

Почвоведение, 2023, № 2, стр. 184-202
Состав стабильных изотопов углерода органического вещества буроземов предгорий хребта Восточный Саян
В. А. Голубцов a, *, Ю. В. Вантеева a, М. А. Бронникова b, А. А. Черкашина a, Т. И. Знаменская a
a Институт географии им. В.Б. Сочавы СО РАН
664033 Иркутск, ул. Улан-Баторская, 1, Россия
b Институт географии РАН
119017 Москва, Старомонетный пер., 29, Россия
* E-mail: tea_88@inbox.ru
Поступила в редакцию 15.07.2022
После доработки 26.09.2022
Принята к публикации 07.10.2022
- EDN: BKDMWJ
- DOI: 10.31857/S0032180X22600937
Аннотация
Оценка вариабельности состава стабильных изотопов углерода органического вещества буроземов актуальна для понимания реакции лесных почв на изменения условий педогенеза. Исследования проведены в предгорьях северо-восточного макросклона хр. Восточный Саян. Впервые установлено формирование здесь буроземов, являющихся одними из наиболее изотопно-легких почв Байкальского региона, что в наибольшей степени связано с благоприятными условиями увлажнения для растительности, формирующей их органическое вещество (величины δ13С доминирующих видов варьируют от –32.6 до –27.8‰). Для органического вещества поверхностных горизонтов почв вариации значений δ13С от разреза к разрезу не превышают 1‰ (–25.81…–26.81‰), несмотря на существенные различия в характере поступления и последующей трансформации органического вещества. Отмечается значительный внутрипочвенный градиент значений δ13С (4–5‰). Анализ различий интенсивности оборота углерода, выраженной через наклон линейной регрессии (β) значений δ13С от содержания общего углерода в почве, изменяющихся с глубиной, показал, что изотопный профиль исследуемых буроземов складывается под влиянием микробной утилизации органического вещества, проявляющейся с разной интенсивностью в зависимости от строения органопрофиля. Повышенная интенсивность минерализации легкодоступных компонентов растительного материала в верхней части органопрофиля почв с развитой подстилкой и органо-аккумулятивными горизонтами может приводить к заметному росту значений δ13С и более выраженному β в отличие от почв с преобладанием минеральных горизонтов в профиле.
ВВЕДЕНИЕ
Органическое вещество почв является одним из ключевых компонентов глобального углеродного цикла [49]. Важную роль в этом процессе играют почвы бореальных лесов [28], в которых сосредоточено около 13% глобального органического углерода [40]. Органическое вещество таких почв характеризуется относительно низкой скоростью оборачиваемости из-за малой биологической доступности [58] и холодных климатических условий [63]. В то же время лесные почвы умеренного пояса могут стать одним из основных источников поступления углерода в атмосферу в ходе прогнозируемого потепления климата и роста интенсивности минерализации органического вещества [40].
Одними из наиболее широко распространенных почв в почвенном покрове лесов умеренного пояса являются буроземы [25, 29, 57], которые в международной системе почвенной классификации WRB [56] рассматриваются в рамках реферативной группы Cambisols. По мнению ряда исследователей в Байкальском регионе ареал буроземообразования ограничен темнохвойными ландшафтами северных циклонических склонов гор [34, 36], что отмечено на карте почвенно-экологического районирования [21]. В свете последних данных [4, 30, 35] их ареал можно оценивать в более широких географических рамках. Тем не менее, данные о распространении буроземов на обширных территориях северного макросклона хр. Восточный Саян в настоящий момент отсутствуют, хотя по ряду характеристик (прежде всего, распространению темнохвойных ландшафтов, количеству и характеру атмосферного увлажнения) эта территория может рассматриваться как благоприятная для буроземообразования.
Формирование органического вещества буроземов не обнаруживает каких-либо специфических механизмов, однако в значительной степени связано с характером поступающего опада [57] и гидротермическими условиями, определяющими характер его трансформации [29]. Поэтому исследования органопрофиля буроземов, формирующихся в различных ландшафтных условиях, могут способствовать углублению знаний об отклике данных почв на изменения условий среды. Анализ состава стабильных изотопов углерода (δ13С) представляет собой один из важных подходов к исследованию пространственно-временной вариабельности почвенного органического вещества (ПОВ) [48]. δ13С рассматривается в качестве интегрального показателя процессов трансформации ПОВ [46] и имеет значительный потенциал для оценки динамики почвенного углерода [42].
Настоящая работа посвящена оценке вариабельности состава стабильных изотопов углерода органического вещества буроземов предгорий северо-восточного макросклона хр. Восточный Саян в зависимости от характера их органопрофиля.
ОБЪЕКТЫ И МЕТОДЫ
Исследуемая территория расположена в предгорьях хр. Восточный Саян, вдоль его северо-восточного макросклона (рис. 1), в полосе предгорных ступеней и краевых прогибов между платформой и сводовым поднятием [14]. Рельеф эрозионно-тектонический, средне- и низкогорный. Речная сеть глубоко врезана и сложноветвящаяся, что связано в большей степени с густой сетью тектонических разрывов и смещений. Водоразделы северо-восточного, северо-западного и субмеридионального направлений в виде длинных гребней со скальными останцами.
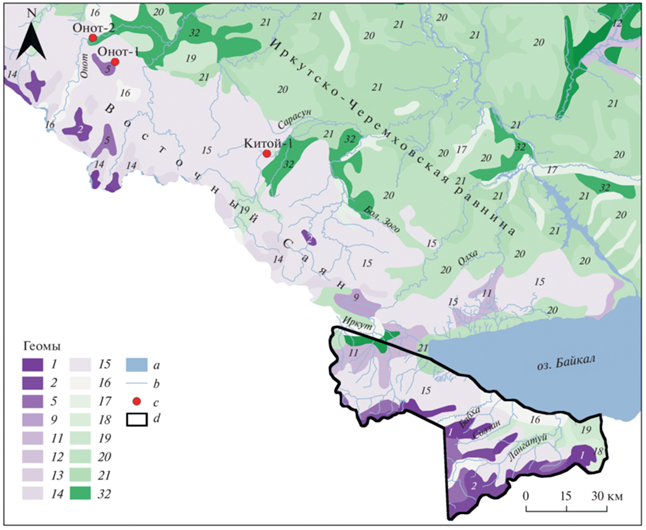
Рис. 1.
Территория исследования. Фрагмент карты “Ландшафты юга Восточной Сибири” [22] на уровне геомов: 1 – гольцовые альпинотипные; 2 – гольцовые тундровые; 5 – подгольцовые темнохвойно-редколесные; 9, 11 – горно-таежные лиственничные; 12 – подгорные и межгорных понижений лиственнично-таежные; 13 – подгорные подтаежные лиственничные; 14, 15, 18 – горно-таежные темнохвойные; 16, 19 – подгорные и межгорных понижений таежные темнохвойные; 17 – подгорные и межгорных понижений таежные кедрово-лиственничные; 20 – горно-таежные сосновые, 21 – подгорные подтаежные сосновые, 32 – подтаежные лиственничные и сосновые. Условные обозначения: a – крупные реки и озера, b – реки, с – расположение почвенных разрезов; d – границы распространения буроземов в качестве доминирующего типа почв в соответствии с почвенно-экологическим районированием Иркутской области [21].
Отрицательные морфоструктуры выполнены карбонатно-терригенными отложениями мотской серии венда [14], несогласно залегающими на различных возрастных уровнях раннедокембрийского фундамента. К последним относятся, в частности, стратифицированные образования савинской серии верхнего архея, слагающие Онотский грабен, расположенный непосредственно в краевой шовной зоне Сибирской платформы. Прогиб выполнен тальк-амфиболит-магнезитовым комплексом камчадальской толщи. В составе вышезалегающих свит – метавулканиты, амфиболиты, кварциты, в том числе железистые. Железистые кварциты представляют собой тонкие (4 мм) чередующиеся слойки кварца, магнетита и гематита, местами отмечаются мономинеральные гематитовые слойки [14]. Коренные породы, как правило, перекрываются маломощной толщей делювиальных легкосуглинистых и супесчаных отложений с многочисленными глыбистыми и щебнистыми включениями.
Климат резко континентальный с продолжительной суровой зимой и прохладным летом, в течение которого выпадает основная масса осадков. По мере увеличения высоты влажность климата возрастает, но в ряде случаев экспозиции склонов сглаживают влияние высоты [15]. Годовая сумма осадков при переходе от Иркутско-Черемховской равнины к предгорной части составляет около 400 мм. При переходе от предгорий к горной части (в районе с. Тальяны, Инга, Онот) годовое количество осадков возрастает до 550–600 мм. В горах Восточного Саяна годовая сумма осадков может достигать 700–900 мм с максимумом в июле–августе и минимумом в марте [26].
Среднегодовая температура воздуха изменяется от –1.4 до –3.9°С (по данным наблюдений метеостанции Дабады). Средние температуры января – от –21.5 до –23.4°С, а июля – от 15.1 до 16°С, самые низкие значения июльских и среднегодовых температур отмечаются в горной части территории. Сумма положительных среднесуточных температур воздуха более 10°С составляет 1400–1500°С. Продолжительность безморозного периода колеблется от 67 до 80 дней [26]. В среднегорьях и низкогорьях распространение многолетней мерзлоты имеет островной характер [3].
Абсолютные высоты от 700 до 1000 м в предгорьях северо-восточного макросклона хр. Восточный Саян занимает горно-таежный высотно-поясной комплекс светлохвойных лесов. Комплекс представлен сосняками и лиственничниками с преобладанием бореальных кустарничков (Vaccinium myrtillus L., V. Vitisidaea L.), таежного мелкотравья (Trientalis europaea L., Pyrola minor L., Mitella nuda L. и др.) и мхов (Pleurozium schreberi (Brid.) Mitt., Rhytidium rugosum (Hedw.) Warnst. и др.) в подчиненных ярусах [27]. Речные долины заняты сосновыми, лиственнично-сосновыми и лиственнично-березовыми травяными лесами.
На абсолютных высотах 900–1300 м господствует горно-таежный высотно-поясной комплекс кедровых (Pinus sibirica Du Tour) лесов. Преобладают кедровники зеленомошной группы, в древостое обычно в значительном количестве присутствуют пихта (Abies sibirica Lebed.) и ель (Picea obovata Lebed.) [27].
В соответствии с почвенно-экологическим районированием [21] исследуемая территория отнесена к среднегорному округу провинции подбуров, подзолов и буроземов Восточного Саяна. Тем не менее, буроземы в качестве господствующего типа почв выделяются лишь у юго-западной оконечности Байкала, тогда как под темнохвойными лесами Присаянья, по мнению Кузьмина [21], доминируют подзолы или подбуры.
Рассмотрим геоморфологическое положение и растительный покров площадок заложения исследуемых разрезов. Разрез Онот-1 (табл. 1, рис. 2а) расположен на высоте 1235 м на пологой приводораздельной поверхности между реками Онот и Мал. Белая. Почва формируется под молодым (60–80 лет) кедровым с примесью сосны и подростом пихты папоротниково (Dryopteris expansa)-мелкотравно (Oxalis acetosella L., Maianthemum bifolium)-кустарничково (Linnaea borealis L., Vaccinium vitis-idaea, V. myrtillus)-зеленомошным (Ptilium crista-castrensis (Hedw.) De Not., Pleurozium schreberi) лесом.
Таблица 1.
Исследуемые почвы и основные характеристики их местоположения
| Точка | Высота, м | Тип растительного сообщества | Вид-доминант | δ13C растительности, ‰ | Почва | Горизонт (глубина, см) | pH | Сорг, % | Nобщ, % | C : N | δ13C, ‰ | Сумма фракций (мм), % | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–0.05 | 0.05–0.001 | <0.001 | ||||||||||||
| Онот-1 | 1235 | Сосново-кедровый с пихтой в подросте папоротниково- мелкотравно-кустарничково-зеленомошный лес | Pinus sibirica Du Tour | –29.58 | Бурозем грубогумусированный эллювиированный глееватый поверхностно турбированный пирогенный (Folic Spodic Cambisol) | O (0–4) | 4.7 | 53.01 | 1.74 | 30.47 | –28.07 | – | – | – |
| Abies sibirica Ledeb. | –30.17 | Aao, pyr, tr (4–12) | 3.8 | 21.45 | 0.91 | 23.57 | –26.82 | 13.9 | 62.8 | 23.2 | ||||
| Dryopteris expansa (C. Presl) Fraser-Jenk. et Jermy | –27.79 | AYao, pyr, tr (12–15) | 3.8 | 12.80 | 0.56 | 22.86 | –25.81 | 18.9 | 58.1 | 23.0 | ||||
| Maianthemum bifolium (L.) F.W. Schmidt | –30.55 | AYao, el, pyr, tr (15–20) | 4.2 | 13.78 | 0.43 | 32.05 | –26.06 | 18.7 | 60.0 | 21.3 | ||||
| Vaccinium myrtillus L. | –32.60 | BMg, pyr (20–25) | 4.4 | 3.31 | 0.24 | 13.79 | –25.12 | 21.7 | 60.0 | 18.4 | ||||
| Pleurozium schreberi (Willd. ex Brid.) Mitt. | –32.54 | BMg, pyr (25–35) | 4.1 | 4.59 | 0.37 | 12.41 | –24.25 | 30.9 | 55.8 | 13.4 | ||||
| BMg (35–44) | 4.6 | 2.51 | 0.25 | 10.04 | –23.38 | 37.4 | 52.6 | 10.0 | ||||||
| BMg (44–50) | 4.7 | 1.59 | 0.18 | 8.83 | –22.90 | 22.8 | 66.3 | 10.9 | ||||||
| C (50–90) | 5.0 | 1.13 | 0.15 | 7.53 | –23.01 | 22.5 | 66.0 | 11.5 | ||||||
| Онот-2 | 1001 | Кедрово-сосново-осиновый с пихтой в подросте бруснично-черничный лес | Pinus sylvestris L. | –29.17 | Бурозем грубогумусированный пирогенный (Sceletic Cambisol) | O (0–3) | 5.6 | 51.80 | 1.43 | 36.22 | –29.11 | – | – | – |
| Aao,pyr (3–9) | 5.2 | 41.67 | 1.82 | 22.90 | –28.75 | – | – | – | ||||||
| Populus tremula L. | –29.99 | AYpyr (9–17) | 4.6 | 7.67 | 0.46 | 16.67 | –26.31 | 38.4 | 47.3 | 14.4 | ||||
| Pinus sibirica | –30.87 | BMpyr (17–20) | 4.8 | 2.63 | 0.21 | 12.52 | –25.62 | 41.7 | 45.0 | 13.2 | ||||
| Vaccinium myrtillus | –32.17 | BM (20–30) | 4.9 | 1.02 | 0.11 | 9.27 | –24.98 | 37.5 | 54.1 | 8.4 | ||||
| C (30–35) | 5.0 | 0.89 | 0.09 | 9.89 | –24.89 | 48.0 | 41.7 | 10.2 | ||||||
| C (35–45) | 5.0 | 0.53 | 0.05 | 10.60 | –24.61 | 44.7 | 47.8 | 7.5 | ||||||
| C (45–55) | 5.3 | 0.37 | 0.04 | 9.25 | –23.70 | 30.0 | 54.8 | 15.1 | ||||||
| C (55–65) | 5.2 | 0.48 | 0.06 | 8.00 | –23.63 | 36.5 | 54.9 | 8.6 | ||||||
| Китой-1 | 740 | Лиственнично-сосновый примесью березы и душекии злаково-мелкотравно-брусничный лес | Pinus sylvestris | –29.80 | Бурозем грубогумусированный пирогенный (Folic Cambisol) | O (0–4) | 4.9 | 49.77 | 0.90 | 55.30 | –27.90 | – | – | – |
| Larix sibirica Ledeb. | –29.60 | Aao (4–7) | 4.9 | 16.22 | 0.75 | 21.63 | –26.81 | 49.2 | 39.3 | 11.5 | ||||
| Betula pubescens Ehrh. | –31.03 | AYao, pyr (7–13) | 4.8 | 10.86 | 0.65 | 16.71 | –26.34 | 55.9 | 34.1 | 10.0 | ||||
| Duschekia fruticose (Rupr.) Pouzar | –30.15 | AYpyr (13–18) | 4.8 | 5.00 | 0.29 | 17.24 | –25.60 | 48.2 | 41.4 | 10.4 | ||||
| Vaccinium vitis-idaea L. | –31.84 | BMpyr (18–20) | 5.0 | 4.40 | 0.27 | 16.30 | –25.33 | 50.5 | 38.4 | 11.1 | ||||
| BMpyr (20–25) | 5.0 | 2.98 | 0.19 | 15.68 | –24.93 | 54.2 | 37.2 | 8.5 | ||||||
| BMpyr (25–30) | 5.3 | 1.90 | 0.13 | 14.62 | –24.53 | 58.6 | 32.7 | 8.7 | ||||||
| C (30–45) | 5.4 | 1.05 | 0.08 | 13.13 | –23.89 | 67.2 | 26.1 | 6.7 | ||||||
Рис. 2.
Общая морфология исследуемых почв (а), их температурные режимы по сравнению с температурными режимами почв в предгорьях Восточного Саяна (Китой (серая)) и на территории Иркутско-Черемховской равнины (Н. Булай и Осиновый (серые метаморфические)) (b). Цифры на линиях – температура, °С. Интенсивность выделения диоксида углерода из почв на площадках Онот-1 и Онот-2 (c).

Разрез Онот-2 заложен на высоте 1001 м в пределах выположенной приводораздельной поверхности между р. Елахой и Онот. Значительные площади занимают производные леса (на разных стадиях сукцессии), возникшие преимущественно вследствие пожаров. На исследуемой площадке сукцессия представлена старовозрастным сосново-осиновым бруснично-черничным лесом, под пологом которого во втором ярусе развивается молодой кедровник с пихтой в подросте.
Разрез Китой-1 заложен на высоте 720 м на покатом западном склоне ложбины, примыкающей к долине р. Китой. В бассейне р. Китой на высотах 700–900 м наблюдается переходная зона от горнотаежных темнохвойных геосистем к таежным светлохвойным и подгорным подтаежным сосновым. В месте заложения разреза светлохвойная тайга представлена лиственнично-сосновым с примесью березы, осины и душекии злаково (Brachypodium pinnatum, Calamagrostis spp.)-мелкотравно (Maianthemum bifolium)-брусничным лесом с признаками антропогенного нарушения (зарастающая старая вырубка) и следами низового пожара.
В 2020 г. площадки Онот-1 и Онот-2 были оборудованы атмосферно-почвенными измерительными комплексами [9] для измерения температурного режима почв и логгерами RC-51H для измерения температуры воздуха. Последние устанавливались на деревьях с северной стороны ствола на высоте 2 м. Точность измерения температуры 0.1°C. Измерения проводятся с периодичностью 1 ч.
На основе анализа описаний видового состава растительности производили отбор надземных частей (свежих зрелых листьев) доминирующих видов, составляющих 80% биомассы на исследуемых площадках (размер площадок 15 × 15 м). Всего на площадках для исследования изотопного состава растительности отобрано 14 видов сосудистых растений и один вид мха. Растительный опад на поверхности почвы отбирали на каждой площадке в пяти местах (четыре угла и центральная точка). Определение активности выделения СО2 из почвы проводили в полевых условиях по [18]. Образцы листьев, подстилки и почв высушивали до воздушно-сухого состояния. Листья дополнительно просушивали при 70°С в течение 60 ч, затем растирали. Образцы минеральных горизонтов почв просеивали через сито диаметром 1 мм с последующим удалением корней. Значения pH почвенных суспензий определяли потенциометрически. Определение гранулометрического состава выполняли по средней пробе в стоячей воде пирофосфатным методом. Содержание органического углерода и азота измеряли на CHN-анализаторе Vario Isotope (Elementar, Германия) методом поточной масс-спектрометрии после пиролиза пробы при температуре 1500°С. Состав стабильных изотопов углерода (δ13С) листьев, опада и минеральных горизонтов почв определяли на масс-спектрометре Isoprime precision IRMS (Elementar, UK). Измерения проводили в ЦКП “Лаборатория радиоуглеродного датирования и электронной микроскопии” Института географии РАН. Полученные результаты выражали в ‰ по отношению к стандарту VPDB. Точность измерений 0.1‰. Микроморфологические исследования выполняли в тонких шлифах с использованием поляризационного микроскопа Nikon Eclipse E200. Типизация почвообразовательных процессов дана по [16].
Статистическую обработку данных осуществляли в программных пакетах PAST 4.03 [55] и MO Excel. Рассчитывали наклон линейной регрессии (β), отражающей степень изменения δ13С от изменения содержания органического углерода с глубиной [39]. Для выявления различий в изотопном составе углерода растительности в зависимости от породы деревьев (хвойные/лиственные) и в зависимости от яруса из-за малого объема выборки применяли непараметрические методы: критерий Манна–Уитни и дисперсионный анализ (тест Краскела–Уоллиса и критерий Данна).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Гидротермические условия почвообразования. Данные микроклиматического мониторинга, проводимого на территории предгорий хр. Восточный Саян и Иркутско-Черемховской равнины, свидетельствуют о меньшей контрастности температурного режима исследуемых площадок в предгорьях по сравнению с почвами равнинной части (рис. 2b). Это явление может быть связано с меньшим прогревом в летние месяцы и меньшим выхолаживанием за счет инверсионного распределения температуры зимой, а также наличием плотного древесного яруса и большей мощностью снежного покрова в предгорьях. Низкая контрастность гидротермических режимов служит одним из основных факторов, благоприятствующих буроземообразованию [34, 36].
Для точки Онот-1, верхней в высотном ряду, отмечается отрицательная средняя годовая температура воздуха (–0.8°С), тогда как в точке Онот-2, расположенной ниже, она составляет 0.7°С. Измерения температуры поверхности почв свидетельствуют о несколько меньшей продолжительности периода с отрицательными температурами в точке Онот-2 (153 дня), нежели чем в точке Онот-1 (179 дней). Сумма положительных температур составляет 1494 и 1283°С для точек Онот-2 и Онот-1, соответственно. В целом исследуемые почвы подвержены кратковременному незначительному промерзанию в верхней толще. Отрицательные температуры в профиле проявляются в январе–феврале. При этом в точке Онот-1 промерзание немного более продолжительное. Хотя на данный момент климатические данные по точке Китой-1 отсутствуют, можно предположить наиболее благоприятный температурный режим и отсутствие промерзания в данной почве, учитывая более низкую абсолютную высоту местности.
Различия в увлажнении местоположений оценивали косвенно по экологической шкале увлажнения почв [37] на основе анализа видового состава растительности [20, 33, 38]. В древесном ярусе на всех площадках представлены виды с широкой экологической амплитудой. Основные различия по экологическим группам прослеживаются в травянисто-кустарничковом ярусе. Расчет средних значений середины амплитуд (табл. 2) показал, что растительные сообщества на площадках Онот-1 и Онот-2 более однородны и относятся к сыровато-лесолуговой экологической группе. На Китое-1 наблюдается более широкий разброс видов по увлажнению.
Таблица 2.
Показатели увлажнения видов на исследуемых площадках
| Вид | ПП, % | Увлажнение почвы, баллы по шкале Цыганова (1983) | Середина амплитуд | Взвешенное бальное значение | |
|---|---|---|---|---|---|
| min | max | ||||
| Онот-1 | |||||
| Pinus sibirica | 40 | 10 | 19 | 14.5 | 580 |
| Pinus sylvestris | 10 | 8 | 20 | 14 | 140 |
| Abies sibirica | 25.5 | 10 | 16 | 13 | 331.5 |
| Sorbus sibirica | 5.5 | 10 | 16 | 13 | 71.5 |
| Lonicera pallasii | 0.5 | 11 | 15 | 13 | 6.5 |
| Vaccinium myrtillus | 20 | 10 | 19 | 14.5 | 290 |
| Maianthemum bifolium | 15 | 11 | 15 | 13 | 195 |
| Linnaea borealis | 5 | 11 | 17 | 14 | 70 |
| Oxalis acetosella | 5 | 10 | 16 | 13 | 65 |
| Vaccinium vitis – idaea | 5 | 10 | 17 | 13.5 | 67.5 |
| Carex iljinii | 3 | – * | – | – | – |
| Trientalis europaea | 1 | 10 | 19 | 14.5 | 14.5 |
| Lycopodium clavatum | 4 | 9 | 15 | 12 | 48 |
| Dryopteris expansa | 10 | – | – | – | – |
| Diphasiastrum complanatum | 0.5 | 9 | 15 | 12 | 6 |
| Lycopodium annotinum | 5 | 9 | 16 | 12.5 | 62.5 |
| Pleurozium schreberi | 55 | 9 | 17 | 13 | 715 |
| Polytrichum commune | 5 | 11 | 19 | 15 | 75 |
| Ptilium crista – castrensis | 20 | 9 | 15 | 12 | 240 |
| Hylocomium splendens | 5 | 11 | 18 | 14.5 | 72.5 |
| Sphagnum sp. | 3 | – | – | – | – |
| Dicranum polysetum | 1 | 8 | 15 | 11.5 | 11.5 |
| Gymnocarpium dryopteris | 0.1 | 11 | 17 | 14 | 1.4 |
| Ledum palustre | 0.1 | 11 | 19 | 15 | 1.5 |
| Среднее (статус описания) | 13.4 | 145.95 | |||
| Онот-2 | |||||
| Populus tremula | 35 | 9 | 19 | 14 | 490 |
| Pinus sylvestris | 15 | 8 | 20 | 14 | 210 |
| Pinus sibirica | 13.5 | 10 | 19 | 14.5 | 195.75 |
| Abies sibirica | 10.5 | 10 | 16 | 13 | 136.5 |
| Sorbus sibirica | 1.5 | 10 | 16 | 13 | 19.5 |
| Rhododendron aureum | 3 | – | – | – | – |
| Vaccinium myrtillus | 60 | 10 | 19 | 14.5 | 870 |
| Gymnocarpium dryopteris | 2 | 11 | 17 | 14 | 28 |
| Diphasiastrum complanatum | 3 | 9 | 15 | 12 | 36 |
| Trientalis europaea | 0.5 | 10 | 19 | 14.5 | 7.25 |
| Maianthemum bifolium | 1 | 11 | 15 | 13 | 13 |
| Vaccinium vitis – idaea | 10 | 10 | 17 | 13.5 | 135 |
| Calamagrostis langsdorffii | 4 | – | – | – | – |
| Linnaea borealis | 2 | 11 | 17 | 14 | 28 |
| Calamagrostis arundinacea | 7 | 7 | 19 | 13 | 91 |
| Lonicera pallasii | 0.5 | 11 | 15 | 13 | 6.5 |
| Dryopteris expansa | 0.1 | – | – | – | – |
| Goodyera repens | 0.1 | 11 | 15 | 13 | 1.3 |
| Pleurozium schreberi | 1 | 9 | 17 | 13 | 13 |
| Ptilium crista – castrensis | 1 | 9 | 15 | 12 | 12 |
| Hylocomium splendens | 1 | 11 | 18 | 14.5 | 14.5 |
| Среднее (статус описания) | 13.5 | 128.2 | |||
| Китой-1 | |||||
| Pinus sylvestris | 20.1 | 8 | 20 | 14 | 281.4 |
| Larix sibirica | 20 | 9 | 16 | 12.5 | 250 |
| Betula pubescens | 7.5 | 10 | 20 | 15 | 112.5 |
| Populus tremula | 3 | 9 | 19 | 14 | 42 |
| Duschekia fruticosa | 20 | 11 | 17 | 14 | 280 |
| Sorbus sibirica | 2.5 | 10 | 16 | 13 | 32.5 |
| Rosa acicularis | 0.5 | 7 | 17 | 12 | 6 |
| Vaccinium vitis – idaea | 30 | 10 | 17 | 13.5 | 405 |
| Sanguisorba officinalis | 0.5 | 7 | 19 | 13 | 6.5 |
| Maianthemum bifolium | 10 | 11 | 15 | 13 | 130 |
| Trientalis europaea | 1 | 10 | 19 | 14.5 | 14.5 |
| Linnaea borealis | 0.5 | 11 | 17 | 14 | 7 |
| Lathyrus humilis | 0.5 | – | – | – | – |
| Rubus saxatilis | 5 | 9 | 19 | 14 | 70 |
| Thalictrum minus | 0.5 | 5 | 15 | 10 | 5 |
| Irirs ruthenica | 0.5 | – | – | – | – |
| Galium boreale | 2 | 7 | 15 | 11 | 22 |
| Vicia baicalensis | 0.1 | – | – | – | – |
| Pteridium pinetorum | 0.5 | 9 | 17 | 13 | 6.5 |
| Atragene sibirica | 0.1 | 9 | 15 | 12 | 1.2 |
| Viola uniflora | 0.5 | 11 | 14 | 12.5 | 6.25 |
| Pinus sibirica | 0.5 | 10 | 19 | 14.5 | 7.25 |
| Abies sibirica | 0.5 | 10 | 16 | 13 | 6.5 |
| Rhododendron dauricum | 2 | – | – | – | – |
| Vaccinium myrtillus | 0.5 | 10 | 19 | 14.5 | 7.25 |
| Veratrum lobelianum | 0.1 | 9 | 19 | 14 | 1.4 |
| Aegopodium alpestre | 0.1 | – | – | – | – |
| Anthriscus sylvestris | 0.5 | 9 | 15 | 12 | 6 |
| Gymnocarpium dryopteris | 0.5 | 11 | 17 | 14 | 7 |
| Equisetum pratense | 0.1 | – | – | – | – |
| Trollius asiaticus | 0.1 | – | – | – | – |
| Anemone crinita | 0.1 | – | – | – | – |
| Brachypodium pinnatum | 20 | 5 | 15 | 10 | 200 |
| Calamagrostis obtusata | 10 | 11 | 15 | 13 | 130 |
| Calamagrostis pavlovii | 10 | – | – | – | – |
| Pleurozium schreberi | 5 | 9 | 17 | 13 | 65 |
| Ptilium crista–castrensis | 5 | 9 | 15 | 12 | 60 |
| Среднее (статус описания) | 13.0 | 77.45 | |||
Расчет взвешенных бальных показателей с учетом проективного покрытия видов показал, что наибольшее увлажнение почв характерно для Онота-1, наименьшее – для площадки Китой-1. Здесь наблюдается значительная доля растений, середина амплитуд которых соответствуют суб-, сухо- и свежелесолуговой группам, доминантом является Brachypodium pinnatum (сублесолуговая группа). На Оноте-1 и Оноте-2 виды последней группы не представлены. Такие различия связаны с особенностями местоположения площадок. Китой-1 располагается на более низких гипсометрических отметках (740 м), где на исследуемой территории происходит смена горнотаежных темнохвойных геосистем светлохвойными, обусловленная, в частности, климатическими изменениями (уменьшением количества осадков и повышением температур). Свою роль играет более легкий гранулометрический состав бурозема в точке Китой-1, что создает лучшие условия для дренажа. Важными факторами являются пожар и рубка на Китое-1, которые оказали влияние на естественный микроклимат. Разреженность древесных крон привела к увеличению освещенности, лучшему прогреву почвы и более высокой испаряемости, что способствовало появлению более светолюбивых и приспособленных к умеренному увлажнению видов.
Описанные различия потенциально могут оказывать значительное влияние на характер трансформации растительных остатков.
Морфология и физико-химические свойства почв. Исследуемые почвы отнесены к типу буроземов. Их объединяет наличие буроокрашенных структурно-метаморфических диагностических горизонтов и в различной степени развитых серогумусовых горизонтов (рис. 2а). В органопрофиле всех трех почв между подстилкой и серогумусовым горизонтом залегает малый горизонт Аaо мощностью менее 10 см, представляющий собой материал, состоящий из смеси растительных остатков с минеральным суглинистым материалом. В разрезах Онот-1 и Китой-1 серогумусовые горизонты также содержат включения грубогумусового материала. Наличие малых горизонтов Ааo и примеси грубогумусового материала в серогумусовых горизонтах позволяют отнести все три профиля к грубогумусированному подтипу. Из важных различий органопрофиля трех исследованных почв следует отметить снижение совокупной мощности горизонтов опада, подстилки и грубогумусированного материала с уменьшением абсолютной высоты в ряду Онот-1 – Онот-2 – Китой-1.
Серогумусовые горизонты всех трех почв морфологически сходны: рыхлые, буровато-серые с неравномерной прокраской дисперсным органически веществом, непрочной мелкокомковатой структурой. Примесь грубогумусового материала в части серогумусовых горизонтов (горизонт AYao в профилях Онот-1 и Китой-1) отражается в более высоком, чем характерно для серогумусового горизонта, содержании органического углерода (7.67 и 10.86% соответственно). Во всех трех разрезах до глубины 20–35 см встречаются угли, что дает основания отнести все три профиля к пирогенному подтипу. При этом отмечается больший объем пирогенного материала в разрезах Онот-2 и Китой-1.
Вместе с тем в исследованных почвах наблюдаются следующие различия подтипового уровня. Органо-аккумулятивные горизонты профиля Онот-1 имеют признаки турбационных нарушений, вероятно, связанных с пирогенезом и/или ветровалами: прерывания сплошности горизонтов, интрузии материала нижележащих минеральных горизонтов. Это дает основание присвоить почве Онот-1 подтиповое название поверхностно-турбированной. В этом профиле горизонт AY в нижней части (15–20 см) имеет неясные седоватые пятна оподзоливания, что позволяет добавить к почве Онот-1 подтиповое название элювиированная.
Различия срединных частей почвенных профилей менее существенны. Так, наиболее интенсивно окрашенный в бурые тона и мощный горизонт BM характерен для разреза Онот-1, тогда как в значительно меньшей степени эти горизонты выражены в разрезах Онот-2 и Китой-1. Во всех исследуемых почвах кровля горизонта BM располагается на глубинах 17–20 см.
Микростроение изучали для двух из трех исследованных почв (Онот-1 и Онот-2). В целом оно сходно и в основных чертах соответствует характерному микростроению, ранее описанному для буроземов различных регионов [10]. Для диагностического структурно-метаморфического горизонта характерны проявления оксидогенеза в виде ожелезнения зерен скелета в результате процессов выветривания (рис. 3а–3c, 3e), накопления окислов-гидроокислов железа в составе тонкодисперсной части общей массы: железисто-глинистый, гумусово-железисто-глинистый состав плазмы и мелких железистых нодулей с резко очерченными границами (рис. 3d, 3e). Эти горизонты имеют хорошо развитую коагуляционную микроструктуру. Микроагрегаты одного, или двух порядков, округлые, иногда собранные в линзовидные агрегаты второго порядка (рис. 3e–3g). Суровые климатические условия определяют развитие криогенных признаков, таких как компактное внутреннее сложение и резкие границы агрегатов, гранулярный и линзовидный мотивы в структуре, элементы вокругагрегатной и вокругскелетной ориентации плазмы (рис. 3b, 3g), сортировка скелета, приводящая к аккумуляции пылеватого материала в межагрегатном пространстве (рис 3e, 3h).
Рис. 3.
Микростроение гор. ВМ; а, c–f, h – ник. ||, b, g – ник. Х; a–c, e: ожелезнение зерен скелета; железисто-глинистый, гумусово-железисто-глинистый состав плазмы; d, e – железистые нодули; d–h – коагуляционная микроструктура: округлые, овальные, линзовидные агрегаты; b, e, g – вокругскелетная и вокругагрегатная ориентации плазмы; c, h – несвязный пылеватый материал в межагрегатном пространстве.

Ключевое отличие микростроения буроземов в разрезах Онот-1 и Онот-2, особенно важное с точки зрения фракционирования изотопов углерода и формирования изотопного состава и его изменений по профилю, заключается в особенностях серогумусовых горизонтов, связанных со степенью минерализации их органического вещества. В профиле Онот-1 видим грубогумусированный серогумусовый горизонт, основной чертой микростроения которого является богатство растительными остатками, в различной степени трансформированными микробиотой и почвенной фауной. Здесь также встречаются углистые частицы. Для этого горизонта характерны мелкая копрогенная структура, обязанная деятельности микрофауны, свежие выбросы фауны, а также тонколистоватое сложение (рис. 4а, 4b), связанное либо с первичной микрослоистой текстурой накапливающегося опада, либо с криогенезом, либо с совокупным влиянием обоих факторов. В серогумусовом горизонте органо-аккумулятивной части профиля Онот-2 органическое вещество существенно сильнее гумифицировано. Здесь преобладает сгустковый, пропиточный гумус. Доля органического вещества значительно меньше, чем в грубогумусированных горизонтах AYao. Для горизонта AY характерны губчатое сложение, обилие биогенных каналовидных пор и пор-камер, выбросы мезо- и микрофауны, более крупная копрогенная и копрогенно-коагуляционная структура двух–трех порядков: округлые агрегаты формируют более крупные округлые и призмовидные (рис. 4c, 4d). В обоих случаях в гумусовых горизонтах обнаруживается много мелких железистых нодулей (рис. 4a–4d).
Рис. 4.
Микростроение гумусовых и органогенных горизонтов, ник. || а, b: гор. Аao, растительные остатки, углистые частицы; копрогенная структура, тонколистоватое сложение, железистые нодули; c, d: гор. АY, пропиточный гумус, губчатое сложение, копрогенная и копрогенно-коагуляционная структура, биогенные поры, выбросы мезо- и микрофауны, железистые нодули.

Основываясь на выполненных морфогенетических исследованиях, можно заключить, что ведущими почвообразовательными процессами в исследуемых буроземах являются процессы метаморфизации минерального вещества и переорганизации почвенной массы: разрушения и трансформации силикатов с накоплением оксидов железа, биогенного, коагуляционного и криогенного оструктуривания, криогенной сортировки, процессы внутригоризонтного перераспределения оксидов железа, в том числе окислительной сегрегации. При этом не наблюдается никаких микроморфологических признаков подвижности ПОВ: гумусовых, глинисто-гумусовых кутан, инфиллингов, интеркалляций.
Физико-химические характеристики исследованных почв отвечают известным особенностям буроземов грубогумусированных. В гумусовых горизонтах отмечается наиболее кислая реакция среды, что особенно ярко выражено в разрезе Онот-1 (pH 3.8). В структурно-метаморфических горизонтах ВМ наблюдается некоторое увеличение значений рН. Содержание органического углерода в горизонтах опада колеблется от 50 до 53%. В грубогумусированных горизонтах Aao содержание углерода также очень высокое и составляет от 16 до 42%. В серогумусовых горизонтах с примесью грубогумусированного материала (AYao) Сорг составляет от 10.9 (Китой-1) до 13.8% (Онот-1), что существенно больше, чем предписано диагностикой для обычных горизонтов AY. Горизонты AYpyr разреза Онот-2 и Китой-1, хотя в них и не наблюдается примеси грубогумусового материала, также содержат повышенное количество органического углерода (5.0 и 7.7% соответственно), вероятно, за счет обогащения пирогенным материалом. Вниз по профилю содержание Сорг во всех разрезах уменьшается, что характерно и для содержания общего азота. Однако в разрезе Онот-2 наибольшее содержание общего азота отмечается для нижней части подстилки (1.8%). Соотношение С : N максимально широкое в опаде и подстилке. Вниз по профилю почв оно, как правило, сужается.
Максимальное содержание илистой фракции приурочено к гумусовым горизонтам почв. Вниз по профилю доля фракции <0.001 мм в целом снижается, демонстрируя отдельные пики в горизонтах С, что не характерно для разреза Китой-1. Кровля структурно-метаморфического горизонта почв характеризуется повышенным содержанием ила по сравнению с нижележащими горизонтами. Вариации песчаных и пылеватых фракций не обнаруживают каких-либо ясных закономерностей, заметно изменяясь в пределах горизонтов. Как правило, рост доли песчаных фракций совпадает с пропорциональным снижением участия фракций пыли.
Состав стабильных изотопов углерода растительности. Изотопный состав углерода доминирующих видов растительности на исследуемой территории легкий и варьирует в пределах от –32.6 до –27.8‰, что характерно для растений с С3-фотосинтезом. Статистически значимых различий по вариациям значений δ13C в хвойных и лиственных породах деревьев и крупных кустарников не выявлено (критерий Манна–Уитни U = 4, p = 0.25). Однако в среднем хвойные на 0.5‰ тяжелее, чем лиственные (рис. 5а), что в целом согласуется с проведенными ранее исследованиями, в ходе которых выявлены более низкие значения δ13С в мелколиственных породах из-за их меньшей эффективности использования воды в ходе транспирации по сравнению с хвойными [23, 46].
Рис. 5.
Диапазоны значений δ13C для пород (а): хвойных (1) и лиственных (2), и по ярусам (b): 1 – древесному, 2 – древесно-кустарниковому, 3 – травяно-кустарничковому, 4 – моховому. Точками показаны фактические значения, черта внутри ящика – среднее, края ящика – квартили.

Внутри растительных сообществ в пределах исследованных площадок значения δ13C существенно варьируют, самый широкий разброс наблюдается на Оноте-1 (4.8‰), самый узкий – на Китое-1 (2.2‰). При этом среднее значение δ13C доминирующей растительности для всех трех площадок практически одинаковое –30.5‰. Явных зависимостей состава стабильных изотопов углерода растительности от высотно-поясного положения и микроклиматических условий не прослеживается.
Состав стабильных изотопов углерода опада и ПОВ. Наиболее легким изотопным составом углерода характеризуются горизонты опада и подстилки разреза Онот-2 (δ13С от –28.75 до –29.11‰). Бóльшие значения δ13С характерны для опада в разрезе Китой-1 (–27.9‰). Разница в изотопном составе углерода при переходе от растительности к опаду на площадках сильно варьирует. Максимальное утяжеление в опаде по сравнению со средним значением δ13C доминирующих видов растительности наблюдается на Китое-1 (2.6‰), чуть меньше на Оноте-1 (2.4‰), минимальное на Оноте-2 (1.39‰).
Изотопный состав опада на Оноте-2 очень близок по составу с основными доминатами – сосной и осиной, хвоя и листья которых составляют основную массу опада.
На Китое-1 опад неоднородный, смешанного состава со значительной долей листьев и стеблей коротконожки перистой (Brachypodium pinnatum (L.) Beauv.), данные об изотопном составе углерода которой пока отсутствуют. Можно предположить, что участие растительных остатков коротконожки повлияло на существенное утяжеление опада по сравнению с доминирующими древесными видами. Опад злаковых растений за счет примеси циннамиловых фенолов отличается более тяжелым изотопным составом по сравнению с древесными [19].
На Оноте-1 состав опада также смешанный: значительна доля хвои кедра и мха (Pleurozium schreberi), а также присутствуют растительные остатки папоротника – щитовника распростертого (Dryopteris expansa). Вероятно, одной из причин столь значительного утяжеления опада (в 1.5‰ по сравнению с живой хвоей кедра и 4.5‰ по сравнению с живым мхом) является присутствие самого тяжелого по составу щитовника, так как степень разложения растительных остатков довольно слабая. Еще одним фактором, влияющим на утяжеление состава стабильных изотопов углерода опада, может являться обогащение 13С отмерших (уже нефотосинтезирующих) органов плевроциума [51].
Серогумусовые горизонты разрезов Онот-2 и Китой-1 близки по своему изотопному составу, тогда как грубогумусовый горизонт разреза Онот-1 демонстрирует несколько бóльшие значения δ13С. Во всех исследуемых разрезах состав стабильных изотопов углерода утяжеляется с глубиной, что совпадает с уменьшением содержания общего органического углерода вниз по профилю. Градиент значений δ13С (Δ) более выражен в разрезах Онот-1 и Онот-2 (5.48 и 5.06‰ соответственно), по сравнению с разрезом Китой-1 (4.01‰).
Значение β наиболее выражено по модулю в точке Онот-2 (–2.8), в точках Онот-1 и Китой-1 оно составило –2.5 и –2.4 соответственно.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Экологические факторы изменчивости состава стабильных изотопов углерода. В целом исследуемые буроземы являются одними из наиболее изотопно-легких почв Байкальского региона [11]. Основываясь на факте наибольшей дискриминации 13С в ходе фотосинтеза, нежели чем в ходе последующих процессов [42, 45], и учитывая ранее выявленную региональную зависимость состава стабильных изотопов углерода ПОВ от влагообеспеченности [12], данное обстоятельство в наибольшей степени связано с благоприятными условиями увлажнения для растительности, формирующей органическое вещество почв.
По составу стабильных изотопов углерода исследуемые почвы отличаются незначительно. Одной из причин этого можно считать относительную схожесть климатических условий. Стоит отметить, что факт сходства величин δ13С в почвах с различным органическим веществом, формирующихся при этом в сходных гидротермических условиях, отмечался ранее [12].
Несмотря на слабое проявление, различия присутствуют. Так, в подстилочных горизонтах отмечаются вариации значений δ13С в пределах 1‰. С учетом достаточного увлажнения на всех площадках логично предположить влияние других факторов на фракционирование изотопов углерода в исследуемых почвах.
В основном углерод подстилки формируется за счет углерода растительного опада, поэтому частично подобные различия можно объяснить субстратной неоднородностью изотопного состава растительности. При близких средних значениях δ13С для доминирующих видов, изотопный состав углерода видов, составляющих большую часть проективного покрытия на разных участках, различается. Так, осина на площадке Онот-2 характеризуется значением δ13С –29.99‰, в то время как δ13С для кедра на площадке Онот-1 составляет –29.58‰.
Дополнительным фактором наблюдаемых изотопных различий ОВ почв исследуемого ряда может служить структура растительных сообществ. Площадки Онот-1 и Онот-2 представляют собой различные сукцессионные стадии темнохвойной тайги. Поэтому кедр принимает заметное участие в сообществах, произрастающих не только в точке Онот-1, но и на площадке Онот-2. Однако на последней он моложе, располагается в более низких ярусах и характеризуется облегченным изотопным составом углерода. Изотопные различия, во-первых, могут быть связаны с эффектом лесного полога, когда в плотных древостоях концентрация 13С в листьях растений имеет выраженный вертикальный градиент [32, 43]. Эффект связан с особенностями фотосинтеза в условиях затенения и с фиксацией растениями обедненной 13C углекислоты, высвобождаемой почвой и подстилкой. Его проявление подтверждается анализом изменений δ13C в растениях разных жизненных форм (рис. 5b), а также результатами дисперсионного анализа (тест Краскела–Уоллиса H = 9.76, p = 0.008), согласно которым между первым древесным и травяно-кустарничковым (без учета папоротника) ярусами есть статистически значимые различия (p = 0.002 по критерию Данна). Исключением из описанной закономерности является щитовник распростертый с самыми высокими значениями δ13C в листьях (рис. 5b) несмотря на произрастание в нижнем ярусе. В данном случае повышение δ13С, возможно, связано с повышенной интенсивностью фотосинтеза за счет частичной микогетеротрофии (обогащение микоризными грибами), свойственной папоротникам [41, 54].
Во-вторых, активность ряда физиологических процессов кедра изменяется в зависимости от высоты произрастания. В частности, эффективность использования воды кедром, являющаяся одним из факторов изотопного фракционирования [46, 50], значительно увеличивается с высотой [5]. Вариации данного показателя на исследуемых площадках могут объяснить значительное утяжеление состава стабильных изотопов углерода кедра на площадке Онот-1, находящейся гипсометрически выше.
Внутрипочвенные факторы фракционирования изотопов углерода. Изменение соотношения 12С/13С с глубиной в профилях исследуемых почв существенно по сравнению со степными и лесостепными почвами Байкальского региона [11] и, как правило, превышает 4–5‰. Одними из основных факторов естественных внутрипрофильных вариаций состава стабильных изотопов углерода ПОВ считаются: процессы внутрипрофильного перемещения водорастворимых форм органического вещества [47, 61], различное участие опада/корневых остатков в формировании ПОВ в различных частях профиля [66], фракционирование изотопов в ходе микробной трансформации ПОВ, остаточное накопление в ПОВ устойчивых к разложению компонентов [45, 47, 48].
В таежно-лесной зоне водная миграция является основным механизмом, обусловливающим транспорт и пространственную дифференциацию веществ в почве. В частности, это относится к водорастворимым формам ПОВ [17]. Преимущественно нисходящая миграция гидрофильных веществ, обогащенных 13С, и физико-химическая сорбция гидрофобных соединений, обедненных 13С, может влиять на изотопный профиль почв [47, 61]. Несмотря на то, что некоторая часть таких растворов может теоретически достичь срединной и более глубоких частей профиля при быстром движении влаги по ходам корней или трещинам, все они могут быть отнесены к быстроразлагаемым веществам, время существования которых в растворе составляет не более нескольких месяцев [17]. Согласно данным исследований состава стабильных изотопов углерода различных пулов органического вещества, выявлено, что изотопный состав почвы определяется изотопной подписью органического вещества фракции остатка, представленного органическим веществом микроагрегатов илистых частиц [2]. Согласно микроморфологическим наблюдениям как минеральное, так и органическое вещество в исследуемых почвах остается стабильным: отсутствуют диагностические морфологические признаки его активной межгоризонтной миграции. Это означает, что большая часть ПОВ ниже органо-аккумулятивных горизонтов поступает преимущественно с корневыми остатками. Поэтому при теоретически возможном влиянии данного фактора на изотопный профиль исследуемых буроземов обнаружить его сложно.
Одной из основных закономерностей при формировании органопрофиля лесных почв является различное участие опада/корневых остатков в формировании ПОВ различных частей профиля [7]. Растительный опад, формируя органическое вещество верхних частей профиля, характеризуется облегченным изотопным составом углерода по сравнению с корнями [66], принимающими заметное участие в поступлении ПОВ в срединные и нижние части профиля [1, 31]. В биомассе корней содержится меньшая часть запасов органического вещества (общая масса корней в лесах таежно-лесной зоны составляет до 25% наземной биомассы). Корни в основном многолетние и не рассматриваются как главный источник гумуса в лесных почвах. Более важна роль тонких корней (d < l мм). Высокая скорость синтеза и низкая скорость разложения предполагают большое значение тонких корней в формировании запасов ПОВ в профиле лесных почв умеренного пояса [62, 64].
Как правило изотопный состав опада и тонких корней близок. Например, в органах и тканях березы наибольшая дискриминация тяжелого изотопа 13С (δ13C до –29‰) отмечена как в листьях, так и в тонких корнях [8]. Несколько более тяжелым составом изотопов отличаются скелетные корни. Такое сходство δ13C листьев и тонких корней в результате прямого и быстрого переноса сахаров по флоэме из листьев в корни представляет подпись изотопной дискриминации углерода фотосинтеза как в ходе отложения “свежего” углерода в камбиальной ткани, так и в ходе углеродного питания корневой системы.
В исследуемых почвах отличия состава стабильных изотопов углерода в верхней 20 см толще могут достигать 2–3‰, что явно перекрывает диапазон различий изотопного состава корней и опада [46]. На наш взгляд, различное соотношение в поступлении надземной фитомассы и корней не может считаться существенным фактором наблюдаемых изотопных вариаций. Это логично с позиции того, что почва не может рассматриваться как статичное тело, и процессы физического перемешивания органического вещества с различным изотопным составом [39, 48] отражаются в большей степени в общем изотопном профиле почв, чем вклад отдельных компонентов поступающей биомассы.
Фракционирование изотопов углерода в ходе микробного разложения ПОВ является одной из наиболее распространенных гипотез для объяснения утяжеления изотопного состава углерода ПОВ с глубиной [39, 44, 52]. В ходе большинства биологических реакций происходит дискриминация изотопов, то есть для ферментативных реакций используются более легкие изотопы, оставляя в субстрате более тяжелые. Степень обогащения ПОВ 13C увеличивается по мере увеличения циклов микробной утилизации [66]. Одним из показателей, позволяющих оценить интенсивность данного процесса и его роль в изотопной дифференциации почв, является наклон линейной регрессии (β) между значениями δ13С и изменением содержания органического углерода (в логарифмическом масштабе) с глубиной [39, 52, 65]. Несмотря на то, что данный показатель служит хорошим индикатором интенсивности оборота углерода в почвах, формирующихся в широком спектре климатических условий [65], а также в контрастных ландшафтно-климатических условиях [13], в рассматриваемых буроземах значения β оказались практически идентичны. На наш взгляд, это можно связать с относительно однородными климатическими условиями, в которых формируются почвы. Однако данная трактовка не объясняет сходство значений коэффициента при настолько выраженных различиях в строении органопрофиля почв. Такие различия, безусловно, отражают существенную разницу в условиях поступления и последующей трансформации органического вещества.
Строение органопрофиля почв как фактор изотопных различий. Исходя из морфогенетических различий, прежде всего, строения органопрофилей, очевидно, что процессы метаморфизма органического вещества: биогенной и абиогенной трансформации растительных остатков, гумификации и закрепления in situ продуктов гумификации, а также минерализации органического вещества наиболее активно протекают в профиле Китой-1 и меньше в профиле Онот-2. Наименьшей активностью этих процессов характеризуется разрез Онот-1. Можно предположить, что в почвах, где активность трансформации растительных остатков наименьшая в рассматриваемом ряду, продукция СО2 будет также низкой. Но полученные данные свидетельствуют об обратном: при низкой интенсивности трансформации органического вещества в разрезе Онот-1, продукция СО2 здесь повышена по сравнению с разрезом Онот-2, где процессы трансформации идут активнее, а СО2 продуцируется меньше.
Вероятно, именно неоднородное строение органопрофиля вносит основной вклад в наблюдаемую противоречивую картину. Значительная часть органопрофиля разреза Онот-1 представлена нестабилизированным органическим веществом в отличие от преимущественно минерально-ассоциированного относительно стабильного ОВ гумусовых горизонтов в разрезах Онот-2 и Китой-1. Активность минерализации углерода незащищенного ОВ подстилочных горизонтов (определяемая по активности выделения СО2), больше в 3–5 раз по сравнению с минеральной почвой [24, 53]. При этом под кедром активность минерализации в подстилке оценивается как одна из наиболее высоких [24]. С учетом выявленной повышенной интенсивности продуцирования СО2 в почве Онот-1 по сравнению с Онот-2 данный фактор может рассматриваться как один из значимых при объяснении повышенных значений δ13С в почве Онот-1 и выраженного наклона β, появляющегося даже при наблюдаемой здесь аккумуляции органического вещества в виде подстилки и опада. Расширенное соотношение C : N в органогенных горизонтах разрезов Онот-2 и Китой-1 также говорит в пользу сниженной интенсивности минерализации ПОВ по сравнению с разрезом Онот-1. Дополнительной причиной активной минерализации ПОВ в подстилке кедрового леса и значительного утяжеления изотопного состава органогенных горизонтов в точке Онот-1 может являться деятельность грибов, обеспечиваемая за счет развития облигаторной микоризы у кедра [6] и частичной у папоротника [54, 59].
ЗАКЛЮЧЕНИЕ
Первичным фактором, обусловливающим изотопную неоднородность почв, являются климатические условия. Достаточное увлажнение на исследуемой территории обеспечивает выраженную дискриминацию 13С в растительных тканях при фиксации углерода. Важно отметить, что в связи с благоприятными условиями увлажнения исследуемые буроземы являются одними из наиболее изотопно-легких почв Байкальского региона.
Модулирующими факторами служат видовые различия, физиологические особенности растений и их ярусное положение. Совокупное влияние перечисленных факторов обеспечивает исходную (субстратную) неоднородность изотопного состава углерода продуцируемого опада. Тем не менее, вариации значений δ13С в связи с изменчивостью указанных факторов в исследуемых буроземах не превышают 1‰. Наиболее логично связывать такую незначительную вариативность с относительно узким диапазоном климатических условий формирования почв.
Значительные отличия в строении органопрофиля исследуемых почв, отражающие существенные различия в условиях поступления и последующей трансформации органического вещества, не находят отражения в составе стабильных изотопов углерода ПОВ. Это может привести к заключению о слабом влиянии процессов внутрипочвенного фракционирования изотопов углерода на изотопный состав почв. Однако такое заключение противоречит значительному внутрипочвенному градиенту значений δ13С, превышающему 4–5‰ и сильно отличающему исследуемые почвы от почв степей и лесостепей Байкальского региона.
Наиболее логичным на данном этапе кажется предположение, что изотопный профиль исследуемых буроземов складывается под влиянием микробной утилизации ОВ, проявляющейся с разной интенсивностью в зависимости от строения органопрофиля. В почвах с преобладанием минеральных горизонтов в органопрофиле (Онот-2, Китой-1) утяжеление изотопного состава углерода происходит за счет прохождения многочисленных циклов микробной утилизации ОВ и его ассоциирования с минеральной фазой. Учитывая невысокие значения коэффициента наклона линейной регрессии (β), интенсивность оборота углерода при этом относительно мала.
Повышенная интенсивность минерализации легкодоступных компонентов растительного материала в верхней части органопрофиля почв с развитой подстилкой и органо-аккумулятивными горизонтами (Онот-1) может приводить к заметному росту значений δ13С и выраженному наклону β, появляющемуся даже при наблюдаемой здесь аккумуляции органического вещества в виде подстилки и опада.
Несмотря на то, что микробная утилизация ОВ рассматривается нами в качестве ведущего фактора в создании внутрипрофильных вариаций величин δ13С, существует множество других факторов, которые потенциально могут влиять на состав стабильных изотопов углерода органического вещества исследуемых почв. Определить степень их участия на данном этапе исследования довольно проблематично. Она может быть оценена при анализе изотопного состава углерода отдельных пулов органического вещества.
Список литературы
Александрова Л.Н. Органическое вещество почвы и процессы его трансформации. Л.: Наука, 1980. 288 с.
Артемьева З.С., Данченко Н.Н., Зазовская Э.П., Колягин Ю.Г., Кириллова Н.П., Когут Б.М. Изотопный состав углерода и химическая структура органического вещества типичного чернозема в условиях контрастного землепользования // Почвоведение. 2021. № 6. С. 686–700.
Байкал. Атлас. М.: Федеральная служба геодезии и картографии России, 1993. 168 с.
Балсанова Л.Д., Гынинова А.Б., Цыбикдоржиев Ц.Ц., Гончиков Б.-М.Н., Шахматова Е.Ю. Генетические особенности почв бассейна озера Котокельское (Восточное Прибайкалье) // Почвоведение. 2014. № 7. С. 781–789.
Бендер О.Г., Зотикова А.П., Бендер А.Г. Состояние фотосинтетического аппарата разновозрастной хвои кедра сибирского на южном пределе произрастания в горах Алтая // Лесные биогеоценозы бореальной зоны: география, структура, функции, динамика. Новосибирск: Изд-во СО РАН, 2014. С. 380–383.
Вайшля О.Б., Карбышева К.С., Бендер О.Г. Особенности микотрофности Pinus sibirica Du Tour в кедровниках Томской области // Известия Санкт-Петербургской лесотехнической академии. 2019. Вып. 229. С. 7–22.
Ведрова Э.Ф. Интенсивность деструкции органического вещества серых почв в лесных экосистемах южной тайги Центральной Сибири // Почвоведение. 2008. № 8. С. 973–982.
Воронин П.Ю., Мухин В.А., Веливецкая Т.А., Игнатьев А.В., Кузнецов Вл.В. Изотопный состав углерода и азота в органах и тканях Betula pendula // Физиология растений. 2017. Т. 64. № 2. С. 127–132.
Воропай Н.Н., Киселев М.В., Черкашина А.А. Мониторинг температуры почв на многолетнемерзлых породах в естественных и антропогенно нарушенных условиях Тункинской котловины // Лед и снег. 2019. Т. 59. № 4. С. 517–528. https://doi.org/10.15356/2076-6734-2019-4-421
Герасимова М.И., Губин С.В., Шоба С.А. Микроморфология почв природных зон СССР. Пущино: Пущинский научный центр РАН, 1992. 214 с.
Голубцов В.А. Состав стабильных изотопов углерода органического вещества позднеплейстоценовых и голоценовых почв Байкальского региона // Почвоведение. 2020. № 6. С. 673–690.
Голубцов В.А., Вантеева Ю.В., Воропай Н.Н. Влияние влагообеспеченности на состав стабильных изотопов углерода органического вещества почв Байкальского региона // Почвоведение. 2021. № 10. С. 1182–1194.
Голубцов В.А., Вантеева Ю.В., Воропай Н.Н., Василенко О.В., Черкашина А.А., Зазовская Э.П. Состав стабильных изотопов углерода (δ13С) как показатель динамики органического вещества в почвах западного побережья озера Байкал // Почвоведение. 2022. № 12. С. 1489–1504. https://doi.org/10.31857/S0032180X22600597
Государственная геологическая карта Российской Федерации. М-б 1 : 1 000 000 (третье поколение). Серия Ангаро-Енисейская. Лист N-48 – Иркутск. Объяснительная записка. СПб.: Картфабрика ВСЕГЕИ, 2009. 574 с.
Жуков В.М. Климат // Предбайкалье и Забайкалье. М.: Наука, 1965. С. 91–126.
Караваева Н.А., Таргульян В.О., Черкинский А.Е., Целищева Л.К., Грачева Р.Г., Марголина Н.Я., Ильичев Б.А., Горячкин С.В., Александровский А.Л., Хитров Н.Б., Замотаев И.В., Панкова Е.И., Айдаров И.П., Ромашкевич А.И., Черняховский А.Г., Бирина А.Г., Гоголев А.И., Розанов А.Б. Элементарные почвообразовательные процессы. Опыт концептуального анализа, характеристика, систематика. М.: Наука, 1992. 184 с.
Караванова Е.И., Золовкина Д.Ф. Влияние состава подстилок на характеристики их водорастворимых органических веществ // Вестник Моск. ун-та. Сер. 17, почвоведение. 2020. № 2. С. 17–23.
Карпачевский Л.О., Киселева Н.К. О методике определения и некоторых особенностях выделения СО2 из почв под широколиственно-еловыми лесами // Почвоведение. 1969. № 7. С. 32–42.
Ковалева Н.О., Ковалев И.В. Лигниновые фенолы в почвах как биомаркеры палеорастительности // Почвоведение, 2015. № 9. С. 1073–1086.
Королюк А.Ю. Использование экологических шкал в геоботанических исследованиях // Актуальные проблемы геоботаники. Петрозаводск, 2007. С. 176–197.
Кузьмин В.А. Почвенно-экологическое районирование Иркутской области // Почвоведение. 2002. № 12. С. 1436–1444.
Ландшафты юга Восточной Сибири [Карты]: [физическая карта] / Aвт. В.С. Михеев, В.А. Ряшин, 1 : 1 500 000. М.: ГУГК, 1977. 4 л.
Меняйло О.В., Макаров М.И., Ченг Ш.-К. Изотопный состав углерода (δ13C) и азота (δ15N) в хвое/листьях и почве в модельных древостоях разных пород // Доклады АН. 2014. Т. 456. С. 738–740. https://doi.org/10.7868/S0869565214180285
Меняйло О.В., Матвиенко А.И., Макаров М.И., Ченг Ш.-К. Положительный отклик минерализации углерода на внесение азота в лесных почвах Сибири // Доклады АН. 2014. Т. 456. № 1. С. 117–120. https://doi.org/10.7868/S0869565214130271
Мировая коррелятивная база почвенных ресурсов: основа для международной классификации и корреляции почв. М.: Товарищество научных изданий КМК, 2007. 278 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 22. Иркутская область и западная часть Бурятской АССР. Л.: Гидрометеоиздат, 1991. 604 с.
Прудникова А.Ю., Чепинога В.В. Флора бассейна реки Тойсук (предгорья Восточного Саяна, Иркутская область). Иркутск: Изд-во ИГУ, 2012. 189 с.
Пулы и потоки углерода в наземных экосистемах России. М.: Наука, 2007. 315 с.
Смоленцев Б.А., Смоленцева Е.Н. Буроземы Кузнецкого Алатау, их свойства и разнообразие // Вестник Том. гос. ун-та. Биология. 2020. № 50. С. 6–27. https://doi.org/10.17223/19988591/50/1
Сымпилова Д.П., Бадмаев Н.Б. Почвообразование в ландшафтах контакта тайги и степи Селенгинского среднегорья (Западное Забайкалье) // Почвоведение. 2019. № 2. С. 140–151.
Титлянова А.А., Булавко Г.И., Миронычева-Токарева Н.П., Хвощевская М.Ф. Запасы органического углерода в почвах Западной Сибири // Почвоведение. 1994. № 10. С. 49–53.
Тиунов А.В. Стабильные изотопы углерода и азота в почвенно-экологических исследованиях // Известия РАН. Сер. биологическая. 2007. № 4. С. 475–489.
Тиходеева М.Ю., Лебедева В.Х. Практическая геоботаника (анализ состава растительных сообществ). СПб.: Изд-во СПбГУ, 2015. 166 с.
Убугунова В.И., Цыбжитов Ц.Х., Большаков В.А. Бурые горно-лесные почвы Прибайкалья // Почвоведение. 1985. № 7. С. 15–21.
Убугунов Л.Л., Гынинова А.Б., Белозерцева И.А., Доржготов Д., Убугунова В.И., Сороковой А.А., Убугунов В.Л., Бадмаев Н.Б., Гончиков Б.Н. Географические закономерности распределения почв на водосборной территории оз. Байкал (к карте “почвы бассейна оз. Байкал”) // Природа Внутренней Азии. 2018. № 2(7). С. 7–26. https://doi.org/10.18101/2542-0623-2018-2-7-26
Цыбжитов Ц.Х., Убугунова В.И., Ермакова О.Д. Водно-физические свойства и гидротермический режим буроземов Прибайкалья // Почвоведение. 1989. № 4. С. 65–70.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 198 с.
Экологические шкалы и методы анализа экологического разнообразия растений. Йошкар-Ола: Мар. гос. ун-т., 2010. 368 с.
Acton P., Fox J., Campbell E., Rowe H., Wilkinson M. Carbon isotopes for estimating soil decomposition and physical mixing in well-drained forest soils // J. Geophys. Res. Biogeosci. 2013. V. 11(4). P. 1532–1545. https://doi.org/10.1002/2013JG002400
Amundson R. The carbon budget in soils // Annual Rev. Earth Planetary Sci. 2001. V. 29. P. 535–562.
Ahulu E.M., Nakata M., Nonaka M. Arum-and Paris-type arbuscular mycorrhizas in a mixed pine forest on sand dune soil in Niigata Prefecture, central Honshu, Japan // Mycorrhiza. 2005. V. 15. P. 129–136.
Bowling D.R., Pataki D.E., Randerson J.T. Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes // New Phytologist. 2008. V. 178. P. 24-40. https://doi.org/10.1111/j.1469-8137.2007.02342.x
Brooks J.R., Flanagan L.B., Varney G.T., Ehleringer J.R. Vertical gradients in photosynthetic gas exchange characteristics and refixation of respired CO2 within boreal forest canopies // Tree Physiol. 1997. V. 17. P. 1–12.
Brunn M., Spielvogel S., Sauer T., Oelmann Y. Temperature and precipitation effects on δ13C depth profiles in SOM under temperate beech forests // Geoderma. 2014. V. 235–236. P. 146–153. https://doi.org/10.1016/j.geoderma.2014.07.007
Camino-Serrano M., Tifafi M., Balesdent J., Hatté C., Peñuelas J., Cornu S., Guenet, B. Including stable carbon isotopes to evaluate the dynamics of soil carbon in the land-surface model ORCHIDEE // J. Adv. Modeling Earth Systems. 2019. V. 11. P. 3650–3669. https://doi.org/10.1029/2018MS001392
Dawson T.E., Mambelli S., Plamboeck A.H., Templer P.H., Tu K.P. Stable isotopes in plant ecology // Annual Reviews of Ecology and Systematics. 2002. V. 33. P. 507–559. https://doi.org/10.1146/annurev.ecolsys.33.020602.095451
Dümig A., Rumpel C., Dignac M.F., Kögel-Knabner, I. The role of lignin for the δ13C signature in C-4 grassland and C-3 forest soils // Soil Biol. Biochem. 2013. V. 57. P. 1–13. https://doi.org/10.1016/j.soilbio.2012.06.018
Ehleringer J.R., Buchmann N., Flanagan L.B. Carbon isotope ratios in belowground carbon cycle processes // Ecol. Appl. 2000. V. 10. P. 412–422.
Eswaran H., Reich P.F., Kimble J.M., Beinroth F.H., Padmanabhan E.M.P. Global carbon stocks // Global Change and Pedogenic Carbonate. CRC Press, Boca Raton, FL, 2000. P. 15–25.
Farquhar G.D., Ehleringer J.R., Hubick K.T. Carbon isotope discrimination and photosynthesis // Annual Review Plant Physiology and Plant Molecular Biology. 1989. V. 40. P. 503–537.
Flanagan L.B., Kubien D.S., Ehleringer J.R. Spatial and temporal variation in the carbon and oxygen stable isotope ratio of respired CO2 in a boreal forest ecosystem // Tellus B: Chem. Phys. Meteoro. 1999. V. 51(2). P. 367–384. https://doi.org/10.3402/tellusb.v51i2.16306
Garten C.T., Cooper L.W., Post W.M., Hanson P.J. Climate Controls on Forest Soil C Isotope Ratios in the Southern Appalachian Mountains // Ecology. 2000. V. 81(4). P. 1108–1119.
Garten C.T., Post W.M., Hanson P.J., Cooper L.W. Forest soil carbon inventories and dynamics along an elevation gradient in the southern Appalachian Mountains // Biogeochemistry. 1999. V. 45. P. 115–145.
Giesemann P. Understanding plant-fungal nutritional strategies using stable isotopes. Bayreuth, 2021. 165 p.
Hammer Ø., Harper D.A.T., Ryan P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. V. 4(1). 4. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
IUSS Working Group WRB. World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports. № 106. Rome, 2015. 192 p.
Kögel-Knabner I., Amelung W. Soil organic matter in major pedogenic soil groups // Geoderma. 2021. V. 384. 114785. https://doi.org/10.1016/j.geoderma.2020.114785
Leifeld J., von Lützow M. Chemical and microbial activation energies of soil organic matter decomposition // Biol Fertil Soils. 2014. V. 50. P. 147–153. https://doi.org/10.1007/s00374-013-0822-6
Lyu M., Xie J., Giardina C.P., Vadeboncoeur M.A., Feng X., Wang M., Ukonmaanaho L., Lin T-Ch., Kuzyakov Y., Yang Y. Understory ferns alter soil carbon chemistry and increase carbon storage during reforestation with native pine on previously degraded sites // Soil Biology and Biochemistry. 2019. V. 132. P. 80–92. https://doi.org/10.1016/j.soilbio.2019.02.004
Nadelhoffer K.F., Fry B. Controls on natural 15N and 13C abundances in forest soil organic matter // Soil Sci. Soc. Am. J. 1988. V. 5(6). P. 1633–1640.
Nakanishi T., Atarashi-Andoh M., Koarashi J., Saito-Kokubu Y., Hirai K. Carbon isotopes of water-extractable organic carbon in a depth profile of forest soil imply a dynamic relationship with soil carbon // Eur. J. Soil Sci. 2012. V. 63(4). P. 495–500. https://doi.org/10.1111/j.1365-2389.2012.01465.x
Persson H. The high input of soil organic matter from dead tree fine roots into the forest soil // Int. J. Forestry Res. 2012. P. 217402. https://doi.org/10.1155/2012/217402
Pregitzer K.S., Euskirchen E.S. Carbon cycling and storage in world forests: biome patterns related to forest age // Global Change Biol. 2004. V. 10. P. 2052-2077. https://doi.org/10.1111/j.1365-2486.2004.00866.x
Wang C., Chen Z., Brunner I., Zhang Z., Zhu X., Li J., Yin H., Guo W., Zhao T., Zheng X., Wang S., Geng Z., Shen S., Jin D., Li M.-H. Global patterns of dead fine root stocks in forest ecosystems // J. Biogeography. 2018. V. 45. P. 1378–1394. https://doi.org/10.1111/jbi.13206
Wang C., Houlton B.Z., Liu D., Hou J., Cheng W., Bai E. Stable isotopic constraints on global soil organic carbon turnover // Biogeosciences. 2018. V. 15. P. 987–995. https://doi.org/10.5194/bg-15-987-2018
Werth M., Kuzyakov Y. 13C fractionation at the root–microorganisms–soil interface: a review and outlook for partitioning studies // Soil Biol. Biochem. 2010. V. 42(9). P. 1372–1384. https://doi.org/10.1016/j.soilbio.2010.04.009
Дополнительные материалы
- скачать ESM.docx
- Приложение 1.
Таблица S1. Основные характеристики исследуемых площадок
Рисунок S2. Коэффициенты наклона линейной регрессии (β) для исследуемых почв.