

Журнал общей биологии, 2019, T. 80, № 4, стр. 307-320
Микробиота – третий компонент системы хищник–жертва
А. С. Северцов 1, А. В. Шубкина 2, *
1 Московский государственный университет им. М.В. Ломоносова,
биологический факультет, кафедра биологической эволюции
119991 Москва, Ленинские горы, 1, Россия
2 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: annashubkina@rambler.ru
Поступила в редакцию 06.09.2018
После доработки 07.04.2019
Принята к публикации 29.04.2019
Аннотация
Пищевые пирамиды – это основа экосистем, а хищничество – одна из сил, формирующих их структуру. Принято считать, что система “хищник–жертва” включает два основных элемента, т.е. является бинарной. Исследование элиминации свободноживущих травоядных позволяет утверждать, что во взаимодействии консументов 1–2 порядков принимают участие прокариоты. В многократных естественных экспериментах регистрировали поведение поиска и преследования модельного хищника (борзые собаки трех пород). Сравнивали добычу (Lepus europeus, Saiga tatarica) борзых и ружейных охотников-резидентов. Показано отсутствие различий добытых животных по массе тела и внешним признакам. Присутствуют отличия благополучия: жертвы борзых в значимо большем числе случаев обладают нежелательными признаками (патологии состояния внутренних органов, часто полиорганные; старые подкожные шрамы; сниженная величина жировой капсулы; повышенное количество кожной микрофлоры). Возрастной состав добычи собак и охотников не различается у сайгака, но среди зайцев содержит значимо меньшую долю очень молодых особей из числа добытых собаками. С помощью посекундной GPS-регистрации показано, что успешность охоты не зависит от максимальной скорости преследования. Результаты преследования значимо различаются у представителей разных генетических групп борзых. Существует низкая успешность и высочайшая селективность отлова относительно отстрела. Изучение динамики количества непатогенной микрофлоры у животных в разном физиологическом состоянии показало, что ее повышение характерно для неблагополучных особей. У зайцев, пойманных борзыми, значимо увеличено количество микрофлоры поверхности тела, что усиливает и изменяет запах этой части особей. Изменение запаха служит сигналом для продолжения преследования. Микрофлора является третьим компонентом во взаимодействии хищника и жертвы, т.е. система “хищник–жертва” – триада.
Проблема взаимодействия хищников и их жертв имеет как фундаментальное, так и практическое значение. Фундаментальный аспект включает исследование трофических связей как процессов взаимодействия, практический же актуален для установления оптимального соотношения численностей хищников и их жертв, что необходимо для поддержания биоразнообразия, регуляции популяций, охраны природы и управления промыслом.
Хищничество – основная сила, формирующая структуру экосистем (Nilsson, 2001). Исключение воздействия хищника на популяции жертв ведет к накоплению изменений паттернов сложения, поведения и использования территории животными видов-жертв (Саблина, 1959; Борискин, Чирков, 1978; Близнюк, 1983; Филонов, 1989; Owen-Smith, Mills, 2008а, b; Preisser et al., 2007; Preisser, 2009).
Постановка проблемы. Хищничество – избирательное изъятие определенных фенотипов в результате взаимодействия – влияет на генетическую структуру популяции жертвы и внутривидовую конкуренцию. Но методы полевой зоологии ограничивают системное изучение паттернов элиминации, что остается основой многочисленных дискуссий (Павлов, 1982; Бибиков и др., 1985; Филонов, 1989; Latham et al., 2011; Hayward et al., 2011). Это обусловлено целым рядом причин. Прежде всего, невозможностью зарегистрировать и количественно описать все охоты хищника как несостоявшиеся (прекращенные в ходе тропления или ограниченные прогоном), так и состоявшиеся (с результатом или без него), т.е. исключает реальную оценку успешности охоты. Впервые эта проблема была сформулирована Макналти с соавт. (MacNulty et al., 2007) и остается актуальной, несмотря на успехи применения GPS-регистрации (Khater et al., 2015). Во-вторых, крайне ограничены методы характеристики добычи хищников, невозможно применение размерных, морфологических, патолого-анатомических и иных критериев для описания состояния жертвы в момент гибели. В лучшем случае при использовании останков добычи хищников применяется оценка bone marrow analysis (метод bma, впервые упомянут Cheatum, 1949, цит. по: Franzmann, Arneson, 1976; Mech, Delgiudice, 1985; O’Gara, Harris, 1988; Okarma, 1991; Mech, 2007) либо проводится сравнение гормональных показателей в популяции жертв до и после периода воздействия хищника (Barber-Meyer et al., 2007а, b, 2008; Barber-Meyer, Mech, 2008). Трудность детального описания процесса охоты и сравнения добычи хищников с иными животными существенно ограничивает возможность понять причины и значение изъятия фенотипов, т.е. установить механизмы элиминации жертв и воздействия хищника на популяции.
Для решения проблемы применили систему естественных экспериментов: изъятие свободноживущих травоядных (Saiga tatarica, Lepus europeus) модельным хищником – борзыми собаками. Добычу борзых сравнивали с результатами отстрела сотрудниками государственного охотничьего надзора и/или резидентными охотниками. Естественные эксперименты обеспечили регистрацию событий при многократном воспроизведении охотничьего поведения хищника (поиска, преследования и изъятия жертвы) и проведение исследований добытых животных для сравнительной характеристики их морфофизиологических особенностей (Соколов и др., 1991; Северцов, Шубкина, 2015а, б; Шубкина, 2017; Severtsov et al., 2017).
Уже в первых экспериментах (начало 1980-х годов, Калмыцкая АССР, сайгак в качестве жертвы) был доказан факт избирательного изъятия (Соколов и др., 1991), т.е. значимо разных долей благополучных (без отклонений в состоянии внутренних органов) и неблагополучных (с отклонениями) животных в выборках “отлов” и “отстрел”. При этом наблюдатели оказались не способны прогнозировать изъятие по визуально определяемым признакам жертвы (размерные и половозрастные отличия, размещение в движущейся группировке, относительная скорость и т.д.). Более того, отсутствовал единообразный состав отклонений состояния, позволявший надеяться на сходные признаки жертв, поддающиеся простому маркированию (например, изменение движений при суставных травмах или изменение запаха при заболевании почек и т.п.). Единственным общим признаком добычи было неблагополучие, возникшее существенно раньше преследования, т.е. наличие долговременного генерализованного адаптационного синдрома. Однако его прямая оценка по состоянию надпочечников либо уровню кортикостероидов не могла применяться из-за несомненного наличия стресса преследования и необходимости валидации измерений на большом поголовье, т.е. дополнительного изъятия нескольких сотен особей. Более того, общепризнанно, что индивидуальная реакция на повышение уровня кортикостероидов у разных особей неодинакова: стресс-реакция может приводить и к усилению, и к ослаблению защитных свойств и целостных реакций организма.
Поэтому были проведены лабораторные эксперименты для установления возможности существования единой системы маркеров, свидетельствующих о наличии долговременной адаптационной реакции. Было доказано существование упорядоченных изменений численности микроорганизмов элементов поверхности кожных покровов при индуцированных и естественных изменениях морфофизиологического состояния (Shubkina, Ushakova, 1994, 1996; Ерофеева, 2014, 2016).
Эти данные и соображения предопределили логику исследований и позволили сформулировать цель и задачи данной статьи.
Цель – системное изучение паттернов элиминации жертвы.
Задачи: 1) Сравнение внешних, половых, морфологических, морфофизиологических, микробиологических, поведенческих признаков особей свободноживущих травоядных, являющихся добычей модельного хищника (“отлов”) и отстрелянных резидентными охотниками (“отстрел”); 2) Выделение общих признаков животных группы “отлов”, необходимых и достаточных для их изъятия модельным хищником.
Этические ограничения при сборе материалов были установлены с первых дней и не изменились до настоящего времени. Они включают сбор материала, проводимый в сроки охоты, в соответствии с государственными лицензиями, при отсутствии негативных для жертвы абиотических факторов, т.е. минимизация ущерба для популяций свободноживущих животных и использование сезонов работы, в которые исключена добыча молодняка, беременных или лактирующих самок, животных, ослабленных засухой, морозами и/или другими абиотическими факторами.
МЕТОДЫ
Период полевых работ – октябрь–январь.
Многократные эксперименты: описание поиска и преследований, регистрация визуально, в баллах и c помощью GPS-трекеров (Шубкина и др., 2008; Severtsov et al., 2017).
Сравнение животных в выборках “отлов” и “отстрел”: описание полового и возрастного состава, полное и частичное патологоанатомическое исследование, оценка количества микро-организмов (Соколов и др., 1991; Шубкина, 2006; Северцов, Шубкина, 2014а, б, 2015а, б; Шубкина, 2017).
Эксперименты на полигоне: оценка возможности изменения параметров движения борзых при наличии обонятельных стимулов микробного происхождения (Шубкина и др., 2012).
Статистическую обработку выполняли с помощью программ Microsoft Excel и Statistica 7.0. При сравнении выборок использовали критерий Стьюдента (St), критерий Вилкоксона (Wi), критерий Хи-квадрат (χ2). Значимыми считались различия на уровне p < 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Успешность охоты
Взаимодействие хищника и жертвы направлено на реализацию энергетических потребностей хищника. Успешность охоты является важнейшим результатом охотничьего поведения хищников. Она существенно ниже 50% у диких наземных хищных и совсем не высока у модельного вида (Северцов, Шубкина, 2014а, б). Для оценки успешности охоты борзых рассчитывали доли поимок относительно числа состоявшихся преследований (по зайцу на состязаниях борзых при наличии большого числа наблюдателей). Для оценки успешности охоты человека применяли опросные данные и прямое наблюдение за охотниками (Блохина, 2016), рассчитывали доли отстрелянных зайцев относительно поднявшихся в радиусе до 50 м.
Успешность охоты борзых существенно ниже, чем у человека с ружьем (табл. 1). Представляется, что при регистрации всех попыток охоты у диких хищных доля поимок у них находится в сопоставимом диапазоне, хотя, несомненно, бывает сочетание условий, при которых успешность охоты хищника возрастает, например, при воздействии абиотических или антропогенных (содержание в загонах) факторов. Однако такие факторы действуют не всегда и не везде. Следовательно, для реализации энергетических потребностей хищника необходимы множественные попытки охоты. Во многих случаях они завершаются при предварительном изучении потенциальной добычи, наблюдении за ней – при скрадывании и прогонах. Прекращение охоты хищника возможно после начала преследования, после приближения к жертве, после попыток атаки, что может быть обусловлено как развитием процесса охоты, так и иными факторами (Капланов, 1948; Mech, 1970; Бологов, 1980; Павлов, 1982; Крушинский и др., 1985; Корытин, Бибиков, 1985; Шубкина, 2017; Hilborn et al., 2018).
Сравнение добычи борзых и охотников
Половой состав выборок “отлов” и “отстрел” у сайгаков не различается (табл. 2). Это соответствует опубликованным данным на тот период для Северо-Западного Прикаспия, Калмыкии и Казахстана (Банников и др., 1961; Жирнов, 1982). В выборке “отлов” среди зайцев-русаков значимо превалируют самцы, в отличие от выборки “отстрел” (Ерофеева, 2019). Это отличается от данных европейских исследователей, полагающих, что соотношение полов у зайцев-русаков близко к единице или в нем превалируют самки (Jezirsky, 1965; Sokos et al., 2015).
Средняя масса тела добытых борзыми зайцев значимо превосходит таковую в группе “отстрел”. Анализ возрастной изменчивости, проведенный И. Фоминой (2016) и Е. Ерофеевой (2019), объясняет причины этих различий: в выборке “отстрел” выше доля очень молодых особей (менее полугода), что также подтверждается нижними пределами изменчивости веса (табл. 3). Вероятно, та же причина объясняет несколько больший средний вес зайцев-русаков юга РФ по сравнению с европейскими странами: 4.1 кг – юг РФ, 3.9 кг – европейские страны (Suchentrunk et al., 2000; Thulin et al., 2012; Misiorowska et al., 2014).
Таблица 3.
Масса тела добытых животных
| Вид | Выборка | Число особей |
Средняя масса (кг) | Минимальное/ максимальное значения | Доверительный интервал |
|---|---|---|---|---|---|
| Заяц-русак | Отлов | 170 | 4.13* | 2.2/5.7 | 4.13 ± 4.06 |
| Отстрел | 59 | 3.79* | 1.2/5.4 | 3.79 ± 3.54 | |
| Сайгак (по: Соколов и др., 1991) | Отлов | 48 | 23 | ||
| Отстрел | 40 | 22.5 | |||
Для оценки возрастной изменчивости зайца-русака использовали критерий массы тела. Это допущение основано на двух исследованиях. В. Архипчук и О. Белова (1987) показали, что в 3–4 мес. зайцы-русаки, содержащиеся на ферме, в среднем достигают массы тела около 3.5 кг и внешне почти не отличаются от взрослых. И. Фомина (2016) провела сравнение массы тела и сухой массы хрусталика глаза свободноживущих русаков. Она показала, что в возрасте около 4 мес. масса сухого хрусталика достигает 70–90 мг. Используя два критерия (масса тела и масса хрусталика), мы разделили зайцев на две группы: заведомо молодые и старше.
В выборке “отлов” доля заведомо молодых зайцев существенно ниже. Возможно, это связано с тем, что значительная доля особей выборки “отлов” собрана на испытаниях, где пуск по молодому зверю запрещен, тогда как в правилах охоты добыча молодых и взрослых зайцев не оговорена (как в РФ, так и в Европейских странах). Не менее вероятно, что стереотипы поведения молодых и взрослых зайцев различны, что позволяет несформированным особям избегать преследования борзыми, но не выстрела охотника.
Данные, представленные на рис. 1, показывают, что возрастная избирательность изъятия жертвы борзыми не отличается от “отстрела” по сайгаку и различна по зайцу. Доли молодых (оценка по массе тела и развитию рогов у самцов) и взрослых сайгаков не различаются в двух выборках. Различия добытых сайгаков по возрастным и половым признакам не установлены. Полученные данные позволяют утверждать, что зайцы выборок “отлов” и “отстрел” различаются по возрастным и половым параметрам: в выборке “отлов” выше доля самцов, но ниже доля очень молодых особей. Следует отметить, что визуально, при наблюдении движущихся зайцев, различить самок и самцов невозможно. Различить молодых (менее 2–2.5 кг) и взрослых (3.5–5.7 кг) зайцев человек может на небольшой дистанции (до 20–30 м).
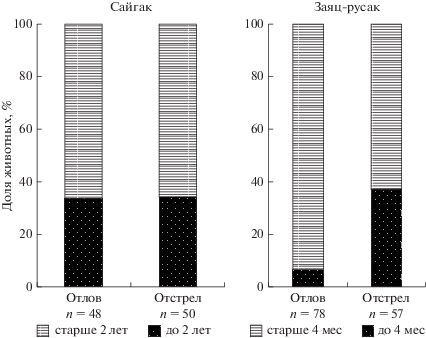
Таким образом, анализ размерных показателей и полового состава добытых травоядных позволяет утверждать, что эта группа свойств не является доминирующей при выборе жертвы борзыми, но избирательность зайцев-русаков по этим признакам возможна.
Особенности преследования
Разумеется, невозможно обсуждать особенности изъятия жертвы борзыми – быстрыми собаками, не учитывая особенностей их движения. Следовало установить, как параметры преследования влияют на успешность охоты. Для характеристики параметров преследования (скорость движения, длина, длительность преследования) применяли GPS-регистрацию высокой частоты (1 с) со специально разработанным оборудованием (Шубкина и др., 2008). Установлено, что длина, длительность и максимальная скорость (рис. 2) успешных и безуспешных преследований не различаются. Результаты подробно описаны ранее (Шубкина и др., 2012).
Рис. 2.
Диапазон максимальной скорости борзых при успешных (поимка) и безуспешных (без поимки) преследованиях (результат GPS-регистрации высокой частоты).
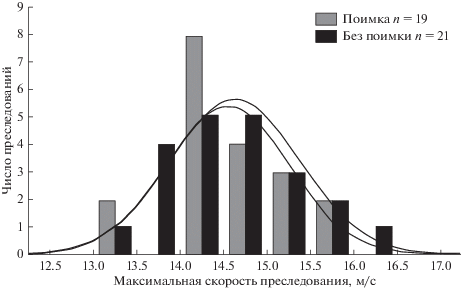
Самый важный из них: влияние повышения скорости на успешность преследования несущественно. Способность развить высокую скорость необходима для поимки борзыми зайца-русака, но это не единственный и даже не основной фактор, обусловливающий это событие. Скорость борзых варьирует в разные дни, при разных преследованиях и в течение каждого преследования (Северцов, Шубкина, 2015а, б). Существование изменений скорости свидетельствует о наличии механизмов ее корректировки. Предварительные данные дают основания полагать, что отбор исключительно на скорость бега ослабляет способность корректировать параметры движения. При этом, как и у гепардов (Wilson et al., 2013), более высокая скорость не повышает частоту поимок. Этот факт установлен для двух видов хищников и разных видов жертвы и, вероятно, является общим для наземных хищников-преследователей. Последнее подтверждают исследования гепардов: сила мотивации хищника (голод, наличие детенышей) и тип места охоты не оказывают прямого влияния на ее успешность (Hilborn et al., 2012).
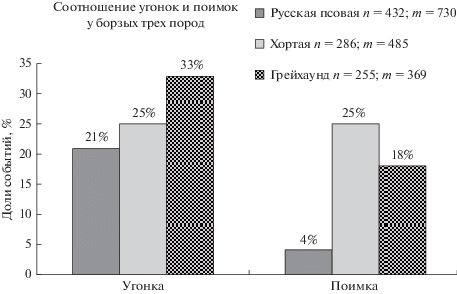
Свидетельством ограниченного влияния механических параметров преследования (скорости, длины, длительности) на успешность охоты являются результаты преследования зайцев борзыми генетически различных групп – представителями трех пород (968 борзых, 1584 преследования). Применяли определение “угонка”, означающее превосходство борзых в скорости по отношению к зайцу. По нашим данным (Северцов, Шубкина, 2015а, б), полученным на большом статистическом материале, не менее чем в 24% преследований борзые, начавшие преследование зайца с дистанции от 25 м, приближаются к нему, вынуждая менять ритм и направление движения (т.е. делают ему “угонку”), что подробно описано ранее. Для сравнения борзых трех пород использованы данные эксперта по испытаниям борзых Н.П. Семигорелова (личное сообщение, предоставлен материал) об успешности преследования на испытаниях (при ограничении поискового поведения собак и дистанции пуска). Представленные данные (рис. 3) показывают отсутствие прямой связи превосходства в скорости (группа “угонка”) и долей поимок от числа преследований у борзых трех генетических групп (пород) при наличии существенных различий успешности охоты (группа “поимка”).
Рис. 3.
Превосходство в скорости (угонки) и поимки у борзых трех пород. n – число собак; m – число преследований, в которых участвовали собаки данной породы (часть собак участвовала в нескольких преследованиях).
Визуальное наблюдение преследований зайцев и сайгаков борзыми и сравнение внешнего вида добытых животных не позволили выделить признаки, отличающие доступных жертв. Наблюдатели не способны прогнозировать успешность охоты борзых. Очень редко (5 из 43 жертв – 12% или 5 из 210 преследований – 2%) удавалось распознать признаки сайгака, влияющие на вероятность его поимки. Увеличение группы сайгаков сверх некоторого предела негативно влияет на успешность преследования, что косвенно свидетельствует о факте выбора. Человек не может распознать признаки зайца, определяющие его поимку до того, как она произойдет.
В начале экспериментальной работы мы предполагали, что выбор борзыми жертвы будет основан преимущественно на особенностях ее маршрута, длительности преследования и соотношениях скоростей движения хищника и жертвы. Оказалось, что это не так: жертвами борзых часто оказывались сайгаки из головки стада, т.е. животные, способные к длительному движению с высокой скоростью. GPS-регистрация показала, что максимальная и средняя скорость собаки, а также протяженность и длительность удачных преследований зайца несколько ниже, чем неудачных.
Показатели благополучия
Для оценки благополучия добытых животных проводили патологоанатомические вскрытия и описания состояния внутренних органов (Соколов и др., 1991; Северцов, Шубкина, 2014а, б, 2015а, б) (рис. 4).
Рис. 4.
Результаты патологоанатомического исследования сайгаков и зайцев, добытых борзыми (отлов) и отстрелянных резидентными охотниками (отстрел). а – частота регистрации отклонений состояния по группам органов у сайгаков; б – частота регистрации подкожных рубцов у сайгаков; в – величина жировой капсулы почек у зайцев-русаков.
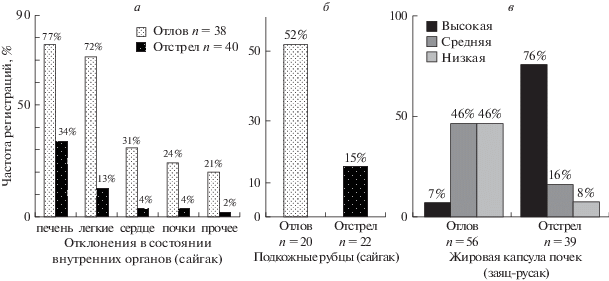
У всех сайгаков, взятых борзыми, присутствовали отклонения в состоянии внутренних органов, но среди отстрелянных доля таких животных составляла около трети. У сайгаков описаны многообразные патологии состояния, часто полиорганные нарушения и подкожные рубцы, следы нелетальных травм (рис. 4б), свидетельствующие о недостаточном благополучии – прямое доказательство сниженной приспособленности.
Сайгаки, пойманные борзыми, пусть и нездоровые (следовательно, в состоянии стресса), пребывали не в терминальном состоянии – это жизнеспособные особи, не отличающиеся по весу, размерам и внешнему виду от конспецификов. Их положение при движении стада – в головке или в центре группировок, а не в шлейфе отстающих – прямо говорит о способности адекватно реагировать и успешно двигаться.
Патанатомическое исследование зайцев (как взятых борзыми, так и отстрелянных) обычно затруднено обширными гематомами – результатами хватки собак или стрельбы дробью. Поэтому с зайцами использовали частичное исследование – сравнение величины жировой капсулы почек (рис. 4в) и встречаемости старых подкожных рубцов. Старые подкожные рубцы, не сопровождаемые проникающими ранениями мягких тканей, впервые описаны у зайцев-русаков Т.В. Блохиной (2016). Доля зайцев с такими следами травм по ее данным составляет около 27% в выборке “отлов”. В выборке “отстрел” такие животные не отмечены.
Наши данные подтверждают результаты исследований (Sinclair, Arcese, 1995), показавших отсутствие прямой связи между состоянием жертвы (как показателем благополучия) и ее подверженностью хищничеству. Эти исследователи установили, что у жертв хищников упитанность лучше, чем у животных, умерших своей смертью, т.е. в популяции жертвы обычно присутствуют особи более истощенные, чем те, кого добыл хищник. Следовательно, состояние жертвы влияет на ее подверженность хищничеству, но хищники обеспечивают исключение из популяций не самых слабых особей. Количество и частота патологий внутренних органов и/или нелетального прижизненного травматизма (подкожные рубцы) не являются признаком какой-либо половозрастной или размерной группы. Аналогичным образом высокий уровень гормонов стресса может служить как причиной, так и следствием неблагополучия (Werner, Morgan, 2017).
Сравнение благополучия изъятых травоядных двух видов позволяет утверждать, что модельный хищник способен к избирательному изъятию менее благополучных особей, в отличие от охотника. При этом такие животные не отличаются от остальных на взгляд наблюдателя или охотника и не отклоняются по размерным показателям. Среди зайцев борзые преимущественно элиминируют взрослых животных и самцов. Выбор жертвы борзыми не включает преимущественное изъятие более мелких и/или молодых особей.
Преследование – затратный процесс, а выраженное снижение благополучия жертвы в норме присутствует лишь у части животных. Однако у многих особей в популяции состояние хуже, чем у прочих. Именно среди них хищник выбирает доступную жертву, поимка которой обеспечивает адекватную компенсацию его энергетических затрат. При прогонах и преследовании хищники вынуждают жертву реализовывать свои физические и психические возможности, выбирая маршрут и скорость бегства и доказывая адекватность этого выбора. Неблагополучное, даже не совсем здоровое или находящееся в плохой физической или психической форме животное все равно может увернуться от хищника, избегнув хватки. Жертва, способная компенсировать свою слабость адекватным поведением (выбор маршрута, убежищ и т.п.), уходит, в отличие от менее приспособленной. Многокомпонентность реакции хищника на жертву детерминирует его способность производить прямой отбор жертв по приспособленности. Однако необходимость выбора наименее адаптированных особей оплачивается снижением успешности охоты.
Проблема определения доступности жертвы
Результаты многократных естественных экспериментов показали, что существуют низкая успешность и высочайшая селективность. Что может обеспечить селективность изъятия при многообразии причин снижения приспособленности и отсутствии внешних различий жертвы (размеров, окраса, скорости и пр.)? Дистантное различение признаков доступности жертвы – запах (хотя этот способ передачи информации имеет множество недостатков).
Когда мы видим хищника, преследующего хорошо видимого зверя, у нас складывается впечатление, что его охотничье поведение основано на зрительной реакции. Доказать использование обоняния при этом трудно, но возможно (рис. 5). Наличие чутья у борзых – давно известный факт. Собственные наблюдения за борзыми в полевых условиях (тренировка, охота, испытания, состязания) вынуждают задуматься о значительной роли обоняния в их поведении. По данным наблюдений и GPS-регистрации борзые активно применяют верхнее чутье: нюхают потоки воздуха и могут двигаться по градиенту. Наиболее наглядно использование обоняния подтверждают случаи, когда борзые обнаруживали зайца до того, как он поднялся с лежки или мог быть замечен (рис. 5).
Рис. 5.
Активация ориентировочной реакции (изменения ритма движения и повороты при ограничении поисковой активности) двух борзых на основе обонятельных стимулов (данные высокочастотной GPS-регистрации). Скобками отмечено проявление поисковой активности борзых за несколько десятков секунд до подъема зайца; стрелка указывает момент подъема зайца и его появления в поле зрения борзых.
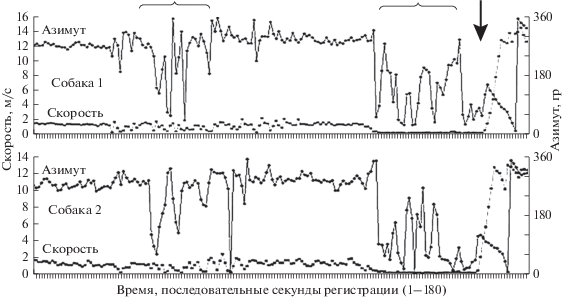
Как и наземные хищники, борзые активно применяют обоняние при реализации охотничьего поведения. Однако запаховые сигналы во время преследования, когда хищник ориентируется верхним чутьем, т.е. по воздушным потокам, а не по фиксированным на субстрате частицам (на грунте, растительности и т.п.), ненадежны. Вероятно, это одна из причин низкой эффективности охоты диких хищников (Северцов, Шубкина, 2015а, б).
Использование GPS-регистрации позволило доказать, что борзые активно используют обоняние, хотя преследование начинается после визуального распознавания. Однако следовало определить, что может служить обонятельным маркером, дифференцированно работающим у животных с разным уровнем благополучия.
Микробиота – маркер состояния макроорганизма
Еще в конце прошлого века теоретически и экспериментально были обоснованы положения об участии кожной микрофлоры в формировании запаха макроорганизмов и о влиянии стресс-реакции на ее численность и состав (Соколов и др., 1990а, б, в; Brown, Schellink, 1994; Shubkina, Ushakova, 1994; Braks et al., 1995; Brown, 1995; Schellink, Brown, 2000; Moshkin et al., 2002; Voigt et al., 2005; Мошкин и др., 2006; Goodacre et al., 2009; Марков, 2009, 2010; Rosenthal et al., 2011).
Е.В. Ерофеева (2016), используя модельный вид (собаки), показала, что количество колоний микроорганизмов, формирующихся на отпечатке (мясопептонный агар, МПА), изменяется в зависимости от состояния особей. При этом не выражена сезонная, географическая, половая изменчивость количества микрофлоры поверхности кожи (носового зеркальца). В течение онтогенеза количество колоний изменяется в зависимости от состояния особи. Существуют значимые различия клинически здоровых и неблагополучных животных (рис. 6).
Рис. 6.
Количество колоний микроорганизмов в пробах здоровых и неблагополучных животных (по: Ерофеева, 2016).
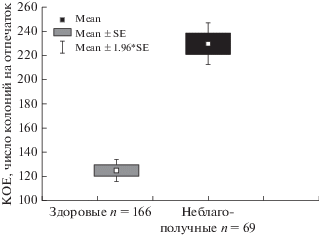
В работе Е.В. Ерофеевой показано наличие тесной взаимосвязи численности кожных бактерий хозяина и его состояния и доказан факт их изменений при заболевании и/или иной форме стресса. Результаты современных лабораторных исследований вскрывают конкретные механизмы модуляции численности кожных бактерий гормонами стресса и изменениями продукции защитных средств организма, опосредованными действием этих гормонов. Таким образом, стресс-зависимое изменение микробиоты жертвы служит основой изменения запаха потенциально доступной особи и обеспечивает переключение внимания на его источник.
У животных многих видов скрещивание с конспецификами, больными или зараженными паразитами, модулируется благодаря использованию обонятельных ключей. Различение запахов здоровых и больных, включая пораженных паразитами, – важнейший модулятор социального поведения, в том числе селективности размножения, избегания распространения инфекций и инвазий в популяциях и других аспектов, прямо влияющих на микроэволюционные процессы.
Животные реагируют на запах измененной микробиоты. Например, комары распознают больных малярией, и у них существует предпочтение культуральной микрофлоры больных людей. После антималярийной терапии количество москитов, кусающих больных, перестает отличаться от числа москитов, атакующих здоровых (Verhulst et al., 2009, 2011).
В соответствии с современными представлениями (Penn, Potts, 1998; Steinert et al., 2000; Fischbach, Bromley, 2001; Alexander, Hudson, 2001; Akmaev, Grinevich, 2001; Krokan, Slupphaug, 2004; Lyte, 2004; Мошкин и др., 2006; Goodacre et al., 2009; Pennisi, 2010; Rosenthal et al., 2011) индивидуальный запах организма является результатом переработки микробиотой продуктов его секреции. Было высказано предположение об участии продуктов бактериальной переработки в активации реакции хищника. Наличие реакции борзых на запах микроорганизмов было показано в совместной работе с К.В. Чепелевой (Шубкина и др., 2010, 2012). Установлено, что у борзых существует врожденная, не нуждающаяся в дополнительном подкреплении, способность реагировать на некоторые компоненты запаха их потенциальных жертв, что соответствует не только старинным описаниям охотничьего поведения этих собак (Губин, 1891), но и последним данным о морфологии их обонятельного аппарата (Wagner, Ruf, 2018).
Врожденная реакция борзых на запах культуральной микрофлоры существует и наблюдается значимо чаще, чем ее отсутствие (n тестов = 566, n собак = 40), и проявляется у подавляющего большинства (~90%) борзых. Реакция включает значимо более длительное принюхивание к пробе с культуральной микрофлорой (сравнение с помощью критерия Вилкоксона (Wi), выявляет значимость на уровне p = 0.003) и/или изменение траектории движения по направлению к ней (сравнение с помощью критерия Стьюдента (St), уровень значимости p < 0.05). Она эндогенна, т.е. не нуждается в подкреплении (Чепелева, 2007; Шубкина и др., 2012).
Распознавание запаха микрофлоры и его предпочтение борзыми показало, что даже запах культуральной микрофлоры достаточен для влияния на поведение борзых: он детерминирует изменение траектории и ритмики их движения. Установлено существование и сделано описание врожденной реакции на компоненты запаха потенциальных жертв, представляющие собой результат переработки и трансформации выделений жертвы комменсальными и патогенными бактериями, обитающими на поверхностях ее кожи и слизистых, – микробиотой. Существует особый механизм распознавания доступности или недоступности потенциальной жертвы с участием поверхностной микрофлоры как единого маркера множественных отклонений состояния. Однако для наземных хищных привлекателен запах не всех микроорганизмов, что показывают наблюдения в природе: описаны случаи избегания жертвы, пораженной некоторыми видами паразитов (например, стригущим лишаем).
Микрофлора зайца-русака
Для доказательства наличия повышенного количества микроорганизмов, модулирующих запах особи – потенциальной добычи модельного хищника, проведено сравнение микрофлоры зайцев русаков, пойманных борзыми, и отстрелянных. Установлено, что количество микроорганизмов на единицу площади отпечатка значимо выше в группе “отлов” (оценка тремя методами). В группе “отстрел” значимо чаще встречаются особи с низким количеством микроорганизмов. Также использовали особую группу русаков – животных, отловленных для расселения, ограниченных в подвижности и транспортируемых, что является достаточными факторами для развития состояния стресса (Boonstra et al., 2002). Зайцы, пойманные борзыми, обладают более высокой численностью кожных бактерий. Она сопоставима с таковой у зайцев, подвергнутых стрессу ограничения подвижности, и значимо превышает уровень, типичный для отстрелянных (рис. 7).
Рис. 7.
Встречаемость зайцев с разным количеством микроорганизмов в выборках отлов, отстрел и стресс (частичная иммобилизация).
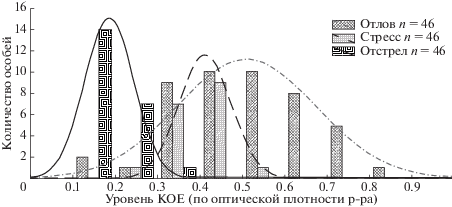
Оценка численности кожной микрофлоры зайцев позволяет утверждать, что у диких животных существует форма изменчивости, которую можно установить с помощью микробиологических методов. Ее наличие маркируется изменением микробиоты поверхности кожи. Микробиота является маркером состояния. Изменение состояния – результат развития стресса различной этиологии, включая заболевания и иные факторы. Таким образом, жертвами борзых становятся животные нездоровые и/или находящиеся в состоянии долговременного стресса.
Существует разнокачественность диких животных, которую нельзя описать по отличиям внешнего вида: различия по благополучию, состоянию здоровья. Одним из маркеров различий является объем кожной микробиоты, что позволяет человеку устанавливать эти отличия неинвазивно, а хищнику – дистантно.
Для хищника менее затратно изъятие неблагополучных особей, но таковыми не всегда являются умирающие. Более того, элиминация заведомо больной жертвы неоднозначна и может иметь негативный эффект. Обеспечивая экономию энергии на получение добычи, она может вести а) к заражению хищника, б) к получению низкого качества продукции (Flick et al., 2016).
До последних лет существовала явная недооценка участия микрофлоры в функционировании макроорганизмов. В последние годы количество исследований возрастает лавинообразно. Принято определять млекопитающих как метагеномных, включающих собственные и прокариотные геномы, т.е. эволюционирующих совместно с ассоциированными микроорганизмами (Ley et al., 2008). Прежде всего, такие исследования подчеркивают значение микрофлоры для макроорганизмов, например кишечной микробиоты. Исследования проходят не только на человеке, но и на животных: например, установлено участие кишечной микрофлоры в гибернации – спячке медведей (Sommer et al., 2016). Значение микрофлоры других полостей и поверхностей тела менее изучено. Однако несомненно, что объединения микробов, эпителиальных клеток и нейрохимические сигналы тесно связаны с развитием стресс-реакции, участвуют в регуляции эффективности метаболизма и формировании устойчивости к заболеваниям (Lyte et al., 2018). Это позволяет утверждать, что подавляющее большинство реакций макроорганизмов опосредуется участием микрофлоры. Менее изучено прямое участие микроорганизмов во взаимодействиях животных, хотя роль бактериальной компоненты в формировании запахов общепризнана.
ЗАКЛЮЧЕНИЕ
Таким образом, комплекс проведенных исследований позволил установить низкую успешность охоты модельного хищника, не превышающую таковую у диких видов. Она сочетается с селективностью (не включающей размерные признаки) по многообразным формам неблагополучия. У жертвы отмечена гипертрофия количества поверхностных микроорганизмов, что изменяет запах особи, а у борзых – выраженная врожденная реакция на запах микрофлоры. Лабораторные эксперименты и результаты современных исследований позволяют утверждать, что маркирование микрофлорой необходимо и достаточно, чтобы обеспечить селективность изъятия жертвы при низкой успешности охоты.
Низкая успешность охоты наземных хищников обусловлена необходимостью множественных преследований, при которых коадаптации жертвы и хищника исключают изъятие наиболее приспособленных особей. При каждом преследовании хищник нуждается в подтверждениях его перспективности, что достигается с помощью запаха жертвы, формируемого при участии сообществ микроорганизмов. Специфическим и важнейшим эволюционным приобретением макроорганизмов является то, что микробиота работает как маркер выраженности приспособительных качеств, важный во внутривидовых и межвидовых взаимодействиях, выполняющий медиаторную функцию во взаимодействии консументов двух трофических уровней.
Все живые существа разнокачественны, а их приспособленность выражена в разной степени. Проблема выбора доступной пищи, требующей минимальных затрат на изъятие и потребление, возникла одновременно с появлением консументов и является их имманентным свойством. Вещества, маркирующие относительную доступность пищевых объектов, вероятно, присутствуют уже у прокариот. Обоняние, продукт эволюции хемотаксиса, является, несомненно, древнейшей сенсорной системой. Таким образом, механизм, сформировавшийся на ранних стадиях эволюции, сохранился и обеспечивает эффективное изъятие наименее приспособленных особей у наземных позвоночных. Поэтому при анализе взаимодействий в пищевых цепях нельзя обойти вниманием микробиоту, опосредующую взаимодействие хищника и жертвы, являющуюся третьим участником и неотъемлемым компонентом этой системы.
Работа выполнена при поддержке грантов РФФИ “Исследование взаимодействия хищника и жертвы в естественном эксперименте” № 13-04-00179, “Механизмы определения доступности жертвы” № 16-04-00732.
Список литературы
Архипчук В.А., Белова О.П., 1987. Разведение зайца-русака на дичефермах // Методические рекомендации. М.: Изд. Союз обществ охотников и рыболовов РСФСР. 24 с.
Банников А.Г., Жирнов Л.В., Лебедева Л.С., Фандеев А.А., 1961. Биология сайгака. М.: Сельхозлит. 335 с.
Бибиков Д.И., Кудактин А.Н., Филимонов А.Н., 1985. Использование территории, перемещение // Волк: история, систематика, морфология, экология / Под ред. Бибикова Д.М. М.: Наука. С. 415–443.
Близнюк А.И., 1983. Смертность сайгаков в Северо-Западном Прикаспии // Периодические явления в жизни животных. М. С. 24–37.
Блохина Т.В., 2016. Сравнительная характеристика состояния зайцев-русаков, добытых двумя способами // Состояние среды обитания и фауна охотничьих животных России и сопредельных территорий. Мат-лы конф., Балашиха, РГАЗУ 2016. С. 34–41.
Бологов В.П., 1980. Поведение волков при нападениях на домашних животных и профилактика ущерба животноводству // Поведение волка. М.: ИЭМЭЖ АН СССР. С. 147–156.
Борискин Г., Чирков М., 1978. Промысел лося в Свердловской области // Охота и охот. х-во. № 8. С. 18–20.
Губин П.М., 1891. Полное руководство ко псовой охоте. М. 347 с.
Ерофеева Е.В., 2014. Неинвазивный метод оценки состояния собак // Человек и животные. Мат-лы VII междунар. заочной конф. Астрахань: Изд-во нижневолжского экоцентра. С. 72–75.
Ерофеева Е.В., 2016. Неинвазивный метод оценки благополучия животных // Состояние среды обитания и фауна охотничьих животных России и сопредельных территорий. Мат-лы конф., Балашиха, РГАЗУ 2016. С. 152–161.
Ерофеева Е.В., 2019. Изменчивость зайца-русака (Lepus europeus) южных регионов РФ // Вестник охотоведения, №3, в печати
Жирнов Л.В., 1982. Возвращенные к жизни: Экология, охрана и использование сайгаков. М.: Лес. Пром. 244 с.
Капланов Л.Г., 1948. Тигр, изюбрь, лось // Мат-лы к познанию фауны и флоры СССР. М.: МОИП. Отд. зоол. Вып. 14. 128 с.
Корытин С.А., Бибиков Д.И., 1985. Охотничье поведение волка // Волк. М.: Наука. С. 295–311.
Крушинский Л.В., Мычко Е.Н., Сотская М.Н., Шубкина А.В., 1985. Элементарная рассудочная деятельность и сложные формы поведения волков // Волк. М.: Наука. С. 284–295.
Марков А., 2009. Иммунная система помогает животным отличать больных сородичей от здоровых по запаху. http://elementy.ru/news/431061
Марков А., 2010. Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы. М.: Астрель, CORPUS. 528 с.
Мошкин М.П., Герлинская Л.А., Нагатоми Р., 2006. Запах, который не лжет. Химическая коммуникация полов и физическое здоровье // Российская наука. М.: “Октопус”. С. 228–239.
Павлов М.П., 1982. Волк. М.: Изд. Лес. Пром. 208 с.
Саблина Т.Б., 1959. Влияние некоторых экологических факторов на физиологию и динамику численности копытных // Совещ. по экол. физиол. 2. Л.: Наука.
Северцов А.С., Шубкина А.В., 2014а. Хищник и жертва – взаимодействие на индивидуальном уровне. Роль хищников в процессе естественного отбора // Зоол. журн. Т. 93. № 6. С. 1–11.
Северцов А.С., Шубкина А.В., 2014б. Хищник и жертва – взаимодействие на индивидуальном уровне. Хищник – механизмы избирательности // Зоол. журн. Т. 93. № 8. С. 1010–1019.
Северцов А.С., Шубкина А.В., 2015а. Эволюционная роль хищных млекопитающих и механизмы ее реализации // Журн. общ. биологии. Т. 76. № 5. С. 355–368.
Северцов А.С., Шубкина А.В., 2015б. Эволюционное значение хищников // Природа. № 6. С. 18–27.
Соколов В.Е., Северцов А.С., Шубкина А.В., 1990а. Моделирование селективного воздействия хищника на жертву: использование борзых собак для отлова сайгаков // Зоол. Журн. Т. 69. № 10. С. 117–125.
Соколов В.Е., Ушакова Н.А., Абрамова О.Ю., Шубкина А.В., 1990б. Реакция микробных ассоциаций участков кожи млекопитающих на изменения физиологического состояния животных // Изв. Акад. наук СССР. Сер. Биол. Вып. 5. С. 694–700.
Соколов В.Е., Ушакова Н.А., Шубкина А.В., Неклюдова Т.И., 1991. Стресс – как фактор, нарушающий стабильность кожных микробных ассоциаций // ДАН СССР. Т. 317. № 3. С. 764–768.
Соколов В.Е., Ушакова Н.А., Приходько В.И., Неклюдова Т.И., Громов В.С., Белоусова И.П., 1990в. Особенности микробных ассоциаций в секретах некоторых кожных желез млекопитающих // Микробиол. Т. 59. № 3. С. 472–481.
Филонов К.П., 1989. Копытные животные и крупные хищники на заповедных территориях. М.: Наука. 252 с.
Фомина И., 2016. Половозрастная структура зайца-русака по результатам псовых охот. Магистерская дис. М.: РГАУ МСХА. 89 с.
Чепелева К.В., 2007. Реакция борзых на запах бактерий // Сб. студенческих науч. работ. Вып. 13. Изд. ФГОУ ВПО РГАУ МСХА им. К.А. Тимирязева. С. 234–237.
Шубкина А.В., 2006. Изучение реакции хищника на жертву с использованием борзых собак в качестве модели // Проблемы исследования домашней собаки. Мат-лы совещания. ИПЭЭ РАН. С. 53–67.
Шубкина А., 2017. Хищник и жертва: взаимодействие особей. Моделирование с помощью борзых. М.: ЛЕНАНД. 264 с.
Шубкина А.В., Северцов А.С., Чаянов Н.В., 2008. Возможности применения аппаратно-программного комплекса на базе GPS для характеристики передвижений и поведения животных степной и лесостепной зон // Зоол. журн. Т. 87. № 11. С. 1–11.
Шубкина А.В., Северцов А.С., Чепелева К.В., 2010. Хищник и жертва: проблема избирательного изъятия на уровне особей // Вестник охотоведения. Т. 7. № 1. С. 5–22.
Шубкина А.В., Северцов А.С., Чепелева К.В., 2012. Факторы, влияющие на изъятие жертвы хищником: моделирование с использованием борзых собак // Изв. РАН. Сер. Биол. № 1. С. 1–13.
Akmaev I.G., Grinevich V.V., 2001. From neuroendocrinology to neuroimmunoendocrinology // Bull. Exp. Biol. Med. V. 131. № 1. P. 15–23.
Alexander E.H., Hudson M.C., 2001. Factors influencing the internalization of Staphylococcus aureus and impacts on the course of infections in humans // Appl. Microbiol. Biotechnol. V. 56. № 3–4. P. 361–366.
Barber-Meyer S.M., Mech L.D., 2008. Factors influencing predation on juvenile ungulates and natural selection implications // Wildl. Biol. Pract. V. 4. № 1. P. 8–29.
Barber-Meyer S.M., White P.J., Mech L.D., 2007a. Survey of selected pathogens and blood parameters of Northern Yellowstone elk: Wolf sanitation effect implications // Am. Midl. Nat. V. 158. P. 369–381.
Barber-Meyer S.M., Mech L.D., White P.J., 2008. Elk calf survival and mortality following wolf restoration to Yellowstone National Park // Wildl. Monogr. № 169. P. 1–30.
Barber-Meyer S.M., Johnson C.R., Murtaugh M.P., Mech L.D., White P.J., 2007b. Interleukin 6 and tumour necrosis alpha factor values in elk neonates // J. Mammol. V. 88. № 2. P. 421–426.
Boonstra R., Takagi N., Bissoon N., Vij S., Gurd J.W., 2002. Trapping-induced changes in expression of the N-methyl-D-aspartate receptor in the hippocampus of snowshoe hares // Neurosci. Lett. V. 324. № 3. P. 173–176.
Braks M.A.H., Anderson R.A., Knols B.G.J., 1995. Infochemicals in mosquito host selection: Human skin microflora and Plasmodium parasites // Parasitol. Today. V. 15. № 10. P. 409–413.
Brown R.E., 1995. What is the role of the immune system in determining individually distinct body odors // Int. J. Immunopharmacol. V. 17. № 8. P. 655–661.
Brown R.E., Schellinck H.M., 1994. Effects of selective depletion of gut bacteria on the odours of individuality in rats // Advances in the Biosciences. V. 93: Chemical Signals in Vertebrates VII. Oxford: Pergamon. P. 267–271.
Cheatum E. L., 1949. The use of corpora lutea for determin-ing ovulation incidence and variations in the fertility of white-tailed deer // Cornell Vet. V. 39. P. 282–291.
Goodacre S.L., Martin O.Y., Bonte D., Hutchings L., Woolley C. et al., 2009. Microbial modification of host long-distance dispersal capacity // BMC Biol. V. 7. № 32. https://doi.org/10.1186/1741-7007-7-32
Fischbach M., Bromley P., 2001. Recombinant cell lines for stress reporter assays // Cell Biol. Toxicol. V. 17. № 4–5. P. 335–351.
Flick A.J., Acevedo M.A., Elderd B.D., 2016. The negative effects of pathogen-infected prey on predators: A meta-analysis // Oikos. V. 125. № 11. P. 1554–1560.
Franzmann A.W., Arneson P.D., 1976. Marrow fat in Alaskan moose femurs in relation to mortality factors // J. Wildl. Manag. V. 40. № 2. P. 336–339.
Hayward M.W., Hayward G.J., Tambling C.J, Kerley G.I., 2011. Do lions Panthera leo actively select prey or do prey preferences simply reflect chance responses via evolutionary adaptations to optimal foraging? // PLoS One. V. 6. № 9. P. e23607.
Hilborn A., Pettorelli N., David C., Orme L., Durant S.M., 2012. Stalk and chase: How hunt stages affect hunting success in Serengeti cheetah // Anim. Behav. V. 84. № 3. P. 701–706.
Hilborn A., Pettorelli N., Caro T., Kelly M.J., Laurenson M.K., Durant S.M., 2018. Cheetahs modify their prey handling behavior depending on risks from top predators // Behav. Ecol. Sociobiol. V. 72. № 4. P. 74.
Jezirsky W., 1965. Studies on the European hare // Acta Theriol. V. 10. № 2. P. 11–25.
Khater M., Murariu D., Gras R., 2015. Predation risk tradeoffs in prey: Effects on energy and behaviour // Theor. Ecol. V. 9. № 3. P. 251–268.
Krokan H.E., Slupphaug B.K.G., 2004. Novel aspects of macromolecular repair and relationship to human disease // J. Mol. Med. V. 82. № 5. P. 280–297.
Latham A.D.M., Latham M.C., McCutchen N.A., Boutin S., 2011. Invading white-tailed deer change wolf–caribou dynamics in Northeastern Alberta // J. Wildl. Manag. V. 75. № 1. P. 204–212.
Ley R.E., Hamady M., Lozupone C., Turnbaugh P.J., Ramey R.R. et al., 2008. Evolution of mammals and their gut microbes // Science. V. 320. № 5883. P. 1647-1651.
Lyte M., 2004. Microbial endocrinology and infectious disease in the 21st century // Trends Microbiol. V. 12. № 1. P. 14–20.
Lyte M., Villageliú D.N., Crooker B.A., Brown D.R., 2018. Symposium review: Microbial endocrinology – Why the integration of microbes, epithelial cells, and neurochemical signals in the digestive tract matters to ruminant health // J. Dairy Sci. V. 101. № 6. P. 5619–5628.
MacNulty D.R., Mech L.D., Smith D.W., 2007. A proposed ethogram of large–carnivore predatory behavior, exemplified by the wolf // J. Mammal. V. 88. № 3. P. 595–605.
Mech L.D., 1970. The Wolf: The Behavior and Ecology of an Endangered Species. N.Y.: Natural History Press. 384 p.
Mech L.D., 2007. Femur-marrow fat of white-tailed deer fawns killed by wolves // J. Wildl. Manag. V. 71. № 3. P. 920–923.
Mech L.D., Delgiudice G.D., 1985. Limitations of the marrow-fat technique as an indicator of body condition // Wildl. Soc. Bull. V. 13. № 2. P. 204–206.
Misiorowska M., Ludwisiak L., Nasiadka P., 2014. Population parameters of brown hare (Lepus europaeus L.) in regions of the species highest density in Poland // Sylwan. V. 158. № 12. P. 901–910.
Moshkin M.P., Gerlinskaya L.A., Morozova O.V., Bakhvalova V.N., Evsikov V.I., 2002. Behaviour, chemosignals, and endocrine functions in male mice infected with tick-borne encephalitis virus // Psychoneuroendocrinol. V. 27. № 5. P. 603–608.
Nilsson P.A., 2001. Predator behaviour and prey density: Evaluating density-dependent intraspecific interactions on predator functional responses // J. Anim. Ecol. V. 70. P 14–19.
O’Gara B., Harris R., 1988. Age and conditions of deer killed by predators and automobiles // J. Wildl. Manag. V. 52. № 2. P. 316–320.
Okarma H., 1991. Marrow fat-content, sex and age of red deer killed by wolves in winter in the Carpathian mountains // Holarctic Ecol. V. 14. № 3. P. 169–172.
Owen-Smith N., Mills M.G., 2008a. Shifting prey selection generates contrasting herbivore dynamics within a large-mammal predator-prey web // Ecology. V. 89. № 4. P. 1120–1133.
Owen-Smith N., Mills M.G., 2008b. Predator-prey size relationships in an African large-mammal food web // J. Anim. Ecol. V. 77. № 1. P. 173–183.
Penn D., Potts W.K., 1998. Chemical signals and parasite-mediated sexual selection // Trends Ecol. Evol. V. 13. № 10. P. 391–396.
Pennisi E., 2010. Body’s hardworking microbes get some overdue respect // Science. V. 330. № 6011. P. 1619.
Preisser E.L., 2009. The physiology of predator stress in free-ranging prey // J. Anim. Ecol. V. 78. № 6. P. 1103–1105.
Preisser E.L., Orrock J.L., Schmitz O.J., 2007. Predator hunting mode and habitat domain alter nonconsumptive effects in predator-prey interactions // Ecology. V. 88. № 11. P. 2744–2751.
Rosenthal M., Goldberg D., Aiello A., Larson E., 2011. Skin microbiota: Microbial community structure and its potential association with health and disease // Infect. Genet. Evol. V. 11. № 5. P. 839–848.
Schellinck H.M., Brown R.E., 2000. Selective depletion of bacteria alters but does not eliminate odors of individuality in Rattus norvegicus // Physiol. Behav. V. 70. № 3–4. P. 261–270.
Severtsov A.S., Rosenzweig M.L., Shubkina A.V., 2017. Predators detect the welfare of their potential prey and cull those that are poorly // Evol. Ecol. Res. V. 18. P. 556–569.
Shubkina A., Ushakova N., 1994. The possible role of odors of skin bacteria in mammalian prey selection // Adv. Biosci. V. 93. P. 259–266.
Shubkina A., Ushakova N., 1996. Mammalian predator-prey system and the possible role of odour of skin bacteria // Lutreola. № 7. P. 13–18.
Sinclair A.R.E., Arcese P., 1995. Population consequences of predation-sensitive foraging: The Serengeti wildebeest // Ecology. V. 76. № 3. P. 882–891.
Sokos C., Touloudi A., Iakovakis C., Papaspyropoulos K., Giannakopoulos A. et al., 2015. Hare demography, physiology and European Brown Hare Syndrome virus infection: Is there an association? // J. Zool. V. 298. № 2. P. 150–157.
Sommer F., Stahlman M., Ilkayeva O., Arnemo J.M., Kindberg J. et al., 2016. The gut microbiota modulates energy metabolism in the hibernating brown bear Ursus arctos // Cell Rep. V. 14. № 7. P. 1655–1661.
Steinert M., Hentschel U., Hacker J., 2000. Symbiosis and pathogenesis: Evolution of the microbe-host interaction // Naturwissenschaften. V. 87. № 1. P. 1–11.
Suchentrunk F., Michailov C., Markov G., Haiden A., 2000. Population genetics of Bulgarian brown hares Lepus europaeus allozymic diversity at zoogeographical crossroads // Acta Theriol. V. 45. № 1. P. 1–12.
Thulin C.-G., Malmsten J., Laurila A., 2012. Differences in body mass, health status and genetic variation between insular and mainland brown hares (Lepus europaeus) in Sweden // Eur. J. Wildl. Res. V. 58. № 6. P. 897–907.
Verhulst N., Mukabana W.R., Takken W., Smallegange R., 2011. Human skin microbiota and their volatiles as odour baits for the malaria mosquito Anopheles gambiae s.s // Entomol. Exp. Appl. V. 139. № 2 P. 170–179.
Verhulst N.O., Beijleveld H., Knols B.G.J., Takken W., Schraa G. et al., 2009. Cultured skin microbiota attracts malaria mosquitoes // Malaria J. V. 8. № 302. P. 1–12.
Voigt C.C., Caspers B., Speck S., 2005. Bats, bacteria, and bat smell: Sex-specific diversity of microbes in a sexually selected scent organ // J. Mammal. V. 86. № 4. P. 745–749.
Wagner F., Ruf I., 2019. Who nose the borzoi? Turbinal skeleton in a dolichocephalic dog breed (Canis lupus familiaris) // Mammal. Biol. V. 94. P. 106–119.
Werner J., Morgan A., 2017. An approach to investigate ‘apparent starvation’ in moose populations: A preliminary scoping // Technical Report. https://doi.org/10.13140/RG.2.2.28077.90085
Wilson M., Lowe J.C., Roskilly K., Hudson P.E., Golabek K.A., McNutt J.W., 2013. Locomotion dynamics of hunting in wild cheetahs // Nature. V. 498. № 7453. P. 185–189.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии