

Растительные ресурсы, 2020, T. 56, № 2, стр. 138-150
Фитоценотическая характеристика, онтогенетическая структура и оценка состояния ценопопуляций Scutellaria scordiifolia (Lamiaceae) в Сибири
В. А. Черёмушкина 1, *, А. А. Гусева 1, Н. И. Макунина 1, А. Ю. Асташенков 1, Г. Р. Денисова 1
1 Центральный сибирский ботанический сад СО РАН
г. Новосибирск, Россия
* E-mail: cher.51@mail.ru
Поступила в редакцию 19.01.2020
После доработки 20.02.2020
Принята к публикации 18.03.2020
Аннотация
Дана эколого-фитоценотическая характеристика приуроченности клонального, явнополицентрического вида Scutellaria scordiifolia Fisch. ex Schrank в Азиатской России, выявлен потенциальный ареал вида в Алтае-Саянской горной области. Показано, что в сообществах луговых и настоящих степей вид встречается в небольшом обилии, очень редко заходит в остепненные травяные леса и на остепненные луга. В разных условиях обитания (остепненные луга, луговые и настоящие степи) изучено 8 ценопопуляций. Установлено, что все ценопопуляции нормальные, устойчивые, вегетативно неполночленные. Самоподдержание ценопопуляций происходит исключительно вегетативным путем. Экологическая плотность варьирует от 10.4 до 145 экземпляров на 1 м2. Максимальная плотность достигается в деградированных степных сообществах и в ценозах с большим обилием ветоши. Выявлено два типа онтогенетического спектра: левосторонний с максимумом на виргинильной или молодой генеративной группах и центрированный с максимумом на средневозрастной генеративной группе. Формирование различных спектров связано с особенностями вегетативного размножения, задернованностью почвы и антропогенной нагрузкой. Дана оценка состояние трех ценопопуляций, расположенных в контрастных условиях обитания. Установлено, что в условиях луговой и деградированной настоящей степях организменные и популяционные показатели максимальны или высокие, что позволяет считать эти условия оптимальными для вида. Остепненные луга менее благоприятны для произрастания S. scordiifolia, большинство показателей здесь имеют низкие значения.
Scutellaria scordiifolia Fischer ex Schrank (сем. Lamiaceae) шлемник скордиелистный – полиморфный вид с азиатским типом ареала [1]. Он распространен от юга Западной Сибири до Дальнего Востока. Северная граница ареала проходит по северу Алтае-Саянской горной области, южной части Забайкальского края, Дальнего Востока и Японии. Южная граница простирается по северной Монголии и северо-востоку Китая. Вид произрастает на степных и лугово-степных склонах, засоленных лугах, по берегам рек, в Китае встречается в широколиственных и сосновых лесах [2–4].
S. scordiifolia – многолетнее травянистое поликарпическое растение, образующее тонкие столоны и относящееся к клональным видам [5]. В естественных местообитаниях вид существует в виде рамет разного онтогенетического состояния. Вегетативное размножение преобладает, а семенное подавлено.
Как и многие виды сем. Lamiaceae, S. scordiifolia рассматривается как перспективное лекарственное растение, содержащее биологически активные вещества [6]. Лекарственная ценность вида определяется наличием широкого спектра природных соединений, в том числе эфирных масел с высоким содержанием монотерпеновых соединений, прежде всего, пулегона, изопулегона, изоментона, лимонена и скутеллареина [7–11]. Получаемые экстракты обладают антибактериальным [12] и противовоспалительным эффектами [13]. В тибетской медицине надземная часть растения используется как жаропонижающее средство [14]. Несмотря на широкое распространение этого вида на территории азиатской части России, его ресурсный потенциал практически не изучен. Недостаточно сведений о фитоценотической приуроченности, отсутствуют данные о структуре ценотических популяций. В связи с этим, целью настоящего исследования является оценка состояния ценопопуляций S. scordiifolia в естественных условиях обитания.
МАТЕРИАЛЫ И МЕТОДЫ
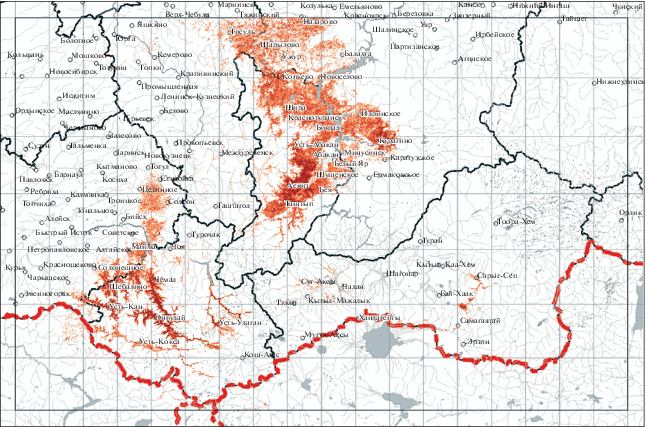
Фитоценотическая характеристика S. scordiifolia дана на основании анализа 430 описаний, сделанных Н.И. Макуниной, и литературных источников. Потенциальный ареал вида построен в программе MaxEnt [15], ячейка растра составляет 3 arc-sec (90 м). Результатом работы алгоритма является модель пригодности местообитаний – карта с прогнозными вероятностями присутствия сообществ в каждой ячейке растра. Карта построена на основе координат 430 описаний со S. scordiifolia. На рис. 1 красным окрашены ячейки, где вероятность присутствия вида превышает 0.5. Экологическая характеристика дана с использованием шкалы Л.Г. Раменского [16].
Рис. 1.
Потенциальный ареал Scutellaria scordiifolia. Красным окрашены ячейки, где вероятность присутствия вида превышает 0.5. Fig. 1. Potential distribution area of Scutellaria scordiifolia. Red indicates areas where the probability of the species presence exceeds 0.5.
Исследования онтогенетической структуры S. scordiifolia проводились в разных районах Сибири: Республиках Хакасия, Бурятия, Алтай и в Забайкальском крае. Всего изучено 8 ценопопуляций в разных местообитаниях (табл. 1).
Таблица 1.
Характеристика ценопопуляций (ЦП) Scutellaria scordiifolia Table 1. Characteristics of Scutellaria scordiifolia coenopopulations (CP)
| Номер ЦП Number of CP | Местонахождение ЦП Location of CP |
Координаты Coordinates |
Сообщество (Community): Доминирующие виды Dominant species |
ОПП, % GPC, % |
|---|---|---|---|---|
| 1 | Хакасия, Усть-Абаканский район,
окр. п. Красный Абакан Khakassia, Ust-Abakan district, Red Abakan village |
N 53о40′34.3″,
E 091o24′32.1″,
h = 245 м над ур. м. h = 245 m. a.s.l. |
Стоповидноосоковый остепненный луг (Carexpediformis steppe meadow): Carex pediformis C.A. Mey, Artemisia commutata Bess., Agropyron cristatum (L.) Beauv., Achillea millefolium L.
Сообщество подвержено выпасу Community is affected by grazing |
90 |
| 2 | Хакасия, Ширинский район, окр. д. Катюшкино,
река Сон Khakassia, Shirinsky district, Katyushkino village, the river Son |
N 54о16′36.2″,
E 090o14′35.9″,
h = 589 м над ур. м. h = 589 m. a.s.l. |
Злаковый остепненный долинный луг (Grass steppe valley meadow): Poa angustifolia L., Phleum phleoides (L.) Karst, Festuca valesiaca Gaudin s.l., Carex pediformis, Pulsatilla patens (L.) Mill. | 90 |
| 3 | Забайкальский край,
Чернышевский район,
окр. с. Улей. Zabaykalsky Krai, Chernyshevsky District, Ulej village. |
N 52о38′40.7″,
E 117o08′03.0″,
h = 594 м над ур. м. h = 594 m. a.s.l. |
Типчаковая луговая степь (Festuca meadow steppe): Festuca valesiaca Gaudin. s.l., Galium verum L., Elytrigia repens (L.) Nevski, Scutellaria scordiifolia Fischer ex Schrank, Phlomoides tuberosa (L.) Moench. | 85 |
| 4 | Республика Алтай,
Шебалинский р-н,
Ануйский хр., дол. р. Курзун Altai Republic, Shebalinskij district, Anujskij ridge, Kurzun river valley |
N 50о55′57.6″,
E 084o52′47.8″,
h = 1050 м над ур. м. h = 1050 m. a.s.l. |
Разнотравная луговая степь (Forb meadow steppe): Stipa capillata L., Achnaterum sibiricum (L.) Keng ex Tzvel., Carex pediformis, Helictotrichon altaica Tzvel., Iris ruthenica Ker-Gawler, Phlomoides tuberosa, Artemisia gmelinii Web. | 80 |
| 5 | Хакасия, Ширинский район, окр. п. Джирим Khakassia, Shirinsky district, Dzhirim village |
N 54о46′64.9″,
E 090o26′90.7″,
h = 453 м над ур. м. h = 453 m. a.s.l. |
Разнотравно-злаковая луговая степь (Forb-grass meadow steppe): Helictotrichon schellianum (Hack.) Kitag., Achnaterum sibiricum, Carex pediformis, Pulsatilla patens. | 95 |
| 6 | Хакасия, Аскизский район, окр. д. Кызлас Khakassia, Askiz district, Kyzlas village |
N 53o00′54.0″,
E 089o52′17.5″,
h = 569 м над ур. м. h = 569 m. a.s.l. |
Овсецово-ковыльная петрофитная луговая степь (Helictotrichon and Stipa meadow steppe): Stipa capillata, Helictotrichon altaicum, Achnaterum sibiricum, Carex pediformis, Iris ruthenica Ker-Gawler, Bupleurum scorzonerifolium Willd. | 90 |
| 7 | Хакасия, Боградский район, окр. с. Бородино. Khakassia, Bogradsky district, Borodino village. |
N 54о04′43.5″,
E 091o09′54.1″,
h = 392 м над ур. м. h = 392 m. a.s.l. |
Овсецово-ковыльная настоящая степь (Helictotrichon and Stipa steppe): Stipa capillata, Helictotrichon altaica, Festuca valesiaca s.l., Achnaterum sibiricum, Koeleria cristata (L.) Pers. s. str. | 85 |
| 8 | Республика Бурятия, Селенгинский район, надпойменная терраса р. Темник,
окр. п. Удунга. Republic of Buryatia, Selenginsky district, floodplain terrace of the river Temnik, Udunga village. |
N 51о08′65.9″,
E 105o56′76.6″,
h = 655 м над ур. м. h = 655 m. a.s.l. |
Разнотравно-житняковая настоящая степь (Forb and Agropyron steppe): Agropyron cristatum (L.) Baeuv., Cleistogenes squarrosa (Trin.) Keng, Allium ramosum L., Allium senescens L. s. str., Potentilla bifurca L. | 70 |
Онтогенетическая структура ценопопуляций исследована по общепринятым методикам [17]. Онтогенетические состояния особей S. scordiifolia выделяли на основе описанного ранее онтогенеза [5]. При изучении онтогенетической структуры за счетную единицу принималась рамета, представляющая собой парциальный побег или систему парциальных побегов. Онтогенетический спектр строили на основе учета особей не менее чем на 10–20 площадках размером 1 м2, заложенных регулярным способом на трансектах шириной 1 м. Тип ценопопуляций определяли по классификации О.В. Смирновой и Н.А. Тороповой [18] и классификации “дельта–омега” Л.А. Животовского [19]. Экологическую плотность устанавливали исходя из численности экземпляров на единицу обитаемого пространства [20]. Эффективная плотность популяции рассчитывали как произведение индекса эффективности (омега) на ее физическую плотность [19]. Состояние ценопопуляций оценивали согласно методике, разработанной Л.А. Заугольновой, суть которой заключается в выборе наиболее информативных организменных и популяционных признаков. Весь диапазон каждого признака организма и ценопопуляции разбивался на 5 классов с одинаковым объемом по равномерной шкале. Затем каждому классу присваивался балл; наименьший балл соответствовал худшему состоянию биосистемы. Результаты отображены в виде полигональных диаграмм [21, 22]. Их сопоставление позволяет оценивать особи и популяции одновременно по комплексу признаков. Для исследования выбраны 3 ценопопуляции, расположенные в контрастных условиях обитания. При сравнении значений организменных признаков использовали значения с достоверными отличиями по t-критерию Стьюдента (при 95%-ном уровне значимости). Измерение признаков производилось у 25 особей средневозрастного генеративного состояния. В качестве организменных признаков были выбраны: высота генеративного побега, число боковых побегов, длина столона I порядка, биомасса раметы; популяционных – экологическая плотность, эффективная плотность, число столонов I порядка, доля v–g1(%).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
S. scordiifolia в азиатской части России в естественных растительных сообществах содоминирует очень редко, в основном встречается единично. В Алтае-Саянской горной области он наиболее широко распространен в луговых степях гумидного и семигумидного секторов (рис. 1). В Северном и Центральном Алтае, Хакасии и прилежащей части Красноярского края вид встречается в лесостепном поясе и входит в состав травостоя луговых степей и их петрофитных вариантов, обнаружен также на остепненных лугах. В степном поясе S. scordiifolia обычен в дерновиннозлаковых степях и их петрофитных вариантах, очень редко заходит в остепненные травяные леса. В Туве его распространение ограничено лесостепным поясом: встречается в луговых степях (Центральная Тува) и в составе травостоя остепненных лесов (Западная Тува) [23]. В предгорных северных лесостепных районах (правобережье р. Оби, Кузнецкая лесостепь) вид практически отсутствует. В южной лесостепи (Бийско-Чумышская лесостепь) S. scordiifolia присутствует в луговых степях. В Забайкалье его произрастание также приурочено к степным районам (бассейны рек Ингода, Шилка, Онон, Аргунь). Он обычен в луговых степях: леймусовых (Leymus chinensis + Poa atteniata + Carex duriuscula), в леспедецево-нителистниковых (Lespedeza hedysaroides + Filifolium sibiricum), ерниково-лапчатково-полынных (Carex korshinskyi + + Carex pediformis + Potentilla acervata + Betula fusca), серобородниково-лапчатково-полынных (Spodiopogon sibiricus + Potentilla acervata + Carex korshinskyi + Carex pediformis) и володушково-полынно-валлисскотипчаковой (Bupleurum sibiricum + Artemisia commutata + Festuca valesiaca), байкальскоковыльных (Stipa baicalensis + Artemisia laciniata), реже присутствует в настоящих крыловоковыльных (Stipa. krylovii + Koeleria cristata + Bupleurum scorsonerifolium) степях [24]. В Якутии S. scordiifolia, помимо луговых степей, произрастает на остепненных и суходольных лугах [25]. По данным П.А. Гоголевой с соавт. [26], вид иногда встречается на засоленных лугах аласов Центральной Якутии (ассоциация Artemisio rupestris–Hordeetum). На Дальнем Востоке он отмечен в луговых сообществах [27].
Проведенный экологический анализ S. scordiifolia на основе геоботанических описаний для Алтае-Саянской горной области и прилежащей к ней части Западно-Сибирской равнины показал, что вид относится к ксеромезофитам и предпочитает в основном сухолуговое увлажнение, реже луговостепное, его диапазон находится в пределах 50–63 ступеней (оптимум 57.2 ± 2.59). В Якутии диапазон ближе к влажнолуговому, но значение оптимума сохраняется на уровне 61.9 ступени [28]. S. scordiifolia в пределах ареала произрастает на достаточно богатых почвах (черноземах и черноземно-луговых), реже встречается на малогумусных солонцеватых черноземах и каштановых почвах [29].
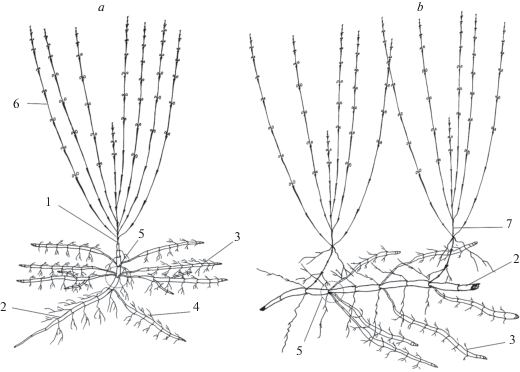
Во всех исследованных местообитаниях у особей S. scordiifolia формируется одна жизненная форма – столонообразующая. Для этого вида характерно преобладание вегетативного размножения, семенных особей в природе не обнаружено, что скорее всего связанно с тем, что у большинства клональных столонообразующих растений происходит стерилизация и утрата семенного размножения [30, 31]. Развитие особей разных онтогенетических состояний происходит по одной схеме. На столоне материнской раметы из боковых почек развиваются удлинённые гипогеогенные столоны I порядка со сближенными узлами в апикальной части. За счет накопления в междоузлиях питательных веществ они почти полностью утолщаются по всей длине (не утолщенными остаются лишь один-два метамера в базальной части) и напоминают вытянутый клубень, в пазухах чешуевидных листьев которого закладываются почки. В этот же год столоны I порядка могут ветвиться за счет раскрытия почек в базальной части клубневидного участка. Новые столоны II порядка удлиненные, полностью утолщенные. По всей длине столонов разного порядка в узлах развиваются придаточные корни. В конце вегетационного сезона неутолщенные метамеры столонов I порядка и междоузлия в базальной части столонов II порядка перегнивают, каждый столон с придаточной корневой системой продолжает существовать самостоятельно. На следующий год за счет вытягивания сближенных метамеров в апикальной части столона формируется апогеотропный участок, а из верхушечной почки развивается надземный ортотропный удлиненный годичный побег с укороченной базальной частью, состоящий из 2–5 сближенных метамеров с чешуевидными и 15–20 метамеров с зелеными листьями. Образуется дициклический побег, который ветвится. В надземной сфере из боковых почек развиваются вегетативные побеги обогащения и паракладии, в подземной сфере на столоне формируются дочерние столоны, дающие начало новому поколению рамет. Материнский столон в течение года постепенно отмирает с базальной части. Иногда верхушечная почка столона отмирает после перезимовки, в этом случае в рост трогаются боковые почки на плагиотропной части столона, дающие начало моноциклическим анизотропным генеративным побегами, образуется система парциальных побегов. Подобные структуры могут формироваться в средневозрастном и старом генеративном состоянии. В средневозрастном генеративном состоянии в одном или двух метамерах столона закладываются сериальные почки, что значительно увеличивает интенсивность вегетативного размножения. Таким образом, S. scordiifolia существует в виде парциальных дициклических побегов, в редких случаях в виде системы парциальных побегов (рис. 2).
Рис. 2.
Схема строения особей Scutellaria scordiifolia средневозрастного генеративного состояния. а – парциальный побег; b – система парциальных побегов; 1 – дициклический анизотропный генеративный побег; 2 – клубневидный столон n-го порядка; 3 – клубневидный столон n + 1-го порядка; 4 – придаточные корни; 5 – метамер с сериальными почками; 6 – паракладий; 7 – моноциклический генеративный побег. Fig. 2. The structure of a Scutellaria scordiifolia middle-aged reproductive individual. a – partial shoot; b – system of partial shoots; 1 – bicyclic anisotropic reproductive shoot; 2 – tuberoid n-th order stolon; 3 – tuberoid stolon of (n + 1)th order; 4 – secondary roots; 5 – metamer with accessory buds; 6 – paracladia; 7 – monocyclic reproductive shoot.
Изучение онтогенетической структуры ценопопуляций S. scordiifolia показало, что по классификации О.В. Смирновой и Н.А. Тороповой [18] они нормальные, вегетативно неполночленные. Самоподдержание ценопопуляций происходит исключительно вегетативным путем. Отсутствие ювенильных, имматурных, субсенильных и сенильных особей во всех ценопопуляциях определяется особенностями развития. У этого вида редко происходит семенное размножение, а онтогенез вегетативных особей (рамет) начинается с виргинильного состояния и заканчивается, как правило, после старого генеративного, изредка субсенильного состояния. Омоложение рамет происходит, как правило, на 1–2 онтогенетических состояния. Эти биологические особенности дают основание рассматривать левосторонний онтогенетический спектр с максимумом на виргинильных особях в качестве характерного для вида.
В изученных ценопопуляциях S. scordiifolia формируется два типа спектра: левосторонний и центрированный (рис. 3). Для большинства ценопопуляций установлен левосторонний одновершинный тип спектра. Абсолютный максимум приходится на виргинильные (ЦП 1, 2, 4, 5, 7), реже молодые генеративные особи (ЦП 6). Преобладание этих онтогенетических групп особей связано с особенностями вегетативного размножения в разных условиях обитания и антропогенной нагрузкой. Так, пик на виргинильных особях отмечен на остепненном лугу (ЦП 2), в луговой (ЦП 1, 4, 5) и настоящей (ЦП 7) степях. В ценопопуляциях 2, 4, 7 он определяется высокой степенью задернованности в сообществе, в ЦП 1 – выпасом. В этих условиях раскрытие почек на материнском столоне приводит к образованию рамет виргинильного состояния. Доля генеративных особей в ценопопуляциях с левосторонним спектром низкая и не превышает 41.6%. По классификации “дельта–омега” ЦП 1, 2, 4 относятся к молодым, ЦП 7 к зреющей (табл. 2). Экологическая плотность рамет в ценопопуляциях колеблется в больших пределах от 10.4 до 145 особей на м2. В большинстве ценопопуляций она составляет от 10 до 30 особей на м2. Присутствие в этих сообществах видов, приводящих к задернению почвы, и выпас сдерживают интенсивность вегетативного размножения, что отражается на относительно низкой численности вида. Наибольшая плотность отмечена в ЦП 5, где она достигает 145 особей на м2. Наличие в разнотравной луговой степи (ЦП 5) ветоши, создающей влажный микроклимат в приземном слое, способствует интенсивному ветвлению столонов до II порядка и раскрытию не только пазушных, но и сериальных почек на столонах средневозрастных особей. При этом, значительно увеличивается общее число рамет, доля генеративных растений и формируется двухвершинный спектр с абсолютным пиком на виргинильных и локальным на средневозрастных генеративных особях. Ценопопуляция по классификации “дельта–омега” – переходная.
Рис. 3.
Онтогенетические спектры ценопопуляций Scutellaria scordiifolia. По горизонтали – онтогенетические состояния; по вертикали – количество особей на единицу площади, %. Fig. 3. Ontogenetic spectra of Scutellaria scordiifolia coenopopulations. X-axis – ontogenetic states; y-axis – the number of individuals per unit area, %.
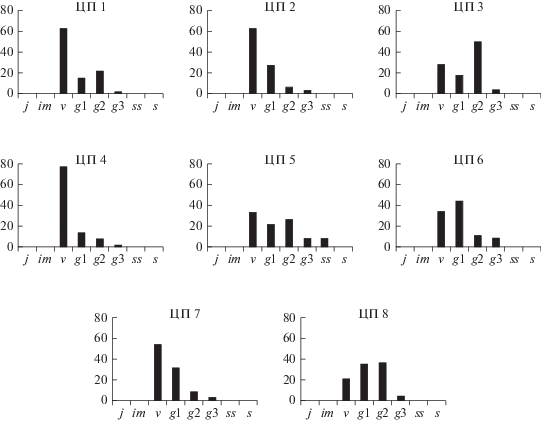
Таблица 2.
Демографические показатели ценопопуляций Scutellaria scordiifolia Table 2. Demographic parameters of Scutellaria scordiifolia coenopopulations (CP)
| № ЦП № CP |
Плотность, экз/м2 Density, ind/m2 |
ω | ∆ | Тип ЦП по классификации “дельта–омега” Type of CP by “Delta–omega” classification |
|---|---|---|---|---|
| 1 | 57.8 ± 20.3 | 0.59 | 0.23 | Молодая/young |
| 2 | 18.6 ± 3.7 | 0.57 | 0.2 | Молодая/young |
| 3 | 28.8 ± 8.5 | 0.79 | 0.36 | Зрелая/mature |
| 4 | 10.4 ± 1.6 | 0.51 | 0.17 | Молодая/young |
| 5 | 145 ± 32.6 | 0.69 | 0.37 | Переходная/transitional |
| 6 | 14.5 ± 1.8 | 0.68 | 0.28 | Зреющая/maturing |
| 7 | 14.8 ± 3.6 | 0.60 | 0.22 | Зреющая/maturing |
| 8 | 99.7 ± 28.2 | 0.78 | 0.35 | Зрелая/mature |
По соотношению онтогенетических групп спектр ЦП 6 также характеризуется значительной долей рамет виргинильного состояния (33.8%), но абсолютный максимум приходится на молодую генеративную фракцию (44.7%). Преобладание молодой генеративной фракции связано с уменьшением интенсивности вегетативного размножения виргинильных особей и образованием в молодом генеративном состоянии неомоложенных рамет. В условиях петрофитной луговой степи до 30% виргинильных особей не способны к образованию вегетативного потомства, так как они представлены моноциклическими анизотропными побегами со сниженной жизненностью. Ценопопуляция характеризуется как зреющая.
Центрированный онтогенетический спектр формируется в ЦП 3 в луговой степи и ЦП 8 в настоящей деградированной степи (рис. 3). В обеих ценопопуляциях на столонах средневозрастных генеративных растений закладываются не только пазушные, но и сериальные почки, раскрытие которых приводит к образованию рамет того же онтогенетического состояния и преобладанию их в ценопопуляции. В результате увеличения генеративных рамет в ценопопуляциях, по классификации “дельта–омега” они становятся зрелыми (табл. 2). Экологическая плотность особей в изученных ценопопуляциях отличается значительно. В ЦП 8 в условиях рыхлого субстрата (почва с песком и мелкой галькой) у особей отмечена высокая интенсивность вегетативного размножения, что отразилось на резком увеличении экологической плотности особей (99.6 экз/м2), по сравнению с ЦП 3 (28.8 экз/м2).
Изучение онтогенетической структуры 8 ценопопуляций S. scordiifolia в разных эколого-ценотических условиях показало динамичность распределения онтогенетических групп и в некоторых случаях несовпадение онтогенетических спектров с теоретическим характерным. Причина заключается в высокой морфологической пластичности и реагировании особей S. scordiifolia на изменение условий произрастания. Колебания влажности почвы и освещения, задернение и структура субстрата приводят к лабильности онтогенетических спектров – появление максимума на молодой или средневозрастной генеративной группе, а не на виргинильной. Такая ответная реакция связана с особенностями онтогенеза явнополицентрического вида S. scordiifolia и спецификой его вегетативного размножения.
Проведена комплексная оценка состояния трех ценопопуляций, изученных в контрастных местообитаниях: на остепненном долинном лугу (ЦП 2), в луговой (ЦП 5) и настоящей (ЦП 8) степях (рис. 4, табл. 3, 4).
Рис. 4.
Оценка состояния ценопопуляций Scutellaria scordiifolia (в баллах). Организменные признаки: 1 – высота генеративного побега, 2 – число боковых побегов, 3 – длина столона I-го порядка и 4 – биомасса раметы; популяционные признаки: 5 – экологическая плотность, 6 – эффективная плотность, 7 – число столонов I-го порядка и 8 – доля v–g1. 1–5 – баллы. Fig. 4. Assessment of the status of Scutellaria scordiifolia coenopopulations (in scores). Characters of organism: 1 – height of reproductive shoot, 2 – number of side shoots, 3 – length of 1st order stolon, 4 – biomass of a ramet; population characteristics: 5 – ecological density, 6 – effective density, 7 – number of 1st order stolons, 8 – proportion of v–g1. 1–5 – score.
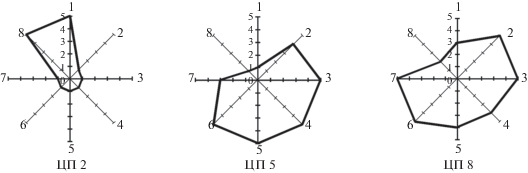
Таблица 3.
Организменные и популяционные показатели Scutellaria scordiifolia Table 3. Organismal and population characters of Scutellaria scordiifolia
| Признаки Character |
Ценопопуляция Coenopopulation |
||
|---|---|---|---|
| 2 | 5 | 8 | |
| Организменные параметры особей средневозрастного состояния (g2) Organismal characters of mature reproductive individual (g2) |
|||
| Высота генеративного побега, см Height of reproductive shoot, cm |
$\frac{{26.8 \pm 0.53{\text{*}}}}{{22{\kern 1pt} --{\kern 1pt} 32}}$ | $\frac{{12.2 \pm 0.64}}{{6{\kern 1pt} --{\kern 1pt} 17}}$ | $\frac{{17.6 \pm 0.59}}{{10{\kern 1pt} --{\kern 1pt} 25}}$ |
| Число боковых побегов, шт. Number of side shoots, pcs |
$\frac{{2 \pm 0.15}}{{1{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{5.3 \pm 0.52}}{{2{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.8 \pm 0.47}}{{4{\kern 1pt} --{\kern 1pt} 13}}$ |
| Длина столона I-го порядка, см Length of 1st order stolon, cm |
$\frac{{3.6 \pm 0.36}}{{2{\kern 1pt} --{\kern 1pt} 7}}$ | $\frac{{12 \pm 0.87}}{{5{\kern 1pt} --{\kern 1pt} 20}}$ | $\frac{{10.4 \pm 0.58}}{{6{\kern 1pt} --{\kern 1pt} 19}}$ |
| Биомасса раметы (сух), г Biomass of a ramet, g |
$\frac{{0.4 \pm 0.01}}{{0.3{\kern 1pt} --{\kern 1pt} 0.52}}$ | $\frac{{0.76{\text{ }}{\kern 1pt} \pm 0.02}}{{0.45{\kern 1pt} --{\kern 1pt} 1.02}}$ | $\frac{{0.65 \pm 0.03}}{{0.37{\kern 1pt} --{\kern 1pt} 0.99}}$ |
| Популяционные признаки Population characters |
|||
| Экологическая плотность, экз/м2 Ecological density, ind/m2 |
$\frac{{18.6 \pm 3.7}}{{2{\kern 1pt} --{\kern 1pt} 59}}$ | $\frac{{145 \pm 32.6}}{{63{\kern 1pt} --{\kern 1pt} 280}}$ | $\frac{{99.7 \pm 28.2}}{{57{\kern 1pt} --{\kern 1pt} 168}}$ |
| Эффективная плотность Effective density |
10.6 | 100 | 77.8 |
| Число столонов I-го порядка. шт Number of 1st order stolons, pcs |
$\frac{{3.4 \pm 0.23}}{{1{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.8 \pm 0.36}}{{2{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.3 \pm 0.78}}{{2{\kern 1pt} --{\kern 1pt} 2}}$ |
| Доля v–g1, % Proportion of v–g1, % |
90.3 | 56.1 | 57.5 |
Таблица 4.
Балловые оценки величины признаков Scutellaria scordiifolia Table 4. Character scores of Scutellaria scordiifolia
| Признаки Character |
Балл Score |
||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| Организменные параметры Organismal characters |
|||||
| Высота генеративного побега, см Height of reproductive shoot, cm |
<12.2 | 12.3–15.8 | 15.9–19.5 | 19.6–23.2 | 23.3–26.8 |
| Число боковых побегов, шт Number of side shoots, pcs |
<2 | 2.1–3.4 | 3.5–4.9 | 5–6.3 | 6.4–7.8 |
| Длина столона I-го порядка, см Length of 1st order stolon, cm |
<3.6 | 3.7–5.7 | 5.8–7.8 | 7.9–9.9 | 10–12 |
| Биомасса раметы (сух), г Biomass of a ramet, g |
<0.4 | 0.41–0.49 | 0.5–0.58 | 0.59–0.67 | 0.68–0.76 |
| Популяционные признаки Population characters |
|||||
| Экологическая плотность, экз/м2 Ecological density, ind/m2 |
18.6 | 18.7–50.2 | 50.3–81.8 | 81.9–113.4 | 113.5–145 |
| Эффективная плотность Effective density |
<10.6 | 10.7–32.9 | 33–55.3 | 55.4–77.7 | 77.8–100 |
| Число столонов I-го порядка, шт. Number of 1st order stolons, pcs |
<3.4 | 3.5–4.62 | 4.63–5.85 | 5.86–7.07 | 7.1–8.3 |
| Доля v–g1, % Proportion of v–g1, % |
<56.1 | 56.2–64.6 | 64.7–73.1 | 73.2–81.6 | 81.7–90.3 |
Оценка состояния ценопопуляций по организменным признакам показала, что высота растений с максимальными значениями (5 баллов) оказалась на остепненном долинном лугу (ЦП 5). Это связано с произрастанием особей в условиях достаточного освещения в сообществе с невысоким травостоем и высокой влажностью почвы. В ЦП 8, расположенной в настоящей разнотравно-житняковой степи, высота побега имеет среднее значение (3 балла). Хорошее развитие особей обеспечивается освещенностью (ОПП 70%) и меньшей задернованностью. Наименьшая высота (1 балл) установлена в луговой степи (ЦП 5), где сильная задернованность и наличие ветоши сдерживают рост особей S. scordiifolia.
В благоприятных условиях обитания, как правило, активизируется большое число почек в надземной и подземной частях растений, тем самым увеличивается биомасса всей особи. Накопление биомассы является одним из самых информативных показателей жизненности растения, который отражает оптимальные условия произрастания растений под действием биотических и абиотических факторов окружающей среды. В условиях луговых и настоящих степей у средневозрастных генеративных растений S. scordiifolia число надземных боковых побегов достигает максимальных величин в ЦП 8 и составляет в среднем 7.8 шт., несколько ниже этот показатель в ЦП 5 (5.5 шт.). Практически не отличаются величины длины столонов I порядка (12 см в ЦП 5 и 10.4 см в ЦП 8, значение tст = 1.53). Однако близость этих показателей обусловлена разными факторами: в луговой степи влажностью почвы, в настоящей степи слабой задернованностью и произрастанием на почве, содержащей песок и мелкую гальку. Эти условия позволяют боковому столону свободно развиваться. Значение двух последних показателей определяют биомассу особи: в ЦП 5 она максимальная (5 баллов), а в ЦП 8 высокая (4 балла). На остепненном лугу (ЦП 2), несмотря на большую высоту побегов (22–32 см), сильная задернованность и более плотная луговая почва приводят к резкому снижению числа боковых побегов и столонов I порядка у рамет, что отражается на уменьшении биомассы в 1.5–2 раза (1 балл).
Оценка ценопопуляций по популяционным признакам выявила максимальную и высокую экологическую плотность в ЦП 5 и ЦП 8 (5 и 4 балла соответственно). Такие значения признака связаны со средними и максимальными показателями числа столонов I порядка (4.8 и 8.3 шт. соответственно), а также заложением сериальных почек и развитием из них разветвленных репродуктивных побегов. Реализация всех почек отразилась на доле генеративных особей в этих ценопопуляциях и, как следствие, на увеличении значений эффективной плотности (77.8 и 100 соответственно). Низкие показатели экологической и эффективной плотности отмечены в ЦП 2 (по 1 баллу), где задернованность почвы снижает интенсивность образования вегетативного потомства. Таким образом, максимальные значения экологической и эффективной плотности отмечены в луговой степи, в настоящей степи показатель экологической плотности высокий, эффективной плотности максимальный, а на остепненном лугу оба показателя минимальны, они в 6–10 раз ниже (табл. 3). Изученные ценопопуляции также отличаются долей молодой фракции (v–g1). В ЦП 2 она составляет 90% (5 баллов), причем основной вклад вносят виргинильные особи, их преобладание определяется особенностями вегетативного размножения в условиях сильного задернения на остепненном лугу. Снижение и небольшая доля (1 и 2 балла) молодой группы особей в ЦП 5 и ЦП 8 связано с увеличением численности средневозрастных генеративных растений в более благоприятных условиях произрастания.
Диагностика состояний ценопопуляций S. scordiifolia показала, что из трех исследованных ценопопуляций две (ЦП 5 и ЦП 8) имеют близкие значения организменных (15 и 17 балла) и популяционных (14 и 16 баллов) признаков. Высокие показатели отмечены в ЦП 8, что, скорее всего, связано с ее нахождением в деградированном сообществе, где слабая конкуренция способствует формированию более мощных средневозрастных генеративных особей и увеличению их численности. Минимальные значения отмечены в ценопопуляции на остепненном лугу (ЦП 2), для которого характерно сильное задернение. Анализ изученных популяций S. scordiifolia в эколого-ценотическом ряду от остепненных лугов до настоящих степей по совокупности признаков показал, что в градиенте увлажнения, изученные ценопопуляции распределились от пессимального (остепненные луга) до оптимального (луговые и настоящие степях). В настоящих степях оптимальное состояние достигается только в сукцессионных (антропогенных) вариантах растительных сообществ.
Подобная динамичность спектров достаточно хорошо изучена для ряда явнополицентрических трав [32–35], у которых изменение разных онтогенетических групп связано с конкретным фитоценотическим окружением. На сегодняшний день имеются только отрывочные сведения по популяционной биологии некоторых столонообразующих растений для Европейской части России, среди которых изучены в основном лесные виды [33]. Так, исследованные ценопопуляции Trientalis europaea L. в трех фитоценозах национального парка “Марий Чодра” характеризовались как молодые, нормальные, неполночленные с различными значениями плотности особей. По онтогенетическому составу и способу самоподдержания популяций T. europaea совпадают с биологическим особенностями и популяционным поведением S. scordiifolia. Динамичность численности и состава онтогенетических групп T. europaea приходится на прегенеративную фракцию и связаны с особенностями фитоценоза, и рассматриваются автором в качестве флюктуаций. Подобная динамика отмечена нами для популяций S. scordiifolia с один отличием в том, что исследования S. scordiifolia проводили в различных участках его ареала. Анализ оригинальных и имеющихся данных по травянистым растениям позволяет предположить, что для столонообразующего S. scordiifolia характерен толерантный тип популяционной стратегии [32]. Устойчивость ценопопуляций большей степенью определяется интенсивностью омоложения рамет и ветвлением столонов, сопряженных с влиянием конкретных биотических и абиотиченских факторов.
ЗАКЛЮЧЕНИЕ
В результате исследования ценопопуляций в широтном направлении, охватывающем р-ны Сибири и Забайкальского края, установлено, что на остепненных лугах, в луговых и настоящих степях ценопопуляции S. scordiifolia нормальные, устойчивые, вегетативно неполночленные. Неполночленность ценопопуляций связано с особенностями вегетативного размножения, не глубоким омоложением рамет, быстрыми темпами старения и отмирания взрослых особей. В большинстве ценопопуляций выявлен одновершинный левосторонний тип спектра, который формируется в условиях умеренного увлажнения и высокой задернованностью почвы в луговых степях, центрированный – деградированной настоящей степи. Колебание численности от 10.4 до 145 экз./м2 отражает градиент увлажнения и влияние антропогенной нагрузки. Проведенный анализ распространения и эколого-фитоценотической приуроченности S. scordiifolia показал, что произрастание вида в Азиатской части России приурочено в основном к луговым и настоящим степям и их петрофитным вариантам, его оптимум находится в пределах 57.2 ± 2.59 ступеней. Вид относится к ксеромезофитам и предпочитает сухолуговое увлажнение, реже луговостепное. Преферентность S. scordiifolia к умеренному увлажнению отражается в комплексной оценке ценопопуляций на протяжении всего ареала. Выявлено, что в луговой степи и деградированной настоящей степи организменные и популяционные показатели максимальные и высокие, эти условия можно считать оптимальными для вида. Полученные результаты по биологии S. scordiifolia, онтогенетическому составу и основным демографическим показателям в пределах Азиатской части России характеризует его как вид с толерантным типом популяционной стратегии.
Список литературы
Пешкова Г.А. 2010. Даурская лесостепь. Барнаул. 144 с.
Грубов В.И. 1982. Определитель сосудистых растений Монголии. Л. 443 с.
Li Xi-wen, Ian C. Hedge. 1994. Flora of China. Beijing, China. V. 17. P. 75–103. http://flora.huh.harvard.edu/china/mss/volume17/Lamiaceae.published.pdf
Доронькин В.М. 2012. Семейство Lamiaceae Martinov, или Labiatae Juss. – В кн.: Конспект флоры Азиатской России: Сосудистые растения. Новосибирск. С. 413–429. http://www.csbg.nsc.ru/uploads/sistematic/Conspect_Flora.pdf
Guseva A. 2018. Features of development of the clonal plant Scutellaria scordiifolia Fischer ex Schrank (Lamiaceae) in Siberia. – Prospects of Development and Challenges of Modern Botany. 11. https://doi.org/10.1051/bioconf/20181100019
Дикорастущие полезные растения России. 2001. СПб. 663 с.
Чемесова И.И. 1993. Флавоноиды видов рода Scutellaria L. – Растительные ресурсы. 29(2): 89–99.
Malikov V.M., Yuldashev M.P. Penolic compounds of plant of the Scutellaria L. genus. Distribution, structure, and properties. 2002. – Chem Nat Compd. 38(4): 361–406. https://doi.org/10.1023/A:1021638411150
Дудетская Н.А., Теслов Л.С., Анизимова Н.А. 2010. Флавоноидный состав видов рода Scutellaria (Lamiaceae) флоры России. – Растительные ресурсы. 46(2): 159–174.
Каримов А.М., Ботиров Э.Х. 2016. Структурное разнообразие и степень изученности флавоноидов рода Scutellaria L. – Химия растительного сырья. 1: 5–28.
Olennikov D.N., Chirikova N.K. 2013. Phenolic compounds and cinnamamide from Scutellaria scordiifolia. – Chem Nat Compd. 49 (1): 124–126. https://doi.org/10.1007/s10600-013-0528-x
Keun-Dol Yook, Nayoung Ha. 2018. Effects of Scutellaria scordiifolia Fisch. ex Schrank exstracts on biofilm formation and the activities of Klebsiella pneumoniae. – Korean Journal of Clinical Laboratory Science. 50(4): 438–443. https://doi.org/10.15324/kjcls.2018.50.4.438
Sonoda M., Nishiyama T., Matsukawa Y., Moriyasua M. 2004. Cytotoxic activities of flavonoids from two Scutellaria plants in Chinese medicine. – J. Etnopharmacol. 91(1): 65–68. https://doi.org/10.1016/j.jep.2003.11.014
Тибетская медицина у бурят. Новосибирск. 2008. 323 с.
Phillips S.J., Anderson R.P., Schapire R.E. 2006. Maximum entropy modeling of species geographic distributions. – Ecol Model. 190(3–4): 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. 1956. Экологическая оценка кормовых угодий по растительному покрову. М. 472 с
Ценопопуляции растений (Основные понятия и структура). 1976. М. 217 с.
Смирнова О.В., Торопова Н.А. 2004. Общие представления популяционной биологии и экологии растений – В кн.: Восточно-европейские леса. Т. 1. М. С. 154–164. http://rjee.ru/wp-content/uploads/2016/03/Kniga_1.pdf
Животовский Л.А. 2001. Онтогенетические состояния, эффективная плотность и классификация популяций растений. – Экология. 1: 3–7.
Одум Ю. 1986. Экология. Т. 2. М. 376 с.
Заугольнова Л.Б., Денисова Л.В., Никитина С.В. 1993. Подходы к оценке состояния ценопопуляций растений. – Бюллетень Московского общества испытателей природы. Отдел биологический. – 98(5): 100–108. http://ashipunov.info/russian/journals/bmsn/ar-chive/moip_1993_098_5.djvu
Заугольнова Л.Б. 1994. Структура популяций семенных растений и проблемы их мониторинга: Дис. … д-ра. биол. наук в форме научного доклада. СПб. 70 с.
Макунина Н.И. 2016. Растительность лесостепи Западно-Сибирской равнины и Алтае-Саянской горной области. Новосибирск. 183 с. http://ashipunov.info/shipunov/school/books/makunina2016_rastit_lesost_zap_sib_ravn.djvu
Дулепова Б.И. 1993. Степи горной лесостепи Даурии и их динамика. Чита. 395 с.
Конспект флоры Якутии: Сосудистые растения. 2012. Новосибирск. 272 с.
Гоголева П.А., Миркин Б.М., Кононов К.Е., Миронова С.И. 1987. Синтаксономия и симфитосоциология растительности аласов Центральной Якутии. Иркутск. 176 с.
Флора российского Дальнего Востока. Дополнения и изменения к изданию “Сосудистые растения советского Дальнего Востока”. 2006. Владивосток. Т. 1–8. (1985–1996). 456 с.
Флора Якутии: Географический и экологический аспекты. 2010. Новосибирск. 192 с.
Куминова А.В. 1960. Растительный покров Алтая. Новосибирск. 449 с.
Eckert C.G. 2002. The loss of sex in clonal plants. – Evol Ecol. 15: 501–520. https://doi.org/10.1023/A:1016005519651
Herben T., Šerá B., Klimešová J. 2015. Clonal growth and sexual reproduction: tradeoffs and environmental constraints. – OIKOS. 124: 469–476. https://doi.org/10.1111/oik.01692
Смирнова О.В. 1987. Структура травяного покрова широколиственных лесов. М. 206 с.
Полянская Т.А. 2005. Седмичник европейский (Trientalis europaea L.) – В кн.: Биоразнообразие растений в экосистемах национального парка “Марий Чодра”. Йошкар-Ола. С. 132–137).
Водолазова С.В., Черемушкина В.А, Колегова Е.Б., Мяделец М.А. 2010. Онтогенез, структура ценопопуляций и эколого-ценотическая характеристика Nepeta sibirica (Lamiaceae) в Хакасии – Растительные ресурсы. 46(1): 3–18.
Черемушкина В.А., Басаргин Е.А. 2011. Структура ценопопуляций степных длиннокорневищных растений – В сб.: Отечественная геоботаника: основные вехи и перспективы: Матер. Всероссийской научной конференции. Т. 2. Структура и динамика растительных сообществ. Экология растительных сообществ. СПб. С. 495–496. https://docplayer.ru/55050727-Otechestvennaya-geobotanika-osnovnye-vehi-i-perspektivy.html
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы