

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 2, стр. 33-54
Зоопроблематики Mobergella radiolata как вид-индекс ярусных подразделений нижнего кембрия
П. Ю. Пархаев 1, *, Ю. Е. Демиденко 1, М. А. Кульша 1
1 Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: pparkh@paleo.ru
Поступила в редакцию 29.12.2018
После доработки 28.02.2019
Принята к публикации 28.03.2019
Аннотация
Обсуждается проблема биостратиграфического обоснования нижних границ атдабанского яруса Общей стратиграфической шкалы России и яруса 3 Международной хроностратиграфической шкалы. Принято считать, что основной характеристикой этих ярусных подразделений является первое появление трилобитов. Продемонстрирована сложность в выборе корреляционного уровня для установления нижней границы яруса 3 Международной хроностратиграфической шкалы по трилобитам, обусловленная различным таксономическим составом комплексов древнейших представителей этой группы членистоногих на разных палеоконтинентах и очевидным диахронизмом уровней их первого появления. Для этих целей предлагается использовать вид зоопроблематик Mobergella radiolata Bengtson, 1968, географическое распространение которого шире, чем у любого вида древнейших трилобитов. Показано, что на территории Сибирской платформы в стратотипическом для нижнекембрийских ярусов регионе (междуречье Лена–Алдан) M. radiolata появляется в разрезах на том же уровне, что и первые атдабанские археоциаты зоны Retecoscinus zegebarti. Это позволяет использовать M. radiolata как вид-индекс нижней границы атдабанского яруса. Находки M. radiolata в других регионах Сибирской платформы (запад, север и юго-восток, внутренние районы платформы) делают этот вид микрофауны крайне ценным инструментом для биостратиграфии нижнекембрийских толщ, корреляционный потенциал которого значительно выше, чем у локально распространенных раннеатдабанских археоциат и трилобитов.
ВВЕДЕНИЕ
Кембрийская часть Международной хроностратиграфической шкалы (МХШ) за последние 20–30 лет претерпела значительные изменения. Это касается как абсолютных датировок границ подразделений (например, в 1980–1990-е гг. начало кембрия определялось в интервале 570–590 млн, сейчас – 541 млн), так и внутреннего наполнения шкалы и ее номенклатуры (Zhu et al., 2018, рис. 1). От традиционного трехчленного деления системы на нижний, средний и верхний отделы мы пришли к четырехчленному с собственными топонимическими названиями отделов (Geyer, Shergold, 2000; Babcock et al., 2005; Babcock, Peng, 2007). Ярусные подразделения, основанные на региональных шкалах (Cowie, Basset, 1989), но использовавшиеся многие годы и на других палеоконтинентах, были упразднены, взамен них предложены формальные ярусы с номерами от 1 до 10 (Geyer, Shergold, 2000; Babcock et al., 2005; Babcock, Peng, 2007). В качестве основных причин в пользу разработки новой шкалы кембрия указывалась невозможность прослеживания границ большинства региональных отделов и ярусов в глобальном масштабе (Palmer, 1998; Geyer, Shergold, 2000; Shergold, Geyer, 2003) и существенное различие в продолжительности традиционных отделов (Landing et al., 1998; Geyer, Shergold, 2000). Новая конструкция кембрийской части МХШ была поддержана не всеми специалистами; в частности, отечественные стратиграфы высказали аргументированные возражения против деления системы на четыре отдела и упразднения нижнекембрийских ярусов МХШ (Розанов и др., 2008; Варламов и др., 2013).
Рис. 1.
Корреляция стратиграфических подразделений средней части нижнего кембрия в ключевых разрезах мира, распространение комплексов древнейших трилобитов (серая заливка) и распространение зоопроблематик Mobergella radiolata Bengtson, 1968 (черная звезда) и Mobergella turgida Bengtson, 1968 (серая звезда). Сокращения: МХШ – Международная хроностратиграфическая шкала, ОСШ – Общая стратиграфическая шкала России.
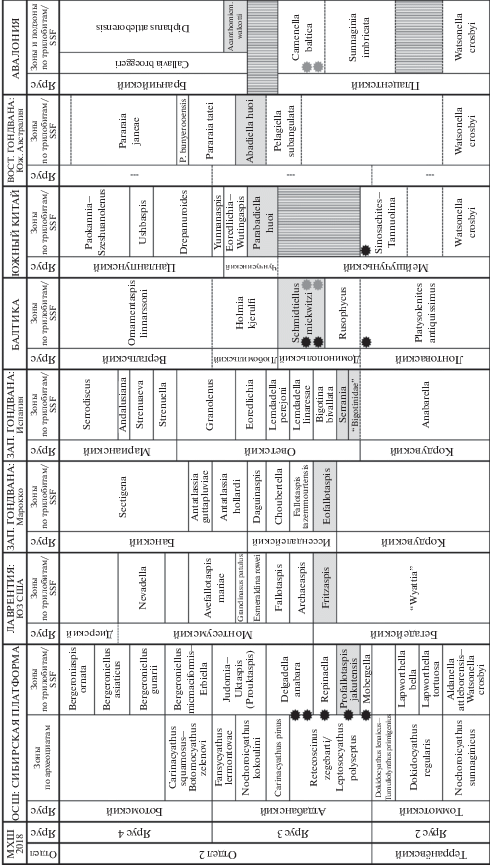
За последние годы в средне-верхнекембрийской части МХШ прогресс по установлению GSSP и соответствующих ярусов вполне очевиден (здесь установлены ярусы 5–9: вулиунский, драмский, гужанский, пайбийский и цзяншаньский), но в традиционном нижнем кембрии (терранёвский отдел и отдел 2) до сих пор присутствует только единственный ярус – форчунский (Landing et al., 2007), непосредственно связанный с нижней границей кембрийской системы МХШ, ратифицированной еще в 1992 г. (Brasier et al., 1994; Landing, 1994). Здесь мы не будем обсуждать проблему нижней границы системы, объективные дефекты выбранного стратотипа, его слабый корреляционный потенциал и возникающие в этой связи стратиграфические проблемы – всем этим вопросам посвящен ряд публикаций (например, Розанов и др., 1997, 2008; Babcock et al., 2014; Zhu et al., 2018). Отметим, что в связи с многочисленными обращениями специалистов, неудовлетворенных утвержденным стратотипом нижней границы кембрия, Международная комиссия по стратиграфии поручила кембрийской подкомиссии рассмотреть вопрос о возможном пересмотре разреза-стратотипа, вида-индекса либо и того, и другого вместе (Babcock et al., 2014). Для работы по данной проблеме создана Международная рабочая группа по нижней границе кембрийской системы, которая должна подготовить соответствующие решения.
По еще неустановленным ярусам МХШ также созданы рабочие группы подкомиссии, успех деятельности которых различен. Так, если по самому верхнему подразделению системы, ярусу 10, значительная часть работы и дискуссий уже проведена, определены уровни корреляции и возможные разрезы-кандидаты (Terfelt et al., 2008; Lazarenko et al., 2011; Peng et al., 2015; Zhu et al., 2018) и в ближайшее время, после проведения ряда формальных процедур, можно ожидать решения подкомиссии, то с нижними ярусами 2, 3 и 4 ситуация складывается иначе. По ярусу 2, который соответствует томмотскому ярусу Общей стратиграфической шкалы России (ОСШ) (Rozanov et al., 2008, fig. 5), мнение большинства специалистов сходится на возможности использования уровня первого появления (FAD) одного из видов моллюсков – Aldanella attleboensis или Watsonella crosbyi для обоснования корреляционного уровня и проведения по нему нижней границы яруса (Li et al., 2011; Parkhaev et al., 2011, 2012; Parkhaev, 2014; Peng et al., 2012). Но пока до детального обсуждения возможных разрезов-кандидатов на GSSP дело еще не дошло. По ярусу 4 (~ботомский ярус ОСШ) высказываются предложения использовать FAD различных трилобитов, например э-одисцид Triangulaspis annio и Hebediscus attleborensis (Geyer, Palmer, 1995; Geyer, 1998; Korovnikov, 2012), Serrodiscus и Calodiscus (Zhuravlev, 1995), ориктоцефалид Arthricocephalus chauveaui и Oryctocarella duyunensis или видов родов Redlichia (s.l.), Olenellus (s.l.), Judomia и Bergeroniellus (Peng et al., 2012; Zhu et al., 2018). При этом уровни появления этих таксонов отнюдь не близки, и, как следствие, потенциальная подошва яруса 4 “плавает” от середины атдабана (Judomia) до второй половины тойона (Arthricocephalus chauveaui) в терминах ОСШ.
Наиболее сложным для обоснования оказался ярус 3 (Zhang et al., 2017; Zhu et al., 2018), который связывают с важнейшим эволюционным событием кембрийского периода – появлением в геологической летописи трилобитов и типа членистоногих в целом.
СТРАТИГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ДРЕВНЕЙШИХ ТРИЛОБИТОВ
Одной из главных характеристик яруса 3 считается появление трилобитов (Peng et al., 2012), поэтому для обоснования нижней границы яруса специалисты пытаются выбрать FAD (first appearing datum, уровень первого появления) одного из древнейших видов трилобитов (Zhang et al., 2017). Основная проблема – очевидная стратиграфическая диахронность уровней появления первых трилобитов и их различный систематический состав (рис. 1).
Предполагается, что самыми древними (Palmer, Repina, 1997) и наиболее примитивными (Jell, 2003) трилобитами являются археаспидиды рода Profallotaspis – P. jakutensis Repina, 1965, P. privica Repina, 1965 и P. tyusserica Bushuev et Goryaeva, 2014, появляющиеся с низов атдабанского яруса на территории Сибирской платформы (Хоментовский, Репина, 1965; Ярусное…, 1984; Репина и др., 1988; Astashkin et al., 1991; Репина, Розанов, 1992; Bushuev et al., 2014). Эти виды отмечены в четырех регионах платформы: в стратотипическом районе атдабанского яруса в среднем течении р. Лена (P. jakutensis и P. privica), на юго-востоке платформы в Учуро-Майском районе (P. jakutensis и P. privica), на севере платформы в нижнем течении р. Лена, хр. Хараулах (P. tyusserica) и в Западном Прианабарье (P. privica), где они характеризуют зону Profallotaspis jakutensis (далее – P. jakutensis). Выше зоны P. jakutensis на Сибирской платформе установлены зоны Repinaella и Delgadella anabara (далее – D. anabara) (Ярусное…, 1984; Astashkin et al., 1991; Варламов и др., 2008). Виды и роды трилобитов, входящие в комплексы этих зон, важны для межрегиональной корреляции.
Возможно, профаллотасписы также присутствуют и в Лаврентии, где описан Profallotaspis? sp. из зоны Fritzaspis формации Кампито, округ Эсмеральда, шт. Невада, США (Hollingsworth, 2005, 2007), однако плохая сохранность материала не позволяет уверенно идентифицировать находки. Совместно найденные археаспидиды Fritzaspis generalis Hollingsworth, 2007, F. ovalis Hollingsworth, 2007, Amplifallotaspis keni Hollingsworth, 2007 и Repinaella sp. позволяют уверенно сопоставлять зону Fritzaspis Лаврентии с зоной Repinaella Сибирской платформы (Hollingsworth, 2007). Ниже этого уровня трилобиты в Лаврентии не обнаружены. В вышележащей зоне Archaeaspis отмечены формы, крайне близкие к типовому виду рода Archaeaspis – A. hupei Repina, 1965, известному из комплекса сибирской зоны Delgadella anabara, поэтому корреляция зоны Archaeaspis Лаврентии и зоны Delgadella anabara Сибирской платформы (Hollingsworth, 2005, 2007) представляется обоснованной.
Другим регионом, возможно, с древнейшими трилобитами является Западная Гондвана (Марокко и Иберийский массив). В Марокко в слоях с первыми трилобитами (пачка Тиут формации Игудин) установлены зоны (снизу вверх): Eofallotaspis, Fallotaspis tazemmourtensis, Choubertella и Daguinaspis, составляющие вместе иссендалейский ярус (Geyer, Landing, 1995, 2006). В зоне E-ofallotaspis появляются трилобиты – биготиниды Hupetina antiqua Sdzuy, 1978, Bigotina bivallata Cobbold, 1935 и фаллотаспидиды Eofallotaspis tioutensis Sdzuy, 1978, E. prima Sdzuy, 1978. Выше появляются редлихииды Lemdadella tioutensis Sdzuy in Liñán et Sdzuy, 1978 из комплекса зоны Fallotaspis tazemmourtensis. Различный таксономический состав древнейших трилобитов Марокко и Сибирской платформы не позволяет проводить прямую корреляцию отложений этих регионов. Однако находки археоциат из той же пачки Тиут, откуда описаны древнейшие марокканские трилобиты (Sdzuy, 1978), позволяют датировать эти толщи средним (Rozanov, Debrenne, 1974; Debrenne, Debrenne, 1978) или даже ранним атдабаном (Debrenne et al., 2015; Yang et al., 2016). Поэтому, вероятно, зону Eofallotaspis следует коррелировать с зоной Repinaella. В пользу такого варианта могут свидетельствовать также находки трилобитов рода Bigotina – B. malykanica Suvorova, 1960 в отложениях зоны Repinaella Сибирской платформы (Ярусное…, 1984; Репина и др., 1988) и B. bivallata в зоне Eofallotaspis Марокко (Pillola, 1993).
На Иберийском полуострове древнейшие трилобиты появляются в нижней половине оветского яруса хребта Сьерра-Морена, пров. Кордова, Южная Испания (Liñán et al., 2008, 2015). Здесь выделяют трилобитовые зоны (снизу вверх): “Bigotinidae”, Serrania, Bigotina bivallata, Lemdadella linaresae, Lemdadella perejoni и Eoredlichia. В зоне “Bigotinidae” отмечены неопределимые обломки биготинидных трилобитов (Zhang et al., 2017), из вышележащей зоны Serrania описаны (Liñán et al., 2008) биготиниды Serrania palaciosi Liñán et al., 2008 и Serrania? gordaensis Liñán et al., 2008. Выше отмечены Bigotina bivallata из отложений одноименной зоны, которая может быть сопоставлена с зоной Eofallotaspis Марокко (Liñán et al., 2015) и, следовательно, с зоной Repinaella Сибирской платформы. В таком случае возможно коррелировать зоны “Bigotinidae” и Serrania с зоной Profallotaspis jakutensis. Значит, зоны Lemdadella linaresae и Lemdadella perejoni могут считаться возрастными аналогами зоны Delgadella anabara, а вышележащая зона Eoredlichia – условно аналог сибирской зоны Judomia–Uktaspis верхнего атдабана. Корреляция низов оветского яруса Иберийского массива с низами атдабанского яруса Сибирской платформы подтверждается также данными по биостратиграфии археоциат (Perejon, Moreno-Eiris, 2006). Недавно из Песчаников Тамамес в провинции Саламанка Центральной Испании описан аграулидный трилобит Lunagraulos tamamensis Liñán, Gamez Vintaned et Gozalo, 2015, стратиграфическое положение вмещающих отложений оценивается как основание оветского яруса (Liñán et al., 2015), что делает этот вид, возможно, древнейшим трилобитом Западной Гондваны.
Древнейшие трилобиты Балтики представлены эндемичными хольмиидами из зоны Schmidtiellus mickwitzi (Ahlberg et al., 1986; Ahlberg, 1991; Lieberman, 1999): Baltobergstroemia mobergi (Bergström, 1973), Schmidtiellus mickwitzi Schmidt, 1888 и Elliptocephala lundgreni (Moberg, 1892) из верхов формации Норреторп Южной Швеции и S. reetae Bergström, 1973 из люкатиских слоев, зоны Volborthella Эстонии. В связи с эндемичностью трилобитов прямая корреляция зоны Schmidtiellus mickwitzi с Сибирской платформой не представляется возможной, однако с учетом филогенетических построений возраст этих видов оценивается (Palmer, Repina, 1993) как середина атдабана–первая половина времени Delgadella anabara. Сопоставление зоны Schmidtiellus mickwitzi с подразделениями шкалы по акритархам, а именно с верхами зоны Skiagia ornata (Moczydłowska, 1991, 2011; Nielsen, Schovsbo, 2011), на наш взгляд, не обеспечивает необходимой точности при корреляции в связи с большой длительностью акритарховых зон (~ярус). Однако находки проблематик Volborthella tenuis Schmidt, 1888 на территории Балтики и Лаврентии позволяют связать стратиграфические последовательности этих двух палеоконтинентов. На территории Эстонии V. tenuis известна из люкатиских слоев, где установлена одноименная зона Volborthella. Этот интервал уверенно коррелируется с зоной Schmidtiellus mickwitzi по трилобитам и акритархам (Moczydłowska, 1991, 2011; Yochelson, Kisilev, 2003). В США (шт. Калифорния, горы Уайт-Иньо) фольбортеллы распространены начиная с верхов пачки Эндрюс-Маунтин (верхи формации Кампито, верхи зоны Fritzaspis; Hollingsworth, 2007, рис. 2) и выше в формации Полета и низах формации Харклесс (Hagadorn, Waggoner, 2002). Если допустить, что первое появление Volborthella в Балтике и Лаврентии одновременно, то зона Schmidtiellus mickwitzi может быть сопоставлена с верхами зоны Fritzaspis и зоной Archaeaspis, которые (см. выше) соответствуют зонам Repinaella и Delgadella anabara Сибирской платформы.
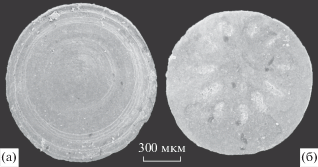
Рис. 2.
Склериты зоопроблематик Mobergella radiolata Bengtson, 1968. а – экз. ПИН, № 5279/5059, склерит снаружи; б – экз. ПИН, № 5279/5056, склерит изнутри; нижний кембрий, атдабанский ярус, зона Retecoscinus zegebarti, Сибирская платформа, среднее течение р. Лена, разрез Журинский мыс, слой 4.
На Южно-Китайской платформе самые ранние трилобиты происходят из отложений зоны Parabadiella huoi в основании яруса чунчуси, выше выделяется зона Eoredlichia–Wutingaspis (Zhang, 1987; Steiner et al., 2001). Корреляция зоны Parabadiella huoi с подразделениями ОСШ России различна, ее сопоставляли (обзор см. Yuan et al., 2011) с зоной Profallotaspis jakutensis (Zhang, 2003; Yuan et al., 2011), с зоной Repinaella (Peng et al., 2006), с зоной Delgadella anabara (Репина, 1986; Shergold, 1997), с зоной Judomia–Uktaspis (Zhuravlev, 1995; Paterson, Brock, 2007) и даже с основанием ботомского яруса (Landing, 1994). Корреляция зон Parabadiella huoi и Profallotaspis jakutensis основана на том, что в вышележащей южнокитайской зоне Eoredlichia–Wutingaspis присутствуют трилобиты рода Yunnanocephalus, который, в свою очередь, встречен в известняке Шэклтон Антарктиды (Palmer, Rowell, 1995) совместно с трилобитом Lemdadella antarctica Palmer et Rowell, 1995. По мнению сторонников рассматриваемой корреляции (Yuan et al., 2011), род Lemdadella – один из древнейших трилобитов и встречается в Марокко (L. spectabilis Sdzuy, 1978 и L. tioutensis Sdzuy in Liñán et Sdzuy, 1978) в зоне Fallotaspis tazemmourtensis, следовательно, южнокитайскую зону Eoredlichia–Wutingaspis следует коррелировать с зоной Fallotaspis tazemmourtensis Западной Гондваны, тогда нижележащая зона Parabadiella huoi – эквивалент зоны Eofallotaspis, а она, в свою очередь, сопоставляется с зоной Profallotaspis jakutensis.
Во-первых, как показано выше, зона Eofallotaspis Марокко коррелируется с зоной Repinaella ОСШ. Во-вторых, вид Lemdadella antarctica известен только из Антарктиды, а сам род Lemdadella распространен существенно шире, чем указывают авторы. Так, L. linaresae Liñán Guijarro, 1978 и L. perejoni Liñán, 2005 – виды-индексы одноименных зон оветского яруса Иберии, которые коррелируются с зоной Delgadella anabara Сибирской платформы, тогда как испанская зона Eoredlichia – вероятный аналог зоны Judomia–Uktaspis. Также отметим, что Lemdadella sp. встречается в отложениях зоны Sectigena формации Иссафен и зоны Antatlassia guttapluviae формации Лемдад Высокого Атласа (Geyer, Landing, 2006), а это очень высокие уровни, сопоставимые с ботомско-тойонским интервалом Сибири (Sundberg et al., 2016; Zhang et al., 2017).
Очевидно, что прямая корреляция по трилобитам нижнекембрийских толщ Южно-Китайской и Сибирской платформ невозможна. Для решения проблемы необходимо обратиться к данным по другим группам фауны – археоциатам и SSF. Недавняя ревизия археоциат Южного Китая (Yang et al., 2016) показывает, что в нижнем кембрии региона можно выделить пять зональных подразделений (снизу вверх): зона Dailycyathus xiuqiensis (середина яруса чунчуси), зона Dictyocyathus shaanxiensis (верхи яруса чунчуси), зона Spirillicyathus duplex (низы яруса цанланпу), зона Sibirecyathus meitanensis (середина яруса цанланпу), слои с Archaeocyathus yanjiaoensis (верхи яруса цанланпу). Зона Dailycyathus xiuqiensis (~середина зоны Eoredlichia–Wutingaspis) включает роды археоциат, характерные для середины или конца атдабана, при этом типично ботомские формы здесь отсутствуют. Зона Dictyocyathus shaanxiensis (=верхи зоны Eoredlichia–Wutingaspis) содержит роды, встречающиеся в средне-позднеатдабанских отложениях Западной Гондваны. В зоне Spirillicyathus duplex (=зона Yunnanaspis–Yiliangella) отмечены несколько родов, появляющихся на уровне атдабан-ботомской границы в различных регионах мира. Зона Sibirecyathus meitanensis (=зона Drepanuroides) содержит роды, характерные для низов ботомского яруса Монголии, Алтае-Саянской складчатой области и Колымского блока Сибирской платформы (Yang et al., 2016).
Таким образом, трилобитовая зона Eoredlichia–Wutingaspis соответствует среднему–верхнему атдабану, зона Yunnanaspis–Yiliangella, по-видимому, терминальному атдабану, а зона Drepanuroides – нижнеботомскому подъярусу. В связи с этим возраст самой нижней трилобитовой зоны Parabadiella huoi Южно-Китайской платформы следует оценивать не моложе середины атдабана.
Дополнительную привязку к зональной шкале атадабанского яруса Сибирской платформы могут дать данные по распространению моллюсков. В нижней части яруса чунчуси установлена зона Pelagiella subangulata, охватывающая почти целиком (без базальной части) формацию Юаньшань на востоке Юньнани и верхнюю пачку формации Цзюлаодун в центральной части Сычуани (Steiner et al., 2007). Это интервал-зона, соответствующая стратиграфическому распространению вида Pelagiella emeishanensis He in Xing et al., 1984, позднее (Gravestock et al., 2001) синонимизированного с P. subangulata (Tate, 1892). Низам этой зоны соответствует зона Parabadiella huoi.
Виды рода Pelagiella распространены на Сибирской платформе начиная с верхов зоны Delgadella anabara второй половины атдабанского яруса и выше до середины амгинского яруса (Ярусное…, 1984; Розанов и др., 2010; Kouchinsky et al., 2011; Коровников, Новожилова, 2012). Самые ранние пелагиеллы Сибири обычно выделяются в вид P. adunca Missarzhevsky, 1966 (иногда их ошибочно определяют как P. lorenzi Kobayashi, 1939), который по морфологии весьма близок с P. subangulata. Из низов ботомского яруса Восточного Прианабарья (верхи эмяксинской свиты, зона Calodiscus–Erbiella) недавно описана Pelagiella sp. (Kouchinsky et al., 2015), крайне схожая с типичной P. subangulata из нижнего кембрия Австралии. Несомненно, род Pelagiella нуждается в ревизии. Сейчас можно уверенно утверждать, что самые древние пелагиеллы – среднеатдабанские. На этом основании зону Parabadiella huoi Китая можно коррелировать с верхами зоны Delgadella anabara ОСШ, что не противоречит вышеприведенным данным по археоциатам.
В Восточной Гондване наиболее полная последовательность нижнекембрийских отложений известна на территории штата Южной Австралии. Здесь установлены аналоги всех ярусных подразделений нижнего кембрия ОСШ России (Daily, 1972; Дэйли, 1976; Zhuravlev, Gravestock, 1994; Gravestock et al., 2001). Древнейшие трилобиты происходят из терригенно-карбонатных отложений бассейнов Стэнсбери и Эрроуи, где установлены четыре последовательные трилобитовые зоны (снизу вверх): Abadiella huoi, Pararaia tatei, Pararaia bunyerooensis и Pararaia janeae (Bengtson et al., 1990, fig. 8 ; Jago et al., 2006). По таксономическому составу комплексы этих зон близки к комплексам из нижнекембрийских толщ Южного Китая – 10 родов и 6 видов общие (Paterson, Brock, 2007), поэтому наименьшие затруднения возникают при корреляции зональных шкал этих регионов.
По наличию трилобитов Eoredlichia sp. и Abadiella huoi Chang, 1966 зона Abadiella huoi Австралии может быть сопоставлена с верхами зоны Parabadiella huoi–низами зоны Eoredlichia–Wutingaspis Китая. В пользу такой корреляции говорят и находки моллюсков P. subangulata в более древних отложениях, чем трилобиты зоны Abadiella huoi, – в средней части формации Кулпара бассейна Стэнсбери и низах Известняка Виррапоуи бассейна Эрроуи (Пархаев, 2019а, рис. 3, 2019 б, рис. 3). Таким образом, сама зона Abadiella huoi, а точнее, ее нижняя половина соответствует лишь верхам зоны Parabadiella huoi (Gravestock et al., 2001; Jago et al., 2006; Kruze et al., 2017).
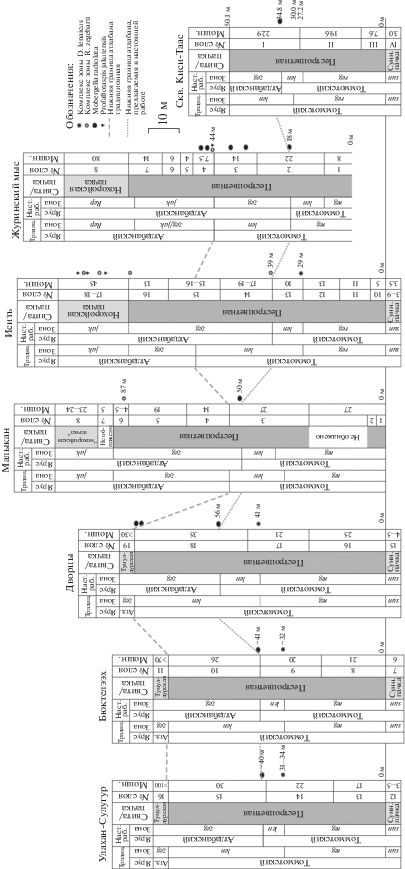
Рис. 3.
Стратиграфическое распространение зоопроблематик Mobergella radiolata, археоциат зон D. lenaicus и R. zegebarti, трилобитов зоны P. jakutensis в стратотипических разрезах томмотского и атдабанского ярусов в среднем течении рек Алдан (Улахан-Сулугур, Бюктелээх, Дворцы) и Лена (Малыкан, Исить, Журинский мыс, скв. Киси-Таас) и положение нижней границы атдабанского яруса: традиционнное (традиц.) и предлагаемое в настоящей работе (наст. раб.). Обозначения зон: sun – Nochoroicyathus sunnaginicus, reg – Dokidocyathus regularis, len – Dokidocyathus lenaicus–Tumuliolynthus primigenius, zeg – Retecoscinus zegebarti/Leptosocyathus polyseptus, jak – Profallotaspis jakutensis, Rep – Repinaella. Сокращения: атд. – атдабанский, сунн. пачка – суннагинская пачка.
Зона Pararaia tatei сопоставляется со второй половиной зоны Eoredlichia–Wutingaspis по наличию вида Eoredlichia shensiensis (Chang, 1966) в обоих регионах. В связи с присутствием в составе комплекса зоны Pararaia tatei таких родов трилобитов, как Prouktaspis (низы зоны) и Egyngolia (середина зоны) (Bengtson et al., 1990, рис. 8 ), возраст подразделения может быть оценен как терминальный атдабан–основание ботомского яруса, так как род Prouktaspis характерен для позднего атдабана Сибирской платформы, а Egyngolia – для ботомских толщ Монголии (Astashkin et al., 1991, 1995; Zhuravlev, 1995; Сухов и др., 2016). В связи с этим, помимо верхов зоны Eoredlichia–Wutingaspis китайской зональной шкалы, эквивалентом зоны Pararaia tatei следует считать зону Yunnanaspis и нижнюю половину зоны Drepanuroides (рис. 1).
Трилобиты зоны Pararaia bunyerooensis представлены шестью видами: P. bunyerooensis Jell, 1990, Yorkella australis (Woodward, 1884), Yunnanocephalus macromelos Paterson et Brock, 2007, Wutingaspis euryoptilos Paterson et Brock, 2007, Eoredlichia sp., Redlichia sp. (Paterson, Brock, 2007). Наличие представителей родов Yunnanocephalus, Wutingaspis и Eoredlichia предполагает сопоставление этой зоны с зоной Eoredlichia–Wutingaspis Китая (средний–верхний атдабан), но находки Redlichia свидетельствуют о ботомском возрасте (Сухов и др., 2016) или даже тойонском (Astashkin et al., 1991; Sundberg et al., 2016) зоны Pararaia bunyerooensis. Встречающиеся совместно моллюски зоны Stenotheca drepanoida указывают на ботомский возраст отложений (Gravestock et al., 2001; Topper et al., 2009; Пархаев, 2019а, 2019б).
Комплекс зоны Pararaia janeae включает около 20 видов трилобитов, среди которых отмечены представители рода Serrodiscus и вид Redlichia endoi Lu, 1950 (Bengtson et al., 1990; Paterson, Brock, 2007). Род Serrodiscus характерен для зоны Bergeroniellus micmaciformis–Erbiella основания ботомского яруса Сибирской платформы (Yang et al., 2016), R. endoi – для зоны Palaeolenus Китая (Paterson, Brock, 2007), которая сопоставляется с верхами ботомского яруса Сибири, с зоной Bergeroniaspis ornata (Zhuravlev, 1995). Таким образом, Pararaia janeae отвечает значительной части ботомского яруса, вероятно, без самых его низов и верхов. Нижней части зоны соответствуют слои с археоциатами Syringocnema favus, возраст которых оценивается от средне- до позднеботомского (Zhuravlev, Gravestock, 1994).
На территории Авалонии (ЮВ Ньюфаундленд) древнейшие трилобиты представлены видами Callavia broeggeri (Walcott, 1890), Acanthomicmacca walcotti (Matthew, 1899), Acanthomicmacca sp., Comluella sp. из пачки Броад-Коув верхов формации Смит-Пойнт (Fletcher, 2006). Эти виды образуют комплекс подзоны Acanthomicmacca walcotti зоны Callavia broeggeri. Стратиграфически выше в пачке Редлэнд-Коув низов формации Бригус разнообразие трилобитов существенно выше; здесь установлена подзона Dipharus attleborensis, в которой присутствуют (Fletcher, 2006): Dipharus attleborensis (Shaler et Foerste, 1888) [=Hebediscus attleborensis], Delgadella plana (Hutchinson, 1962), Calodiscus lobatus (Hall, 1847), C. meeki (Ford, 1876), C. schucherti (Matthew, 1896), Weymouthia nobilis (Ford, 1872), Acimetopus helena (Walcott, 1889), Mallagnostus llarenai (R. Richter et E. Richter, 1941), Serrodiscus bellimarginatus (Shaler et Foerste, 1888), Callavia broeggeri, ?Kjerulfia sp., Triangulaspis meglitzkii (Toll, 1899), T. vigilans (Matthew, 1889), Strenuella strenua (Billings, 1874), Strenuaeva cf. spinosa Ahlberg et Bergström, 1978, Comluella pustulata (Cobbold, 1910), C. protolenoides (Cobbold, 1931), Acanthomicmacca walcotti, Avalonia manuelensis Walcott, 1891, “Bonnia” bombifrons (Matthew, 1886) и Pseudatops reticulatus (Walcott, 1890).
Нижнюю подзону зоны Callavia broeggeri условно коррелируют с низами зоны Antatlasia hollardi Марокко (Fletcher, 2006), что соответствует верхам атдабана ОСШ России. Верхняя подзона по наличию таких родов, как Serrodiscus и Strenuella, имеет уже несомненно ботомский возраст и сопоставляется с зонами Antatlassia guttapluviae и Sectigena Марокко и зонами Granolenus, Strenuella, Strenuaeva, Andalusiana и Serrodiscus Иберийского массива.
Таким образом, первые трилобиты на различных палеоконтинентах появляются неодновременно (рис. 1), причем везде – это разные виды, относящиеся как минимум к четырем семействам (Zhang et al., 2017). Поэтому выбрать уровень появления единственного вида и использовать его для межрегиональной корреляции, как это требуют правила Международной стратиграфической комиссии (Remane et al., 1996), не представляется возможным. Трилобиты более высоких уровней также бесперспективны вплоть до появления в геологической летописи в начале ботомского века, вероятно, пелагических эодисцидых форм (например, Serrodiscus, Calodiscus) с широким географическим распространением.
Очевидно, следует обратиться к другим группам организмов, распространение которых во временных интервалах появления трилобитов не ограничивается одним континентом. Нам представляется, что для данной цели наиболее удачным будет выбор зоопроблематик Mobergella radiolata Bengtson, 1968 (рис. 2). Перспективность мобергелл для стратиграфических построений уже отмечалась ранее (например, Репина, Розанов, 1992; Brasier et al., 1992; Rozanov, Zhuravlev, 1992; Rozanov et al., 2011; Demidenko et al., 2012; Варламов и др., 2013).
Ниже рассмотрим возможность использования вида Mobergella radiolata для определения нижней границы атдабанского яруса ОСШ и нижней границы яруса 3 МХШ.
АТДАБАНСКИЙ ЯРУС ОБЩЕЙ СТРАТИГРАФИЧЕСКОЙ ШКАЛЫ РОССИИ
Атдабанский ярус предложен в 1969 г. как подразделение ярусного ранга, заполняющее интервал между ранее установленными томмотским (снизу) и ботомским (сверху) ярусами и в значительной степени соответствующее по объему атдабанскому горизонту. Решением Межведомственного стратиграфического комитета СССР от 24 мая 1982 г. ярус введен в Общую стратиграфическую шкалу (Постановления…, 1983; Решения…, 1983). Первоначально ярус был определен как последовательность из археоциатовых зон и слоев с археоциатами (снизу вверх): зона Leptosocyathus polyseptus–Retecoscinus zegebarti, зона Porocyathus pinus, слои с Nochoroicyathus kokoulini и слои с Fansycyathus lermontovae (Журавлева и др., 1969, табл. 1 ). Позднее слоям с археоциатами был придан ранг зон. Важной характеристикой яруса является первое появление трилобитов; археоциаты проходят вторую ступень эволюционного развития. Трилобиты разнообразны: в нижней части яруса – Profallotaspis, Repinaella, первые Bigotina и другие роды, в средней части – Nevadella, Archaeaspis, Triangullina, Hebediscus, в верхней части – Judomia и Uktaspis, которые в самых верхах яруса встречаются вместе с Bonnia, Kootenia и другими формами. В верхней части яруса встречаются очень характерные и широко распространенные SSF – Pelagiella, Hadimopanella и Microdictyon (Ярусное…, 1984).
В качестве стратотипа атдабанского яруса указана серия обнажений в среднем течении р. Лена между разрезом Ой-Муран на левом берегу и разрезом Аччагый-Кыыры-Таас на правом берегу (Ярусное…, 1984). Гипостратотип яруса и стратотип его нижней границы расположены в разрезе Журинский мыc, где обнажается подошва яруса и его нижняя часть (Путеводитель…, 1973, рис. 11 ; Решения…, 1983; Ярусное…, 1984, рис. 13 ; Guidebook…, 1990, рис. 5; Варламов и др., 2008, рис. 27, 34, 35 ). В этом разрезе граница находится в основании слоя 4, который маркируется появлением древнейших трилобитов и сложно организованных археоциат и соответствует нижней границе зон Profallotaspis jakutensis и Retecoscinus zegebarti11.
Рассмотрим распространение ключевых для томмот-атдабанского переходного интервала таксонов ископаемых организмов в серии разрезов нижнего кембрия стратотипического Лено-Алданского района (рис. 3) и других регионов Сибирской платформы (рис. 4–6).
Рис. 4.
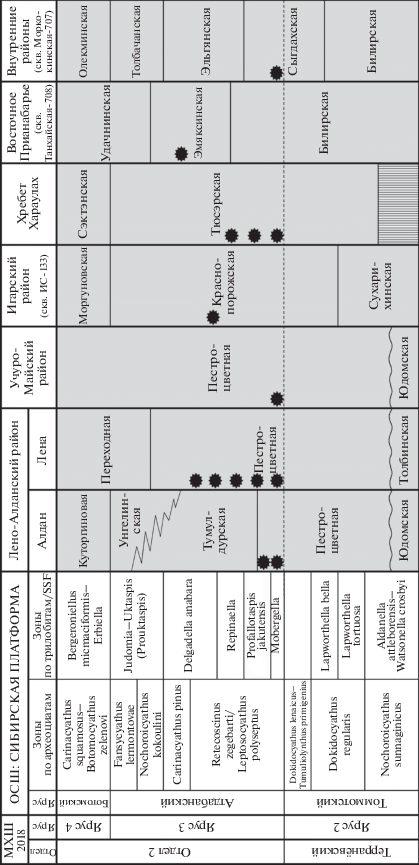
Распространение зоопроблематик Mobergella radiolata Bengtson, 1968 на территории Сибирской платформы (контур платформы показан серой линией). Разрезы: 1–8 – Лено-Алданский район: 1 – Дворцы, 2 – Бюктелээх и Улахан-Сулугур, 3 – Малыкан, 4 – Исить, 5 – Журинский мыс, 6 – скв. Киси-Таас, 7 – Ой-Муран, 8 – Бачык; 9 – скв. Улу-3; 10, 11 – хр. Хараулах: 10 – Чекуровская антиклиналь, Чекуровка, 11 – Булкурская антиклиналь, Улахан-Алдьярхай и Тюсэр; 12 – Учуро-Майский район, С-элиндэ; 13 – Игарский район, скв. ИС-133; 14 – Восточное Прианабарье, скв. Танхайская-708; 15 – внутренние районы, скв. Моркокинская-707.
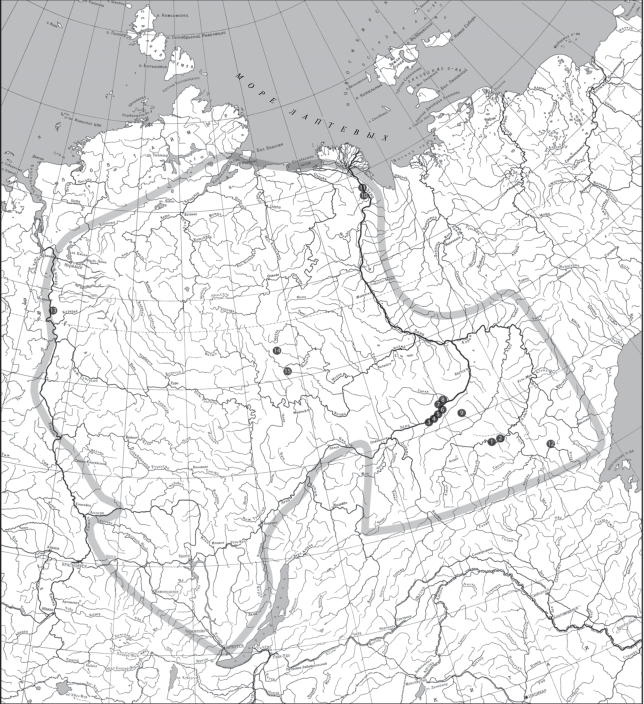
Рис. 5.
Стратиграфическое распространение зоопроблематик Mobergella radiolata Bengtson, 1968, трилобитов и археоциат в разрезах Хараулаха (Репина и др., 1974; Коровников, Новожилова, 2012; Bushuev et al., 2014). Обозначения зон: reg – Dokidocyathus regularis, len – Dokidocyathus lenaicus–Tumuliolynthus primigenius, zeg – Retecoscinus zegebarti/Leptosocyathus polyseptus, jak – Profallotaspis jakutensis, Rep – Repinaella, anab – Delgadella anabara, Jud – Judomia/Prouktaspis, Nev – Nevadella, tum – Majatheca tummefacta. Сокращения: зона арх. – зона по археоциатам, зона трил. – зона по трилобитам.
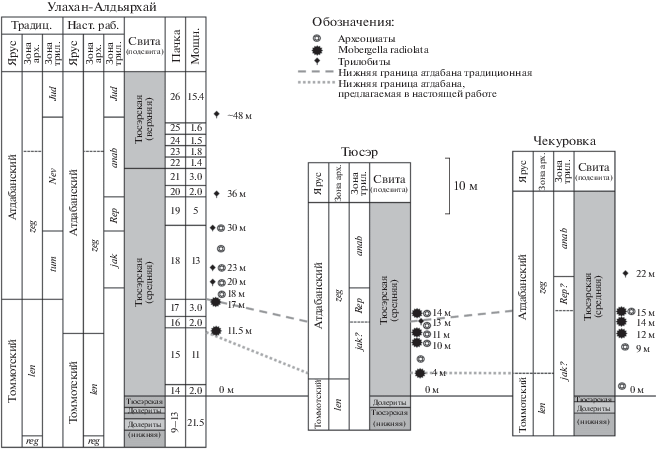
Рис. 6.
Стратиграфическое распространение зоопроблематик Mobergella radiolata (черная звезда) в литолого-стратиграфических подразделениях нижнего кембрия различных структурно-фациальных регионов Сибирской платформы. Пунктиром отмечена нижняя граница атдабанского яруса (свиты по Astashkin et al., 1991; Сухов и др., 2016). Сокращения: МХШ – Международная хроностратиграфическая шкала, ОСШ – Общая стратиграфическая шкала России.
Лено-Алданский район. Разрез Улахан-Сулугур. Разрез расположен на левом берегу р. Алдан в 7 км выше устья руч. Улахан-Сулугур (Путеводитель…, 1973, рис. 3, 7 ; Репина, Розанов, 1992). На доломитах юдомской свиты (слои 1–11) залегают разнообразные по цвету и составу известняки пестроцветной свиты (слои 12–15) и серые доломиты и известняки тумулдурской свиты (слой 16). В пределах пестроцветной свиты установлены зоны по археоциатам: Nochoroicyathus sunnaginicus, верхи (слой 12), Dokidocyathus regularis (слои 13–14) и Dokidocyathus lenaicus–Tumuliolynthus primigenius (слои 14–15)22. Общая мощность пестроцветной свиты (без суннагинской пачки, слой 12) в разрезе – 69 м. Нижняя граница зоны D. lenaicus проводится по одноименному комплексу археоциат в верхних 5–8 метрах слоя 14. Таким образом, мощность нижележащей зоны D. regularis в данном разрезе – 31–34 м, мощность зоны D. lenaicus – 35–38 м. В соседнем обнажении в аналогах нижней части слоя 15 обнаружены зоопроблематики Mobergella radiolata (Розанов и др., 1969, с. 28, как Discinella brastadi в обн. 407; Путеводитель…, 1973).
Разрез Бюктэлээх. Разрез расположен на левом берегу р. Алдан в 12 км выше разреза Улахан-Сулугур (Путеводитель…, 1973, рис. 6). На доломитах юдомской свиты (слои 1–6) залегают разнообразные по цвету и составу известняки пестроцветной свиты (слои 7–10) и серые доломиты и известняки тумулдурской свиты (слой 11). В пределах пестроцветной свиты установлены зоны по археоциатам: N. sunnaginicus (слой 7), D. regularis (слои 8–9) и D. lenaicus (слои 9–10). Общая мощность пестроцветной свиты (без суннагинской пачки, слой 7) в разрезе ~67 м. Нижняя граница зоны D. lenaicus проводится по одноименному комплексу археоциат в ~11 м от подошвы слоя 9. Таким образом, мощность нижележащей зоны D. regularis в данном разрезе ~32 м, мощность зоны D. lenaicus – 39 м. В основании слоя 10 найдены зоопроблематики Mobergella radiolata (Путеводитель…, 1973).
Разрез Дворцы. Разрез расположен на левом берегу р. Алдан в 4 км выше устья руч. Дьялхах. Разрез выбран в качестве стратотипа томмотского яруса, его нижней границы и зоны N. sunnaginicus, в связи с этим он неоднократно описывался в литературе (см. обзор в Репина, Розанов, 1992, рис. 2; Варламов и др., 2008, рис. 7 ). Разрез представлен скальными выходами высотой более 300 м и протяженностью 4.5 км, в которых обнажаются доломиты юдомской свиты (слои 1–14), разнообразные известняки пестроцветной свиты (слои 15–18) и доломиты и известняки тумулдурской (слой 19) свиты. В пределах пестроцветной свиты установлены зоны по археоциатам: N. sunnaginicus, верхи (слой 15), D. regularis (слои 16–17) и D. lenaicus (слои 17–18). Общая мощность пестроцветной свиты (без суннагинской пачки, слой 15) в разрезе – 81 м. Нижняя граница зоны D. lenaicus проводится в 16 м от подошвы слоя 17 по появлению археоциат, характерных для комплекса одноименной зоны. Таким образом, мощность нижележащей зоны D. regularis в данном разрезе 41 м, мощность зоны D. lenaicus – 40 м. В 10 м от подошвы слоя 18 обнаружены зоопроблематики Mobergella radiolata (Ярусное…, 1984).
В вышеупомянутых разрезах среднего течения р. Алдан нижняя граница атдабанского яруса проводится условно по подошве тумулдурской свиты, руководящие формы атдабана (трилобиты зоны P. jakutensis или археоциаты зоны R. zegebarti) в данных разрезах не установлены.
Разрез Исить. Разрез расположен на правом берегу р. Лена напротив пос. Исить. Разрез является гипостратотипом томмотского яруса и стратотипом зон D. regularis и D. lenaicus и неоднократно описан в литературе (см. обзор в Ярусное…, 1984; Варламов и др., 2008, рис. 22 ). В урезе воды выходят серые доломиты толбинской свиты (слой 1), а после скрытого осыпью интервала мощностью 9 м (слой 2) обнажается пестроцветная свита (слои 3–16), выше перекрытая нохоройской пачкой (слои 17–18). Слои 3–9 (вскрыты шурфом) представлены зоной N. sunnaginicus, слои 10–12 – стратотип зоны D. regularis, слой 13 – стратотип зоны D. lenaicus. Нижняя граница атдабанского яруса (и зоны R. zegebarti) ранее проводилась условно в середине слоя 15, исходя из представлений о примерно 40-метровой мощности нижележащей зоны D. lenaicus (Путеводитель…, 1973; Ярусное…, 1984). Достоверные археоциаты зоны R. zegebarti описаны в 20 м выше, в верхнем метре слоя 16 (Иситский биостром), а первые трилобиты Profallotaspis jakutensis обнаружены еще выше – в нохоройской пачке (слой 17). Новые исследования (Скорлотова, 2013) показали наличие представительного зонального комплекса археоциат зоны R. zegebarti (включая и вид-индекс)33 в 0.7 м выше основания слоя 14, что позволяет обоснованно опустить нижнюю границу зоны R. zegebarti на 26 м. В результате мощность зоны D. regularis в разрезе Исить составляет ~29 м, а мощность зоны D. lenaicus сокращена до 10–11 м.
Разрез Журинский мыс. Разрез расположен на правом берегу р. Лена напротив устья руч. Негюрчюне (Путеводитель…, 1973, рис. 11 ; Ярусное…, 1984; Варламов и др., 2008, рис. 27 ). Разрез является стратотипом атдабанского яруса и его нижней границы, а также стратотипом зон R. zegebarti, P. jakutensis, Repinaella и D. anabara. От уреза воды обнажаются глинистые известняки с биогермами (слой 1), в различной степени глинистые известняки (слои 2–6) и доломиты (слой 7) пестроцветной свиты, перекрытые доломитизированными известняками нохоройской пачки (слой 8). В слое 1 установлен комплекс зоны D. regularis, подошва зоны D. lenaicus условно проводится по первому появлению зоопроблематик Mobergella radiolata в 10 м выше основания слоя 2. Нижняя граница атдабанского яруса и основание зон R. zegebarti и Profallotaspis проводятся по первому появлению соответствующих видов-индексов археоциат и трилобитов в основании слоя 4.
Разрез скв. Киси-Таас. Скважина пробурена на правом берегу р. Лена в 200 м ниже устья руч. Киси-Таас (Журавлева и др., 1983; Варламов и др., 2008). Скважиной вскрыты 50.1 м пестроцветной свиты и более 120 м толбинской свиты. В интервале 0–19.5 м вскрыта толща делювия. В пределах пестроцветной свиты выделены четыре пачки: I – интервал 19.5–42.4 м, II – интервал 42.4–62.0 м, III – интервал 62.0–69.6 м, IV – интервал 69.6–72.5 м (суннагинская пачка). В интервале 39.6–69.6 м отмечены археоциаты зоны D. regularis. Нижняя граница зоны D. lenaicus проводится условно на глубине 39.6 м по исчезновению археоциат нижележащей зоны. Таким образом, в скв. Киси-Таас мощность зоны D. regularis – 30 м, мощность зоны D. lenaicus – более 20 м. С глубины 34.8 м определена Mobergella radiolata.
Разрезы Ой-Муран–Бачык. Ниже по течению р. Лена в восточных разрезах стратотипического района M. radiolata встречается на нескольких уровнях в пределах зоны Leptosocyathus polyseptus (аналог зоны R. zegebarti в восточных разрезах). Так, в разрезе Ой-Муран мобергеллы отмечены непосредственно в обнажениях у устья р. Гостинная (Демиденко, 2016) и в скв. Гостинная на гл. 0–4 и 8–15 м в верхней части зоны L. polyseptus и в интервале 52.5–79 м выше уровня с археоциатами зоны D. lenaicus (Ярусное…, 1984, рис. 16 ), что, по-видимому, соответствует уже низам зоны L. polyseptus. В разрезе Бачык M. radiolata обнаружена (Варламов и др., 2008) в основании слоя 2 еще в пределах зоны L. polyseptus (зона Carinacyathus pinus в разрезе начинается выше – с бачыкского водорослево-археоциатового биострома, слой 3), но уже вместе с трилобитами Delgadella anabara (Laz.), Archaeaspis hupei Rep., Nevadella subgroenlandica (Rep.), что позволяет сопоставлять этот уровень с низами зоны Delgadella anabara по трилобитам.
Междуречье Лена–Алдан. Mobergella radiolata отмечена здесь в пестроцветной свите скв. Улу-3 в фаунистических комплексах V и VI пограничного томмот-атдабанского интервала (Васильева, 1998, как Mobergella brastady (Poulsen, 1932) и Brastadella brastady (Poulsen, 1932)). Подробного описания разреза скважины и распространения в нем органических остатков автор не приводит и дает лишь весьма краткую характеристику комплексов. Для комплексов V и VI характерно наличие мобергелл и зоопроблематик рода Rhombocorniculum, комплекс VI отличается от комплекса V наличием спикул губок Heterostella. Здесь мы приводим сведения о находке M. radiolata в данном районе только для дополнения географического распространения вида.
Разрез Малыкан. Нельзя оставить без внимания разрез пестроцветной свиты на правом берегу р. Лена напротив пос. Малыкан (Розанов, Миссаржевский, 1966; Розанов и др., 1969; Путеводитель…, 1973; Мешкова, 1974). Разрез обнажен не полностью, и комплексы ископаемых не настолько полны, как в других разрезах стратотипического района, однако он весьма важен для нашего анализа, так как представляет собой самый западный разрез из серии обнажений переходной фациальной области и в нем отмечены археоциаты зон D. regularis (низы пестроцветной свиты у уреза воды) и R. zegebarti (пестроцветная свита под нохоройской пачкой), а также находки Mobergella radiolata (на уровне ~50 м от уреза воды и ~45 м ниже нохоройской пачки). Очевидно, что находки археоциат комплекса R. zegebarti в разрезе Малыкан по уровню соответствуют иситскому биострому, а мобергеллы обнаружены на уровне основания слоя 14 разреза Исить (45–47 м ниже нохоройской пачки), где отмечены самые нижние находки археоциат комплекса R. zegebarti.
Проанализируем мощности зон D. regularis и D. lenaicus в рассмотренных разрезах. Мощность зоны D. regularis: Улахан-Сулугур ~31 м, Бюктэлээх ~32 м, Дворцы – 41 м, Исить – 29 м, Журинский мыс (неполная мощность) – 18 м, Киси-Таас – 30 м. Мощность зоны D. lenaicus: Улахан-Сулугур ~38 м, Бюктэлээх – 39 м, Дворцы – 40 м, Исить (с учетом новых находок археоциат) – 10 м, Журинский мыс – 26 м, Киси-Таас – 20 м (неполная мощность). Легко заметить, что мощность зоны D. regularis варьирует незначительно, в среднем составляя около 30 м. Мощность же зоны D. lenaicus изменяется четырехкратно – от 9 до 40 м (рис. 3). Данное обстоятельство не согласуется с выдержанными по мощности нижнекембрийскими толщами среднего течения рек Алдан и Лена, конечно, кроме рифовых и прилегающих к ним фаций (Ярусное…, 1984). Легко заметить, что в значительной степени завышенная мощность зоны D. lenaicus приходится на разрезы, в которых нижняя граница вышележащей зоны R. zegebarti проводится условно, в связи с отсутствием достоверных находок ископаемых данного комплекса. Так, во всех разрезах среднего течения р. Алдан подошва зоны R. zegebarti и атдабанского яруса совмещается с подошвой тумулдурской свиты, так как в верхней части пестроцветной свиты находки ископаемых редки.
Важнейшими являются находки зоопроблематик Mobergella radiolata в средней части пестроцветной свиты алданских разрезов Улахан-Сулугур, Бюктэлээх и Дворцы, в 9–14 м выше подошвы зоны D. lenaicus (рис. 3). Примечательно, что этот уровень, так же как и в разрезе Малыкан, совпадает с новыми находками археоциат комплекса R. zegebarti в разрезе Исить в 10 м выше подошвы зоны D. lenaicus. Целенаправленные поиски Mobergella radiolata в нескольких пересечениях разреза Исить в ходе полевых работ 2006, 2007, 2009, 2014, 2016–2018 гг. результатов не дали. Однако находки данного вида в 30 км ниже по течению р. Лена в разрезах Журинский мыс (наши сборы 2007 г.), Негюрчене (Ярусное…, 1984; наши сборы 2009 г.) и скв. Киси-Таас приходятся примерно на этот же уровень (рис. 3).
Нам представляется целесообразным включить вид Mobergella radiolata в состав комплекса зоны R. zegebarti и использовать первое появление данного вида в качестве надежного биостратиграфического инструмента для определения нижней границы зоны и, как следствие, нижней границы атдабанского яруса. Большое значение Mobergella radiolata приобретает для разрезов, в которых отсутствуют находки раннеатдабнских археоциат и трилобитов, распространение которых весьма локально.
Если мы принимаем уровень первого появления Mobergella radiolata за нижнюю границу атдабанского яруса, снимается противоречие с четырехкратным варьированием мощности зоны D. lenaicus, и ее мощность устанавливается в интервале 9–15 м: Улахан-Сулугур ~9 м, Бюктэлээх ~9 м, Дворцы – 15 м, Исить (по археоциатам) – 10 м, Журинский мыс – 10 м. В скв. Киси-Таас M. radiolata отмечена примерно в 5 м выше последних археоциат зоны D. regularis, однако, учитывая ограниченность материала из керна скважин, нельзя исключить, что археоциаты зоны D. lenaicus здесь пропущены, и нижнюю границу одноименной зоны в действительности следует проводить немного ниже.
Важно напомнить, что атдабанский ярус был установлен как последовательность комплексных подразделений по археоциатам, а уровень первого появления трилобитов рассматривался лишь как дополнительная характеристика нижней границы яруса (Журавлева и др., 1969). Таким образом, использование M. radiolata в качестве вида-индекса яруса не противоречит его оригинальной концепции, так как первое появление этого вида надежно увязано с подошвой первой археоциатовой зоны атдабана – R. zegebarti. Тот факт, что первые трилобиты обнаружены заведомо выше рассматриваемого уровня (рис. 3), лишь подтверждает устоявшуюся идею о том, что появление древнейших трилобитов связано с атдабаном.
Изменение положения границы между томмотским и атдабанским ярусами не существенно меняет объем ярусов (менее чем на треть) и, следовательно, не влечет за собой номенклатурных изменений (Стратиграфический…, 2006). Так, в стратотипе томмотского яруса в разрезе Дворцы в связи с уменьшением мощности зоны D. lenaicus на 25 м (рис. 3) мощность яруса уменьшается с 86 до 61 м (на ~29%). В гипостратотипе атдабанского яруса в разрезе Журинский мыс в связи с увеличением мощности зоны R. zegebarti на 26 м мощность яруса увеличивается с примерно 200 до 226 м (на ~11%). Важным положительным моментом является сдвиг стратотипа границы атдабанского яруса и зоны R. zegebarti с границы между слоями 3 и 4 разреза Журинский мыс, которая резко подчеркнута литологически, на 26 м вниз, внутрь слоя 2 того же разреза, где не наблюдается видимых литологических изменений в монотонной толще вишнево-красных глинистых известняков. В таком случае стратотип границы яруса ОСШ России будет удовлетворять и международным требованиям, предъявляемым к лимитотипам (Remane et al., 1996; Стратиграфический…, 2006).
Рассмотрим распространение вида Mobergella radiolata за пределами стратотипического Лено-Алданского региона (рис. 4–6).
Учуро-Майский район. В Учуро-Майском районе M. radiolata отмечена в разрезе по р. Сэлиндэ примерно в 95 м выше подошвы пестроцветной свиты (в верхах слоя 36; Репина и др., 1988, рис. 3). В 6–7 м выше этого уровня (низы слоя 37) встречены первые трилобиты Profallotaspis privica Rep., еще в 4 м выше – P. jakutensis Rep. (низы слоя 38). Выше, в верхах слоя 38, отмечены трилобиты Repinaella explicata (Rep.), в верхах слоя 41 – Repinaella sibirica (Rep.) и Bigotina malykanica Suv. В разрезе Сэлиндэ выделяются зоны томмотского яруса N. sunnaginicus, D. regularis и D. lenaicus по комплексам SSF, соответствующим стратотипическому региону, и зоны атдабанского яруса Profallotaspis jakutensis, Repinaella, Delgadella anabara–Nevadella по трилобитам. Нижняя граница атдабана проводится по первому появлению трилобита Profallotaspis privica, хотя в нескольких метрах ниже отмечаются неопределимые остатки трилобитов в шлифах (Репина и др., 1988). На наш взгляд, нижнюю границу яруса следует опустить на 6–7 м вниз до FAD вида M. radiolata, в таком случае все неопределимые фрагменты трилобитов будут заведомо атдабанские. Определимые археоциаты в разрезе Сэлиндэ появляются на более высоких уровнях – только в зоне Delgadella anabara.
Под различными названиями (Discinella brastadi Poulsen, 1932, Discinella sp., Brastadella brastadi (Poulsen, 1932), Mobergella braastadi (Poulsen, 1932), M. radiolata Bengtson, 1968, M. sibirica Skovsted, 2003, M. hexactina Skovsted, 2003, Mobergella sp. и др.; синонимику и ревизию см. Демиденко, 2016) вид описан и в других регионах Сибирской платформы: из средней подсвиты тюсэрской свиты на севере платформы в нижнем течении р. Лена (Розанов и др., 1969; Мешкова, 1974; Репина и др., 1974; Коровников, Новожилова, 2012); из верхов краснопорожской свиты скв. ИС-133 в Игарском районе на западе платформы (Сухов и др., 2016); в эмяксинской свите, скв. Танхайская-708 Восточного Прианабарья (Сухов и др., 2016); в сыгдахской свите скв. Моркокинская-707 внутренних районов платформы (Сухов и др., 2016).
Хребет Хараулах. В нижнем течении р. Лена на хр. Туора-Сис (Хараулахские горы) M. radiolata отмечена в трех разрезах в пределах Булкурской и Чекуровской антиклиналей: разрез выше устья руч. Улахан-Алдьярхай (Репина и др., 1974, разрез 1), разрез возле устья руч. Тюсэр на правом берегу р. Лена (Репина и др., 1974, разрез 3) и разрез в 4 км ниже пос. Чекуровка на левом берегу р. Лена (Репина и др., 1974, разрез 4). В разрезе Улахан-Алдьярхай мобергелла обнаружена в интервале44 11.5–17 м от основания средней подсвиты тюсэрской свиты. В интервале 18–23 м из биогермов определен раннеатдабанский комплекс археоциат (Репина и др., 1974; Репина, Розанов, 1992). Здесь же в интервале 20–23 м из биогермов описаны трилобиты Profallotaspis tyusserica Bushuev et Goryaeva, 2014 (Bushuev et al., 2014). Выше на уровне ~30 м отмечены Repinaella sp., а на уровне 36 м и выше уже в верхней подсвите тюсэрской свиты – трилобиты зоны Delgadella anabara–Nevadella: Delgadella pervulgata (Lazarenko, 1962), D. anabara (Lazarenko, 1962), D. sakhaica (Nikiforov, 1974), Nevadella effusa Repina, 1974, Paranevadella subgroenlandica (Repina, 1965) (Репина и др., 1974; Bushuev et al., 2014).
В разрезе Тюсэр мобергелла отмечена на нескольких уровнях: 4, 10, 11 и 14 м от основания средней подсвиты тюсэрской свиты, здесь же на уровне 13 м определены трилобиты Repinaella explicata, а в интервале 7–14 м – находки археоциат, распространенных в зонах D. lenaicus–R. zegebarti. В разрезе Чекуровка мобергелла присутствует на трех уровнях: 12, 14 и 15 м от основания средней подсвиты тюсэрской свиты, а выше на уровне ~22 м от основания подсвиты определены D. anabara и Nevadella sp. (Репина и др., 1974).
Биостратиграфическое расчленение разреза тюсэрской свиты Хараулаха выглядит следующим образом (рис. 5). Нижняя подсвита (мощность 26 м) по находкам вида зоопроблематик Lapworthella tortuosa в 2.5 м выше основания свиты относится к низам зоны D. regularis, подзоне Lapworthella tortuosa (Коровников, Новожилова, 2012). В 16–20 м от основания свиты фиксируется подошва зоны D. lenaicus по находкам археоциат и SSF (Astashkin et al., 1991). В средней подсвите (мощность 41 м) начиная с отметки 12 м выше ее подошвы отмечены находки мобергелл, что позволяет этот уровень считать подошвой атдабанского яруса и зоны R. zegebarti. Расположенные выше в 6 м (или в 1 м по Bushuev et al., 2014, рис. 3) биогермы с археоциатами принадлежат этой же зоне (Репина, Розанов, 1992). Трилобиты зоны Profallotaspis jakutensis появляются в 24 м выше подошвы атдабанского яруса, определенного по Mobergella radiolata. Основание зоны Delgadella anabara фиксируется на уровне 38 м от подошвы яруса.
Таким образом, характер зонального расчленения и даже мощности зон Хараулахских разрезов близки к таковым стратотипического района. Здесь также M. radiolata и первые атдабанские археоциаты появляются в разрезе практически одновременно, а первые трилобиты – чуть позже.
Игарский район. В Игарском районе на западе платформы M. radiolata отмечена на уровне ~42 м выше подошвы краснопорожской свиты в разрезе скв. ИС-133 вместе с зоопроблематиками Rhombocorniculum insolutum Miss., мощность свиты здесь ~58 м (Сухов и др., 2016, рис. 28 ). В этом же районе в разрезе по р. Сухариха нижняя граница атдабанского яруса по фауне археоциат фиксируется на уровне 12–13 м (Rowland et al., 1998) или 15–17 м (Розанов и др., 1969) выше подошвы краснопорожской свиты, а мощность самой свиты оценивается от 135 м (Kouchinsky et al., 2007) до 150–200 м (Розанов и др., 1969; Репина, Розанов, 1992). Поэтому находки мобергелл в скв. ИС-133, по-видимому, не раннеатдабанские, а должны быть датированы серединой века, временем Delgadella anabara, что не противоречит совместному нахождению с зоопроблематиками Rhombocorniculum insolutum, которые распространены в первой половине атдабана от зоны P. jakutensis до зоны D. anabara (Kouchinsky et al., 2015)55. Отсутствие находок археоциат и трилобитов в разрезе скважины не позволяет более точно установить биостратиграфические границы.
Восточное Прианабарье. В Восточном Прианабарье M. radiolata отмечена в эмяксинской свите (мощность 171 м) скв. Танхайская-708 (Сухов и др., 2016, табл. 2 , лист 22). Вид встречается на уровне 16–26 м от подошвы свиты вместе со среднеатдабанскими археоциатами Geocyathus latini (Zhur.) и G. botomaensis Zhur. зоны Carinacyathus pinus. Выше на уровне 131–142 м от подошвы свиты определены трилобиты Delgadella sp., Triangulaspis sp. Поэтому находки мобергелл в скв. Танхайская-708 следует также датировать временем Delgadella anabara/Carinacyathus pinus.
Внутренние районы платформы. В центральных областях Сибирской платформы мобергелла определена в 24–36 м от подошвы сыгдахской свиты (мощность 326 м) скв. Моркокинская-707 на границе томмотского и атдабанского ярусов (Сухов и др., 2016, табл. 2 , лист 15). Совместно встреченный комплекс SSF (Majatheca sp., Hyolithellus tenuis Miss., H. vladimirovae Miss., Lapworthella sp., Camenella admiranda Miss., Archiasterella sp.) близок к таковому из зоны Profallotaspis jakutensis середины средней подсвиты тюсэрской свиты Хараулаха (Коровников, Новожилова, 2012, рис. 3).
ЯРУС 3 МЕЖДУНАРОДНОЙ СТРАТИГРАФИЧЕСКОЙ ШКАЛЫ
Рассмотрим распространение вида Mobergella radiolata за пределами Сибирской платформы и определим возможность использования этого таксона как вида-индекса нижней границы яруса 3 МХШ.
Балтика. Это еще один палеоконтинент, где широко распространена Mobergella radiolata. Здесь этот вид встречен на территории современных Швеции, Дании, Эстонии, Польши и Европейской России. На юге Швеции M. radiolata известна из низов формации Норреторп Скании и датского острова Борнхольм (Bengtson, 1968; Bergström, 1981; Nielsen, Schovsbo, 2007, 2011), из низов формации Филе-Хайдар (Песчаники Калмарсунд и Песчаники Mickwitzia) полуострова Скяггенас и балтийских островов Эланд и Готланд (Bengtson, 1968; Jensen, 1997; Nielsen, Schovsbo, 2007, 2011). Эти толщи коррелируются с низами зоны Schmidtiellus mickwitzi (Bengtson, 1968, 1970, 1977; Bergström, 1981; Jensen, 1997; Kouchinsky et al., 2012), в которой встречаются древнейшие трилобиты Балтики (см. выше), но есть мнение и о более молодом возрасте песчаников Mickwitzia – их иногда сопоставляют с зоной Holmia kjerulfi (Moczydłowska, 1991; Nielsen, Schovsbo, 2007, 2011). В Западной Эстонии M. radiolata описана из люкатиских слоев и низов тискреских слоев (Менс, Пиррус, 1977), которые коррелируются с зоной Schmidtiellus mickwitzi по трилобитам и акритархам (Bergström, 1973; Менс, Пиррус, 1977; Mens et al., 1990; Moczydłowska, 1991, 2011, рис. 2; Meidla, 2017, рис. 2). В Восточной Польше M. radiolata известна из вскрытых скважинами завишинских слоев, а на севере Польши – из мобергелловых слоев (Lendzion, 1972, 1975, 1983; Арень, Лендзён, 1974; Dzik, Lendzion, 1988). В завишинских слоях совместно с мобергеллами найдены слабоминерализованные артроподы – аномалокариды Cassubia infercambriensis (Lendzion, 1975), трилобиты Liwia plana (Lendzion, 1975) и L. convexa (Lendzion, 1975). Завишинские и мобергелловые слои Польши также коррелируют с зоной Schmidtiellus mickwitzi Скандинавии (Moczydłowska, 1991, рис. 8 ) или чуть более древними толщами (Dzik, Lendzion, 1988). Находки мобергелл в глебовских слоях верхов лонтоваского горизонта Московской синеклизы, верхи зоны Platysolenites antiquissimus (Розанов, 1973, рис. 134 ; Кирсанов, 1974; Rozanov, Zhuravlev, 1992)66, вероятно, являются самыми древними в Балтике, и их следует сопоставлять с базальным атдабаном Сибирской платформы (рис. 1).
Таким образом, в Балтике Mobergella radiolata распространена крайне широко, но подавляющее большинство находок приурочено к зоне Schmidtiellus mickwitzi, что соответствует верхней половине зоны Repinaella и значительной части зоны Delgadella anabara на Сибирской платформе. Как отмечалось выше, в Сибири на этом уровне M. radiolata также широко распространена. И только для Московской синеклизы Восточно-Европейской платформы можно говорить о вероятной синхронности первого появления мобергелл на палеоконтинентах Балтики и Сибири.
Гондвана. На крупнейшем кембрийском континенте Гондвана достоверных находок Mobergella radiolata пока не отмечено. Однако следует обратить внимание на ископаемых из нижнего кембрия пров. Гуйчжоу (Южно-Китайская платформа), описанных как Mobergella? guizhouensis Qian et Yin, 1984 (Qian, Yin, 1984, p. 112, pl. 4, fig. 6, pl. 5, figs. 12, 13 ; Wang et al., 1984, pl. 4, fig. 8 ). На фототаблицах изображена мобергелла удовлетворительной сохранности с грубыми, радиально расходящимися складками на поверхности склерита. Близкий облик имеют частично разрушенные склериты M. radiolata из сибирских разрезов (Демиденко, 2016, рис. 2, в–е; Демиденко, Пархаев, 2016, рис. 2, а). M.? guizhouensis происходит из верхов формации Гежонву и самых низов формации Нютитан в разрезе Гежонву, округ Жицзинь (Wei et al., 2018, рис. 2). Выше в разрезе с низов верхней пачки формации Нютитан встречены трилобиты Tsunyidiscus niutitangensis (Chang, 1964), T. qingzhenensis Yin in Yin et Li, 1978, Tsunyidiscus sp. (=Guizhoudiscus sp.). Очевидно, что пачка с трилобитами относится к подзоне Tsunyidiscus (Peng et al., 2012) низов зоны Eoredlichia–Wutingaspis (Steiner et al., 2007), которая соответствует среднему–верхнему атдабану (см. выше). Поэтому нижележащие толщи могут быть датированы ранним–средним атдабаном, а именно в этом временном интервале следует ожидать находки M. radiolata. В этой связи дополнительное изучение палеонтологической характеристики разреза Гежонву, пров. Гуйчжоу, Южный Китай крайне желательно.
Авалония. В литературе встречаются упоминания о находках Mobergella radiolata на территории Авалонии (Великобритания, разрез Комли в гр. Шропшир) (Ярусное…, 1984, с. 152). Однако следует признать, что они относятся к другому виду мобергелл – M. turgida Bengtson, 1968 (Bengtson, 1977; Brasier, 1989; Демиденко, 2016), а вид Mobergella radiolata в Авалонии до сих пор не найден. Но тот факт, что M. turgida встречается вместе с M. radiolata в серии разрезов Балтики (Bengtson, 1968, 1977; Арень, Лендзён, 1974), не оставляет сомнений в том, что слои с Obolella groomi основания песчаников Комли (слой Ab1) с мобергеллами следует сопоставлять с зоной/слоями с Mobergella Балтики и с верхней частью интервала распространения M. radiolata на Сибирской платформе.
Палеонезия. Палеонезией называют группу террейнов или микроконтинентов, существовавших в Палеоазиатском океане в позднем докембрии–раннем палеозое к востоку от территории Сибирской платформы (Levashova et al., 2011, рис. 9 ). В их число входили казахстанский и монгольский блоки. Находки Mobergella sp. в низах ботомского яруса хаирханской свиты разреза Саланы-Гол Западной Монголии (Коробов, Миссаржевский, 1977; Воронин и др., 1982, с. 40, рис. 3) были переописаны (Миссаржевский, 1989) как новый вид Discinella mongolica Missarzhevsky, 1989, а впоследствии эту форму отнесли к роду Hippoklosma (Conway Morris, Chapman, 1997). На настоящий момент виды рода Mobergella в кембрии Монголии достоверно не известны.
Mobergella radiolata была описана из мобергелловых слоев Центрального Казахстана (Конева, 1983; Гридина, 1990), разрез р. Селета, керимбайская свита. Однако встречающиеся совместно трилобиты Labradoria sp., Kootenia sp. и Bajanaspis sp. свидетельствуют о ботомском возрасте вмещающих отложений (Гридина, 1990). Переизучение материала может уточнить вопрос – действительно ли M. radiolata в Казахстане существовала в ботомском веке, или здесь мы имеем дело с ошибочным определением, и находки должны быть отнесены к другому виду.
Подводя итог анализа распространения вида Mobergella radiolata в мире, можно констатировать следующее. На территории Сибирской платформы (практически все основные структурно-фациальные районы) и Балтики (Восточно-Европейская платформа, Скандинавия) появление вида в разрезах четко свидетельствует о раннеатдабанском возрасте вмещающих отложений. Наличие региональных и локальных перерывов различного масштаба на границе возрастных аналогов томмотского и атдабанского ярусов в разрезах Балтики (Mens et al., 1990) и, соответственно, возможное отсутствие здесь самых базальных горизонтов атдабана не препятствует использованию таксона M. radiolata как вида-индекса ярусного подразделения. На территории Гондваны возможные находки мобергелл в дотрилобитовых толщах пров. Гуйчжоу, Южный Китай служат важным дополнением к корреляционному потенциалу обсуждаемого вида. Поэтому нам представляется целесообразным использовать M. radiolata для установления нижней границы яруса 3 МХШ. В любом случае, данный вид зоопроблематик обеспечивает несравнимо более широкую и надежную корреляцию, чем какой-либо из видов древнейших трилобитов или представителей других групп ископаемых.
ЗАКЛЮЧЕНИЕ
1. Мы предлагаем считать стратотипом нижней границы комплексной зоны R. zegebarti и нижней границы атдабанского яруса ОСШ уровень первого появления вида зоопроблематик Mobergella radiolata в средней части пестроцветной свиты в 10 м выше подошвы слоя 2 разреза Журинский мыс. В таком случае (а) решается проблема четырехкратного разброса в мощности зоны D. lenaicus в разрезах среднего течения рек Алдан и Лена, и мощность фиксируется в пределах 9–15 м; (б) серия разрезов, в которых отсутствуют находки археоциат и нижняя граница атдабанского яруса проводится условно, получает надежную биостратиграфическую характеристику для обоснования границы яруса; (в) представитель микрофауны Mobergella radiolata, как биостратиграфический инструмент, имеет очевидные преимущества перед макрофауной – археоциатами и трилобитами, находки которых имеют узкую фациальную приуроченность и довольно редки даже в пределах стратотипического региона. Mobergella radiolata встречается за пределами стратотипического региона (Учуро-Майский район, Хараулах, Игарский район, Восточное Прианабарье, внутренние районы платформы), что позволяет проводить надежную межрегиональную корреляцию внутри Сибирской платформы. Использование Mobergella radiolata для обоснования подошвы атдабанского яруса не противоречит оригинальной концепции яруса и сохраняет атдабан как время первого появления трилобитов в геологической летописи.
2. Мы предлагаем использовать Mobergella radiolata как вид-индекс для обоснования нижней границы яруса 3 МХШ. В таком случае (а) нижняя граница яруса 3 уверенно распознается в разрезах нескольких палеоконтинентов: Сибирии, Балтики и, возможно, Западной Гондваны (Южный Китай); (б) век 3 остается как время первого появления трилобитов в глобальном масштабе, так как уровень первого появления Mobergella radiolata на всех палеконтинентах заведомо ниже уровня появления первых трилобитов; (в) использование одного и того же вида ископаемых для определения нижней границы атдабанского яруса ОСШ и яруса 3 МХШ позволяет однозначно сопоставлять эти подразделения и, фактически, использовать атдабанский ярус как подразделение МХШ.
В заключение отметим, что в одной из последних крупных сводок по стратиграфии и фауне кембрия Сибирской платформы (Сухов и др., 2016, рис. 2, 4) в базальной части атдабанского яруса присутствует подразделение слои с “Mobergella radiolata”, соответствующее одной зоне по археоциатам Retecoscinus zegebarti/Leptosocyathus polyseptus и двум зонам по трилобитам Profallotaspis jakutensis и Repinaella. Таким образом, идея об использовании M. radiolata как важной формы для определения нижней границы атдабанского яруса ОСШ России уже находит поддержку среди специалистов.
Источники финансирования. Работа выполнена при поддержке РФФИ, проекты №№ 16-05-00651, 19-04-01027.
Список литературы
Арень Б., Лендзён К. Распространение и стратиграфия климонтовского яруса нижнего кембрия на платформе в Польше // Биостратиграфия и палеонтология нижнего кембрия Европы и Северной Азии. М.: Наука, 1974. С. 30–35.
Варламов А.И., Розанов А.Ю., Хоментовский В.В. и др. Кембрий Сибирской платформы. Книга 1: Алдано-Ленский регион. М.: ПИН РАН, 2008. 300 с.
Варламов А.И., Розанов А.Ю., Демиденко Ю.Е. и др. Проект кембрийской части общей стратиграфической шкалы России // Матералы Всероссийского совещания “Общая стратиграфическая шкала России: состояние и перспективы обустройства”, Москва, 23–25 мая 2013 г. Ред. Федонкин М.А. М.: ГИН РАН, 2013. С. 79–87.
Васильева Н.И. Мелкая раковинная фауна и биостратиграфия нижнего кембрия Сибирской платформы. СПб.: ВНИГРИ, 1998. С. 1–139.
Воронин Ю.И., Воронова Л.Г., Григорьева Н.В. и др. Граница докембрия и кембрия в геосинклинальных областях (опорный разрез Саланы-Гол, МНР). М.: Наука, 1982. 152 с. (Тр. ССМПЭ. Вып. 18).
Гридина Н.М. Фосфатные проблематики в отложениях нижнего кембрия реки Селеты (Центральный Казахстан) // Тезисы докладов 3-го Междунар. симпозиума по кембрийской системе. Новосибирск: ИГиГ СО АН СССР, 1990. С. 25.
Демиденко Ю.Е. Морфология, систематическое положение и стратиграфическое распространение раннекембрийских скелетных проблематик Mobergella radiolata Bengtson, 1968 // Палеонтол. журн. 2016. № 5. С. 10–22.
Демиденко Ю.Е., Пархаев П.Ю. Мелкораковинная фауна, внедрение химического препарирования кембрийских ископаемых и новые данные по микроструктуре склеритов Mobergella radiolata // Палеонтология. Стратиграфия. Астробиология. К 80-летию академика А.Ю. Розанова. Ред. Рожнов С.В. М.: ПИН РАН, 2016. С. 188–194.
Дэйли Б. Новые данные об основании кембрия в Южной Австралии // Изв. Акад. наук СССР. Сер. геол. 1976. № 3. С. 45–52.
Журавлева И.Т., Коршунов В.И., Розанов А.Ю. Атдабанский ярус и его обоснование по археоциатам в стратотипическом разрезе // Биостратиграфия и палеонтология нижнего кембрия Сибири и Дальнего Востока. М.: Наука, 1969. С. 5–59.
Журавлева И.Т., Коршунов В.И., Лучинина В.А. и др. Опорные скважины верхнего докембрия–нижнего кембрия в стратотипическом районе среднего течения р. Лены // Биостратиграфия и палеонтология нижнего и среднего кембрия Азиатской части СССР. М.: Наука, 1983. С. 3–45.
Кирсанов В.В. К вопросу о стратиграфии пограничных слоев венда и кембрия в центральных районах Восточно-Европейской платформы // Биостратиграфия и палеонтология нижнего кембрия Европы и Северной Азии. М.: Наука, 1974. С. 5–21.
Конева С.П. Mobergella из нижнего кембрия Селетинского синклинория // Стратиграфия и палеонтология нижнего палеозоя Казахстана. Алма-Ата: Наука, 1983. С. 110–112.
Коробов М.Н., Миссаржевский В.В. О пограничных слоях кембрия и докембрия Западной Монголии (хребет Хасагт-Хайрхан) // Беспозвоночные палеозоя Монголии. М.: Наука, 1977. С. 7–9.
Коровников И.В., Новожилова Н.В. Новые данные по биостратиграфии нижнего и низов среднего кембрия Хараулахских гор (северо-восток Сибирской платформы, Чекуровская антиклиналь) // Геология и геофизика. 2012. Т. 53. № 8. С. 1014–1026.
Менс К., Пиррус Э. Стратотипические разрезы кембрия Эстонии. Таллин: Валгус, 1977. 68 с.
Мешкова Н.П. Хиолиты нижнего кембрия Сибирской платформы. Новосибирск: Наука, 1974. 110 с.
Миссаржевский В.В. Древнейшие скелетные окаменелости и стратиграфия пограничных толщ докембрия и кембрия. М.: Наука, 1989. 237 с. (Тр. Геол. ин-та АН СССР. Вып. 443).
Пархаев П.Ю. Кембрийские моллюски Австралии: обзор таксономии, биостратиграфии и палеобиогеографии // Стратиграф. Геол. корреляция. 2019а. Т. 27. № 2. С. 52–79.
Пархаев П.Ю. Находка моллюсков Watsonella crosbyi Grabau (Gastropoda: Helcionelliformes) в ботомских отложениях Китая // Докл. АН. 2019б. Сер. геол. Т. 488. № 6. С. 624–629.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Л.: ВСЕГЕИ, 1983. Вып. 21. 74 с.
Путеводитель экскурсии по рекам Алдану и Лене (Международная экскурсия по проблеме границы кембрия и докембрия). Якутск: ВИЭМС, 1973. 118 с.
Решения Всесоюзного стратиграфического совещания по докембрию, палеозою и четвертичной системе Средней Сибири. Часть 1 (верхний протерозой и нижний палеозой). Новосибирск: СибРМСК, 1983. 216 с.
Репина Л.Н. Планетарная корреляция ярусных подразделений нижнего кембрия по трилобитам // Биостратиграфия и палеонтология кембрия Северной Азии. М.: Наука, 1986. С. 4–25 (Тр. Ин-та геологии и геофизики СО АН СССР. Вып. 669).
Репина Л.Н., Розанов А.Ю. (ред.). Кембрий Сибири. Новосибирск: Наука, 1992. 135 с.
Репина Л.Н., Лазаренко Н.П., Мешкова Н.П. и др. Биостратиграфия и фауна нижнего кембрия Хараулаха. М.: Наука, 1974. 300 с. (Тр. Ин-та геологии и геофизики СО АН СССР. Вып. 235).
Репина Л.Н., Бородаевская З.В., Ермак В.В. Опорный разрез по р. Сэлиндэ (юго-восточная окраина Алданского щита) // Кембрий Сибири и Средней Азии. М.: Наука, 1988. С. 3–31 (Тр. Ин-та геологии и геофизики СО АН СССР. Вып. 720).
Розанов А.Ю. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука, 1973. 164 с. (Тр. Геол. ин-та АН СССР. Вып. 241).
Розанов А.Ю., Миссаржевский В.В. Биостратиграфия и фауна нижних горизонтов кембрия. М.: Наука, 1966. 127 с. (Тр. Геол. ин-та АН СССР. Вып. 148).
Розанов А.Ю., Миссаржевский В.В., Волкова Н.А. и др. Томмотский ярус и проблема нижней границы кембрия. М.: Наука, 1969. 380 с. (Тр. Геол. ин-та АН СССР. Вып. 206).
Розанов А.Ю., Семихатов М.А., Соколов Б.С. и др. Решение о выборе стратотипа границы докембрия и кембрия: прорыв в проблеме или ошибка? // Стратиграфия. Геол. корреляция. 1997. № 1. С. 21–31.
Розанов А.Ю., Хоментовский В.В., Шабанов Ю.Я. и др. К проблеме ярусного расчленения нижнего кембрия // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 1. С. 3–21.
Розанов А.Ю., Пархаев П.Ю., Демиденко Ю.Е. и др. Ископаемые стратотипов ярусов нижнего кембрия. М.: ПИН РАН, 2010. 228 с.
Скорлотова Н.А. Археоциаты стратотипических разрезов томмотского и атдабанского ярусов. Автореф. дисс. … канд. геол.-мин. наук. М.: ПИН РАН, 2013. 16 с.
Стратиграфический кодекс России. Изд. 3-е. СПб.: ВСЕГЕИ, 2006. 96 с.
Сухов С.С., Шабанов Ю.Я., Пегель Т.В. и др. Стратиграфия нефтегазоносных бассейнов Сибири. Кембрий Сибирской платформы. Т. 1. Стратиграфия. Новосибирск: ИНГГ СО РАН, 2016. 498 с.
Хоментовский В.В., Репина Л.Н. Нижний кембрий стратотипического разреза Сибири. М.: Наука, 1965. 200 с.
Ярусное расчленение нижнего кембрия. Стратиграфия. Ред. Розанов А.Ю., Соколов Б.С. М.: Наука, 1984. 184 с.
Ahlberg P. Trilobites in the Lower Cambrian of Scandinavia // GFF. 1991. V. 113. P. 74–75.
Ahlberg P., Bergström J., Johansson J. Lower Cambrian olenellid trilobites from the Baltic faunal province // GFF. 1986. V. 108. Pt. 1. P. 39–56.
Astashkin V.A., Pegel T.V., Repina L.N. et al. The Cambrian System on the Siberian Platform. Correlation chart and explanatory notes // Int. Union Geol. Sci. Spec. Publ. 1991. № 27. P. 1–133.
Astashkin V.A., Belyaeva G.V., Esakova N.V. et al. The Cambrian System of the foldbelts of Russia and Mongolia. Correlation chart and explanatory notes // Int. Union Geol. Sci. Spec. Publ. 1995. № 32. P. 1–132.
Babcock L.E., Peng S. Cambrian chronostratigraphy: current state and future plans // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007. V. 254. P. 62–66.
Babcock L.E., Peng S., Geyef G., Shergold J.H. Changing perspectives on Cambrian chronostratigraphy and progress toward subdivision of the Cambrian System // Geosci. J. 2005. V. 9. P. 101–106.
Babcock L.E., Peng S., Zhu M. et al. Proposed reassessment of the Cambrian GSSP // J. African Earth Sci. 2014. V. 98. P. 3–10.
Bengtson S. The problematic genus Mobergella from the Lower Cambrian of the Baltic area // Lethaia. 1968. V. 1. P. 325–351.
Bengtson S. The Lower Cambrian fossil Tommotia // Lethaia. 1970. V. 3. P. 363–392.
Bengtson S. Aspects of problematic fossils in the early Palaeozoic // Acta Univ. Upsaliensis. Abstr. Uppsala Dissert. Faculty of Sci. 1977. V. 415. P. 1–71.
Bengtson S., Conway Morris S., Cooper B. et al. Early Cambrian fossils from South Australia // Mem. Assoc. Austral. Palaeontol. 1990. № 9. P. 1–364.
Bergström J. Classification of olenellid trilobites and some Balto-Scandian species // Norsk Geol. Tidsskrift. 1973. V. 53. P. 283–314.
Bergström J. Lower Cambrian shelly faunas and biostratigraphy in Scandinavia // Short Pap. 2nd Int. Sympos. on Cambrian system. Golden: Colorado School of Mines, 1981. P. 22–25.
Brasier M.D. Towards a biostratigraphy of the earliest skeletal biotas // The Precambrian–Cambrian Boundary. Eds. Cowie J.W., Brasier M.D. Oxford: Clarendon Press, 1989. P. 117–165.
Brasier M.D., Anderson M.M., Corfield R.M. Oxygen and carbon isotope stratigraphy of Early Cambrian carbonates in southeastern Newfoundland and England // Geol. Mag. 1992. V. 129. P. 265–279.
Brasier M.D., Cowie J., Taylor M. Decision on the Precambrian–Cambrian boundary // Episodes. 1994. V. 17. P. 3–8.
Bushuev E., Goryaeva I., Pereladov V. New discoveries of the oldest trilobites Profallotaspis and Nevadella in the northeastern Siberian Platform, Russia // Bull. Geosci. 2014. V. 89. № 2. P. 347–364.
Conway Morris S., Chapman A.J. Mobergellans from the Lower Cambrian of Mongolia, Sweden, and the United States: Molluscs or opercula of incertae sedis? // J. Paleontol. 1997. V. 71. № 6. P. 968–984.
Cowie J.W., Basset M.G. Global Stratigraphic Chart with geochronometric and magnetostratigraphic calibration // Episodes. 1989. V. 12. № 2. Supplement.
Daily B. The base of the Cambrian and the first Cambrian faunas // Stratigraphic problems of the Later Precambrian and Early Cambrian. Eds. Jones J.B., McGowran B. Univ. Adelaide Centre Precambr. Res. Spec. Pap. 1972. № 1. P. 13–41.
Debrenne F., Debrenne M. Archaeocyathid fauna of the lowest fossiliferous levels of Tiout (Lower Cambrian, Southern Morocco) // Geol. Mag. 1978. V. 115. № 2. P. 101–120.
Debrenne F., Zhuravlev A.Yu., Kruse P.D. General features of the Archaeocyatha. Systematic descriptions: Archaeocyatha // Treatise on Invertebrate Paleontology. Part E: Porifera revised, Hypercalcified Porifera, Volume 5. Ed. Selden P.A. Lawrence: Univ. of Kansas Press, 2015. P. 845–1084.
Demidenko Yu.E., Parkhaev P.Yu., Rozanov A.Yu. Morphological variability and types of preservation of Mobergella radiolata – a potential index species for the GSSP of Cambrian Stage 2 // J. Guizhou Univ. 2012. V. 29. Suppl. 1. P. 157–158.
Dzik J., Lendzion K. The oldest arthropods of the East European Platform // Lethaia. 1988. V. 21. P. 29–38.
Fletcher T.P. Bedrock geology of the Cape St. Mary’s Peninsula, Southwest Avalon Peninsula, Newfoundland. St. John’s: Gov. of Newfoundland and Labrador, 2006. P. 1–117.
Geyer G. Intercontinental trilobite-based correlation of the Moroccan early Middle Cambrian // Canad. J. Earth Sci. 1998. V. 35. P. 374–401.
Geyer G., Landing E. The Cambrian of the Moroccan Atlas regions // Beringeria. 1995. Spec. № 2. P. 7–46.
Geyer G., Landing E. Morocco-2006: Ediacaran–Cambrian depositional environments and stratigraphy of the western Atlas regions // Beringeria. 2006. Spec. № 6. P. 1–121.
Geyer G., Palmer A.R. Neltneriidae and Holmiidae (Trilobita) from Morocco and the problem of Early Cambrian intercontinental correlation // J. Paleontol. 1995. V. 69. P. 459–474.
Geyer G., Shergold J. The quest for internationally recognized divisions of Cambrian time // Episodes. 2000. V. 23. № 3. P. 188–195.
Gravestock D.I., Alexander E.M., Demidenko Yu.E. et al. The Cambrian biostratigraphy of the Stansbury Basin, South Australia. Moscow: MAIK Nauka/Interperiodica, 2001. 344 p. (Trans. Paleontol. Inst. RAS. V. 282).
Guide book for excursion on the Aldan and Lena rivers. The Siberian Platform. 3rd Int. Sympos. on Cambrian System. Eds. Zhuravlev A.Yu., Repina L.N. Novosibirsk: IGiG SO AN SSSR, 1990. 120 p.
Hagadorn J.W., Waggoner B. The Early Cambrian problematic fossil Volborthella: new insights from the basin and range // Proterozoic–Cambrian of the Great Basin and Beyond. Ed. Corsetti F.A. SEPM, Pacific Section. 2002. Book 93. P. 135–150.
Hollingsworth J.S. The earliest occurrence of trilobites and brachiopods in the Cambrian of Laurentia // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005. V. 220. P. 153–165.
Hollingsworth J.S. Fallotaspidoid trilobite assemblage from the Esmeralda Basin (western Nevada, USA.): the oldest trilobites from Laurentia // Mem. Austral. Assoc. Palaeontol. 2007. V. 32. P. 123–140.
Jago J.B., Zang W., Sun X. et al. A review of the Cambrian biostratigraphy of South Australia // Palaeoworld. 2006. V. 15. P. 406–423.
Jell P.A. Phylogeny of Early Cambrian trilobites // Spec. Pap. Palaeontol. 2003. V. 70. P. 45–57.
Jensen S. Trace fossils from the Lower Cambrian Mickwitzia sandstone // Fossil and Strata. 1997. V. 42. P. 1–111.
Korovnikov I.V. Correlation potential FADs of the Triangulaspis annio and Hebediscus attleborensis on the Siberian platform (a possible GSSP for the lower boundary of the Cambrian Stage 4) // J. Guizhou Univ. 2012. V. 29. Suppl. 1. P. 169–170.
Kouchinsky A., Bengtson S., Pavlov V. et al. Carbon isotope stratigraphy of the Precambrian–Cambrian Sukharikha River section, northwestern Siberian platform // Geol. Mag. 2007. V. 144. № 3. P. 1–10.
Kouchinsky A., Bengtson S., Clausen S. et al. A middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia // Alcheringa. 2011. V. 35. № 1. P. 123–189.
Kouchinsky A., Bengtson S., Runnegar B. et al. Chronology of early Cambrian biomineralization // Geol. Mag. 2012. V. 149. № 2. P. 221–251.
Kouchinsky A., Bengtson S., Clausen S., Vendrasco M.J. An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia // Acta Palaeontol. Polon. 2015. V. 60. № 2. P. 421–512.
Kruse P.D., Zhuravlev A.Yu., Parkhaev P.Yu., Zhu M. Comment: a new lower Cambrian shelly fossil biostratigraphy for South Australia by Marissa J. Betts, John R. Paterson, James B. Jago, Sarah M. Jacquet, Christian B. Skovsted, Timothy P. Topper & Glenn A. Brock // Gondwana Res. 2017. V. 44. P. 258–261.
Landing E. Precambrian–Cambrian boundary global stratotype ratified and a new perspective of Cambrian time // Geology. 1994. V. 22. P. 179–182.
Landing E., Bowring S.A., Davidek K.L. et al. Duration of the Cambrian: U–Pb ages of the volcanic ashes from Avalon and Gondwana // Canad. J. Earth Sci. 1998. V. 35. P. 329–338.
Landing E., Peng S., Babcock L.E. et al. Global standard names for the Lowermost Cambrian Series and Stage // Episodes. 2007. V. 30. № 4. P. 287–289.
Lazarenko N.P., Gogin I.Y., Pegel T.V., Abaimova G.P. The Khos-Nelege section of the Ogon’or Formation: a potential candidate for the GSSP of Stage 10, Cambrian System // Bull. Geosci. 2011. V. 86. № 3. P. 555–568.
Lendzion K. Kambr subholmiowy W północno-wschodniei Polsce // Kwartalnik Geol. 1972. T. 16. № 3. P. 557–575.
Lendzion K. Fauna of the Mobergella zone in the Polish Lower Cambrian // Kwartalnik Geol. 1975. T. 19. № 2. P. 237–244.
Lendzion K. Mobergella // Upper Precambrian and Cambrian Palaeontology of the East European platform. Eds. Urbanek A., Rozanov A.Yu. Warszawa: Publ. House Wydawnictwa Geol., 1983. P. 121–123.
Levashova N.M., Meert J.G., Gibsher A.S. et al. The origin of microcontinents in the Central Asian Orogenic Belt: constraints from paleomagnetism and geochronology // Precambrian Res. 2011. V. 185. P. 37–54.
Li G., Zhao X., Gubanov A. et al. Early Cambrian Mollusc Watsonella crosbyi: A Potential GSSP Index Fossil for the Base of the Cambrian Stage 2 // Acta Geol. Sin. 2011. V. 85. № 2. P. 309–319.
Lieberman B.S. Systematic Revision of the Olenelloidea (Trilobita, Cambrian) // Bull. Peabody Museum Nat. Hist. 1999. V. 45. P. 1–150.
Liñán E., Gozalo R., Dies Álvarez M. E. et al. Nuevos trilobites del Ovetiense inferior (Cámbrico Inferior bajo) de Sierra Morena (España) // Ameghiniana. 2008. V. 45. № 1. P. 123–138.
Liñán E., Gamez Vintaned J.A., Gozalo R. The middle lower Cambrian (Ovetian) Lunagraulos n. gen. from Spain and the oldest trilobite records // Geol. Mag. 2015. V. 152. № 6. P. 1123–1136.
Meidla T. Ediacaran and Cambrian stratigraphy in Estonia: an updated review // Estonian J. Earth Sci. 2017. V. 66. № 3. P. 152–160.
Mens K., Bergström J., Lendzion K. The Cambrian System on the East European Platform. Correlation chart and explanatory notes // Int. Union Geol. Sci. Spec. Publ. 1990. № 25. P. 1–74.
Moczydłowska M. Acritarch biostratigraphy of the Lower Cambrian and the Precambrian–Cambrian boundary in southeastern Poland // Fossils and Strata. 1991. № 29. P. 1–127.
Moczydłowska M. The early Cambrian phytoplankton radiation: acritarch evidence from the Lükati Formation, Estonia // Palynology. 2011. V. 35. № 1. P. 103–145.
Nielsen A.T., Schovsbo N.H. Cambrian to basal Ordovician lithostratigraphy in southern Scandinavia // Bull. Geol. Soc. Denmark. 2007. V. 53. P. 47–92.
Nielsen A.T., Schovsbo N.H. The Lower Cambrian of Scandinavia: depositional environment, sequence stratigraphy and palaeogeography // Earth Sci. Rev. 2011. V. 107. P. 207–310.
Palmer A.R. A proposed nomenclature for stages and series for the Cambrian of Laurentia // Can. J. Earth Sci. 1998. V. 35. P. 323–328.
Palmer A.R., Repina L.N. Through a glass darkly: taxonomy, phylogeny, and biostratigraphy of the Olenellina // Univ. Kansas Paleontol. Contrib. 1993. № 3. P. 1–35.
Palmer A.R., Repina L.N. Suborder Olenellina // Treatise on Invertebrate Paleontology. Pt. O. Arthropoda 1. Trilobita. Rev. V. 1. Introduction, Order Agnostida, Order Redlichiida. Lawrence: Univ. Kansas Press, 1997. P. 404–428.
Palmer A.R., Rowell A.J. Early Cambrian trilobites from the Shackleton Limestone of the Central Transantarctic mountains // Mem. Paleontol. Soc. 1995. V. 45. P. 1–28.
Parkhaev P.Yu. On the stratigraphy of Aldanella attleborensis – potential index species for defining the base of Cambrian Stage 2 // Ext. Sum. IGCP Project 591 Field Workshop 2014. Nanjing: Nanjing Univ. Press, 2014. P. 102–105.
Parkhaev P.Yu., Karlova G.A., Rozanov A.Yu. Taxonomy, stratigraphy and biogeography of Aldanella attleborensis – a possible candidate for defining the base of Cambrian Stage 2 // Bull. Museum Northern Arizona. 2011. № 67. P. 298–300.
Parkhaev P.Yu., Karlova G.A., Rozanov A.Yu. Stratigraphic distribution of two potential species for the GSSP of Cambrian Stage 2 – Aldanella attleborensis and Watsonella crosbyi // J. Guizhou Univ. 2012. V. 29. Suppl. 1. P. 179–180.
Paterson J.R., Brock G.A. Early Cambrian trilobites from Angorichina, Flinders Ranges, South Australia, with a new assemblage from the Pararaia bunyerooensis Zone // J. Paleontol. 2007. V. 81. № 1. P. 116–142.
Peng J., Sun H., Zhao Y. et al. Introduction of Cryogenian to Cambrian Taijiangian stratigraphy from Central and Eastern Guizhou // J. Guizhou Univ. 2012. V. 29. Suppl. 1. P. 59–65.
Peng S., Babcock L.E., Zuo J. et al. Proposed GSSP for the base of Cambrian Stage 7, coinciding with the first appearance of Lejopyge laevigata, Hunan, China // Palaeoworld. 2006. V. 15. P. 367–383.
Peng S., Babcock L.E., Cooper R.A. Chapter 19. The Cambrian Period // The Geologic Time Scale 2012. Elsevier, 2012. P. 437–488.
Peng S., Babcock L.E., Zhu X. et al. Intraspecific variation and taphonomic alteration in the Cambrian (Furongian) agnostoid Lotagnostus americanus: new information from China // Bull. Geosci. 2015. V. 90. № 2. P. 281–306.
Perejon A., Moreno-Eiris E. Biostratigraphy and paleobiogeography of the archaeocyaths on the southwestern margin of Gondwana // Z. dt. Ges. Geowiss. 2006. V. 157. P. 611–627.
Pillola G.L. The Lower Cambrian trilobite Bigotina and allied genera // Palaeontology. 1993. V. 36. Pt. 4. P. 855–881.
Qian Y., Yin G. Small Shelly Fossils from the Lowerest Cambrian in Guizhou // Prof. Pap. Stratigr. Palaeontol. 1984. V. 13. P. 91–123.
Remane J., Bassett M.G., Cowie J.W. et al. Revised guidelines for the establishment of global chronostratigraphic standards by the International Commission on Stratigraphy (ICS) // Episodes. 1996. V. 19. № 3. P. 77–81.
Rowland S.M., Luchinina V.A., Korovnikov I.V. et al. Biostratigraphy of the Vendian–Cambrian Sukharikha River section, northwestern Siberian Platform // Canad. J. Earth Sci. 1998. V. 35. № 2. P. 339–352.
Rozanov A.Yu., Debrenne F. Age of archaeocyathid assemblages // Am. J. Sci. 1974. V. 274. P. 833–848.
Rozanov A.Yu., Zhuravlev A.Yu. Chapter 7. The Lower Cambrian fossil record of the Soviet Union // Origin and early evolution of the Metazoa. Eds. Lipps J.H., Signor P.W. New York: Plenum Press, 1992. P. 205–282.
Rozanov A.Yu., Parkhaev P.Yu, Shabanov Yu.Ya. et al. The 13th International Conference of the Cambrian Stage Subdivision Working Group // Episodes. 2008. V. 31. № 4. P. 440–441.
Rozanov A.Yu., Parkhaev P.Yu., Demidenko Yu.E., Skorlotova N.A. Mobergella radiolata – a possible candidate for defining the base of Cambrian Series 2 and Stage 3 // Bull. Museum Northern Arizona. 2011. № 67. P. 304–306.
Sdzuy K. The Precambrian–Cambrian boundary beds in Morocco (Preliminary Report) // Geol. Mag. 1978. V. 115. № 2. P. 83–94.
Shergold J. Explanatory notes for the Cambrian correlation chart // Treatise on Invertebrate Paleontology. Pt. O. Arthropoda 1. Trilobita. Revised. Lawrence: Univ. Kansas Press, 1997. P. 303–311.
Shergold J., Geyer G. The subcommission on Cambrian stratigraphy: the status quo // Geol. Acta. 2003. № 1. P. 5–9.
Steiner M., Weber B., Geyer G. The Lower Cambrian of Eastern Yunnan: trilobite-based biostratigraphy and related faunas // Acta Palaeontol. Sin. 2001. V. 40. Suppl. P. 63–79.
Steiner M., Li G., Qian Y. et al. Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China) // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007. V. 254. P. 67–99.
Sundberg F.A., Geyer G., Kruse P.D. et al. International correlation of the Cambrian Series 2–3, Stages 4–5 boundary interval // Mem. Assoc. Austral. Palaeontol. 2016. V. 49. P. 83–124.
Terfelt F., Eriksson M.E., Ahlberg P., Babcock L.E. Furongian Series (Cambrian) biostratigraphy of Scandinavia – a revision // Norw. J. Geol. 2008. V. 88. P. 73–87.
Topper T.P., Brock G.A., Skovsted C.B., Paterson J.R. Shelly fossils from the lower Cambrian Pararaia bunyerooensis Zone, Flinders Ranges, South Australia // Mem. Assoc. Austral. Palaeontol. 2009. V. 37. P. 199–246.
Yang A., Zhu M., Zhuravlev A.Yu. et al. Archaeocyathan zonation of the Yangtze Platform: implications for regional and global correlation of lower Cambrian stages // Geol. Mag. 2016. V. 153. № 3. P. 388–409.
Yochelson E.L., Kisselev G.N. Early Cambrian Salterella and Volborthella (Phylum Agmata) re-evaluated // Lethaia. 2003. V. 36. P. 8–20.
Yuan J., Zhu X., Lin J., Zhu M. Tentative correlation of Cambrian Series 2 between South China and other continents // Bull. Geosci. 2011. V. 86. № 3. P. 397–404.
Wang Y., Yin G., Zheng S., Qian Y. Stratigraphy of the Boundary Sinian–Cambrian in the Yangzi Area of Guizhou // The Upper Precambrian and Sinian–Cambrian Boundary in Guizhou. Guiyang: The Peoples’s Publ. House of Guizhou, 1984. P. 1–36.
Wei S., Fu Y., Liang H. et al. Re–Os geochronology of the Cambrian stage-2 and -3 boundary in Zhijin County, Guizhou Province, China // Acta Geochim. 2018. V. 37. № 2. P. 323–333.
Zhang W. World’s oldest Cambrian trilobites from eastern Yunnan // Stratigraphy and Palaeontology of Systemic Boundaries in China. Precambrian–Cambrian Boundary 1. Nanjing: Nanjing Univ. Publ. House, 1987. P. 1–18.
Zhang W. Cambrian biostratigraphy of China // Biostratigraphy of China. Eds. Zhang W., Chen P., Palmer A.R. Beijing: Science Press, 2003. P. 55–119.
Zhang X., Ahlberg P., Babcock L.E. et al. Challenges in defining the base of Cambrian Series 2 and Stage 3 // Earth-Sci. Rev. 2017. V. 172. P. 124–139.
Zhu M., Yang A., Yuan J. et al. Cambrian integrative stratigraphy and timescale of China // Sci. China Earth Sci. 2018. V. 61. P. 1–36.
Zhuravlev A.Yu. Preliminary suggestions on the global Early Cambrian zonation // Beringeria. 1995. Spec. № 2. P. 147–160.
Zhuravlev A.Yu., Gravestock D.I. Archaeocyaths from Yorke Peninsula, South Australia and archaeocyathan Early Cambrian zonation // Alcheringa. 1994. V. 18. P. 1–54.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция