

Журнал высшей нервной деятельности им. И.П. Павлова, 2020, T. 70, № 5, стр. 696-720
Автоотождествление и сенсорно-моторное повторение как физиологические механизмы сознания
В. Я. Сергин *
Институт математических проблем биологии РАН
г. Пущино, Россия
* E-mail: v.sergin@mail.ru
Поступила в редакцию 28.10.2019
После доработки 12.01.2020
Принята к публикации 26.02.2020
Аннотация
В статье рассматриваются природа и нейрофизиологические механизмы сенсорного осознания, сознательного восприятия, порождения мысли, воображения, интуиции и дискурсивного мышления. Показано, что в основе сенсорного осознания и порождения мысли лежат высокочастотные циклические процессы автоотождествления. В основе воображения и дискурсивного мышления лежат циклические процессы сенсорно-моторного повторения. Повторение является низкочастотным процессом (3–6 Гц), и его содержание доступно обозрению с помощью высокочастотных процессов автоотождествления (30–70 Гц). Череда мысленных образов, слов или символов управляема посредством моторной системы и доступна обозрению посредством процессов автоотождествления. Взаимодействующие механизмы сенсорно-моторного повторения и автоотождествления позволяют нам формировать образы, сцены и диалоги, наблюдать и изменять их, создавая подвижный и управляемый мир сознательного опыта. Показано, что механизмы автоотождествления и сенсорно-моторного повторения позволяют объяснить природу широкого спектра свойств сознательной активности мозга, включая такие пока еще загадочные явления, как гипноз и медитация.
ВВЕДЕНИЕ
В начале статьи полагается давать определение предмета исследования, в нашем случае – определение сознания. Но описать сущность или дать точное определение сознания до того, как станут известны порождающие его психофизиологические механизмы, вероятно, невозможно. К этой проблеме мы еще вернемся, а пока в качестве исходной посылки примем представления, существующие в повседневной медицинской практике относительно возникновения или исчезновения сознания у пациентов. Признаком сознательного состояния пациента является его способность отвечать на вопросы врача о своем состоянии и об окружающей обстановке. То есть признаком сознательного состояния субъекта является его способность сообщить свое знание другому [Симонов, 1987]. Но чтобы передать свое знание другому, необходимо сначала представить его себе. Тогда представление собственного знания самому себе, возможно, лежит в основе процесса осознания.
Что значит представить собственное знание самому себе? Пока знание хранится в памяти, оно существует в неявной (скрытой) форме. Неявное знание – это знание существующее, но не явленное, не представленное себе. Извлеченное из памяти знание, представленное себе подобно внешним данным, становится явным. Тогда механизм представления внутренних данных в явной форме может лежать в основе процесса осознания.
Действительно, способность осуществлять явное представление хранящихся в памяти данных подобно представлению внешних объектов является хорошо известным свойством сознания. Функционирование сознания всегда связано с явными представлениями образов, символов, звуков, запахов, прикосновений и т.п., и неотделимо от них. Никакая сознательная деятельность мозга не возможна без явного представления данных, которое, возможно, является ключевым свойством осознания. Если это действительно так, то какой нейробиологический механизм может осуществлять явное представление данных и тем самым порождать психически переживаемый феномен их осознания? Ответу на этот вопрос посвящены недавние работы [Сергин, 2016; Sergin, 2017]. Здесь мы ограничимся только кратким изложением механизмов осознания, которые необходимы для понимания более широких аспектов сознательной активности мозга.
Можно назвать две частные проблемы, решение которых могло бы пролить свет на природу и механизмы сознания. Важнейшей из них является выяснение нейронных механизмов, порождающих феномен осознания внешних и внутренних событий. Необходимо понять, как мы осознаем что бы то ни было, например, вспышку света, запах или боль. Другая проблема связана со способностью человека к произвольному оперированию знаниями. Мы оперируем знаниями в процессах рассудочного и образного мышления. Феноменологически эти процессы выражаются мыслями. Что такое мысль и какие нейрофизиологические механизмы лежат в основе наших способностей к мышлению?
Целью данной статьи является построение концептуальной модели сознания, основанной на рассмотрении двух названных частных проблем. Представляется интересным и важным выяснить, как далеко можно продвинуться в понимании природы широкого спектра феноменов сознательной активности мозга на основе решения этих частных проблем.
АВТООТОЖДЕСТВЛЕНИЕ КАК КЛЮЧЕВОЙ МЕХАНИЗМ ОСОЗНАНИЯ
В этом разделе нас будет интересовать нейробиологический механизм, который может осуществлять явное представление данных и тем самым порождать психически переживаемый феномен их осознания. Для начала зададимся более простым, но более фундаментальным вопросом. Каким образом нейронные структуры головного мозга приобретают способность отображать сенсорные объекты? Естественным и самым простым способом отображения специфического сенсорного возбуждения является формирование нейронной структурой такого выходного паттерна нейронной активности, который был бы тождественным паттерну входного возбуждения. Условие тождественности выходного и входного паттернов нейронной активности открывает возможность обучения нейронной структуры без учителя. В таком процессе об-учения выходной паттерн нейронной активности может передаваться на собственный вход посредством положительных обратных связей. Если паттерн обратной связи совпадает с паттерном входного возбуждения, то они суммируются, что увеличивает интенсивность специфического возбуждения. Увеличение интенсивности возбуждения на входе ведет к росту выходного возбуждения. По правилу Хебба [Hebb, 1949], интенсивное пред- и постсинаптическое возбуждение увеличивают эффективность тех синапсов, которые обеспечивают порождение паттерна данной специфичности. Интенсивный рост специфического возбуждения в нейронной структуре с положительными обратными связями обеспечивает интенсивное увеличение эффективности отобранных синапсов. В результате такого обучения нейронная структура приобретает способность отвечать избирательно на сенсорное возбуждение данной специфичности (на данный сенсорный объект).
Аналогичным образом может происходить обучение нейронной структуры отвечать избирательно на паттерны возбуждения других специфичностей. Набор специфических паттернов входного возбуждения, на которые данная нейронная структура отвечает тождественной выходной активностью, характеризует долговременную память этой структуры. Память нейронной структуры анатомически закрепляется специфическим распределением эффективности синапсов, которое может меняться в процессах дальнейшего обучения.
Механизм автоотождествления мог возникнуть в процессе эволюции как способ самообучения и самоорганизации нейронных структур. Эффективность этого механизма вела к кардинальному расширению разнообразия его функций в растущем мозге. Нас будет интересовать пока роль этого механизма в порождении субъективных феноменов.
Как известно, в процессе восприятия стимул порождает в одной или нескольких областях коры специфический паттерн возбуждения. Можно предположить, что выходные нейроны этих областей коры формируют паттерн нейронной активности, тождественный паттерну входного возбуждения. Выходной паттерн нейронной активности может передаваться на входы тех же областей коры через автономные массированно-параллельные обратные связи. Тождественные (совпадающие в основных чертах) паттерны возбуждения – порождаемый стимулом и передаваемый по обратным связям – складываются на одних и тех же нейронных структурах (рис. 1), вызывая разрядку все большего числа нейронов и увеличивая интенсивность возбуждения этих структур. Такой циклический процесс с положительной обратной связью взрывообразно увеличивает интенсивность специфического паттерна возбуждения. Кроме того, порождаемое стимулом интенсивное возбуждение может вызывать динамическое торможение окружающих нейронных структур [Mountcastle 1978, 1998]. В результате паттерн возбуждения может приобретать высокую контрастность. Специфичность пространственного возбуждения коры акцентирует специфические характеристики стимула, обеспечивая его категоризацию [Сергин, 2009а; 2016; Sergin, 2000; 2017].
Рис. 1.
Локальная схема процесса автоотождествления. Связи с другими областями коры и подкорковыми структурами не показаны [Сергин, 2009а].
Fig. 1. The local scheme of the process of autoidentification. Connections with other areas of the cortex and subcortical structures are not shown [Sergin, 2009a].
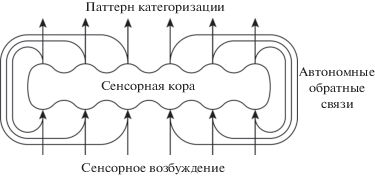
Результат категоризации – цвет, звук, запах – выражает субъективный смысл сенсорного стимула. Категоризация является ответом распределенной долговременной памяти на входное возбуждение и физиологически выражается специфическим паттерном нейронной активности коры. Паттерн категоризации передается на вход и включается в цикл автоотождествления, обеспечивая интенсивное отображение субъективного смысла стимула.
Отображение сенсорных категорий (внутренних данных) паттернами входной нейронной активности коры есть представление этих категорий субъекту в качестве элементов отображения внешнего мира. В результате внешний мир оказывается представленным субъекту не в объективных характеристиках физического мира, а в сенсорных категориях: цвет, вкус, запах, тактильные ощущения и т.п. Это и есть первичные субъективные качества, субъективные ощущения.
Формирование выходного паттерна нейронной активности (паттерна категоризации), тождественного специфическому паттерну входного возбуждения, является процессом автоотождествления. Паттерны нейронной активности являются тождественными, если мозг интерпретирует их как одинаковые сенсорные объекты. Необходимым условием реализации процесса автоотождествления является наличие автономных массированно-параллельных положительных обратных связей, которые могут порождать взрывообразный рост интенсивности специфического паттерна возбуждения коры.
Механизм автоотождествления отбирает, посредством итерационной процедуры, такой специфический паттерн сенсорного возбуждения, на который данная нейронная структура отвечает наиболее высокой активностью [Сергин, 2016; Sergin, 2017]. В результате данное сенсорное возбуждение интерпретируется наиболее подходящим вариантом сенсорной категории, которой располагает долговременная память. Это значит, что в процессе восприятия доминирует прошлый опыт, который делает возможным адекватную биологическую интерпретацию сенсорного объекта даже в условиях помех или по частичным сенсорным данным. Но, с другой стороны, это значит, что мы воспринимаем скорее то, что знаем, чем то, что объективно регистрируют рецепторные органы.
Следует подчеркнуть, что осознается не входное возбуждение, а сенсорные категории, которые порождаются нейронной структурой коры в ответ на входное возбуждение. Сенсорные категории – это внутренние данные, которые содержатся в памяти, а процесс автоотождествления является способом представления внутренних данных в явной форме. Это значит, что внешнее событие сначала должно быть воспринято, т.е. представлено в сенсорных категориях, и только потом мозг сможет осознать его. Осознание является формой вторичной обработки данных, а процессы неосознаваемого восприятия и осознания оказываются разделенными по времени.
Сенсорное осознание – это осознание внешних событий, которые отображаются паттернами нейронной активности коры и становятся внутренними сигналами мозга. В процессе мышления операционная активность мозга также отображается паттернами нейронной активности коры. Поэтому нейрофизиологический механизм осознания результатов операционной активности мозга может быть аналогичен механизму осознания сенсорных сигналов. Механизм автоотождествления оказывается универсальным аппаратом осознания сигналов, как порождаемых сенсорным входом, так и генерируемых самим мозгом.
Существует ли механизм автоотождествления в действительности? Если этот циклический механизм действительно существует, то его функционирование должно порождать вполне определенные и предсказуемые свойства процесса осознания. Тогда сопоставление теоретически предсказанных и экспериментально установленных свойств сознательного восприятия позволит подтвердить или опровергнуть существование механизма автоотождествления.
Автоотождествление осуществляется за время циркуляции сигнала по замкнутому контуру, и его результатом является единичное событие – осознание сигнала. Следовательно, механизм автоотождествления порождает дискретные события с интервалом дискретности, равным времени цикла.
Можно дать априорную оценку вероятной длительности цикла автоотождествления, исходя из экологических условий восприятия. Из постулируемого механизма следует, что любой сигнал должен пройти, по крайней мере, один цикл автоотождествления, чтобы мозг мог осознать его. Тогда время цикла должно быть меньше характерных временных интервалов изменений в окружающей среде, существенных для выживания организма. В противном случае важные изменения в окружающей обстановке будут происходить, а их осознание не будет успевать за событиями. Витально значимые события, такие как быстрые движения животных, погоня, прыжки, удары лапой, а также время оборонительной реакции, характеризуются спектрами, высокочастотные компоненты которых имеют периоды около 0.1 сек. Измеренные спектры турбулентности в приземном слое атмосферы, связанные с вихрями и ливнями, колебаниями веток деревьев, кустарников, травы и другими вариациями окружающей среды, имеют резкий спад около 0.1 сек. Время цикла – это период квантования непрерывного сигнала. Представление непрерывного сигнала дискретной выборкой требует, по крайней мере, двух отсчетов за период самой высокочастотной компоненты спектра непрерывного сигнала. Тогда период квантования должен быть меньше или приблизительно равным 50 мс, что является теоретической оценкой времени цикла автоотождествления.
Время цикла 50 мс соответствует частоте циклических процессов около 20 Гц. Тогда нейронные структуры сенсорной коры могут обладать собственными частотами, близкими к этой величине. Эксперименты по транскраниальной магнитной стимуляции (ТМС) мозга подтверждают наше предположение. В этих экспериментах катушка ТМС размещается на поверхности головы над зрительной затылочной долей. Переменный ток в катушке порождает магнитное поле, которое индуцирует электрическую активность в зрительной коре. Испытуемый с закрытыми глазами ощущает (видит) слабые вспышки света – фосфены [Walsh, Cowey, 2000]. Интенсивность фосфенов зависит от частоты стимуляции и имеет четкий максимум на частоте около 20 Гц [Баарс, Гейдж, 2014].
Осознание посредством автоотождествления паттернов нейронной активности может происходить только в той области коры, которая отображает соответствующее сенсорное качество. Эксперименты показывают, что ТМС проекционной зрительной коры V1 вызывает маленькие неподвижные фосфены, а ТМС области МТ порождает движущиеся фосфены [Pascual-Leone, Walsh, 2001]. Таким образом, результаты экспериментов по транс-краниальной магнитной стимуляции подтверждают теоретические предсказания.
АВТООТОЖДЕСТВЛЕНИЕ И ЭКСПЕРИМЕНТАЛЬНЫЕ ДАННЫЕ
Циклическая обработка информации должна проявляться в особенностях восприятия непрерывности движения (действительного или кажущегося). Примером может служить восприятие кинофильма. Время цикла является минимально различимым временем, поэтому дискретная последовательность кадров кинофильма должна восприниматься как непрерывная, если последовательные кадры попадают в каждый цикл автоотождествления. Если кадры фильма предъявляются через цикл или реже, то такая последовательность может восприниматься как дискретная. Таким образом, из модели автоотождествления следует необходимость существования некоторой критической частоты, выше которой наблюдается непрерывное движение, а ниже – дискретное. Причем численное значение этой критической частоты должно соответствовать частоте циклов. Такая частота действительно существует, она известна как критическая частота слияния мельканий и составляет 10–50 Гц, что соответствует времени цикла 20–100 мс. Эта экспериментальная оценка времени цикла хорошо соответствует нашей теоретической оценке.
Гипотеза автоотождествления влечет за собой точно формулируемые следствия в отношении временных характеристик осознания сигналов. Анализ обширных психофизиологических данных, относящихся к таким феноменам как движение, временной порог, временная суммация, обратная маскировка, слияние мерцаний и многих других, позволяет оценить время цикла автоотождествления величиной порядка нескольких десятков миллисекунд, которая может варьировать в диапазоне от 10 до 100 мс [Сергин, 1994; 1998; 2016; Sergin, 1991; 1992; 1994; 2000; 2017]. Следовательно, соответствующую полосу частот циклических процессов можно оценить диапазоном приблизительно от 10 до 100 Гц.
Частота циклических процессов автоотождествления является физиологической характеристикой, она может быть различной у разных людей и зависеть от состояния субъекта и интенсивности переживаемых событий, аналогично тому, как это имеет место для других периодических процессов, например, частоты пульса и дыхания. На увеличение потока информации $I$ по любому перцептивному каналу механизм автоотождествления должен отвечать увеличением частоты циклов ${{f}_{c}}$, чтобы успевать включать ее в процесс осознания. Следовательно, ${{f}_{c}}\sim I$. Это соотношение имеет две асимптотики. Если поток информации очень велик, частота может достигать значения, увеличение которого физиологически невозможно. При дальнейшем росте входной информации механизм автоотождествления не может справляться с нагрузкой, что означает функциональный отказ сознания как системы обработки данных и физиологически может выражаться стрессом. Если поток входной информации очень мал, частота падает до минимума, соответствующего состоянию релаксации, что может выражаться ощущениями замедленного течения мыслей, ленивой созерцательности, скуки и пустоты. Следовательно, зависимость частоты циклических процессов от потока входной информации можно представить возрастающей функцией с двумя асимптотиками, около 10 и 100 Гц [Сергин, 1994; 2016; Sergin, 1994; 2017], как это показано на рис. 2.
Рис. 2.
Зависимость частоты циклических процессов автоотождествления ${{f}_{c}}$ от потока входной информации I (сплошная линия). Пунктиром показаны особые режимы функционирования сознания 1 и 2, которые поясняются в конце статьи.
Fig. 2. The dependence of the frequency of cyclic autoidentification on the input information stream I (solid line). The dotted line shows the special modes of functioning of consciousness 1 and 2, which are explained at the end of the article.
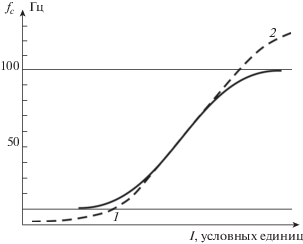
Процессы автоотождествления реализуются в обширных областях коры головного мозга. Если эти циклические процессы действительно существуют, то они могут быть обнаружены прямыми измерениями электрической активности коры. Ф. Крик и К. Кох [Crick, Koch, 1990; Koch, Crick, 1994] выполнили анализ обширных электрофизиологических данных и обнаружили, что зрительное осознание коррелирует с высокочастотной электрической активностью коры головного мозга в диапазоне 30–70 Гц. Этот диапазон точно соответствует центральной полосе частотной характеристики аппарата осознания, представленной на рис. 2.
Данные электро- и магнитоэнцефалографии показывают, что интенсивная сознательная активность мозга характеризуется высокочастотной электрической активностью коры в бета- (14–30 Гц) и гамма- (30–100 Гц) диапазонах. В состоянии релаксации, в отсутствие внешних стимулов и при закрытых глазах электрическая активность смещается в низкочастотную область с преобладанием альфа-ритма (8–13 Гц) [Кропотов, 2010]. Таким образом, высокочастотные колебания электрической активности в диапазоне от 10 Гц до 100 Гц, которые должны порождаться процессами автоотождествления в коре головного мозга, не только существуют в действительности, но и являются доминирующими.
Анатомическим базисом осознания являются автономные обратные связи коры головного мозга. Обратные связи являются автономными, если они начинаются с выходных популяций нейронов данной области коры и заканчиваются на входных популяциях этой же области. Автономные обратные связи могут замыкаться через ядра подкорковых структур, но не через другие области коры.
Из анатомических и нейрофизиологических данных известно, что структура коры головного мозга состоит из сотен миллионов вертикально ориентированных популяций нейронов (мини-колонок) с множеством связей по вертикали и с относительно небольшим числом горизонтальных связей [Mountcastle, 1978; 1998]. Эти модули обработки содержат массированные корко-корковые обратные связи, передающие возбуждение непосредственно или через ядра подкорковых структур. Установлено также, что подавляющее большинство обратных связей являются положительными. Такая анатомическая организация вполне обеспечивает формирование автономных обратных связей в распределенных структурах коры.
Система таламо-кортикальных связей кажется вероятным кандидатом на роль аппарата осознания внешних и внутренних сигналов. Реализация процессов автоотождествления требует массированно-параллельных замкнутых цепей, и это объясняет тот факт, что связи в таламо-кортикальной системе всегда двусторонние. Такие замкнутые связи должны охватывать все области коры, которые участвуют в осознании внешних или внутренних сигналов.
Другим кандидатом на роль анатомической основы аппарата автоотождествления может быть распределенная система “кора–базальные ганглии–таламус–кора” [Силькис, 2006; 2011; 2017]. Мы не будем останавливаться здесь на проблемах, которые обсуждаются в работах [Сергин, 2016; Sergin, 2017] и требуют дальнейших исследований.
Процессы автоотождествления могут порождать основную часть наблюдаемых высокочастотных осцилляций электрической активности коры в бета- и гамма-диапазонах, как во время активного бодрствования, так и в быстрой фазе сна. Таким образом, природа высокочастотных колебаний электрической активности головного мозга, возможно, связана с циклическими процессами в нейронных структурах с положительными обратными связями, а не с предполагаемыми осцилляциями в нейронных сетях коры. Если это действительно так, то осознание достигается дорогой ценой: на это может расходоваться значительная часть всей доступной энергии мозга.
Компьютерные исследования нейронных сетей показывают, что тормозные нейроны играют определяющую роль в генерации осцилляций [Кропотов, 2010]. Но 90% нейронов новой коры являются возбуждающими. Тормозных нейронов совсем немного, и их функции могут быть совершено иными, например, такими, как латеральное торможение. Это, конечно, не означает, что не может существовать нейронных структур с отрицательными обратными связями, построенными на тормозных нейронах. Такие структуры существуют и могут вносить свой вклад в электрическую активность коры.
Завершая краткое рассмотрение модели автоотождествления, следует указать на более ранние исследования, которые предлагались для объяснения феномена осознания. А.М. Иваницкий [А.М. Иваницкий, 1976, 1987, 1996; Иваницкий и др., 1984] высказал идею о том, что синтез информации о физических свойствах стимула и извлекаемых из памяти сведений о его значимости на нейронах проекционной коры составляет ключевой механизм возникновения ощущения как психического феномена. Синтез осуществляется путем сопоставления сенсорных сигналов и сигналов обратных связей, поступающих из подкорковых структур и ассоциативной коры в проекционную область. Этот процесс с обратными связями повторяется, образуя кольцо ощущений с периодом 100–150 мс.
Дж.M. Эделмен [Edelman 1978, 1981, 1989] предложил идею, согласно которой осознание может порождаться повторной передачей на вход фиксируемых в памяти сигналов. При этом сигналы, фиксируемые в памяти, поступают опять на вход так же, как и внешние сигналы. Повторный вход сигналов позволяет ассоциировать их с содержимым памяти. Тогда осознание может представлять собой циклический процесс ассоциативного воспоминания – “вспоминаемое настоящее”.
Непреходящая ценность пионерских работ А.М. Иваницкого и Дж. Эделмена состоит в том, что научное объяснение феномена сенсорного осознания стало возможным без редукции к “гомункулусу”. В этих работах двумя различными способами показано, как введение обратных связей делает “гомункулус” не нужным.
Модель автоотождествления внешне напоминает модель повторного входа Дж. Эделмена, поэтому следует указать их отличия.
– Автоотождествление осуществляется посредством массированно-параллельных автономных обратных связей: выходы данной области коры замыкаются на ее собственный вход через подкорковые структуры. Повторный вход сигналов в модели Дж. Эделмена осуществляется с помощью реципрокных обратных связей между различными областями коры [Edelman, 1978; 1989].
– Автоотождествление порождает высокочастотные колебания электрической активности коры в диапазоне от 10 до 100 Гц. Время повторного входа сигналов в модели Дж. Эделмена оценивается величиной от 100 до 300 мс, что соответствует частотам от 10 до 3 Гц. [Edelman, 1989; Edelman, Tononi, 2000].
– Автоотождествление является циклическим процессом, который порождает дискретный процесс осознания. Повторный вход сигналов в модели Дж. Эделмена формирует непрерывный процесс осознания [Edelman, 1989; Edelman, Tononi, 2000].
ОРГАНИЗАЦИЯ ПРОЦЕССОВ ВОСПРИЯТИЯ
Систему сознательного восприятия, по одному из сенсорных путей, можно представить схемой, показанной на рис. 3. Хотя процессы неосознаваемого восприятия и осознания тесно связаны между собой и протекают в одних и тех же областях коры, их нейронная организация различна. Из механизма автоотождествления следует, что осознание является формой вторичной обработки данных, поэтому процессы неосознаваемого восприятия и осознания разделены по времени, а их нейронные аппараты должны быть функционально обособлены.
Рис. 3.
Функциональная схема системы сознательного восприятия. Восходящее сенсорное возбуждение и обратные связи показаны одинарными линиями. Двойными линиями показаны автономные обратные связи аппарата автоотождествления.
Fig. 3. Functional diagram of the system of conscious perception. Upward sensory stimulation and feedbacks are shown by single lines. The double lines show the autonomous feedbacks of the autoidentification device.
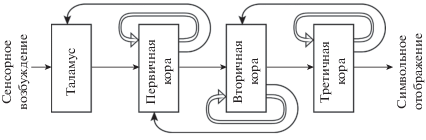
Если из системы сознательного восприятия, представленной схемой на рис. 3, исключить автономные обратные связи аппарата автоотождествления, получим схему неосознаваемого восприятия. Модель системы восприятия описана в нескольких статьях [Сергин, 2002, 2009б; Sergin, 2003; A. Sergin, V. Sergin, 2008; В. Сергин, А. Сергин, 2019]. В ее основе лежит известный нейробиологический факт: размеры рецептивных полей нейронов растут вверх по сенсорным путям. В такой модели на каждом уровне восприятия формируется характеристика, объемлющая отобранные данные нижележащего уровня. Объемлющая характеристика – это перцептивная гипотеза о том, что данная специфическая комбинация признаков соответствует определенному сенсорному объекту. Объемлющая характеристика выражает некоторую целостность, обладающую качественной специфичностью на более высоком уровне восприятия. Например, наборы линий образуют решетку, последовательность звуков – ритм, глаза и нос – лицо и т.п.
Следует подчеркнуть отличие объемлющей характеристики от интегральной. Интегрирование в своей сущности является процессом суммирования (определяемое как предел аппроксимирующей суммы фрагментов при их неограниченном измельчении). Интегральная форма представления – это представление суммарных данных. Интегральные характеристики (виды интегралов и процедуры их вычисления) формально определены, что обеспечивает объективное представление данных.
Объемлющая характеристика не обязательно выражает объективно важные или физически реальные свойства сенсорного объекта, которые организму не известны. Она выражает биологически значимые свойства объекта и представляет собой адаптивную реакцию организма на этот объект. Это значит, что объемлющая характеристика изначально субъективна и должна быть чувствительной к индивидуальным различиям.
В системе восприятия сенсорное возбуждение передается по иерархии корковых зон от проекционных к ассоциативным областям. Обратные связи системы восприятия передают нисходящее возбуждение. Нисходящее возбуждение избирательно активирует нейронные ансамбли нижележащих уровней, облегчая передачу тех компонентов сенсорного возбуждения, которые соответствуют объемлющей характеристике. Например, если объемлющей характеристикой является решетка, то могут активироваться нейронные популяции нижележащих уровней, реагирующие на линии определенных ориентаций.
Последовательность объемлющих характеристик составляет иерархию от сенсорных признаков до целостных образов и сцен. Объемлющая характеристика не содержит в себе данных нижележащего уровня, а только связывает их в некоторую целостность. Чем выше уровень в системе восприятия, тем меньше физических подробностей содержит объемлющая характеристика и тем более уникальные семантические свойства окружающего мира она выражает. Высшая объемлющая характеристика выражает ключевые черты целостного образа, его схему, концепцию.
Важно заметить, что высшая объемлющая характеристика – это не образ и не сцена, а уникальная схема сочетания объектов нижележащих уровней, порождающих образ или сцену. Образы и сцены могут отображаться иерархиями вложенных сенсорных характеристик, что обеспечивает детальность (за счет нижних уровней) и целостность отображений (за счет верхних уровней). Высшая объемлющая характеристика единственна в каждом акте восприятия.
Возбуждение сверху (через вставочные нейроны или через подкорковые структуры) переводит нейроны отобранных популяций нижележащего уровня в состояние, близкое к порогу разрядки [Сергин, 2007; 2009б]. Известно [Кропотов, 2010], что состояние, близкое к порогу разрядки, характерно для нейронов коры. Ответ нейронного ансамбля в предпороговом состоянии на сенсорное возбуждение может быть высоко когерентным [König et al., 1996].
Возбуждение сверху уменьшает время ответа отобранных нейронов нижележащего уровня до миллисекундного диапазона. В результате популяции нейронов нижележащего уровня, участвующие в данном акте восприятия, на короткое время становятся детекторами совпадения. Они интенсивно отвечают на синхронное сенсорное возбуждение, вызванное стимулом, и слабо реагируют на сигналы, распределенные во времени [König et al., 1996].
Объемлющая характеристика посредством активации сверху отбирает популяции нейронов нижележащих уровней, которые могут отображать признаки данного объекта по прошлому опыту. Те признаки данного объекта в данном месте, которые действительно обнаружены рецепторами, отображаются когерентной разрядкой активированных популяций. Тем самым стимул отбирает ансамбль популяций, участвующих в данном акте восприятия, из совокупности, активированной сверху. Посредством итераций по всем иерархическим уровням сверху вниз и снизу вверх синхронизируется уникальный набор нейронных ансамблей. Этот набор ансамблей из детекторов совпадений формирует быстрый сенсорный путь, единственный и уникальный для каждого акта восприятия. Одновременное (параллельное во времени) функционирование нейронных ансамблей вдоль сенсорного пути составляет конвейерную систему, которая многократно увеличивает быстродействие восприятия.
Другим важным свойством системы обработки данных является то, что она ничего не вычисляет. Уже на первых уровнях обработки система восприятия порождает перцептивные гипотезы о содержании стимульного поля. Перцептивные гипотезы – это готовые решения, которые являются результатом эволюционного и прижизненного опыта и порождаются нейронными структурами соответствующих уровней обработки в ответ на специфическое сенсорное возбуждение. Иерархия объемлющих характеристик быстро уменьшает количество возможных сенсорных объектов при движении вверх по сенсорному пути. Единственность высшей объемлющей характеристики обеспечивает быструю сходимость процесса. Иерархия объемлющих характеристик – это алгоритм синтеза данных, отличный от интегрирования. Мозг отбирает и синтезирует данные, а не распознает объекты посредством решения вычислительных задач [В. Сергин, А. Сергин, 2019].
Иерархии объемлющих характеристик образуют разветвленное дерево: от множества рецепторов данной модальности (листьев дерева) на вышележащие уровни обработки (ветви и ствол). Сходящиеся ветви на средних уровнях обработки формируют специализированные объемлющие характеристики, которые передаются вверх и формируют субмодальности разных специфичностей. Иерархии разных сенсорных и моторных модальностей взаимодействуют на всех уровнях обработки посредством взаимных и обратных связей. Высшие объемлющие характеристики охватывают весь спектр сигналов кроны дерева и порождают образы и сцены.
Взаимодействующие иерархии объемлющих характеристик – это мощный ассоциативный аппарат, обладающий в силу многосвязности сенсорных и моторных путей практически неограниченными возможностями выделения значимых ассоциаций и обобщений. Ассоциативные процессы реализуются посредством взаимодействия нейронных ансамблей, что обеспечивает статистически устойчивую повторяемость ассоциаций. Но в разных ситуациях (контекстах) один и тот же стимул может вызывать активность различных ансамблей нейронов и порождать неожиданный образ или чувство.
Ассоциативный аппарат, возникнув вместе с системой восприятия и моторной системой организмов, породил принципиально новые возможности мозга. Многомерный и многоуровневый ассоциативный аппарат создал широкие возможности формирования сенсорных, моторных и когнитивных объектов, которые не являются результатом непосредственного сенсорного или моторного опыта. Эти виртуальные объекты, порождаемые комбинаторной активностью мозга, доступны опытной проверке, как и любые перцептивные или моторные гипотезы. Способности мозга порождать виртуальные (возможные) объекты, их свойства и отношения лежат в основе воображения, предвидения и прогноза. Ассоциативный аппарат генерирует множество перцептивных, моторных и когнитивных гипотез, которые неизмеримо расширяют наши способности к восприятию и новым формам поведения.
Архитектура системы неосознаваемой обработки данных – это архитектура основного процессора головного мозга [В. Сергин, А. Сергин, 2019]. В то же время – это архитектура иерархической долговременной памяти. Архитектура в виде дерева иерархий объемлющих характеристик дает адреса любых данных: по качественной специфичности (модальности), степени обобщения или абстракции (уровня обработки), значимости (частоты востребования), актуальности (соответствия текущей задаче) и т.п.
СОЗНАТЕЛЬНОЕ ВОСПРИЯТИЕ
В каких областях коры головного мозга происходит осознание сенсорных данных? Согласно гипотезе автоотождествления, осознание осуществляется посредством автономных обратных связей. Поэтому осознание любой сенсорной характеристики может происходить только в той области коры, которая отображает данное сенсорное качество. Например, осознание цвета должно происходить в основном в зрительной области V4, специализация которой связана с отображением цвета, а осознание движения – преимущественно в специализированной области MT.
Данные первичных зрительных областей коры V1 и V2 также должны осознаваться. Только в этом случае возможно рассматривание деталей зрительного поля с высоким пространственным разрешением. Известно, что больные с пораженной престриарной корой и сохранной областью V1 способны идентифицировать отдельные линии и простые контуры и могут аккуратно их копировать. Это значит, что сенсорные признаки не только отображаются, но и осознаются посредством механизмов зрительной области V1. Но больные не способны воспринимать совокупность линий как единое целое, например, как дом или собор [Zeki, 1993]. Осознание целостных образов возможно только посредством процессов автоотождествления в престриарной и ассоциативной коре, в которых эти образы формируются, и которые оказались разрушенными.
Осознание осуществляется в тех же областях коры, в которых происходит неосознаваемое восприятие и формирование моторного ответа. Поэтому автономные обратные связи должны существовать в проекционных и специализированных областях коры всех сенсорных модальностей, а также в ассоциативных областях, включая височную, теменную и лобную доли. Во всех случаях осознается то содержание, которое соответствует функциям данной области коры.
Адаптивное значение имеет целостный образ, а не его отдельные сенсорные признаки. Поэтому первичное осознание должно происходить в ассоциативной коре, отображающей объемлющую характеристику объекта. С этого уровня обработки возбуждение может передаваться по каскаду обратных связей вниз, и в следующий момент времени могут осознаваться среднемасштабные сенсорные характеристики. В последнюю очередь должны осознаваться мелкомасштабные детали и всякого рода подробности. Экспериментально установлено, что во время фиксации взора зрительный образ обрабатывается последовательно, по этапам, с выделением сначала общих характеристик, потом все более мелкомасштабных и детальных [Хофман, 1986; Барабанщиков, 2000]. Из повседневного опыта известно: то, что мы видим мельком, мы видим в общих чертах. Необходимо некоторое, хотя и небольшое, время, чтобы увидеть подробности.
Осознание высшей объемлющей характеристики на первом этапе восприятия объясняет нашу способность схватывать осмысленное целое еще до осознания составляющих признаков. Это и есть “гештальт” – явление, породившее в начале прошлого века гештальтпсихологию.
Гештальтом принято называть целостное одномоментное восприятие объекта без расчленения его на части, которое противопоставляется восприятию по частям с последующим синтезом целого. Целостность рассматривается как интегральность, не разделенная на отдельные компоненты. В соответствии с этими воззрениями выделяют два разных процесса узнавания: целостный – по интегральным образам, что соответствует гештальтпсихологии, и признаковый – по набору элементов, признаков, деталей, что соответствует психофизиологии [Шамис, 2005].
Согласно модели объемлющих характеристик, восприятие осуществляется посредством иерархии сенсорных признаков и характеристик объекта, в полном соответствии с психофизиологической теорией. Осознание (распознавание) объекта начинается с формирования его высшей объемлющей характеристики. Объемлющая характеристика порождает возбуждение сверху и акцентирует отображение тех сенсорных признаков и характеристик нижележащих уровней, которые принадлежат данному объекту. В полном соответствии с гештальтпсихологией целое предшествует своим частям.
Таким образом снимаются давние противоречия между психофизиологией и гештальтпсихологией. Психофизиология описывает процессы восприятия, гештальтпсихология – процессы осознания (распознавания). Обе совокупности процессов протекают одновременно, взаимодействуют между собой на каждом уровне обработки и завершаются распознаванием сенсорного объекта со всеми его деталями и специфическими особенностями.
Сенсорный объект может отображаться и после его исчезновения, если циклические процессы автоотождествления поддерживаются неспецифической активирующей системой мозга на протяжении хотя бы нескольких циклов. Сохранение специфических паттернов нейронной активности, отображающих стимул на протяжении нескольких десятков или сотен миллисекунд после его исчезновения, может формировать ультракратковременную (иконическую, эхоическую) сенсорную память.
Экспериментально установлено, что ультракратковременная сенсорная память существует в самом деле. Сенсорная память характеризуется большой емкостью и малым сроком хранения. Например, зрительные образы на матрицах размерностью 32 × 32 запоминаются почти без ошибок на срок около 100 мс [Phillips, 1983]. Принято считать, что зрительная (иконическая) память позволяет фиксировать всю поступающую информацию, но только на доли секунды [Клацки, 1978].
Тактовая частота перцептивного сознания как системы обработки данных составляет 10–100 Гц. Большая емкость сенсорной памяти и повторная фиксация поступающей информации на каждом цикле автоотождествления обеспечивают полноту, детальность и точность образов сознательного восприятия. Краткий срок хранения обусловливает быструю деградацию не подтвержденной сенсорным опытом информации и готовность к восприятию новых сенсорных данных.
Не все сенсорные данные требуют точного и детального осознания. Если некоторые данные не представляют актуальной ценности, процесс может быть прерван на стадии осознания общего (качественного) характера сенсорного объекта или только факта его наличия. Это предосознание, возможно, и является первичным ощущением. Ощущение может быть ранней стадией осознания, на которой отображаются только качественные свойства стимула. Некоторые стимулы, например, слишком слабые или быстро изменяющиеся, физически недоступны детальному осознанию и могут только ощущаться. Данные многих рецепторных органов, например, равновесия, температуры или давления, а также данные о внутренней среде организма, в противоположность зрительной или слуховой информации, всегда отображаются в качественной форме. В результате подавляющее большинство стимулов окружающей среды и внутреннего мира субъекта только ощущается. Но именно этот многомодальный и многообразный поток предосознаваемых стимулов и создает чувство жизни, ощущение бытия в окружающем мире.
Осознание является формой вторичной обработки, назначение которой состоит в явном представлении актуально значимых данных. Универсальная система явного представления данных достигает наибольшей эффективности в зрительном восприятии, порождая психический феномен вѝдения. Вѝдение – это явное внутреннее представление сенсорного объекта, которое порождает понимание, поскольку является его субъективной интерпретацией. Осознание придает новое качество процессу восприятия в психическом плане, позволяя не только смотреть и адекватно реагировать, но смотреть и видеть, не только слушать и действовать, но слушать и слышать. Осознание вовлекает сенсорные данные в процессы рассудочного и образного мышления и в психическую активность субъекта.
Выполненные исследования [Сергин, 2016; Sergin, 2017] позволяют утверждать, что осознание – это процесс представления внутренних данных в явной форме. Явное символьное представление сенсорных данных порождает сенсорное осознание. Явное символьное представление результатов операционной активности мозга порождает мысль. Если зрительное осознание – это вѝдение внешних событий, то мысль – это вѝдение результатов операционной активности мозга.
Мысль изначально имеет характер внутреннего вѝдения (“внутренний взор”). Возникновение мысли обычно означает понимание чего-либо. В русском языке слово “видеть” часто используется в значении “понимать”. В английском языке слово “see” означает и “видеть”, и “понимать”. В древней Греции, на заре математики, доказательством утверждения (теоремы) считалось соответствующее геометрическое построение: “теперь смотри”.
СЕНСОРНО-МОТОРНОЕ ПОВТОРЕНИЕ И МЫШЛЕНИЕ
Мы способны осознавать результаты операционной активности мозга и можем произвольно управлять этой активностью, т.е. думать. Как нам удается управлять операционной активностью мозга и какие нейрофизиологические механизмы лежат в основе наших способностей к мышлению?
Ключевая идея, позволяющая ответить на этот вопрос, состоит в том, что явное символьное представление внутренних данных делает их относительно обособленными объектами. В результате возникает возможность управлять ими так же, как мы управляем объектами. Примером произвольного управления может служить вербальное повторение: человек произносит слово, слышит его, запоминает, произносит вновь и т.д. Это циклический вербальный процесс сенсорно-моторного повторения. Слова не обязательно произносить вслух, человек может повторять их про себя, что составляет процесс внутреннего сенсорно-моторного повторения. Повторение – управляемый процесс, его можно прерывать и возобновлять вновь, его содержание можно произвольно изменять.
Передача информации в мозге имеет унитарный характер. Информация передается от одной области коры к другой посредством паттернов нейронной активности. Для моторной системы не только движения, но и слова, образы или символы – это всего лишь специфические паттерны нейронной активности. Поэтому программный аппарат моторной системы, управляющий двигательными актами, точно так же может управлять словами, символами или образами.
Вербальная система имеет наиболее мощный аппарат сенсорно-моторного повторения, который включает височную речевую зону коры (область Вернике), осуществляющую рецептивный подбор слов, и лобную область формирования речи (область Брока) [Лурия, 2003; Баарс, Гейдж, ч. 1, 2014; Standring, 2005]. Когда мы хотим высказать некоторую мысль, мы еще не знаем, какие слова будем произносить. Мысль – инвариантная характеристика высказывания, и может быть выражена различными комбинациями слов. Мысль как объемлющая характеристика высказывания порождает симультанную семантическую схему, которая выражает отношения между словами, соответствующие прошлому опыту. Эта симультанная схема передается в область Брока, которая порождает последовательность паттернов возбуждения, отображающих временную последовательность слов. Последовательные паттерны возбуждения передаются вниз, в речедвигательные области коры.
Вместе с тем эти последовательные паттерны возбуждения могут одновременно передаваться и в область Вернике (височно-теменно-затылочную область), которая порождает симультанную семантическую схему, соответствующую программируемой последовательности слов. Симультанные семантические схемы, порождаемые мыслью и программируемые последовательностью слов, могут сравниваться, обнаруживая, насколько точно временная последовательность слов выражает мысль. Если фактически порождаемые слова или их расстановка не вполне соответствуют мысли, возбуждение сверху может повторяться, и мысль получает иное, более точное выражение.
Итерационный процесс выражения мысли – это и есть внутренняя речь, которая реализуется посредством паттернов нейронной активности и протекает очень быстро. Процесс высказывания мысли (физическое воплощение слов) требует включения мышечного аппарата речедвигательной системы, что занимает значительно больше времени. Поэтому высказывание начинается с запаздыванием и может содержать такую расстановку слов, которая является финальным результатом нескольких быстрых итераций процесса выражения мысли. Слова возникают как бы сами собой, и нас иногда даже удивляет, насколько метко и точно они выражают мысль.
Зрительное повторение представляет собой циклический процесс сенсорно-моторного воспроизведения зрительных образов. Многие зрительные образы, их признаки, свойства и операции над ними выражаются словами и, следовательно, отображаются паттернами речевой моторной системы. Чтобы визуализировать образ, достаточно назвать его про себя или вслух. Называя определенное слово, человек активирует речевую моторную систему, паттерны которой передаются в ассоциативную зрительную кору. Специфическая реакция долговременной зрительной памяти порождает соответствующий образ.
Процедуры преобразования фигур (декомпозиция, сдвиги, повороты и т.п.) также управляются паттернами речевой моторной системы. Эти процессы образного мышления осуществляются посредством механизмов зрительно-вербального повторения. Б. Кохен [Cohen, 1986], анализируя обширный экспериментальный материал, находит, что внутренняя речь играет основную роль в вызове и управлении образами разных модальностей.
Отображение образов разных модальностей паттернами речевой моторной системы позволяет управлять ими посредством программного аппарата. Обширная и хорошо структурированная база данных речевой системы и ее непосредственная управляемость программным аппаратом порождают универсальность процедур вызова и управления. Поэтому любые виды сознательной активности мозга, например, формирование мысленных образов или логические рассуждения, должны порождать специфическую активность в левой речевой височной области. А.М. Иваницкий [2009], обозревая опыт обширных экспериментальных исследований, приходит к выводу, что возникновение очага возбуждения в левой височной области происходит при выполнении любых видов мыслительных задач.
Рассудочное мышление непосредственно связано с отображением информации паттернами нейронной активности речевой моторной системы. Процедура повторения может обеспечивать последовательность и непрерывность процесса рассуждений, его связанность и однозначность. В этих свойствах рассудочного мышления легко просматриваются характерные черты моторной системы, функционирование которой осуществляется посредством выбора единственной из множества потенциальных команд, их последовательного выполнения и строгой связанности последовательных действий. Программный аппарат моторной системы является источником рациональности мышления. Рациональность порождается тем, что действия моторной системы причинно об-условлены, здесь нет места фикциям. Эта причинная обусловленность воспроизводима посредством аппарата сенсорно-моторного повторения.
Программный аппарат моторной системы отбирает небольшие координированные наборы действий из громадного множества потенциально возможных. Строгая последовательность небольших наборов координированных действий позволяет осуществлять целенаправленное движение. Программный аппарат, формирующий связанную однозначную последовательность двигательных актов, точно так же формирует связанную однозначную последовательность слов или символов. Мы можем управлять последовательностью мыслей так же, как мы управляем последовательностью движений. Схематизм, жесткая однозначность и логичность рассудочного мышления порождаются программным аппаратом моторной системы.
Аппарат программирования трехмерного движения является одним из наиболее фундаментальных достижений биологической эволюции. Естественный отбор выметал из программного аппарата все, что не отвечало логике реализации двигательных актов, временной последовательности и пространственной координации движений и другим рациональным критериям целенаправленного поведения. Формальная логика, похоже, всего лишь малая выборка из арсенала средств программного аппарата моторной системы. Источником рациональности и логичности рассудочного мышления является моторная система. Рассудочное мышление оказывается механистичным по своей природе. Это накладывает ограничения на его творческие возможности, а также объясняет ту легкость, с которой дискурсивные процедуры реализуются на компьютере.
ОПЕРАЦИОННОЕ СОЗНАНИЕ
Систему оперирования данными, представленными в явной форме, которая осуществляет такие высшие функции, как рассудочное и образное мышление, будем называть операционным сознанием [Сергин, 1994; Sergin, 1994, 1999]. Механизм сенсорно-моторного повторения является базовым средством операционного сознания. Экспериментальные данные показывают, что типичная частота вербального повторения составляет 3–6 Гц, частота зрительного повторения несколько ниже [Клацки, 1978]. Частота повторения является тактовой частотой операционного сознания как системы обработки данных. Следовательно, процессы рассудочного и образного мышления реализуются посредством циклических механизмов, работающих с тактовой частотой порядка нескольких герц, что приблизительно соответствует тета-ритму. Заметим для сравнения, что тактовая частота аппарата осознания на порядок выше и составляет 30–70 Гц, а частота неосознаваемой обработки, вероятно, составляет сотни герц.
Процедура сенсорно-моторного повторения доступна обозрению и управляема. Физиологически это возможно потому, что повторение является низкочастотным процессом, и его содержание доступно обозрению с помощью высокочастотного аппарата автоотождествления (осознания), который надстроен над контуром повторения (рис. 4). Череда мысленных образов, слов или символов управляема посредством программного аппарата моторной системы и доступна обозрению посредством аппарата автоотождествления. Взаимодействующие механизмы сенсорно-моторного повторения и автоотождествления позволяют нам формировать образы, сцены и диалоги, наблюдать и изменять их, порождая подвижный и управляемый мир сознательного опыта [Сергин, 1994; Sergin, 1994; 1999].
Рис. 4.
Схема взаимодействия процессов сенсорно-моторного повторения и осознания данных.
Fig. 4. Scheme of interaction of sensory-motor rehearsal processes and data awareness.
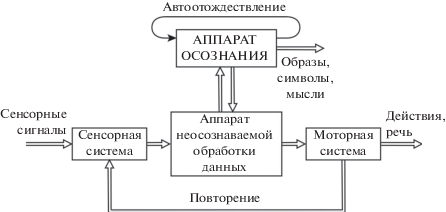
Время хранения данных в оперативной (кратковременной) памяти определяется временем поддержания циклических процессов повторения (например, когда мы повторяем про себя номер телефона). Объем оперативной памяти невелик и составляет приблизительно от 4 до 7 условных единиц [Клацки, 1978; Баарс, Гейдж, ч. 2, 2014; Halford, Cowan, 2007; Cowan, 2008]. Единицами хранения данных в оперативной памяти могут быть числа, буквы, слоги, слова, символы, образы и что угодно еще. В процессе повторения мы можем исключать некоторые элементы или добавлять другие, менять местами элементы выборки, объединять некоторые элементы в группы или разделять их.
Повторение нескольких элементов в различных сочетаниях, их сопоставление, манипулирование ими – это и есть процесс мышления. В этом процессе некоторые сочетания вызывают ассоциативно связанные данные из долговременной памяти и включают их в процесс повторения, в то время как другие данные исключаются из цикла повторения. Воспроизводимость и наблюдаемость позволяют отбирать причинно-следственные отношения. Способность устанавливать причинно-следственные связи является ключевым свойством человеческого разума. Знание причинно-следственных связей обеспечивает человеку важные адаптивные преимущества, которые могли играть решающую роль в процессе его эволюции.
Операционное сознание осуществляет такие высшие функции, как рассудочное и образное мышление, воображение и рефлексию. Операционное сознание работает во времени последовательно, шаг за шагом. Система последовательного действия является универсальным средством реализации логических высказываний и рассуждений, что определяет ее широкие операционные возможности. Однако ее производительность ограничена низкой тактовой частотой (3–6 Гц) и небольшим объемом оперативной памяти. Универсальность, низкая производительность и небольшая память являются главными характеристиками операционного сознания как системы обработки данных. Отсюда следует, что сознанию доступно решение практически любых задач, не требующих большого объема вычислений.
Мы имитируем образы объектов, сцен и событий или конструируем их произвольно, используя базу данных долговременной памяти. Если образы отображают реальности окружающей среды, имеет место мысленное рассмотрение внешнего мира. Если образы отображают собственные свойства субъекта, имеет место рассмотрение своего внутреннего мира (самонаблюдение). Самонаблюдение позволяет создавать описания самих себя, что порождает самосознание.
Если субъект мысленно имитирует сцены или события, выражающие его взаимодействия с другим субъектом, например, диалог, то функционирование такой модели и ее анализ представляют собой еще одну область рефлексии. Управляемые изменения модели позволяют субъекту имитировать свое поведение, проигрывать варианты поступков и оценивать их результаты, не совершая самих действий. Этот аппарат операционного сознания открывает возможности планировать свою деятельность, прогнозировать ее результаты и осуществлять ретроспективный анализ.
Оперативная память содержит только те данные, которые человек в данный момент осознает. Сохранение данных в оперативной памяти – это психический процесс, который физиологически выражается в поддержании взаимно скоординированных специфических паттернов нейронной активности в тех сенсорных, ассоциативных и моторных областях коры, которые составляют замкнутый контур передачи возбуждения. Тогда топографическая локализация и специфичность паттернов нейронной активности коры должны соответствовать когнитивному содержанию оперативной памяти. Например, удержание в памяти зрительного образа должно вызывать избирательную активацию соответствующих областей зрительной, теменно-височной и лобной коры, временно связанных этой специфической активностью в замкнутый контур сенсорно-моторного повторения. Удержание в памяти слов или чисел должно вызывать активацию левой височной коры, лобной и моторной коры, ответственных за восприятие и порождение речи.
Многочисленные экспериментальные данные подтверждают специфическую связь когнитивного содержания мысленных операций с топографией функционально активных областей коры. Эта связь оказывается достаточно хорошо выраженной и устойчивой, так что по рисунку электроэнцефалограммы удается определять тип задачи, которую решает испытуемый [Г. Иваницкий и др., 2007, 2009; Таротин и др., 2017].
ПРИРОДА ИНТУИЦИИ
Кора больших полушарий головного мозга имеет модульную организацию [Hubel, Wiesel, 1974; 1977; Szentagothai, 1975; Creutzfeldt, 1977; Mountcastle, 1978; 1998; Батуев, 1984; Савельев, 2010; и др.]. Неосознаваемая обработка данных осуществляется массированно-параллельной системой, которая включает миллионы параллельно работающих модулей (мини-колонок) новой коры. Параллельная работа модулей распределенной системы сопровождается конвейерными (параллельными во времени) процессами обработки данных в модулях, составляющих последовательные структуры. Поэтому мозг в целом является параллельно-параллельной системой обработки данных. Частота обработки определяется временем переключения нейрона и может составлять сотни герц. Архитектуру системы неосознаваемой обработки данных формирует сходящийся многосвязный пучок иерархий объемлющих характеристик, который порождает практически неограниченные ассоциативные возможности мозга.
В областях коры головного мозга, охваченных циклическими процессами автоотождествления и сенсорно-моторного повторения, конвейерные процессы замещаются дискретной последовательной обработкой данных, и сознание можно охарактеризовать как систему параллельно-последовательного действия. Такая архитектура кардинально уменьшает производительность сознания как системы обработки данных. Ее тактовая частота определяется частотой сенсорно-моторного повторения, что снижает скорость обработки еще на 2 порядка.
Выборка информации из долговременной памяти мозга осуществляется в соответствии с целью и содержанием конкретной задачи. Минимальное время выборки определяется временем переключения нейрона и составляет величину порядка 1 мс. Минимальное время выборки из памяти сознания определяется временем цикла автоотождествления и составляет величину порядка нескольких десятков миллисекунд. То есть, в отличие от компьютеров, оперативная (кратковременная) память мозга является более медленной, чем долговременная. Результаты такой организации мозга проявляются в том, что неосознаваемые действия происходят много быстрее, чем те же действия, управляемые сознанием.
Таким образом, в мозге воплощены две взаимодействующие, но относительно обособленные и функционально различные системы обработки данных: система неосознаваемой обработки и операционное сознание. Объем памяти и быстродействие системы неосознаваемой обработки данных на много порядков больше, чем аналогичные характеристики операционного сознания.
Из этой модели с необходимостью следует вполне определенная схема творческого процесса. В сознании может формироваться лишь постановка задачи (цели, вопросы, проблемы). Процесс решения осуществляется системой неосознаваемой обработки, которая обладает обширной базой данных и высокой производительностью. Затем полученное решение выдается в область сознания. В этом и состоит природа интуиции [Sergin, 1991; 1992]. В область сознания может выдаваться лишь окончательное решение проблемы. Промежуточные операции отыскания решения не могут передаваться в сознание из-за его малой информационной емкости. Поэтому мы не можем осознавать аргументацию (или процедуры), приведшие мозг к интуитивному решению.
Интуиция характеризуется тем, что позволяет устанавливать новые связи и отношения в условиях видимого недостатка данных. В работе П.В. Симонова [1987] указывается, что принципиальной особенностью проявления интуиции является дефицит информации, необходимой и достаточной для логически безупречного заключения. Другие важные свойства интуиции состоят в том, что она непредсказуема и в то же время неслучайна.
Перечисленные свойства интуиции естественным образом вытекают из предлагаемой модели. Действительно, интуитивное решение есть функция системы неосознаваемой обработки данных, поэтому осознаваемая информация может быть сколь угодно недостаточной. Далее, интуитивное решение непредсказуемо для сознания так же, как решение суперкомпьютера непредсказуемо для персонального компьютера. В то же время интуитивное решение неслучайно, оно определяется объемом и специфичностью знаний, накопленных долговременной памятью индивида, операционной структурой его мозга, сознательно сформулированной постановкой задачи (целью исследования) и интенсивностью намерения решить проблему.
Таким образом, определение цели исследования, сбор данных и формулировка задачи являются сознательными процессами. Затем следует интуитивное решение проблемы. И лишь после того, как ответ (не обязательно правильный) стал известен, производится его аргументированное доказательство (или опровержение).
В науке широко распространено представление о том, что правильно поставить задачу – значит наполовину решить ее. Постановка задачи, будучи преимущественно сознательным процессом, содержит важные эвристические компоненты, что свидетельствует об участии неосознаваемой системы в этой работе. В правильно поставленной задаче содержится неявное знание характера ее решения, что свидетельствует об уже выполненной (хотя и неосознанной) предварительной работе.
Интуиция имеет характер непосредственного вѝдения. Это позволяет предположить, что возможна глубокая аналогия процессов восприятия и интуитивного мышления. Иерархия характеристик, объемлющая сенсорный опыт, порождает перцептивные гипотезы [В. Сергин, А. Сергин, 2019]. Иерархия характеристик, объемлющая опыт рассудочной и ассоциативной работы мозга, может порождать когнитивные гипотезы. Формирование иерархии объемлющих характеристик является неосознаваемым процессом, осознается лишь конечный результат, его высшая объемлющая характеристика. Поэтому результаты рассудочной и ассоциативной работы мозга могут быть неожиданными для размышляющего субъекта. Внезапное решение проблемы, на которую затрачено исключительно много сил и времени, – это и есть озарение (инсайт).
Мы способны осознавать лишь окончательные результаты неосознаваемой работы мозга, такие как понимание ситуации или решение проблемы. Промежуточные результаты, которые привели мозг к решению проблемы, непосредственно не осознаются. Мы можем знать решение проблемы, но мы не знаем, как это решение получено. Эта будто бы парадоксальная ситуация не более загадочна, чем любой случай восприятия. Мы можем обнаружить сенсорный объект, но мы не знаем, как этот объект обнаружен (хотя теоретические предположения на этот счет имеются).
ГЛОБАЛЬНАЯ МОДЕЛЬ СОЗНАНИЯ
Подводя итоги предыдущих разделов, глобальную модель сознания можно представить схемой, показанной на рис. 5. Большими прямоугольниками выделены три относительно обособленные системы мозга. Эти системы могут работать одновременно на отчасти совпадающих нейронных структурах. Но они четко разделяются различными частотными диапазонами, в которых формируются их функции.
Рис. 5.
Блок-схема глобальной модели сознания.
Fig. 5. The block diagram of the global model of consciousness.
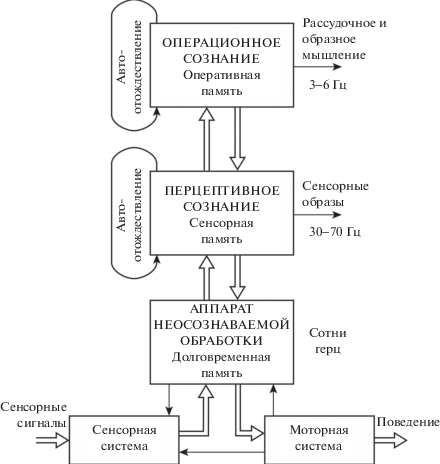
Операционное сознание является носителем постоянно обновляющейся модели внешнего и внутреннего мира субъекта. Эта модель глобальна в том смысле, что охватывает весь мир до границ чувственного и духовного опыта. Причем все ее компоненты определенным образом связаны, образуя единое непротиворечивое целое. Но из-за малой информационной емкости сознания модель имеет мало деталей и в этом смысле минимальна. Модель сознания весьма схематична, она верна лишь в общих чертах, это имитационная модель внешнего и внутреннего мира. Но именно потому, что модель невелика, с ней легко работать, проигрывая различные ситуации и сопоставляя альтернативы для выбора целесообразного способа действия.
Эта модель перестраивается. В зависимости от содержания проблемы те или иные явления или объекты воссоздаются с большей подробностью. Анализ отдельного явления может быть чрезвычайно детальным, но лишь за счет исключения из рассмотрения всего остального. Во всех случаях объем модели сознания остается небольшим, что обеспечивает высокую подвижность и оперативность ее использования.
Построенная модель сознания дает морфологический критерий, который позволяет четко определить области, известные по психиатрическим терминам: бессознательное, неосознаваемое и осознаваемое. Нейронные структуры, не имеющие автономных обратных связей, не могут порождать осознание ни при каких условиях. Их совокупность составляет область бессознательного.
Нейронные структуры, имеющие автономные обратные связи, физиологически могут порождать осознание. Но процессы осознания требуют больших затрат доступной энергии и снижают производительность нейронных структур как систем обработки. Все это ограничивает повторяемость процессов осознания на множестве таких структур. Совокупность нейронных структур с автономными обратными связями, систематически не вовлекаемых в процессы осознания, составляет область неосознаваемого. Совокупность нейронных структур, имеющих автономные обратные связи и систематически вовлекаемых в процессы осознания, составляет область осознаваемого.
Область осознания – это множество нейронных структур коры больших полушарий, охваченных циклическими процессами авто-отождествления. Циклические процессы создают относительную обособленность сознания от остальной психики и формируют его подвижную организацию.
С точки зрения функционирования мозга как системы обработки данных, состояние сознания достигается дорогой ценой, за счет потери производительности и высоких затрат доступной энергии. Поэтому естественно предположить, что сознание постоянно реализуется лишь на небольшом числе структур головного мозга и быстро расширяется в процессах активного восприятия или интенсивной рассудочной деятельности. Расширение области сознания в новой коре может происходить только за счет системы неосознаваемой обработки данных. Следовательно, одни и те же ресурсы используются поочередно сознанием и системой неосознаваемой обработки. Конкуренция за ресурсы мозга наблюдается и непосредственно: каждому знакомо побуждение отключиться от внешней информации, чтобы плодотворно поразмышлять.
Область сознания сокращается до минимума во время сна (если не принимать во внимание краткие эпизоды сновидений), и мозг освобождается для более производительной работы, чем, по-видимому, и определяется физиологическая необходимость сна [Sergin, 1991]. При этом высвобождаются громадные ресурсы обработки, которые могут использоваться для решения задач, поставленных ранее в процессе взаимодействия сознания с системой неосознаваемой обработки. Это подтверждается хорошо известными фактами, когда решение трудных проблем приходит в утреннюю пору без видимых усилий, как бы самопроизвольно: система неосознаваемой обработки транслирует результаты ночной работы. Пословица “утро вечера мудренее” отражает богатейший экспериментальный материал, накопленный в результате разносторонней человеческой деятельности.
Наличие в мозге двух функционально различных типов мышления порождает двойственность в мироощущении человека. Эта двойственность наблюдается непосредственно, когда логические рассуждения подсказывают нам один способ действия, а интуиция побуждает к другому. Такие явления бывают столь часто, что их не имеет смысла даже подробно обсуждать. Приведем только один нетривиальный пример из рассказа А.И. Куприна “Лунной ночью”.
“Случалось вам видеть во сне, будто вы сдаете трудный экзамен? Вам задают вопрос и вы на него никак не можете ответить. Вы усиленно думаете, ломаете голову, но ответ, как нарочно, не идет на ум. Тогда учитель обращается к одному из ваших товарищей, тот отвечает самым правильным и блестящим образом, и вам становится стыдно за свое незнание.
…Во сне на экзамене отвечает ваш товарищ. Но ведь товарища-то на самом деле нет, отвечаете вы же и вы же удивляетесь тому, что говорите. Видите, какая двойственность?”
Здесь ответ системы неосознаваемой обработки трактуется сознанием, в соответствии со сценарием экзамена, как ответ товарища. В бодрствующем состоянии ответ системы неосознаваемой обработки иногда бывает столь неожиданным и радикальным, что трактуется сознанием как подсказка со стороны или внушение свыше. Так, например, Сократ был уверен, что его идеи были ему продиктованы его личным демоном, а Помпилий часто “консультировался” у нимфы по имени Этерия [Адамар, 1970]. Таинственные голоса Жанны д’Арк, откровения пророков, небесные послания основателей сект и верований имеют вполне материальную природу. Их информационным источником является система неосознаваемой обработки данных их собственного мозга.
Массированно-параллельная организация и ассоциативная структура системы неосознаваемой обработки данных определяют ее фундаментальные свойства. Операционный аппарат системы неосознаваемой обработки основан на механизмах отбора ассоциативных связей. Это универсальный аппарат обработки данных, эффективность которого неизмеримо превышает возможности логических построений.
Система неосознаваемой обработки осуществляет отбор и синтез данных посредством иерархий объемлющих характеристик, сходящихся от уровня рецепторов до уровня формирования целостных объектов в высшей ассоциативной коре. Она охватывает все сенсорные модальности, двигательные структуры и когнитивную систему. Аппарат неосознаваемой обработки данных обращен вовне и осуществляет взаимодействие организма с окружающим миром. Но чем занят этот аппарат в ночное время, когда мы спим без сновидений? Только ли решением поставленных ранее интеллектуальных задач?
Можно предположить, что во время сна система неосознаваемой обработки данных обращена вовнутрь организма и работает совместно с автономной нервной системой. Система неосознаваемой обработки, обращенная вовнутрь, сохраняет свою архитектуру и основные свойства. Рецепторы внутренних органов обеспечивают детальные данные, объемлющие характеристики которых могут отображать состояния тканей и органов тела. Такой аппарат, совместно с автономной нервной системой, как кажется, может обнаруживать и восстанавливать повреждения и функциональные расстройства на всех уровнях соматической организации, от клеток и их ансамблей до органов и их систем.
Эта гипотетическая схема выглядит оптимистично, но существуют ли связи между автономной системой и системой неосознаваемой обработки данных? Автономная нервная система является эволюционно древним образованием. Именно в процессе формирования автономной нервной системы отрабатывались организация и структуры иерархических систем. И только в ходе дальнейшей эволюции эти приобретения могли переноситься на решение проблем взаимодействия организма с окружающей средой. Таким образом, иерархические структуры автономной нервной системы являются оригиналами, а структуры соматической нервной системы – их аналогами. Следовательно, связи между автономной системой и нарождающейся новой корой существовали изначально. Если это действительно так, то крепкий сон является критически важным условием здоровья и долголетия.
Заметим, что операционное сознание, – это аппарат программирования и, в этом качестве, оно может ориентировать взаимодействие системы неосознаваемой обработки данных и автономной системы на специфическую активность по восстановлению собственных органов и тканей, чем бы ни были они повреждены. Несгибаемое намерение выздороветь, подтвержденное специфической физической (моторной) активностью субъекта (например, приемом лекарств или бегом трусцой) может активировать такие средства автономной нервной системы, которые выходят далеко за горизонты современной фармакологии.
Обретение здоровья – это проблема, и как и всякая сложная проблема, она может решаться средствами системы неосознаваемой обработки данных. Операционное сознание как аппарат программирования формирует намерение и возможные пути восстановления здоровья, а система неосознаваемой обработки данных порождает физически реализуемые процессы выздоровления.
Выздоровление – это инсайт, который порождается системой неосознаваемой обработки и реализуется посредством ее взаимодействия с автономной нервной системой. Мы не будем углубляться далее в эту проблему, поскольку она может увести нас далеко от основной темы статьи. Заметим только, что проблема взаимодействия коры головного мозга с висцеральной системой обсуждается со времен К.М. Быкова [1954] и представлена современными теоретическими и экспериментальными исследованиями И.Н. Пигарева и соавт. (см. обзор в [Пигарев, 2013]).
ИЗМЕНЕННЫЕ СОСТОЯНИЯ СОЗНАНИЯ
В основе построенной модели сознания лежат нейрофизиологические механизмы автоотождествления и сенсорно-моторного повторения. Вместе с тем психика человека характеризуется чрезвычайно широким разнообразием феноменов сознательной деятельности, которые простираются от восприятия и мышления до таких пока еще загадочных явлений, как гипноз и медитация. Если окажется, что построенная модель позволяет объяснить психофизиологические механизмы столь различных явлений, не прибегая к дополнительным предположениям для каждого частного случая, то это станет важным свидетельством в пользу ее адекватности.
К интригующим свойствам человеческой психики относится гипноз. В состоянии глубокого гипноза субъект не способен не только осознавать ситуацию и свое поведение или критически воспринимать внешние сигналы, но не может адекватно ощущать даже физиологические воздействия. Например, в состоянии гипнотической анальгезии субъект не чувствует боли [Буль, 1975; Kihlstrom, Hoyt, 1988].
Многие явления гипнотических состояний можно объяснить частичным или полным прекращением процессов автоотождествления и сенсорно-моторного повторения. Если воздействия гипнотизера расстраивают циклические процессы автоотождествления, то осознание поступающей информации становится невозможным, хотя ее неосознаваемое восприятие происходит. Так, при гипнотической анальгезии субъект сообщает об отсутствии болевых ощущений, в то время как измерения кожно-гальванической реакции и частоты сердечных сокращений демонстрируют адекватную физиологическую реакцию на болевые воздействия [Kihlstrom, Hoyt, 1988].
Внушенная слепота, глухота или нечувствительность к боли должны сопровождаться прекращением высокочастотных синхронных колебаний в соответствующих специализированных областях сенсорной коры, что доступно прямой экспериментальной проверке.
Любые практические способы гипноза должны включать воздействия, которые снижают или расстраивают активацию процессов автоотождествления и сенсорно-моторного повторения. В результате сознание гипнотизируемого субъекта оказывается подавленным, что делает невозможным осознание ситуации и собственного поведения. В этих условиях указания гипнотизера могут восприниматься мозгом гипнотизируемого субъекта как программа, подобная той, что обычно поступает от собственного сознания и потому выполняется. Мозг человека, сознание которого подавлено, не способен рассуждать и критически оценивать внешние сообщения, поэтому он воспринимает инструкцию гипнотизера как достоверную информацию, что вызывает немедленную поведенческую реакцию. Гипнотизер управляет гипнотизируемым субъектом словесно. В этой форме человек получает программу и от собственного сознания, поскольку внутренняя речь является основным средством вызова представлений и образов разных модальностей. Только наличие универсального речевого канала передачи информации делает в принципе возможной ситуацию, когда сознание одного человека управляет сознанием другого.
Гипнотизер может посредством слова вызвать образ из долговременной памяти гипнотизируемого субъекта, активируя тем самым процессы автоотождествления в ограниченных областях коры. Происходит осознание, т.е. вторичная обработка информации, не существующей в первичных структурах восприятия. Субъект галлюцинирует, воспринимая объект, которого нет в окружающем мире. Вызов информации из долговременной памяти субъекта и включение ее в процесс автоотождествления (визуализация образа) осуществляются так же, как это происходит в процессе воображения, когда субъект посредством слова вызывает образ из собственной памяти. В гипнозе собственное сознание субъекта замещено сознанием гипнотизера, который произвольно вызывает образы, хранящиеся в долговременной памяти гипнотизируемого субъекта. Из этой теории следует, что гипнотизер, не владеющий языком гипнотизируемого субъекта, не может вызвать у него произвольные галлюцинации. Это предсказание доступно экспериментальной проверке.
Сознание обладает чертами искусственной системы, оно управляемо и относительно обособлено от остальной психики. Состояние гипноза характеризуется контролируемым сокращением области осознания до физиологически возможного минимума и избирательным поддержанием изолированного очага осознания. Теоретически возможно и обратное состояние – контролируемое расширение области осознания до физиологически возможного максимума. Такое состояние мозга известно как состояние медитации.
Медитация представляет собой практику использования аппарата осознания для проникновения в свой собственный внутренний мир [Sergin, 1992: 1999]. Обязательным условием состояния медитации является изоляция от внешних событий при поддержании высокого уровня бодрствования. В этом состоянии мозг целиком обращен внутрь себя и все его ресурсы направлены на восприятие внутренних сигналов. Другим важным условием медитации является глубокое расслабление моторной системы, мышечная релаксация, которая минимизирует поток сигналов от собственного тела. Глубокое расслабление моторной системы позволяет прервать процедуру сенсорно-моторного повторения, что прекращает внутренний диалог. Прекращение процедуры повторения означает прекращение потока мыслей и образов, что ведет к “пустоте” сознания. Активированный и свободный от интенсивной рутинной работы мозг обладает более высокой чувствительностью и низким уровнем шума. Поэтому слабые сигналы неосознаваемой обработки, которые в обычных условиях являются подпороговыми, могут вызывать активацию аппарата осознания. Возникает возможность осознания сигналов из глубин бессознательного, что позволяет реализовать предельные возможности аппарата осознания как системы внутреннего видения.
Техника медитации предъявляет к субъекту противоречивые требования. С одной стороны, необходима глубокая мышечная релаксация, прекращение внутреннего диалога и изоляция от внешних событий, что уменьшает приток сигналов и по этой причине снижает уровень активации мозга. С другой стороны, необходим высокий уровень активации мозга для восприятия слабых, обычно неосознаваемых сигналов. Высокий уровень активации достигается специальными методами. Например, известна техника дыхания [Кацуки, 1993], которая переводит организм в критическое состояние (вероятно, подобное гипоксии), что вызывает высокую неспецифическую активацию мозга. Эта изощренная и, в сущности, противоестественная техника дыхания трудна и опасна. Тем не менее она обеспечивает высокий уровень активации мозга и вызывает субъективные переживания необыкновенной ясности и внутренней силы.
Исходя из построенной модели сознания, можно предсказать некоторые необычные психические феномены, которые должны происходить, если выполняются обычные условия медитативной практики: обособленность от сигналов внешнего окружения и собственного тела; прекращение потока мыслей и образов; поддержание высокого уровня неспецифической активации мозга.
Частота процессов автоотождествления выражает интенсивность психической жизни субъекта и определяет темп субъективного времени. Это текущее время, переживаемое настоящее. Хотя аппарат осознания позволяет определять новизну и отличать предшествующее событие от последующего, он не содержит в себе средств крупномасштабного сравнения настоящего с прошлым и будущим. Такими средствами обладает рассудочное мышление, которое основано на процессах сенсорно-моторного повторения. В глубокой фазе медитации повторение отсутствует, и сопоставление настоящего с прошлым и будущим становится невозможным. Текущие события должны выпадать из общего хода времени, превращаясь в “непрерывное настоящее”. События могут возникать и завершаться, но, как ни странно, все это должно происходить в настоящем.
Пространственное мышление, в соответствии с характером конкретной задачи, формирует функциональное пространство и пространственные отношения, основанные на опыте восприятия внешнего мира и собственного тела. То есть мы воспринимаем и конструируем пространство. В глубокой фазе медитации нет ни восприятия из-за обособленности от внешних событий и ощущений собственного тела, ни воображения, основанного на процессах сенсорно-моторного повторения. Следовательно, не может быть и привычного пространства с его атрибутами. События происходят, но они, как это ни удивительно, должны протекать “вне пространства”.
Если в глубокой фазе медитации нет времени и пространства, то невозможно определение различий, основанных на временных и пространственных сравнениях. А поскольку нет рассуждений, то нет и операционных средств различения вообще. Это может порождать странный мир, где нет различий и где царит “единство всех вещей”.
Мы устанавливаем причины и следствия посредством рассуждений. Если нет рассудочного мышления, то в нашем ментальном мире нет и причинности. События происходят, но они протекают “сами по себе”. Отсутствие причинности определяет полную свободу и независимость явлений. Возникает необычный мир, в котором господствует “абсолютная независимость”.
Такого рода психические феномены в самом деле известны по отчетам практикующих медитацию, которые описаны в обширной литературе. “В абсолютном самадхи время исчезает полностью, то же самое происходит и с пространством. Исчезает также и причинность. Существует только ряд событий.” “… распадается привычный способ сознания. Внезапно мы постигаем мир, где нет противоположностей и мы переживаем единство всех вещей.” “… в каждое мгновение приходит и уходит только настоящее, струится непрерывный поток настоящего.” “… жизнь в настоящем абсолютно независима, это и есть наше истинное существование” [Кацуки, 1993].
В глубокой фазе медитации процесс повторения прерван, нет ни рассудочного мышления, ни рефлексии, “мысли исчезают”. Нет никаких средств самонаблюдения или контроля за процессом осознания. “Просмотр внутреннего мира осуществляется непосредственно и безотчетно. Внутренний мир постигается как таковой, без замечаний, сомнений, анализа или рассуждений. Постигается внутреннее неосознаваемое существование, глубинная природа субъекта” [Кацуки, 1993].
Уменьшение сигналов из внешнего мира и от собственного тела и прекращение процедуры повторения снижают поток данных на входе аппарата осознания. Физиологически это должно выражаться в уменьшении частоты циклических процессов автоотождествления ниже ее нижнечастотной границы (10–12 Гц) и замедлении субъективного времени. Экспериментально установлено, что медитация в самом деле характеризуется низкочастотным альфа- и тета-ритмом [West, 1982]. При этом в отчетах испытуемых сообщается о субъективных переживаниях замедления мыслей и времени, вплоть до их полной “остановки”.
Состояние медитации характеризуется расширением области осознания до физиологически возможного максимума. Сознание, распространившееся на сферу неосознаваемого, теряет обособленность от него, поскольку воплощается во всем мыслящем мозге. Это выражается в характерном для медитации ощущении слияния со всем миром или какими-то его аспектами или частями (запечатленными, конечно, в долговременной памяти). Такие состояния всегда сопровождаются замедлением собственного внутреннего времени. В высшей фазе медитации время и мысли замедляются настолько, что почти “останавливаются”, наступает “пустота” сознания, состояние “без мыслей и без времени” [Кацуки, 1993]. В этом пике сознание охватывает весь мыслящий мозг и почти прекращает свое собственное функционирование (частота автоотождествления стремится к нулю), что с необходимостью вызывает “остановку” субъективного времени и “пустоту”. Это особое состояние сознания, известное глубоко мистическими толкованиями, получает теперь нейрофизиологическое объяснение.
Частота циклических процессов автоотождествления характеризует темп субъективного времени, зависящий от интенсивности психической жизни субъекта. Это текущее время, переживаемое настоящее. Возвращаясь к частотной характеристике сознания, представленной на рис. 1, заметим, что режим 1 соответствует состоянию медитации [Сергин, 1994; Sergin, 1992, 1994]. Низкая частота циклических процессов соответствует низкой интенсивности психической жизни субъекта. Это объясняет известное по отчетам испытуемых ощущение глубокого отдыха, которое возникает после выхода из состояния медитации. Порождаемое медитацией состояние глубокого покоя и отдыха, как показывает практика, является эффективным способом целительства психики. Именно в этом качестве медитация широко известна с древнейших времен. Теперь, кроме того, становятся более прозрачными и нейрофизиологические причины терапевтического действия медитации.
Теоретически возможно также существование особого режима функционирования сознания (кривая 2 на рис. 1), который характеризуется высокими частотами циклических процессов [Сергин, 1994; Sergin, 1992]. Этот форсированный режим, требующий аномально высокой активности мозга, может включаться в моменты крайней опасности. Известны многочисленные случаи, описанные в литературе, когда за краткие мгновения перед внутренним взором субъекта проносится вся его жизнь. Описаны случаи, когда в условиях неотвратимой угрозы, вызывающей сверхактивацию мозга, внешние события вдруг начинают казаться замедленными, в то время как мысли протекают чрезвычайно быстро и с исключительной ясностью. На краткий миг открывается возможность сверхбыстрых и адекватных решений в опасной для жизни ситуации [Sergin, 1992].
ЗАКЛЮЧЕНИЕ
Явные события – это события вне воспринимающей системы, т.е. внешние события, что отличает их от неявных событий, происходящих внутри воспринимающей системы. Механизм автоотождествления позволяет представлять внутренние данные на входы нервной системы, что делает возможным их восприятие в явной форме.
Осознание – это процесс представления данных в явной форме. Явное символьное представление сенсорных данных порождает сенсорное осознание. Явное символьное представление результатов операционной активности мозга порождает мысль. Если зрительное осознание – это вѝдение внешних событий, то мысль – это вѝдение результатов операционной активности мозга.
Сознание – система оперирования данными, представленными в явной форме. Явное символьное представление внутренних данных позволяет управлять ими как обособленными объектами. Произвольное управление данными возможно посредством механизма сенсорно-моторного повторения. Процедура повторения может обеспечивать последовательность и непрерывность процесса рассуждений, его связанность и однозначность. Программный аппарат моторной системы, формирующий связанную однозначную последовательность двигательных актов, так же формирует связанную однозначную последовательность образов, слов или символов.
Cенсорно-моторное повторение является низкочастотным процессом, и его содержание доступно обозрению (осознанию) с помощью высокочастотного процесса автоотождествления. Череда мысленных образов, слов или символов управляема посредством программного аппарата моторной системы и доступна обозрению посредством процессов автоотождествления. Взаимодействующие процессы сенсорно-моторного повторения и автоотождествления позволяют нам формировать образы, сцены и диалоги, наблюдать и изменять их, создавая подвижный и управляемый мир сознательного опыта. Сознание, которое выглядит как непостижимая данность, оказывается хотя и глобальной, но все же постижимой системой оперирования данными, представленными в явной форме.
В мозге воплощены две взаимодействующие, но относительно обособленные и функционально различные системы обработки данных: операционное сознание и система неосознаваемой обработки. Объем памяти и быстродействие системы неосознаваемой обработки на много порядков больше, чем аналогичные характеристики операционного сознания. В процессе мышления сознание осуществляет постановку и логический анализ проблемы. Решение проблемы осуществляет система неосознаваемой обработки. Затем полученное решение выдается в область сознания. В этом и состоит природа интуиции (инсайта).
Построенная модель сознания позволяет с единых позиций анализировать широкий спектр феноменов сознательной активности мозга, обнаруживая единство природы, казалось бы, не связанных друг с другом психических явлений и универсальность нейронных механизмов их формирования. Модель доступна проверке: из нее следует ряд принципиальных утверждений о функционировании мозга, которые могут быть опровергнуты или подтверждены экспериментально.
Список литературы
Адамар Ж. Исследования психологии процесса изобретения в области математики. М.: Советское радио. 1970. 150 с.
Баарс Б., Гейдж Н. Мозг, познание, разум: введение в когнитивные нейронауки. Ч. 1. Пер. с английского. БИНОМ, Лаборатория знаний. М., 2014. 544 с.
Баарс Б., Гейдж Н. Мозг, познание, разум: введение в когнитивные нейронауки. Ч. 2. Пер. с английского. БИНОМ, Лаборатория знаний. М., 2014. 464 с.
Барабанщиков В.А. Системогенез чувственного восприятия. М.: Инст. практич. психол., 2000. 464 с.
Батуев А.С. Нейрофизиология коры головного мозга: Модульный принцип организации. Л.: ЛГУ, 1984. 216 с.
Буль П. Гипноз и внушение. М.: Медицина. 1975. 328 с.
Быков К.М. Избранные произведения. Т. 2. Кора головного мозга и внутренние органы. М.: Гос. изд-во мед. лит. 1954. 415 с.
Иваницкий А.М. Мозговые механизмы оценки сигналов. М.: Медицина, 1976. 264 с.
Иваницкий А.М., Стрелец Б.В., Корсаков М.А. Информационные процессы мозга и психическая деятельность. М.: Наука, 1984. 200 с.
Иваницкий А.М. Психическая деятельность и организация мозговых процессов. Вестник АМН СССР. 1987. 8: 14–23.
Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза. Журн. высш. нерв. деят. им. И.П. Павлова. 1996. 46 (2): 241–252.
Иваницкий А.М. Проблема сознания и физиология мозга. Проблема сознания в философии и науке. Под ред. Д.И. Дубровского. М.: “Канон” РООИ “Реабилитация”. 2009. С. 383–394.
Иваницкий Г.А., Наумов Р.А., Иваницкий А.М. Технология определения типа совершаемой в уме мыслительной операции по рисунку электроэнцефалограммы. Технология живых систем. 2007. 4 (5–6): 20–27.
Иваницкий Г.А., Наумов Р.А., Роик А.О., Иваницкий А.М. Как определить, чем занят мозг, по его электрическим потенциалам? Устойчивые паттерны ЭЭГ при выполнении когнитивных заданий. Вопросы искусственного интеллекта. 2009. 1: 93–102.
Кацуки С. Практика Дзэн. Киев: Пресса Украины, REFL-book. 1993. 336 с.
Клацки Р. Память человека. М.: Мир. 1978. 319 с.
Кропотов Ю.Д. Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия. Донецк; издатель Заславский, 2010. 506 с.
Лурия А.Р. Основы нейропсихологии. М.: Издательский центр “Академия”, 2003. С. 384.
Пигарев И.Н. Висцеральная теория сна. Журн. высш. нерв. деят. им. И.П. Павлова. 2013. 63 (1): 86–104.
Савельев С.В. Возникновение мозга человека. М.: Веди, 2010. 323 с.
Сергин В.Я. Перцептивное связывание сенсорных событий: гипотеза объемлющих характеристик. Журн. высш. нерв. деят. им. И.П. Павлова. 2002. 52 (6): 645–655.
Сергин В.Я. Сознание как система внутреннего видения. Журн. высш. нерв. деят. им. И.П. Павлова. 1994. 44 (4–5): 627–639.
Сергин В.Я. Психофизиологические механизмы осознания: гипотеза самоотождествления. Журн. высш. нерв. деят. им. И.П. Павлова. 1998. 48 (3): 558–571.
Сергин В.Я. Биологически правдоподобная модель зрительного восприятия: иерархия объемлющих сенсорных характеристик. Лекция на IX Всероссийской научно-технической конференции “Нейроинформатика-2007”. Лекции по нейроинформатике, ч. 2. Ред. Тюменцев Ю.В. М.: МИФИ, 2007, 77–120 с.
Сергин В.Я. Нейронные механизмы сознательного восприятия: гипотеза автоотождествления. Лекция на XI Всероссийской научно-технической конференции “Нейроинформатика-2009”. Лекции по нейроинформатике. Ред. Тюменцев Ю.В. М.: МИФИ, 2009а. 35–65 с.
Сергин В.Я. Психофизиологические механизмы восприятия: концепция объемлющих характеристик. Успехи физиологических наук. 2009б. 40 (4): 42–63.
Сергин В.Я. Автоотождествление паттернов нейронной активности как физиологический механизм осознания. Журн. высш. нерв. деят. им. И.П. Павлова. 2016. 66 (2): 1–20.
Сергин В.Я., Сергин А.В. Иерархическая модель восприятия без комбинаторного взрыва. Журн. высш. нерв. деят. им. И.П. Павлова. 2019. 69 (5): 633–658. DOI: 0044467719050113https://doi.org/10.1134/S
Силькис И.Г. Вклад синаптической пластичности в базальных ганглиях в обработку зрительной информации (гипотетический механизм). Журн. высш. нерв. деят. им. И.П. Павлова. 2006. 56 (6): 742–756.
Силькис И.Г. Возможный механизм участия цепей кора–базальные ганглии–таламус–кора в восприятии времени. Успехи физиологических наук. 2011. 42 (2): 41–56.
Силькис И.Г. Унифицированный нейронный механизм влияний внимания, эмоций и неврологических заболеваний на восприятие субсекундных временных интервалов (роль базальных ганглиев). Успехи физиологических наук. 2017. 48 (4): 78–94.
Симонов П.В. Мотивированный мозг. М.: Наука. 1987. 267 с.
Таротин И.В., Атанов М.С., Иваницкий Г.А. Макет устройства слежения за когнитивной деятельностью человека в реальном времени (“когновизор”). Журн. высш. нервн. деят. им. И.П. Павлова. 2017. 67 (4): 493–503.
Хофман И. Активная память. М.: Прогресс, 1986. 310 с.
Шамис А.Л. Поведение, восприятие, мышление. М.: Едиториал УРСС. 2005. С. 224.
Cohen B.H. The Motor Theory of Voluntary Thinking. Consciousness and self-regulation. Eds R.J. Davidson et al. N.Y.: Plenum Press, 1986. V. 4. 19–37 pp.
Cowan N. What are the differences between long-term, short-term, and working memory? Progress in Brain Research. 2008. 169: 323–338.
Crick F., Koch C. Some reflections on visual awareness. Gold Spring Harbor Symp. on Quantit. Biol. 1990. Vol. LV: 953–962.
Creutzfeldt O.D. Generality of the Functional Structure of the Neocortex. Naturwissenschaften. 1977. 64: 507–517.
Edelman G.M. Group selection and phasic reentrant signaling: A theory of higher brain function. In The Mindful Brain. Cambridge: MIT Press, 1978. 50–100 pp.
Edelman G.M. The remembered present. A biological theory of consciousness. N.Y.: Basic Books, 1989. 346 p.
Edelman G.M., Tononi G.A. Universe of Consciousness. N. Y.: Basic Books, 2000. 274 p.
Halford G.S., Cowan N., Andrews G. Separating cognitive capacity from knowledge: A new hypothesis. Trends in Cognitive Sciences. 2007. 11 (6): 236–242.
Hebb D.O. The Organization of Behavior. New York: Wiley, 1949. 335 p.
Hubel D.H., Wiesel T.N. Sequence, regularity and geometry of orientation columns in the monkey striate cortex. J. Comp. Neurol. 1974. 158: 267–294.
Hubel D.H., Wiesel T.N. Functional architecture of macaque monkey cortex. Proc. R. Soc. Lond. 1977. B198: 1–59.
Kihlstrom J.F., Hoyt I.P. Hypnosis and the Psychology Delusions. Delusional Beliefs. Eds. T.F. Oltmanns & Brendan A.M. New York: Wiley. 1988. Chapter 4. 66–107 pp.
Koch C., Crick F. Some Further Ideas Regarding the Neuronal Basis of Awareness. Large-Scale Neuronal Theories of the Brain. Eds. Koch, C. and Davis, J. Cambridge, MA: MIT Press. 1994. 93–110 pp.
König P., Engel A.K., Singer W. Integrator or coincidence detector? The role of the cortical neuron revisited. Trends Neurosci. 1996. 19: 130–137.
Mountcastle V.B. An Organizing Principle for Cerebral Function: The Unit Model and the Distributed System. Eds. Edelman G.M. and Mountcastle V.B. The Mindful Brain. Cambridge, Mass.: MIT Press, 1978. 7–50 pp.
Mountcastle V.B. Perceptual Neuroscience: The Cerebral Cortex. Cambridge, Mass.: Harvard University Press, 1998. 512 p.
Pascual-Leone F., Walsh V. Fast back projections from the motion to the primary visual area necessary for visual awareness. Science. 2001. 292 (5516): 510–512.
Phillips W.A. Short-term visual memory. Phil. Trans. Roy. Soc. Lond. 1983. B 302. 295–309 pp.
Sergin V.Ya. Brain as Neurocomputer. The Macrostructure of Inteligence. In A.V. Holden & V.I. Kryukov (Eds.) Neurocomputers and Attention. Manchester: Manchester University Press. 1991. 2: 771–781.
Sergin V.Ya. A Global Model of Human Mentality. “Cybernetics and Systems’ 92”. World Scientific Publishing Co. 1992. 1: 883–890 pp.
Sergin V.Ya. Consciousness as a data-processing system. Neural Network World. 1994. 4 (5): 601–608.
Sergin V.Ya. Self-Identification and Sensori-Motor Rehearsal as Key Mechanisms of Consciousness. International Journal of Computing Anticipatory Systems. 1999. 4: 81–99.
Sergin V.Ya. Nature of Sensory Awareness: The Hypothesis of Self-Identification. In Complex Brain Function: Conceptual Advances in Russian Neurosciences. Eds. Miller R., Ivanitsky A.M., Balaban P.M. Harwood Academic Publishers. 2000. 97–112 pp.
Sergin V.Ya. Perceptual binding of sensory events: the hypothesis of inclusive characteristics. Neuroscience and Behavioral Physiology. 2003. 33 (8): 753–761.
Sergin A.V., Sergin V.Ya. Model of perception: The hierarchy of inclusive sensory characteristics and top-down cascade transfer of excitation. Neural Network World. 2008. 18 (3): 227–244.
Sergin V.Ya. Auto-Identification of Neuronal Activity Patterns as a Physiological Mechanism of Awareness. Neuroscience and Behavioral Physiology. 2017. 47 (7): 1–14. https://doi.org/10.1007/s11055-017-0462-7
Standring S. (Ed.). Gray’s anatomy: The anatomical basis of clinical practice (39th ed.). Edinburgh: Churchill Livingstone. 2005. 1627 p.
Szentagothai J. The 'module-concept' in cerebral cortex architecture. Brain Res. 1975. 95: 475–496.
Walsh V., Cowey A. Transcranial magnetic stimulation and cognitive neuroscience. Nature Reviews. Neuroscience. 2000. I (1): 73–79.
West M. Meditation and self-awareness: psychological and phenomenological approaches. Aspects of Consciousness. Eds. G. Underwood, R. Stevens. L: Acad. Press. 1982. V. 3. P. 199.
Zeki S.M. A vision of the brain. Blackwell Scientific Publications. 1993. 366 p.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова