

Журнал общей биологии, 2022, T. 83, № 4, стр. 243-255
Особенности сообществ почвенных нематод в различных зонах фитогенного поля дерева
Д. С. Калинкина 1, *, А. А. Сущук 1, Н. В. Геникова 2
1 Институт биологии Карельского научного центра РАН, ФИЦ “КарНЦ РАН”
185910 Петрозаводск, Пушкинская, 11, Россия
2 Институт леса Карельского научного центра РАН, ФИЦ “КарНЦ РАН”
185910 Петрозаводск, Пушкинская, 11, Россия
* E-mail: kalinkinads@gmail.com
Поступила в редакцию 21.04.2022
После доработки 18.06.2022
Принята к публикации 04.07.2022
- EDN: IXIMYZ
- DOI: 10.31857/S0044459622040054
Аннотация
Представлены первичные данные об особенностях сообществ почвенных нематод во внутренних зонах фитогенного поля (приствольное повышение и подкроновое пространство) лиственницы сибирской Larix sibirica Ledeb. и ели обыкновенной Picea abies (L.) H. Karst., а также в межкроновом пространстве между деревьями. С помощью общепринятых нематологических методов были изучены следующие параметры: таксономическое разнообразие, плотность популяций, эколого-трофическая структура сообществ нематод. В результате исследования показано повышение общей численности, а также абсолютной и относительной плотности нематод, трофически связанных с растениями (паразиты растений, нематоды, ассоциированные с растениями), в фитогенном поле лиственницы по сравнению с елью. В эколого-трофической структуре сообществ нематод для лиственницы показана смена группы-доминанта при продвижении от ствола к периферии (от бактериотрофов к микотрофам), для ели тенденция не выявлена, вышеупомянутые группы имели одинаково высокий процент в обеих зонах. Отмечено повышение общей численности нематод и относительного обилия политрофов в приствольных зонах обоих деревьев по сравнению с подкроновыми пространствами и межкроновой зоной. Таким образом, полученные результаты косвенно позволяют говорить о том, что отличительные черты, характеризующие сообщества почвенных нематод, связаны с микроклиматическими условиями в зонах фитогенных полей, обусловленных влиянием деревьев.
Деревья выполняют значительную средообразующую роль в экосистеме. Обладая сильным фитогенным полем, они воздействуют на все элементы прилегающего пространства (освещенность, количество осадков, почвенные характеристики, растительность, почвенных беспозвоночных и др.), создавая тем самым гетерогенную среду обитания для организмов. Почвенные животные в значительной степени связаны с составом, структурой и продуктивностью растительных сообществ, а также с физической средой, сформированной растительным компонентом (Безкоровайная и др., 2017).
Под фитогенным полем понимается часть пространства, в пределах которого среда обретает отличительные свойства, определяемые присутствием в ней растения (Уранов, 1965). Для многих древесных растений наблюдаются сходные тенденции изменения абиотических и биотических факторов в различных зонах фитогенного поля. Так, для некоторых древесных пород показано, что температура и влажность почвы плавно понижаются по направлению от ствола дерева к периферии (Черняева, Викторов, 2016). Однако в зависимости от силы фитогенного поля деревья в разной степени трансформируют окружающую среду. Характер и степень влияния на прилегающее пространство обусловлены возрастными различиями, а также качеством и количеством опада (Ипатов, 2007). Например, для сосны обыкновенной показано, что сквозистость кроны увеличивается от ствола к краю кроны и отражается на количестве осадков и опада, а также на температурном режиме в подкроновом пространстве. Наиболее низкая температура наблюдается в приствольной зоне, она на 4–5°C ниже температуры на фоне. Также в приствольной зоне наблюдается снижение количества проникающих осадков по сравнению с фоновым и подкисление почвы (Журавлева и др., 2012). Для липы характерно снижение микробиологической активности под кроной по сравнению с сосновым древостоем. Взрослые липы образуют сильное затенение, что препятствует развитию живого напочвенного покрова и приводит к снижению микробной массы (Ашек, Тиходеева, 2006). Лиственница и ель являются мощными эдификаторами, они обладают сильными фитогенными полями (Ипатов и др., 2009; Прокушкин, Зырянова, 2013). В подкроновом пространстве данных пород, где мало света и влаги, создаются неблагоприятные условия для растений напочвенного покрова. Показано, что радиус фитогенного поля ели составляет 2–3 м (Демьянов, 1990). Ель возрастом 100 лет не оказывает подкисляющего эффекта на почвы, а возрастом 50 лет, напротив, способствует ее подкислению. Это связывают с несформированной кроной у молодого дерева и, соответственно, меньшей аккумуляцией кальция в опадающей хвое, а также большей пропускной способностью кроны, что содействует вымыванию оснований из почвы (Лукина и др., 2010). Лиственница, в частности, формирует широко разветвленную поверхностную корневую систему и характеризуется специфическим фитогенным полем. Также для почвы, сформированной в фитогенных полях отдельных деревьев, показана изменчивость физико-химических свойств (Карпачевский и др., 1989). Помимо косвенного трансформирующего воздействия дерева-эдификатора на окружающую среду, существуют значительные прямые связи корней растений с почвой и почвенными организмами. Физиологические и биохимические механизмы у растения, например корневые выделения, воздействуют на почвенную фауну и изменяют ее численность, видовой состав и эколого-трофическую структуру сообществ (Иванов, 1973; Bonkowski et al., 2009; Ларикова, Волобуева, 2021).
Таким образом, вследствие трансформации факторов окружающей среды и изменения различных характеристик травяного покрова происходят перестройки в составе и структуре фауны почвенных беспозвоночных. Интерес ученых при изучении нематод главным образом сосредоточен на характеристике сообществ в однородных древостоях, лиственных и хвойных лесах. Исследованиями, проведенными в различных областях мира, показано, что в структуре сообществ почвенных нематод лесных экосистем преобладает группа бактериотрофов, высокая степень доминирования отмечена для микотрофов и нематод, ассоциированных с растениями, тогда как группа паразитов растений и хищников представлена слабо (Boag, 1974; Magnusson, 1983; Ruess, 1995; Alphei, 1998; Yeates, 2007; Hanel, 2010; Hanel, Cerevkova, 2010; Renco et al., 2012; Kitagami, Matsuda, 2015; Sushchuk, Matveeva, 2021). Исследования сообществ почвенных нематод в различных зонах фитогенного поля дерева единичны. Так, в результате сравнения сообществ нематод в почве подкронового пространства пихты и на открытых участках показано изменение трофической структуры сообщества, в то время как для таксономического разнообразия и численности нематод различий отмечено не было (Shen et al., 2019). Влияние многих видов интродуцированных деревьев на сообщества почвенных нематод изучено в ботанических садах. В подкроновых пространствах деревьев обнаружено увеличение таксономического разнообразия паразитов растений, их численности и доли в сообществе по сравнению с естественными условиями произрастания (Калинкина и др., 2016, 2019; Сущук и др., 2016). Полевой эксперимент по опоясыванию елей, направленный на выявление воздействия корневых выделений дерева на почвенных нематод, показал снижение численности микотрофов, повышение бактериотрофов, при этом другие трофические группы, а также родовое богатство и численность не изменялись (Kudrin et al., 2021).
Согласно литературным данным, для других групп почвенных беспозвоночных также отмечаются микропространственные различия в численности, видовой и трофической структуре их сообществ, вызванные влиянием фитогенного поля дерева (Захаров и др., 1989; Таскаева, Долгин, 2010; Мазей и др., 2011; Гончаров и др., 2014, 2016).
Изучение сообществ почвенных нематод в зонах фитогенного поля различных видов деревьев позволит выявить природные закономерности формирования сообществ педобионтов. В связи с этим целью данной работы является изучение особенностей сообществ почвенных нематод в разных зонах фитогенного поля с акцентом на нематод, трофически связанных с растениями (паразиты растений и нематоды, ассоциированные с растениями), так как, предположительно, данные группы будут наиболее подвержены влиянию фитогенного поля.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования выполнены на территории Лахденпохского района, вблизи пос. Элисенваара, Республика Карелия (61°27.421′ с.ш., 29°39.228′ в.д.) в июле 2015 г. Территория располагается на границе южной и средней подзон тайги и характеризуется умеренным климатом, теплым и влажным летом, холодной и облачной зимой, что обусловлено географической близостью Ладожского озера (Назарова и др., 2004; Гнатюк и др., 2011). Исследования выполнены в лиственничнике кисличном, возраст которого насчитывает 80 лет. Лиственничник кисличный создан культурами лиственницы и характеризуется преобладанием в напочвенном покрове Oxalis acetosella L. (30%), Hepatica nobilis Schreb. (20%), Calamagrostis arundinacea (L.) Roth (10%) и Maianthemum bifolium (L.) F.W. Schmidt (10%). Почва в месте исследования подзолистая супесчаная на озерно-ледниковых отложениях и характеризуется следующими агрохимическими характеристиками с учетом генетических горизонтов A0, A1A2, A2B: С (%) на глубине 0–1 см – 7.8, 1–2 см – 0.7, 2–17 см – 0.6; N (%) на глубине 0–1 см – 0.5, 1–2 см – 0.3, 2–17 см – 0.1; pН на глубине 0–1 см – 5.8, 1–2 см – 5.5, 2–17 см – 4.9 (Рыжкова и др., 2016). Почвенные образцы были отобраны по трансекте от лиственницы сибирской Larix sibirica Ledeb. к ели обыкновенной Picea abies (L.) H. Karst. на расстоянии от соседних деревьев, чтобы исключить их влияние. Расстояние между деревьями (стволами) насчитывало около 10 м. Исследованы следующие зоны фитогенного поля деревьев: приствольное повышение (ПП_1) и подкроновое пространство (ПП_2), а также межкроновое пространство (МП, пространство между кронами деревьев). Зона ПП_1 для обоих деревьев составляла 20–30 см, зона ПП_2 для лиственницы – около 450 см, для ели – 300 см. МП насчитывало около 200–250 см. Напочвенный покров в зонах фитогенного поля деревьев представлен в табл. 1.
Таблица 1.
Состав травяно-кустарничкового яруса (проективное покрытие) в различных зонах фитогенного поля лиственницы и ели
| Лиственница | МП | Ель | |||
|---|---|---|---|---|---|
| ПП_1 | ПП_2 | ПП_1 | ПП_2 | ||
| Общее проективное покрытие, % | 15 | 40 | 70 | 50 | 40 |
| Травяно-кустарничковый ярус, % | 15 | 40 | 70 | 50 | 40 |
| Calamagrostis arundinacea (L.) Roth | 3 | 5 | 20 | 2 | 7 |
| Campanula rotundifolia L. | – | – | 1 | – | – |
| Carex globularis L. | – | – | – | – | 3 |
| Deschampsiaflexuosa P. Beauv. | – | 3 | – | – | – |
| Dryopteris cathusiana (Vill) | 2 | 10 | 10 | – | – |
| Equisetum sylvaticum L. | 0.5 | 0.5 | 1 | 0.5 | – |
| Fragaria vesca L. | – | – | 1 | 0.5 | – |
| Hepatica nobilis Mill | 0.5 | 5 | 15 | 15 | 15 |
| Luzula pilloza (L.) Willd. | – | 1 | 1 | – | 1 |
| Melampyrum sylvaticum L. | – | 1 | 0.5 | 0.5 | – |
| Mianthemum bifolium (L.) F.W. Schmidt | 2 | 10 | 5 | 5 | 15 |
| Orthilia secunda (L.) House | – | 0.5 | 1 | – | – |
| Oxalisacetosella L. | 5 | 10 | 30 | 10 | 5 |
| Rubus saxatilis L. | 0.5 | 2 | 3 | 15 | 1 |
| Trientaliseuropaea L. | 3 | 2 | 1 | – | 3 |
| Vaccinium myrtillus L. | – | – | 2 | – | – |
| Vaccinium vitis-idaea L. | – | – | 1 | – | – |
| Veronica chamaedrys L. | – | – | 0.5 | – | – |
| Мохово-лишайниковый ярус, % | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Pleurozium schreberi Willd. Ex Brid | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Dicranum scoparium Hedw. | – | – | – | – | 0.5 |
| Возобновление древесных растений, % | – | – | – | 15 | – |
| Sorbus aucuparia L. | – | – | – | 15 | – |
| Опад, % | 95 | 85 | 60 | 85 | 90 |
Примечания. Названия растений приводятся по: The Plant List, 2013 (http://www.theplantlist.org/). Здесь и в табл. 2, 3: ПП_1 – приствольное повышение, ПП_2 – подкроновое пространство, МП – межкроновое пространство.
Отбор почвенных образцов массой 30 г производили в девяти повторностях для каждой исследованной зоны фитогенного поля с использованием почвенного бура (d = 2 см) из верхнего слоя на глубину 10 см. Каждый почвенный образец был помещен в пластиковый пакет с замком (zip-lock) для предотвращения высыхания и в течение 48 ч доставлен в лабораторию паразитологии животных и растений ИБ КарНЦ РАН. В лабораторных условиях до экстракции почвенные пробы хранили при температуре 4°С не более одной недели. Для выделения подвижных форм нематод из почвы был использован вороночный метод Бермана. Экспозиция выделения нематод из почвы составляла 48 ч. В качестве фиксатора использовали ТАФ (триэтаноламин + формалин + вода, в соотношении 2 : 7 : 91) (Bezooijen, 2006). Идентификацию нематод (не менее 100 особей из каждой пробы) до рода (для половозрелых особей – до вида) проводили на временных глицериновых микроскопических препаратах с помощью светового микроскопа Olympus серии CX41 при увеличении в 400–800 раз. Таксономическая структура нематод приведена в соответствии с классификацией, предложенной Де Леем и Блэкстером (De Ley, Blaxter, 2004; Abebe et al., 2006). Оценены общая численность почвенных нематод и численность паразитов растений и нематод, ассоциированных с растениями в отдельности (экз./100 г почвы), таксономическое разнообразие, эколого-трофическая структура, индекс Шеннона на основе родов нематод (H') (Одум, 1975), индекс Чао второго уровня (Chao2) (Chao, 1987; Hammer et al., 2001).
Каждый таксон нематод относили к одной из эколого-трофических групп: бактериотрофы (Б), микотрофы (М), политрофы (П), хищники (Х), паразиты растений (Пр) и нематоды, ассоциированные с растениями (Аср) (Yeates et al., 1993). Кроме того, при анализе разнообразия фауны учтена трофическая группа энтомопатогенных нематод, но группа была исключена из дальнейшего анализа сообществ, так как данные виды не принимают участия в почвенных процессах. Особое внимание в исследовании уделено нематодам, трофически связанным с растениями (паразиты растений и нематоды, ассоциированные с растениями).
Статистическая обработка проведена с использованием программы PAST 4.0 (Hammer et al., 2001). Формат представления данных в таблицах M ± SD (среднее значение ± стандартное отклонение). Значимость различий оценивали с помощью непараметрического U-критерия Манна–Уитни (Mann–Whitney, U-test) для парных сравнений, вследствие несоответствия распределения признаков нормальному. Различия между группами считали достоверными при р < 0.05. Для выявления специфики сообществ почвенных нематод, сформировавшихся в различных зонах фитогенного поля дерева, использовали метод главных компонент и кластерный анализ. Тенденции изменения таксономической структуры сообществ изучали с помощью компонентного анализа. Исходная матрица состояла из n объектов (зоны фитогенного поля), охарактеризованных по m признакам (средние значения относительного обилия таксонов, уровень плотности которых достигал 9%). Кластерный анализ проведен для иерархической классификации данных с использованием индекса сходства Жаккара. Исходная матрица состояла из n объектов (зоны фитогенного поля), охарактеризованных по m признакам (таксономическое разнообразие). Бутстреп-поддержка составляла 1000 итераций. Для выполнения исследования почвенных нематод использовано научное оборудование Центра коллективного пользования Федерального исследовательского центра “Карельский научный центр Российской академии наук”.
РЕЗУЛЬТАТЫ
Таксономическое разнообразие
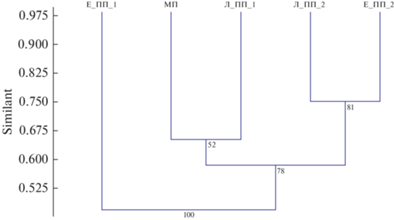
В результате проведенного исследования обнаружено 39 таксонов, принадлежащих к 25 семействам, 10 отрядам. Значительных различий в общем таксономическом разнообразии между зонами фитогенного поля отмечено не было (табл. 2). Количество родов нематод оставалось на уровне 24–26. Следующие 12 родов были обнаружены во всех точках: бактериотрофы (Alaimus, Plectus, Rhabditis, Acrobeloides), микотрофы (Tylencholaimus, Aphelenchoides, Ditylenchus), политрофы (Eudorylaimus), нематоды, ассоциированные с растениями (Malenchus, Coslenchus), паразиты растений (Paratylenchus), энтомопатогенные нематоды (Steinernema). Единично обнаружены таксоны Mesodorylaimus, Ceratoplectus, Cephalobus (приствольное повышение ели), Seinura (подкроновое пространство ели), Tylocephalus (межкроновое пространство), Pratylenchus, Sphaeronema, Monhystrella (подкроновое пространство лиственницы), Bunonema (приствольное повышение лиственницы). Группа паразитов растений во всех зонах фитогенного поля деревьев была представлена одним таксоном Paratylenchus straeleni de Conick, 1931, только в подкроновом пространстве лиственницы помимо вышеупомянутого вида были обнаружены еще два таксона – Pratylenchus sp., Sphaeronema sp. Нематоды, ассоциированные с растениями, напротив, характеризовались достаточно высоким разнообразием и насчитывали от трех до пяти таксонов. В зоне ПП_2 обоих деревьев обнаружено по пять таксонов данной эколого-трофической группы, количество таксонов снижается до трех в межкроновом пространстве. Расчет индекса Chao2, прогнозирующего потенциальное видовое богатство, показал, что видовой состав изучен на высоком уровне (более чем на 83%). Индекс Шеннона имел средние значения, минимальным показатель оказался в подкроновом пространстве ели, что при отсутствии значительных различий по числу таксонов между выборками говорит о наименьшей выравненности фауны нематод в данной точке (табл. 3). Применение кластерного анализа с использованием общего таксономического разнообразия нематод позволило оценить сходство между различными зонами фитогенного поля дерева. Один кластер образуют пробы из подкроновых пространств лиственницы и ели, второй объединил приствольное повышение лиственницы и межкроновое пространство. Достоверно обосабливается приствольное повышение ели (рис. 1).
Таблица 2.
Список таксонов нематод и их относительное обилие (%) в различных зонах фитогенного поля лиственницы и ели
| Таксон | Лиственница | МП | Ель | ||
|---|---|---|---|---|---|
| ПП_1 | ПП_2 | ПП_1 | ПП_2 | ||
| Отряд Enoplida Filipjev, 1929 | |||||
| Семейство Alaimidae Micoletzky, 1922 | |||||
| Alaimus primitivus de Man, 1880 | 0.1 | 0.1 | 0.3 | 3.5 | 0.4 |
| Отряд Triplonchida Cobb, 1920 | |||||
| Семейство Prismatolaimidae Micoletzky, 1922 | |||||
| Prismatolaimus intermedius Butschli, 1873 | – | 1.5 | 0.8 | – | 0.3 |
| Отряд Dorylaimida Pearse, 1942 | |||||
| Семейство Tylencholaimidae Filipjev, 1934 | |||||
| Tylencholaimus mirabilis Butschli, 1873 | 9.4 | 24.5 | 29.1 | 11.3 | 42.2 |
| Tylencholaimus stecki Steiner, 1914 | – | 1.0 | 6.3 | 21.8 | 0.4 |
| Семейство Qudsianematidae Jairajpuri, 1963 | |||||
| Eudorylaimus Andrassy, 1959 | 9.5 | 4.9 | 2.7 | 7.6 | 1.9 |
| Семейство Aporcelaimidae Heyns, 1965 | |||||
| Aporcelaimellus sp. Heyns, 1965 | – | 0.3 | 0.3 | 1.9 | 0.2 |
| Семейство Dorylaimidae de Man, 1876 | |||||
| Mesodorylaimusbastiani Andrássy, 1959 | – | – | – | 0.7 | – |
| Отряд Mononchida Jairajpuri, 1963 | |||||
| Семейство Mononchidae Jairajpuri, 1963 | |||||
| Clarkus papillatus Bastian, 1970 | 0.1 | 0.3 | – | 0.8 | 0.2 |
| Отряд Plectida Malakhov, 1982 | |||||
| Семейство Metateratocephalidae Eroshenko, 1973 | |||||
| Metateratocephalus Eroshenko, 1973 | 0.4 | – | 0.3 | – | 0.4 |
| Семейство Teratocephalidae Andrassy, 1958 | |||||
| Teratocephalus de Man, 1876 | 0.8 | 0.2 | 0.2 | – | 0.1 |
| Семейство Plectidae Orley, 1880 | |||||
| Plectus Bastian, 1865 | 22.5 | 8.4 | 16.4 | 19.5 | 34.2 |
| Plectus parietinus Bastian, 1865 | – | – | – | 1.7 | – |
| Ceratoplectus armatus Andrassy, 1984 | – | – | – | 0.7 | – |
| Wilsonemaotophorum Cobb, 1913 | 0.1 | – | – | 1.4 | 0.1 |
| Tylocephalus auriculatus Crossman, 1933 | – | – | 0.5 | – | – |
| Отряд Desmodorida De Coninck, 1965 | |||||
| Семейство Desmodoridae Filipjev, 1922 | |||||
| Prodesmodora Micoletzky, 1923 | 0.4 | – | 0.8 | 0.8 | – |
| Отряд Rhabditida Chitwood, 1933 | |||||
| Семейство Panagrolaimidae Filipjev, 1931 | |||||
| Panagrolaimusrigidus Fuchs, 1930 | 0.7 | 0.2 | 0.7 | – | 0.3 |
| Семейство Rhabditidae Orley, 1880 | |||||
| Rhabditis Dujardin, 1845 | 1.1 | 0.5 | 2.7 | 2.2 | 0.8 |
| Семейство Cephalobidae Filipjev, 1931 | |||||
| Cervidellus Thorne, 1937 | – | 1.5 | 0.9 | 2.0 | 0.8 |
| Acrobeloides butschli de Man, 1884 | 14.2 | 17.1 | 11.3 | 3.5 | 8.9 |
| Cephalobus persegnis Butschli, 1973 | – | – | – | 0.8 | – |
| Eucephalobus oxyuroides de Man, 1876 | 0.1 | – | 1 | 0.8 | – |
| Семейство Seinuridae Husain, Khan, 1967 | |||||
| Seinura Fuchs, 1931 | – | – | – | – | 0.2 |
| Семейство Aphelenchoididae Skarbilovich, 1947 | |||||
| Aphelenchoides Fischer, 1894 | 7.3 | 8.7 | 7.6 | 2.0 | 2.3 |
| Семейство Anguinidae Nicoll, 1935 | |||||
| Ditylenchus Filipjev, 1936 | 0.2 | 1.1 | 0.6 | 3.0 | 0.8 |
| Семейство Tylenchidae Orley, 1880 | |||||
| Filenchus Andrassy, 1954 | – | 0.3 | – | 2.1 | 0.4 |
| Malenchus Andrassy, 1968 | 24.2 | 3.8 | 3.4 | 3.2 | 2.1 |
| Aglenchus agricola Andrassy, 1954 | – | 0.1 | – | 0.4 | 0.2 |
| Coslenchus costatus de Man, 1921 | 2.6 | 1.1 | 2.7 | 7.7 | 0.1 |
| Lelenchus leptosome Andrassy, 1954 | 4.4 | 9.7 | 5.6 | – | 1.6 |
| Семейство Steinernematidae Filipjev, 1934 | |||||
| Steinernema Travassos, 1927 | 0.4 | 1.3 | 0.2 | 0.1 | 0.5 |
| Семейство Tylenchulidae Skarbilovich, 1947 | |||||
| Paratylenchus straeleni de Coninck, 1931 | 0.1 | 2.5 | 1.3 | 0.4 | 0.1 |
| Семейство Pratylenchidae Thorne, 1949 | |||||
| Pratylenchus Thorne, 1949 | – | 0.1 | – | – | – |
| Семейство Sphaeronematidae Raski, Sher, 1952 | |||||
| Spaeronema Raski, Sher, 1952 | – | 0.5 | – | – | – |
| Семейство Bunonematidae Micoletzky, 1922 | |||||
| Bunonema Jagerskold, 1905 | 0.3 | – | – | – | – |
| Отряд Triplonchida Cobb, 1919 | |||||
| Семейство Diphtherophoridae Micolatzky, 1922 | |||||
| Diphtherophora de Man, 1880 | 0.1 | 7.8 | – | – | 0.4 |
| Отряд Chromadorida Chitwood, 1933 | |||||
| Chromadoridae Filipjev, 1917 | – | – | – | 0.1 | – |
| Отряд Monhysterida Filipjev, 1929 | |||||
| Семейство Monhysteridae De Man, 1876 | |||||
| Monhystera stagnalis Bastian, 1865 | 0.8 | – | 4 | – | – |
| Monhystrella Cobb, 1918 | – | 2.5 | – | – | – |
| Nematoda (поврежденные особи) | 0.2 | – | 0.3 | – | 0.1 |
| Общее количество родов: 38 | 25 | 25 | 24 | 24 | 26 |
| Общее количество видов: 40 | 25 | 26 | 25 | 26 | 27 |
Таблица 3.
Параметры сообществ почвенных нематод в различных зонах фитогенного поля лиственницы и ели
| Показатель | Лиственница | МП n = 9 |
Ель | ||
|---|---|---|---|---|---|
| ПП_1 n = 9 |
ПП_2 n = 9 |
ПП_1 n = 9 |
ПП_2 n = 9 |
||
| Таксономическое разнообразие | |||||
| Общее количество родов | 25 | 25 | 24 | 24 | 26 |
| Паразиты растений | 1 | 3 | 1 | 1 | 1 |
| Нематоды, ассоциированные с растениями | 3 | 5 | 3 | 4 | 5 |
| H' | 2.14 | 2.43 | 2.38 | 2.60 | 1.63 |
| Chao2 | 29.90 | 27.77 | 25.14 | 24.6 | 28.45 |
| Численность, экз./100 г почвы | |||||
| Общая | 2318 ± 277c | 1513 ± 265bc | 1283 ± 121b | 1618 ± 353bc | 733 ± 50 a |
| Паразиты растений | 2 ± 2 a | 47 ± 19b | 17 ± 8ab | 7 ± 6a | 1 ± 1a |
| Нематоды, ассоциированные с растениями | 725 ± 48 c | 226 ± 66b | 151 ± 46b | 32 ± 72a | 216 ± 7b |
| Эколого-трофическая структура, % | |||||
| Бактериотрофы | 42.1 ± 19.1ab | 33.3 ± 6.5a | 40.4 ± 8.9ab | 37.1 ± 10.5ab | 46.9 ± 12.7b |
| Микотрофы | 17.0 ± 4.8a | 43.1 ± 9.2b | 43.6 ± 12.7b | 38.1 ± 15.7b | 46.1 ± 10.1b |
| Политрофы | 9.5 ± 8.5ab | 5.2 ± 4.8ab | 3.0 ± 3.0 a | 10.2 ± 6.5b | 2.1 ± 1.6a |
| Хищные нематоды | 0.1 ± 0.2a | 0.3 ± 0.4a | 0a | 0.8 ± 0.8a | 0.4 ± 0.4a |
| Нематоды, ассоциированные с растениями | 31.2 ± 18.3c | 15.0 ± 7.3b | 11.7 ± 8.1ab | 13.4 ± 9.7ab | 4.4 ± 2.9a |
| Паразиты растений | 0.1 ± 0.1a | 3.1 ± 2.2b | 1.3 ± 0.6ab | 0.4 ± 0.4a | 0.1 ± 0.1a |
Рис. 1.
Иерархическая кластеризация сообществ почвенных нематод из различных зон фитогенного поля лиственницы и ели на основе таксономического разнообразия, используя коэффициент Жаккара (Jaccard). Обозначения зон фитогенного поля: Л_ПП_1 – приствольное повышение лиственницы, Л_ПП_2 – подкроновое пространство лиственницы, МП – межкроновое пространство, Е_ПП_1 – приствольное повышение ели, Е_ПП_2 – подкроновое пространство ели. В узлах указана bootstrap-поддержка.
Плотность популяций нематод
Наибольшая численность нематод отмечена для приствольного повышения лиственницы, наименьший показатель отмечен для подкронового пространства ели (табл. 3). Род Tylencholaimus преобладал во всех исследованных точках отбора проб, в фитогенном поле лиственницы также многочисленными были бактериотрофы родов Acrobeloides, Plectus (приствольное повышение), а также нематоды, ассоциированные с растениями, родов Malenchus (приствольное повышение), Lelenchus (подкроновое пространство). Вышеупомянутый р. Plectus также характеризовался высокими значениями численности для фитогенного поля ели. Численность паразитов растений характеризовалась низкими значениями во всех исследованных точках, самые низкие значения показателя отмечены под елью по сравнению с МП и ПП_2 лиственницы. В зоне ПП_2 лиственницы численность паразитов была наибольшей, в 2.8 раза выше, чем в межкроновом пространстве. Группа нематод, ассоциированных с растениями, напротив, характеризовалась высокими значениями численности, особенно в фитогенном поле лиственницы.
Эколого-трофическая структура сообществ
В эколого-трофической структуре сообществ нематод в большинстве точек доминировали микотрофы и бактериотрофы, группа нематод, ассоциированных с растениями, также имела достаточно высокую долю, главным образом в зонах фитогенного поля лиственницы. Эти три эколого-трофические группы вместе составляли до 97% от видового состава фауны (табл. 3). Наблюдались некоторые отличия в позициях группы-доминанта: для лиственницы показана смена доминанта при продвижении от ствола к периферии (от бактериотрофов к микотрофам), для ели тенденция не выявлена, и вышеупомянутые группы имели одинаково высокий процент в обеих зонах. Среди микотрофов наиболее высоким относительным обилием характеризовались виды р. Tylencholaimus (от 9.4 до 42.6% состава фауны), среди бактериотрофов – р. Plectus. Наиболее низкие показатели относительного обилия отмечены для хищников и паразитов растений. Для приствольных повышений лиственницы и ели характерно повышение доли политрофов и нематод, ассоциированных с растениями, по сравнению с другими точками. Межкроновое пространство характеризовалось неполночленной эколого-трофической структурой (полностью отсутствовала группа хищников).
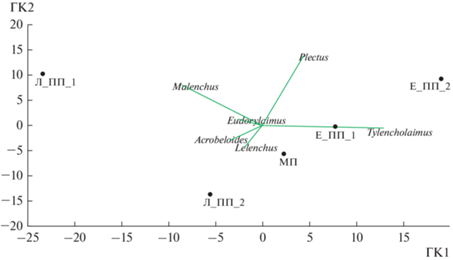
Ординация биотопов методом главных компонент на основе видов с относительным обилием более 9% наглядно демонстрирует смену доминирующих таксонов в зонах фитогенного поля лиственницы и ели (рис. 2). Так, таксоны нематод, ассоциированных с растениями, – Malenchus и Lelenchus, а также бактериотроф р. Acrobeloides тяготеют к зонам фитогенного поля лиственницы, р. Tylencholaimus – к ели.
Рис. 2.
Ординация доминирующих таксонов нематод (виды с относительным обилием более 9%), обитающих в зонах фитогенного поля лиственницы и ели. Первая главная компонента (ГК1) объясняет 67.15% различий, вторая главная компонента (ГК2) – 26.18%. Обозначения зон фитогенного поля те же, что на рис. 1.
ОБСУЖДЕНИЕ
В целом, обнаруженное в результате исследования таксономическое разнообразие нематод является типичным для естественных хвойных биоценозов Республики Карелия. Однако, согласно варьирующим значениям индекса Шеннона (1.63–2.60) в сочетании со сходным количеством родов, фауна оценивается как имеющая низкую выравненность, т.е. с неравномерным распределением доминирования по численности между таксонами, что нехарактерно для бореальных лесов (Груздева и др., 2011; Matveeva, Sushchuk, 2016). Среди паразитов растений отмечены три таксона, среди которых вид P. straeleni, обнаруженный во всех точках, является широко распространенным по всему миру, наиболее часто обнаруживается в лесных почвах и паразитирует на корнях деревьев и кустарников (Brzeski, 1998; Akyazi et al., 2015). Седентарный паразит р. Sphaeronema, найденный в подкроновом пространстве лиственницы, единично обнаруживается на территории республики. Согласно результатам кластерного анализа, основанного на таксономическом разнообразии нематод, высокий достоверный уровень обособленности отмечен для приствольного повышения ели, здесь обнаружено пять таксонов, которые не были найдены в других зонах, в каждой из которых найдено по одному–два специфичных таксона. Для этой зоны также отмечается высокий индекс Шеннона, что с учетом низкого числа таксонов указывает на высокий уровень равномерности распределения видов по обилию, а наиболее низкое значение индекса Chao2 среди исследованных зон свидетельствует о наименьшей степени выявленности таксономического разнообразия.
Численность нематод в приствольном повышении лиственницы характеризовалась наиболее высокими значениями по сравнению с другими точками, главным образом за счет группы нематод, ассоциированных с растениями, и бактериотрофов. Лиственница, по сравнению с елью, обладает более высокой сквозистостью кроны, существенно увеличивает плодородие почвы за счет высокого содержания химических веществ в хвое, в частности азота, характеризуется высокой интенсивностью биологического круговорота биогенных элементов в подстилке, сформированной в ее подкроновом пространстве (Рыжкова и др., 2016). Кроме того, для лиственничной подстилки, по сравнению с другими породами, характерно высокое содержание почвенных микроорганизмов и беспозвоночных (Безкоровайная, Яшихин, 2003). В то же время низкие значения численности нематод под елью в зоне приствольного повышения, по-видимому, обуславливаются крайне низкой пропускной способностью кроны ели, крупные ели задерживают до 70% осадков (Ипатов и др., 2009). Для горизонтального размещения коллембол под елью показаны сходные закономерности – наибольшая численность беспозвоночных отмечена для прикомлевого участка у основания деревьев (Таскаева, Долгин, 2010). Подобная ситуация отмечена для ельника-кисличника в Подмосковье, где для прикомлевых участков показана наибольшая плотность и разнообразие микроартропод (Захаров и др., 1989). Полученные результаты авторы связывают со стабильным гидротермическим режимом в данной зоне. Кроме того, приствольные воды характеризуются более высоким содержанием органического углерода, ионов ${\text{NO}}_{3}^{ - }$, ${\text{NH}}_{4}^{ + }$, а также более кислой реакцией почвенного раствора по сравнению с водами подкронового пространства (Арчегова и др., 2015).
Полученные данные, характеризующие эколого-трофическую структуру сообщества нематод в исследованных точках, согласуются с литературными данными (Ruess, 1995; Alphei, 1998; Yeates, 2007; Hanel, 2010; Hanel, Cerevkova, 2010; Renco et al., 2012; Kitagami, Matsuda, 2015; Matveeva, Sushchuk, 2016; Shen et al., 2019). Все зоны фитогенного поля деревьев характеризовались сходным рядом доминирования, однако в некоторых случаях наблюдалась перестройка трофической структуры (смена группы доминанта). Среди нематод, трофически связанных с растениями, преимущество получают виды, ассоциированные с растениями, вследствие экологической пластичности и широкого спектра питания они могут питаться как на эпидермальных клетках корневых волосков, так и содержимым гифов грибов. Высокое относительное обилие данной трофической группы в сообществе в целом характерно для северных лесов (Hanel, 2004, 2010; Matveeva, Sushchuk, 2016). В литературных данных также отмечается некоторое возрастание доли ассоциированных с растениями нематод в почве под лиственницей (Boag, 1974). Такие микропространственные изменения в эколого-трофической структуре отмечаются для многих других почвенных беспозвоночных. Показано, что трофическая структура для многих групп беспозвоночных почвы значительно варьировала в межкроновом пространстве, в подкроновом эти различия были менее выражены (Гончаров и др., 2014). Так, при переходе от прикомлевых пространств через подкроновые к межкроновым изменяется состав субдоминирующих видов почвообитающих раковинных амеб (Мазей и др., 2011). Фитофаги среди почвенных беспозвоночных увеличивали плотность по направлению от ствола к краю кроны (Гончаров и др., 2016). Невысокая доля паразитов растений в сообществах нематод под лиственницей, а также под елью, не согласуется с данными, полученными нами ранее для деревьев, интродуцированных в ботанических садах Северо-Запада России (Калинкина и др., 2016, 2019; Сущук и др., 2016). Это, вероятно, связано с тем, что высаженные в Лахденпохском районе деревья росли и развивались спонтанно, не испытывая антропогенных воздействий (включая отсутствие планомерного ухода за посадками), и сформировали лесной массив с соответствующей напочвенной растительностью, не предпочтительной для нематод, трофически связанных с растениями.
ЗАКЛЮЧЕНИЕ
Таким образом, в результате проведенного исследования получены первичные данные об особенностях сообществ почвенных нематод в зонах фитогенного поля двух видов деревьев – лиственницы сибирской и ели обыкновенной. В целом полученные данные, описывающие сообщества нематод в почве под деревьями, соотносятся с таковыми, характеризующими естественные хвойные леса по всему миру. Однако наблюдались некоторые отличия между фитогенными полями ели и лиственницы. Показано повышение общей численности, а также абсолютной и относительной плотности нематод, трофически связанных с растениями (паразиты растений, нематоды, ассоциированные с растениями), под лиственницей по сравнению с елью, в то же время под елью возрастает относительное обилие микотрофов и бактериотрофов в подкроновом пространстве, менее требовательных к условиям среды групп. Вероятно, большая сквозистость кроны лиственницы, а также богатый органическими и минеральными веществами опад создают более благоприятную среду для травянистых растений и, соответственно, для нематод, трофически связанных с растениями, чем в фитогенном поле ели.
При сравнении исследованных показателей внутри фитогенных полей отмечено повышение общей численности нематод и относительного обилия политрофов в приствольных зонах обоих деревьев по сравнению с подкроновыми пространствами и межкроновой зоной. Хорошая воздухо- и водопроницаемость в данной зоне, а также сток приствольных вод, обладающих высоким содержанием питательных веществ, вероятно, привели к формированию таких закономерностей.
Таким образом, полученные результаты косвенно позволяют говорить о том, что отличительные черты, характеризующие сообщества почвенных нематод, связаны с микроклиматическими условиями в зонах фитогенных полей, обусловленных влиянием деревьев. Можно предположить, что пестрота экологических условий, формирующихся в зонах фитогенного поля дерева, отражается на реакции почвенных нематод под воздействием изменяющихся абиотических и биотических факторов среды.
Список литературы
Арчегова И.Б., Кузнецова Е.Г., Лиханова И.А., Панюков А.Н., Хабибуллина Ф.М., Виноградова Ю.А., 2015. Формирование лесных экосистем на посттехногенных территориях в таежной зоне / Под ред. Арчеговой И.Б. Сыктывкар: Институт биологии Коми НЦ УрО РАН. 140 с.
Ашек Е.В., Тиходеева М.Ю., 2006. Исследование фитогенного поля Tilia cordata Mill. в посадках в заповеднике “Белогорье” Белгородской области // Вестн. СПбГУ. Сер. 3. Биол. Вып. 3. С. 64–73.
Безкоровайная И.Н., Яшихин Г.И., 2003. Влияние гидротермических условий почвы на комплексы беспозвоночных в хвойных и лиственных культурах // Экология. № 1. С. 56–62.
Безкоровайная И.Н., Егунова М.Н., Таскаева А.А., 2017. Почвенные беспозвоночные и их трофическая активность в 40-летних лесных культурах // Сиб. экол. журн. № 5. С. 609–620. https://doi.org/10.15372/SEJ20170507
Гнатюк Е.П., Крышень А.М., Кузнецов О.Л., 2011. Биогеографическая характеристики приграничной Карелии // Тр. КарНЦ РАН. № 2. С. 12–22.
Гончаров А.А., Храмова Е.Ю., Тиунов А.В., 2014. Микростациальные различия трофической структуры сообществ почвенных беспозвоночных в лесах Печоро-Илычского заповедника // Биология почв. № 5. С. 571–579. https://doi.org/10.7868/S0032180X14050074
Гончаров А.А., Короткевич А.Ю., Кузнецова Н.А., 2016. Влияние фитогенного поля дуба грузинского на структуру населения почвенных беспозвоночных // Мат-лы Всеросс. (с междунар. участием) науч. шк.-конф., посвященной 115-летию со дня рождения А.А. Уранова (10–14 мая 2016 г.). Пенза: Изд-во ПГУ. С. 324–326.
Груздева Л.И., Матвеева Е.М., Сущук А.А., 2011. Разнообразие фауны нематод естественных биоценозов Карелии // Нематоды естественных и трансформированных экосистем. Сб. статей по мат-лам IX симп. Росс. об-ва с междунар. участием. Петрозаводск: КарНЦ РАН. С. 54–56.
Демьянов В.А., 1990. Ценотическая роль ели сибирской в сообщества лесного пояса на северо-западе плато Путорана // Тр. III молодежной конф. ботаников Ленинграда. С. 160–172.
Журавлева Е.Н., Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю., 2012. Изменение растительности на лугах под влиянием сосны обыкновенной (Pinus sylvestris L.) // Вестн. СПбГУ. Сер. 3. Бот. Вып. 2. С. 3–12.
Захаров А.А., Бызова Ю.Б., Уваров А.В., 1989. Почвенные беспозвоночные рекреационных ельников Подмосковья. М.: Наука. 224 с.
Иванов В.П., 1973. Растительные выделения и их значение в жизни фитоценозов. М.: Наука. 293 с.
Ипатов В.С., 2007. Фитогенное поле одиночных деревьев некоторых пород в одном экотопе // Бот. журн. Т. 92. № 8. С. 1186–1191.
Ипатов В.С., Журавлева Е.Н., Лебедева В.Х., Тиходеева М.Ю., 2009. Фитогенное поле Picea abies, P. obovata // Бот. журн. Т. 94. № 4. С. 558–568.
Калинкина Д.С., Сущук А.А., Матвеева E.М., 2016. Особенности сообществ почвенных нематод в условиях интродукции древесных растений // Экология. № 5. С. 360–367. https://doi.org/10.1134/S1067413616050052
Калинкина Д.С., Сущук А.А., Матвеева Е.М., Зенкова И.В., 2019. Сообщества почвенных нематод подкронового пространства деревьев, интродуцированных на территории Полярно-альпийского ботанического сада // Сиб. экол. журн. № 1. С. 71–85. https://doi.org/10.15372/SEJ20190106
Карпачевский Л.О., Холопова Л.Б., Просвирина В.П., 1989. О динамике строения почвенного покрова в лесных биоценозах // Почвоведение. № 5. С. 94–103.
Ларикова Ю.С., Волобуева О.Г., 2021. Современные представления об эколого-физиологической роли корневых экссудатов растений // Зернобобовые и крупяные культуры. № 4. С. 93–101. https://doi.org/10.24412/2309-348X-2021-4-93-101
Лукина Н.В., Орлова М.А., Исаева Л.Г., 2010. Плодородие лесных почв как основа взаимосвязи почва-растительность // Лесоведение. № 5. С. 45–56.
Мазей Ю.А., Блинохватова Ю.В., Ембулаева Е.А., 2011. Особенности микропространственного распределения почвообитающих раковинных амеб в лесах Среднего Поволжья // Аридные экосистемы. Т. 17. № 1 (46). С. 37–46.
Назарова Л.Е., Сало Ю.А., Филатов Н.Н., 2004. Особенности изменений климата в водосборе Ладожского озера // Климат Карелии: Изменчивость и влияние на водные объекты / Отв. ред. Филатов Н.Н. Петрозаводск: КарНЦ РАН. С. 160–170.
Одум Ю., 1975. Основы экологии. М.: Мир. С. 186–187.
Прокушкин С.Г., Зырянова О.А., 2013. О влиянии деревьев лиственницы Гмелина на послепожарное восстановление лесного фитоценоза в криолитозоне Средней Сибири // Сиб. экол. журн. № 5. С. 645–652.
Рыжкова Н.И., Крышень А.М., Геникова Н.В., Преснухин Ю.В., Ткаченко Ю.Н., 2016. Сравнительный анализ структуры напочвенного покрова в культурах лиственницы и зональных ельниках на границе средней и южной подзон тайги // Тр. КарНЦ РАН. № 12. С. 25–З8. https://doi.org/10.17076/eco517
Сущук А.А., Калинкина Д.С., Платонова Е.А., 2016. Сообщества почвенных нематод в условиях интродукции древесных растений на территории Ботанического сада Петрозаводского государственного университета // Hortus Botanicus. Т. 11. С. 157–170.
Таскаева А.А., Долгин М.М., 2010. Пространственное распределение коллембол и их динамика в среднетаежном ельнике черничном (Республика Коми) // Изв. Самар. науч. центра РАН. Т. 12. № 1. С. 103–107.
Уранов А.А., 1965. Фитогенное поле // Проблемы соврем. ботаники. Т. 1. С. 251–254.
Черняева Е.В., Викторов В.П., 2016. История и современное состояние изучения фитогенных полей // Соц.-экол. технологии. № 1. С. 89–105.
Abebe E., Andrassy I., Traunspurger W., 2006. Freshwater Nematodes: Ecology and Taxonomy. Wallingford: CABI Publishing. 752 p.
Akyazi F., Felek A.F., Cermak V., Cudejkova M., Foit J., et al., 2015. Description of Paratylenchus (Gracilacus) straeleni (De Coninck, 1931) Oostenbrink, 1960 (Nematoda: Criconematoidea, Tylenchulidae) from hazelnut in Turkey and its comparison with other world populations // Helminthologia. V. 52. № 3. P. 270–279.
Alphei J., 1998. Differences in soil nematode community structure of beech forests: Comparison between a mull and a modern soil // Appl. Soil Ecol. V. 9. P. 9–15.
Bezooijen J., 2006. Methods and Techniques for Nematology. Wageningen: Wageningen Univ. Press. 112 p.
Boag B., 1974. Nematodes associated with forest and woodland trees in Scotland // Ann. Appl. Biol. V. 77. P. 41–50.
Bonkowski M., Villenave C., Griffiths B., 2009. Rhizosphere fauna: The functional and structural diversity of intimate interactions of soil fauna with plant roots // Plant Soil. V. 321. P. 213–233. https://doi.org/10.1007/s11104-009-0013-2
Brzeski M.W., 1998. Nematodes of Tylenchina in Poland and Temperate Europe. Warszawa: Museum and Institute of Zoology Polish Academy of Sciences. 397 p.
Chao A., 1987. Estimating the population size for capture-recapture data with unequal catchability // Biometrics. V. 43. P. 783–791.
De Ley P., Blaxter M., 2004. A new system for Nematoda: Combining morphological characters with molecular trees, and translating clades into ranks and taxa // Nematology Monographs and Perspectives. V. 2. Leiden: E.J. Brill. P. 633–653.
Hammer Ø., Harper D.A.T., Ryan P.D., 2001. Past: Paleontological statistics software package for education and data analysis // Palaeontol. Electron. V. 4. № 1. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hanel L., 2004. Response of soil nematodes inhabiting spruce forests in the Šumava Mountains to disturbance by bark beetles and clear-cutting // For. Ecol. Manag. V. 202. P. 209–225.
Hanel L., 2010. An outline of soil nematode succession on abandoned fields in South Bohemia // Appl. Soil Ecol. V. 46. P. 355–371.
Hanel L., Cerevkova A., 2010. Species and genera of soil nematodes in forest ecosystems of the Vihorlat Protected Landscape Area, Slovakia // Helminthologia. V. 47. № 2. P. 123–135.
Kitagami Y., Matsuda Y., 2015. Community and trophic structure of soil nematode in Japanese coastal pine forest // Russ. J. Nematol. V. 23. № 2. P. 160.
Kudrin A.A., Zuev A.G., Taskaeva A.A., Konakova T.N., Kolesnikova A.A. et al., 2021. Spruce girdling decreases abundance of fungivorous soil nematodes in a boreal forest // Soil Biol. Biochem. V. 155. https://doi.org/10.1016/j.soilbio.2021.108184
Magnusson C., 1983. Abundance and trophic structure of pine forest nematodes in relation to soil layers and ground cover // Holarctic Ecol. V. 6. № 2. P. 175–182.
Matveeva E.M., Sushchuk A.A., 2016. Features of soil nematode communities in various types of natural biocenoses: Effectiveness of assessment parameters // Biol. Bull. V. 43. № 5. P. 474–482.
Renco M., Cermak V., Cerevkova A., 2012. Composition of soil nematode communities in native birch forests in Central Europe // Nematology. V. 14. № 1. P. 15–25.
Ruess L., 1995. Nematode fauna in spruce forest soils: a qualitative/quantitative comparison // Nematologica. V. 41. P. 106–124.
Shen Y., Yang W., Zhang J., Xu Z., Zhang L. et al., 2019. Forest gap size alters the functional diversity of soil nematode communities in alpine forest ecosystems // Forests. V. 10. https://doi.org/10.3390/f10090806
Sushchuk A.A., Matveeva E.M., 2021. Soil nematodes of coniferous forests in the Finnish-Russian Friendship Nature Reserve // Nat. Conserv. Res. V. 6. P. 76–88. https://doi.org/10.24189/ncr.2021.015
Yeates G.W., 2007. Abundance, diversity, and resilience of nematode assemblages in forest soils // Can. J. For. Res. V. 37. P. 216–225.
Yeates G.W., Bongers T., Goede R.G.M., de, Freckman D.W., Georgieva S.S., 1993. Feeding habits in soil nematode families and genera: An outline for soil ecologists // J. Nematol. V. 25. № 3. P. 315–331.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии