

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 3, стр. 148-160
Развитие растительности и микротериофауны в позднеледниковье и голоцене на территории национального парка “Югыд-Ва”
Ю. В. Голубева 1, *, И. В. Кряжева 1
1 Институт геологии им. акад. Н.П. Юшкина Коми научного центра УрО РАН
Сыктывкар, Россия
* E-mail: yvgolubeva@geo.komisc.ru
Поступила в редакцию 10.09.2018
После доработки 31.10.2018
Принята к публикации 28.03.2019
Аннотация
Обсуждаются результаты изучения позднеплейстоценовых и голоценовых спорово-пыльцевых спектров и микротериофауны рыхлых отложений четырех пещерных местонахождений на западном склоне Приполярного Урала, расположенных в бассейнах рек Щугер и Кожым на территории национального парка “Югыд-Ва”. Определения возраста осадков карстовых полостей базируются на данных радиоуглеродного датирования костей млекопитающих методом AMS. Спелеогенные образования изученных местонахождений содержат слои завершающего этапа деградации полярноуральского (поздневалдайского) оледенения со среднего по поздний дриас и трех периодов голоцена – пребореального, бореального и субатлантического. Для аллерёда выявлен ископаемый комплекс микротериофауны с преобладанием криоксерофильных тундровых видов (88.3%). Спорово-пыльцевыми спектрами зафиксированы холодный и теплый периоды позднеледниковья – средний дриас и аллерёд соответственно. К концу позднего дриаса на западном склоне Приполярного Урала микротериофауна остается достаточно холодолюбивой. Для этого времени установлено распространение ерниковых зарослей и марево-злаково-разнотравных сообществ. В позднепребореальное похолодание растительность западных предгорий Приполярного Урала представляла собой травяно-кустарниковую тундру с единичными деревьями ели. Для сообщества микротериофауны также был характерен тундроподобный облик. Установлено, что в бореальное время на юге Приполярного Урала произрастали таежные темнохвойные леса, а на севере – редкостойные елово-березовые леса с участием сосны и карликовой березкой в подлеске. Фауна грызунов к этому времени становится лесной. К середине субатлантического периода на западном склоне Приполярного Урала получили развитие северотаежные темнохвойные леса из ели со значительной примесью сосны, а к концу этого периода в составе древостоев увеличилось количество тундровых элементов. Микротериофауна исследуемого региона имела типичный лесной облик, характерный для всех таежных позднеголоценовых сообществ северо-востока Европы.
ВВЕДЕНИЕ
Палинологическое изучение рыхлых отложений пещер и других карстовых образований позволяет получить информацию о характере растительного покрова горных территорий, а исследования костных остатков дают представление о состоянии фаунистических сообществ прошлого и значительно облегчают датировку отложений и интерпретацию полученных аналитических данных. В связи с этим спелеогенные осадки вызывают все больший интерес.
Полигоном для такого рода исследований выступает Урал. Здесь в карстующихся палеозойских породах образуется большое количество пещер, гротов, навесов и ниш.
В общих чертах позднечетвертичная история формирования микротериофауны западного склона Приполярного Урала была опубликована в ряде работ (Пономарев, Кряжева, 2011; Кряжева, 2012; Кряжева и др., 2012; Кряжева, Пономарев, 2014). Результаты палинологического исследования этого района приводятся лишь С. Култти с соавторами (Kultti et al., 2003). Палинологическая характеристика отложений других регионов запада Урала, в том числе спелеогенного генезиса, отсутствует. Сведения о растительности позднеледниковья и голоцена получены преимущественно для восточного склона Урала (Федорова, 1951; Маковский, 1966; Сурова и др., 1975; Кошкарова и др., 1999; Панова и др., 2003; Jankovska et al., 2006 и др.). Интересные материалы получены Е.Г. Лаптевой непосредственно по рыхлым спелеогенным осадкам (Лаптева, 2005, 2009).
Работы по изучению динамики развития растительности и климата в голоцене в горных районах западного склона Приполярного Урала были проведены С. Култти с соавторами (Kultti et al., 2003) с применением спорово-пыльцевого и диатомового методов, анализа состава рецентных остатков кладоцер и растительных макроостатков, а также радиоуглеродного датирования. На основе реконструкции перемещения вертикальной границы лесной растительности сделан вывод о наиболее теплом времени голоцена в бореальном периоде, когда летние температуры были на 2°С и более выше, чем сейчас. Атлантическое и суббореальное потепления, по мнению С. Култти с соавторами (Kultti et al., 2003), проявились слабее. Другой группой исследователей (Кошкарова и др., 1999) бореальный период также выделяется как наиболее теплый в голоцене для восточного склона Полярного Урала.
В то же время это предположение не находит подтверждения в публикациях по палинологии сопредельных районов Полярного Урала (Сурова и др., 1975; Панова и др., 2003; Jankovska et al., 2006), Северного Урала (Маковский, 1966; Лаптева, 2009), Северного Предуралья (Немкова, 1976) и других более южных уральских регионов (Немкова, 1976), характеризующих атлантический период как самый влажный и теплый.
Цель настоящей работы – комплексные палеонтологические исследования, которые, учитывая недостаточный объем данных по палинологии и микротериологии, а также имеющиеся расхождения в интерпретации климатических условий в голоцене на территории исследований, представляются весьма актуальными и позволят дополнить и уточнить представления о ходе развития природы Приполярного Урала в позднеледниковье и голоцене.
РАЙОН, ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Были исследованы поздненеоплейстоценовые и голоценовые отложения четырех гротов, расположенных на западном макросклоне Урала, в бассейнах рек Щугер и Кожым (Приполярный Урал) на территории, принадлежащей национальному парку “Югыд-Ва” (рис. 1).
Рис. 1.
Географическое положение местонахождений. a – пещерные местонахождения, б – озерные и речные местонахождения, в – национальный парк “Югыд-Ва”. Местонахождения: 1 – Кожым-1, Кожым-2; 2 – Щугер-1, Щугер-2, Щугер-4; 3 – Соколиный (Пономарев, 2005); 4 – Вангыр, Межгорное (Kultti et al., 2003); 5 – Уса-2 (Кряжева и др., 2018); 6 – Шарью-1 (Кряжева и др., 2016); 7 – Пымва-Шор (Смирнов и др., 1999); 8 – Черемухово-1, Усольцевская пещера, Каква-4 (Лаптева, 2009); 9 – Дутово (Немкова, 1976); 10 – Карпушовка (Никифорова, 1979); 11 – бугристые торфяники (Пастухов и др., 2017); 12 – Малая Хадата, Большая Лагорта (Сурова и др., 1975), Черная горка (Jankovska et al., 2006), оз. Перевальное (Панова и др., 2003), Рай-Из (Кошкарова и др., 1999). Звездочкой выделены впервые изученные в настоящей работе местонахождения.
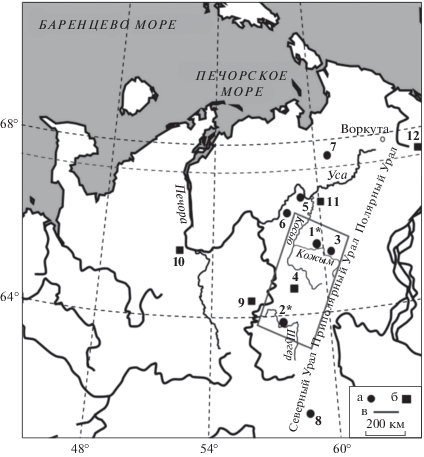
Местонахождения Щугер-2 и Щугер-4 располагаются на правом берегу р. Щугер в скальном выходе каменноугольных известняков (64°21.395′ с.ш., 58°28.401′ в.д., 200 м над уровнем моря (н. у. м.)), называемом Верхние Ворота, в 20 м выше по течению от устья ручья Велдор-Кыртаель на расстоянии 50 м друг от друга.
Местонахождение Щугер-4 представляет собой грот протяженностью 9 м, шириной 4 м, высотой 3 м и находится на высоте 7 м от уреза воды. В строении разреза рыхлых отложений выделены четыре слоя, представленные суглинистыми (слой 4), супесчано-суглинистыми осадками с большим количеством грубообломочного материала (слой 3), переходящими вверх по разрезу в дресвяно-щебнисто-глыбовые (слой 2) и алевритовые (слой 1) отложения. Палеонтологические остатки содержатся в слоях 1, 3 и 4.
Местонахождение Щугер-2 представляет собой нишу шириной 6 м и глубиной 1.5 м и находится на высоте 25 м от уреза воды. Раскопом площадью 1 м2 вскрыт слой, представленный супесчаными отложениями с большим содержанием корней растений и грубообломочного материала, мощностью 0.55 м.
Местонахождения Кожым-1 и 2 расположены на правом берегу р. Кожым в 7 км выше устья р. Сывью и в 1 км ниже по течению от скалы Каюк-Нырд в скальном выходе известняков ордовика на расстоянии 50 м друг от друга (65°40.642′ с.ш., 59°55.434′ в.д., 200 м н. у. м.).
Местонахождение Кожым-1 – маленькая карстовая полость длиной 2.4 м, шириной 1.1 м, высотой 1.1 м. Разрез рыхлых осадков состоит из трех слоев, представленных глыбово-щебнисто-дресвяной смесью почти без заполнителя (слой 3), переходящей вверх по разрезу в суглинок и алеврит (слои 2 и 1). Два верхних слоя содержат остатки позвоночных животных.
Местонахождение Кожым-2 представлено нишей шириной 2.5 м, высотой 1.6 м и глубиной 1.7 м. В разрезе вскрываются два горизонта песчано-алевритовых (слой 2) и супесчаных осадков (слой 1) с большим количеством грубообломочного материала и костными остатками позвоночных.
Современная растительность в районе исследований представлена предгорными еловыми и березовыми лесами. По склонам вдоль р. Щугер распространены елово-пихтовые кустарничково- или кислично-зеленомошные и папоротниковые леса, иногда с примесью кедра. К востоку, где начинает проявляться высотная поясность растительности, эти леса располагаются на высоте до 550 м н. у. м. Выше произрастают редкостойные елово-березовые и березовые крупнотравные леса и горные луга, поднимающиеся до 700 м н. у. м. Еще выше местообитания заняты ивняковыми, ерниковыми и можжевельниковыми зарослями, горными тундрами. Согласно климатическому районированию (Атлас…, 1964), в пределах северотаежной провинции климат характеризуется очень холодной длительной зимой и прохладным летом. Средняя температура января –18…–21°С, июля 12–16°С, годовая сумма осадков 600–700 мм и более. В бассейне р. Кожым, вследствие менее благоприятных климатических и почвенных условий для древесной растительности, развиты угнетенные редкостойные таежные леса. Заболоченность составляет 70% территории. По приречным склонам в растительном покрове преобладают елово-лиственничные леса, на междуречьях – ельники сфагновые, березовые и березово-еловые заболоченные редколесья (Леса…, 1999). Крайнесеверотаежная провинция расположена на границе приполярного равнинного и центрального горного климатических районов, характеризующихся длительной суровой зимой и прохладным летом (Атлас…, 1964). Средняя температура января ‒18…–20°С, июля 10–12°С, годовая сумма осадков 500–700 мм.
Всего из отложений четырех гротов определенно более 9000 зубов мелких млекопитающих (табл. 1).
Таблица 1.
Количество щечных зубов и соотношение остатков (в скобках, %) мелких млекопитающих из местонахождений западного склона Приполярного Урала
| Вид | Щугер-4 | Щугер-2 | Кожым-1 | Кожым-2 | Биом* | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Сл. 1 | Сл. 2 | Сл. 3 | Сл. 1 | Сл. 1 | Сл. 2 | Сл. 1 | Сл. 2 | с | т | л | и | |
| Степная пищуха – Ochotona pusilla | – | 20(1) | 6(1.7) | – | – | 1(0.1) | – | – | + | – | – | – |
| Белка – Sciurus vulgaris | – | – | – | 165(11.6) | 4(1.8) | – | 3(0.5) | – | – | – | + | – |
| Лесная мышовка – Sicista betulina | – | – | – | – | – | – | 5(1) | – | – | – | + | – |
| Красно-серая полевка – Craseomys rufocanus | 77(4.6) | 21(0.8) | – | 222(15.7) | 14(6.5) | 28(1.7) | 43(6.2) | 34(7) | – | – | + | – |
| Рыжая полевка – Myodes glareolus | 44(2.6) | 18(0.7) | – | 66(4.7) | 15(7.3) | 15(0.9) | 46(6.7) | 62(13) | – | – | + | – |
| Красная полевка – Myodes rutilus | 104(6.2) | 32(1.2) | 4(1.2) | 74(5.2) | 19(8.9) | 48(2.9) | 78(11.4) | 94(19.5) | – | – | + | – |
| Копытный лемминг – Dicrostonyx sp. | 234(13.9) | 416(15.8) | 53(15.7) | – | 3(1.5) | 205(12.8) | – | – | – | + | - | – |
| Сибирский лемминг – Lemmus sibiricus | 407(24.2) | 954(36.2) | 139(43.3) | 46(3.7) | 6(2.8) | 163(10.3) | 11(1.6) | 24(5) | – | + | - | – |
| Лесной лемминг – Myopus schisticolor | 90(5.4) | 173(6.4) | 15(4.4) | 82(5.8) | 13(6) | 27(1.6) | 34(5) | 14(2.9) | – | – | + | – |
| Водяная полевка – Arvicola amphibius | 130(7.6) | 49(1.8) | 2(1) | 594(41) | 83(38.7) | 493(30.6) | 85(12.3) | 141(29.3) | – | – | – | + |
| Узкочерепная полевка – Lasiopodomys gregalis | 318(18.9) | 666(25.3) | 99(29.3) | 17(1.8) | 13(6) | 198(12.3) | 7(1) | 13(2.7) | + | + | – | – |
| Полевка экономка – Alexandromys oeconomus | 122(7.4) | 95(3.6) | 8(2.2) | 65(4.6) | 21(9.8) | 263(16.3) | 83(12.1) | 24(5) | – | – | – | + |
| Темная полевка – Microtus agrestis | 155(9.2) | 190(7.2) | 4(1.2) | 84(5.9) | 23(10.7) | 170(10.5) | 289(42.2) | 75(15.6) | – | – | + | – |
| Всего | 1680 | 2634 | 337 | 1415 | 214 | 1611 | 684 | 481 | ||||
Раскопки в гротах проводили по стандартным методикам (Методическое…, 1955, 1987; Методы…, 1957). Отбор проб из осадков для спорово-пыльцевого анализа и их лабораторную обработку осуществляли по общепринятым методикам (Гричук, Заклинская, 1948). Расчет результатов анализа проводили групповым способом.
Реконструкции климатических параметров (средних температур июля, января и года, среднего годового количества осадков) проводили с помощью зонально-формационного метода (Савина, Хотинский, 1982), успешно примененного Е.Г. Лаптевой (2009) для получения палеоклиматических показателей на восточном склоне Северного Урала.
Для расчета доли каждого вида полевок все зубы, определенные как Microtus sp., разделяли по видам в соответствии с распределением первых нижних коренных. Морфологически сходные виды, такие как полевка Миддендорфа–темная полевка и лесной лемминг–сибирский лемминг, диагностика которых представляет определенные трудности, были разделены по методике, описанной Н.Г. Смирновым с соавторами (Смирнов и др., 1997). Моляры трех видов родов Myodes и Craseomys идентифицировались согласно методу, предложенному А.В. Бородиным с соавторами (Бородин и др., 2005).
Группировку видов грызунов по биологическим и экологическим свойствам осуществляли на основе общеизвестных особенностей биологии и экологии видов, как, например, это сделано в работе Т.В. Фадеевой и Н.Г. Смирнова (2008).
Радиоуглеродное (14С) датирование прямым масс-спектрометрическим методом (AMS) по костям грызунов (табл. 2) было выполнено Й. ван дер Плихтом (Центр изотопных исследований университета г. Гронинген, Нидерланды). В данной работе приведены некалиброванные значения радиоуглеродного возраста.
Таблица 2.
Результаты АМS радиоуглеродного датирования спелеогенных отложений на территории национального парка “Югыд-Ва”
| № п/п |
Местонахождение | Лаб. индекс и номер образца | Радиоуглеродный возраст, лет | σ | Калиброванная дата, лет |
|---|---|---|---|---|---|
| 1 | Щугер-4, слой 3 | GrA- 49439 | 11 850 | 60 | 13 599–13 733 |
| 2 | Щугер-4, слой 1 | GrA-49440 | 10 090 | 60 | 11 408–11 439 |
| 3 | Кожым-1, слой 2 | GrA-66908 | 9360 | 190 | 10 267–10 786 |
| 4 | Кожым-2, слой 2 | GrA-49355 | 1655 | 35 | 1528–1574 |
| 5 | Щугер-2, слой 1 | GrA-49352 | 950 | 35 | 799–816 |
Примечание. Радиоуглеродные датировки калибровали в компьютерной программе “Calib” (http://calib.qub.ac.uk/calib/), используя шкалу intcal13 (Reimer et al., 2013).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В местонахождении Щугер-4 на основании видового состава микротериофауны и морфологии коренных зубов копытного лемминга и узкочерепной полевки установлены два ископаемых комплекса. Первый комплекс (слои 2 и 3) – умеренный криоксерофильный с преобладанием тундровых видов (88.3%). Отложения в основании разреза (слой 3, гл. 0.9 м) датированы 11850 ± 60 лет (обр. GrA-49439) (Пономарев, Кряжева, 2011), что соответствует границе среднедриасового и аллерёдского периодов позднеледниковья. По палинологическим данным (обр. 12, гл. 1 м), накопление осадков слоя, скорее всего, происходило в среднем дриасе (рис. 2). Этот период характеризовался развитием тундроподобных ерниковых зарослей (содержание пыльцы Betula nana – 72% от суммы пыльцы древесных растений, Betula sect. Fruticosae – 13%) и травянистых сообществ из разнотравья, злаков и маревых. Климат был значительно холоднее, чем сейчас, и, вероятно, напоминал современные климатические условия в пределах предгорных ерниковых тундр Полярного Урала (средние температуры июля 8–10°С, января ‒20°С, года ниже –7°С, сумма годовых осадков 400–500 мм). Выше по разрезу спорово-пыльцевыми спектрами слоя 2 (обр. 11, гл. 0.4–0.5 м) фиксируется интерстадиальный период позднеледниковья – аллерёд. Улучшение климатических условий способствовало распространению по речным долинам елово-сосновых редколесий, на что указывает увеличение количества пыльцы хвойных пород – ели и сосны (с единичных зерен до 9 и 19% соответственно). Также выявлены следующие особенности спорово-пыльцевых спектров аллерёда. В общем составе преобладают споры (до 70%). В группе трав снижается участие пыльцы ксерофитов (Chenopodiaceae и Ephedra distachya) за счет возрастания роли пыльцы разнотравья (до 86%), среди которой определено повышенное содержание пыльцы цикориевых (52%), характерных для нарушенных или несформированных почв после пожаров или освободившихся от ледников. В видовом составе спор, как и в предыдущем спектре, доминирует Polypodiaceae (75%).
Рис. 2.
Сводная спорово-пыльцевая диаграмма по разрезам спелеогенных отложений на Приполярном Урале. 1 – алеврит, 2 – суглинок, 3 – супесь, 4 – песок, 5 – валуны, 6 – дресва и щебень, 7 – номер слоя.
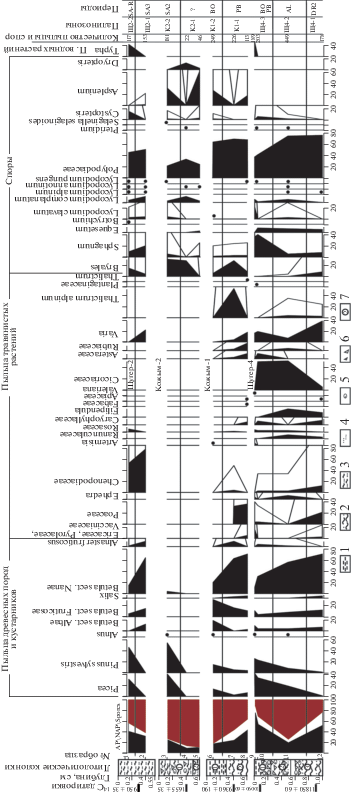
Второй комплекс фауны выделен в слое 1. Здесь остатки тундровых видов также преобладают, но их доля составляет только 57%, на лесные виды приходится 28% и на интразональные – 15%. По нижней челюсти копытного лемминга из верхней части слоя (слой 1, гл. 0.08 м) получена радиоуглеродная датировка 10090 ± 60 лет (обр. GrA-49440), что соответствует раннему пребореалу. Ранее нами было установлено, что фауна географически близко расположенного грота Соколиный (т. 3 на рис. 1; Пономарев, 2005) практически идентична по составу и структуре ископаемому комплексу из слоя 1 грота Щугер-4 и имеет радиоуглеродную датировку 10390 ± 50 лет (обр. GrA-42215), что соответствует позднему дриасу. В фауне грота Соколиный на тундровые виды приходится 54%, на лесные – 35.5% и на околоводные – 10.5% (Пономарев, Кряжева, 2011; Кряжева, 2012; Кряжева и др., 2012). Учитывая, что датировка в гроте Щугер-4 получена по материалу из самой верхней части слоя 1 при его мощности 0.4 м, а также полное сходство комплекса фауны этого грота с позднедриасовым фаунистическим комплексом слоя 2 грота Соколиный, мы считаем, что фаунистический комплекс слоя 1 грота Щугер-4 отражает позднедриасовую фазу истории микротериофауны. Позднедриасовая растительность не охарактеризована вследствие отсутствия материала для палинологического анализа.
Выше по разрезу по спорово-пыльцевому спектру (обр. 10, гл. 0.05 м) фиксируется потепление климата. Рост содержаний пыльцы хвойных пород Picea sp. и Pinus sylvestris и падение количества пыльцы Betula nana указывают на увеличение площадей, занимаемых древесными формациями. В группе трав содержание пыльцы цикориевых остается повышенным (50%). Среди спор доля Polypodiaceae (45%) сокращается за счет увеличения содержания Sphagnum (41%). Вероятно, полученный спорово-пыльцевой спектр отражает произрастание еловых редколесий в первой половине пребореального периода, когда на европейском северо-востоке России происходило массовое распространение лесных формаций (Никифорова, 1979; Непомилуева, Дурягина, 1990).
Спорово-пыльцевой спектр кровли разреза (обр. 9, гл. 0.01 м) с преобладанием пыльцы хвойных пород Picea sp. и Pinus sylvestris (42 и 26% соответственно) и со спорами лесных видов плаунов свидетельствует о произрастании таежных темнохвойных лесов и климате, близком к современному. На дальнейшее потепление, по сравнению с предыдущей фазой, указывает и присутствие пыльцы Typhа latifolia (рис. 2). В пределах территории наших исследований в настоящее время ареал этого вида охватывает бассейны рек Вычегда и Вымь. Учитывая, что сомкнутый лесной покров на сопредельных территориях, в Северном Предуралье и на западном склоне Приполярного Урала, был сформирован в бореале (Немкова, 1976; Kultti et al., 2003), можно предположить, что осадконакопление в гроте Щугер-4 закончилось в этот период.
По данным микротериологических исследований и пыльцевого анализа, в местонахождении Кожым-1 выделены два ископаемых фаунистических комплекса и две соответствующие им палинозоны. Первый фаунистический комплекс, выделенный в слое 2, состоит из достаточно криофильных видов. В нем на тундровые и степные виды приходится 36%, на интразональные – 47% и на лесные – 17% (Кряжева, Пономарев, 2014). К сожалению, данное сообщество не имеет датированных аналогов в регионе. Оно заметно отличается от позднедриасового фаунистического комплекса грота Щугер-4 более низкой долей тундровых видов, а от бореального сообщества грота Уса-2 (т. 5 на рис. 1; Кряжева и др., 2018), наоборот, более высоким содержанием тундровых видов и очень низким количеством лесных (рис. 1).
Спорово-пыльцевые спектры нижней части разреза (обр. 7, гл. 0.4 м; обр. 8, гл. 0.6 м) свидетельствуют о преобладании в растительном покрове тундроподобных кустарниковых зарослей. Значительная роль принадлежала кустарниковым березам (Betula nana 64–72%, Betula sect. Fruticosae 11–16%), ивам, ольховнику и травянистым сообществам из разнотравья (64%), злаков (36%) и маревых. Присутствие в спектрах пыльцы ели (3–7% от суммы пыльцы древесных растений) указывает на произрастание лишь отдельных ее деревьев. Растительность была подобна позднеледниковому растительному комплексу с преобладанием ерниковых зарослей и травянистых группировок из разнотравья, злаков и маревых, установленному нами в разрезе Щугер-4. Выявленные особенности развития растительности, состава и структуры микротериофауны, а также радиоуглеродная датировка 9360 ± 190 лет (обр. GrA-66908) позволяют связать накопление осадков в основании разреза с концом пребореального периода голоцена. В пребореале на северо-востоке Русской равнины и Полярном Урале растительный покров еще сохранял черты перигляциального комплекса с преобладанием травяно-кустарниковых сообществ в сочетании с редколесьями (Никифорова, 1979; Панова и др., 2003). Ухудшение климатических условий во второй половине пребореального периода способствовало увеличению в составе растительности тундровых элементов (Сурова и др., 1975; Никифорова, 1979). Можно предположить, что реконструированные климатические параметры были схожи с таковыми в современной зоне распространения горных тундр Приполярного Урала: средние температуры июля 9–10°С, января –18…–19°С, года –5…–6°С, сумма годовых осадков 600–700 мм и более.
По сравнению с ископаемым комплексом из слоя 2 в сообществе мелких млекопитающих слоя 1 доля тундровых и степных видов сокращается до 10%, а доля лесных видов увеличивается до 41%. На интразональные виды приходится почти 50% от всех остатков. Похожая картина наблюдается в сообществе слоя 2 грота Уса-2 и слоя 3 местонахождения Пымва-Шор, которые имеют радиоуглеродные датировки 8470 ± 45 лет (обр. GrA-66466) и 8500 ± 250 лет (обр. ГИН-9005), что отвечает середине бореального периода (Смирнов и др., 1999; Кряжева и др., 2018). Фауна там имеет типичный лесной облик, где доминируют лесные и интразональные виды (44.8 и 44.9% соответственно), а на тундровые и степные виды приходится всего 10%.
Состав пыльцы и спор в верхнем слое разреза Кожым-1 (обр. 6, гл. 0.1 м) также изменяется и, вероятно, отражает потепление климата в бореале, которое способствовало распространению редкостойных елово-березовых лесов. На это указывает более частая встречаемость в группе древесных (20%) пыльцы деревьев, в основном представленной Betula sect. Albae (19%), Pinus sylvestris (15%) и Picea sp. (13% от суммы пыльцы древесных). В растительном покрове значительно снизилась роль криофильных кустарников, о чем свидетельствует сокращение их пыльцы в спектре с 70 до 20% от суммы пыльцы древесных. Травяной покров был представлен папоротниками и разнотравно-злаковыми ассоциациями.
В сообществе мелких млекопитающих Кожым-2 на долю тундровых видов приходится 7.7%, на лесные – 58% и на интразональные – 34.3%. Выше по разрезу участие тундровых и интразональных видов сокращается до 2.6 и 24.4% соответственно, а содержание остатков лесных видов увеличивается до 73%. В составе лесных видов появляется белка. Одновозрастные фауны, датируемые субатлантиком и имеющие схожий состав и структуру, известны из местонахождений Кожым-4 (1620 ± 30 лет, обр. GrA-66465) и Шарью-1 (1510 ± 35 лет, обр. GrA-66862). Там также преобладают лесные виды (60–70% от всех остатков), околоводные виды составляют 27–40%, а тундровые – всего 2–5% (Кряжева, Пономарев, 2014; Кряжева и др., 2016). Из представленных материалов видно, что в субатлантическое время микротериофауна Приполярного Урала имела типичный лесной облик. Этот вывод не противоречит полученным палинологическим данным. Отложения верхней части разреза (обр. 3, гл. 0.12 м) содержат комплекс спор и пыльцы с доминированием спор (67%) и пыльцы древесных растений (31%). В группе пыльцы древесных преобладают Picea sp. (36%) и Pinus sylvestris (59%). В составе спор доминируют споры папоротников (54%) и зеленых мхов (31%). Обнаружены споры сфагновых мхов, таежных видов плаунов Lycopodium clavatum и L. complanatum, северотаежно-лесотундрового вида Selaginella selaginoides и тундрового арктоальпийского вида L. pungens. Состав комплекса указывает на произрастание на территории исследований редкостойных лесов из ели со значительной примесью сосны при обилии скальных (родов Asplenium и Cystopteris) и лесных (рода Dryopteris и др.) папоротников. Климатические условия были приближены к современным.
В палеотериологическом материале местонахождения Щугер-2 выделен лесной тип фауны. Наряду с остатками лесных видов (49.8%) отмечено значительное участие интразональных видов (44.7%), что объясняется тафономическими причинами, в основном пищевой специализацией хищника. Остатки тундровых видов грызунов (Lemmus sibiricus, Lasiopodomys gregalis) были обнаружены только в нижней части разреза, их доля составляет 5.5% от всего числа остатков (Пономарев, Кряжева, 2011; Кряжева, 2012; Кряжева и др., 2012) (табл. 1). Спорово-пыльцевой спектр нижней части слоя (обр. 2, гл. 0.4 м) свидетельствует о преобладании в растительном покрове кустарниковых зарослей из карликовой березки (Betula nana 65%, Betula sect. Fruticosae 15%) и ольховника (9%) с березовыми редколесьями (Betula sect. Albae 11%). Распространение тундровой и лесотундровой растительности на месте современной северной тайги указывает на более холодный климат, чем в настоящее время, близкий к нынешним условиям в районах распространения горных тундр на Северном и Приполярном Урале (средние температуры июля 10–14°С, января –18…–21°С, года ‒4…–5°С, сумма годовых осадков 600–700 мм и более). Палинологическая характеристика и датировка 950 ± 35 лет (обр. GrA-49352) позволяют связать формирование рыхлых отложений в основании разреза с периодом SA-3, так называемой “малой ледниковой эпохой”. Состав спектра верхней части слоя (обр. 1, гл. 0.15 м) указывает на значительное сокращение площадей, занятых ерниковыми зарослями (Betula nana 15%, Betula sect. Fruticosae 6%), и распространение в предгорьях Северного Урала елово-сосновых лесов (Picea sp. 31% и Pinus sylvestris 46%), что обусловлено потеплением климата в настоящее время (рис. 2).
ВЫВОДЫ
Рыхлые осадки изученных карстовых полостей, несмотря на их малую мощность по сравнению с разрезами торфяников, оказались достаточно информативными при реконструкциях палеогеографических обстановок позднеледниковья и голоцена. Спелеогенные отложения содержат позднеледниковые слои аллерёда и среднего дриаса, которые не были ранее палинологически охарактеризованы при исследованиях торфяных образований на изученной территории, а также слои трех периодов голоцена – пребореального, бореального и субатлантического. Наиболее древние, позднеплейстоценовые, фаунистические и спорово-пыльцевые комплексы обнаружены нами в местонахождении Щугер-4. Осадконакопление происходило в ледниковых условиях, в течение заключительной стадии деградации полярноуральского (поздневалдайского) оледенения. Для аллерёда выявлен умеренный криоксерофильный ископаемый комплекс микротериофауны с преобладанием тундровых видов (88.3%) (Пономарев, Кряжева, 2011; Кряжева, 2012; Кряжева и др., 2012). Спорово-пыльцевыми спектрами фиксируются холодный и теплый периоды позднеледниковья – средний дриас и аллерёд соответственно. В среднем дриасе в западных предгорьях Приполярного Урала произрастали ерниковые заросли и марево-злаково-разнотравные сообщества. Потепление в аллерёде вызвало некоторое вытеснение ерниковых зарослей елово-сосновыми редколесьями (рис. 3).
Рис. 3.
Развитие растительности и микротериофауны на территории национального парка “Югыд-Ва” в позднеледниковье и голоцене. Пунктирной линией показано развитие растительности в пределах современной подзоны северной тайги, сплошной линией – в пределах современной подзоны крайнесеверной тайги. Тайга крайнесеверная: 1 – елово-березовые и сосновые редкостойные леса, 2 – сосново-еловые редкостойные леса; тайга северная: 3 – с преобладанием березовых и лиственничных лесов, 4 – с преобладанием еловых лесов.
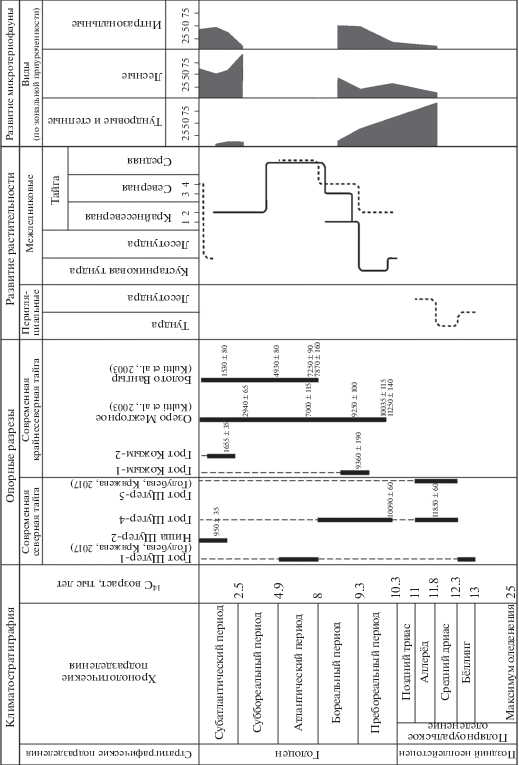
К концу позднего дриаса на западном склоне Приполярного Урала микротериофауна оставалась достаточно криофильной. Остатки тундровых видов этого времени также доминируют, но их доля снижается с 88 до 57%. По сравнению с предыдущим периодом в составе фауны заметно увеличилась доля видов лесных (с 5 до 18.8%) и околоводных (с 3 до 15%) местообитаний.
По данным Е.Г. Лаптевой (2009), южнее, на восточном макросклоне Северного Урала, в течение позднего дриаса характерным ландшафтом была перигляциальная лесостепь: на плакорах произрастали разнотравно-злаковые сообщества, на северных склонах и заболоченных участках – ерники, а по речным долинам – березовые и сосновые редколесья. Учитывая эти сведения, изученные нами среднедриасовые спорово-пыльцевые спектры, а также состав и структуру позднедриасового фаунистического комплекса, можно предположить, что на территории исследований ерниковые заросли и марево-злаково-разнотравные сообщества также имели распространение в этот период.
Отличительной особенностью спорово-пыльцевых спектров позднеледниковья Приполярного Урала, по сравнению с аналогичными спектрами северных районов Восточно-Европейской равнины (Никифорова, 1979; Zaretskaya et al., 2014), является меньшее содержание пыльцы травянистых растений (7–25%), в том числе и ксерофильных видов, и преобладание среди спор Polypodiaceae (51–75%), что свидетельствует о менее засушливых условиях в предгорьях Приполярного Урала.
Полученные в настоящей работе и ранее опубликованные данные (Kultti et al., 2003; Голубева, Кряжева, 2017) позволили выявить особенности развития фауны и растительности на протяжении голоцена в пребореальном, бореальном, атлантическом и субатлантическом периодах на юге Приполярного Урала и в течение пребореального, бореального, атлантического, суббореального и субатлантического периодов на севере этого региона. Охарактеризована биота первой и второй половины пребореального периода, с которыми соотносят раннепребореальное потепление и позднепребореальное похолодание в Тимано-Печоро-Вычегодском регионе (Никифорова, 1979), являющиеся аналогами половецкого потепления и переславльского похолодания на Русской равнине (Хотинский, 1987) и фиксирующиеся на спорово-пыльцевых диаграммах, полученных как для сопредельных равнинных, так и для горных территорий: южной части Полярного Урала (Сурова и др., 1975), междуречья Косъю–Кочмес–Лемва–Кожым (Пастухов и др., 2017) и долины р. Печора (Никифорова, 1979). Растительность западных предгорий Приполярного Урала в первой половине пребореального периода представляла собой еловые редколесья, сменившиеся во второй половине периода травяно-кустарниковой тундрой с единичными деревьями ели, возможно, наподобие растительного покрова в южной части восточных предгорий Полярного Урала (Сурова и др., 1975). Для сообщества микротериофауны также был характерен криофильный состав: тундровые и степные виды составляют 36% (с почти равным количеством остатков узкочерепной полевки, сибирского и копытного леммингов), на интразональные виды также приходится почти половина от всех остатков. Климатические условия были близки к современным.
Спорово-пыльцевые спектры бореального периода указывают на произрастание в южной части национального парка “Югыд-Ва” таежных темнохвойных лесов, а в северной части – редкостойных елово-березовых лесов с участием сосны и карликовой березкой в подлеске. По данным С. Култти с соавторами (Kultti et al., 2003), первой по склонам Приполярного Урала расселялась древовидная береза, затем распространились и заняли свои современные местообитания лиственница, пихта и ель. Фауна приобретает лесной облик: доля тундровых и степных видов сокращается до 10%, содержание остатков лесных видов, напротив, увеличивается до 41%, интразональные виды составляют 49%. Климатические условия, на наш взгляд, были близки к современным, что согласуется с палинологическими данными по восточному склону Северного Урала (Лаптева, 2009) и Полярного Урала (Сурова и др., 1975; Панова и др., 2003; Jankovska et al., 2006), но противоречит выводу другой группы исследователей (Kultti et al., 2003) о том, что на западном склоне Приполярного Урала климат в бореале был теплее, чем сейчас, а возможно, и наиболее теплым в голоцене.
С атлантическим и суббореальным периодами соотнесена фаза сфагновых сосново-еловых лесов с примесью пихты, выделенная в кровле ранее изученного разреза спелеогенных осадков Щугер-1 (Голубева, Кряжева, 2017), так как в Северном Предуралье и на Приполярном Урале пихта в составе сосново-еловых массивов, распространенных начиная с бореального периода, появилась в атлантике (Немкова, 1976; Kultti et al., 2003). К сожалению, отложения в разрезе Щугер-1 не датированы, поэтому корректное сопоставление с ландшафтно-климатическими условиями других периодов пока не представляется возможным. Некоторые исследователи (Kultti et al., 2003) объясняют увеличение доли пихты и лиственницы в составе лесов снижением границ высотных растительных поясов в результате постепенного похолодания. Однако, по нашему мнению, максимальное содержание пыльцы широколиственных пород (вяза, липы, дуба и орешника) на полученных ими диаграммах свидетельствует о внедрении в состав лесов в южной части национального парка представителей неморальной флоры, характерном и для сопредельных районов (Немкова, 1976; Лаптева, 2009), и указывает на климат существенно теплее современного.
Спорово-пыльцевые спектры субатлантического периода отражают три ландшафтно-климатических этапа, относящиеся к середине (Кожым) и концу периода (Щугер). В местонахождении Кожым-2 отложения датированы 1655 ± 35 лет (обр. GrA-49355). Здесь получили развитие редкостойные темнохвойные леса из ели со значительной примесью сосны при обилии папоротников. Климат был близок к современному. Отложения, вскрытые на юге Приполярного Урала в местонахождении Щугер-2, датированы 950 ± 35 лет (обр. GrA-49352). Данные их палинологического изучения свидетельствуют о том, что к концу субатлантика в составе древостоев увеличилось количество тундровых элементов, что, вероятно, было вызвано похолоданием. Затем в условиях современного потепления площади, занятые ерниковыми зарослями, значительно сократились. В субатлантическое время микротериофауна исследуемого региона имела типичный лесной облик, характерный для всех таежных позднеголоценовых сообществ северо-востока Европы, что хорошо согласуется с результатами пыльцевого анализа.
Источники финансирования. Работа выполнена в рамках темы “Биогеологические события и стратиграфия фанерозоя субарктической зоны Баренцевоморского региона, Тимана и западного склона Урала” ГР № AAAA-A17-117121140081-7 и при частичной поддержке Программы РАН № 15-18-5-41.
Список литературы
Атлас Коми АССР. М.: Изд-во Главного управления геодезии и картографии Государственного геологического комитета СССР, 1964.
Бородин А.В., Коурова Т.П., Маркова Е.А. Размерные характеристики щечных зубов лесных полевок Clethrionomys (Craseomys) rufocanus, Cl. (Clethrionomys) glareolus, Cl. (Cl.) rutilus (Arvicolinae, Rodentia) и их использование для видовой идентификации // Зоологический журн. 2005. Т. 84. № 2. С. 236–244.
Голубева Ю.В., Кряжева И.В. Развитие растительных и фаунистических сообществ голоцена на Приполярном Урале (по результатам изучения осадков пещерных местонахождений Щугер и Кожым) // Фундаментальные проблемы квартера: итоги изучения и основные направления дальнейших исследований. Материалы X Всеросс. совещания по изучению четвертичного периода. М.: ГЕОС, 2017. С. 101–103.
Гричук В.П., Заклинская Е.Д. Анализ ископаемых пыльцы и спор и его применение в палеогеографии. М.: ОГИЗ ГЕОГРАФГИЗ, 1948.
Кошкарова В.Л., Карпенко Л.В., Орлова Л.А. Динамика растительности и верхней границы леса в голоцене на Полярном Урале // Экология. 1999. № 2. С. 121–260.
Кряжева И.В. Мелкие млекопитающие позднего плейстоцена и голоцена Приполярного Урала. Автореф. дисс. … канд. геол.-мин. наук. Сыктывкар, 2012. 18 с.
Кряжева И.В., Пономарев Д.В. Микротериофауна западного склона Приполярного Урала в позднем плейстоцене и голоцене (р. Кожым) // Вестник Института геологии Коми НЦ УрО РАН. 2014. № 5. С. 3–9.
Кряжева И.В., Пономарев Д.В., ван Кольфсхотен Т., ван дер Плихт Й. История формирования современных сообществ микромаммалий Приполярного Урала // Экология. 2012. № 6. С. 434–441.
Кряжева И.В., Пономарев Д.В., ван Кольфсхотен Т., ван дер Плихт Й. Позднеголоценовые мелкие млекопитающие из местонахождений долины реки Шарью (гряда Чернышева) // Вестник Института геологии Коми НЦ УрО РАН. 2016. № 12. С. 16–22.
Кряжева И.В., Пономарев Д.В., ван Кольфсхотен Т., ван дер Плихт Й. Мелкие млекопитающие голоцена из пещерных местонахождений бассейна р. Усы (гряда Чернышева, северо-восток европейской части России) // Вестник Института геологии Коми НЦ УрО РАН. 2018. № 7. С. 34–40.
Лаптева Е.Г. Реконструкция этапов развития растительности Северного Урала в позднем неоплейстоцене и голоцене // Экология: от генов до экосистем. Материалы конференции молодых ученых. Екатеринбург: Академкнига, 2005. С. 141–151.
Лаптева Е.Г. Ландшафтно-климатические изменения на восточном склоне Северного Урала за последние 50 тыс. лет // Экология. 2009. № 4. С. 284–290.
Леса Республики Коми. Ред. Козубов Г.М., Таскаев А.И. М.: Издательско-продюсерский центр “Дизайн. Информация. Картография”, 1999.
Маковский В.И. О возрасте торфяников и формировании лесной растительности в подзоне северной тайги (междуречье Лозьвы и Пелыма) // Вопросы физиологии и геоботаники (Свердловск). 1966. Вып. 4. С. 53–63.
Методическое руководство по изучению и геологической съемке четвертичных отложений. М.: Госгеолтехиздат, 1955. Ч. 2.
Методическое руководство по изучению и геологической съемке четвертичных отложений. Л.: Недра, 1987.
Методы изучения осадочных пород. М.: Госгеолтехиздат, 1957. Т. 2.
Немкова В.К. История растительности Предуралья за поздне- и послеледниковое время // Актуальные вопросы современной геохронологии. М.: Наука, 1976. С. 259–276.
Непомилуева Н.И., Дурягина Д.А. К истории лиственничников Среднего Тимана в голоцене (Коми АССР) // Ботан. журн. 1990. Т. 75. С. 326–335.
Никифорова Л.Д. Изменение природной среды в голоцене на северо-востоке европейской части СССР. Дисс. … канд. географ. наук. М., 1979. 154 с.
Панова Н.К., Янковска В., Корона О.М., Зиновьев Е.В. Динамика растительности и экологических условий на Полярном Урале в голоцене // Экология. 2003. № 4. С. 248–260.
Пастухов А.В., Марченко-Вагапова Т.И., Каверин Д.А., Кулижский С.П., Кузнецов О.Л., Панов В.С. Динамика развития бугристых торфяников на южной границе Восточно-Европейской криолитозоны // Почвоведение. 2017. № 5. С. 544–557.
Пономарев Д.В. Остатки мелких млекопитающих из грота Соколиный (Приполярный Сыктывкар: Геопринт, 2005. Урал) // Квартер-2005. Материалы IV Всеросс. совещания по изучению четвертичного периода. С. 334–336.
Пономарев Д.В., Кряжева И.В. Мелкие млекопитающие позднего плейстоцена и голоцена из местонахождений на реке Щугер (Приполярный Урал) // Вестник Института геологии Коми НЦ УрО РАН. 2011. № 7. С. 9–13.
Савина С.С., Хотинский Н.А. Зональный метод реконструкции палеоклиматов голоцена // Развитие природы территории СССР в позднем плейстоцене и голоцене. М.: Наука, 1982. С. 231–244.
Смирнов Н.Г., Головачев И.Б., Бачура О.П., Кузнецова И.А., Чепраков М.И. Сложные случаи определения зубов грызунов из отложений позднего плейстоцена и голоцена тундровых районов Северной Евразии // Материалы по истории и современному состоянию фауны севера Западной Сибири. Челябинск: Рифей, 1997. С. 60–90.
Смирнов Н.Г., Андреичева Л.Н., Корона О.М., Зиновьев Е.В., Головачев И.Б., Павлов П.Ю., Хуфтхаммер А.-К. Материалы к характеристике биоты Приуральской Субарктики в голоценовом оптимуме // Биота Приуральской Субарктики в позднем плейстоцене и голоцене. Екатеринбург: Изд-во “Екатеринбург”, 1999. С. 23–60.
Сурова Т.Г., Троицкий Л.С., Пуннинг Я.М. Палеогеография и абсолютная хронология голоцена Полярного Урала // Изв. АН ЭССР. Химия–геология. 1975. Т. 24. № 2. С. 152–159.
Фадеева Т.В., Смирнов Н.Г. Мелкие млекопитающие Пермского Предуралья в позднем плейстоцене и голоцене. Екатеринбург: Изд-во “Гошицкий”, 2008.
Федорова Р.В. Материалы палеоботанического исследования погребенных торфяников Приполярного Урала // Известия Всесоюзн. географического общества. 1951. Т. 83. Вып. 6. С. 635–640.
Хотинский Н.А. Радиоуглеродная хронология и корреляция природных и антропогенных рубежей голоцена // Новые данные по геохронологии четвертичного периода. М.: Наука, 1987. С. 39–45.
Jankovska V., Andreev A., Panova N. Holocene environmental history on the eastern slope of the Polar Ural Mountains, Russia // Boreas. 2006. V. 35. P. 650–661.
Kultti S., Väliranta M., Sarmaja-Korjonen K., Solovieva N., Virtanen T., Kauppila T., Eronen M. Palaeoecological evidence of changes in vegetation and climate during the Holocene in the pre-Polar Urals, northeast European Russia // J. Quaternary Sci. 2003. V. 18. P. 503–520.
Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Bronk Ramsey C., Buck C.E., Cheng H., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P., Haflidason H., Hajdas I., Hatté C., Heaton T.J., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., Manning S.W., Niu M., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Turney C.S.M., van der Plicht J. IntCal13 and Marine13 radiocarbon age calibration curves 0–50.000 years cal BP // Radiocarbon. 2013. V. 55. № 3. P. 1869–1887.
Zaretskaya N.E., Panin A.V., Golubeva Yu.V., Chernov A.V. Sedimentation settings and the Late Pleistocene–Holocene geochronology in the Vychegda River Valley // Doklady Earth Sci. 2014. V. 455. Iss. 1. P. 223–228.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция