

Палеонтологический журнал, 2020, № 5, стр. 49-61
Микроструктура раковины у древних патеринид (брахиоподы) и ее преобразования в ходе захоронения
Г. Т. Ушатинская *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: gushat@paleo.ru
Поступила в редакцию 03.12.2019
После доработки 12.12.2019
Принята к публикации 12.12.2019
Аннотация
Изучена микроструктура раковин и ее посмертные преобразования у ранне–среднекембрийских представителей двух семейств, составляющих отряд Paterinida. При жизни их раковины имели органо-фосфатный скелет, в котором значительная часть принадлежала органическому веществу; последнее при захоронении оказывало существенное влияние на конечный результат фоссилизации. Большую роль в посмертных преобразованиях микроструктуры играли условия, в которых раковины оказывались после смерти.
Класс Paterinata (отряд Paterinida) – раннепалеозойские брахиоподы, при жизни обладавшие органо-фосфатной раковиной. Как и у органо-фосфатных брахиопод подтипа Linguliformea, у патеринид отсутствовало замковое сочленение в виде минеральных выростов на заднем крае брюшных створок и соответствующих им ямок в спинных. При этом, в отличие от остальных лингулиформей, они обладали настоящими интерареями с дельтирием и нототирием, частично или полностью прикрытыми, соответственно, дельтидием и хилидием, и прямым или почти прямым задним краем, что сближает их с подтипом замковых брахиопод Rhynchonelliformea (или Articulata). Кроме того, у некоторых представителей древнейших патеринид у макушки брюшной створки имелось округлое отверстие для прохода ножки, прикрытое сзади вогнутой пластиной – коллеплаксом. Такая структура известна еще у представителей раннекембрийского отряда Chileida из класса Rhynchonelliformea (Holmer et al., 2009). Сочетание этих признаков обусловило сложности при отнесении патеринид к таксону высокого ранга. В “Основах палеонтологии” (Горянский, 1960) они были объединены в один отряд Kutorginida с надсемействами Paterinacea и Kutorginacea. В последнее входили брахиоподы с карбонатными раковинами, а весь отряд рассматривался в составе класса Inarticulata (беззамковые). В первом издании “Treatise …” (Rowell, 1965) патериниды были выделены в отдельный отряд Paterinida Rowell, 1965, который также вместе с отрядами карбонатных брахиопод Craniida и Obolellida был включен в класс Inarticulata.
Позже В.Е. Горянский и Л.Е. Попов (1985) выделили класс Lingulata Gorjansky et Popov, 1985, к которому они отнесли все отряды брахиопод, обладавшие органо-фосфатной раковиной, в том числе и патеринид. И, наконец, А. Вильямс и др. (Williams et al., 1996), проанализировав сочетание таких признаков ранних патеринид, как органо-фосфатный состав раковины, строение заднего края створок, присутствие интерарей на обеих створках, устройство мускулатуры, мантийных каналов, и присутствие у некоторых патеринид форамена, прикрытого частично коллеплаксом, выделили патеринид в отдельный класс Paterinata, вошедший вместе с классом Lingulata в подтип Linguliformea. Такая классификация принята в последних изданиях “Treatise …” (Laurie, 2000; Holmer, Popov, 2000, 2007) и сейчас широко используется в литературе. Класс Paterinata состоит из отряда Paterinida Rowell, 1965, который включает два семейства – Cryptotretidae Pelman, 1977 и Paterinidae Schuchert, 1893 (табл. 1). Наиболее древним является семейство Cryptotretidae, первые входящие в его состав роды появились в основании томмотского яруса, а последние вымерли в течение ботомского века. Представители семейства Paterinidae известны с ботомского века, к концу ордовика они вымерли (Williams et al., 1998, рис. 7 ).
Таблица 1.
Развитие взглядов на положение патеринид в системе брахиопод
| Тип Brachiopoda | |||
|---|---|---|---|
| Горянский, 1960 | Rowell, 1965 | Горянский, Попов, 1985 | Williams et al., 1996 |
| Класс Inarticulata | Класс Inarticulata | Класс Lingulata | Класс Paterinata |
| Отряд Kutorginida | Отряд Paterinida + отряды Obollelida, Craniida |
Отряд Paterinida + отряды Lingulida, Acrotretida, Siphonotretida | Отряд Paterinida |
| Н/сем. Kutorginacea Paterinacea | Н/сем. Paterinacea | Н/сем. Paterinacea | Н/сем. Paterinoidea |
| Сем. Paterinidae | Сем. Paterinidae | Сем. Paterinidae | Сем. Paterinidae Cryptotretidae |
Раковины патеринид состояли из периостракума, первичного и вторичного слоев (Williams et al., 1998). Микроскульптура и микроструктура раковин обоих семейств патеринид изучались неоднократно (Попов, Ушатинская, 1987; Ушатинская, 1987, 1993, 1995; Williams et al., 1998; Holmer et al.; 2009; Skovsted et al., 2014, 2015). В первых работах была описана микроструктура вторичного слоя у представителей семейства Cryptotretidae (роды Pelmanotreta, Salanygolina, Dzunarzina), состоящая из тонких пластин (ламин) от 15–20 до 20–40 мкм толщиной, приблизительно параллельных наружной поверхности, сложенных плотно прилегающими друг к другу угловатыми призмами, высота которых совпадает с толщиной ламин, а ширина равна 7–8 мкм. Такая микроструктура была названа фавозитной (Ушатинская, 1995). Вильямс с соавт. (Williams et al., 1998) подробно описали микроскульптуру и строение первичного и вторичного слоев у обоих семейств патеринид на примере большинства известных в этом отряде родов из разных регионов. По их данным, вторичный слой у представителей семейства Cryptotretidae состоит из чередования тонких, горизонтально расположенных фосфатизированных “мембран” 80–100 нм толщиной и апатитовых ламин до 2 мкм толщиной. На внутренней поверхности створок иногда можно видеть, что “мембраны” построены прилегающими друг к другу округленно-гексагональными призмами–куполами, отражающими клеточное строение внешнего эпителия мантии. Апатитовые ламины состоят из сферолитов около 100 нм в диаметре и из игольчатых кристаллов апатита. Позже Л. Хольмер и Х. Сковстед с соавт. (Holmer et al., 2009; Skovsted et аl., 2015) подтвердили присутствие фавозитного типа микроструктуры во вторичном слое у Pelmanotreta и Salanygolina, но подробно на причинах столь существенной разницы в описаниях различных авторов не останавливались. Как показало изучение створок патеринид из обоих семейств, происходящих из различных местонахождений нижнего и среднего кембрия Сибирской платформы и из нижнего кембрия Монголии, вероятно, из-за высокого содержания при жизни органики в их раковинах первичное строение – собственно минеральный каркас створок – сохраняется редко. Гораздо чаще попадаются створки, в которых слагавшее их при жизни органо-фосфатное вещество преобразовано в плотный, с плохо различимой структурой, фосфатный агрегат, или же в скопление сферолитов, игольчатых или гантелеобразных кристаллов. Но иногда на сколах остаются участки, которые, вероятно, отражают первичную микроструктуру апатитовой части створок.
Статья написана на основе переизучения строения стенок раковин представителей обоих семейств отряда Paterinida из нижнего и среднего кембрия Сибирской платформы и из нижнего кембрия Монголии, и в ней рассмотрены варианты посмертного преобразования первичной микроструктуры в зависимости от условий захоронения.
СТРОЕНИЕ СТЕНКИ РАКОВИНЫ БРАХИОПОД ОТРЯДА PATERINIDA
Семейство Cryptotretidae. Описания даны на основании изучения родов Aldanotreta Pelman, 1977 и Tumulduria Missarzhevsky in Rozanov et al., 1969 из томмотского яруса, Pelmanotreta Skovsted et al., 2015 из томмотского и атдабанского ярусов Сибирской платформы, и родов Dzunarzina Ushatinskaya, 1993 из аналогов атдабанского и Salanygolina Ushatinskaya, 1987 из аналогов ботомского ярусов Монголии (Ушатинская, 1987, 1993, 1995) (рис. 1). Следует отметить, что створки криптотретид больше по размерам створок представителей семейства Paterinidae, также встречающихся на Сибирской платформе. Так, по данным Ю.Л. Пельмана (1977), брюшные створки Aldanotreta достигают 21 мм в длину и 25 мм в ширину, хотя в среднем их размеры колеблются в пределах 7–10 мм. Створки Pelmanotreta обычно помельче – они имеют размеры около 6–8 мм в ширину и 3–5 мм в длину. Длина и ширина створок у Dzunarzina и Salanygolina колеблется в пределах 2–5 мм. Соответственно, и толщина створок у Aldanotreta достигает 80–100 мкм, а у остальных криптотретид – от 20–30 до 50 мкм.
Рис. 1.
Изученные представители отряда Paterinida из нижнего и среднего кембрия Сибирской платформы (прямой шрифт) и из нижнего кембрия Монголии (курсив).
Микроскульптура поверхности у криптотретид подробно описана Вильямсом и соавт. (Williams et al., 1998). Авторы отмечают, что хотя периостракум на створках не сохранился, его слепок во многих случаях отчетливо отпечатан на наружной поверхности первичного слоя, и это позволяет достаточно определенно судить о прижизненном рельефе поверхности. Снаружи взрослые створки криптотретид были покрыты многочисленными тонкими, иногда прерывистыми концентрическими струйками. У Aldanotreta (самого крупного из известных среди криптотретид родов) струйки широкие (до 50–80 мкм), частые, хорошо видные, тесно расположенные, с высотой около 10 мкм. Иногда видно, что поверхность струек покрыта еще более тонкими нитевидными часто расположенными концентрическими линиями (табл. IV , фиг. 1а, 1б; см. вклейку). У Tumulduria поверхностная скульптура тоже состоит из тонких, слегка уплощенных и близко расположенных концентрических струек, но они пересекаются частыми нитевидными радиальными бороздками, которые превращают струйки в короткие волнообразные (фестончатые) отрезки, создавая видимость мелкосетчатой скульптуры (табл. IV , фиг. 8, 10 а, 10б). Поверхности створок Salanygolina и Pelmanotreta тоже несут многочисленные концентрические прерывисто-волнистые струйки (табл. V , фиг. 2а, 2б; см. вклейку; рис. 2, а–в). Выделяются струйки более широкие и высокие первого порядка, их на поверхности створок средней величины бывает 7–12, они имеют ширину около 10–25 мкм, и отстоят одна от другой на 30–50 мкм. Между ними находятся по 4–5 более мелких струек второго порядка, шириной 3–5–8 мкм, тоже волнистых и прерывистых. У Dzunarzina струйки второго порядка еще более мелкие, шириной 1–2 мкм, тесно прилегают друг к другу, между двумя соседними более широкими концентрическими струйками бывает до 10–12 мелких (табл. V , фиг. 5а–5в). Прерывистость струек на поверхности у патеринид, как и у многих органо-фосфатных брахиопод, по мнению Вильямса и Хольмера (Williams, Holmer, 1992), объясняется появлением на их протяжении “точек сужения” (nickpoints), возникающих из-за временного присутствия у переднего края периостракума клеток, заключающих щетинки – сета; по мере роста раковины клетки с сета передвигаются кпереди. Эти “точки сужения” нарушают непрерывность концентрических струек. У Tumulduria прерывистость скорее связана с развитием очень тонких радиальных бороздок, равномерно пересекающих концентрические струйки (табл. IV , фиг. 10 б).
Рис. 2.
Pelmanotreta negeurtchenensis (Pelman, 1977): а – экз. ПИН, № 5486/551, спинная створка снаружи; Сиб. платформа, левый берег р. Лены, местонахождение Негюрчене; б–г – экз. ПИН, № 5486/ 552, спинная створка снаружи: б – задняя часть, в – поверхность на участке взрослой части, г – поверхность на участке брефической раковины той же створки, продольное углубление – след от проходившей при жизни здесь щетинки, по бокам углубления – срез первичного слоя, поверхность которого сложена мелкими сферолитами; Сиб. платформа, правый берег р. Лены, разрез Журинский мыс, слой 6 (Путеводитель …, 1973); нижний кембрий, атдабанский ярус, пестроцветная свита; д – экз. ПИН, № 5486/553, участок наружной поверхности створки, уступ образован сохранившимся первичным слоем; е – экз. ПИН, № 4290/9, косой скол обломка створки, на котором видны ступенчатые ламины, сложенные призмами, наружная сторона створки в нижней части снимка; ж, и – экз. ПИН, № 5486/554, обломок спинной створки: ж – скол сбоку; и – то же, увеличено; к – экз. ПИН, № 5486/555, сильно увеличенный участок скола створки, на котором видны пустые пространства, оставшиеся от разрушенных оболочек призм, образующих столбики; л – экз. ПИН, № 5486/556, участок поперечного скола створки с линзой, частично заполненной мелкими сферолитами, вверху снимка первичный слой; м – экз. ПИН, № 5486/559, обломок спинной створки, внутренняя поверхность со слепками клеток внешнего эпителия; н – экз. ПИН, № 5486/557, неполная спинная створка, скол сбоку, видны два перекристаллизованных слоя, состоящие из слившихся ламин, они разделены двумя тонкими бесструктурными слоями, вероятно, оставшимися от органических прослоев, в левой части снимка видны небольшие участки, состоящие из сохранившихся призматических ламин; о – экз. ПИН, № 5486/558, наружная поверхность первичного слоя, состоящая из гантелеобразных кристаллов и мелких сферолитов; Сиб. платформа, правый берег р. Лены, разрез Журинский мыс, слой 3 (Путеводитель…, 1973); нижний кембрий, верхи томмотского яруса; п – экз. ПИН, № 5486/560, брюшная створка, скол сбоку, пачки тонких “мембран”, пространствo между которыми заполнено мелкими сферолитами; правый берег р. Лены, разрез Ой-Муран, нижняя часть осыпи, бечевник; нижний кембрий, атдабанский ярус, пестроцветная свита.
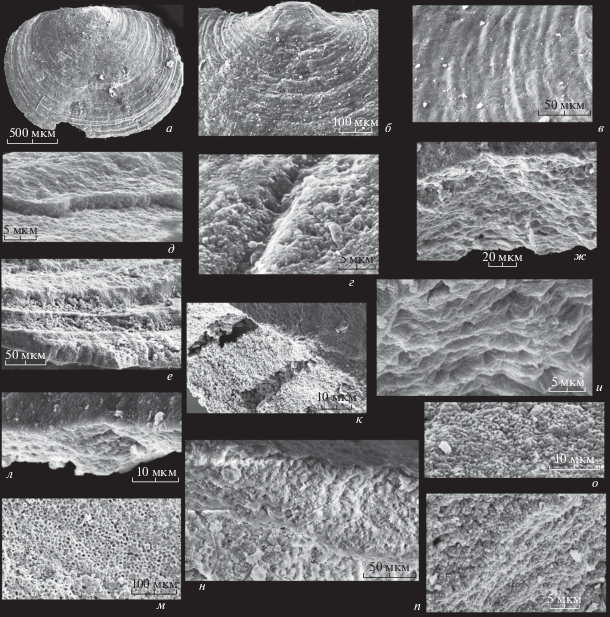
Первичный слой редко сохраняется на всей поверхности створок. Его толщина около 5 мкм, в местах пересечения с наружными струйками он немного изгибается и утолщается (табл. IV , фиг. 5а; табл. V , фиг. 10 а; рис. 2, г, д, к, л). Иногда он совсем тонкий – около 1 мкм, и сливается с лежащим под ним вторичным слоем. На сколе слой чаще всего гомогенный, отдельные элементы в нем не различимы, но бывает, что он разбит вертикальными трещинками и выглядит столбчатым, вероятно, за счет вторичного преобразования (табл. IV , фиг. 5а; рис. 2, д). При жизни первичный слой, скорее всего, был органическим по составу, хотя мог содержать мелкие фосфатные включения, как у современных лингулид, и был минерализован в ходе фоссилизации (Williams et al., 1994). При большом увеличении поверхность первичного слоя, несмотря на присутствие концентрической скульптуры, вся состоит из мелких (менее 1 мкм) или из более крупных (3–5 мкм) тесно прилегающих друг к другу сферолитов, что как раз является результатом его, вероятно, очень быстрой минерализации (табл. IV , фиг. 3; рис. 2, г, о).
Рис. 3.
а, б – Dzunarzina elenae Ushatinskaya, 1993, экз. ПИН, № 3302/5060, спинная створка: а – участок наружной поверхности в левой стороне снимка с частично содранным первичным слоем, под ним находится поверхность вторичного слоя, состоящая из оболочек призм, которыми была сложена верхняя ламина данного слоя; б – то же, увеличено; Монголия, гора Дзун-Арца; нижний кембрий, ботомский ярус, нижняя часть (пересечение D-105; Воронин и др., 1983); в, г – Salanygolina obliqua Ushatinskaya, 1987: в – экз. ПИН, № 3302/5025, поверхность спинной створки изнутри с частично отсутствующим внутренним слоем, под ним видно строение поверхности ламины вторичного слоя из тесно прилегающих друг к другу призм, от которых остались одни оболочки; г – экз. ПИН, № 3302/5026, участок внутренней поверхности створки с остатками перегородок между призмами; Монголия, разрез Саланы-Гол (пересечение С-205; Граница докембрия и кембрия…, 1982); нижний кембрий, ботомский ярус, хаирханская свита; д, е – Pelmanotreta negeurtchenensis (Pelman, 1977), экз. ПИН, № 4290/214, обломок спинной створки: д – внутренняя поверхность, несущая призмы, от которых сохранились только оболочки; е – то же, увеличено; Сиб. платформа, правый берег р. Лены, разрез Журинский мыс, слой 3 (Путеводитель…, 1973); нижний кембрий, томмотский ярус, пестроцветная свита.

Вторичный слой у всех изученных криптотретид составляет основную часть створок и часто сложен плохо раскристаллизованным апатитом с неотчетливо видной тонкой слоистостью (табл. IV , фиг. 7 ; табл. V , фиг. 10 а; рис. 2, п). Иногда можно рассмотреть на сколах мелкие сферолиты. Вероятно, это результат посмертной фоссилизации вещества раковины. Но в некоторых местонахождениях удалось наблюдать не полностью измененную в ходе посмертных преобразований первичную микроструктуру. У таких экземпляров вторичный слой или его участки состоят из пакетов ламин, почти параллельных наружной поверхности створок. У родов Pelmanotreta и Salanygolina толщина отдельных ламин составляет 10–20 мкм, сами ламины сложены плотно прилегающими друг к другу 4-х, 5-ти, 6-ти угольными призмами, полыми внутри, от них сохраняются только оболочки, высота призм совпадает с толщиной ламин, а ширина составляет 7–10 мкм (табл. V , фиг. 1в, 2в, 2г; рис. 2, е). Именно такой тип микроструктуры у патеринид был назван мною фавозитным (Ушатинская, 1995). Но, как показали последующие наблюдения, подобное строение стенки раковины у патеринид наблюдается нечасто. Вильямс и др. (Williams et al., 1998) написали, что в строении раковин патеринид принимают участие пакеты тонких протяженных “мембран”. Действительно, на боковых стенках призм при большом увеличении можно разглядеть очень тонкую параллельную слоистость (табл. V , фиг. 2г; рис. 2, к). Представляется, что “мембраны” являются сейчас минерализованными, а при жизни были органо-фосфатными отдельными слойками, составлявшими стенку раковины и отложившимися в ходе деятельности внешнего эпителия мантии. Клеточное строение эпителия определило тонкопризматическую природу “мембран”. Несколько таких “мембран”, образовавшихся в короткий промежуток времени, составляют ламину, а расположенные друг под другом клетки–призмы слились в столбики. Внутри столбиков при жизни, вероятно, находилась органо-фосфатная смесь, а друг от друга они были отделены органическими стенками окружавших их с боков клеток. Столбики на сколах и на поверхностях ламин часто бывают полыми, а разделяющие их стенки толщиной около 1 мкм состоят из очень мелких сферолитов или из почти не раскристаллизованного апатита (табл. V , фиг. 2в, 2г). Но бывает, что призмы и столбики выглядят как бы негативными слепками этих структур; их внутреннее пространство состоит из тонких параллельных “мембран”, а друг от друга они отделены тонкими полыми щелями, в которых при жизни могли располагаться разделявшие их стенки (рис. 2, к). Образование пакета “мембран” прерывалось отложением тонкого, вероятно, органического слоя, как это происходит и у современных лингулид. Затем опять откладывался пакет “мембран”. В результате ламины отделялись одна от другой тонкими слойками–пленками, которые в ходе фоссилизации тоже превратились в тонкие ламины (рис. 2, н), но иногда они разрушались, и тогда на их месте образовывались узкие полые щели (табл. IV , фиг. 6 б; рис. 2, и). У рода Dzunarzina ламины, слагающие вторичный слой, гораздо тоньше, чем у Pelmanotreta и Salanygolina, всего 3–4 мкм, и тоже состоят из нескольких “мембран” (табл. V , фиг. 6 а–6в, 7, 10б). Последние сложены 4-х, 5-ти и 6-ти угольными, плотно прилегающими друг к другу клетками–призмами 8–12 мкм в поперечнике (табл. V , фиг. 6 в). Перегородки между ними, вероятно, были совсем тонкими, при большом увеличении видна лишь узкая полоска, разделяющая призмы. Призмы состоят из очень мелких, менее 1 мкм, сферолитов или удлиненных кристаллов (табл. V , фиг. 6 в, 9а, 9б). Так же, как у Pelmanotreta и Salanygolina, ламины, состоящие из нескольких “мембран”, отделялись друг от друга органическими слойками–пленками, сейчас между ними иногда видны полые щели (табл. V , фиг. 10 б).
Внутренняя поверхность вторичного слоя всех трех родов часто покрыта сферолитами, вероятно, образовавшимися в ходе растворения органо-минерального вещества и последующего минералообразования из возникшего раствора (табл. V , фиг. 8, 10 а, 10в). Но встречаются экземпляры, у которых на внутренней стороне створок сохранились отпечатки клеток наружного эпителия мантии, плотно прилегавшего к ней при жизни. Это полые внутри, угловатые, часто неправильной формы призмы, ограниченные стенками, выстилающие значительную часть внутренней поверхности створок. Стенки клеток–призм могут быть тонкими, менее 1 мкм, а могут быть сильно утолщенными за счет вторичного отложения апатита (табл. V , фиг. 3; рис. 2, м; 3, г–е). Размеры призм в поперечнике 8–12 мкм. Представляется, что при жизни оболочки были более плотными, чем содержимое клеток, и были минерализованы при быстрой фоссилизации, в том числе за счет растворения содержимого клеток. Иногда отпечатки клеток внешнего эпителия мантии сохраняются на поверхностях ламин вторичного слоя. На рис. 4, д–ж показан участок поверхности вторичного слоя под отсутствующим первичным. На нем видны немного округленные, но еще угловатые призмы, разделенные полыми промежутками. Скорее всего, это не доведенный до конца процесс преобразования органо-фосфатного вещества, содержащегося в клетках, слагавших “мембраны”, в полые сферолиты. Последние начали образовываться в ячейках призм, из которых состояла данная “мембрана”. Пустые промежутки – растворенные оболочки между призмами.
Рис. 4.
а, б – Aldanotreta sunnaginensis Pelman, 1977, экз. ПИН, № 5486/508, обломок спинной створки изнутри: а – внутренняя поверхность, полностью покрытая сферолитами; б – то же, увеличено; разрез по левому берегу р. Алдан; нижний кембрий, томмотский ярус, нижняя часть пестроцветной свиты, в, г – Pelmanotreta sp., экз. ПИН, № 5486/561, внутренняя поверхность обломка спинной створки: в – участок со сферолитами; г – отдельный сферолит; Сиб. платформа, левый берег р. Лены, разрез Ой-Муран, осыпь в нижней части склона; д–ж – Pelmanotreta negeurtchenensis (Pelman, 1977), экз. ПИН, № 5486/562, спинная створка снаружи: д – участок под отсутствующим первичным слоем; е, ж – увеличенные фрагменты рис. 4, д; Сиб. платформа, правый берег р. Лены, разрез Журинский мыс, слой 6 (Путеводитель …, 1973); нижний кембрий, атдабанский ярус, пестроцветная свита; и, к – Tumulduria incomperta Missarzhevsky, 1969, экз. ПИН, № 5486/522, спинная створка: и – участок скола близ переднего края, видны слоистые сферолитоподобные структуры в пустотах, оставшихся после растворения центральных частей призм; к – другой участок, вверху лежат радиально-лучистые сферолиты, выстилающие створку изнутри, под которыми видна поверхность одной из ламин вторичного слоя, состоящая из мелких призм; л – экз. ПИН, № 5486/524, участок поперечного скола с сохранившимися слоями отдельных “мембран”, слившихся в ламины и с перегородками между призмами, некоторые призмы замещены полыми внутри сферолитоподобными образованиями; север Сиб. платформы, бассейн р. Оленек; нижний кембрий, томмотский ярус, нижняя часть еркекетской свиты.

Строение стенок раковин двух других родов, относимых к семейству Cryptotretidae, Aldanotreta и Tumulduria, до сих пор описывалось как состоящее целиком из апатитовых сферолитов, что объяснялось посмертными преобразованиями микроструктуры (Ушатинская, 1995; Williams et al., 1998; Skovsted et al., 2015). Изучение имеющегося материала показало, что в отдельных случаях на некоторых участках створок у обоих родов можно видеть реликты микроструктуры вторичного слоя, при жизни слагавшего основную часть раковин. И она мало отличается от микроструктуры других криптотретид. На сколах створок Aldanotreta, происходящих из самых нижних горизонтов томмотского яруса разрезов по р. Алдан, видно, что створки состояли из протяженных ламин, параллельных их поверхностям; в пространстве между ламинами, сейчас неплотно заполненном сферолитами и мелко раскристаллизованным апатитом, располагались перпендикулярные к ним перегородки, вероятно, оставшиеся от оболочек призм, слагавших ламины. Толщина ламин – 8–10 мкм, расстояние между перегородками – 6–8 мкм, толщина перегородок – около 1 мкм (табл. IV , фиг. 5б, 6а, 6б). На другом образце такое же призматическое строение поверхности вторичного слоя видно на поверхности обломка створки под отсутствующим первичным слоем (табл. IV , фиг. 4а, 4б).
Похожие участки слоистой микроструктуры обнаружены и на сколах створок у рода Tumulduria, встреченного как в тех же слоях нижнего кембрия в среднем течении р. Алдан, что и Aldanotreta, так и на севере Сибирской платформы в бассейне р. Оленек в основании еркекетской свиты, коррелируемом с томмотским ярусом (Сухов и др., 2016). Чаще всего вся внутренняя поверхность створок у Tumulduria и их обломков сложена сферолитами до 10–20 мкм в диаметре (табл. IV , фиг. 9 а, 9б). На сколах тоже видны отдельные или слившиеся сферолиты, радиально-лучистые или полые, имеющие однослойную оболочку или же состоящие из нескольких слоев (рис. 4, и–л). Но сохранились небольшие участки, где можно видеть реликты первичного строения (табл. IV , фиг. 11 а, 11б; рис. 4, к, л). Как и у других криптотретид, створки тумулдурий были, вероятно, сложены тонкими “мембранами”, параллельными наружной поверхности, “мембраны” состояли из призм, от которых местами остались перегородки, и объединялись в ламины, разделенные органическими прослоями. Толщина ламин и размеры призм примерно такие же, как и у Aldanotreta (табл. IV , фиг. 11 б; рис. 4, л). На внутренней поверхности одного из образцов сохранились следы призматического строения отдельных “мембран” (табл. IV , фиг. 11 а).
Иногда на поперечных сколах створок у всех изученных родов встречаются небольшие линзовидные участки, вытянутые параллельно поверхностям створок, длиной 30–40 мкм и толщиной 10–20 мкм (табл. IV , фиг. 6, 7 ). Они могут быть полыми, а могут быть частично заполнены мелкораскристаллизованным апатитом. По мнению Вильямса и его коллег (Williams et al., 1998), эти линзы при жизни были сложены органическим веществом, которое при захоронении растворилось, а пустоты иногда оставались незаполненными, а иногда в них откладывались мелкие кристаллы.
Семейство Paterinidae. Из семейства Paterinidae были изучены представители родов Paterina Beacher, 1891, Micromitra Meek, 1873 и Dictyonina Cooper, 1942 из среднего кембрия Сибирской платформы. Длина и ширина их створок редко превышает 1–2 мм, их толщина в задней части близ макушки около 50 мкм, а у переднего края – около 10–20 мкм.
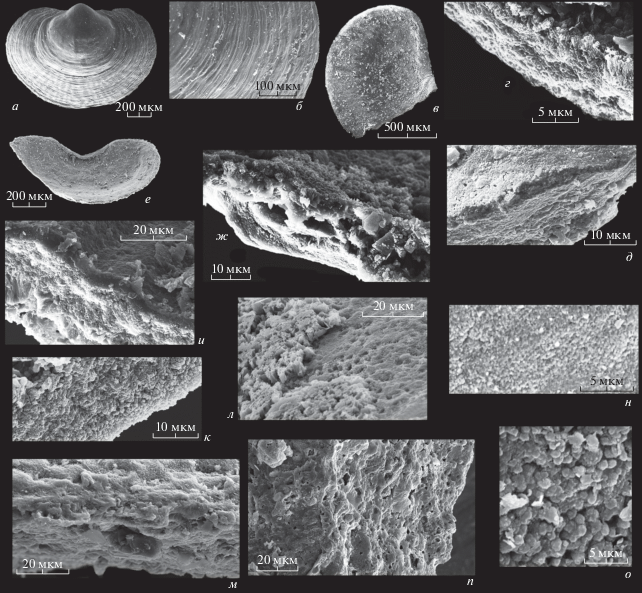
Скульптура поверхности у перечисленных родов патеринид на примере образцов из среднего и верхнего кембрия нескольких регионов мира детально уже описана (Williams et al., 1998), так что на ней останавливаться не будем. Первичный слой, как и у криптотретид, тонкий – около 1–2 мкм; его наружная поверхность при большом увеличении состоит из мелких сферолитов около 0.1–0.2 мкм в диаметре (рис. 5, н). На сколе он часто гомогенный, но бывает, что построен мелкими кристаллами апатита (рис. 5, г, и). Вторичный слой у патеринид тоже часто выглядит скрытокристаллическим, иногда на сколе можно разглядеть мелкие (менее микрона в диаметре) сферолиты или игольчатые кристаллы (рис. 5, г, к). Но у некоторых образцов, особенно если раковина очень тонкая (около 10–20 мкм), видно, что вторичный слой состоял из тонких протяженных слабоизвилистых “мембран” (около 0.4–0.5 мкм толщиной), иногда собранных в линзовидные пачки по несколько мембран, образующих параллельные поверхностям створок ламины толщиной в 3–4 мкм. Друг от друга такие ламины, вероятно, были отделены тонкими органическими пленками, определявшими их нижнюю и верхнюю границы (рис. 5, г, д, п). Как и у криптотретид, отдельные “мембраны” сложены мелкими, тонкими (в толщину “мембраны”) призмами. Об этом говорят угловатые выступы краев “мембран” на сколах створок и их поверхности в тех местах, где отсутствуют вышележащие слои (рис. 5, л, п). Иногда на таких поверхностях сохраняются минерализованные оболочки призм (рис. 5, л, с правой стороны), в середине они полые. На сколах створок встречаются мелкие участки, где ламины сложены призматическими столбиками (рис. 5, м). Имеются пустоты–линзы длиной 10–15 мкм, частично заполненные кристаллами, которые, вероятно, возникли на месте включений органического вещества (рис. 5, ж, м). Часто внутренняя поверхность створок покрыта мелкими сферолитами (рис. 5, о), но в некоторых случаях на внутренней поверхности сохранился слепок наружного слоя мантийного эпителия, состоявший при жизни из призматических клеток около 5–6 мкм в диаметре (рис. 3, д, е; 5, д, л).
Рис. 5.
а–ж, н, о – Paterina sp.: а, б – экз. ПИН, № 5486/601, брюшная створка снаружи: а – общий вид, б – скульптура на поверхности взрослой части створки; в–д – экз. ПИН, № 5486/602, обломанная спинная створка изнутри: в – общий вид, г – поперечный скол в средней части, д – участок внутренней поверхности, ступенька образована краем внешнего эпителия мантии; е, ж – экз. ПИН, № 5486/603, спинная створка изнутри: е – общий вид, ж – поперечный скол у переднего края; н – экз. ПИН, № 5486/604, наружная поверхность первичного слоя на спинной створке, сложенная мелкими сферолитами; о – экз. ПИН, № 5486/605, участок внутренней поверхности спинной створки, покрытый сферолитами; и – Micromitra sp., экз. ПИН, № 5486/606, участок поперечного скола спинной створки, наружная сторона с первичным слоем вверху снимка; верхи нижнего–средний кембрий, юнкюлябит-юряхская свита; к–м, п – Dictyonina sp.: к – экз. ПИН, № 5486/607, участок поперечного скола обломка спинной створки, наружная поверхность с первичным слоем внизу снимка; Сиб. платформа, район нижнего течения р. Оленек; средний–низы верxнего кембрия, чомурдахская свита; л, м – экз. ПИН, № 5486/608, спинная створка: л – участок внутренней поверхности, в левой стороне снимка – слепки клеток внешнего эпителия мантии, под которыми (справа) поверхность ламины вторичного слоя с углублениями, оставшимися от призм; м – участок поперечного скола той же створки, видны полая линза, при жизни, вероятно, заполненная органикой, и остатки перегородок между призмами; низовья р. Лены, Булкурская антиклиналь; средний кембрий, майский ярус, маяктахская свита; п – экз. ПИН, № 5486/609, участок поперечного скола спинной створки; Сиб. платформа, район нижнего течения р. Оленек, средний–низы верхнего кембрия, чомурдахская свита.
Из описания и приведенных фотографий можно видеть, что типы строения микроструктуры стенок раковин у брахиопод из семейств Cryptotretidae и Paterinidae очень близки. Различия имеются в толщине ламин, в величине слагающих их призм, что могло быть связано с гораздо меньшими размерами представителей семейства Paterinidae.
* * *
Раковины патеринид при жизни состояли из тонких параллельных друг другу органо-фосфатных “мембран”, объединяемых в ламины (пластины), почти параллельные поверхностям створок и отделенные друг от друга тонкими органическими слоями, которые, возможно, включали и небольшое количество фосфатного материала. В раковинах современных лингулиформей содержится от 40 до 62% органической составляющей (вместе с водой), которая включает протеин, хитин и гликозаминогликаны (GAGs); остальная часть минеральная и представлена апатитом (франколитом) (Williams et al., 1998). Можно предположить, что в раковинах впервые появившихся на Земле древнейших органо-фосфатных брахиопод количество органики еще сильнее превышало содержание минерального вещества. По данным Х. Эмига (Emig, 1986), створки современных лингулид, оказавшиеся на поверхности осадка, под воздействием биохимических и механических сил разрушаются в течение нескольких дней, а сохраниться в ископаемом состоянии имеют шанс лишь те, что погибли в норках или после смерти были быстро занесены осадками в результате штормов, придонных течений, оползней и пр. Вероятно, так же происходило и в раннем палеозое. И сохраниться в ископаемой летописи могли только те раковины или створки, у которых вскоре после смерти была хотя бы частично минерализована заключенная в них органическая составляющая, скреплявшая минеральную часть скелета. Слои, из которых состояли раковины патеринид, очень тонкие, толщина створок, особенно у переднего края, могла достигать всего 10–20 мкм. По предположению Вильямса и др. (Williams et al., 1998), развитие в ходе фоссилизации в раковинах патеринид большого количества тонких игольчатых кристаллов может говорить о более легкой деградации органической составляющей, отличавшейся по составу от органики, характерной для других лингулиформей. Такое предположение тоже нельзя исключать.
Чаще всего ископаемые остатки фосфатных брахиопод, в том числе и патеринид, извлекаются путем растворения содержащих их пород в слабой уксусной кислоте, поэтому важным является присутствие в исследуемой породе известняка. Но неоднократно отмечено, что лучше сохраняются створки, извлеченные из тех образцов, где, помимо карбонатной составляющей, присутствует и тонкий терригенный материал (Ушатинская, 2018). Например, первичная микроструктура у створок Pelmanotreta лучше сохранилась в образцах из пестроцветной свиты в местонахождениях Журинский мыс и Ой-Мурен, из слоев, которые сложены сильно глинистыми вишнево-красными средне- и тонкоплитчатыми известняками (Ярусное расчленение …, 1983). Вероятно, их образование происходило на небольшой глубине при преобладании карбонатного осадконакопления в неспокойных условиях, постоянном перемещении и перераспределении на дне турбидных или штормовых потоков, содержащих тонкий глинистый материал. Присутствующие среди слоистых глинистых известняков водорослево-археоциатные биогермы, по краям которых могли обитать брахиоподы, и неспокойные условия близ поверхности дна способствовали сносу и расчленению их раковин и их быстрому захоронению под тонкими глинисто-карбонатными породами. Примечательно, что наряду с несколькими десятками плоских спинных створок, иногда хорошей сохранности, встречено не более пяти брюшных, и то частично разрушенных; брюшные створки у Pelmanotreta были сильно выпуклыми, потому они скорее обламывались и разрушались даже при небольшом перемещении. Нельзя исключать, что количество органического вещества в брюшных створках было больше, чем в спинных, они были более тонкими у макушки, где проходила ножка, что также могло препятствовать захоронению.
Вероятно, похожие условия осадконакопления были в это же время на территории Западной Монголии в районе гор Дзун-Арца (Воронин и др., 1983), где в течение атдабанского–раннеботомского веков накопилась толща малиновых глинистых плитчатых известняков с отдельными водорослево-археоциатовыми постройками, рядом с которыми обитали патериниды рода Dzunarzina (семейство Cryptotretidae). Встречающиеся здесь их створки нередко имеют хорошо сохранившуюся микроструктуру (Ушатинская, 1993).
В статье, посвященной посмертной минерализации органики в раковинах фосфатных брахиопод из кембрия Сибирской платформы, были рассмотрены более подробно причины сохранения в ископаемом состоянии органических структур (Ушатинская, 2018). Такие брахиоподы, при жизни обитавшие у дна в местах накопления глинисто-карбонатных осадков, после смерти погружались в илистые поровые воды ниже аэробно-анаэробной границы. В последующих преобразованиях определяющую роль играло несколько факторов: ограниченный доступ кислорода, что замедляло разложение органики, присутствие бактерий, способствовавших частичному переходу содержавшегося в мягких телах и раковинах ионов фосфора и кальция в растворимое состояние, небольшое повышение щелочности. В результате при росте их концентрации начиналось замещение вновь образующимся апатитом тонких органических структур: наружных покровов створок, органических слоев и включений в органоминеральных слоях, клеток внешнего эпителия мантии, прилегающих к внутренней поверхности створок. Все это способствовало консервации створок и, в результате, сохранению микроструктуры в ископаемом состоянии.
На примере створок Pelmanotreta можно также видеть, что, вероятно, даже небольшие отклонения в условиях захоронения приводили к разному типу сохранности. Чаще всего встречаются створки, у которых вся внутренняя поверхность покрыта сферолитами (рис. 4, в, г; 5, о). Вероятно, разложение–растворение мягкого тела и органического содержимого створок при содействии бактерий происходило достаточно быстро, и в условиях перенасыщения раствора ионами кальция и фосфора осуществлялось осаждение апатита, который, не успев осесть на быстро растворившиеся тонкие органические слои, кристаллизовался в виде сферолитов в освободившемся пространстве. Последние состоят из радиально-ориентированных игольчатых кристаллов, расходящихся лучами от центра (рис. 4, в, г). По данным А.А. Годовикова и др. (1989), для образования сферолитов как раз необходимы условия перенасыщения, способствующие успешному росту и расщеплению зародышевых кристаллов. Но в этой же пестроцветной свите имеются образцы Pelmanotreta, у которых на внутренней поверхности створок сохранился фосфатизированный наружный слой мантийного эпителия со слепками оболочек клеток, из которых он состоял (рис. 2, м; 3, д, е). Похожие слепки клеточных оболочек мантийного эпителия, тесно прилегавшего к внутренней поверхности створок, сохранились и у монгольских родов Dzunarzina и Salanygolina (табл. V , фиг. 3; рис. 3, а–г). Призмы, образованные клетками мантийного эпителия в ходе секреции “мембран” вторичного слоя створок, тоже нередко сохраняются (табл. V , фиг. 1в, 2в, 2г; рис. 4, л). Но встречаются створки Pelmanotreta, у которых участки отдельных ламин вторичного слоя сложены не плотными, тесно прилегающими друг к другу оболочками полых внутри призм, а сохранилось только их минерализованное содержимое. На рис. 4, д–ж можно видеть, что поверхности ламин вторичного слоя, лежащие под отсутствующим первичным, состоят из мелких, слегка округленных призм 5–6 мкм в поперечнике, которые окружены тонкими пустыми пространствами, соответствующими прежним их оболочкам. Отдельные призмы снаружи сложены плотным тонким слоем из мелких округлых или пластинчатых кристаллов, а в середине призмы полые. Возможно, растворенного содержимого призм хватило только на образование наружного слоя (рис. 4, ж). Скорее всего, такие различия в типах сохранности зависели от очень тонких колебаний рН, концентрации фосфата кальция, наличия кислорода в микроокружении, активности бактерий при захоронении створок. Также и тонкие при жизни, скорее всего органические прослои, разделявшие пачки ламин, сложенных призмами, в одних случаях состоят из очень мелких плотных сферолитов, а в других от них остались лишь полые протяженные щели.
Створки родов Aldanotreta и Tumulduria происходят из местонахождений Улахан-Сулугур и Дворцы, расположенных на левом берегу р. Алдан. Их остатки известны не из тонкослоистых глинистых известняков (как Pelmanotreta и Dzunarzina), а из известняков со значительной примесью обломочного материала и с многочисленными зернами глауконита (Ярусное расчленение …, 1983). Скорее всего, в данном районе была более высокая динамическая активность воды у дна, и створки брахиопод после смерти подвергались более существенному разрушению, перемещению и вторичным изменениям. Вероятно, близкие условия накопления были и во время формирования нижней части еркекетской свиты на севере Сибирской платформы, которая в низах сложена известковистыми песчаниками и гравелитами (Сухов и др., 2016). Оттуда происходят несколько образцов Tumulduria sp. Чаще всего в ходе растворения обогащенных терригенным материалом известняков в слабой уксусной кислоте выделяются мелкие многочисленные обломки створок. Несколько почти целых створок родов Aldanotreta и Tumulduria были обнаружены Ю.Л. Пельманом и И.В. Коровниковым в разрезах р. Алдан и любезно переданы автору (табл. IV , фиг. 1а, 8). У целых створок обычно наружный слой сильно поврежден, многие обломки несут на поверхности мелкие сферолиты, а на сколе по всей толщине его и на внутренней поверхности видны более крупные сферолиты. Как уже говорилось, скорее всего, у этих, самых древних из известных брахиопод, количество органического вещества в раковинах могло быть больше, чем у более поздних, и разлагавшийся и растворявшийся органо-фосфатный материал создавал перенасыщенный раствор, из которого и осаждались скопления сферолитов. На одном обломке створки, принадлежащей роду Tumulduria, можно видеть, как могло происходить отложение новообразованного апатита на отдельных участках, располагавшихся на внутренней или наружной поверхностях створок, или в их середине. С наружной и внутренней сторон створки для кристаллизации имелось достаточно свободного пространства, и там апатит сформировался в виде радиально лучистых, сплошных сферолитов (табл. IV , фиг. 9 а, 9б; рис. 4, к). А в центральной части скола створки, там, где она была построена призмами, в их замкнутых пространствах, кристаллизовались тоже округлые сферолитоподобные образования, но часто полые внутри. Их наружная оболочка одно-, двух- или многослойная; отдельные слои имеют толщину около 1–2 мкм и состоят из мелких игольчатых кристаллов. Россыпь мелких (менее 0.5 мкм) кристаллов иногда видна на внутренней поверхности (рис. 4, и, л). Рядом с этими округлыми структурами лежат небольшие участки с сохранившимися минерализованными перегородками между призмами. В “Геологическом словаре” (1973, с. 208) подобные образования, рост которых происходит от периферии к центру (а не от центра к периферии, как у сферолитов), названы секрециями, если они крупные, или миндалинами, если их размеры достигают первых миллиметров. Кристаллизация этих образований происходит от периферии к центру путем заполнения пустот, начиная со стенок. И хотя в нашем случае величина описываемых структур еще на три порядка меньше, чем указано для миндалин, их появление тоже, скорее всего, связано с частичным или полным заполнением призм, слагавших вторичный слой створок, начиная с их стенок.
Возможно, фоссилизация створок и их обломков в нестабильных условиях проходила в несколько этапов. Сначала хорошо сформированные сферолиты образовывались на внутренних и внешних поверхностях створок, где была высокая концентрация органо-фосфатного раствора и больше свободного места для их осаждения; во многих случаях мелкими сферолитами замещалась и вся стенка раковины. Но иногда органо-фосфатный раствор оставался внутри замкнутого пространства призм, частично уже ставших полыми, и тогда мелкие игольчатые кристаллы апатита могли оседать рядом с полуразрушенными их стенками; для полного заполнения призм, вероятно, было недостаточно материала. Но, по мнению Э.М. Спиридонова (устн. сообщ.), не исключено, что сферолитоподобные по форме, полые внутри структуры могли появиться и в результате отложения из раствора органо-фосфатного материала в виде игольчатых кристаллов апатита на поверхностях возникших в микропространствах внутри створок пузырьков газа (например, СО2), или же на образовавшихся из раствора мельчайших зернах кальцита, позднее растворившихся.
Таким образом, изучение микроструктуры раковин и ее посмертных преобразований у ранне–среднекембрийских представителей обоих семейств отряда Paterinida показывает, что секреция раковины у них происходила сходным образом. Значительная часть вещества створок принадлежала органической составляющей, которая при захоронении оказывала существенное влияние на результат фоссилизации. Большую роль в посмертных преобразованиях микроструктуры играли условия, в которых раковины оказывались после смерти.
* * *
Данная работа поддержана Грантом РФФИ 19-04-01027, Программой Президиума РАН № 17 “Эволюция органического мира. Роль и влияние планетарных процессов” (Подпрограмма 1 “Развитие жизненных биосферных процессов”) и Министерством высшего образования и науки РФ. Автор очень благодарен И.В. Коровникову (ИНГиГ СО РАН, Новосибирск), передавшему материал с остатками раковин Aldanotreta из разрезов низов кембрия по р. Алдан и коллекцию патеринид из кембрия Сибирской платформы Ю.Л. Пельмана, сотрудникам ПИН РАН Е.А. Жегалло и Р.А. Ракитову за большую помощь при изучении материала с использованием электронного сканирующего микроскопа, а также проф. кафедры минералогии геологического фак-та МГУ Э.М. Спиридонову за консультации по проблемам минералообразования.
В работе были изучены брахиоподы из отряда Paterinida, собранные в разные годы на Сибирской платформе Ю.Л. Пельманом, И.В. Коровниковым (ИНГГ СО РАН), Р.О. Галабалой (ВАГТ), А.Ю. Журавлевым, Я.Е. Малаховской, А.Ю. Иванцовым и автором статьи (ПИН РАН), а также брахиоподы из нижнего кембрия Монголии, собранные Е.А. Жегалло, А.Ю. Журавлевым и автором в 80-е годы XX века. Весь изображенный материал хранится в Отделе фондов ПИН РАН, Сибирская колл. ПИН, № 5486, Монгольская колл. ПИН, № 3302.
Список литературы
Воронин Ю.И., Воронова Л.Г., Дроздова Н.А. и др. Докембрий–кембрийские отложения разреза Дзун-Арца в Западной Монголии // Бюлл. МОИП. Отд. геол. 1983. Т. 58. Вып. 2. С. 53–66.
Геологический словарь. Т. 2. М.: Недра, 1973. 455 с.
Годовиков А.А., Рипинен О.И., Степанов В.И. Сферолиты, сферокристаллы, сфероидолиты, ядросферолиты // Новые данные о минералах СССР. М.: Наука, 1989. С. 24–36 (Тр. Минерал. музея АН СССР. Вып. 36).
Горянский В.Ю. Класс Inarticulata. Беззамковые // Основы палеонтологии. Мшанки, Брахиоподы. М.: Изд‑во АН СССР, 1960. С. 172–182.
Горянский В.Ю., Попов Л.Е. Морфология, систематическое положение и происхождение беззамковых брахиопод с карбонатной раковиной // Палеонтол. журн. 1985. № 3. С. 3–14.
Граница докембрия и кембрия в геосинклинальных областях (опорный разрез Саланы-Гол, МНР). М.: Наука, 1982. 152 с. (Тр. Совм. Сов.-Монгол. палеонтол. экспед. Вып. 18).
Пельман Ю.Л. Ранне- и среднекембрийские беззамковые брахиоподы Сибирской платформы. Новосибирск: Наука, 1977. 168 с. (Тр. ИГиГ СО АН СССР. Вып. 316).
Попов Л.Е., Ушатинская Г.Т. Новые данные о микроструктуре раковины беззамковых брахиопод отряда Paterinida // Докл. АН СССР. 1987. Т. 293. № 5. С. 1228–1230.
Путеводитель экскурсии по рекам Алдану и Лене (Международная экскурсия по проблеме границы кембрия и докембрия). М.-Якутск: ВИЭМС, 1973. 118 с.
Сухов С.С., Шабанов Ю.Я., Пегель Т.В. и др. Стратиграфия нефтегазоносных бассейнов Сибири. Кембрий Сибирской платформы. Т. 1 Стратиграфия. Новосибирск: ИНГГ СО РАН, 2016. 497 с.
Ушатинская Г.Т. Необычные беззамковые брахиоподы из нижнего кембрия Монголии // Палеонтол. журн. 1987. № 2. С. 62–68.
Ушатинская Г.Т. Новый род патеринид (брахиоподы) из нижнего кембрия Западной Монголии // Палеонтол. журн. 1993. № 1. С. 115–118.
Ушатинская Г.Т. Древнейшие лингуляты. М.: Наука, 1995. 89 с. (Тр. Палеонтол. ин-та РАН. Т. 262).
Ушатинская Г.Т. Сохранение минерализованных мягких тканей и их отпечатков в раковинах древних фосфатных брахиопод // Палеонтол. журн. 2018. № 5. С. 23–27.
Ярусное расчленение нижнего кембрия. Стратиграфия. М.: Наука, 1983. 184 с.
Emig Ch.C. Conditions de fossilization du genera Lingula (Brachiopoda) et implication palaeoecologiques // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1986. V. 53. № 2–4. P. 245–253.
Holmer L.E., Popov L.E. Class Lingulata // Treatise on Invertebrate Paleontology: Part H. Brachiopoda, Revised. V. 2: Linguliformea, Craniiformea, and Rhynchonelliformea (Part) / Ed. Kaesler R.L. Boulder–Lawrence: Geol. Soc. Amer. – Univ. Kansas Press, 2000. P. 30–146.
Holmer L.E., Popov L.E. Paterinata // Treatise on Invertebrate Paleontology: Part H. Brachiopoda, Revised. V. 6: Suppl. / Ed. Kaesler P.A. Boulder–Lawrence: Geol. Soc. Amer. – Univ. Kansas Press, 2007. P. 2578–2579.
Holmer L.E., Stolk S.P., Skovsted Ch. B. et al. The enigmatic Early Cambrian Salanygolina – a stem group of rhynchonelliform chileate brachiopods? // Palaeontology. 2009. V. 52. Pt 1. P. 1–10.
Laurie J.R. Class Paterinata // Treatise on Invertebrate Paleontology: Part H. Brachiopoda, Revised. V. 2: Linguliformea, Craniiformea, and Rhynchonelliformea (Part) / Ed. Kaesler R.L. Boulder–Lawrence: Geol. Soc. Amer. – Univ. Kansas Press, 2000. P. 147–156.
Rowell A.J. Inarticulata // Treatise of Invertebrate Paleontology: Part H. Brachiopoda. V. 1 / Ed. Moore R.C. Boulder–Lawrence: Geol. Soc. Amer. – Univ. Kansas Press, 1965. P. 260–297.
Scovsted Ch., B., Kuchinsky A., Bengtson S., Holmer L.E. The problematic early Cambrian fossil Tumulduria compecta represents the detached ventral interarea of a paterinid brachiopod // Acta Palaeontol. Pol. 2014. V. 59. № 2. P. 359–365.
Skovsted Ch.B., Ushatinskaya G., Holmer L.E. et al. Taxonomy, morphology, shell structure and early ontogeny of Pelmanotreta nom. nov. from the Lower Cambrian of Siberia // GFF. 2015. V. 137. № 1. P. 1–8.
Williams A., Carlson S.J., Brunton C.H.C. et al. A supra-ordinal classification of the Brachiopoda // Phil. Trans. Roy. Soc. Lond. B. 1996. V. 351. P. 1151–1193.
Williams A., Cusack M., Mackay S. Collagenous chitinophosphatic shell of the brachiopod Lingula // Phil. Trans. Roy. Soc. Lond. B. 1994. V. 346. P. 223–263.
Williams A., Holmer L.E. Ornamentation and shell structure of acrotretoid brachiopods // Palaeontology. 1992. V. 35. Pt 2. P. 657–692.
Williams A., Popov L.E., Holmer L.E., Cusack M. The diversity and phylogeny of the paterinate brachiopods // Palaeontology. 1998. V. 41. Pt 2. P. 221–262.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал