, где они представлены в той или иной степени деформированными (раздавленными) целыми раковинами.
Собранная и изученная коллекция насчитывает 12 целых створок и около 20 фрагментов раковин ретроцерамов и >150 преимущественно отдельных створок раковин унионид.
Оригиналы хранятся в Государственном геологическом музее им. В.И. Вернадского, г. Москва (ГГМ) и Геологическом музее им А.А. Чернова Института геологии Коми Научного Центра Уральского отделения РАН, г. Сыктывкар (ИГ КНЦ).
ПАЛЕОНТОЛОГИЧЕСКОЕ ОПИСАНИЕ
ОТРЯД PTERIOIDA NEWELL, 1965
НАДСЕМЕЙСТВО INOCERAMOIDEA C. GIEBEL, 1852
СЕМЕЙСТВО RETROCERAMIDAE POLUBOTKO ET SEY IN NEVESSKAJA ET AL., 2013
Хотя название “Retroceramidae” широко используется в отечественной литературе уже почти 50 лет (Кошелкина, 1971; Полуботко, 1992 и мн. др.), попадая в том числе и в заголовки публикаций (как “ретроцерамиды”; см. Кошелкина, 1971, 1974), диагноз семейства, необходимый для обеспечения пригодности названия (Международный кодекс зоологической номенклатуры (МКЗН), ст. 13.1, 13.2), был впервые опубликован лишь в 2013 г. И.В. Полуботко и И.И. Сей (в Невесская и др., 2013). Согласно положениям статей 50.1 и 10.1 МКЗН, авторство семейства в этом случае принадлежит указанным авторам.
Род Retroceramus Koshelkina, 1957
Подрод Retroceramus s. str.
До 1957 г. большинство видов иноцерамид, широко распространенных в юрских отложениях Бореальной области, включалось в состав рода Inoceramus J. Sowerby, 1822. Специфика строения, в частности, лигамента послужила основанием для выделения самостоятельных подродов I. (Retroceramus) и I. (Fractoceramus) (Кошелкина, 1957). Позднее подрод Retroceramus был возведен в ранг рода, и в его составе были выделены новые подроды R. (Boreioceramus) Kosh., 1971 и R. (Mennericeramus) Kosh., 1971. Кроме того, в качестве самостоятельного рода Arcticeramus Kosh., 1962 были обособлены верхнеюрские представители группы, имеющие неравностворчатую раковину.
Долгое время род Retroceramus и его подроды не признавались советскими палеонтологами в качестве валидных таксонов, несмотря на то, что они были включены в американский справочник “Treatise on invertebrate paleontology” (1969, p. N320). Например, в публикациях 1980-х годов (Сей, Калачева, 1980; Меледина и др., 1987 и мн. др.) род именуется Mytiloceramus Rollier, 1914. Лишь в 1990-х годах валидное наименование рода, наконец, прочно вошло в палеонтологическую практику. Подроды, впрочем, по-прежнему не используются в публикациях, хотя и включены в современные справочники по двустворкам (Невесская и др., 2013). В какой-то мере это объясняется тем, что до сих пор не проведена глубокая ревизия видового состава ретроцерамов на основе популяционного, филетического и палеобиогеографического анализа (см. также раздел “Обсуждение”).
Виды описаны ниже в порядке их первого появления в изученной последовательности, снизу вверх.
Retroceramus (R.) elongatus Koshelkina, 1960
Табл. I , фиг. 1; табл. II, фиг. 1
Retroceramus (R.) elongatus: Кошелкина, 1960, с. 37, табл. 7 , фиг. 5; 1961, с. 149, табл. 1 , фиг. 4; 1963, с. 142, табл. 5 , фиг. 4; Полуботко в Ефимова и др., 1968, с. 70, табл. 74 , фиг. 3(?); табл. 75 , фиг. 1; табл. 76 , фиг. 1; табл. 77 , фиг. 1.

Retroceramus (R.) sublimus: Кошелкина, 1969а, с. 90, табл. XX , фиг. 1–3.
?Retroceramus porrectus: Сей, Калачева, 1980, с. 121, табл. XXVIII , фиг. 1–2.
Материал. Две створки удовлетворительной сохранности (экз. № ГГМ Рц1, экз. № ИГ КНЦ 415/55) и несколько крупных фрагментов.
Описание. В оригинальном описании отмечается крупный размер раковины вида, достигающей в длину 157 мм, с превышением длины над шириной в два раза; умеренная выпуклость, неширокое крыло. Выпуклость в области макушки круто опускается к переднему и заднему краям и постепенно – к нижнему. Макушки широкие, наклонены к замочному краю и слегка повернуты вперед. Передний край всегда выпуклый, а замочный – прямой. Задний край описывает слабо выпуклую дугу. Поверхность створок с 12–13 асимметричными, редко расположенными и неясно ветвящимися концентрическими складками, сглаживающимися к макушке. Некоторые складки дихотомируют. Отдельные резкие складки проходят и на крыло. Область заднего края, а также примыкающая к нему часть крыла обычно гладкие. К переднему краю концентрические складки подходят под очень острым углом. Близкое к изложенному, но более детальное морфологическое описание раковины приводит И.В. Полуботко (в Ефимова и др., 1968, с. 70).
По всем перечисленным в этих описаниях признакам два наиболее крупных экземпляра в нашей коллекции, представленных узкими сильно удлиненными правой и левой створками (табл. I , фиг. 1; табл. II, фиг. 1), соответствуют идентифицированному виду. На обеих створках полностью сохранился иризирующий раковинный слой, покрывающий их по всей длине по линии наибольшего роста, превышающей 160 мм. Скульптура представлена четкими концентрическим складками, закономерно расположенными в верхней части створки, и грубыми неправильными складками, резче выраженными по нижнему краю. Очертания верхнего края правой створки скрыты под перекрывающим его фрагментом другой раковины. Примакушечная часть сужена, острый кончик макушки утопает в породе.
Таблица I . Ретроцерамы из байоса разрезов на реках Ижма и Дрещанка. Здесь и в табл. II –IV изображенные экземпляры приведены в натуральную величину, за исключением особо отмеченных случаев. 1 – Retroceramus (R.) elongatus Koshelkina, 1960; экз. № ГГМ Рц1, а – левая створка, б – замок, связочные ямки, обн. 24, сл. 1; нижний (?) байос; 2 – Retroceramus (R.) cf. lungershauseni Koshelkina, 1962; экз. № ГГМ Рц7, левая створка на ранней стадии онтогенеза; обн. 24, сл. 1; нижний (?) байос; 3–5 – Retroceramus (R.) alliensis Zakharov, sp. nov., нижний (?) байос: 3 – экз. № ГГМ Рц3, правая створка, обн. 19; 4 – экз. № ГГМ Рц4, левая створка на ранней стадии онтогенеза, обн. 24, сл. 7; 5 – экз. № ГГМ Рц5, левая створка, обн. 24, сл. 7.

Таблица II. Ретроцерамы из байоса разрезов на реках Ижма и Дрещанка. 1 – Retroceramus (R.) elongatus Koshelkina, 1960, экз. № ИГ КНЦ 415/55, правая створка, обн. 29', нижний (?) байос; 2 – Retroceramus (R.) startensis Polubotko, 1992, экз. № ГГМ Рц6, правая створка, обн. 14, сл. 1а, верхний байос, зона Arctocephalites arcticus; 3 – Retroceramus (?) inopinatus Zakharov, sp. nov., экз. № ГГМ Рц9 (голотип), левая створка на ранней стадии онтогенеза, а – вид с наружной стороны, б – вид со стороны смычного края, в – детали строения связочной полоски, ×2; обн. 25, сл. 1а, верхний байос, зона Arctocephalites arcticus; 4 – Retroceramus (R.) ex gr. porrectus Polubotko, 1992, экз. № ГГМ Рц8, левая створка, обн. 19; нижний (?) байос.

Таблица III. Ретроцерамы из верхнего байоса–нижнего бата разрезов на р. Дрещанка. 1 – Retroceramus (R.) alliensis Zakharov, sp. nov.; экз. № ГГМ Рц10 (голотип), обн. 26, сл. 1 (осыпь на бечевнике, ~1.5 м вниз от кровли слоя), верхний байос, зона Arctocephalites arcticus: 1а – левая створка; 1б – правая створка; 1в – вид со стороны заднего края; 1г – вид со стороны переднего края; 2–3 – Retroceramus cf. eximius (von Eichwald, 1871), безаммонитовый пограничный интервал байоса и бата: 2 – экз. № ГГМ Рц11, обломок левой створки, обн. 26, осыпь верхней части сл. 1; верхний байос, зона Arctocephalites arcticus; 3 – экз. № ГГМ Рц12, обломок левой створки, обн. 12 (D-5), 45–50 см от подошвы сл. 4а; 4 – Retroceramus (R.) sp. ind.; экз. № ГГМ Рц13, левая створка; обн. 12 (D-5), сл. 8 (средняя часть); нижний бат, зона Arcticoceras ishmae.

Таблица IV. Mezounio zverkovi Zakharov, gen. et sp. nov. из байоса разрезов на реках Ижма и Дрещанка. 1 – экз. № ГГМ Un1, правая створка; а – снаружи, б, в – изнутри; г – прорисовка изнутри; 2 – экз. № ГГМ Un2, правая створка; 3 – экз. № ГГМ Un3, целая раковина, голотип, а – левая створка, б – вид спереди, в – вид сверху, г – правая створка; 4 – экз. № ГГМ Un4, а – правая створка с обломанным задним краем, б – левая створка с обломанной частью заднего края; 5 – экз. № ГГМ Un5 с хорошо сохранившимся отпечатком переднего аддуктора, а – правая створка, б – левая створка; 6 – экз. № ГГМ Un6, отпечаток переднего аддуктора правой створки; 7 – экз. № ГГМ Un7 с отпечатоком заднего аддуктора левой створки; 8 – экз. № ГГМ Un8, правая створка (внизу) и крупный фрагмент левой створки той же раковины. Все образцы – обн. 19; нижний (?) байос.

Сравнение. Изображенные в публикациях раковины этого вида перечислены в синонимике. Судя по изображению и описанию морфологии раковины, наши образцы больше всего напоминают находки из байосcкого яруса Северо-Востока России (Ефимова и др., 1968, с. 70, табл. 75, 76 , фиг. 1, табл. 77 ). От R. (R.) bulunensis Koshelkina, 1961 описываемый вид отличается узкой вытянутой сильно скошенной раковиной, удлиненной (оттянутой) примакушечной частью и крупными резкими округленными концентрическими складками, покрывающими всю поверхность створки. Следует обратить внимание на R. porrectus, который определен И.И. Сей из верхнего байоса Дальнего Востока (Сей, Калачева, 1980, с. 121, табл. XXVIII , фиг. 1–2) и по очертаниям раковины, характеру наружного рельефа и особенностям скульптуры напоминает R. elongatus c р. Левый Кедон (Ефимова и др., 1968, табл. 75 , фиг. 1).
Местонахождение: р. Ижма, обн. 24, сл. 1; р. Дрещанка, устьевая часть, обн. 29'.
Геологический возраст и географическое распространение. Предполагаемый возраст – байос. Оригинальный материал происходит с р. Лена и собран у мыса Кыстатым и в 13 км севернее него, а также в нижнем течении р. Молодо (сборы З.В. Кошелкиной, 1954, 1958 гг.). Большая часть помещенных в синонимику таксонов происходит из байосских отложений, но стратиграфический диапазон некоторых включает и бат. В разрезах средней юры Омолонского массива (Северо-Восток Азии), для которых Retroceramus elongatus используется в качестве зонального индекса, возраст одноименного подразделения определен как поздний байос под вопросом (Кошелкина, 1980б).
Retroceramus (R.) cf. lungershauseni Koshelkina, 1962
Табл. I , фиг. 2
Материал. Одна левая створка молодого экземпляра удовлетворительной сохранности (экз. № ГГМ Рц7).
Описание и сравнение. Интерес представляет сравнение морфологических особенностей экземпляров, находящихся на относительно ранней стадии индивидуального развития, изображенных в настоящей работе (табл. I , фиг. 2) и у И.В. Полуботко (Ефимова и др., 1968, табл. 64 , фиг. 2; 1992, табл. 29 , фиг. 9 ). Хотя наш экземпляр сохранился не полностью, характер очертания краев раковины (плавно изогнутый передний, почти прямой задний, относительно длинный смычной край), слабо выраженная скульптура, небольшая выпуклость, проходящая от макушки к нижнему краю и исчезающая вблизи его, почти идентичны образцу с р. Левый Кедон (басс. р. Оленек).
Голотип R. (R.) lungershauseni (Кошелкина, 1962, с. 66–67, табл. IX , фиг. 1) из нижнего (?) аалена Приверхоянского прогиба похож по очертаниям и скульптуре верхней части раковины на указанные выше экземпляры, но явно отличается коротким связочным (замочным) краем. Ограниченный материал, привлеченный при первоописании вида, не позволяет судить о его характерных признаках. Этим можно объяснить редкость упоминания R. (R.) lungershauseni в публикациях. Странно, что автор вида сравнивает его с R. elongatus (Кошелкина, 1961, с. 149, табл. I , фиг. 4), который по всем признакам отличается от R. (R.) lungershauseni. Скорее сравнения заслуживает изображенный на той же таблице, что и R. (R.) lungershauseni, R. (R.) elegans (Кошелкина, 1962, табл. IX , фиг. 4), единственная левая створка которого не слишком отличается от левой же створки рассматриваемого вида.
Поскольку морфологически близкие виды группы R. elegans–R. jurensis крайне важны для датирования нижней части средней юры, им следует уделить особое внимание. К этой группе, помимо номинативных видов, И.В. Полуботко (1992, с. 69) относила ряд менее известных видов, включая и R. lungershauseni Kosh., который выделяется, по ее мнению, “более длинным связочным краем, меньшей скошенностью, большей шириной раковины и более широкой площадкой необособленного крыла”. Это описание отличается от изображения вида (Полуботко, 1992, табл. 29 , фиг. 9 ), которое лучше соответствует экземпляру из нашей коллекции (табл. I , фиг. 2).
Следует заметить, что как только появляются фотографии экземпляров видов из близких по возрасту стратиграфических интервалов, среди них всегда находятся весьма сходные. Так, по свидетельству И.В. Полуботко, ключевые виды “характеризуются широкой индивидуальной изменчивостью и образуют гомологичные ряды форм на различных возрастных уровнях” (Полуботко, 1972б). Такие формы встречаются совместно и связаны непрерывными переходами признаков. Выделяемые в этих рядах виды, основанные на отдельных характерных признаках, И.В. Полуботко предложила именовать “условными”; и понятно, что в таких ситуациях возможно лишь субъективное таксономическое решение. Например, при сравнении “условных” видов R. anilis (G. Pčelinceva, 1962) (Полуботко, 1992, с. 70, табл. 29 , фиг. 3) и R. lungershauseni (Полуботко, 1992, с. 70, табл. 29 , фиг. 9 ) из одного и того же ясчанского горизонта, нам не удалось обнаружить достойных для обособления межвидовых различий. В сходной ситуации оказалась и И.И. Сей (Сей, Калачева, 1980, с. 104). Описывая Mytiloceramus [= Retroceramus] ex gr. elegans Kosh., она обратила внимание на то, что “среди… образцов выделяются менее скошенные экземпляры, близкие к M. anilis (G. Pĉel.), и более скошенные, приближающиеся к M. tugurensis Sey и M. elegans Kosh.”.
Местонахождение: р. Ижма, обнажение 24, слой 1.
Геологический возраст и географическое распространение. Предполагаемый возраст образца – ранний байос. Голотип вида происходит из разреза темно-серых и коричневато-серых алевролитов нижнего (?) аалена в среднем течении р. Молодо Приверхоянского прогиба (Кошелкина, 1962). Этот возраст не подкреплен находками аммонитов. Вид R. lungershauseni указан из ясчанского горизонта, что соответствует b-зоне R. elegans–R. jurensis верхнего аалена–основания байоса Северо-Востока России и Северной Сибири (Полуботко, 1992).
Retroceramus (R.) alliensis Zakharov, sp. nov.
Табл. I , фиг. 3–5; табл. III, фиг. 1
Название вида от alliensis (лат.) – чужой.
Голотип: экз. № ГГМ Рц10, полная сочлененная раковина, внутри заполненная пиритом; р. Дрещанка, обн. 26, сл. 1, с бечевника.
Материал. Три целые створки (экз. № ГГМ Рц3–5), целый экземпляр (голотип экз. № ГГМ Рц10) и несколько фрагментов ядер различной сохранности с остатками раковинного слоя.
Диагноз. Раковина среднего размера (длина наиболее крупного экземпляра достигает 100 мм по оси роста), клиновидная (скошенность составляет 35°), с вытянутой примакушечной частью (апикальный угол 40°), узкая (длина более чем в два раза превышает ширину), передний связочный угол чуть более 60°, задний – около 140°, слабо до умеренного выпуклая, равностворчатая, c почти идеально параллельными передним и задним краями. Скульптура представлена относительно правильными, широко расположенными, редкими концентрическими складками с мягко сглаженными гребнями, постепенно укрупняющимися к нижнему краю.
Сравнение и замечания. В комплексе, собранном на р. Дрещанка, нет форм, морфологически близких к описываемому новом виду. По размеру раковины ближе всего стоит R. (R.) startensis. Однако на этом сходство заканчивается: раковины отличаются по очертаниям, скошенности, соотношению всех краев, характеру скульптуры. По некоторым признакам – узкой, клиновидной, относительно удлиненной раковине с вытянутой заостренной примакушечной частью и почти идеально параллельными передним и задним краями – новый вид сближается с группой видов, происходящих преимущественно из ретроцерамовой зоны R. elegans–R. jurensis, таких как R. (R.) mongkensis Koshelkina, 1969а; R. (R.) elegans Koshelkina, 1962; R. (R.) jurensis Koshelkina, 1969а; R. (R.) priscus (Sey, 1972); R. (R.) subtilis (Sey, 1972), а также с видами из слоев с R. (R.) merengensis и R. (R.) gizhigensis кровли ясчанского горизонта Северного Приохотья (Полуботко, 1992, с. 70–71, табл. 30 , фиг. 2, 3). Экземпляры R. (R.) alliensis Zakharov, sp. nov. существенно уступают по размеру (прежде всего, по длине раковины) наиболее крупным формам, перечисленным выше. Отличаются указанные виды и по характеру скульптуры, которая у большинства из них сглаженная либо представлена слабо рельефными, неправильными, густо расположенными гребневидными складками. От похожего R. acutus Polubotko, 1992 (Полуботко, 1992, с. 74–75, табл. 35 , фиг. 1, 2; табл. 36 , фиг. 1; табл. 37 , фиг. 2) экземпляры с р. Дрещанка четко отличаются субпрямоугольной раковиной, которая у R. acutus каплевидная и существенно более крупная у взрослых форм (Полуботко, 1992, с. 74–75, табл. 35 , фиг. 1; табл. 36 , фиг. 1). Помимо того, у наших экземпляров, как и у R. startensis, параллельные передний и задний края и, конечно, не столь сильно вытянутая примакушечная часть. Имеются существенные отличия и в типе скульптуры, более упорядоченной и “изящной” у R. (R.) alliensis Zakharov, sp. nov.
Местонахождение: р. Ижма, обн. 19, 24 (из прослоев карбонатных алевролитов) и р. Дрещанка, обн. 26 (осыпь на бечевнике).
Геологический возраст и географическое распространение. Наиболее древние находки, возможно, имеют раннебайосский возраст (см. ниже), а голотип происходит из верхнебайосской зоны A. arcticus. R. (R.) alliensis Zakharov, sp. nov. пока известен только на севере Европейской России, в басс. р. Ижма (Тимано-Печорский регион).
Retroceramus (R.) ex gr. porrectus (von Eichwald, 1871)
Табл. II, фиг. 4
Материал. Примакушечный фрагмент левой створки взрослого экземпляра удовлетворительной сохранности (экз. № ГГМ Рц8).
Описание и таксономическая трактовка. Судя по сохранившемуся контуру фрагмента, раковина достигала крупного размера и имела клиновидную сильно скошенную форму, характерную для видов группы R. (R.) porrectus. Покрытая грубыми концентрическими складками, вытянутая верхняя и примакушечная части сходны по форме, например, с изображенным экземпляром правой створки R. porrectus из басс. р. Армань (Ефимова и др., 1968, табл. 85 , фиг. 2). Впрочем, грубые рельефные складки характерны для многих видов среднеюрских крупноразмерных ретроцерамов. Точная диагностика вида затруднена из-за недостаточного материала.
Местонахождение: р. Ижма, обн. 19.
Геологический возраст и географическое распространение. Похожие формы в Северном Приохотье помещаются в татынгычанский горизонт, который имеет условные границы и согласно современной аммонитовой схеме (Меледина, 2014) примерно соотносится с верхами нижнего байоса.
Retroceramus (R.) startensis Polubotko, 1992
Табл. II, фиг. 2
Retroceramus startensis: Полуботко, 1992, с. 72, табл. 31 , фиг. 5.
Материал. Единственная правая створка (экз. № ГГМ Рц6).
Описание. Раковина 90 мм по оси роста, слабо выпуклая в верхней части и умеренно – в средней, ширина превышает длину примерно на 15%, скошенность составляет 45°, апикальный угол 55°, передний связочный угол чуть более 70°, задний связочный угол 125°. Связочный край равен примерно половине длины и почти равен ширине. Передний край почти прямой. Крыло довольно широкое и практически не обособлено от створки. Скульптура представлена тесно неправильно расположенными, изменчивыми по длине раковины, но хорошо выраженными на всей ее поверхности складками, продолжающимися на крыло, перед которым некоторые складки, несколько ослабевая, исчезают. Макушка четкая, слегка заостренная, почти не выступающая.
Сравнение. По мнению И.В. Полуботко, R. startensis напоминает R. merengensis Polubotko, 1992 в начальной стадии онтогенеза последнего (Полуботко, 1992, с. 71, табл. 30 , фиг. 2). Однако указанные ею отличия – меньшие размеры, слабо выраженная клиновидная форма, иной характер макушек и резко выраженные складки – не позволяют считать данные виды близкородственными. Рассматривая довольно многочисленный в видовом отношении комплекс ретроцерамов, заключенных в стратиграфическом интервале двух горизонтов, меренгинского и татынгычанского, соответствующих нижнему байосу в терминах современной версии зональной шкалы по аммонитам (Меледина, 2014), нам не удалось обнаружить виды, сходные с Retroceramus (R.) startensis Polubotko, 1992.
Местонахождение: р. Дрещанка, обн. 14, сл. 1.
Геологический возраст и географическое распространение. На Северо-Востоке России образцы вида найдены в основании меренгинского горизонта (ретроцерамовая зона R. lucifer, нижний байос – аналог аммонитовой зоны Arkelloceras tozeri; см. Меледина и др., 2011) в междуречье Омолона и Коркодона, в бассейне р. Левый Кедон, а также в Печорской впадине в бассейне р. Ижма (верхний байос, зона Arctocephalites arcticus).
Retroceramus (?) inopinatus Zakharov, sp. nov.
Табл. II, фиг. 3
Название вида от inopinatus (лат.) – неожиданный.
Голотип: экз. № ГГМ Рц9, левая створка хорошо сохранившейся раковины с обломанным нижним краем.
Материал. Только голотип.
Диагноз. Наиболее четкий признак нового вида – весьма существенная скошенность раковины (угол скошенности 34°–36°) и длинный замочный край, до 30 мм при длине створки 40 мм. Передний край очерчен слабо изогнутой дугой. Имеется еще одна особенность: это необычное для рода расположение связочной полоски перпендикулярно плоскости смыкания створок (табл. II, фиг. 3в). Поскольку ямки для связки обращены “вверх”, ямки противоположной створки должны их накрывать сверху. Это означает, что правая створка должна быть более выпуклой, чем левая. Следовательно, раковина была неравностворчатая, что не характерно для ретроцерамов. У описываемого вида фиксируется конечное расположение макушки на раковине, в то время как “макушка у юрских форм [рода Retroceramus] не является конечной и отделена от переднего края выступом или площадкой” (Кошелкина, 1963, с. 135). На сохранившейся части раковины почти идеально правильно расположены 15 четких, слегка поднимающихся над поверхностью изящных концентрических складок. Судя по этому признаку, мы имеем дело с раковиной взрослого экземпляра.
Сравнение. В публикациях отсутствуют взрослые раковины столь малого размера. Учитывая сказанное, не представляется возможным провести сравнительный анализ с какими-либо видами в роде Retroceramus.
Местонахождение: р. Дрещанка, обн. 25, сл. 1a.
Геологический возраст и географическое распространение. Тимано-Печорский регион; верхний байос, зона Arctocephalites arcticus.
Retroceramus (R.) cf. eximius (von Eichwald, 1871)
Табл. III, фиг. 2–3
Материал. Два обломка левых створок (экз. № ГГМ Рц11–12).
Описание и таксономическая трактовка. Судя по сохранившимся обломкам, раковины имели средний для рода размер. Длина фрагмента, изображенного в табл. III, фиг. 3, составляет 70 мм по оси роста. О степени скошенности трудно судить; скорее всего, она не слишком значительная. Из диагностических признаков сохранилась лишь скульптура, представленная правильными плотно расположенными концентрическими складками, слегка приостренными на гребнях. Детальное описание морфологии складок на краях раковины, приведенное И.В. Полуботко (см. ниже), не противоречит признакам, наблюдаемым у вышеупомянутого фрагмента. На основе описанного типа складок рассматриваемые образцы можно сопоставить с R. eximius (von Eichw.) из байос-батских отложений побережья Охотского моря (Ефимова и др., 1968, с. 70, табл. 78 , фиг. 1–2).
Местонахождение: р. Дрещанка, обн. 26, из осыпи, верхи слоя 1; обн. 12 (D-5), 45–50 см выше подошвы слоя 4а.
Геологический возраст и географическое распространение. Тимано-Печорский регион, бассейн р. Ижма, пограничные отложения верхнего байоса и бата.
Retroceramus (R.) sp. indet.
Табл. III, фиг. 4
Материал. Почти полностью сохранившаяся левая створка (экз. № ГГМ Рц13).
Описание. Судя по отпечатку, раковина представлена левой створкой, умеренно скошенной (угол скошенности около 40о) и умеренно выпуклой по линии наибольшего роста в ее средней части. Концентрические складки сглаженные, слабо выраженные в рельефе и проявленные лишь в задне-нижней части. Внешних признаков недостаточно для идентификации вида даже в открытой номенклатуре.
Местонахождение: р. Дрещанка, обн. 12 (D-5), средняя часть слоя 8.
Геологический возраст. Нижний бат, зона Arcticoceras ishmae.
ОТРЯД UNIONOIDA STOLICZKA, 1871
НАДСЕМЕЙСТВО UNIONOIDEA RAFINESQUE, 1820
СЕМЕЙСТВО UNIONIDAE RAFINESQUE, 1820
Род Mezounio Zakharov, gen. nov.
Название рода от комб. лат. mezo- – средний и Unio – родового названия современных двустворчатых моллюсков.
Типовой вид – Mezounio zverkovi Zakharov, gen. et sp. nov.
Видовой состав: только типовой вид.
Диагноз. Раковина среднего размера, равностворчатая, существенно неравносторонняя, сильно удлиненная, овальная; длина превышает высоту до 4 раз. Верхний и нижний края не строго параллельны, поскольку плавно изгибаются наружу. Раковина постепенно сужается по направлению и к переднему, и к заднему краю. Передний край обычно более заужен и иногда заострен. Макушки маленькие, необособленные, слабо выступающие над замочным краем и на треть длины створки приближены кпереди. Концентрические линии нарастания очень тонкие, часто расположенные и равномерно покрывают обе створки. Ядра гладкие. Кардинальные (псевдокардинальные?) зубы не наблюдались. Имеются два латеральных зуба на левой створке и единственный латеральный – на правой створке. Под макушкой заметна неглубокая, плохо очерченная (лигаментная?) ямка.
Сравнение. В международном справочнике по двустворкам (Treatise…, 1969) среди семейства Unionidae и одноименного подсемейства Unioninae можно обнаружить немалое число родов, напоминающих по форме раковины Mezounio gen. nov. Эти роды большей частью происходят из современных и кайнозойских отложений в основном Юго-Восточной Азии: Вьетнама, Китая, Японии. Не имея возможности провести анализ всех родовых таксонов, да и будучи ограниченными материалом лишь из одного местонахождения, рассмотрим наиболее часто упоминаемые в публикациях роды, отмечавшиеся ранее в мезозойских, в частности среднеюрских, отложениях.
Род Cuneopsis Simpson, 1900 (Treatise…, 1969, p. N421, fig. D20 (2)) по очертаниям створок, скульптуре и суженному заднему концу, иногда встречающемуся у наших экземпляров, а также по характеру латеральных зубов и расположению отпечатков мускулов-замыкателей похож на описываемый род, отличаясь местоположением макушек, на ¼ длины створки отодвинутых от конца переднего края, отсутствием псевдокардинальных зубов и наличием небольшой слегка утопленной (?резилиферной) ямки под макушкой. Род Cuneopsis был идентифицирован в составе многочисленных, сведенных авторами в синонимику видов из среднеюрских отложений Южного Китая (Pan, Sha, 2009). Однако это заключение было поставлено под сомнение С. Шнайдером, поскольку исследованный китайскими авторами материал имеет плохую сохранность, не позволяющую судить о характере замка (Schneider et al., 2013). С. Шнайдер, основываясь на мнении К. Сузуки (Suzuki, 1949) и результатах исследования минералогии юрских и современных раковин, проведенных Джин-уа Ченом (Chen, 1987), заявил, что нет прямых генетических связей между мезозойскими и кайнозойскими унионидами из Восточной Азии. Учитывая существенные различия в строении раковинного слоя у юрских и современных Unionidae, Чен (Chen, 1987) ранее предложил для юрских таксонов новое семейство Qiyangiidae Chen, 1983 и рекомендовал использовать названия существующих юрских родов юнионид только в кавычках. Таким образом, среди специалистов отсутствует единое мнение о систематическом составе ископаемых Unionidae, притом не только мезозойских, но и более молодых.
Сходство с Mezounio gen. nov. обнаруживают также представители рода Lanceolaria Conrad, 1853, известного с плиоцена и обитающего ныне в Восточной Азии (Treatise…, 1969, p. N424, fig. D23 (1)). Недавно этот род по одной левой створке был установлен в палеогене Северного Вьетнама (Schneider et al., 2013, p. 8, fig. 5A). По сильно удлиненной ланцетовидной раковине (L/H = 3.5), ее закругленным концам, приближенной к переднему краю слегка прозогирной макушке, отсутствию следов аллометрического роста некоторые из более чем 100 экземпляров Mezounio gen. nov. сходны с палеогеновыми Lanceolaria. Однако детали внутреннего строения Mezounio gen. nov. остаются неизвестными, поскольку нам не удалось отпрепарировать замок. Следовательно, полноценное сравнение рассматриваемых таксонов на данный момент не может быть осуществлено.
Некоторые современные виды рода Unio также обнаруживают сходство с описываемым таксоном. Например, Unio с территорий Восточной Европы (верхнее и среднее течение р. Дунай), западной части Европейской России и Украины по контуру раковины, характеру закругления передней и задней ее частей, параллельным вентральным и дорзальным краям очень напоминают многие экземпляры из нашей коллекции (Nesemann, 1994; Klishko et al., 2017, p. 103, figs. 4A, 4B, 4C). Внутренние структуры – латеральные зубы и их расположение, отпечатки обоих мускулов, местоположение макушки и разнообразие псевдокардинальных зубов – имеют сходство с ископаемыми формами. Существенные различия заключаются в гораздо меньших размерах раковин и пропорциях длина/высота современных видов. Поскольку выборки современных экземпляров могут быть значительными, они позволяют наблюдать широкую изменчивость всех признаков.
Поиски возможных ближайших предков среднеюрского рода приводят к триасовым унионидам Северо-Восточной Азии и Японии. По форме раковины, ее габаритам и очертанию Mezounio gen. nov. напоминает Palaeopharus Kittl, 1907 из норийского яруса, но у первого отсутствуют псевдокардинальные зубы, на месте которых имеется мелкое углубление, и характерные для Palaeopharus радиальные ребра в скульптуре (Treatise…, 1969; Курушин, 1992). Ранее Т. Кобаяши и К. Ичикава (Kobayashi, Ichikawa, 1951) подчеркивали сходство псевдокардинального зуба у Unio и Palaeopharus. Однако, по мнению А. Токуяма (Tokuyama, 1958, p. 293), сходство может быть поверхностным, потому что псевдокардинальный зуб у унионид “в действительности является своего рода кардинальным зубом, тогда как у Palaeopharus он образуется из переднего бокового зуба”. Указанный автор считает, что “присутствие или отсутствие “псевдокардинального зуба” не так важно для классификации пелеципод”. На этом основании А. Токуяма (Tokuyama, 1958, p. 293) полагает, что род Palaeopharus следует отделять от группы настоящих унионид, которая включает типовой род Unio и, возможно, является потомком другого триасового рода – Myophoria Bronn in von Alberti, 1834. Тем не менее род Palaeopharus наряду c другими позднетриасовыми таксонами – Cardinioides, Pachycardia, Trigonodus, Unionites – в более поздних сводках включается в состав отряда Unionoida (Hallam, 1981).
Геологический возраст и географическое распространение. Север Европейской России, Тимано-Печорский регион, нижний? байос.
Mezounio zverkovi Zakharov, sp. nov.
Название вида в честь Н.Г. Зверькова, нашедшего массовое скопление раковин этого вида в разрезах средней юры на р. Ижма в 2018 г.
Голотип: экз. № ГГМ Un3, целая раковина; р. Ижма, обн. 19.
Материал. Более 100 преимущественно разрозненных створок и крупных фрагментов с частично сохранившимися раковинами (в том числе экз. № ГГМ Un1–8).
Описание. Раковина равностворчатая, существенно неравносторонняя, сильно удлиненная, овальная. У наиболее крупных форм длина превышает высоту иногда в 4 раза. Вентральный и дорзальный края плавно изгибаются во внешнюю сторону, поэтому строго не параллельны. Передний край более заужен и иногда заострен. Макушки маленькие, необособленные, приближены кпереди на треть длины створки. Тонкие, часто расположенные концентрические линии равномерно покрывают обе створки. На ядрах они не видны. Отпечаток мускула-замыкателя, иногда каплевидно очерченный, расположен впереди макушки и приближен к переднему краю.
Имеется один (возможно, два) латеральных зуба на левой створке и единственный латеральный – на правой створке. Кардинальные зубы не наблюдались. Под макушкой имеется неглубокая плохо очерченная ямка, возможно, для размещения лигаментной (?) связки.
Местонахождение: р. Дрещанка, обн. 14, основание слоя 1, обн. 29'', 29'; р. Ижма, обн. 19 и 24 (сл. 7).
Геологический возраст и географическое распространение. Нижний?–верхний байос, Тимано-Печорский регион (басс. р. Ижма).
ОБСУЖДЕНИЕ
Зональная шкала по ретроцерамам: принципы построения и перспективы совершенствования
Казалось бы, постановка такого вопроса некорректна, поскольку ранее уже была предложена зональная шкала, основанная на последовательностях видов рода Retroceramus, для Омолонского массива (Кошелкина, 1980а, 1980б). В публикациях текущего столетия она же представлена в актуализированном виде (Шурыгин, 2005; Меледина и др., 2011; Никитенко и др., 2013). Тем не менее с момента выделения горизонтов и слоев с Retroceramus в начале 1960-х и 1970-х годов (Полуботко, 1972а, 1972б; Кошелкина, 1974) [в публикациях род часто именовался Mytiloceramus] и до настоящего времени нет публикаций, в которых бы на единых разрезах были выделены и описаны последовательности слоев в ранге зон с соблюдением необходимых требований. Если речь идет о зонах, требуется установить, по крайней мере, смыкаемость биостратонов в едином разрезе (Стратиграфический…, 2019, с. 34).
В 1960-х и 1970-х годах опубликованы десятки статей и монографий с описанием новых видов рода Retroceramus из наиболее известных разрезов средней юры Вилюйской синеклизы, Приверхоянского краевого прогиба (Кошелкина, 1963), Северо-Востока СССР (побережье Охотского моря, западное и восточное Приколымье и Корякское нагорье) (Кошелкина, 1969а; Полуботко, 1972а, 1972б), Дальнего Востока (Сей, Калачева, 1980) и других районов. Согласно подсчетам И.В. Полуботко (1972б), “общее количество видов, встречающихся в средней юре региона [Северо-Восток СССР – примечание авторов], достигло цифры 90” (!). Геологический возраст многих видов был установлен благодаря их совместным находкам с аммонитами. Таким образом, уже в то время стало очевидным биостратиграфическое значение ретроцерамов. Однако И.В. Полуботко (1972б) лишь перечисляются виды ретроцерамов, тогда как формальные биостратоны по ним не были зафиксированы (Кошелкина, 1969б). Идеи о выделении биостратонов в ранге слоев с видами Retroceramus к началу 1970-х годов, так сказать, “витали в воздухе”.
По-видимому, впервые формальные биостратоны по ретроцерамам в ранге слоев были выделены для среднеюрских отложений Северо-Востока СССР и смежных территорий зарубежной части Бореальной области З.В. Кошелкиной (1970). Виды-индексы этих слоев, их стратиграфические объемы и, конечно, возрастные датировки позднее были существенно скорректированы. Почти одновременно с З.В. Кошелкиной серьезный анализ последовательностей комплексов среднеюрских ретроцерамов по материалам с Северо-Востока СССР провела И.В. Полуботко (1972б). На приведенной ею схеме показаны виды-индексы ретроцерамов для всех подъярусов средней юры. С современных позиций привлекает внимание то, что “горизонты с иноцерамами” были выделены автором автономно, независимо от “последовательностей аммонитов”, хотя геологический возраст “горизонтов” определялся по совместным находкам видов-индексов ретроцерамов с аммонитами, часто в географически удаленных разрезах. Оценка “качества” слоев, выделенных в то время, с позиций классических зональных шкал по ортостратиграфическим группам, не имеет смысла, прежде всего, из-за существенной разницы в темпах морфогенеза аммонитов и ретроцерамов. Кроме того, ретроцерамы нуждались в подходящем типе грунта для прикрепления личинки и, таким образом, зависели от фаций. Наконец, географические ареалы некоторых (многих?) из них контролировались климатически. На эти факторы среды справедливо обратила внимание И.В. Полуботко (1972б). Тем не менее, сравнивая последовательности слоев с ретроцерамами в публикациях начала 1970-х годов у разных авторов, нетрудно видеть их значительное сходство в выборе видов-индексов. Например, слои с jurensis, lucifer, elongatus, kystatymensis, retrorsus в приведенной последовательности даны З.В. Кошелкиной (1970), а слои с popovi, elegans, jurensis, menneri, lucifer, elongatus, kystatymensis, bulunensis в указанной последовательности предложены И.В. Полуботко (1972б). Со временем становилось все более и более очевидным, что ретроцерамы могут иметь важное значение не только для региональной, но и для межрегиональной корреляции. И.В. Полуботко и И.И. Сей предприняли совместную попытку осуществить корреляцию разрезов средней юры азиатской части современной России по ретроцерамам. Им удалось проследить отдельные ретроцерамовые стратоны в пределах Северо-Востока и Дальнего Востока СССР (Полуботко, Сей, 1981). Это был первый шаг к признанию слоев как зон. Такой шаг был сделан Б.Н. Шурыгиным по материалам изучения разрезов средней юры на севере Сибири (Меледина и др., 1987; Шурыгин, 1989). Важно заметить, что выделение зон по ретроцерамидам (b-зон) было осуществлено на наиболее богатом аммонитами разрезе средней юры на полуострове Юрюнг-Тумус, что позволило существенно уточнить объемы b-зон по аммонитам. В то же время следует согласиться с мнением Б.Н. Шурыгина о том, что “подход к выделению биостратонов [по ретроцерамам – примечание авторов] различен: границы устанавливаются (и) по появлению вида-индекса, (и) по эпиболе вида-индекса, (и) по зоне совместного распространения двух-трех видов” (Шурыгин, 1989, с. 76).
Таким образом, ретроцерамовая шкала, как и все биостратиграфические шкалы начиная с XIX века, формируется опытным путем. За время, прошедшее после первых попыток создания шкалы, она детализировалась с помощью включения новых уровней, замены мало информативных видов другими, уточнения стратиграфического объема зон путем перемещения их границ, часто следом за изменением границ зон по аммонитам. Многие из первоначально установленных слоев, обычно в исходной последовательности, можно обнаружить и в современной версии зональной шкалы по ретроцерамам. Несмотря на то, что указанным “путем проб и ошибок” зональная шкала по ретроцерамам постепенно совершенствуется, она непременно станет более “стерильной” после ревизии таксономического состава. В настоящее время в составе рода Retroceramus (s.l.) насчитывается около 120–130 видов. Число их в результате ревизии должно вдвое, если не втрое, сократиться. Такой сценарий, и вряд ли следует в этом сомневаться, лежит в основе совершенствования биостратиграфической шкалы по ретроцерамам в будущем.
К установлению в разрезах границ зон по парастратиграфическим группам не следует относиться так же строго, как к определению зональных границ по ортостратиграфическим группам. Первые не столь жестко фиксируются на основании первого появления вида в разрезе, поскольку нередко смена видов, характерных для смежных зон, происходит внутри некоего стратиграфического интервала с переходными формами. В таких ситуациях граница назначается либо по формальному признаку (например, по горизонту конкреций), либо строго посередине “переходного” интервала.
Оценка возраста изученных разрезов на реках Ижма и Дрещанка по ретроцерамам
Распределение аммонитов и двустворок в изученной последовательности показано на рис. 5. Поскольку в слоях, где были найдены наиболее древние ретроцерамы, аммониты не обнаружены, возраст вмещающих слоев может быть оценен только по самим ретроцерамам. Произведенная нами корреляция обнажений предполагает, что верхняя часть интервала с блинообразными конкрециями соотносится с аммонитовой зоной Arctocephalites arcticus верхнего байоса (примерно на этот уровень экстраполируются находки из разреза 14 (~D-3)). Обнажения 19, 29' и 29'' соответствуют нижней и средней частям интервала с блинообразными конкрециями и, таким образом, непосредственно подстилают интервал, охарактеризованный находками аммонитов A. arcticus.
Учитывая запутанную и местами противоречивую ситуацию с классификацией ретроцерамов (см. выше), определение возраста вмещающих толщ по ним может быть сделано с известной долей осторожности. По-видимому, нужно говорить о возможном раннебайосском возрасте безаммонитовых слоев, подстилающих интервал с находками A. arcticus. О близости к указанному возрасту свидетельствуют находки в самых нижних слоях изученной последовательности форм, ранее отмечавшихся именно в нижнем байосе Северо-Запада Азии и восточной части Сибири: R. (R.) elongatus и R. (R.) cf. lungershauseni. С другой стороны, тот факт, что R. (R.) elongatus в обнажениях 29' и 29'' встречен совместно с Mezounio zverkovi, а последний, в свою очередь, отмечен в разрезе 14 в интервале с аммонитами Arctocephalites arcticus, свидетельствует о близости наиболее древнего комплекса изученных двустворок и к верхнему байосу.
Историко-геологическое значение находок унионид на Ижме
В северных районах Западной Сибири в керне скважин, вскрывающих переходные слои между лайдинским и вымским горизонтами (верхний аален), а также вымский горизонт (верхний аален–нижний байос), совместно с морскими двустворками Arctotis lenaensis (Lahusen, 1886) и Arctica humiliculminata Schurygin in Zakharov et Schurygin, 1978 упоминаются находки “Unionidae (?)”, которые южнее становятся довольно многочисленными (Шурыгин и др., 2000). В кровле вымской толщи расположен пласт угля У10 – хорошо прослеженный маркирующий уровень юга Западной Сибири, который восточнее относится к верхам верхнего аалена–низам нижнего байоса (Шурыгин и др., 2000, с. 133–134). Скопления раковин Unionidae, встреченных в обн. 19 на р. Ижма совместно с Retroceramus (R.) alliensis и R. (R.) ex gr. porrectus в породе с обильными остатками обугленной древесины, возможно, фиксируют следы события, сходного с северосибирским и связанного с начальной фазой трансгрессии среднеюрского моря на территорию Тимано-Печорского региона. Скопление унионид на отдельной плоскости напластования в нижней части разреза (обнажение 19) свидетельствует об их весьма кратковременном существовании в условиях крайнего (прибрежного) мелководья.
ЗАКЛЮЧЕНИЕ
В нижней части морской терригенной толщи среднеюрского возраста Тимано-Печорского региона впервые детально описаны двустворчатые моллюски Retroceramus (Retroceramidae) и Mezounio Zakharov, gen. nov. (Unionidae). R. (R.) cf. lungershauseni Kosh., 1962 и R. (R.) elongatus Kosh., 1960, найденные в последовательности значительно ниже находок аммонитов Arctocephalites arcticus, свидетельствуют о возможном раннебайосском возрасте вмещающих пород. Такая датировка несколько удревняет время начала бореальной морской среднеюрской трансгрессии в регионе и, таким образом, открывает перспективу уточнения палеогеографических реконструкций Печорского моря для среднеюрского времени и восстановления его палеобиогеографических связей через сибирские акватории с бореально-пацифическими бассейнами.
Благодарности. Авторы искренне благодарны рецензентам И.В. Полуботко и Е.М. Тесаковой за конструктивные отзывы, способствовавшие повышению качества статьи. Некоторые важные замечания к рукописи были сделаны М.А. Роговым и В.С. Вишневской, которым мы также выражаем глубокую признательность.
Источники финансирования. Работа выполнена при финансовой поддержке Программы фундаментальных исследований Президиума РАН № 8 (координатор академик РАН Н.С. Бортников), проекта РФФИ 18-05-01070_А, а также по теме госзадания № 0135-2018-0035 ГИН РАН.
Список литературы
Бодылевский В.И. [Бассейн р. Печоры] // Атлас руководящих форм ископаемых фаун СССР. Т. IX. Верхняя юра. М.: ГОНТИ, 1949. С. 28–31.
Бодылевский В.И. Юрская система // Геология СССР. Том II. Архангельская, Вологодская области и Коми АССР. Часть I. Геологическое описание. М.: Гос. научно-техническое изд-во литературы по геологии и охране недр, 1963. С. 631–666.
Ефимова А.Ф., Кинасов В.П., Паракецов К.В., Полуботко И.В., Репин Ю.С., Дагис А.С. Полевой атлас юрской фауны и флоры Северо-Востока СССР. Магадан: Магаданское книжное изд-во, 1968. 379 с.
Захаров В.А., Шурыгин Б.Н. Биогеография, фации и стратиграфия средней юры Советской Арктики (по двустворчатым моллюскам). Новосибирск: Наука, 1978 (Труды Института геологии и геофизики. Вып. 352).
Захаров В.А., Богомолов Ю.И., Ильина В.И., Константинов А.Г., Курушин Н.И., Лебедева Н.К., Меледина С.В., Никитенко Б.Л., Соболев Е.С., Шурыгин Б.Н. Бореальный зональный стандарт и биостратиграфия мезозоя Сибири // Геология и геофизика. 1997. Т. 38. № 5. С. 927–956.
Ипполитов А.П., Киселев Д.Н., Зверьков Н.Г. О положении границы байоса и бата в опорных разрезах р. Ижмы (Тимано-Печорская область) // ПАЛЕОСТРАТ-2019. Годичное собрание (научная конференция) секции палеонтологии МОИП и Московского отделения Палеонтологического общества при РАН. Москва, 28–30 января 2019 г. Программа и тезисы докладов. Ред. Алексеев А.С., Назарова В.М. М.: Палеонтологический ин-т им. А.А. Борисяка РАН, 2019. С. 32–33.
Киселев Д.Н. Изменчивость скорости морфогенеза и проявление модулярности развития у юрских аммонитов рода Arcticoceras Spath // Палеонтол. журн. 2020а. № 1. С. 20–32.
Киселев Д.Н. Род Аrcticoceras Spath, 1924 (Аmmonoidea) и инфразональная биостратиграфия нижнего бата Бореальной области // Стратиграфия. Геол. корреляция. 2020б. № 2. С. 77–102.
Кошелкина З.В. Палеонтологическое обоснование ярусного расчленения морских юрских отложений Вилюйской впадины и Приверхоянского краевого прогиба // Труды Межведомственного совещания по разработке унифицированных стратиграфических схем Сибири 1956 г. Доклады по стратиграфии мезозойских и кайнозойских отложений. Л.: Гостоптехиздат, 1957. С. 38–45.
Кошелкина З.В. Новые среднеюрские иноцерамы Северной Сибири // Новые виды древних растений и беспозвоночных СССР. Ч. 2. М.: Госгеолтехиздат, 1960. С. 36–38.
Кошелкина З.В. Иноцерамы и их стратиграфическое значение для расчленения юрских отложений Сибири // Тр. Всесоюзного аэрогеологического треста. 1961. Вып. 7. С. 140–166.
Кошелкина З.В. Полевой атлас руководящих фаун юрских отложений Вилюйской синеклизы и Приверхоянского краевого прогиба. Магадан: Магаданское книжное изд-во, 1962. 133 с.
Кошелкина З.В. Стратиграфия и двустворчатые моллюски юрских отложений Вилюйской синеклизы и Приверхоянского краевого прогиба // Тр. СВКНИИ Сибирского отделения АН СССР. 1963. Вып. 5. 218 с.
Кошелкина З.В. Иноцерамы юры Охотского побережья, Западного и Восточного Приколымья и Корякского нагорья // Иноцерамы юры и мела Северо-Востока СССР. Магадан: Изд-во СВКНИИ, 1969а. С. 14–117 (Тр. СВКНИИ. Вып. 32).
Кошелкина З.В. Новые данные по стратиграфии юрских отложений бассейна реки Анадырь // Иноцерамы юры и мела Северо-Востока СССР. Магадан: Изд-во СВКНИИ, 1969б. С. 173–189 (Тр. СВКНИИ. Вып. 32).
Кошелкина З.В. Среднеюрские отложения Северо-Востока СССР, смежных территорий зарубежной части Бореальной области // Палеомагнитная и биостратиграфическая характеристика некоторых опорных разрезов мезозоя и кайнозоя севера Дальнего Востока. Магадан: Изд-во СВКНИИ, 1970. С. 157–175 (Тр. СВКНИИ. Вып. 37).
Кошелкина З.В. К вопросу о номенклатуре и систематике юрских ретроцерамид // Колыма. 1971. № 5. С. 43–45.
Кошелкина З.В. Региональная схема стратиграфии средней юры Северо-Восточной Сибири по ретроцерамидам // Основные проблемы биостратиграфии и палеогеографии Северо-Востока СССР: Мезозой. Том 2, часть 2. Магадан: Изд-во СВКНИИИ ДВНЦ АН СССР, 1974. С. 90–181 (Тр. СВКНИИ. Вып. 63).
Кошелкина З.В. Средняя юра Гижигинской структурно-фациальной зоны (ааленский ярус) // Биостратиграфия и корреляция мезозойских отложений Северо-Востока СССР. Магадан: Изд-во СВКНИИ ДВНЦ АН СССР, 1980а. С. 62–75.
Кошелкина З.В. Корреляция среднеюрских отложений некоторых регионов бореального пояса (Омолонский массив) и анализ комплексов // Биостратиграфия и корреляция мезозойских отложений Северо-Востока СССР. Магадан: Изд-во СВКНИИ ДВНЦ АН СССР, 1980б. С. 76–90.
Кравец В.С., Месежников М.С., Слонимский Г.А. Строение юрско-нижнемеловой толщи в бассейне р. Печоры // Биостратиграфия отложений мезозоя нефтегазоносных областей СССР. Л.: Изд-во ВНИГРИ, 1976. С. 27–41 (Тр. ВНИГРИ. Вып. 338).
Курушин Н.И. Триасовые палеогетеродонтные и гетеродонтные двустворки Сибири. Новосибирск: Наука, Сиб. отд., 1992 (Тр. ИГиГ. Вып. 742).
Леонова М.Н. Государственная геологическая карта. Лист P-39-VI. Масштаб 1 : 200 000. 1964 г.
Меледина С.В. Бореальная средняя юра России // Тр. ИГиГ СО РАН. 1994. Вып. 819. 184 с.
Меледина С.В. О корреляции зон байоса и бата Сибири в свете новых палеонтологических данных // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 6. С. 45–56. https://doi.org/10.7868/S0869592X14060040
Меледина С.В., Нальняева Т.И., Шурыгин Б.Н. Юра Енисей-Хатангского прогиба. Нордвикская зона, типовой разрез. Новосибирск: Институт геологии и геофизики СО АН СССР, 1987. 127 с.
Меледина С.В., Шурыгин Б.Н., Злобина О.Н., Левчук М.А., Нальняева Т.И., Никитенко Б.Л. Чекуровская свита (бат-келловей) в стратотипе // Детальная стратиграфия и палеонтология юры и мела Сибири. Новосибирск: Наука, 1991. С. 5–37 (Тр. ИГиГ СО АН СССР. Вып. 769).
Меледина С.В., Никитенко Б.Л., Шурыгин Б.Н., Дзюба О.С., Князев В.Г. Современная комплексная зональная шкала юры Сибири и бореальный стандарт // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2011. Т. 52. Вып. 16–17. С. 17–40.
Митта В.В. Первая находка Arctocephalites (Cardioceratidae, Ammonoidea) в средней юре бассейна Печоры // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, эволюция и биостратиграфия. Материалы Всероссийского совещания (Москва, 8–10 ноября 2006 г.). Ред. Барсков И.С., Леонова Т.Б. М.: ПИН РАН, 2006. С. 82–84.
Митта В.В. Верхний байос и нижний бат бассейна Печоры и бореально-тетическая корреляция // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 1. С. 77–87.
Митта В.В., Сельцер В.Б. Первые находки Arctocephalitinae (Ammonoidea) в юре юго-востока Русской платформы и корреляция бореального батского яруса со стандартной шкалой // Тр. НИИ геологии СГУ. Нов. сер. 2002. Т. X. С. 12–39.
Митта В.В., Захаров В.А., Барсков И.С., Сельцер В.Б., Иванов А.И. Верхний байос и нижний бат окрестностей Саратова: малакологическая характеристика и биостратиграфия // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 5. С. 32–45.
Митта В.В., Костылева В.В., Глинских Л.А. О байосе и бате (средняя юра) бассейна р. Ижма // Проблемы геологии Европейской России. Сборник научных трудов Всероссийской научной конференции, посвященной 130-летию со дня рождения профессора Бориса Александровича Можаровского. Саратов, 11–14 сентября 2013 г. Саратов: СГТУ, 2013. С. 93–103.
Митта В.В., Глинских Л.А., Костылева В.В. Предварительный отчет по полевым работам 2013 г. в бассейнах рек Сысола и Ижма // Геология и минеральные ресурсы Европейского Северо-Востока России. Материалы XVI Геологического съезда Республики Коми. Т. 2. Сыктывкар: Геопринт, 2014а. С. 188–190.
Митта В.В., Костылева В.В., Глинских Л.А., Шурыгин Б.Н., Стародубцева И.А. Стратиграфия средней юры юго-запада Республики Татарстан // Стратиграфия. Геол. корреляция. 2014б. Т. 22. № 1. С. 31–46. https://doi.org/10.7868/80869592X14010049
Невесская Л.А., Попов С.В., Гончарова И.А., Гужов А.В., Янин Б.Т., Полуботко И.В., Бяков А.С., Гаврилова В.А. Двустворчатые моллюски России и сопредельных стран в фанерозое // Тр. ПИН РАН. 2013. Т. 294. 524 с.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г., Меледина С.В., Дзюба О.С., Лебедева Н.К., Пещевицкая Е.Б., Глинских Л.А., Горячева А.А., Хафаева С.Н. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный зональный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082. https://doi.org/10.1016/j.rgg.2013.07.005
Полуботко И.В. О стратиграфическом значении иноцерамов из группы Inoceramus lucifer Eichwald в средней юре Северо-Востока СССР // Тр. Всесоюзного коллоквиума по иноцерамам. Вып. 1. М.: ВИНИТИ, 1972а. С. 15–24.
Полуботко И.В. К вопросу об объеме видов среднеюрских иноцерамов Северо-Востока СССР // Тр. Всесоюзного коллоквиума по иноцерамам. Вып. 1. М.: ВИНИТИ, 1972б. С. 38–50.
Полуботко И.В. Иноцерамовые двустворки нижней и средней юры Северо-Востока СССР и севера Сибири // Атлас руководящих групп фауны мезозоя Юга и Востока СССР. СПб.: Недра, 1992. С. 56–79.
Полуботко И.В., Сей И.И. Расчленение среднеюрских отложений восточной части СССР по митилоцерамам // Изв. АН СССР. Сер. геол. 1981. № 12. С. 63–70.
Пчелинцева Г.Т. Стратиграфия и фауна пластинчатожаберных Западного Приохотья // Труды Геол. музея им. Карпинского АН СССР. 1962. Вып. IX. 88 с.
Репин Ю.С. Новые данные по стратиграфии юры Печорской впадины // Бюлл. МОИП. Отд. геол. 2005. Т. 80. Вып. 3. С. 17–25.
Репин Ю.С., Захаров В.А., Меледина С.В., Нальняева Т.И. Атлас моллюсков Печорской юры. СПб.: Недра, 2006. 262 с.
Сазонов Н.Т. Юрские отложения Центральных областей Русской платформы. Л.: Гостоптехиздат, 1957.
Сей И.И. Представители раннеааленских иноцерамид Западного Приохотья (Дальний Восток) // Записки Ленинградского горного ин-та. 1972. Т. LXIII. Вып. 2. С. 120–130.
Сей И.И., Калачева Е.Д. Биостратиграфия нижне- и среднеюрских отложений Дальнего Востока // Труды ВСЕГЕИ. Новая серия. 1980. Т. 285. 187 с.
Сей И.И., Калачева Е.Д. Граница батского и келловейского ярусов в бореальных и суббореальных районах России // Отечественная геология. 1992. № 10. С. 47–52.
Соколов Д.Н. К аммонитовой фауне Печорской юры // Тр. Геол. Ком. Нов. Сер. 1912. Вып. 76. С. i, 1–65, 3 табл.
Стратиграфический кодекс России. Издание третье, исправленное и дополненное. СПб.: Изд-во ВСЕГЕИ, 2019.
Чирва С.А., Куликова Н.К. Сопоставление среднеюрских–келловейских песчаных отложений бассейнов рек Сысолы, Яренги и Печоры // Юрские отложения Русской платформы (сборник научных трудов). Л.: ВНИГРИ, 1986. С. 87–102.
Чирва С.А., Яковлева С.П. Строение и биостратиграфия пограничных слоев континентальной и морской юры Тимано-Уральской области // Стратиграфия триасовых и юрских отложений нефтегазоносных бассейнов СССР (сборник научных трудов). Л.: ВНИГРИ, 1982. С. 57–65.
Шурыгин Б.Н. Проблемы создания зональных шкал юры на севере Сибири по двустворкам // Ярусные и зональные шкалы бореального мезозоя СССР. М.: Наука, 1989. С. 74–82 (Тр. ИГиГ СО АН СССР. Вып. 722).
Шурыгин Б.Н. Биогеография, фации и стратиграфия нижней и средней юры Сибири по двустворчатым моллюскам. Новосибирск: ГЕО, 2005. 154 с.
Шурыгин Б.Н., Никитенко Б.Л., Девятов В.П., Ильина В.И., Меледина С.В., Гайдебурова Е.А., Дзюба О.С., Казаков А.М., Могучева Н.К. Стратиграфия нефтегазоносных бассейнов Сибири. Юрская система. Новосибирск: Изд-во СО РАН, филиал “ГЕО”, 2000. 480 с.
Alberti F., von. Beiträge zu einer Monographie des bunten Sandsteins, Muschelkalkes und Keupers und die Verbindung dieser Gebilde zu einer Formation (Trias). Stuttgart: Cotta, 1834. 366 s.
Callomon J.H. The evolution of the Jurassic ammonite family Cardioceratidae // Palaeontology. Spec. Pap. 1985. № 33. P. 49–90.
Chen J. Microstructures of Lower–Middle Jurassic unionid (Bivalvia) shells // Acta Palaeontologica Sinica. 1987. V. 26. P. 8–17.
Conrad T.A. Synopsis of the family of Naiades of North America; with notes and a table of some of the genera and subgenera // Proc. Acad. Nat. Sci. Philadelphia. 1853. V. 6. P. 243–269, 449.
Eichwald E., von. Geognostisch-palaeontologische Bemerkungen über die Halbinsel Mangischlak und die Aleutischen Inseln. St. Petersburg: Kaiserlichen Akademie der Wissenschaften, 1871. https://doi.org/10.5962/bhl.title.15765
Giebel C.G. Allgemeine Palaeontologie. Entwurf einer systematischen Darstellung der Fauna und Flora der Vorwelt. (Abtheilung 1: Palaeozoologie). Edition 2. Leipzig: Ambrosius Abel, 1852. viii + 414 p.
Hallam A. The end-Triassic bivalve extinction event // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1981. V. 35. P. 1–44. https://doi.org/10.1016/0031-0182(81)90092-4
Keyserling A. Geognostische Beobachtungen // Wissenschaftliche Beobachtungen auf Einer Riese in das Petschora-Land, im Jahre 1843. St. Petersburg: C. Kray, 1846. P. 149–406.
Kittl E. Die Triasfossilien von Heureka sund // Report of the Sec. Norwegian Arctic Expedition in the “Fram” 1898–1902. 1907. V. II. № 7. P. 1–44.
Klishko O., Lopes-Lima M., Froufe E., Bogan A., Vasilieva L., Yanovich L. Taxonomic reassessment of the freshwater mussel genus Unio (Bivalvia: Unionidae) in Russia and Ukraine based on morphological and molecular data // Zootaxa. 2017. V. 4286. № 1. P. 93–112. https://doi.org/10.11646/zootaxa.4286.1.4
Kobayashi T., Ichikawa K. On Palaeopharus, a Late Triassic pelecypod genus // Trans. Proc. Palaeont. Soc. Japan, n.s. 1951. Iss. 1. P. 7–12. https://doi.org/10.14825/prpsj1951.1951.1_7
Lahusen I.I. Die Inoceramen-Schichten an dem Olenek und der Lena // Mém. Acad. St.-Petérsb. VII sér. 1886. T. 33. № 7. P. 1–13.
Mitta V., Kostyleva V., Dzyuba O., Glinskikh L., Shurygin B., Seltzer V., Ivanov A., Urman O. Biostratigraphy and sedimentary settings of the Upper Bajocian–Lower Bathonian in the vicinity of Saratov (Central Russia) // N. Jb. Geol. Paläont. Abh. 2014. V. 271. Nr. 1. S. 95–121. https://doi.org/10.1127/0077-7749/2014/0380
Mitta V., Glinskikh L., Kostyleva V., Dzyuba O., Shurygin B., Nikitenko B. Biostratigraphy and sedimentary settings of the Bajocian–Bathonian beds in the Ishma River basin (European North of Russia) // N. Jb. Geol. Paläont. Abh. 2015. V. 277. Nr. 3. S. 307–335. https://doi.org/10.1127/njgpa/2015/0507
Nesemann H. Die Subspezies von Unio crassus Philipsson 1788 im Einzugsgebiet der mittleren Donau (Mollusca: Bivalvia, Unionidae) // Lauterbornia. 1994. Hft. 15. S. 59–77.
Newell N.D. Classification of the Bivalvia // Am. Museum Novitates. 1965. № 2206. P. 1–25.
Pan Y.-H., Sha J.-G. Middle Jurassic unionids (non-marine Bivalvia) from the Shiwandashan Basin, southern China, with special emphasis on Cuneopsis Simpson // GFF. 2009. V. 131. № 1–2. P. 183–194. https://doi.org/10.1080/11035890902867456
Poulton T.P. Zonation and correlation of Middle Boreal Bathonian to Lower Callovian (Jurassic) ammonites, Salmon Cache Canyon, Porcupine river, Northern Yukon // Bull. Geol. Surv. Canada. 1987. № 358. P. 1–155.
Rafinesque C.S. Monographie des coquilles bivalves fluviatiles de la rivière Ohio, contenant douze genres et soixante-huit espèces // Ann. Gén. Sci. Phys. Bruxelles. 1820. V. 5. № 13. P. 287–322.
Rollier L. Fossiles nouveaux ou peu connus des terrains secondaires (mésozoiques) du Jura et des contrées environnantes. Partie 4 // Mém. Soc. paléont. Suisse. 1914. V. 40. P. 321–440.
Schneider S., Böhme M., Prieto J. Unionidae (Bivalvia; Palaeoheterodonta) from the Palaeogene of northern Vietnam: exploring the origins of the modern East Asian freshwater bivalve fauna // J. Systematic Palaeontology. 2013. V. 11. Iss. 3. P. 1–21. https://doi.org/10.1080/14772019.2012.665085
Simpson C.T. Synopsis of the naiades, or pearly fresh-water mussels // Proc. United States National Museum. 1900. V. 22. № 1205. P. 501–1044. https://doi.org/10.5479/si.00963801.22-1205.501
Sowerby J. On a fossil shell of a fibrous structure, the fragments of which occur abundantly in the Chalk Strata and in the flints accompanying it // Trans. Linn. Soc. London. 1822. V. 13. № 2. P. 453–458. https://doi.org/10.1111/j.1095-8339.1821.tb00067.x
Stoliczka F. The Pelecypoda, with a review of all known genera of this class, fossil and Recent // Paleontologia Indica. Mem. Geol. Surv. India. Ser. 6. 1870–1871. V. 3. P. i–xxii, 1–537, pl. 1–50 (P. i–xxii, 223–537, pl. 23–50 for 1871).
Suzuki K. Development of the fossil non-marine molluscan faunas in Eastern Asia // Jap. J. Geol. Geogr. 1949. V. 21. P. 91–133.
Tokuyama A. Late Triassic Palaeopharus in Japan // Trans. Proc. Palaeont. Soc. Japan. N. ser. 1958. Iss. 32. P. 291–298. https://doi.org/10.14825/prpsj1951.1958.32_291
Treatise on Invertebrate Paleontology. Part N. Volume 1 (of 3) Mollusca 6. Bivalvia. N.Y.: Geological Society of America and The University of Kansas, 1969. xxxviii + 1224 p.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция