

Палеонтологический журнал, 2022, № 6, стр. 14-19
Новые раннепермские наутилиды из семейства Rhiphaeoceratidae рифа Шахтау (Башкортостан)
А. Ю. Щедухин *
Палеонтологический институт им. А.А. Борисяка РАН
117647 Москва, Россия
* E-mail: d_alsch2017@mail.ru
Поступила в редакцию 17.04.2022
После доработки 06.05.2022
Принята к публикации 06.05.2022
- EDN: ALXJOT
- DOI: 10.31857/S0031031X22060095
Аннотация
Рассмотрена история изучения семейства Rhiphaeoceratidae и приведены сведения о стратиграфическом и географическом распространении его представителей. В составе Rhiphaeoceratidae описаны новый род Eximioceras с типовым видом E. venustum sp. nov. и новый вид Pararhiphaeoceras parvum, также относящийся к этому семейству. Находки новых форм расширяют наши представления о таксономическом разнообразии рифового сообщества цефалопод Шахтау и дополняют его экологическую характеристику.
ВВЕДЕНИЕ
Первые представители Rhiphaeoceratidae Ruzhencev et Shimansky, 1954 были обнаружены в середине XX в. В.Е. Руженцевым и В.Н. Шиманским (1954) в сакмарских и артинских отложениях Казахстана. В составе нового семейства авторами было описано четыре рода: Rhiphaeoceras (два вида), Pararhiphaeoceras (три вида), Sholakoceras (три вида) и Rhiphaeonautilus (один вид). В качестве характерных особенностей этого семейства была указана скульптура из наклонных ребер, расположенных на латеральных сторонах раковины, перегородочная линия с широким вентральным седлом, иногда осложненным небольшой лопастью, и глубокая воронкообразная лопасть на дорсальной стороне. Строение дорсальной части перегородочной линии значительно отличает рифеоцератид от других известных пермских наутилид. По мнению Руженцева и Шиманского (1954), наиболее близким к рифеоцератидам является семейство Aktubonautilidae Ruzhencev et Shimansky, 1954. Это семейство было описано по двум выделенным ими родам: Aktubonautilus Ruzhencev et Shimansky, 1954 (единственный экземпляр из артинского яруса Казахстана) и Basleonautilus Ruzhencev et Shimansky, 1954 [описан К. Ганиэлем (Haniel, 1915) как Pleuronautilus dyadicus Haniel, 1915 из слоев Баслео (средняя пермь) о-ва Тимор]. Сходный по скульптуре раковины и общей форме перегородочной линии с рифеоцератидами, Aktubonautilus отличается строением эмбриональной раковины – крупным первым оборотом, а также более широкой и менее глубокой дорсальной лопастью.
Спустя почти десятилетие в сводке “Основы палеонтологии” (Шиманский, 1962) Rhiphaeoceratidae были объединены с Aktubonautilidae в надсемейство Rhiphaeocerataceae Ruzhencev et Shimansky, 1954. К рифеоцератидам в этой работе был отнесен род Tungkuanoceras Hayasaka, 1947. Зарубежные исследователи (Kummel, 1964) признали самостоятельность семейства Rhiphaeoceratidae, но не Actubonautilidae. Роды, входящие в состав актюбонаутилид, и род Tungkuanoceras из рифеоцератид были сведены в синонимику, а сами рифеоцератиды отнесены к надсемейству Tainocerataceae Hyatt, 1883 (Kummel, 1964). В конце 70-х гг. прошлого века Шиманский (1979) описал новый вид Pararhiphaeoceras probum Shimansky, 1979 из верхней перми Азербайджана. Долгое время никакой новой информации об этом семействе не появлялось. Только спустя 40 лет были описаны новые находки рифеоцератид из Южноуральского местонахождения Шахтау (Leonova, Shchedukhin, 2020; Shchedukhin, Leonova, 2020). В ассельско-сакмарском интервале были определены Pararhiphaeoceras tastubense Ruzhencev et Shimansky, 1954, Pararhiphaeoceras sp. и новый вид Sholakoceras formosum Leonova et Shchedukhin, 2020. В верхнеартинских отложениях этого местонахождения был установлен только один вид – Pararhiphaeoceras aktastense Ruzhencev et Shimansky, 1954. Таким образом, в настоящее время известно 11 видов рифеоцератид всего из двух регионов – с Южного Урала и из Закавказья, все – исключительно из пермских отложений.
МАТЕРИАЛ И МЕТОДИКА
Изученный материал происходит из нижнепермских отложений карьера Шахтау. Местонахождение расположено в республике Башкортостан, в 5 км на восток от г. Стерлитамак. Описываемая часть коллекции была собрана весной 2021 г. из крупных блоков брахиоподово-мшанкового пакстоуна из стенки первого уступа (высотные отметки 127–140 м) в южной части карьера. Согласно геологическим профилям (Mazaev, 2019), эта часть разреза относится к ассельско–сакмарскому интервалу.
Изученный материал представлен 13 экз. хорошей сохранности. На многих из них сохранилась раковина, а на некоторых – устьевой край, что позволяет подробно описать форму гипономического синуса. Описание раковин наутилид производилось по методике, принятой для этой группы отечественными исследователями (Руженцев, Шиманский, 1954). При характеристике общей формы раковины использовалась уже устоявшаяся терминология, применяемая к аммоноидеям (Руженцев, Богословская, 1971). Коллекция хранится в лаб. моллюсков Палеонтологического ин-та им. А.А. Борисяка РАН (ПИН РАН), колл. № 5668. В таблицах измерений приняты следующие сокращения: Д – диаметр раковины, В – высота оборота, Ш – ширина оборота, Ду – диаметр умбилика/умбиликального отверстия (для первого оборота).
Автор искренне признателен А.В. Мазаеву – организатору полевых работ на шихане Шахтау, а также рецензентам статьи за ценные и конструктивные замечания, которые позволили улучшить ее содержание.
Работа поддержана грантом РНФ № 22-24-00099 “Эволюция сообществ моллюсков раннепермского рифа Шахтау”.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ПОДКЛАСС NAUTILOIDEA
ОТРЯД NAUTILIDA
НАДСЕМЕЙСТВО TAINOCERATOIDEA HYATT, 1883
СЕМЕЙСТВО RHIPHAEOCERATIDAE RUZHENCEV ET SHIMANSKY, 1954
Род Pararhiphaeoceras Ruzhencev et Shimansky, 1954
Pararhiphaeoceras parvum Shchedukhin, sp. nov.
Табл. III , фиг. 1, 2
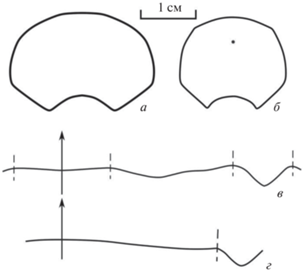
Рис. 1.
Поперечные сечения взрослых оборотов и перегородочные линии новых представителей Rhiphaeoceratidae: а, г – Pararhiphaeoceras parvum sp. nov.; б, в – Eximioceras venustum gen. et sp. nov.
Название вида от parvus лат. – маленький (о размере первого оборота).
Голотип – ПИН, № 5668/5-8; Россия, Башкортостан, шихан Шахтау; нижняя пермь, верхнеассельские–нижнесакмарские отложения.
Объяснение к таблице III
Фиг. 1, 2 . Pararhiphaeoceras parvum sp. nov.: 1 – голотип ПИН, № 5668/5-8: 1а – с вентральной стороны, 1б – со стороны устья, 1в – с латеральной стороны; 2 – экз. ПИН, № 5668/5-11, гипономический синус.
Фиг. 3, 4 . Eximioceras venustum gen. et sp. nov.: 3 – экз. ПИН, № 5668/5-3, гипономический синус; 4 – голотип ПИН, № 5668/5-1: 4а – с латеральной стороны, 4б – со стороны устья, 4в – с вентральной стороны.
Все: Башкортостан, Шахтау; нижняя пермь, пограничные ассельско–сакмарские отложения.

Описание. Форма (табл. III , фиг. 1, 2 ). Раковина эволютная, пахиконовая; состоит из 2.5–3 оборотов. Обороты возрастают заметно больше в ширину, чем в высоту (во взрослом состоянии Ш/В = 1.66). Первый оборот маленький, диаметром 10.1 мм. Умбиликальное отверстие очень маленькое – 4.1–4.7 мм. Поперечное сечение в онтогенезе изменяется от округлого, на протяжении первого оборота, до почковидного – с начала второго оборота и округло-октогонального – во взрослом состоянии. Дорсальный желобок появляется с первой трети второго оборота. Вентральная сторона широкоокругленная. Вентролатеральный край сглаженный. Боковые стороны немного уплощенные, сходятся к вентральной стороне. Умбиликальный перегиб отчетливый. Умбиликальная стенка слабовыпуклая, с плоскостью симметрии образует угол около 55°. Дорсальная сторона вогнута примерно на 1/5 высоты оборота. Умбилик среднего размера (Ду/Д = = 0.37), ступенчатый. У раковин с сохранившимся устьем на вентральной стороне расположен глубокий U-образный гипономический синус, равный примерно трети ширины всей вентральной стороны.
Размеры в мм и отношения:
| Экз. ПИН № | Д | В | Ш | Ду | В/Д | Ш/Д | Ду/Д | Ш/В |
|---|---|---|---|---|---|---|---|---|
| Голотип 5668/5-8 | 38.4 | 15.5 | 25.8 | 14.5 | 0.4 | 0.67 | 0.37 | 1.66 |
| 10.1 | 3 | 7 | 4.1 | 0.29 | 0.69 | 0.4 | 2.3 | |
| 5668/5-9 | 38.8 | 15 | 22.5 | 15 | 0.38 | 0.58 | 0.38 | 1.5 |
| 10.7 | 3.2 | 5.7 | 4.7 | 0.29 | 0.53 | 0.43 | 1.78 | |
| 5668/5-10 | 40.5 | 15.3 | 25.4 | 18 | 0.37 | 0.62 | 0.44 | 1.66 |
| 11 | 2.7 | 5.3 | 4.7 | 0.24 | 0.48 | 0.42 | 1.96 | |
| 5668/5-11 | 46.1 | 11.5 | 18.5 | 16.9 | 0.24 | 0.4 | 0.36 | 1.6 |
| 10 | 3.1 | 5.9 | 4.3 | 0.31 | 0.59 | 0.43 | 1.9 |
Перегородки вогнуты примерно на высоту 1.5 газовых камер. На вторую половину последнего видимого оборота приходятся около 12 камер. На величину, равную ширине оборота, приходятся четыре камеры.
Скульптура на первом полуобороте представлена тонкими прямыми струйками нарастания. С третьей четверти первого оборота на вентральной стороне возникают овальные бугорки, которые быстро сменяются наклонными ребрышками. С возрастом ребра становятся еще более отчетливыми и немного изогнутыми по направлению от устья, лишь ближе к началу третьего оборота они слегка сглаживаются. На одну камеру приходятся одно–два ребра. На последнем видимом обороте наблюдаются 24 ребра.
Перегородочная линия слабо расчлененная (рис. 1, г). На вентральной стороне расположено очень широкое, слабо выраженное седло, которое на латеральной и умбиликальной стороне переходит в практически такую же широкую лопасть. Внутреннее седло узкое. Дорсальная лопасть V‑образная, угол между ее боковыми сторонами от 85° до 95°.
Сифон смещен от центра к вентральной стороне на величину 0.4 высоты оборота от нее.
Сравнение. От всех других видов этого рода отличается значительно меньшим размером первого оборота (10.1 против 14–18 мм). По внешнему облику и очертаниям перегородочной линии P. parvum sp. nov. напоминает P. tastubense Ruzhencev et Shimansky, 1954, но между этими видами имеются и существенные отличия: высота камер у P. parvum меньше (на ширину оборота приходятся 3.5–4 камеры против 2–2.5 камер у P. tastubense), а дорсальная лопасть у P. tastubense более глубокая. Различается и характер скульптуры: у последнего ребра на раковине имеют заметно больший наклон, чем у P. parvum. От P. aktastense Ruzhencev et Shimansky, 1954 новый вид отличается формой поперечного сечения с менее выраженным вентролатеральным краем и более отчетливым умбиликальным перегибом. Наиболее близок к новому виду P. incallidum Ruzhencev et Shimansky, 1954 из артинских отложений Казахстана. Сходство состоит в близких соотношениях В/Д, Ш/Д, Ду/Д, а также положении сифона. Тем не менее, оба вида четко различаются между собой: для P. parvum sp. nov. характерна менее расчлененная внешняя часть перегородочной линии – в ней вентральное седло выражено слабее и не осложнено лопастью в вершине, а боковая лопасть почти не выражена. Кроме этого, у P. incallidum форма поперечного сечения со значительным преобладанием ширины над высотой (Ш/В = 1.6 вместо Ш/В = 1.39 у нового вида). От позднепермского P. probum новый вид отличается положением сифона, смещенного к вентральной стороне, и формой ребер на латеральных сторонах, которые на начальных оборотах не осложнены бугорком на вентральном крае.
Замечание. При изучении экземпляров нового вида с сохранившейся жилой камерой на вентральной стороне ядра выявлено очень узкое ребрышко, проходящее ровно по плоскости симметрии. Функциональное значение этой структуры не совсем ясно.
Материал. Семь экз. из верхнеассельских–нижнесакмарских известняков шихана Шахтау, Башкортостан. Сборы автора.
Род Eximioceras Shchedukhin, gen. nov.
Название рода от eximia лат. – исключительный.
Типовой вид – Eximioceras venustum sp. nov.
Диагноз. Раковина эволютная, субплатиконовая. Первый оборот небольшой, около 15 мм. Умбиликальное отверстие средних размеров (6 мм). Поперечное сечение во взрослом состоянии гексагональное, с небольшим преобладанием ширины над высотой. Умбилик широкий, ступенчатый. Скульптура представлена слабоизогнутыми ребрышками на боковых сторонах, каждое из них имеет утолщения на краевых частях. Край ребрышка, расположенный на умбиликальном перегибе, имеет на своем конце приподнятое серповидное утолщение. Другой край ребра, расположенный ближе к вентролатеральному перегибу, оканчивается округлым бугорком. Перегородочная линия извилистая, с очень широким вентральным седлом, широкой латеральной лопастью и узким умбиликальным седлом. Дорсальная лопасть V-образная, широкая и глубокая. Угол между сторонами дорсальной лопасти составляет от 90° до 105°.
Видовой состав. Типовой вид из Башкортостана, шихан Шахтау; нижняя пермь, верхнеассельские–нижнесакмарские отложения.
Сравнение. От других родов этого семейства отличается гексагональной формой поперечного сечения и характерной скульптурой в виде слегка наклонных, изогнутых ребрышек, с асимметричными утолщениями на краевых частях.
Eximioceras venustum Shchedukhin, sp. nov.
Табл. III , фиг. 3, 4
Название вида от venustus лат. – прелестный.
Голотип – ПИН, № 5668/5-1; Башкортостан, шихан Шахтау; нижняя пермь, верхнеассельские–нижнесакмарские отложения.
Описание. Форма (табл. III , фиг. 3, 4 ). Раковина эволютная, субплатиконовая; во взрослом состоянии состоит из 2.5–3 оборотов. Обороты возрастают в ширину немного больше, чем в высоту (Ш/В = 1.3). Первый оборот небольшой, в диаметре достигает 15 мм. Умбиликальное отверстие около 6.5 мм. Поперечное сечение на протяжении первого полуоборота округлое. С третьей четверти первого оборота начинают обособляться вентральный и умбиликальный края, поперечное сечение становится близким к овальному. Дорсальный желобок возникает со второй половины первого оборота, а поперечное сечение становится гексагональным (рис. 1, б). Вентральная сторона выпуклая, слегка уплощенная, на ней расположен широкий и глубокий U-образный гипономический синус. Вентральный край скругленный, но отчетливый. Боковые стороны слабовыпуклые. Умбиликальный перегиб резкий, узко-округленный. Умбиликальная стенка слабовогнутая, с плоскостью симметрии образует угол около 50°. Дорсальная стенка вогнута чуть больше, чем на одну шестую высоты оборота. Умбилик средних размеров (Ду/Д = 0.35–0.39), ступенчатый.
Размеры в мм и отношения:
| Экз. ПИН № | Д | В | Ш | Ду | В/Д | Ш/Д | Ду/Д | Ш/В |
|---|---|---|---|---|---|---|---|---|
| Голотип 5668/5-1 | 43.5 | 17.4 | 22.5 | 18.2 | 0.4 | 0.51 | 0.41 | 1.29 |
| 15.7 | 5.8 | 8 | 6.6 | 0.36 | 0.5 | 0.42 | 1.37 | |
| 5668/5-2 | 35 | 14.3 | 17.7 | 14.6 | 0.4 | 0.5 | 0.41 | 1.24 |
| 5668/5-3 | 47.5 | 19 | 24.7 | 16.9 | 0.4 | 0.52 | 0.35 | 1.3 |
| 14.4 | 4.2 | 7.2 | 6 | 0.29 | 0.5 | 0.41 | 1.71 | |
| 5668/5-4 | 45.2 | 18.3 | 22.7 | 19.9 | 0.4 | 0.5 | 0.44 | 1.24 |
| 15.2 | 5.2 | 8.4 | 5.3 | 0.34 | 0.55 | 0.34 | 1.61 | |
| 5668/5-5 | 39.1 | 16.5 | 18.4 | 13.6 | 0.42 | 0.47 | 0.34 | 1.11 |
| 14.3 | 4.9 | 8 | 5.8 | 0.34 | 0.55 | 0.4 | 1.6 | |
| 5668/5-6 | 32.7 | 13.2 | 14.7 | 14.2 | 0.4 | 0.44 | 0.43 | 1.11 |
Поверхность раковины покрыта тонкими струйками нарастания. На первом полуобороте струйки слегка наклонены назад. Со второй половины первого оборота характер струйчатости изменяется. На вентральной стороне струйки формируют широкий и глубокий синус, переходящий в широкоокругленный изгиб вперед на вентролатеральном крае. На умбиликальном крае расположен узкоокругленный синус. На умбиликальной стенке струйки образуют небольшой изгиб вперед.
Со второй половины первого оборота на латеральных сторонах появляются отчетливые наклонные ребрышки. С первой половины второго оборота ребрышки становятся более отчетливыми, с характерными асимметричными бугорками на краях. Край ребрышка, приближенный к вентролатеральному перегибу, оканчивается округлым бугорком, приподнятым над поверхностью ребра и слегка скошенным по направлению к устью. На другой оконечности ребра, на умбиликальном перегибе, бугорок имеет серповидную форму и выражен еще сильнее. На одну камеру приходится одно ребро. С возрастом ребрышки становятся более сглаженными. На последнем обороте насчитываются 28 ребер.
Перегородки вогнуты примерно на 1.5–2 камеры. Камеры низкие. На ширину поперечного сечения приходятся около четырех камер. На вторую половину последнего видимого оборота приходятся 14 камер.
Перегородочная линия слабоизвилистая (рис. 1, в). Широкое вентральное седло разделено неглубокой лопастью. Латеральная лопасть широкая. На умбиликальной поверхности перегородочная линия практически прямая. Дорсальная лопасть V-образная, глубокая, угол между ее боковыми сторонами около 90°–105°.
Сифон узкий, смещен от центра к вентральной стороне на 0.39 высоты оборота от нее.
Материал. Голотип и пять экз. из верхнеассельских–нижнесакмарских известняков шихана Шахтау, Башкортостан. Сборы автора.
Список литературы
Руженцев В.Е., Богословская М.Ф. Намюрский этап в эволюции аммоноидей. Ранненамюрские аммоноидеи. М.: Наука, 1971. 382 с. (Тр. Палеонтол. ин-та АН СССР. Т. 133).
Руженцев В.Е., Шиманский В.Н. Нижнепермские свернутые и согнутые наутилоидеи Южного Урала. М.: Наука, 1954. С. 1–150 (Тр. Палеонтол. ин-та АН СССР. Т. 50).
Шиманский В.Н. Отряд Nautilida // Основы палеонтологии. Моллюски-головоногие. I / Ред. В.Е. Руженцев. М.: Изд-во АН СССР, 1962. С. 115–154.
Шиманский В.Н. Новые наутилоидеи из верхнего палеозоя Закавказья // Бюлл. Моск. об-ва испыт. прир. Отд. геол. 1979. Т. 54. № 3. С. 54–61.
Haniel C.A. Die Cephalopoden der Dyas von Timor // Paläontol. Timor. 1915. Lfg. 3. Abh. 6. 153 s.
Kummel B. Nautiloidea–Nautilida // Treatise on Invertebrate Paleontology. Pt K. Mollusca 3. Lawrence: Univ. Kansas Press, 1964. P. K383–K457.
Leonova T.B., Shchedukhin A.Yu. Asselian-Sakmarian nautiloids of the Shakh-Tau reef (Bashkortostan) // Paleontol. J. 2020. V. 54. № 10. P. 1113–1134.
Mazaev A.V. Lower Permian gastropods of Shakhtau (Asselian–Sakmarian boundary beds, Southern Cisuralia) // Paleontol. J. 2019. V. 53. № 12. P. 1237–1345.
Shchedukhin A.Yu., Leonova T.B. Late Artinskian nautiloids of the Shakh-Tau reef (Bashkortostan) // Paleontol. J. 2020. V. 54. № 10. P. 1135–1151.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал