

Палеонтологический журнал, 2019, № 5, стр. 54-62
Magnofossacrinus – новый род кладидных криноидей (Crinoidea, Echinodermata) из московского яруса (пенсильваний) Подмосковья
Г. В. Миранцев *
Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
* E-mail: GMirantsev@gmail.com
Поступила в редакцию 17.08.2018
После доработки 16.09.2018
Принята к публикации 14.11.2018
Аннотация
Из пенсильвания Московской синеклизы описан новый род потериокринидых морских лилий – Magnofossacrinus, представленный типовым видом – M. domodedovoensis sp. nov.. Новый род характеризуется необычно крупными циррусами с полинодальной фасеткой и является наиболее молодым достоверным представителем семейства Poteriocrinidae, известным из московского яруса. Обсуждена и сравнена морфология стеблей Magnofossacrinus со стеблями других близкородственных морских лилий.
Целые экземпляры морских лилий встречаются значительно реже изолированных элементов скелета – члеников и фрагментов стеблей, табличек чашечки и брахиалей. Для отдельных члеников стеблей морских лилий была разработана формальная систематика (паратаксономия), которая широко использовалась для биостратиграфического расчленения палеозойских отложений (Елтышева, 1956, 1959; Стукалина, 1965, 2000; Moore, Jeffords, 1968 и др.). Важной задачей, благодаря решению которой можно получить новые сведения о распространении таксонов криноидей, является сопоставление изолированных фрагментов с целыми скелетами морских лилий, классифицируемыми в естественной системе. Определение изолированных фрагментов стеблей и соотнесение их с кронами обычно затруднены как малым набором диагностических признаков, так и сходством в строении стеблей даже у неродственных таксонов. Поэтому для этих целей легче всего использовать таксоны с необычной, ярко выраженной морфологией стебля.
В подольском горизонте московского яруса, особенно в улитинской и щуровской свитах Подмосковья, где находки целых идентифицируемых чашечек и крон морских лилий редки (Mirantsev, Rozhnov, 2012; Mirantsev, 2015), весьма обычны членики, а также отдельные фрагменты стеблей, отличающиеся от остальных, встречающихся в этих же отложениях, наличием необычно крупных фасеток для прикрепления циррусов. Циррусы у этих морских лилий массивные, примыкают к стеблю под острым углом и, судя по диаметру циррусной фасетки, могут достигать более 3/4 диаметра стебля, чем заметно отличаются от большинства остальных средне-позднекаменноугольных криноидей. Фасетка для прикрепления одного цирруса занимает обычно несколько члеников стебля. Идентичные фрагменты стеблей со сходной морфологией были обнаружены в разрезе Домодедовского карьера. Имеющиеся в коллекции экземпляры происходят из нижней части слоя 26 (нумерация по: Goreva et al., 2009), относящегося к домодедовской свите мячковского горизонта. Слой представлен известковой глиной, листоватой, с биокластами хорошей сохранности. Среди них нередки целые кроны морских лилий [Cromyocrinus simplex Trautschold, Dicromyocrinus ornatus (Trautschold), Mooreocrinus geminatus (Trautschold), Moscovicrinus multiplex (Trautschold), Pegocrinus bijugus (Trautschold) и др.] и целые панцири морских ежей Archaeocidaris rossica (von Buch) и еще не описанного представителя семейства Lepidesthidae. Из этого же слоя происходят несколько крон (4 экз. хранятся в коллекциях ПИН РАН и несколько – в частных коллекциях) криноидей, относящихся к новому роду и найденных в ассоциации с указанными стеблями. На некоторых экземплярах крон сохранились проксимальные части стебля, имеющие сходную морфологию, что и найденные отдельно стебли. Таким образом, с достаточно большой вероятностью можно соотнести изолированные членики и фрагменты стеблей с целыми кронами. Примечательно, что в других одновозрастных местонахождениях мячковского горизонта кроны этих морских лилий, а также фрагменты стеблей (за исключением единичной находки из Касимовского карьера на Окско-Цнинском валу) не обнаружены. Массовые находки этих криноидей в домодедовской свите Домодедовского карьера, по-видимому, связаны не с широким стратиграфическим распространением этого рода, а с уникальным захоронением массового поселения, поскольку все они найдены в одном слое и в одной части карьера (С.В. Гришин, перс. сообщ.).
При написании данной работы был использован материал, хранящийся в Палеонтологическом ин-те им. А.А. Борисяка РАН, собранный в разное время А.П. Ивановым (ПИН, колл. № 137), В.Л. Карчевским (ПИН, колл. № 5348, сборы 1990-х гг.), С.В. Гришиным (ПИН, колл. № 5362, сборы 1995 г.), П.Б. Кабановым, А.В. Ступаченко, Р.П. Широковым и В.А. Горбенко, сотрудниками Московского детско-юношеского центра экологии, краеведения и туризма (МДЭБЦ, ранее МосГорСЮН), а также автором (ПИН, колл. № 5450; сборы 1990–2014 гг.).
КЛАСС CRINOIDEA
ОТРЯД CLADIDA
ПОДОТРЯД DENDROCRININA
СЕМЕЙСТВО POTERIOCRINIDAE AUSTIN ET AUSTIN, 1842
Типовой род – Poteriocrinites Miller, 1821; средний девон – средний миссисипий, имеет широкое распространение (Webster, 2013).
Диагноз. Корона большая. Чашечка от умеренно- до высококонической, с тремя или четырьмя анальными табличками; радиальные фасетки занимают не всю ширину радиальной таблички, с хорошо развитым поперечным гребнем; анальный мешок крупный, удлиненный, обычно с шиповидными табличками на вершине, с порами или щелями на границах табличек. Руки ветвятся изотомически один или несколько раз, с пиннулами.
Состав. Помимо типового, роды Balearocrinus Bourrouilh et Termier, 1973, миссисипий Испании и Алжира; Denariocrinus Schmidt, 1942, ранний девон Германии; Magnofossacrinus gen. nov., пенсильваний Подмосковья; Ornatuscrinites Hauser, 2007, средний девон Германии; Rhabdocrinus Wright, 1944, миссисипий США, Великобритании, Испании и Подмосковья; и Springericrinus Jaekel, 1918, миссисипий США.
Замечания. В традиционной систематике (Moore et al., 1978) семейство потериокринид рассматривают в составе подотряда Poteriocrinina, объединяющего всех пиннульных кладидных криноидей. В последнее время считается, что наличие пиннул не является надежным признаком для объединения кладидных морских лилий в отдельный подотряд, поскольку пиннулы могли возникнуть параллельно у нескольких палеозойских групп морских лилий. Г. Вебстер с соавт. (Webster et al., 2004) включили представителей семейства потериокринид в состав подотряда Cyathocrinina на основании сходной морфологии (строение радиальных фасеток, чашечки, рук и тегмена). В других работах (McIntosh, 2001; Webster, Lane, 2007), потериокриниды рассматривается в составе отряда дендрокринид. В этой работе принята последняя точка зрения.
В сводке “Treatise …” (Moore et al., 1978), помимо вышеуказанных таксонов, в составе семейства Poteriocrinidae также рассматривается род Propoteriocrinus Schmidt, 1942. По мнению современных исследователей, этот род ближе к представителям семейств Glossocrinidae Goldring, 1923 (Webster et al., 1999) или Rutkowskicrinidae (McIntosh, 2001).
Род Magnofossacrinus Mirantsev, gen. nov.
Название рода по крупным циррусным цоколям на стеблях, характеризующих таксон.
Типовой вид – Magnofossacrinus domodedovoensis sp. nov.
Диагноз. Потериокриниды средних размеров. Крона крупная, удлиненная (рис. 1, а). Чашечка высококоническая с вытянутым основанием. Таблички чашечки тонкие. Инфрабазальный венчик состоит из пяти одинаковых, высоких табличек. Пять базальных табличек, базалии примерно одинаковой высоты с инфрабазалиями. Пять радиальных табличек, радиальные фасетки занимают большую часть ширины радиальной таблички. Три анальные таблички в составе чашечки. Анальный мешок крупный, высокий, цилиндрический. Руки однорядные, ветвятся изотомически два раза. Стебель и осевой канал в поперечном сечении округлой формы. Циррусы располагаются на протяжении всего стебля. Нодальный членик с характерным выступом, несущим основание крепления циррусного цоколя (рис. 2, д). Вогнутый циррусный цоколь располагается в верхней кромке оттянутой нодалии и обычно захватывает несколько (до трех) супранодалей. Циррусный цоколь округлой формы. Его размер увеличивается дистально: на дистальных участках стебля диаметр цоколя может составлять более 3/4 диаметра стебля.
Рис. 1.
Magnofossacrinus gen. nov.: a – реконструкция кроны M. domodedovoensis sp. nov.; б – развертка чашечки с проксимальными частями рук M. domodedovoensis sp. nov., oсновано на экз. ПИН, № 5348/190; pеконструированные части (показаны пунктиром) основаны на других экземплярах; pадиальные таблички закрашены черным цветом, радианальная табличка закрашена штриховкой, X- и RX-анальные таблички закрашены точками; в, г – Magnofossacrinus sp., экз. ПИН, № 5450/2422, раздавленная чашечка с проксимальными частями рук и стебля, точное местонахождение и возраст неизвестны; д, е – M. domodedovoensis gen. et sp. nov., экз. ПИН, № 5348/190, раздавленная чашечка с проксимальными частями рук и стебля (экземпляр покрыт хлористым аммонием); Домодедовский карьер; средний карбон, московский ярус, мячковский горизонт, песковская свита. Длина масштабного отрезка – 10 мм.
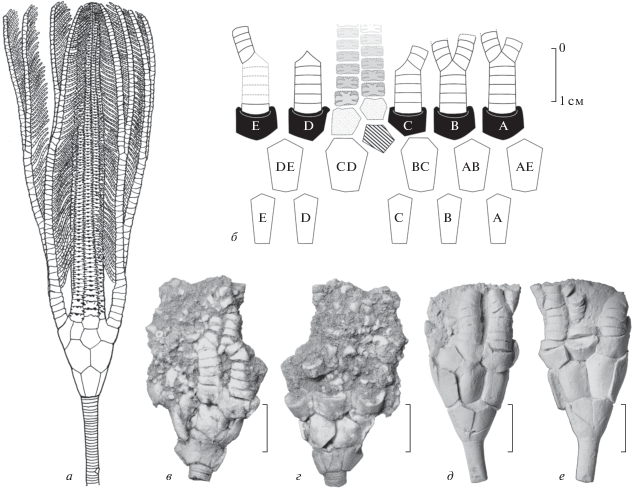
Рис. 2.
Детали строения стебля и циррусов Magnofossacrinus sp.: а – экз. ПИН, № 5450/2502, фрагмент стебля из трех члеников с циррусной фасеткой; б – экз. ПИН, № 5450/2501, нодальный членик, вид сверху; в, г – экз. ПИН, № 5450/2507, фрагмент стебля из двух члеников с циррусной фасеткой; д – экз. ПИН, № 5450/2505, нодальный членик; е – экз. ПИН, № 5450/2504, нодальный членик, вид снизу; ж–и – экз. ПИН, № 5450/2503, фрагмент стебля ?ювенильного экземпляра с циррусом; к, л – экз. ПИН, № 5450/2443, фрагмент стебля с циррусной фасеткой со вздутиями и сверлениями, вид сбоку и сверху; м–о – экз. ПИН, № 5450/2514, фрагмент стебля из четырех члеников, включая нодальный; п, р – экз. ПИН, № 5450/2441, фрагмент стебля из четырех члеников, с проксимальной частью цирруса, сверху и сбоку; с–у – экз. ПИН, № 5450/2440, фрагмент стебля с крупным циррусом; ф – экз. ПИН, № 5450/2508, фрагмент стебля из 11 члеников с циррусом, видна инкрустирующая мшанка; х – тот же экземпляр, вид с другой стороны, видны многочисленные прижизненные сверлениями, сопровождающиеся деформациями стебля и “царапинами”; ц – экз. ПИН, № 5450/2509, пришлифованный фрагмент стебля, c циррусной фасеткой; ч – экз. ПИН, № 5450/2442, фрагмент стебля из девяти члеников; ш – экз. ПИН, № 5450/2510, фрагмент стебля небольшого экземпляра со сверлениями и прижизненными деформациями стереома; карьеры Пирочи (к, л, п–у) и Акатьево (а–и, м–о, ф–ш); cредний карбон, московский ярус, подольский горизонт, улитинская и щуровская свиты. Длина масштабного отрезка – 1 мм (б), 2 мм (в остальных случаях). Все образцы, кроме экз. ПИН, № 5450/2509 (ц), покрыты хлористым аммонием. Oбозначения: a.c. – осевой канал; c.c. – осевой канал цирруса.

Видовой состав. Типовой вид из московского яруса Московской синеклизы и Окско-Цнинского вала.
Сравнение. От остальных родов потериокринид отличается более удлиненной формой чашечки, кроной с меньшим (по четыре ветвления в каждом радиусе) числом ветвлений рук (меньше только у девонского Denariocrinus c 10 ветвями), массивными циррусами.
Замечания. В колл. ПИН РАН имеется чашечка Magnofossacrinus с основанием рук (рис. 1, в, г; экз. ПИН, № 5450/2422: сборы П.Б. Кабанова, экземпляр передан А.С. Алексеевым). К сожалению, этикетка с указанием местонахождения отсутствует; тем не менее, характер сохранности, цвет и тип вмещающей породы данного образца существенно отличаются от таковых у экземпляров, известных из типового местонахождения. Вероятно, что этот экземпляр происходит из более древних отложений подольского горизонта, где стебли этих криноидей весьма обильны. С этим же экземпляром ассоциирована изолированная брюшная створка брахиоподы Tegulispirifer tegulatus (Trautschold). Форма и сохранность раковины этой брахиоподы (толстостенная окремнелая створка со слабой скульптурой) также подтверждают подольский возраст (Сарычева, Сокольская, 1952).
Стебли Magnofossacrinus присутствуют во многих местонахождениях подольского горизонта Московской синеклизы (рис. 2). Однако отсутствие ассоциированных находок крон не позволяет определить данные фрагменты стеблей точнее, чем Magnofossacrinus sp.
Magnofossacrinus – один из наиболее молодых представителей семейства, известный по целым кронам. Остальные пенсильванские и пермские потериокриниды описаны по отдельным чашечкам или фрагментарному материалу (Webster, Lane, 2007). Вероятно, новый род произошел от раннекаменноугольных представителей семейства путем педоморфоза. Об этом свидетельствуют меньшее количество ветвлений в кроне, пропорционально более удлиненные брахиальные членики и общая грацильность в строении кроны.
Magnofossacrinus domodedovoensis Mirantsev, sp. nov.
Табл. VIII , фиг. 1–4; табл. IX, фиг. 1–3 (см. вклейку)
Рис. 3.
Сравнение стеблей Magnofossacrinus gen. nov. со стеблями других каменноугольных кладидных криноидей: а, б – Magnofossacrinus; в, г – Unilineatocrinus (= ?Rhabdocrinus); д – Moscovicrinus. Длина масштабного отрезка – 5 мм.
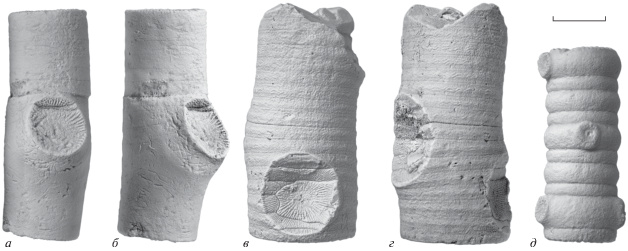
Рис. 4.
Реконструкция внешнего вида Magnofossacrinus domodedovoensis sp. nov. (рисунок Г.А. Анекеевой по наброску автора).

Название вида по типовому местонахождению.
Голотип – ПИН, № 5362/83 (табл. IX , фиг. 2), крона хорошей сохранности с частично сохранившимся анальным мешком и проксимальной частью стебля; Московская обл., Домодедовский карьер возле д. Киселиха и д. Новленское; средний карбон, московский ярус, мячковский горизонт, домодедовская свита, слой 26 (по: Goreva et al., 2009).
Описание (рис. 1, a, б, д, е; 3, а, б; 4). Чашечка высоко-конической формы с удлиненным основанием. В инфрабазальном венчике пять крупных вытянутых табличек одинакового размера. В базальном венчике пять табличек: базалии AB, AE и DE примерно равного размера, шестиугольной формы; базалии CD и BC несколько крупнее остальных, семиугольной формы (рис. 1, б). Пять пятиугольных радиальных табличек. Радиальная фасетка занимает примерно 80% ширины радиалии. Таблички чашечки тонкие, в связи с этим кроны редко сохраняются в недеформированном виде: чашечки у всех имеющихся в коллекции экземпляров раздавлены.
Анальные таблички на одних экземплярах сохранились лишь частично, на других скрыты в породе. Судя по сохранившимся табличкам и фасеткам, в состав чашечки входили три анальные таблички. Анальная табличка X была примерно одинаково размера с радианальной табличкой или, возможно, немного превосходила ее (экз. ПИН, № 5450/2422, табл. IX , фиг. 3). Радианальная табличка пятиугольная, ее длина сопоставима с высотой. Анальный мешок хорошо развит, высокий, цилиндрический, состоит из нескольких рядов мелких складчатых табличек. Размер табличек на задней стороне мешка меньше, чем на передней (табл. IX , фиг. 2), в дистальной части мешка размер табличек значительно уменьшается (табл. IX , фиг. 1б). Высота мешка немного уступает длине рук.
Руки ветвятся изотомически, по-видимому, два раза (дистальные части рук неизвестны). Первый раз ветвление происходит на пятых-седьмых примибрахиалях. У голотипа (табл. IX , фиг. 2) второе ветвление происходит на 29 секундибрахиалях. Точное количество тертибрахиалей неизвестно, вероятно их было не менее 12. Примибрахиали низкие, прямоугольного очертания. Секундибрахиали, начиная с третьих, и тертибрахиали приобретают клиновидное очертание. Аксилярные примибрахиали и секундибрахиали треугольной формы. Пиннулы длинные и тонкие (табл. IX , фиг. 1а), расположены поочередно слева и справа на каждой брахиали, кроме аксилярных.
Скульптура на поверхности табличек чашечки или брахиалей отсутствует.
Стебель в поперечном сечении круглый, в проксимальной части слегка расширяется. Стебель однопорядковый. Экземпляры с целыми стеблями неизвестны, однако, исходя из имеющихся фрагментов, можно предположить, что весь стебель достигал в длину около одного метра. Толщина стебля была неравномерной, наиболее тонкая часть приходилась на проксимальную часть стебля, и самого большого диаметра на дистальную его часть (3.6 и 10 мм соответственно для взрослых экземпляров).
На большинстве из сохранившихся фрагментов стеблей, в том числе и на самых проксимальных, примыкающих к кроне (табл. VIII , фиг. 1; табл. IX , фиг. 2, 3), присутствуют циррусы. Вероятно, циррусы располагались на всем протяжении стебля, но на значительном удалении друг от друга. В районе проксистеллы циррусы имеют “зачаточное” состояние в виде небольших бугорков или маленьких циррусов, состоящих, судя по всему, всего из нескольких цирралей (табл. IX , фиг. 2, 3). Фасетка проксимально расположенных циррусов примыкала к одному нодальному членику стебля; дистально вниз по стеблю размер фасеток (как и величина самих циррусов) увеличивался, захватывая соседние (до 4–5) супранодальные членики. В районе мезостеллы присутствуют более крупные циррусы, состоящие из более чем 20 цирралей с диаметром циррусной фасетки равным или превышающим половину диаметра стебля. Наиболее крупные и длинные циррусы располагались, по всей видимости, в дистальной части стебля (рис. 4). Высота цирралей уменьшается проксимально: наиболее короткие циррали расположены вблизи от фасетки.
Циррусы примыкают под углом к стеблю, основанием, направленным вверх в сторону кроны (табл. VIII ). В области ответвления цирруса стебель слабо расширяется. Густота циррусов увеличивается дистально.
Осевой канал широкий (от 1/4 до 1/3 ширины стебля). Канал цирруса существенно уже осевого канала (рис. 2, б).
Материал. Помимо голотипа, чашечка (экз. ПИН, № 5348/190) и две кроны (экз. ПИН, №№ 5450/2423 и 5450/2424), а также 11 фрагментов стебля из типового местонахождения.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ И ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
Для большинства продвинутых представителей подкласса кладидных морских лилий, являвшихся доминантными во многих пенсильванских и пермских криноидных сообществах, характерны небольшие тонкие циррусы с унинодальным сочленением (рис. 3, д). Этот тип сочленения, при котором фасетка цирруса занимает пространство только одного членика (нодального), является наиболее распространенным среди криноидей, в том числе и современных стебельчатых. Magnofossacrinus, напротив, характеризуется полинодальным сочленением циррусов, при котором циррусная фасетка примыкает к нескольким соседним к нодальному членикам. Массивные циррусы, характеризующиеся полинодальным сочленением, отмечены в большей степени у камерат и флексибилий, а также у раннепалеозойских кладид (Brett, 1981). Большинство криноидей, для которых отмечено полинодальное сочленение циррусов, характеризуются достаточно крупными размерами (стебель более метра в длину). Полинодальный тип сочленения позволяет увеличивать размер циррусов, необходимых для поддержания массивных стеблей крупных морских лилий палеозоя.
Другие представители семейства потериокринид, к которым относится Magnofossacrinus, также обладали циррусами с полинодальным типом сочленения (напр., Rhabdocrinus; Donovan, Birtle, 2011). Однако для этого семейства в целом характерны массивные и сравнительно более толстые стебли со значительно более широким осевым каналом по сравнению с таковым у Magnofossacrinus. С. Донован и М. Байртл (Donovan, Birtle, 2011) указали на наличие в дистальной части стебля Rhabdocrinus ветвящихся радиксов. Между тем, присутствие ветвящихся структур на дистальной части стебля Rhabdocrinus, судя по материалу из нижнего карбона Подмосковья, не отмечено. По-видимому, данное указание ошибочно, и дистальная часть стебля Rhabdocrinus, как и у остальных потериокринид, представляла собой холдфаст ризоидного типа с массивными циррусами (Brett, 1981, рис. 1, E). В стешевском горизонте нижнего карбона Московской синеклизы вместе с чашечками и табличками Rhabdocrinus часто встречаются массивные стебли, отнесенные к паратаксону Unilineatocrinus (Arendt, 2002), по морфологии сходные со стеблями Rhabdocrinus (рис. 3, в, г). Этот паратаксон характеризуется присутствием полинодального типа сочленения циррусов.
Еще одной отличительной особенностью стеблей Magnofossacrinus является редкость в расположении циррусов. На всех имеющихся фрагментах стеблей расстояние между соседними циррусами составляет не менее 24 члеников (табл. VIII ), тогда как у большинства родственных потериокринид (Poteriocrinites, Rhabdocrinus) циррусы располагались довольно густо (рис. 3, в, г). Циррусы, расположенные в проксимальной части стебля, ввиду своего небольшого размера, судя по всему, не несли опорной функции (рис. 4; табл. VIII , фиг. 1). Проксимальная часть стебля вместе с кроной были приподняты и находились в вертикальном положении. По мере роста стебля и появления новых члеников эти циррусы постепенно увеличивались в размере, сдвигаясь в дистальную сторону. В ходе онтогенеза стеблевая фасетка циррусов также увеличивалась, захватывая соседние членики стебля.
На дистальной части стебля циррусы, судя по всему, располагались более компактно, чем в других местах (табл. IX , фиг. 2). Некоторые циррусные фасетки на дистальной части стебля соизмеримы с диаметром самого стебля. Полинодальная фасетка опиралась при этом на 5–6 члеников стебля (рис. 2, с–у). Сами циррусы, расположенные на этих фасетках, вероятно, тоже были массивные и длинные, в отличие от циррусов в проксимальной части стебля.
Скорее всего, Magnofossacrinus обитали на мягких грунтах. Циррусы, расположенные на дистальной части стебля, несли опорную функцию, поддерживая стебель над субстратом и не давая ему погрузиться в осадок (рис. 4).
На некоторых, главным образом, дистальных фрагментах стебля Magnofossacrinus (экз. ПИН, №№ 5450/2443, 5450/2508, 5450/2510; рис 2, к, л, х, ш), а также на одном экземпляре кроны (табл. IX , фиг. 2) имеются неглубокие прижизненные сверления округло-параболической формы, нередко сопровождающиеся разрастанием стереома. Сходные сверления были неоднократно описаны в литературе под разными названиями: Myzostomites (Clarke, 1921), Tremichnus (Brett, 1985), Oichnus (Wisshak et al., 2015) и др. Такие сверления обычно неглубокие и не затрагивают осевой канал. По мнению Ю.А. Арендта (1961), эти сверления могли быть связаны с ихнородом Schizoproboscina, представляющим собой парные отверстия на руках морских лилий с сопровождающимся разрастанием скелетной ткани хозяина. Однако уверенных доказательств, устанавливающих связь сверлений Schizoproboscina с округлыми сверлениями на стеблях криноидей, пока нет. Диаметр сверлений на стеблях Magnofossacrinus несколько меньше аналогичных, встреченных у большинства других подмосковных средне-позднекаменноугольных криноидей (например, у Moscovicrinus). На некоторых экземплярах стеблей Magnofossacrinus сохранились прижизненно прикрепленные инкрустирующие мшанки (экз. ПИН, № 5450/2508; рис. 2, ф).
Автор признателен А.С. Алексееву (ПИН РАН, МГУ), А.В. Ступаченко, Р.П. Широкову, В.А. Горбенко, сотрудникам отдела палеонтологии МДЭБЦ и лично Д.Б. Кучеру и А.С. Шмакову (ПИН РАН) за переданный материал по Magnofossacrinus. Отдельную благодарность выражаю Г.А. Анекеевой (ПИН РАН) за рисунок реконструкции Magnofossacrinus. Автор благодарен А.С. Алексееву и С.В. Рожнову (ПИН РАН) за ценные замечания. Работа выполнена в рамках программы Президиума РАН “Биологическое разнообразие” и при финансовой поддержке РФФИ (проекты № 14-05-31464 и № 12-04-01750-а).
Список литературы
Арендт Ю.А. О повреждениях морских лилий, вызванных Schizoproboscina // Палеонтол. журн. 1961. № 2. С. 101–106.
Елтышева Р.С. Стебли морских лилий и их классификация // Вестн. ЛГУ. Сер. геол. и геогр. 1956. Вып. 2. С. 40–46.
Елтышева Р.С. Принципы классификации, методики изучения и стратиграфическое значение стеблей морских лилий // Вопросы палеобиологии и биостратиграфии. Тр. II сессии Всесоюз. палеонтол. об-ва / Ред. Степанов Д.Л. М.: Госгеолтехиздат, 1959. С. 230–235.
Сарычева Т.Г., Сокольская А.Н. Определитель палеозойских брахиопод Подмосковной котловины // Тр. ПИН АН СССР. 1952. Т. 38. 308 с.
Стукалина Г.А. О таксономическом значении стеблей древних морских лилий // Тр. Всес. н.-и. геол. ин-та, нов. сер. 1965. Т. 115. С. 210–218.
Стукалина Г.А. Криноидеи палеозоя. Практическое руководство по макрофауне России и сопредельных территорий. СПб.: Изд-во ВСЕГЕИ, 2000. 283 с.
Arendt Yu.A. Early Carboniferous echinoderms of the Moscow region // Paleontol. J. 2002. V. 36. Suppl. 2. P. 115–184.
Brett C.E. Terminology and functional morphology of attachment structures in pelmatozoan echinoderms // Lethaia. 1981. V. 14. № 4. C. 343–370.
Brett C.E. Tremichnus, a new ichnogenus of circular-parabolic pits in fossil echinoderms // J. Paleontol. 1985. V. 59. № 3. P. 625–635.
Clarke J.M. Organic dependence and disease: their origin and significance // Bull. New York State Museum. 1921. V. 221–222. P. 1–113.
Donovan S.K., Birtle M. The cladid crinoid Rhabdocrinus Wright from the Namurian of Co. Durham, UK // Proc. Yorkshire Geol. Soc. 2011. V. 58. № 3. P. 167–171.
Goreva N.V., Isakova T.N., Alekseev A.S. et al. Domodedovo Section // Neostratotype of the Moscovian Stage and Myachkovian Substage, in type and reference Carboniferous Section in the south part of the Moscow Basin: Field Trip Guidebook. Moscow, 2009. P. 65–90.
McIntosh G.C. Devonian cladid crinoids: Families Glossocrinidae Goldring, 1923, and Rutkowskicrinidae new family // J. Paleontol. 2001. V. 75. № 4. P. 783–807.
Mirantsev G.V. New data on the distribution of the Upper Carboniferous (Pennsylvanian) flexible crinoid Cibolocrinus (Crinoidea, Flexibilia) // Palaeoworld. 2015. V. 24. № 4. P. 470–478.
Mirantsev G.V., Rozhnov S.V. New data on Carboniferous crinoids from the Moscow Region // Zoosymposia. 2012. V. 7. P. 91–100.
Moore R.C., Jeffords R.M. Classification and nomenclature of fossil crinoids based on studies of dissociated parts of their columns // Univ. Kansas Paleontol. Contrib. 1968. Echinodernata Art. 9. P. 1–86.
Moore R.C., Lane G.N., Strimple H.L. Cladida // Treatise on Invertebrate Paleontology. Pt T. Echinodermata 2. V. 2 / Eds. Moore R.C., Teichert C. Lawrence: Geol. Soc. Amer. and Univ. Kansas, 1978. P. 572–759.
Webster G.D. Bibliography and index of Paleozoic crinoids, coronates, and hemistreptocrinoids. 2013. P. 1758–2011. URL: http://crinoids.azurewebsites.net (дата обращения 22.11.2018).
Webster G.D., Hafley D.J., Blake D.B., Glass A. Crinoids and stelleroids (Echinodermata) from the Broken Rib Member, Dyer Formation (Late Devonian, Famennian) of the White River Plateau, Colorado // J. Paleontol. 1999. V. 73. № 3. P. 461–486.
Webster G.D., Lane N.G. New Permian crinoids from the Battleship Wash patch reef in southern Nevada // J. Paleontol. 2007. V. 81. № 5. P. 951–965.
Webster G.D., Maples C.G., Sevastopulo G.D. et al. Carboniferous (Visean-Moscovian) echinoderms from the Bechar Basin area of western Algeria // Bull. Amer. Paleontol. 2004. № 368. 98 p.
Wisshak M., Kroh A., Bertling M. et al. In defence of an iconic ichnogenus–Oichnus Bromley, 1981 // Ann. Soc. Geol. Pol. 2015. V. 85. № 3. P. 445–451.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал