

Палеонтологический журнал, 2022, № 6, стр. 42-50
Новый вид брахиопод из отряда Pentamerida в ордовике Прителецкого Алтая
Т. А. Щербаненко a, *, Н. В. Сенников a, **
a Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
630090 Новосибирск, Россия
* E-mail: Shcherbanenkota@ipgg.sbras.ru
** E-mail: Sennikovnv@ipgg.sbras.ru
Поступила в редакцию 28.04.2022
После доработки 19.05.2022
Принята к публикации 19.05.2022
- EDN: LORSMN
- DOI: 10.31857/S0031031X22060101
Аннотация
Описан новый вид брахиопод Eoanastrophia tozodoviensis sp. nov. из тозодовской толщи верхнего ордовика Прителецкой структурно-фациальной зоны Горного Алтая. Предложено расширение диагноза рода Eoanastrophia Nikiforova et Sapelnikov.
ВВЕДЕНИЕ
Брахиоподы – одна из основных бентосных групп фауны, на базе которых традиционно проводилось биостратиграфическое обоснование подразделений региональных и местных стратиграфических шкал палеозоя. Исследование плеченогих ордовика Алтае-Саянской складчатой области началось с 50-х гг. прошлого века Л.Г. Севергиной (1960), позднее О.Н. Андреевой (1982, 1985), Н.П. Кульковым (Кульков, Севергина, 1984) и др. К настоящему времени оттуда описано и определено более 200 видов (Кульков, Севергина, 1989).
Настоящее сообщение посвящено алтайским представителям пентамерид – эоанастрофиям. Род Eoanastrophia Nikiforova et Sapelnikov, 1973 известен из верхнего ордовика Тянь-Шаня (Узбекистан, Казахстан), Горного Алтая (Россия), горных массивов Италии, Испании и Франции. Видовые таксоны рассматриваемого рода распространены в узком стратиграфическом интервале (верхний ордовик, сандбиан–катиан), в связи с чем они имеют важное значение для стратиграфии.
О.И. Никифоровой и В.П. Сапельниковым (1973) был выделен из нижнеарчалыкских слоев ашгиллского яруса перевала Шахриомон Южного Тянь-Шаня новый род Eoanastrophia с типовым видом E. antiquata Nikiforova et Sapelnikov. Затем другими специалистами было описано несколько видов в рамках этого рода: 1) E. kurdaica Sapelnikov et Rukavishnikova из кескинтасской свиты андеркенского горизонта среднего–верхнего карадока (Сапельников, Рукавишникова, 1975); 2) E. transversa Severgina из орловского горизонта верхнего ордовика Горного Алтая (Севергина, 1978); 3) E. primordialis Rozman из обикалонских слоев шахриомонской свиты Южного Тянь-Шаня (Розман, 1978); 4) E. pentamera (Meneghini) из карадока–ашгилла о. Сардиния, Карнийских Альп (формация de Uqua) (Италия), Пиринеев (Испания), п-ова Крозон северо-запада Франции, Армориканского массива, Черных Гор (Montagne Noire) Центрального массива Франции (Havlíček, 1981; Melou, 1990; Botquelen, 2003); 5) E. extenuata Nikitin et Popov из саргалдакской свиты еркебидаикского горизонта карадока среднего ордовика хр. Чингиз (Казахстан, Тянь-Шань) (Кленина и др., 1984); 6) E. lebediensis (Severgina) из нижней части гурьяновской свиты ханхаринского горизонта среднего–верхнего ордовика Уйменско-Лебедской структурно-фациальной зоны (СФЗ) Горного Алтая (Кульков, Севергина, 1989).
МАТЕРИАЛ
В Прителецкой СФЗ на востоке Горного Алтая в разрезе Тозодов и в палеонтологической точке (п.т.) Нижний Турочак (Сенников и др., 2018; Sennikov et al., 2019) сотрудниками Ин-та нефтегазовой геологии и геофизики им. А.А. Трофимука (ИНГГ) СО РАН при тематических работах были обнаружены представители рода Eoanastrophia. Находки этих таксонов брахиопод в алтайских изученных разрезах редки. На первом этапе исследования они были определены как E. lebediensis (Shcherbanenko, 2019). В рамках полевых работ последних лет было продолжено изучение разрезов и дополнительный сбор палеонтологических остатков в Прителецкой СФЗ Алтая. Прителецкая коллекция ордовикских брахиопод пополнилась новыми образцами эоанастрофий. На этом дополнительном материале был использован необходимый для современных исследований метод последовательных пришлифовок. Это позволило получить более точную информацию о внутреннем строении эоанастрофий. Представительный материал позволил по серии раковин и створок доказать стабильность признаков в исследуемой выборке. Кроме того, хорошая сохранность одного из экземпляров эоанастрофий дала возможность изучить строение замочного края.
Прителецкие эоанастрофии по облику близки к E. lebediensis из нижней части гурьяновской свиты сандбийского яруса верхнего ордовика Северо-Восточного Алтая (Уйменско-Лебедская СФЗ), однако внешне отличаются меньшими размерами, формой раковины, характером выпуклости створок.
ИЗУЧЕНИЕ ТИПОВОЙ КОЛЛЕКЦИИ EOANASTROPHIA LEBEDIENSIS. ОБОСНОВАНИЕ ВЫДЕЛЕНИЯ НОВОГО ВИДА
E. lebediensis был выделен в рамках рода Camerella Billings (Севергина, 1967), а позже отнесен к роду Eoanastrophia (Кульков, Севергина, 1989). В качестве специфического для рода признака было указано “наличие угловатых ребер, начинающихся от макушки” (Кульков, Севергина, 1989). Критерии, отличающие этот вид от близкородственных видов – меньшие размеры, более грубые ребра, число ребер (10–12 против 20–24 у Е. primordiаlis), меньшие размеры, более грубая скульптура, иное очертание раковины (сравнение с Е. transversa).
Описание внутреннего строения E. lebediensis (Севергина, 1967) довольно кратко: “В брюшной створке спондилиум широкий, поддерживается короткой септой. В спинной створке маленький узкий круралиум поддерживается довольно длинной срединной септой, которая протягивается почти до средины створки”. В монографии (Кульков, Севергина, 1989) описание вида не было пересмотрено, с учетом новой предложенной терминологии пентамерид (Сапельников, 1985).
В коллекции № 1323, хранящейся в Центре коллективного пользования “Коллекции уникальных геологических материалов (палеонтологических, микропалеонтологических и палинологических) Сибири и Арктики (ГЕОХРОН)” ИНГГ СО РАН (ранее – Музей Ин-та геологии и геофизики СО АН СССР, далее – ЦКП “ГЕОХРОН”) в Новосибирске, вид E. lebediensis представлен единственным экземпляром, голотипом № 403/1323. Его фото в четырех положениях приведено в монографии (Кульков, Севергина, 1989, табл. X , фиг. 19 а–г) и при первичном описании вида (Севергина, 1967). Кроме того, при первоописании присутствует изображение ядра брюшной створки, на котором видно строение спондилия (Севергина, 1967, табл. 4 , фиг. 5 ). Этот экземпляр, как и остальные представители вида (а это более 20 разрозненных створок удовлетворительной сохранности) в коллекции музея отсутствует.
Голотипом таксона Е. lebediensis является слегка помятый и надломленный с одного бока экземпляр. Со стороны брюшной створки отсутствует (смята) левая часть примакушечной области. Это затрудняет точную интерпретацию строения спондилия: на каком расстоянии от макушки сходятся зубные пластины, и с какого расстояния они поддерживаются срединной септой. В задней части створок голотипа Е. lebediensis раковинное вещество выщелочено, поэтому примакушечные области раковины, содержащие элементы спондилия и брахиофория, по типу сохранности являются ядром. Внутреннее строение Е. lebediensis было описано Севергиной (1967) по ядрам. В отличие от метода последовательных пришлифовок, изучение внутренних структур по ядрам не показывает всех деталей внутрираковинных образований. В частности, не понятны тип спондилия (сидячий, комбинированный, двойной); высота септы, протягивается ли она вперед за границы спондилия; строение зубных пластин; особенности строения брахиофория – высота дорсальной септы, параметры септальных и брахиальных пластин.
В рамках настоящей работы был изучен голотип Е. lebediensis и сделаны его фотографии (табл. VIII , фиг. 4 ). Следует обратить внимание на то, что при первом описании вида Е. lebediensis (Севергина, 1967) было отмечено: а) “Арея отсутствует. Макушки обеих створок смыкаются” – на голотипе видно, что раковинное вещество в области макушек брюшной и спинной створок выщелочено, сами макушки экземпляра не сохранились; б) “Поверхность створок покрыта 10–12 резкими округлыми складками…” – на голотипе число складок на брюшной и спинной створке одинаково и равно 14.
Первый из выше отмеченных пунктов подчеркивает имеющуюся неопределенность в характеристике замочной области этого таксона (в строении замочного края, вентральной, дорсальной ареи, во взаимоотношении макушек брюшной и спинной створок). Кроме того, неполное для современного уровня описание внутреннего строения Е. lebediensis и отсутствие для изучения большей части типовой коллекции в музее не позволяют даже формально отождествить эоанастрофий из Прителецкой СФЗ с видом Е. lebediensis.
Дополнительно к проведенным лабораторным исследованиям в 2021 г. в типовом местонахождении голотипа таксона E. lebediensis, в разрезе у устья р. Бура (правый приток р. Лебедь) была предпринята попытка пополнения типовой коллекции. Целенаправленно проводился поиск целых экземпляров этого вида, с достаточной сохранностью для изучения внутренней структуры методом последовательных пришлифовок. К сожалению, в пачке, из которой была собрана коллекция Севергиной, в найденных образцах с различными другими таксонами брахиопод, в т.ч. хорошей сохранности, представители эоанастрофий обнаружены не были.
Учитывая все перечисленные обстоятельства, прителецкие эоанастрофии выделены как новый вид.
ПАЛЕОНТОЛОГИЧЕСКОЕ ОПИСАНИЕ
Авторы придерживаются систематики брахиопод по А. Уильямсу и др. (Williams et al., 2002). При характеристике морфологических признаков использована терминология, разработанная Сапельниковым (1985).
ОТРЯД PENTAMERIDA
ПОДОТРЯД SYNTROPHIIDINA
НАДСЕМЕЙСТВО CAMERELLOIDEA HALL ET CLARKE, 1895
СЕМЕЙСТВО PARASTROPHINIDAE SCHUCHERT ET LEVENE, 1929
Род Eoanastrophia Nikiforova et Sapelnikov, 1973
Eoanastrophia tozodoviensis Shcherbanenko, sp. nov.
Табл. VIII , фиг. 1–3, 5, 6
Eoanastrophia lebediensis: Shcherbanenko, 2019, c. 190, табл. I , фиг. 1 .
Название вида – по ручью Тозодов.
Объяснение к таблице VIII
Все: Республика Алтай; верхний ордовик, сандбийский ярус.
Фиг. 1–3, 5, 6 . Eoanastrophia tozodoviensis sp. nov.: 1 – голотип ЦКП “ГЕОХРОН”, № 2110/1: 1а – брюшная створка, 1б – спинная створка, 1в – вид сбоку, 1г – вид сбоку, 1д – передний край, 1е – задний край; Прителецкая СФЗ, разрез Тозодов; тозодовская толща; 2 – экз. ЦКП “ГЕОХРОН”, № 2110/2: 2а – брюшная створка, 2б – спинная створка, 2в – вид сбоку, 2г – передний край, 2д – задний край; Прителецкая СФЗ, п.т. Нижний Турочак (С-163); 3 – экз. ЦКП “ГЕОХРОН”, № 2110/3: 3а – брюшная створка, 3б – спинная створка, 3в – вид сбоку, 3г – передний край, 3д – задний край; Прителецкая СФЗ, разрез Тозодов; тозодовская толща; 5 – экз. ЦКП “ГЕОХРОН”, № 2110/4: 5а – брюшная створка, 5б – спинная створка; Прителецкая СФЗ, разрез Тозодов; тозодовская толща; 6 – экз. ЦКП “ГЕОХРОН”, № 2110/2, серия последовательных пришлифовок; цифрами обозначено расстояние от макушки до изображенного среза; Прителецкая СФЗ, п.т. Нижний Турочак (С-163).
Фиг. 4 . Eoanastrophia lebediensis, голотип ЦКП “ГЕОХРОН”, № 403/1323: 4а – брюшная створка, 4б – спинная створка, 4в – вид сбоку, 4г – вид сбоку, 4д – передний край, 4е – задний край; Уйменско-Лебедская СФЗ, разрез по р. Лебедь; нижняя часть гурьяновской свиты.
Масштабная линейка – 10 мм.
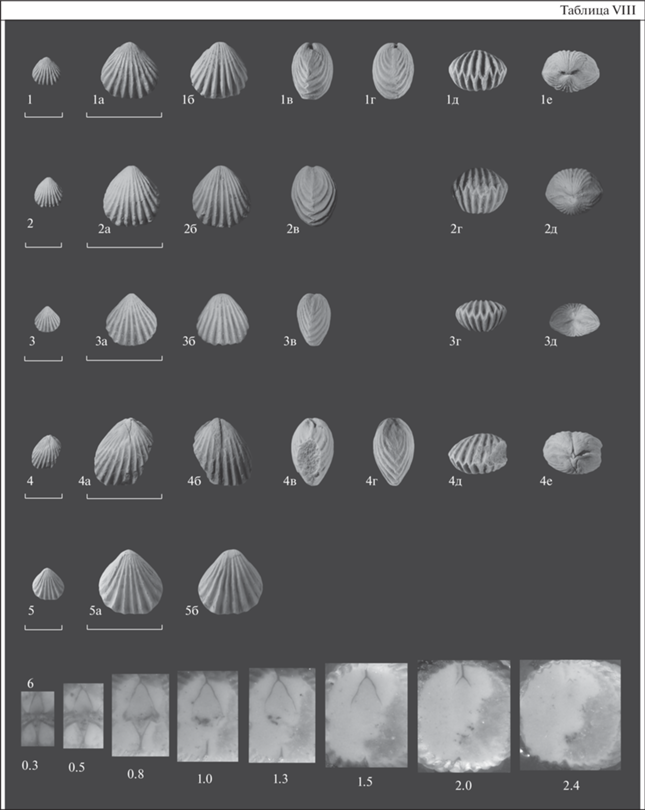
Голотип – ЦКП “ГЕОХРОН”, № 2110-1; Северо-Восточный Алтай, запад северной части Телецкого озера, разрез в правом борту руч. Тозодов, притока р. Иогач; верхний ордовик, сандбийский ярус, тозодовская толща; обозначен здесь, табл. VIII , фиг. 1 .
Описание. Раковина маленькая, не превышает 10 мм в длину и ширину, округло-пятиугольного очертания, изометричная или немного вытянутая в длину, спиннодвояковыпуклая, ребристая от макушек. Ребра простые, угловатые, 14–16 на каждой створке. Замочный край короткий, его длина составляет половину максимальной ширины раковины, которая приходится на переднюю треть длины раковины.
Брюшная створка выпуклая в центральной части. Вентральная арея ортоклинная, дельтириум открытый. Макушка прямая, заостренная, не выдается за замочный край. Синус не выражен, на передней комиссуре имеется слабозаметное понижение в сторону спинной створки.
Спинная створка более выпуклая, чем брюшная. Максимальная выпуклость в задней и средней части створки, к переднему краю равномерно уплощается. Дорсальная арея анаклинная, нототирий открытый. Макушка массивная, изогнутая, вздутая, нависает над замочным краем, прижата к дельтирию. Возвышение не развито, на переднем крае наблюдается изгиб в сторону спинной створки.
Внутреннее строение (табл. VIII , фиг. 6 ). В брюшной створке развит спондилий-комбинариум (=сложный, комбинированный, simplex), колоколообразный в сечении. На ранней стадии онтогенеза разобщенный, затем, с расстояния 0.5–0.9 мм от макушки, поддерживается низкой срединной септой, увеличивающейся по высоте в направлении передней комиссуры. Длина спондилия не превышает 3.0 мм. Септа не продолжается вперед за границы спондилия. Спондилий сложен продольно-волокнистым веществом. Зубы каплевидной формы.
В спинной створке двойной узкий брахиофорий (=круралиум, септалиум), профиль брахиофория Y-образный. Септальные пластины тонкие, тесно сближенные у дна створки в срединную септу. Ее высота составляет половину высоты брахиофория. Септа начинается от макушки, поддерживая брахиофорий на всем его протяжении, и продолжается за нижнюю границу брахиальных пластин вперед на треть длины раковины. Брахиальные пластины массивные, отогнутые к бокам раковины. Основания брахиальных отростков не дифференцируются. Крыловидные отростки отсутствуют. Брахиофорий сложен продольно-волокнистым веществом. Длина брахиофория меньше длины спондиальных образований в брюшной створке.
Размеры в мм (табл. 1).
Таблица 1.
Размеры целых экземпляров E. tozodoviensis sp. nov. и Е. lebediensis
| Экз. № | Длина (Д) | Ширина (Ш) | Толщина (Т) | Д/Ш | Т/Д | Т/Ш |
|---|---|---|---|---|---|---|
| 2110-1, голотип | 7.5 | 7.7 | 5.5 | 0.97 | 0.73 | 0.71 |
| 2110-2 | 8.3 | 7.8 | 4.7 | 1.06 | 0.57 | 0.60 |
| 2110-3 | 7.7 | 7.3 | 5.8 | 1.05 | 0.75 | 0.79 |
| 2110-4 | 8.1 | 7.7 | 5.5 | 1.05 | 0.68 | 0.71 |
| 2110-5 юная раковина | 5.5 | 4.6 | 3.1 | 1.20 | 0.56 | 0.67 |
| E. lebediensis, голотип, 403/1323 | 9 | 8.8 | 5.5 | 1.02 | 0.61 | 0.63 |
Сравнение. От типового вида E. antiquata (Никифорова, Сапельников, 1973, с. 66, табл. I , фиг. 1–7 ) из нижнеарчалыкских слоев ашгиллского яруса перевала Шахриомон Южного Тянь-Шаня описываемый новый вид отличается: изометричной формой раковины, овально-пентагональным очертанием (раковина E. antiquata вытянута по ширине); в два–три раза меньшими размерами раковин; отношение толщины спинной створки к толщине брюшной створки у E. tozodoviensis sp. nov. меньше этого параметра у E. antiquata; характеристиками замочного края: в поперечном профиле у E. tozodoviensis sp. nov. макушка брюшной створки находится на одном уровне по высоте с задним краем спинной створки (примакушечной областью), а макушка спинной створки направлена в сторону дельтирия брюшной створки, арея брюшной створки ортоклинная (в то время как у E. antiquata примакушечный край спинной створки превышает по высоте макушку брюшной створки); отсутствием ярко выраженного синуса и возвышения; взаимоотношением внутренних структур спинной и брюшной створок (у E. tozodoviensis sp. nov. спондилий почти вдвое длиннее, чем брахиофорий, у E. antiquata длина брахиофория, наоборот, превышает длину спондилия); колоколообразной формой спондилия (у E. antiquata – округло-треугольной формы); формой зубов: (у E. tozodoviensis sp. nov. каплевидные, у E. antiquata продолговатые); внутренним строением спинной створки: стабильным по выборке наличием высокой септы, поддерживающей брахиофорий и продолжающейся за границы брахиальных пластин (у E. antiquata септальные пластины либо сходятся на дне створки, либо тесно сближенные, либо сливаются в низкую септу); отсутствием крыловидных отростков (у E. antiquata наблюдаются зачатки крыловидных отростков или их отсутствие); простыми ребрами (у E. antiquata ребра дихотомируют); числом ребер: 14–16 у Е. tozodoviensis sp. nov. против 17–22 (в синусе 5–7, на возвышении 6–7, по сторонам 5–8) у E. antiquata.
От Е. kurdaica из кескинтасской свиты андеркенского горизонта среднего–верхнего карадока (зона Parastrophina hemiplicata) Южного Казахстана, хребта Кендыктас (Сапельников, Рукавишникова, 1975, с. 34, табл. XIX , фиг. 10–12 ), новый вид отличается: в среднем более мелкими раковинами, чем у Е. kurdaica (10–12 мм в длину); отсутствием синуса и возвышения, лишь унипликатной передней комисcурой (у Е. kurdaica синус и возвышение ярко выражены); отсутствием крыловидных отростков (у Е. kurdaica они хорошо развиты); простыми ребрами (у Е. kurdaica – слабо дихотомирующие); строением брахиофория: у Е. tozodoviensis sp. nov. высокая септа, поддерживающая брахиофорий, у Е. kurdaica тесно сближенные септальные пластины создают видимость слияния в низкую септу; взаимоотношением внутренних структур спинной и брюшной створок: у E. tozodoviensis sp. nov. спондилий длиннее, чем брахиофорий, почти вдвое, у E. kurdaica длина брахиофория соответствует длине спондилия.
От Е. transversa из орловского горизонта верхнего ордовика Северо-Запада Алтая (Севергина, 1978, с. 30, табл. IV , фиг. 1, 2 ) описываемый вид отличается: формой раковины, у Е. transversa – вытянутая по ширине; меньшими размерами (у Е. transversa 9–12 мм в длину, 13–19 мм в ширину); отсутствием синуса и возвышения (у Е. transversa они хорошо выражены), лишь изгибом переднего края в сторону спинной створки; характером строения примакушечной области: макушки створок не смыкаются, как у Е. transversa; меньшим числом ребер: 14–16 против 20 ребер у Е. transversa; простыми, нераздваивающимися ребрами (у Е. transversa ребра дихотомируют); высокой септой, поддерживающей брахиофорий. В описании внутренних элементов Е. transversa указано лишь принципиальное строение спондилия и брахиофория, сопровожденное одной фотографией пришлифовки плохого качества. Это затрудняет детальное сравнение параметров внутреннего строения нового вида и Е. transversa.
От близкородственного вида Е. primordialis (Розман, 1978, с. 100, табл. ХVII , фиг. 10–14 ) из шахриомонской свиты Южного Тянь-Шаня описываемый вид отличается: меньшими размерами; строением замочной области (макушка брюшной створки ортоклинная, не нависает над макушкой спинной створки); простыми, не дихотомирующими ребрами и меньшим их числом (14–16 против 24 у Е. primordialis); внутренним строением спинной створки: высокой дорсальной септой, поддерживающей брахиофорий от макушки на всем его протяжении и протягивающейся вперед за границу брахиальных пластин. У Е. primordialis только передний край брахиофория поддерживается низкой срединной септой. Заключение о различиях во внутреннем строении сделаны на основании описания Е. primordialis. В работе Х.С. Розман (1978) приведены фотографии ядер брюшной, спинной створок и целой раковины. Без пришлифовок и латексных слепков сложно оценить некоторые количественные и качественные параметры внутренних структур. В частности, непонятно, протягивается ли вентральная септа вперед за границы спондилия. При описании Е. primordialis в разделе “Замечание” Розман (1978) указывает на наличие в коллекции изменчивых форм – группы вытянутых раковин с немногочисленными ребрами, сходных с Camerella lebediensis Severgina, 1967. При возможности, следует изучить эти экземпляры на предмет отождествления с Е. tozodoviensis sp. nov.
В литературе имеется описание европейских эоанастрофий из трех различных местонахождений, относимых разными авторами к одному виду E. pentamera. Поскольку описания и фотоизображения разнятся друг с другом, авторы сочли необходимым провести сравнение нового вида с каждым из описанных под названием “E. pentamera (Meneghini, 1857)” (см. ниже).
От E. pentamera из карадока Черных Гор (Montagne Noire) Центрального массива Франции (Havlíček, 1981, с. 28, табл. 9 , фиг. 1–4 ) новый вид отличается: вдвое меньшими размерами; меньшим числом ребер (14–16 против 20–22); внутренним строением спинной створки: высокой дорсальной септой, поддерживающей брахиофорий от макушки на всем его протяжении и протягивающейся вперед за границу брахиальных пластин. У Е. pentamera септальные пластины рядом с макушкой не соприкасаются друг с другом, в то время как спереди они соединяются в низкую септу, поддерживающую переднюю часть брахиофория. Внутреннее строение брюшной створки и вентральная арея E. pentamera В. Гавличеком (Havlíček, 1981) не исследовались, а внутреннее строение спинной створки изучено им на ядре, т.к. коллекция состояла из двух экземпляров: одной целой раковины и ядра спинной створки. Поэтому невозможно провести полноценное сравнение Е. tozodoviensis sp. nov. с E. pentamera из Франции. Кроме того, Гавличек (Havlíček, 1981) указывает на возможную тождественность описываемых им экземпляров виду Е. primordialis.
От E. pentamera из формации Розан (Rosan) ашгиллского яруса, п-ова Крозон, северо-запад Франции (Melou, 1990, с. 556, табл. 10 , фиг. 1, 2, 5–12 ), новый вид отличается: формой раковины; меньшими размерами (втрое меньше самых крупных представителей E. pentamera); ортоклинной вентральной ареей (у E. pentamera апсаклинная); анаклинной дорсальной ареей (у E. pentamera апсаклинная); числом ребер [у нового вида их 14–16, у E. pentamera в описании указано 16–18; судя по фототаблицам (Melou, 1990, табл. 10 , фиг. 1, 7–12 ), их 20–22]; внутренним строением брюшной створки: септа, поддерживающая спондилий, не продолжается вперед за его границы, а у E. pentamera она продолжается вперед. В настоящее время это признак родового уровня (Williams et al., 2002, с. 955). Кроме того, у Е. tozodoviensis sp. nov. зубные пластины сливаются в септу в 5 мм по высоте от макушки, т.е. на начальных стадиях онтогенеза спондилий разобщенный. У E. pentamera, по данным M. Мелоу (Melou, 1990), спондилий поддерживается по всей длине тонкой срединной перегородкой. У нового вида отсутствует платформа на спинной створке, а у Е. pentamera мускульные отпечатки расположены на платформе. Высота дорсальной септы Е. pentamera не указана, внутреннее строение изучено на ядрах.
От E. pentamera, описанного в диссертации (Botquelen, 2003, с. 249, табл. 8 , фиг. 9 ), новый вид отличается: меньшими размерами (у E. pentamera длина равна 10 мм, ширина – 12 мм); формой раковины (у Е. tozodoviensis sp. nov. овально-пентагональное очертание, у E. pentamera субтреугольный контур); анаклинной дорсальной ареей (у E. pentamera – апсаклинная); одинаковым размером ребер (у E. pentamera срединное ребро в синусе и два ребра в средней части возвышения выделяются у половины общего числа экземпляров). Внешнее и внутреннее строение брюшной створки не сравнивалось, т.к. его описание в диссертации (Botquelen, 2003) отсутствует. В строении брахиофория не уточняется высота септы и продолжается ли она вперед за пределы брахиальных пластин.
От E. extenuata из саргалдакской свиты, еркебидаикского горизонта среднего ордовика хр. Чингиз (Казахстан, Тянь-Шань) (Кленина и др., 1984, с. 143, табл. XIX , фиг. 1, 2 , рис. 34 ) Е. tozodoviensis sp. nov. отличается: более крупными размерами (у E. extenuata длина – 5–7 мм, ширина – 6–8 мм); изометричной формой раковины (у E. extenuata вытянута по ширине); невыраженными синусом и седлом, лишь изгибом лобного края в сторону спинной створки, тогда как у E. extenuata синус начинается от макушки, четко обосабливается в передней половине, возвышение развито со средней части створки, язычок невысокий; внутренним строением брюшной створки: формой спондилия (у Е. tozodoviensis sp. nov. колоколообразной формы, у E. extenuata – параболовидной); у нового вида септа не продолжается вперед за границы спондилия, у E. extenuata – продолжается; внутренним строением спинной створки: у Е. tozodoviensis sp. nov. брахиофорий поддерживается высокой септой на всем протяжении, у E. extenuata сидячий брахиофорий, на небольшом участке в середине длины поддерживаемый короткой септой (Кленина и др., 1984, рис. 34 ).
От E. lebediensis из нижней части гурьяновской свиты ханхаринского горизонта среднего–верхнего ордовика Уйменско-Лебедской СФЗ Горного Алтая (Севергина, 1967, с. 131, табл. 4 , фиг. 4, 5 ; Кульков, Севергина, 1989, с. 151, табл. X , фиг. 19 ) новый вид отличается: меньшими размерами; очертанием раковины (у E. tozodoviensis sp. nov. – пентагональное, у E. lebediensis – треугольно-пентагональное); характером выпуклости створок (у E. tozodoviensis sp. nov. створки равномерно выпуклые, у E. lebediensis выпуклость довольно резко уменьшается по направлению к переднему краю); числом ребер (14–16 ребер на каждой створке против 10–12).
Кроме того, установлен ряд параметров E. tozodoviensis sp. nov., неизвестных для E. lebediensis в силу степени сохранности и методики изучения внутреннего строения последнего: строение замочного края E. tozodoviensis sp. nov. – макушка брюшной створки находится на одном уровне по высоте с задним краем спинной створки (примакушечной областью), макушка спинной створки направлена в сторону дельтирия брюшной створки, арея брюшной створки ортоклинная. Внутреннее строение брюшной створки E. tozodoviensis sp. nov.: определен тип спондилия, спондилий-комбинариум (=сложный, комбинированный, simplex); форма спондилия в сечении колоколообразная; на ранней стадии онтогенеза спондилиум разобщенный, затем, с расстояния 0.5–0.9 мм от макушки, поддерживается низкой срединной септой; срединная септа увеличивается по высоте в направлении передней комиссуры; срединная септа не продолжается вперед за границы спондилия. Внутреннее строение спинной створки E. tozodoviensis sp. nov.: высота дорсальной срединной септы составляет половину высоты брахиофория, брахиофорий Y-образной формы.
Несмотря на имеющиеся отличия во внешнем и внутреннем строении, E. lebediensis и E. tozodoviensis sp. nov. составляют группу таксонов с близкими морфологическими признаками.
Материал. Шесть целых раковин, 15 разрозненных створок из Прителецкой СФЗ, разреза Тозодов (шестая, седьмая, восьмая пачки), п.т. Нижний Турочак (С-163) (Сенников и др., 2018; Sennikov et al., 2019).
ОБСУЖДЕНИЕ
По внутреннему строению новый вид имеет все характерные признаки рода Еoanastrophia: сложный спондилий, поддерживающийся короткой септой, которая не продолжается за границы спондилия; в спинной створке двойной брахиофорий. От ранее известных эоанастрофий он отличается высокой дорсальной септой, составляющей половину высоты брахиофория. В родовом диагнозе Еoanastrophia строение брахиофория описывается как: “…брахиофорий глубокий, V-образный; септальные пластины тонкие, высокие, сходящиеся у дна створки…” (Никифорова, Сапельников, 1973; Сапельников, Рукавишникова, 1975); “…приямочные пластины сходятся, формируя глубокий узкий септалиум-симплекс, поддерживаемый длинной низкой срединной септой…” (Williams et al., 2002; перевод наш). По наличию у нового вида высокой дорсальной септы следует расширить понимание объема рода: “Брахиофорий глубокий, V-образной формы, либо поддерживаемый низкой или высокой длинной срединной септой”.
По внешнему строению новый вид E. tozodoviensis sp. nov. также соответствует параметрам рода Еoanastrophia: раковины ребристые от макушки, спинная створка более выпуклая, чем брюшная. Специфика нового вида E. tozodoviensis sp. nov. – наличие простых не раздваивающихся ребер, отсутствие развитого синуса и возвышения, лишь слабый изгиб передней комиссуры.
ЗАКЛЮЧЕНИЕ
Изучены представители рода Eoanastrophia из двух местонахождений верхнего ордовика Прителецкой СФЗ Горного Алтая. Сравнительный анализ с типовой коллекцией близкородственного алтайского вида E. lebediensis из Уйменско-Лебедской СФЗ выявил необходимость описания нового таксона – E. tozodoviensis sp. nov. Хорошая сохранность и достаточное количество экземпляров выделенного вида позволили детально изучить внутреннее строение и доказать стабильность признака высокой септы в его спинной створке. Таким образом, род Eoanastrophia включает в себя виды не только с низкой дорсальной септой или V-образным сидячим брахиофорием, но и новый вид с высокой спинной септой, формирующей брахиофорий Y-образной формы.
* * *
Работа выполнена в рамках госзадания РАН по Фундаментальным научным исследованиям (проект FW ZZ-2022-0003). Изученная коллекция прителецких алтайских эоанастрофий хранится в ЦКП “ГЕОХРОН” ИНГГ СО РАН в Новосибирске под № 2110. Авторы благодарят Д.А. Ильина, Е.В. Лыкову, О.Т. Обут, Д.А. Токарева, Р.А. Хабибулину за помощь при сборе изученных коллекций брахиопод.
Список литературы
Андреева О.Н. Среднеордовикские брахиоподы Тувы и Алтая // Палеонтол. журн. 1982. № 2. С. 52–61.
Андреева О.Н. Среднеордовикские брахиоподы и трилобиты Тувы и Алтая // Палеонтол. журн. 1985. № 2. С. 38–47.
Кленина Л.Н., Никитин И.Ф., Попов Л.Е. Брахиоподы и биостратиграфия среднего и верхнего ордовика хребта Чингиз. Алма-Ата: Наука, 1984. 196 с.
Кульков H.П., Севергина Л.Г. Вопросы корреляции и стратиграфии верхнего ордовика Горного Алтая // Палеонтология и биостратиграфия палеозоя Сибири. Новосибирск: Наука, 1984. С. 32–38.
Кульков Н.П., Севергина Л.Г. Стратиграфия и брахиоподы ордовика и нижнего силура Горного Алтая. Новосибирск: Наука, 1989. 223 с.
Никифорова О.И., Сапельников В.П. Некоторые древние пентамериды Зеравшанского хребта // Сборник по вопросам стратиграфии. № 18. Свердловск, 1973. С. 64–88 (Тр. Ин-та геол. и геохим. УНЦ СССР. Вып. 99).
Розман Х.С. Брахиоподы обикалонских слоев // Пограничные слои ордовика и силура Алтае-Саянской области и Средней Азии. М.: Наука, 1978. С. 75–101.
Сапельников В.П. Морфология и таксономическая эволюция брахиопод (отряд пентамериды). Свердловск, 1985. 231 с.
Сапельников В.П., Рукавишникова Т.Б. Верхнеордовикские, силурийские и нижнедевонские пентамериды Казахстана. М.: Наука, 1975. 227 с.
Севергина Л.Г. Брахиоподы ордовика // Биостратиграфия палеозоя Саяно-Алтайской горной области. Т. 1. Новосибирск, 1960. С. 400–409.
Севергина Л.Г. Новые виды и роды ордовикских брахиопод Саяно-Алтайской горной области // Учен. зап. Томск. ун-та. 1967. Вып. 63. С. 120–146.
Севергина Л.Г. Брахиоподы и стратиграфия верхнего ордовика Горного Алтая, Салаира и Горной Шории // Фауна и биостратиграфия верхнего ордовика и силура Алтае-Саянской области. М.: Наука, 1978. С. 3–41.
Сенников Н.В., Обут О.Т., Гонта Т.В. и др. Ордовикские фаунистические ассоциации и осадочные комплексы Прителецкой части Горного Алтая // Тр. Палеонтол. об-ва. Т. I. М.: ПИН РАН, 2018. С. 134–147.
Botquelen A. Impact des variations eustatiques sur les assemblages benthiques à brachiopodes: l’Ordovicien sarde et le Dévonien Ibéro-armoricain. Paléoécologie–Taphonomie–Stratigraphie séquentielle–Systématique. Unpubl. PhD Thesis. Brest: Univ. de Bretagne Occidentale, 2003. 325 p.
Havlíček V. Upper Ordovician brachiopods from the Montagne Noire // Palaeontogr. Abt. A. 1981. V. 176. P. 1–34.
Mélou M. Brachiopodes articulés de la coupe de l’Ile de Rosan (Crozon, Finistère), Formation des Tufs et Calcaires de Rosan (Caradoc-Ashgill) // Geobios. 1990. № 23. Fasc. 5. P. 539–579.
Sennikov N.V., Obut O.T., Lykova E.V. et al. Ordovician sedimentary basins and paleobiotas of the Gorny Altai. Novosibirsk: Publ. House of SB RAS, 2019. 184 p.
Shcherbanenko T.A. Brachiopods from Upper Ordovician sections northeast of the Gorny Altai (Teletskoe Lakeside area) // 13th Intern. Symp. on the Ordovician System: Contributions of Intern. Symp. (Novosibirsk, Russia, July 19–22, 2019). Novosibirsk: Publ. House of SB RAS, 2019. P. 189–191.
Williams A., Bruton C.H.C., Carlson S.J. (Eds). Treatise on Invertebrate Paleontology: Part H, Brachiopods. V. 4. Lawrence, Kans.: Univ. of Kansas Press, 2002. P. 921–1688.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал