

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 3, стр. 114-133
Микропалеонтологическая и литогеохимическая характеристика туртасской свиты (верхний олигоцен), Западная Сибирь
Т. В. Орешкина 1, *, Г. Н. Александрова 1, С. М. Ляпунов 1, П. В. Смирнов 2, Б. В. Ермолаев 1
1 Геологический институт РАН
Москва, Россия
2 Тюменский государственный университет
Тюмень, Россия
* E-mail: oreshkina@ginras.ru
Поступила в редакцию 18.12.2018
После доработки 15.02.2019
Принята к публикации 28.03.2019
Аннотация
Проведено комплексное изучение микрофоссилий и литогеохимических характеристик континентальных отложений туртасской свиты верхнего олигоцена Западной Сибири из скв. Зырянская-1 на юго-западе Тюменской области. Доминирование пресноводных планктонных диатомовых водорослей при подчиненном значении бентоса, а также присутствие гемуллосклер губок свидетельствуют о прибрежных обстановках пресноводного водоема c глубинами от 3 до 50 м. Изученный палинокомплекс аналогичен спорово-пыльцевым спектрам туртасской свиты Нижнего Приобья, но отличается от палинокомплексов этой свиты более южных районов Западной Сибири низким содержанием пыльцы Taxodiaceae/Cupressaceae, Betulaceae, отсутствием Fagaceae, а также цист пресноводных динофлагеллат Pseudokomewuia. Выявленные отличия можно объяснить как особенностями фациальных условий, так и различиями в объеме этой свиты в известных на сегодняшний день местонахождениях. Данные по микроэлементному составу проб, геохимические и петрохимические модули свидетельствуют о формировании туртасской свиты в условиях относительно теплого гумидного климата в прибрежных участках слабоминерализованного бассейна.
ВВЕДЕНИЕ
Туртасская свита была выделена С.Б. Шацким (Гурари и др., 1961) со стратотипом на р. Туртас ниже устья р. Кацис (Шацкий и др., 1973). Туртасская свита распространена на обширных площадях Западно-Сибирской низменности с глубинами залегания 50–100 м (Шацкий и др., 1973). Ее наиболее полные разрезы находятся на правобережье р. Обь севернее устья Иртыша и далее ниже по течению у д. Карымкары, где наблюдаются ее нижняя граница с нижнеолигоценовой новомихайловской свитой и верхняя граница с нижнемиоценовой абросимовской свитой. Свита сложена зеленовато-серыми глинами и алевролитами тонкослоистыми, с прослоями диатомитов, нередко с присутствием глауконита. Положение туртасской свиты в региональной стратиграфической схеме палеогеновых отложений Западной Сибири неоднократно пересматривалось. Первая унифицированная схема расчленения палеогеновых отложений Западной Сибири была разработана в 1956 г., дополнена и утверждена в 1960 г. и принята МСК в 1968 г. В окончательном варианте этой схемы туртасская свита отсутствует и заменяется на журавскую из-за разного понимания объема свиты. В схемах 1976 и 1983 гг., как и в схеме 2001 г. (Унифицированные…, 2001), туртасская свита входит в состав журавского горизонта. Согласно С.Б. Шацкому (1978), туртасская свита представляет собой более глубоководные фации центральных частей бассейна, журавская – прибрежные фации, развитые в Кулундинской и Барабинской степях, Омском и Павлодарском Прииртышье (Зальцман, 1968; Зыкин, 2012).
Туртасская свита по палинологическим данным сопоставляется с морскими образованиями позднего олигоцена (хатт) Северного Устюрта (нижнебайгубекский подгоризонт), охарактеризованных фауной и флорой (Панова, 1971).
Изучение диатомовых комплексов туртасской свиты впервые было проведено Н.В. Рубиной (рис. 1) по материалам 8 скважин и 17 обнажений (1962, 1967, 1968, 1973а, 1976; Диатомовые…, 1974). Обобщая эти данные, Н.В. Рубина установила для туртасской свиты стратиграфическую последовательность диатомовых комплексов. Комплекс нижнего туртаса, предположительно соотнесенный с периферической частью бассейна, представлен редкими ископаемыми планктонными Melosira, относящимися к группе prae-, и обильными переотложенными морскими эоценовыми видами. Следует отметить, что впоследствии большая часть пресноводных планктонных центрических видов диатомей, относящихся к роду Melosira, была переведена в род Aulacoseira (Simonsen, 1979). Второй комплекс, включающий преимущественно те же виды рода Melosira и обильные спикулы губок, фиксируется на всей территории бассейна и характерен для относительно грубозернистых пород. К более тонким осадкам – слоистым алевритам и алевритистым глинам – приурочен комплекс с Melosira группы prae-, разнообразными представителями пеннатного рода Tetracyclus и целым рядом описанных Н.В. Рубиной эндемичных видов. Среди них – Melosira ignota, тонкоареолированные Coscinodiscus, связанные переходными формами. Для последней ассоциации Н.В. Рубина прослеживает последовательность появления описанных ею эндемичных Coscinodiscus снизу вверх по разрезу, выделяя слои I–IV. Для слоя I характерен Coscinodiscus tener c сопутствующими Tetracyclus celatom Okuno и Eunotia aff. revoluta A. Cleve; слой II содержит переходные формы от Сoscinodiscus tener к С. trapeziformis, фиксируется первое появление Сoscinodiscus lobatus. Слой III характеризуется массовым количеством С. trapeziformis. В слое IV появляется типичный С. gorbunovii наряду с присутствием всех вышеперечисленных Coscinodiscus за исключением С. tener. Позже Н.В. Рубина (1973б) описала ряд разновидностей Melosira praegranulata (var. praegranulata, var. anguloporosa, var. cribrosa, var. minor), M. ignota (var. ignota, var. simplex), M. atlymica (var. atlymica, var. minor), которые, по ее мнению, имеют стратиграфическую и фациальную приуроченность.
Рис. 1.
Область распространения туртасской свиты (сплошная черная линия), местонахождения туртасских комплексов диатомей (пунктирная линия), изученных Н.В. Рубиной (1973), и местоположения скв. Зырянская-1 (настоящая работа) и скважин с находками диноцист (по Кузьмина, Шурыгин, 2016) в туртасской и журавской свитах.
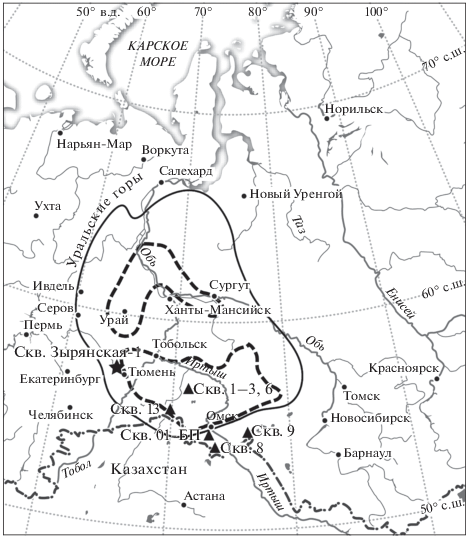
В региональной схеме 2001 г. (Унифицированные…, 2001) комплекс диатомей с Melosira islandica (D7) соответствует атлымскому и новомихайловскому горизонтам, комплекс с Coscinodiscus trapeziformis (D8) – журавскому горизонту, включающему туртасскую свиту. В выделенной Н.А. Добруцкой (1961) у с. Леуши зареченской пачке (зеленовато-серые диатомовые глины с прослоями серых алевритов), залегающей с несогласием на типичных породах туртасской свиты, реперные виды туртасского комплекса не встречены. Индикаторами зареченской пачки Н.В. Рубина (1968) считает виды Melosira atlymica, Coscinodiscus lobatus, Melosira jouseana. Позже Н.В. Рубина (1973а) отнесла этот комплекс к абросимовской свите миоцена. Близкий комплекс с Melosira atlymica, датированный миоценом, описан из глин (мощностью 16–18 м) в составе куполообразных структур, внедренных в песчано-алевритовые прибрежно-морские отложения плейстоцена в береговых обрывах на западном побережье п-ва Ямал вблизи полярной станции Марресале (Полякова, Данилов, 1989).
В последние годы ископаемые пресноводные диатомеи Западной Сибири в стратиграфическом аспекте не изучались, однако ряд характерных эндемичных видов из туртасской свиты подвергcя таксономической ревизии. Так, Coscinodiscus trapeziformis Rub. и C. trapeziformis var. grandis Rub. были переведены в род Actinocyclus (Рубина, Хурсевич, 1990; Хурсевич, Рубина, 1991). Вид Coscinodiscus flabellatus Rub. был переведен в разновидность A. trapeziformis var. flabellatus Rub. et Khurs. Вид Coscinodiscus lobatus переведен в новую комбинацию Actinocyclus lobatus (Rub.) Rub. et Khurs.; Coscinodiscus gorbunovii Sheshuk. и Coscinodiscus tener Rub. также были переведены в род Actinocyclus (Хурсевич и др., 1990; Khursevich, Kociolek, 2012). Melosira аtlymica Rub. (Рубина, 1962), характерная для вышележащей абросимовской свиты, была переведена в род Aulacoseira – A. atlymica (Rub.) Moisseeva (Моисеева, 1981). Вид, описанный Рубиной как Melosira ignota Rub. (Рубина, 1962), до последнего времени оставался в группе видов неясного систематического положения (Диатомовые…, 1992) и позже был отнесен к роду Pseudoaulacosira (Oreshkina, 2018).
С конца 1990-х годов новые данные по олигоцену Западной Сибири получены в основном по результатам изучения палиноморф и макрофлоры. В работах В.С. Волковой и И.А. Кульковой (Волкова, Кулькова, 1988; Волкова и др., 2000, 2005; Кулькова, Волкова, 1997) реконструирована растительность туртасского времени с преобладанием хвойных пород, с Taxodium spp. и примесью широко- и мелколиственных пород деревьев. Климатические реконструкции указывают на теплоумеренный влажный климат со среднегодовой температурой +15°С, отсутствием отрицательных зимних температур и количеством осадков 800–1000 мм в год. Одна из дискуссионных находок последних лет связана с присутствием в журавской и туртасской свитах органикостенного микрофитопланктона рода Pseudokomewuia (Кузьмина, Волкова, 2004; Кузьмина, Шурыгин, 2016), что позволило вернуться к гипотезе о морском или солоноватоводном характере туртасского моря-озера и его связях с Арктикой и байгубекским морем через Тургайский прогиб. В последнее время предпочтение отдается точке зрения о неморском (пресноводном или солоноватоводном) генезисе этих отложений, основанной на данных по переотложенному характеру зерен глауконита (Шумилова, 1963), биомаркерам (Каширцев и др., 2008), литогеохимическим показателям (Смирнов и др., 2017; Кузьмина и др., 2017б). Первые данные по комплексному изучению литологии и микрофоссилий туртасской свиты по материалам скв. Зырянская-1 в районе г. Тюмень (Александрова и др., 2017; Смирнов и др., 2017) также говорят в пользу ее неморского генезиса. Цель настоящей статьи – более детальный анализ ассоциаций биокремнистых микрофоссилий и литогеохимических характеристик туртасской свиты и реконструкция на их основе обстановок седиментации.
МАТЕРИАЛ И МЕТОДЫ
Скважина Зырянская-1 (Александрова и др., 2017; Смирнов и др., 2017) пробурена в 4 км к северо-западу от с. Зырянка Тюменского района Тюменской области (57°10′6.43′′ с.ш.; 64°59′9.93′′ в.д.) в нижнем течении р. Пышма (абс. отметка 106 м) и имеет следующий разрез (рис. 2):
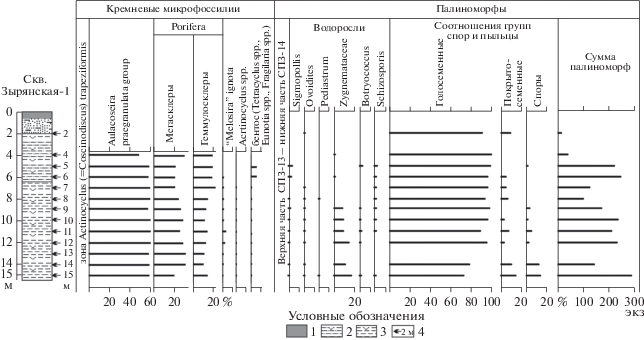
Рис. 2.
Стратиграфическое положение и экологическая структура комплексов микрофоссилий туртасской свиты в скв. Зырянская-1. 1 – почвенно-растительный слой; 2 – глинистый алеврит; 3 – алеврит; 4 – глубина отбора проб.
Инт. 0–0.3 м. Почвенно-растительный слой.
Инт. 0.3–2 м. Алеврит серый, мелкозернистый, неслоистый.
Инт. 2–5 м. Алеврит палево-серый, мелкозернистый, неслоистый, с линзами серого песка и включениями мелкого гравия.
Инт. 5–9 м. Алеврит желто-палевого цвета, массивный, тонкозернистый.
Инт. 9–15 м. Глинистый алеврит желто-палевого цвета, тонкозернистый, плотный, неясно-тонкослоистый.
Химическую обработку 13 проб (рис. 2) для выделения микрофоссилий проводили по стандартной процедуре, принятой в лабораториях палеофлористики и микропалеонтологии ГИН РАН (Александрова и др., 2012).
Для изучения диатомей были сделаны стандартные препараты (Диатомовые…, 1974). При просмотре покровного стекла 18 × 18 мм с объективом ×40 и окуляром ×16 для оценки встречаемости каждого вида диатомовых водорослей в препарате использовалась полуколичественная шкала: единичная встречаемость – 1–10 экземпляров в препарате (<0.1%); редко – 10–30 экземпляров в препарате (1–10%); часто – несколько экземпляров в каждом ряду покровного стекла (10–50%); в массе – несколько экземпляров в каждом поле зрения (>50%). Характерные таксоны кремневых микрофоссилий приведены в табл. I–IV .
Таблица I . Микрофоссилии туртасской свиты, скв. Зырянская-1, юго-запад Тюменской области. 1 – общий вид комплекса; 2–7 – геммулосклеры кремневых губок: 2, 3, 4 – гл. 13 м, 5 – гл. 15 м, 6 – гл. 14 м, 7 – гл. 8 м; 8 – Eunotia nikolskiae Tscher., гл. 6 м; 9 – Ebriopsis crenulata Hovasse, гл. 13 м; 10 – Eunotia arcus Ehr., гл. 8 м; 11 – Eunotia monodon Her., гл. 16 м; 12 – Eunotia majuscula Moiss., гл. 14 м; 13 – циста Chrysophyta, гл. 13 м.
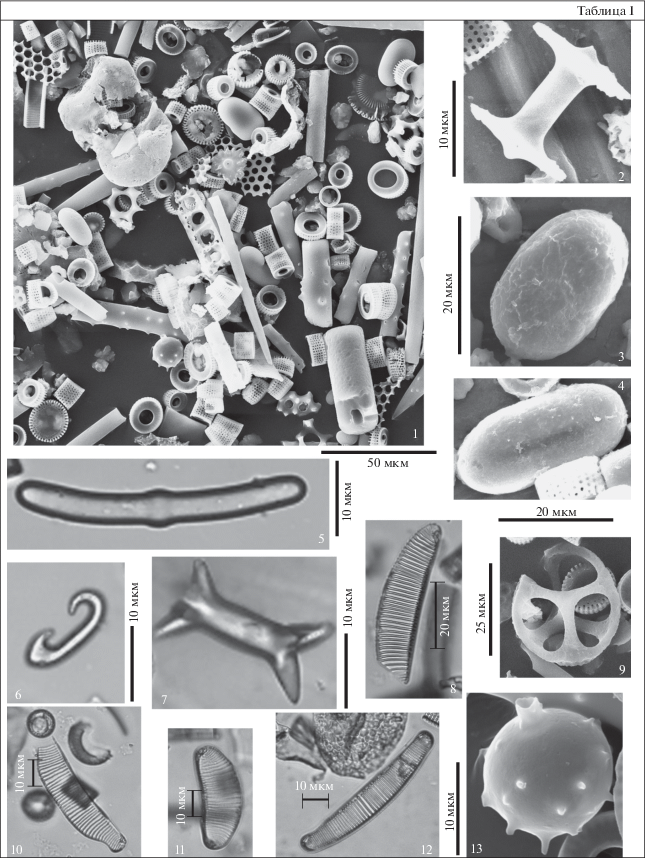
Таблица II. Диатомовые водоросли туртасской свиты, скв. Зырянская-1, юго-запад Тюменской области. 1, 2 – Pseudoaulacosira ignota (Rub.) Oreshkina comb. nov., гл. 13 м; 3, 4 – Aulacoseira praegranulata (Jousé) Simonsen var. praegranulata (Jousé) Moiss.: 3 – гл. 6 м, 4 – гл. 13 м; 5, 6 – Aulacoseira praegranulata var. praeislandica (Jousé) Moiss.: 5 – гл. 13 м, 6 – гл. 11 м; 7, 8 – Actinocyclus tener (Rub.) Rub. et Khurs., гл. 13 м.

Таблица III. Диатомовые водоросли туртасской свиты, скв. Зырянская-1, юго-запад Тюменской области. 1–4, 7, 8 – Actinocyclus tener (Rub.) Rub. et Khurs.: 1 – гл. 15 м, 2, 3 – гл. 8 м, 4 – гл. 6 м, 7 – гл. 10 м, 8 – гл. 13 м; 5, 6 – Actinocyclus trapeziformis (Rub.) Rub. et Khurs.: 5 – гл. 10 м, 6 – гл. 11 м.
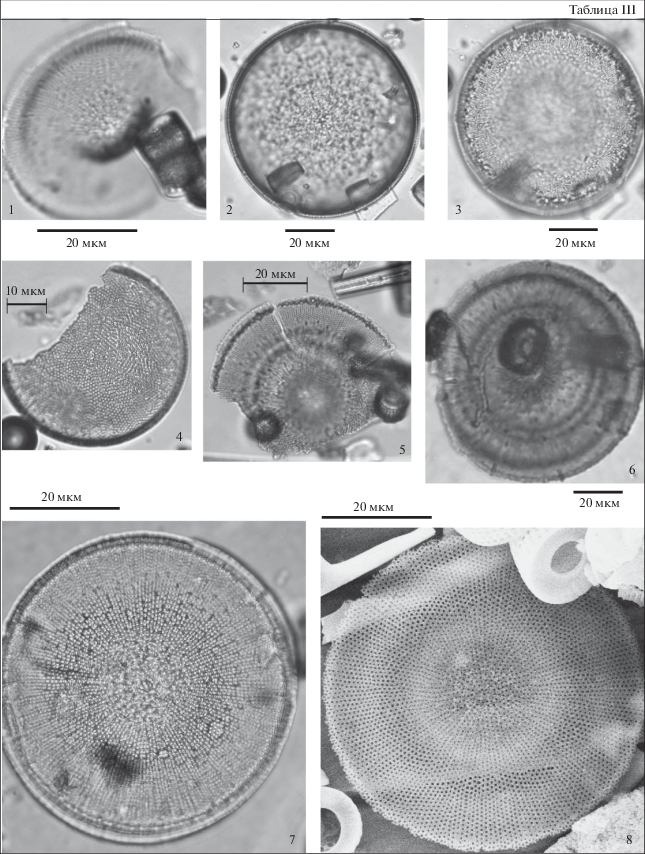
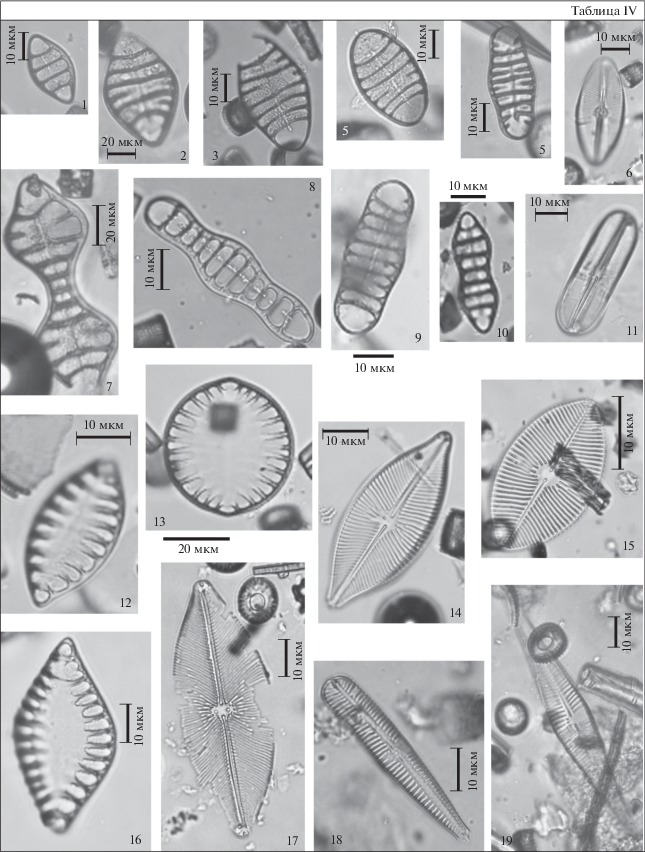
Таблица IV. Диатомовые водоросли туртасской свиты, скв. Зырянская-1, юго-запад Тюменской области. 1–3 – Tetracyclus lacustris var. strumosus (Ehr.) Hust., гл. 15 м; 4 – Tetracyclus ellipticus (Ehr.) Grun., гл. 15 м; 5, 9 – Tetracyclus elongatus Hust.: 5 – гл. 14 м, 9 – гл. 12 м; 6 – Navicula sp., гл. 15 м; 7 – Tetracyclus celatom Okuno, гл. 6 м; 8 – Tetracyclus ellipticus var. latissima Hust., гл. 6 м; 10 – Tetracyclus javanicus Hust., гл. 15 м; 11 – Navicula sp., гл. 14 м; 12, 16 – Surirella turgida W. Sm., гл. 7 м; 13 – Campilodiscus ? sp., гл. 11 м; 14 – Placoneis gastrum (Her.) Meresch., гл. 7 м; 15 – Placoneis sp., гл. 15 м; 17 – Navicula jimboi Pant., гл. 15 м; 18 – Gomphonema sp., гл. 7 м; 19 – Gomphocymbella? sp., гл. 14 м.
Изучение палинологических препаратов и фотографирование таксонов (изображения встреченных в туртасской свите палиноморф приведены в статье (Александрова и др., 2017)) производилось в оптическом микроскопе Axiostar plus (Carl Zeiss) при рабочих увеличениях ×400 и ×600. Определялась таксономическая принадлежность всех встреченных в препарате палиноморф. Минимальное подсчитываемое количественное значение на образец – 200 экземпляров.
Для литогеохимического изучения вмещающих пород в Лаборатории химико-аналитических исследований ГИН РАН были использованы усредненные пробы из следующих интервалов скв. Зырянская-1: 8–9 м, 9–10 м, 10–11 м, 11–12 м, 12–15 м. Из проб, истертых до размерности 100 мкм, прессовали аналитические таблетки диаметром 32 мм, в которых определяли элементы рентгенофлуоресцентным методом с использованием последовательного спектрометра S4 Pioneer фирмы “Bruker” (Германия) и программного обеспечения “Spectra-Plus”. В качестве образцов сравнения использованы стандартные образцы (ГСО, ОСО, МСО), близкие по литологическому составу к анализируемым пробам (алевролиты и песчаники). Для построения градуировочных графиков использовалось более 50 стандартных образцов различного состава. Техника подготовки проб и статистические показатели точности и правильности анализа соответствуют требованиям Методики НСАМ № 439-РС (2010). В качестве сравнительного материала также привлекались данные по современным донным отложениям оз. Байкал (Стандартный…, 2009а, 2009б) с высоким содержанием остатков диатомей.
МИКРОПАЛЕОНТОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ОТЛОЖЕНИЙ СКВ. ЗЫРЯНСКАЯ-1
Биокремнистые микрофоссилии (пресноводные и переотложенные морские диатомовые водоросли, спикулы губок, хризофиты, эбридеи) приурочены к интервалу глубин 15–8 м. В интервале 7–4 м содержание биокремнистых остатков значительно снижается, как и их таксономическое разнообразие. Наиболее многочисленная группа биокремнистых организмов представлена массовым количеством мега- и микросклер пресноводных губок, которые нами специально не изучались. Среди них преобладают морфотипы яйцевидной формы размерностью около 20 мкм по продольной оси (табл. I , фиг. 3, 4). Их кремниевый состав (SiO2 с незначительной примесью алюминия) подтвержден локальным микрозондированием. Отмечены единичные хризофиты и эбридеи. Также небольшую долю составляют переотложенные морские микрофоссилии – редкие обломки и целые створки морских эоценовых диатомей (Coscinodiscus payeri Grun., Paralia sulcata (Ehr.) Cl., Pyxidicula moelleri (A. S.) Streln. et Nik., “Umbilicus” spp.). Пресноводные диатомовые присутствуют в массовом количестве. Среди планктонных форм к числу доминантов относятся центрические Aulacoseira praegranulata gr. и Pseudoaulacosira ignota (Rub.) Oreshkina comb. nov. Крупные тонкоареолированные створки пресноводных диатомей рода Actinocyclus, идентифицированные как Actinocyclus (=Coscinodiscus) tener Rub., A. trapeziformis (Rub.) Rub. et Khurs. и Ellerbeckia kochii (Pant.) Moiss., встречены в незначительном количестве. Бентос представлен видами родов Eunotia (E. monodon Ehr., E. arcus Ehr., E. majuscula Moiss., E. nikolskiae Tscher.) и Tetracyclus (T. ellipticus var. latissimus Hust., T. lacustris var. strumosus (Ehr.) Hust., T. ellipticus (Ehr.) Grun., T. ellipticus var. linearis Hust., T. celatom, T. lacustris Ralfs, T. ellipticus var. lancea f. subrostrata Hust., T. clypeus (Ehr.) Li). Кроме того, определены Navicula jimboi Pant., Placoneis gastrum (Ehr.) Meresch., Placoneis spp., Cymbella aspera (Ehr.) Grun., Surirella turgida W. Sm., Navicula americana Ehr.
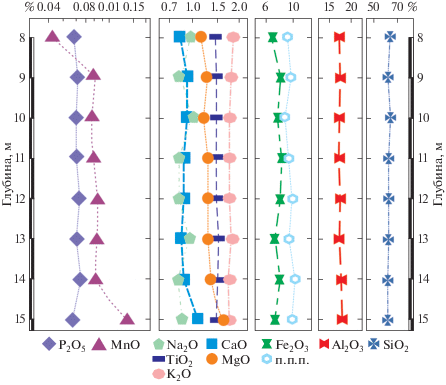
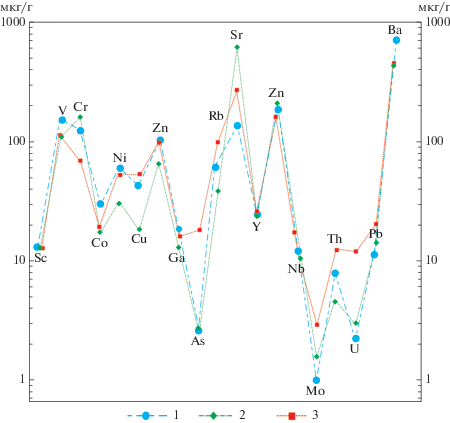
Рис. 4.
Сравнение среднего содержания микроэлементов (мкг/г) в разрезе скв. Зырянская-1 с составом глубоководных (БИЛ-1) и мелководных осадков (БИЛ-2) озера Байкал. 1 – туртасская свита; 2 – мелководные осадки (БИЛ-2) озера Байкал; 3 – глубоководные осадки (БИЛ-1) озера Байкал.
Выделенный комплекс наиболее близок к диатомовой ассоциации, описанной Н.В. Рубиной (1968, 1973а) из нижней части туртасской свиты и включающей обильные переотложенные спикулы, породообразующие Melosira, впоследствии переведенные в род Aulacoseira и относящиеся к группе видов praegranulata. В качестве индикаторов нижней части туртасской свиты Н.В. Рубина приводит эндемичные виды Melosira ignota (=Pseudoaulacosira ignota), а также Coscinodiscus tener, C. lobatus, C. trapeziformis, C. gorbunovii, впоследствии переведенные в род Actinocyclus и связанные между собой переходными формами. В составе бентоса Н.В. Рубина отмечает присутствие разнообразных Tetracyclus (T. celatom, T. lacustris, T. aff. ellipticus (Ehr.) Grun).
Палиноморфы. В палинологических ассоциациях скв. Зырянская-1 (Александрова и др., 2017) преобладает пыльца голосеменных с содержанием не менее 85–90% от общего количества палиноморф. Споры, а также пыльца покрытосеменных немногочисленны. В интервале 15–9 м доминирует пыльца хвойных Pinus s/g Haploxylon et Diploxylon, присутствуют Tsuga canadensis (L.) Carrière, T. crispa Zakl., T. torulosa Zakl., T. sp., Abies sp., Picea sect. Omorica Willk., P. sect. Eupicea Willk., P. tobolica Pan., Taxodiaceae-Cupressaceae, Sciadopitys sp., Podocarpus spp., Cedrus sp., Ephedra sp. В составе покрытосеменных отмечаются редкие Juglans polyporata Vojc., J. sieboldianiformis Vojc., J. nigripites Wodehouse, J. sp., Pterocarya stenopteroides Vojc., P. sp., Ulmus sp., Alnus sp., Sparganium sp., Carya sp., Ilex sp., Liquidambar sp., Tilia sp., Betula sp., Carpinus sp., Corylus sp., Myrica sp., Comptonia sp., Nelumbo sp., Aldrovanda sp., Onargaceae, Ericales. Споры представлены Polypodiaceae, Neogenisporis sp., Sporites durabilis (Dokt.-Hreb.), Lycopodium spp., Osmunda sp. Характерны ископаемые зигоспоры зеленых водорослей как неясного систематического положения, так и близких к Zygnematales: ex gr. Trochlearia sibirica Lub., Diagonalites spp., Megatetrapidites spp., Rundinella spp., Planctonites sp., Sigmopollis sp., Ovoidites spp., Schizosporis sp., что также подтверждает условия пресноводного водоема.
В интервале 8–2 м, на фоне сохраняющегося преобладания пыльцы Pinus s/g Haploxylon et Diploxylon, отмечается исчезновение Tsuga spp., Taxodiaceae-Cupressaceae, Sciadopitys sp., большинства таксонов покрытосеменных, спор наземных растений, а также зигоспор Zygnematales. В пробах с глубины 4–2 м палиноморфы уже единичны и представлены только пыльцой Pinaceae. Синхронная изменениям палинокомплексов смена вмещающих тонких глинисто-алевритовых осадков на песчано-алевритовые свидетельствует о переходе озерных палеообстановок в аллювиальные.
Преобладание пыльцы Pinaceae, присутствие таких стратиграфически важных таксонов, как Picea tobolica, Juglans polyporata, Pterocarya stenopteroides, позволяют сопоставить рассматриваемый комплекс с верхней частью палинозоны СПЗ-13 и нижней частью палинозоны СПЗ-14 (Панова, 1971; Волкова, Кулькова, 1980, 1984; Практическая…, 1990; Унифицированные…, 2001; Кузьмина, Волкова, 2008), что соответствует верхней части новомихайловского горизонта–нижней части журавского горизонта (верхний рюпель–нижний хатт). С учетом данных по диатомовым водорослям, изученный интервал скв. Зырянская-1 отвечает нижней части туртасской свиты (нижний олигоцен). Возраст и генезис серых алевритов в инт. 0.3–2 м нами не установлен.
ЛИТОГЕОХИМИЯ
Проанализирован литохимический состав проб и петрохимические модули для 8 проб туртасской свиты из скв. Зырянская-1 (табл. 1, 2).
Таблица 1.
Состав проб (%) и петрохимические модули для скв. Зырянская-1 и донных отложений оз. Байкал (пробы БИЛ-1 и БИЛ-2)
| Место отбора пробы, шифр пробы | Глубина, м | SiO2 | TiO2 | Al2O3 | Fe2O3 | MnO | MgO | CaO | K2O | Na2O | P2O5 | П.п.п | ТМ | Al2O3/TiO2 | ЩМ | CIА | Fe/Mn | CaO/MgO MgO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| скв. Зырянская-1 | 8 м | 62.94 | 1.39 | 16.46 | 6.34 | 0.043 | 1.13 | 0.82 | 1.79 | 0.95 | 0.06 | 8.06 | 0.084 | 12 | 0.53 | 82 | 134 | 0.73 |
| » | 9 м | 61.08 | 1.40 | 16.66 | 7.36 | 0.077 | 1.23 | 0.90 | 1.74 | 0.86 | 0.06 | 8.62 | 0.084 | 12 | 0.49 | 83 | 86 | 0.73 |
| » | 10 м | 62.49 | 1.41 | 16.30 | 7.03 | 0.076 | 1.17 | 0.90 | 1.70 | 0.92 | 0.06 | 7.94 | 0.099 | 10 | 0.54 | 82 | 83 | 0.77 |
| » | 11 м | 61.45 | 1.40 | 16.55 | 7.55 | 0.078 | 1.24 | 0.88 | 1.69 | 0.85 | 0.06 | 8.26 | 0.085 | 12 | 0.50 | 83 | 88 | 0.71 |
| » | 12 м | 60.91 | 1.41 | 16.70 | 7.37 | 0.083 | 1.24 | 0.88 | 1.68 | 0.82 | 0.06 | 8.84 | 0.084 | 12 | 0.50 | 83 | 81 | 0.71 |
| » | 13 м | 62.00 | 1.44 | 16.53 | 6.70 | 0.083 | 1.25 | 0.86 | 1.76 | 0.92 | 0.06 | 8.39 | 0.087 | 11 | 0.52 | 82 | 73 | 0.69 |
| » | 14 м | 60.49 | 1.42 | 16.94 | 7.21 | 0.081 | 1.30 | 0.86 | 1.67 | 0.82 | 0.07 | 9.15 | 0.084 | 12 | 0.49 | 84 | 80 | 0.66 |
| » | 15 м | 60.46 | 1.41 | 17.25 | 6.84 | 0.131 | 1.52 | 1.08 | 1.66 | 0.84 | 0.06 | 8.74 | 0.082 | 12 | 0.51 | 83 | 47 | 0.71 |
| Среднее по скважине | 61.48 | 1.41 | 16.67 | 7.05 | 0.082 | 1.26 | 0.90 | 1.71 | 0.87 | 0.061 | 8.5 | 0.086 | 11.6 | 0.51 | 83 | 84 | 0.71 | |
| БИЛ-1 | 61.07 | 0.69 | 13.57 | 7.02 | 0.40 | 2.0 | 1.85 | 2.21 | 1.96 | 0.345 | 8.34 | 0.051 | 20 | 0.89 | 69 | 16 | 0.93 | |
| БИЛ-2 | 62.46 | 0.76 | 14.22 | 5.39 | 0.12 | 3.12 | 7.09 | 1.51 | 3.11 | 0.139 | 1.78 | 0.053 | 19 | 2.06 | 55 | 41 | 2.27 |
Таблица 2.
Микроэлементный состав проб (мкг/г) и петрогеохимические модули для скв. Зырянская-1 и донных отложений оз. Байкал (пробы БИЛ-1 и БИЛ-2)
| Место отбора пробы, шифр пробы | Глубина, м | V | Cr | Co | Ni | Cu | Zn | Ga | As | Rb | Sr | Y | Zr | Nb | Mo | Th | U | Pb | Ba | Sr/Ba |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| скв. Зырянская-1 | 8 | 145 | 117 | 20 | 57 | 38 | 90 | 19 | <2 | 61 | 146 | 25 | 238 | 17 | <1 | 6.8 | 3.4 | 12 | 547 | 0.27 |
| » | 9 | 150 | 125 | 29 | 64 | 52 | 104 | 20 | <2 | 61 | 143 | 26 | 195 | 17 | <1 | 6.0 | 2.3 | 13 | 574 | 0.25 |
| » | 10 | 151 | 120 | 27 | 59 | 42 | 93 | 18 | 6.1 | 57 | 142 | 25 | 204 | 17 | <1 | 5.9 | <1 | 13 | 542 | 0.26 |
| » | 11 | 153 | 125 | 29 | 57 | 44 | 97 | 18 | <2 | 59 | 137 | 26 | 199 | 17 | <1 | 7.2 | 2.2 | 12 | 538 | 0.25 |
| » | 12 | 157 | 124 | 30 | 62 | 41 | 103 | 19 | 5.4 | 60 | 137 | 26 | 196 | 17 | <1 | 6.9 | 2.0 | 13 | 546 | 0.25 |
| » | 13 | 152 | 122 | 30 | 56 | 39 | 101 | 19 | <2 | 61 | 144 | 26 | 203 | 16 | <1 | 5.8 | <1 | 13 | 569 | 0.25 |
| » | 14 | 154 | 127 | 32 | 65 | 45 | 107 | 20 | <2 | 60 | 137 | 27 | 199 | 17 | <1 | 6.3 | 2.1 | 13 | 545 | 0.25 |
| » | 15 | 161 | 138 | 34 | 68 | 44 | 109 | 20 | <2 | 63 | 131 | 26 | 201 | 18 | <1 | 7.1 | 3.6 | 13 | 528 | 0.25 |
| Среднее по скважине | 153 | 125 | 29 | 61 | 43 | 101 | 19 | 2.5 | 60 | 139 | 26 | 204 | 17 | <1 | 8 | 2.2 | 13 | 549 | 0.25 | |
| БИЛ-1 | 110 | 66 | 18 | 54 | 52 | 96 | 16 | 18 | 93 | 266 | 30 | 156 | 12 | 2.9 | 13 | 12 | 21 | 710 | 0.37 | |
| БИЛ-2 | 105 | 158 | 17 | 31 | 18 | 64 | 13 | 2.5 | 39 | 580 | 24 | 204 | 10 | 1.6 | 4.8 | 3 | 14 | 530 | 1.09 |
В целом алевриты туртасской свиты характеризуются достаточно устойчивым составом во всех изученных пробах (рис. 3). Вместе с тем в основании разреза наблюдаются синхронные изменения (рост) концентраций MnO (0.043–0.131%), CaO (0.82–1.08%), MgO (1.13–1.52%) относительно его верхней части, что можно интерпретировать как некоторые различия в условиях осадконакопления в начале и в конце временного интервала формирования осадков. Остальные макрокомпоненты ведут себя относительно устойчиво. Концентрации отдельных микроэлементов (табл. 2), в частности элементов семейства железа (Sc, V, Cr, Co, Ni), незначительно возрастают сверху вниз по разрезу, аналогично ведут себя элементы второй группы (Cu, Zn, Pb). В то же время концентрации элементов литофильной группы (Y, Zr, Nb, Th) несколько уменьшаются.
Геохимический состав проб скв. Зырянская-1 сопоставлен с имеющимися данными по современным донным отложениям оз. Байкал, материал из которых отобран на различных участках котловины озера для создания стандартных образцов состава (СОС). Данные по их макро- и микроэлементному составу также представлены в табл. 1 и 2 (Стандартный…, 2009а, 2009б). Материал образца БИЛ-1 отобран из наиболее глубоководной части средней котловины озера Байкал. Здесь донные отложения представлены пелитовыми и мелкоалевритовыми илами с высоким содержанием диатомитовой компоненты (до 16% от массы образца). Место отбора материала БИЛ-2 – внутренняя котловина Малого моря озера Байкал. Материал образца получен как композиция из 13 отдельных проб, поднятых по периферии котловины в прибрежной зоне. Донные отложения представлены песчано-алевро-пелитовым материалом, обогащенным карбонатами и щелочными компонентами.
Сравнение среднего состава микроэлементов осадков озера Байкал и отложений скв. Зырянская-1 (рис. 4) указывает на сходные условия формирования глубоководных диатомитовых илов оз. Байкал и отложений туртасской свиты. Имеющиеся расхождения в содержании отдельных микроэлементов можно объяснить различиями в источниках сноса и глубине бассейнов.
Для реконструкции условий осадконакопления использовался ряд классических расчетных параметров (рис. 5). Одним из наиболее достоверных показателей принято считать соотношение железа и марганца (Fe/Mn). Значения этого модуля уменьшаются с увеличением глубины бассейна и с переходом от прибрежных фаций к более глубоководным (Yanochkina, 1966; Nesbitt, Young, 1982; Розен и др., 1994; Скляров, 2001; Маслов и др., 2003). Тенденция уменьшения этого модуля обусловлена поглощением осадочными отложениями марганца, проявляющимся сильнее по мере увеличения глубины бассейна.
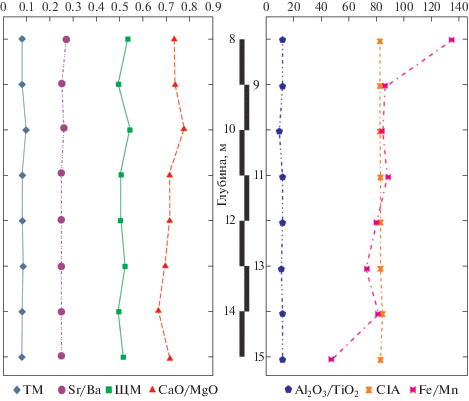
По значениям отношения Fe/Mn осадочные породы классифицируются следующим образом: <40 – глубоководные; 40–80 – мелководные; >160 – мелководно-прибрежные с преимущественно терригенным источником сноса. Кроме того, соотношение Fe/Mn > 13 характеризует освещенный мелководный бассейн, а значения <13 соответствуют глубоководной среде с более слабой освещенностью. В нашем случае в разрезе скв. Зырянская-1 индекс Fe/Mn варьирует в интервале 134–47, что свидетельствует об уменьшении глубины бассейна в ходе осадконакопления. При этом условия седиментации могут колебаться от умеренно глубоководных до мелководных с соответствующим уровнем освещенности. Следуя модели, предложенной в перечисленных выше работах, изученные отложения можно отнести к мелководно-прибрежным.
Таким образом, туртасский бассейн, скорее всего, развивался от относительно глубоководного в начале рассматриваемого интервала времени к более мелководному. Высокие показатели освещенности в туртасском бассейне подтверждаются интенсивной вегетацией планктонных диатомей и развитием губок. На общую пониженную минерализацию вод туртасского бассейна указывают низкие значения фациального индикатора палеосолености Sr/Ba.
Для реконструкции климатических показателей используются несколько модулей. Значения щелочного модуля (ЩМ = Na2O/K2O), равные 0.47–0.54, указывают на повышенную интенсивность химического выветривания и, следовательно, на теплый гумидный климат, что подтверждается и составом палинокомплекса туртасской свиты. Щелочной модуль прибрежных осадков Байкала (БИЛ-2), напротив, говорит о слабом химическом выветривании в условиях аридного климата.
Индекс химического выветривания CIA (Al2О3/(Al2О3 + СаО + Na2О + K2О) × 100) также используется как индикатор климатических параметров (Nesbitt, Young, 1982). Для осадочных отложений гумидных обстановок характерен вынос из полевых шпатов кальция, натрия и калия, что ведет к увеличению соотношения алюминия и щелочей в продуктах выветривания. В качестве критерия для разграничения отложений, формирующихся в обстановках теплого и холодного климата, принято считать значение индекса химического выветривания, равное 70. Невыветрелые породы характеризуются значениями CIA порядка 50, тогда как CIA сильно выветрелых разновидностей стремятся к 100 (Медведева, 2016). Величина индекса для пород туртасской свиты находится в пределах 82–84 (табл. 1). Это соответствует умеренно-теплому климату с промежуточными значениями уровня выветрелости пород.
Еще одним показателем климатических условий является титановый модуль ТМ (ТiO2/А12О3). Его использование основано на двух противоположных тенденциях поведения титана и алюминия в процессах химического выветривания. Величина ТМ возрастает при переходе из аридной зоны в гумидную, а в пределах последней – по мере движения от глубоководных зон к прибрежным (Енгалычев, Панова, 2011). Следует отметить, что имеется вероятность поглощения титана губками и другими организмами, в результате чего его повышенные значения могут отмечаться для кремнистых органогенных пород, например для спонголитов (Юдович, 1981). Величина модуля ТМ для скв. Зырянская-1 составляет 82–99, что, возможно, отражает высокую концентрацию биокремнистых микроорганизмов в туртасском бассейне.
Еще одним индикатором климата в области сноса является отношение А12O3/ТiO2, которое составляет <20 для гумидных и >30 для аридных обстановок. Этот показатель находится в пределах 10–12 для пород туртасской свиты, что соответствует предложенной выше оценке климатической обстановки формирования этих пород.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ И ВЫВОДЫ
С регрессией тавдинского моря в эоцене связано формирование обширной озерно-аллювиальной равнины, в пределах которой протекало осадконакопление на протяжении всего олигоцена (Шацкий, 1978). Образование значительного по площади позднеолигоценового туртасского бассейна, вероятно, было связано с активизацией Иртыш-Обского субрегионального прогиба. Туртасская озерная система во время олигоценового седиментационного цикла активно меняла свою конфигурацию. Рассчитанные значения литохимических модулей свидетельствуют о том, что бассейн в начале исследованного интервала времени развивался от относительно глубоководного к более мелководному. В совокупности с наличием в разрезе песчаных линз и плохо сортированных разнозернистых песков с гравийным материалом (Смирнов и др., 2017) это является признаком существования в краевых частях бассейна постоянно перемещавшейся береговой линии. Периодические прогибания территории Западной Сибири, на фоне ее общего подъема в олигоцене (Астапов и др., 1979), происходили при относительно высокой роли химического выветривания. По-видимому, таким образом обеспечивался трансфер кремния в форме истинных или коллоидных растворов в бассейн седиментации.
В палеогеновом разрезе Западной Сибири осадки, содержащие морские диатомовые комплексы раннего эоцена–начала среднего эоцена, сменяются отложениями тавдинской свиты, в которой не найдены признаки биогенного кремненакопления (Ахметьев и др., 2010). Для олигоцена и миоцена характерны комплексы пресноводных диатомей новомихайловской (ранний олигоцен), туртасской и журавской (поздний олигоцен) и абросимовской (начало миоцена) свит. Туртасская пресноводная флора диатомей, датированная поздним олигоценом, со значительным присутствием эндемичных видов, не имеет аналогов на территории Северной Евразии. Находки донеогеновой пресноводной диатомовой флоры в Северной Евразии ограничены комплексом эоцен-олигоценового возраста, обнаруженным на Камчатке (мыс Астрономический) (Лупикина, Долматова, 1985). Он имеет лишь частичное сходство с туртасской флорой. Комплекс с преобладанием пеннатных диатомей олигоценового возраста известен из Северной Богемии на границе Чехии и Германии (Houk, 2007; Shiller, 2007), однако сходства с туртасской флорой не наблюдается. Наиболее полно развитие донеогеновых пресноводных диатомей можно проследить на Североамериканском континенте, где известно несколько местонахождений возрастом от позднего мела до олигоцена. Их подробная характеристика приведена в работе (Benson et al., 2007). Олигоцен-миоценовый комплекс из местонахождения в штате Монтана (формация Canion Ferry) отличается доминированием диатомей Aulacoseira granulata gr., что характерно и для туртасской флоры. Более детально изучена в последние годы неогеновая диатомовая флора Сибири и Дальнего Востока. Это местонахождения в Приморье, на Витимском плато, Байкале, Камчатке, на подводных возвышенностях в Японском море, однако ассоциации из этих местонахождений лишь отчасти наследуют структуру и таксономический состав туртасской флоры (Моисеева, 1971; Лихачева и др., 2009; Усольцева, Титова, 2015; Цой, Усольцева, 2016).
Реперным видом туртасской флоры является эндемичный вид Pseudiaulacosira ignota. Впервые род Pseudoaulacosira был описан из эрмановской свиты неогена Западной Камчатки (Атлас…, 1984) и представлен видом P. moisseevae. Второй вид этого рода – P. sculpta Usoltseva et Houk установлен в третичных отложениях штата Орегон западного побережья США (Usoltseva, Houk, 2016). Таким образом, присутствие рода Pseudoaulacosira в отложениях верхнего олигоцена Западной Сибири значительно расширяет его стратиграфический и географический диапазон.
Отличительная черта туртасской диатомовой флоры – присутствие эндемичных видов рода Actinocyclus. Близкие по морфологии и возрасту представители этого рода с тонкоареолированными створками известны из морских местонахождений Северного Перитетиса. Так, Actinocyclus mangishlakensis описан А.П. Жузе (1978) из карагиинской свиты верхнего олигоцена во впадине Карагие п-ва Мангышлак. В майкопских отложениях Западной Кубани присутствует Actinocyclus kisselevii Makar. (Шешукова-Порецкая, Глезер, 1962), в менелитовой формации Польских Карпат (олигоцен–ранний миоцен) встречены Actinocyclus octonarius Ehr., A. loczii Pant., A. cholnokyi Van Lan. (Kotlarczyk, Kaczmarska, 1987). Таким образом, для диатомовых комплексов туртасской свиты характерна значительная степень эндемизма на видовом уровне, которая, как отмечала Н.В. Рубина (1968), нивелируется в комплексах вышележащей абросимовской свиты, уже близких по таксономическому составу к неогеновым комплексам Приморья, Прибайкалья, Японского моря (Моисеева, 1971; Лихачева и др., 2009; Усольцева, Титова, 2015; Цой, Усольцева, 2016).
Еще одной особенностью туртасского пресноводного биокремнистого комплекса является значительная доля участия мега- и микросклер пресноводных губок и единичное присутствие эбридей и стоматоцист хризофитовых водорослей. Н.В. Рубина (1968) предполагала, что многочисленные спикулы губок, приуроченные к нижней части туртасской свиты, переотложены из морских эоценовых отложений. Полученный нами материал с разнообразными морфотипами макросклер, высоким содержанием геммулосклер, которые не отмечались в морских эоценовых отложениях, скорее предполагает их инситный генезис. Хотя эта группа биокремнистых микроорганизмов нами специально не изучалась, уже сейчас можно сказать, что отдельные встреченные морфотипы близки к группе современных тропических Potamophlois (Manconi, Pronzato, 2002). Следует отметить, что в Байкале пресноводные губки обитают в широком диапазоне глубин – от 2 до 1000 м. Незначительное присутствие хризофитовых водорослей и эбридей, возможно, связано с повышенной трофностью туртасского бассейна и относительно высокими температурными показателями, неблагоприятными для развития этих групп кремневых микрофоссилий.
Ассоциации палиноморф скв. Зырянская-1 имеют наибольшее сходство с комплексами туртасской свиты Нижнего Приобья (Добруцкая, 1961), но существенно отличаются от ассоциаций палиноморф туртасской и журавской свит более южных районов Западной Сибири отсутствием пыльцы Fagaceae, низким содержанием Taxodiaceae/Cupressaceae и Betulaceae, высоким содержанием голосеменных Pinus s/g Haploxylon et Diploxylon. Особо следует отметить, что в нашем материале не были найдены пресноводные диноцисты рода Pseudokomewuia, присутствующие в палинокомплексах ряда местонахождений журавской и туртасской свит (Кузьмина, Волкова, 2001, 2008; Кузьмина и др., 2003, 2017а; Кузьмина, Шурыгин, 2016; Волкова и др., 2005). Возможно, это связано с несколькими причинами. Во-первых, следует иметь в виду конкурентные отношения органикостенной и кремневой групп планктона, имеющих различные жизненные стратегии. Кроме того, присутствие диноцист в отложениях более южных районов Западной Сибири, возможно, связано с водными массами, проникавшими в эту часть бассейна из области Перитетиса. Вполне вероятен и сценарий, при котором наличие высоких концентраций диноцист рода Pseudokomewuia в журавской и туртасской свитах приходится на максимум позднеолигоценовой трансгрессии (Кузьмина и др., 2017б), в то время как отложения, вскрытые скв. Зырянская-1, относятся к более раннему этапу развития туртасского бассейна – началу позднего олигоцена. Дальнейшее комплексное биостратиграфическое изучение более полных разрезов туртасской и журавской свит с привлечением литогеохимических методов откроет новые возможности для детальной реконструкции этапов развития туртасской озерной системы в позднем олигоцене Западной Сибири.
Благодарности. Авторы искренне признательны С.В. Гришину за помощь в лабораторной обработке пород, Н.В. Горьковой и Е.А. Жегалло за предоставленную возможность изучения микрофоссилий в сканирующем электронном микроскопе. Особая благодарность рецензентам О.Б. Кузьминой, Н.К. Лебедевой и А.И. Яковлевой за качественный и глубокий разбор статьи.
Источники финансирования. Исследования проведены в соответствии с планами научно-исследовательской работы ГИН РАН.
Список литературы
Александрова Г.Н., Орешкина Т.В., Яковлева А.И., Радионова Э.П. Диатомеи и диноцисты в интервале позднего палеоцена–раннего эоцена в биокремнистых фациях Среднего Зауралья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 4. С. 68–94. https://doi.org/10.1134/ S0869593812030021
Александрова Г.Н., Орешкина Т.В., Смирнов П.В., Константинов А.О., Кузьмина О.Б. Данные о возрасте и генезисе отложений верхнего кайнозоя Тюменской области (Западная Сибирь), обогащенных биокремнистым материалом // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 4. С. 62–88. https://doi.org/10.7868/S869592X17040032
Астапов А.П., Дрожащих Н.Б., Генералова Р.С. Палеогеография палеогена и неогена юга Тюменской области в связи с оценкой перспективности территории на нерудное сырье. Тюмень: ЗапсибНИГНИ, 1979. 33 с.
Атлас фауны и флоры палеогеновых и неогеновых отложений Дальнего Востока. Точилинский опорный разрез Западной Камчатки. М.: Наука, 1984. 332 с.
Ахметьев М.А., Запорожец Н.И., Яковлева А.И., Александрова Г.Н., Беньямовский В.Н., Орешкина Т.В., Гнибиденко З.Н., Доля Ж.А. Сравнительный анализ разрезов и биоты морского палеогена Западной Сибири и Арктики // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 6. С. 78–103.
Волкова В.С., Кулькова И.А. Палинологическое обоснование стратиграфического расчленения и корреляция континентальных палеогеновых и неогеновых отложений юга Западной Сибири и Северного Казахстана // Палеопалинология Сибири. М.: Наука, 1980. С. 65–72.
Волкова В.С., Кулькова И.А. Изменение состава палеофлоры Сибири в позднем кайнозое // Среда и жизнь на рубежах эпох кайнозоя в Сибири и на Дальнем Востоке. Новосибирск: Наука, 1984. С. 54–63.
Волкова B.C., Кулькова И.А. Количественная оценка некоторых элементов климата позднего олигоцена и неогена Сибири // Палинология в СССР. Новосибирск: Наука, 1988. С. 31–35.
Волкова В.С., Гнибиденко З.Н., Кулькова И.А. О природе туртасского позднеолигоценового озера-моря в Западной Сибири // Геология и геофизика. 2000. Т. 141. № 1. С. 62–70.
Волкова В.С., Кузьмина О.Б., Кулькова И.А. Морской и континентальный палеоген и миоцен южной части Барабинского фациального района Западной Сибири (стратиграфия, микрофоссилии) // Геология и геофизика. 2005. Т. 46. № 1. С. 60–71.
Гурари Ф.Г., Шацкий С.Б., Хохлова Г.Я., Соколов П.Н. К стратиграфии континентальных верхнепалеогеновых отложений Западно-Сибирской низменности // Решения и труды Межведомственного совещания по доработке и уточнению стратиграфических схем Западно-Сибирской низменности. Л.: Гостоптехиздат, 1961. С. 308–315.
Диатомовые водоросли СССР (ископаемые и современные). Т. 1. Ред. Прошкина-Лавренко А.И. Л.: Наука, 1974. 402 с.
Диатомовые водоросли СССР. Ископаемые и современные. Т. II. Вып. 2. Ред. Глезер З.И., Макарова И.В. СПб.: Наука, 1992. 125 с.
Добруцкая Н.А. Стратиграфия континентальных третичных отложений нижнего Приобья // Труды Межведомственного совещания по доработке и уточнению стратиграфических схем Западно-Сибирской низменности. Л.: Гостоптехиздат, 1961. С. 295–307.
Енгалычев С.Ю., Панова Е.Г. Геохимия и генезис песчаников восточной части Главного девонского поля на северо-западе Русской плиты // Литосфера. 2011. № 5. С. 16–29.
Жузе А.П. Диатомеи и силикофлагеллаты Южного Мангышлака // Морская микропалеонтология (диатомеи, радиолярии, силикофлагелляты, фораминиферы и известковый планктон). Ред. Жузе А.П. М.: Наука, 1978. С. 49–58.
Зальцман И.Г. Стратиграфия палеогеновых и неогеновых отложений Кулундинской степи. Красноярск: Красноярское книжное издательство, 1968. 150 с.
Зыкин В.С. Стратиграфия и эволюция природной среды и климата в позднем кайнозое юга Западной Сибири. Новосибирск: Гео, 2012. 487 с.
Каширцев В.А., Конторович А.Э., Москвин В.И., Кучкина А.Ю., Ким В.Е. Углеводороды-биомаркеры в органическом веществе палеогеновых отложений юга Западной Сибири // Нефтехимия. 2008. Т. 48. № 4. С. 271–276.
Кузьмина О.Б., Волкова В.С. Стратиграфия и палинологическая характеристика олигоцен-миоценовых отложений по материалам бурения (скв. 01-БП) в Омском Прииртышье Западной Сибири // Новости палеонтологии и стратиграфии. 2001. Вып. 4. С. 135–141.
Кузьмина О.Б., Волкова В.С. Первые находки динофлагеллат в континентальных верхнеолигоценовых отложениях журавской свиты (Западная Сибирь) // Докл. АН. 2004. Т. 394. № 2. С. 229–233.
Кузьмина О.Б., Волкова В.С. Палиностратиграфия континентальных олигоцен-миоценовых отложений юга Западной Сибири // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 5. С. 87–100.
Кузьмина О.Б., Шурыгин Б.Н. Первые находки диноцист в туртасской свите (верхний олигоцен) на юге Тюменской области // Докл. АН. 2016. Т. 467. № 6. С. 682–686.
Кузьмина О.Б., Волкова В.С., Гнибиденко З.Н., Лебедева Н.К. Микрофитофоссилии и магнитостратиграфия верхнемеловых и кайнозойских отложений юго-восточной части Западно-Сибирской равнины // Геология и геофизика. 2003. Т. 44. № 4. С. 348–363.
Кузьмина О.Б., Гнибиденко З.Н., Хазин Л.Б., Хазина И.В. Новые данные по стратиграфии (палиноморфы, остракоды, палеомагнетизм) континентальных кайнозойских отложений Ишимской равнины, Западная Сибирь // Стратиграфия. Геол. корреляция. 2017а. Т. 25. № 3. С. 109–128.
Кузьмина О.Б., Кузьмин Д.В., Шурыгин Б.Н. Причины вспышки продуктивности микрофитопланктона в позднеолигоценовом Туртасском озере-море (Западная Сибирь) // Докл. АН. 2017б. Т. 474. № 6. С. 712–716.
Кулькова И.А., Волкова В.С. Ландшафты и климат Западной Сибири в палеогене и неогене // Геология и геофизика. 1997. Т. 38. № 3. С. 581–595.
Лихачева О.Ю., Пушкарь В.С., Черепанова М.В., Павлюткин Б.И. Зональная диатомовая шкала и основные геобиологические события неогена Приморья // Вестник ДВО РАН. 2009. № 4. С. 64–72.
Лупикина Е.Г., Долматова Л.М. О палеогеновой лагунной флоре диатомей Камчатки // Палеонтол. журн. 1985. № 1. С. 120–128.
Маслов А.В., Крупенин М.Т., Гареев Э.З. Литологические, литохимические и геохимические индикаторы палеоклимата (на примере рифея южного Урала) // Литология и полезн. ископаемые. 2003. № 5. С. 502–525.
Медведева С.А. Литохимические и флористические данные для реконструкции мезозойского палеоклимата (на примере Буреинского осадочного бассейна, Дальний Восток, Россия) // Успехи современного естествознания. 2016. № 4. С. 156–161.
Моисеева А.И. Атлас неогеновых диатомовых водорослей Приморского края // Тр. ВСЕГЕИ. Нов. сер. 1971. Т. 171. 152 с.
Моисеева А.И. Ревизия систематического положения пресноводных видов рода Melosira Ag. s.l. (Bacillariophyta) // Систематика, эволюция, экология водорослей и их значение в практике геологических исследований. Тез. докл. II Всесоюзного палеоальгологического совещания. Киев, 1981. С. 124–126.
Методика НСАМ № 439-РС “Определение фтора, натрия, магния, алюминия, кремния, фосфора, калия, кальция, скандия, титана, ванадия, хрома, марганца, железа, кобальта, никеля, стронция, циркония, ниобия в горных породах, рудах и продуктах их переработки рентгеноспектральным флуоресцентным методом”. М.: ВИМС, 2010 (с изм. № 1 от 12.05.2015 г.).
Панова Л.А. Олигоцен Западно-Сибирской низменности. Кайнозойские флоры Сибири по палинологическим данным. М.: Наука, 1971. С. 40–50.
Полякова Е.И., Данилов И.Д. Миоцен крайнего севера Западной Сибири (полуостров Ямал) // Докл. АН СССР. 1989. Т. 308. № 2. С. 428–431.
Практическая палиностратиграфия. Ред. Панова Л.А., Ошуркова М.В., Романовская Г.М. Л.: Недра, 1990. 348 с.
Розен О.М., Журавлев Д.3., Ляпунов С.М. Геохимические исследования осадочных отложений Тимано-Печерской провинции // Разведка и охрана недр. 1994. № 1. С. 18–21.
Рубина Н.В. Новые виды рода Melosira из туртасской свиты Западно-Сибирской низменности // Тр. СНИИГГиМС. 1962. Вып. 23. Материалы по палеонтологии и стратиграфии Западной Сибири. С. 104–107.
Рубина Н.В. Новые виды рода Coscinodiscus Ehr. из туртасской свиты Западно-Сибирской низменности // Новости систематики низших растений. 1967. Т. 4. С. 63–68.
Рубина Н.В. Комплексы диатомей в отложениях туртасской свиты Западно-Сибирской низменности // Ископаемые диатомовые водоросли СССР. М.: Наука, 1968. С. 61–66.
Рубина Н.В. Палеоальгологическое обоснование стратиграфии морских и континентальных палеогеновых и неогеновых отложений Западно-Сибирской низменности // Морской и континентальный палеоген Сибири. Новосибирск: Наука, 1973а. С. 87–96.
Рубина Н.В. Новые разновидности пресноводных диатомовых водорослей из некраcовской серии Западно-Сибирской низменности // Тр. Тюменского индустриального института. 1973б. Вып. 17. С. 104–107.
Рубина Н.В. Новые и редкие виды родов Coscinodiscus Ehr. и Aulacodiscus Ehr. из континентальных верхнеолигоценовых и нижнемиоценовых отложений Западно-Сибирской низменности // Новости систематики низших растений. 1976. Т. 13. С. 54–59.
Рубина Н.В., Хурсевич Г.К. Новые представители рода Actinocyclus (Bacillariophyta) из позднего олигоцена Западной Сибири // Ботан. журн. 1990. Т. 75. № 11. С. 1565–1567.
Скляров Е.В. Интерпретация геохимических данных. Ученое пособие. М.: Интермет Инжиниринг, 2001. 288 с.
Смирнов П.В., Константинов А.О., Александрова Г.Н., Кузьмина О.Б., Шурыгин Б.Н. Новые данные по литологии прибрежных фаций туртасской свиты (верхний олигоцен, юг Западной Сибири) // Докл. АН. 2017. Т. 475. № 6. С. 1–4.
Стандартный образец состава байкальского ила, БИЛ-1 (ГСО 7126-94) // Перечень стандартных образцов состава минеральных веществ, разработанных Институтом геохимии СО РАН. Каталог стандартных образцов состава природных и техногенных сред. Иркутск, 2009а. С. 28–29.
Стандартный образец состава донных отложений оз. Байкал, БИЛ-2 (ГСО 7176-95) // Перечень стандартных образцов состава минеральных веществ, разработанных Институтом геохимии СО РАН. Каталог стандартных образцов состава природных и техногенных сред. Иркутск, 2009б. С. 30–31.
Унифицированные региональные стратиграфические схемы палеогеновых и неогеновых отложений Западно-Сибирской равнины. Объяснительная записка и схема. Новосибирск: Изд-во СНИИГГиМС, 2001. 84 с.
Усольцева М.В., Титова Л.А. Ископаемые пресноводные диатомовые водоросли Тункинской впадины (Прибайкалье) по материалам бурения 2013 г. // Вопросы современной альгологии. 2015. Спец. вып. С. 170–172.
Хурсевич Г.К., Рубина Н.В. Новая таксономическая комбинация в роде Actinocyclus (Bacillariophyta) // Ботан. журн. 1991. Т. 76. № 8. С. 1151–1154.
Хурсевич Г.К., Моисеева А.И., Козыренко Т.Ф., Рубина Н.В. Новые таксоны рода Actinocyclus (Bacillariophyta) из неогеновых пресноводных отложений СССР // Ботан. журн. 1990. Т. 75. № 10. С. 1439–1442.
Цой И.Б., Усольцева М.В. Миоценовые пресноводные диатомеи из отложений подводного плато Уллын (возвышенность Криштофовича) Японского моря // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 3. С. 62–80.
Шацкий С.Б. Основные вопросы стратиграфии и палеогеографии палеогена Западной Сибири // Палеоген и неоген Сибири (палеонтология и стратиграфия). Новосибирск: Наука, 1978. С. 3–21.
Шацкий С.Б., Александрова Л.В., Богдашев В.А. и др. Палеоген центральных районов Западной Сибири // Морской и континентальный палеоген Сибири. Новосибирск: Наука, 1973. С. 4–39.
Шешукова-Порецкая В.С., Глезер З.И. Диатомовые водоросли, силикофлагеллаты и эбриидеи из майкопских отложений из р. Шибик // Ученые записки ЛГУ. Сер. биол. 1962. Т. 313. С. 171–205.
Шумилова Е.В. Терригенные компоненты мезозойских и кайнозойских отложений Западно-Сибирской низменности и их роль в палеогеографических реконструкциях. Новосибирск: Изд-во СО АН СССР, 1963. 314 с.
Юдович Я.Э. Региональная геохимия осадочных толщ. Л.: Наука, 1981. 276 с.
Benson M.L., Kociolek J.P., Spaulding S.A., Smith D.M. Pre-Neogene non-marine diatom biochronology with new data from the late Eocene Florissant Formation of Colorado, USA // Stratigraphy. 2007. V. 9. № 2. P. 131–152.
Houk V. Aulacoseira hibschii (Reichelt) Houk from its type locality and other diatomite localities in North Bohemia (Czech Republic and Germany) // Proc. 1st Central European Diatom Meeting. Eds. Kusber W.-H., Jahn R. Berlin, 2007. P. 55–58. https://doi.org/10.3372/cediatom.111
Khursevich G., Kociolek J.P. A preliminary, worldwide inventory of the extinct, freshwater fossil diatoms from the orders Thalassiosirales, Stephanodiscales, Paraliales, Aulacoseirales, Melosirales, Coscinodiscales, and Biddulphiales // Nova Hedwigia. 2012. Beih. 141. P. 315–364.
Kotlarczyk J., Kaczmarska I. Two diatom horizons in the Oligocene and (?) Lower Miocene of the Polish Outer Carpathians // Ann. Soc. Geol. Pol. 1987. № 57. P. 143–188.
Manconi R., Pronzato R. Spongillina subord. nov. Lubomirskiidae, Malawispongiidae n. fam., Metaniidae, Metschnikowiidae, Potamolepidae, Spongillidae // Systema Porifera: a guide to the classification of sponges. Eds. Hooper J.N.A, van Soest R.W.M. Kluwer Academic/Plenum New York Publishers, 2002. P. 921–1019.
Nesbitt H.W., Young G.M. Early Proterozoic climates and plate motions inferred from major element chemistry of lutites // Nature. 1982. V. 299. P. 715–717.
Oreshkina T.V. Transfer of extant non-marine diatom Melosira ignota Rubina from Late Oligocene of West Siberia to the genus Pseudoaulacosira Lupikina et Khursevich // Nova Hedwigia. 2018. Beih. 147. P. 183–192.
Shiller W. Siliceous microfossils from the Oligocene Tripoli-deposits of Seifhennersdorf // Sbornik narodniho muzea v Praze acta musei nationalis pragae. Series B. Historia Naturalis. 2007. V. 63. № 2–4. P. 195–205.
Simonsen R. The Diatom system: ideas on phylogeny // Bacillaria. 1979. V. 2. P. 9–71.
Usoltseva M., Houk V. Morphology and taxonomy of an almost forgotten fossil diatom, Gaillonella sculpta Ehrenberg (Bacillariophyceae) // Diatom Res. 2016. V. 31. P. 39–49. http://dx.doi.org/10.108 0/0269249X.2016.1143034
Yanochkina E.A. Statistical methods for studying variegated. Moscow: Nedra, 1966. 143 p.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция