

Палеонтологический журнал, 2020, № 6, стр. 55-59
Первая находка сетчатокрылых насекомых (Neuroptera: Mesochrysopidae) в нижнем мелу Южной Кореи
А. В. Храмов a, *, Г. С. Нам b
a Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
b Национальный университет образования
Конджу, Республика Корея
* E-mail: a-hramov@yandex.ru
Поступила в редакцию 22.12.2019
После доработки 09.01.2020
Принята к публикации 09.01.2020
Аннотация
Из нижнемеловой формации Чинджу (Южная Корея) описывается новый представитель семейства Mesochrysopidae (Neuroptera), Tachinymphes koraiensis sp. nov., который является первым ископаемым сетчатокрылым насекомым, найденным на территории Корейского полуострова. Обсуждается эволюционная история семейства Mesochrysopidae, высказывается предположение, что его расцвет в раннем мелу был связан с мирмелеонтоидными модификациями жилкования крыльев, которые позволили улучшить летные качества этих насекомых. Приводится предварительный обзор водных и наземных насекомых, найденных в формации Чинджу, с примерной оценкой относительной численности различных отрядов.
Мезохризопиды – одни из наиболее ярких и узнаваемых представителей мезозойских сетчатокрылых. Вид Mesotermes heros (Hagen, 1862), относящийся к мезохризопидам, был в числе первых ископаемых сетчатокрылых, ставших достоянием науки, несмотря на то, что немецкий энтомолог Г. Хаген, описавший его из верхнеюрских зольнхофенских известняков, первоначально принял это насекомое за термита (Hagen, 1862). Древнейшие мезохризопиды были найдены в нижней юре Люксембурга (Nel, Henrotay, 1994), последняя по возрасту находка этого вымершего семейства происходит из мелового бирманского янтаря (Liu et al., 2016).
Хотя мезохризопид обычно сближают со златоглазками (Chrysopidae) (Nel et al., 2005; Makarkin, Menon, 2005), их жилкование, особенно если речь идет о меловых таксонах, организовано по мирмелеонтоидному типу. Как и у нитекрылок (Nemopteridae), муравьиных львов (Myrmeleontidae) и аскалафид (Ascalaphidae), у типичных мезохризопид в переднем крыле имеется “треугольник”, образованный сближением или слиянием жилок MP и CuA. Но, в отличие от мирмелеонтоидов, у которых MP1 остается свободной, у мезохризопид в состав этого “треугольника” входят обе ветви MP. Кроме того, у многих мезохризопид в крыльях имеются ряды поперечных жилок, выстроенных в единую линию, которые очень напоминают так называемые линии Банкса – продольные складки на крыльях муравьиных львов.
Неудивительно, что некоторых мезохризопид относили к мирмелеонтоидам (Пономаренко, 1992; см. также Engel, Grimaldi, 2007, где мезохризопида Mesascalaphus yangi Ren et al., 2007 фигурирует в списке ископаемых аскалафид), хотя, по-видимому, сходство в их жилковании является результатом конвергентной эволюции. Стоит отметить, что юрские роды Protoaristenymphes Nel et Henrotay, 1994, Macronympha Panfilov, 1980, Aristenymphes Panfilov, 1980, которые считаются древнейшими представителями мезохризопид (Yang et al., 2012), еще не демонстрируют мирмелеонтоидных черт – MP и CuA у этих таксонов не сближены, а вместо многочисленных рядов поперечных жилок, некоторые из которых образуют подобие линий Банкса, в крыльях юрских мезохризопид имеется всего два ступенчатых ряда поперечных жилок, что больше напоминает жилкование златоглазок.
Впервые мезохризопиды с мирмелеонтоидными чертами появляются в верхней юре Зольнхофена, что явствует из жилкования вида Mesochrysopa zitteli (Meunier, 1898), который сохранился лучше, чем вышеупомянутый Mesotermes heros из того же местонахождения. В раннем мелу мезохризопиды становятся очень распространенной и весьма многочисленной группой – например, в испанском меловом местонахождении Лас-Ойас они абсолютно преобладают среди сетчатокрылых, также представители этого семейства обильны в нижнемеловых отложениях формации Крато в Бразилии (Makarkin et al., 2012). Напротив, в юре мезохризопиды были крайне редки – например, только один отпечаток этих насекомых встретился в ходе просмотра коллекции из 2000 сетчатокрылых, найденных в средней юре Даохугоу (Китай) (Yang et al., 2012). Из почти 550 сетчатокрылых, найденных в верхней юре Каратау (Казахстан), к мезохризопидам относится только восемь экземпляров (перс. набл.).
Чтобы лучше понять эволюцию мезохризопид, необходимо обратиться к их современным аналогам – к мирмелеонтоидам. Хотя Джеральд Даррелл называл муравьиных львов “пьяными стрекозами” за неустойчивый, дергающийся полет, в целом мирмелеонтоиды превосходят остальных сетчатокрылых по своим летным качествам. Например, аскалафиды в полете очень маневренны и занимают ту же экологическую нишу, что и стрекозы, хватая добычу прямо в воздухе. С улучшенными летными характеристиками мирмелеонтоидов напрямую коррелируют особенности их крыльев, которые имеют вытянутую форму и сужены к основанию, а также укреплены дополнительными центрами жесткости, такими как линии Банкса и слившиеся CuA и MP. Муравьиные львы – крупнейшее семейство современных сетчатокрылых, вместе с аскалафидами они насчитывают более 2100 видов (Machado et al., 2018), что составляет около трети всего видового разнообразия отряда. Очевидно, что более эффективный полет, необходимый для расселения, поиска добычи и подходящих мест для откладки яиц, был одним из факторов, предопределивших эволюционный успех мирмелеонтоидов.
Вполне возможно, расцвет мезохризопид в первой половине мелового периода объясняется этими же причинами, а именно, появлением форм с более продвинутыми крыльями мирмелеонтоидного типа. О значении летных качеств для эволюции мезохризопид говорит тот факт, что они “экспериментировали” со строением крыльев активнее многих других сетчатокрылых. Если не считать вымерших Dipteromantispidae и некоторых современных Hemerobiidae, мезохризопиды были единственными сетчатокрылыми, кто перешел к практически полной двукрылости – у Allopterus luianus Zhang, 1991 и A. mayorgai Nel et al., 2005 из нижнего мела Китая и Испании, соответственно, задние крылья укоротились по сравнению с передними более чем в 3 раза, а у Pedanoptera arachnophila Liu et al., 2016 из бирманского янтаря они редуцировались до размера еле различимых пластинок.
К настоящему моменту описано 26 видов мезохризопид в составе 15 родов (см. список в: Yang et al., 2012; Liu et al., 2016). Самым разнообразным и широко распространенным из них является род Tachinymphes Ponomarenko, 1992, который насчитывает пять видов: T. ascalaphoides Ponomarenko, 1992 из раннего мела Забайкалья (местонахождение Байса), T. delicatus (Ren et Yin, 2002) и T. magnificus Nel et al., 2005 из раннего мела Китая, а также T. paicheleri Nel et al., 2005 и T. penalveri Nel et al., 2005 из раннего мела Испании (местонахождение Лас-Ойас). В настоящей работе описывается новый вид этого рода, T. koraiensis sp. nov., найденный в нижнемеловых континентальных отложениях формации Чинджу (провинция Кёнсан-намдо), в окрестностях одноименного южнокорейского города. Новый вид устанавливается по изолированному заднему крылу, однако поскольку задние крылья известны для всех пяти ранее описанных видов рода Tachinymphes, это не представляет больших трудностей.
T. koraiensis sp. nov. является первым сетчатокрылым, описываемым из формации Чинджу, а также первым ископаемым сетчатокрылым с территории Корейского полуострова. Возраст формации Чинджу, которую в литературе также иногда называют формацией Донмёнсон, точно не установлен. Руководствуясь таксономическим составом остракод, эту формацию относят к баррему (Hayashi, 2006); уран-свинцовый метод позволяет датировать ее поздним аптом–ранним альбом (Lee et al., 2010). Формация Чинджу частично сложена из черных сланцев, откуда и происходят отпечатки насекомых, а также рыбы и листовая флора. Вероятно, осадконакопление происходило в бескислородных условиях на дне глубокого озера, в поверхностных слоях которого обитала многочисленная водная энтомофауна.
Среди нескольких тысяч насекомых, собранных в формации Чинджу, водные насекомые преобладают. Почти все они целые, без признаков длительного посмертного переноса. Особенно обильны личинки и куколки комаров Chaoboridae, которые часто образуют напластования, в большом количестве попадаются как личинки, так и имаго водных жуков Coptoclavidae (Park et al., 2013), нимфы и имаго стрекоз Hemeroscopidae (Nam, Kim, 2016), личинки и имаго водных жуков Helophoridae, по предварительным данным, относящиеся к тем же родам, что отмечаются в нижнем мелу забайкальского местонахождения Байса (Fikáček et al., 2012). Присутствуют, но в гораздо меньшем количестве, клопы Corixidae. Если не считать структур, описывавшихся как ручейниковые биогермы (Paik, 2005), другие водные насекомые (помимо вышеперечисленных) в формации Чинджу отсутствуют или единичны. Особенно примечательно отсутствие личинок поденок, которые многочисленны во многих нижнемеловых континентальных отложениях российского Забайкалья, Монголии и Китая. Если следовать классификации раннемеловых водных энтомофаун, предложенной Н.Д. Синиченковой, то комплекс водных насекомых из формации Чинджу относится к 8 типу, для которого характерно обилие хаоборид и коптоклавид, при отсутствии личинок поденок (Sinitshenkova, 2002, с. 412).
Среди наземных насекомых в формации Чинджу наиболее массовой группой являются тараканы (Baek, Yang, 2004), за ними по числу находок идут жуки (Sohn et al., 2019) и различные Hemiptera, главным образом, Auchenorrhyncha и Heteroptera. В существенно меньшем количестве представлены Diptera, среди которых одной из массовых групп являются длиннохоботковые мухи-джангсольвиды (Khramov et al., 2020), Hymenoptera, Raphidioptera (Engel et al., 2006), Neuroptera, Orthoptera, стрекозы подотряда Zygoptera, а также Dermaptera (Engel et al., 2002). Остальные отряды отсутствуют или известны в виде единичных находок. Если не считать ранее публиковавшихся описаний отдельных видов, можно констатировать, что в целом энтомофауна формации Чинджу изучена еще недостаточно.
Голотип Tachinymphes koraiensis sp. nov. хранится в коллекции Национального ун-та образования г. Конджу (Южная Корея). Работа поддержана Программой Президиума РАН “Проблемы происхождения жизни и эволюция биосферы” и грантом РФФИ № 18-04-00322.
СЕМЕЙСТВО MESOCHRYSOPIDAE HANDLIRSCH, 1906
Род Tachinymphes Ponomarenko, 1992
Tachinymphes koraiensis Khramov et Nam, sp. nov.
Название вида koraiensis лат. – корейский.
Голотип – GNUE-216001, отпечаток и противоотпечаток заднего крыла; Южная Корея, провинция Кёнсан-намдо; нижний мел, формация Чинджу.
Описание (рис. 1). Крыло вытянутой формы, птеростигма хорошо развита, заканчивается чуть дистальнее слияния Sc и R1, костальное поле в два раза уже, чем поле между R1 и Rs, Rs c 10 ветвями, Sc и R1 сливаются чуть дистальнее места отхождения последней ветви Rs, имеются шесть длинных ступенчатых рядов поперечных жилок, третий (если считать с нижнего края крыла) из них образует складку наподобие линии Банкса, общий ствол MP + CuA разделяется чуть проксимальнее отхождения Rs от R1, MA отходит от Rs на уровне разветвления CuA, CuP с простым развилком на конце, A1 с тремя концевыми ответвлениями.
Рис. 1.
Tachinymphes koraiensis sp. nov. (Neuroptera: Mesochrysopidae), голотип GNUE-216001, заднее крыло: а – отпечаток, б – противоотпечаток (зеркально отражен), в – прорисовка жилкования. Длина масштабной линейки 3 мм.
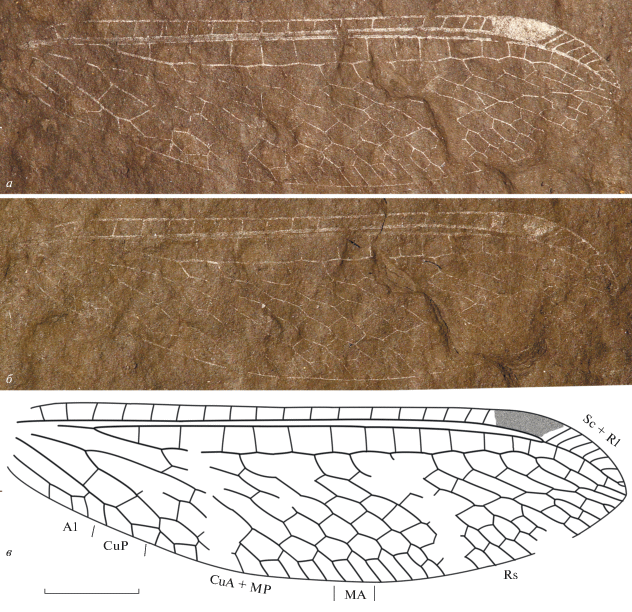
Размеры в мм: длина крыла – 19.5, ширина – 5.4.
Сравнение. О том, что экземпляр представляет собой заднее крыло, говорит отсутствие независимо идущего ствола CuA, который слился со стволом MP. О принадлежности экземпляра к роду Tachinymphes свидетельствует продолговатая, суженная форма крыла, общая конфигурация жилок CuA + MP и CuP, наличие птеростигмы (есть у T. delicatus в обоих крыльях и у T. paicheleri в переднем крыле), слабая выгнутость продольной складки, относительная немногочисленность поперечных жилок. T. koraiensis sp. nov. очень похож на T. ascalaphoides, отличается от него отсутствием темной окраски в дистальной части крыла. От всех остальных четырех видов отличается большим числом рядов поперечных жилок.
Материал. Голотип.
Список литературы
Пономаренко А.Г. Сетчатокрылые (Insecta, Neuroptera) из нижнего мела Забайкалья // Палеонтол. журн. 1992. № 3. С. 43–50.
Baek K.S., Yang S.Y. Cockroaches from the Early Cretaceous of Korea // J. Paleontol. Soc. Korea. 2004. V. 20. P. 71–98.
Engel M.S., Grimaldi D.A. The neuropterid fauna of Dominican and Mexican amber (Neuropterida: Megaloptera, Neuroptera) // Amer. Museum Novit. 2007. № 3587. P. 1–58.
Engel M.S., Lim J.D., Baek K.S. New fossil Mesochrysopidae (Neuroptera) from the Mesozoic of China // N. Jb. Geol. Paläontol. Mh. 2006. № 4. P. 249–256.
Engel M.S., Lim J.D., Baek K.S., Martin L.D. An earwig from the Lower Cretaceous of Korea (Dermaptera: Forficulina) // J. Kans. Entomol. Soc. 2002. V. 75. P. 86–90.
Fikáček M., Prokin A., Angus R.B. et al. Revision of Mesozoic fossils of the helophorid lineage of the superfamily Hydrophiloidea (Coleoptera: Polyphaga) // Acta Entomol. Mus. Natl. Pragae. 2012. V. 52. P. 89–127.
Hagen H.A. Ueber die Neuropteren aus dem lithographischen Schiefer in Bayern // Palaeontogr. 1862. V. 10. P. 96–145.
Hayashi K. Nonmarine ostracode zonation and long-distance correlation based on analysis of regional ostracode successions in China, Korea, Japan, and Mongolia // Cret. Res. 2006. V. 27. P. 168–188.
Khramov A.V., Nam G.S., Vasilenko D.V. First long-proboscid flies (Diptera: Zhangsolvidae) from the Lower Cretaceous of South Korea // Alcheringa. 2020. V. 44. P. 160–168.
Lee T.H., Park K.H., Chun J.H., Yi H.W. SHRIMP U-Pb zircon ages of the Jinju Formation and Silla conglomerate, Gyeongsang Basin // J. Petrol. Soc. Korea. 2010. V. 19. P. 89–101.
Liu X., Zhang W., Winterton S.L. et al. Early morphological specialization for insect-spider associations in Mesozoic lacewings // Curr. Biol. 2016. V. 26. P. 1590–1594.
Machado R.J.P., Gillung J.P., Winterton S.L. et al. Owlflies are derived antlions: anchored phylogenomics supports a new phylogeny and classification of Myrmeleontidae (Neuroptera) // Syst. Entomol. 2018. V. 44. P. 418–450.
Makarkin V.N., Menon F. New species of the Mesochrysopidae (Insecta, Neuroptera) from the Crato Formation of Brazil (Lower Cretaceous), with taxonomic treatment of the family // Cret. Res. 2005. V. 26. P. 801–812.
Makarkin V.N., Yang Q., Peng Y.Y., Ren D. A comparative overview of the neuropteran assemblage of the Early Cretaceous Yixian Formation (China), with description of a new genus of Psychopsidae (Insecta: Neuroptera) // Cret. Res. 2012. V. 35. P. 57–68.
Nam K.S., Kim J.H. Reconstruction and paleoecological implications of dragonfly Hemeroscopus baissicus Pritykina, 1977 (Hemeroscopidae) from the Lower Cretaceous Jinju Formation in the Jinju Area, Korea // Paleontol. Soc. Korea. 2016. V. 52. P. 105–112.
Nel A., Delclos X., Hutin A. Mesozoic chrysopid-like Planipennia: a phylogenetic approach (Insecta: Neuroptera) // Bull. Soc. Entomol. Fr. 2005. V. 41. P. 29–69.
Nel A., Henrotay M. Les Chrysopidae mésozoïques. État actuel des connaissances. Description d’un nouveau genre et nouvelle espèce dans le Jurassique inférieur (Lias) (Insecta: Neuroptera) // Ann. Soc. entomol. Fr. 1994. V. 30. P. 293–318.
Paik I.S. The oldest record of microbial-caddisfly bioherms from the Early Cretaceous Jinju Formation, Korea: occurrence and palaeoenvironmental implications // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2005. V. 218. P. 301–315.
Park T.Y., Kim Y.H., Nam K.S. Preliminary research on the aquatic coleopteran, Coptoclava from the Early Cretaceous Jinju Formation // J. Geol. Soc. Korea. 2013. V. 49. P. 617–624.
Sinitshenkova N.D. Ecological history of the aquatic insects // History of Insects / Eds. Rasnitsyn A.P., Quicke D.L.J. Dordrecht: Kluwer Acad. Publ., 2002. P. 388–417.
Sohn J.C., Nam G.S., Choi S.W., Ren D. New fossils of Elateridae (Insecta, Coleoptera) from Early Cretaceous Jinju Formation (South Korea) with their implications to evolutionary diversity of extinct Protagrypninae // PLoS ONE. 2019. V. 14. e0225502.
Yang Q., Makarkin V.N., Ren D. New fossil Mesochrysopidae (Neuroptera) from the Mesozoic of China // Zootaxa. 2012. V. 3597. P. 1–14.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал